zum einfluß molekularer parameter der pektinsubstrate auf die aktivität von pektinesterasen aus...
TRANSCRIPT

Die Nahrung 28, 5, 1984,507-516
Zentralinstitut fur Ernahrung in Potsdam-Rehbriicke (Direktor: Prof. Dr. H. SCHMANDKE), Forschungszentrum fur Molekularbiologie und Medizin der AdW der DDR
Zum EinfluD molekularer Parameter der Pektinsubstrate auf die Aktivitat von Pektinesterasen aus Aspergillus niger und aus hoheren Pflanzen G. DONGOWSKI und W. BOCK
Die Aktivitat der Pektinesterasen (PE) wird in starkem MaDe vom Veresterungsgrad (VG) sowie von der Verteilung der freien und veresterten Carboxylgruppen im makromolekularen Pektinsubstrat beeinfluDt. An partiell alkalisch entesterten Pektinsubstraten ergibt die PE aus A . niger mit steigendem VG eine zuneh- mende Aktivitat, wahrend fur PE-Praparate aus Orangen, Tomaten und WeiBkohl optimale Aktivitatsbe- reiche bei Veresterungsgraden um 30 %, 50 % bzw. 40 ”/, nachgewiesen werden. Der EinfluB der Molmasse des Pektinsubstrats, gemessen als Viskositatszahl [ q ] , 1st hingegen in einem weiten Bereich relativ gering. Pektinsubstrate mit einer blockweisen Verteilung der Estermethoxylgruppen, die bei einer vorhergehenden Einwirkung von pflanzlicher PE entsteht, werden durch die PE aus Orangen langsamer und die PE aus A . niger schneller als die partiell alkalisch entesterten Pektine gespalten. In umgekehrter Weise wirkt sich eine vorhergehende partielle Entesterung der Pektinsubstrate durch die Pektinesterase aus A . niger auf die Aktivitat der untersuchten PE-Praparate aus.
Einleitung
Die Untersuchungen zur Wirkungsweise von Pektinesterase (PE) (EC 3.1.1.1 1 .) wurden in der Vergangenheit bevorzugt mit mehr oder weniger einheitlichen Enzympraparaten aus hoheren Pflanzen durchgefuhrt [ 1-61, Diese verursachen eine blockweise Entesterung des Pektinsubstrats (Einkettenmechanismus) [7]. Demgegenuber hydrolysieren PE aus ver- schiedenen Aspergillus-Species die Estermethoxylgruppen nach einem statistischen Me- chanismus [8- 1 11, der auch fur die partielle Pektinentesterung durch Alkalilaugen und Sauren charakteristisch ist.
Bei der Bestimmung der Aktivitat von Pektinesterasen wird der Auswahl des Pektin- substrates bisher relativ wenig Aufmerksamkeit gewidmet. Im folgenden wird der EinfluD des Veresterungsgrades (VG), der Verteilung der Estermethoxylgruppen und der Viskosi- tatszahl [q] der Pektinsubstrate auf die Aktivitat von gereinigter PE aus Aspergillus niger und von partiell gereinigten pflanzlichen PE untersucht.
Material und Methoden
Praparierung und Charakterisierung der Substrate
gesetzt [12]. Es werden handelsiibliche und im Labor praparierte, gereinigte Apfel- und Citruspektine als Substrate ein-
Zur partiellen alkalischen Entesterung werden hochveresterte Pektine in waBriger Losung nach der pH-
32’

508 DONG~WSKI/BOCK
stat-Methode bei einem pH-Wert von 10.0 bis zum Verbrauch einer vorher berechneten Menge an Natron- lauge behandelt. Die partielle enzymatische Entesterung erfolgt in gleicher Weise mit PE aus Orangen (pH 7 3 ) bzw. A. niger(pH 4.5). Dann wird aufeinen pH-Wert von i 3.0 angesauert und zur Enzyminaktivierung 15 min auf > 95 ;C erhitzt. Die Praparate werden dit Ethanol gefallt und gewaschen sowie getrocknet.
Durch mechanolytischen Abbau in trockenem Zustand wird die Molmasse der Pektine erniedrigt [ 131. Es werden der Gehalt an Galakturonan nach der Carbazolmethode und der Veresterungsgrad nach der
Chromotropsauremethode bestimmt [l4]. Die Viskositatszahl [q ] , die in Beziehung zur Molmasse steht [15], wird bei hochveresterten Pektinen in 0.155 M Natriumchlorid und bei niederveresterten Pektinen in 0,05 M Natriumchlorid,'0.005 M Natriumoxalat in einem UBBELOHDE-ViSkOSimeter bei (25 & 0,OS)"C gemessen und auf den Gehalt an Galakturonan bezogen.
Prupurierung der Pektinesterasen
Pektinester-use uus Aspergillus niger
Ein Kulturfiltrat von Aspergillus niger wird durch Ethanol- und Ammoniumsulfatfallung vorgereinigt. Bei der anschlieDenden Perkolation iiber DEAE-Sephadex A 50 (0,05 M MCILVAIN-Puffer, pH 6,O) wird im Unterschied zur Hauptmenge der Polygalakturonase (PG) nur die Pektinesterase gebunden. Die Elution der PE erfolgt mittels eines Natriumchloridgradienten. Das an DEAE-Sephadex A 50 rechromatographierte PE-Praparat enthllt nur noch LuDerst geringe PG-Spuren. Fur die kinetischen Untersuchungen wird es durch priparative Elektrofokussierung vollstandig von PG befreit.
Pektinesterusen uus Orungen. Tomuten und Weipkohl
Homogenisierte Navelorangen werden mit I M Natriumacetat bei pH 8,O und 4 "C extrahiert. Der Extrakt wird dreimal durch fraktionierte Ammoniumsulfatfdllung (30 bis 75 9 , Sattigung) vorgereinigt. Die PE wird rnit Wasser sowie 0,25 M Natriumchlorid extrahiert.
Rotreife Tomaten werden homogenisiert und 1 h bei 3000 U m i n zentrifugiert. Der hierbei erhaltene Riick- stand wird mit der gleichen Menge an 1O0.,,iger Natriumchloridlosung 1 h geruhrt und dann 16 h bei 4 "C extrahiert. Der durch Zentrifugieren abgetrennte enzymaktive Extrakt wird stufenweise rnit Ammonium- sulfat (30 bis 70 Sattigung) gefallt. Das dialysierte Rohenzympraparat wird in 0,02 N Dinatriumphosphat- losung, pH 8.0. gelost, mit Calciumphosphatgel 30 rnin unter Riihren behandelt und zentrifugiert. Dieser Vorgang wird bis Zuni negativen Nachweis der PG mindestens fiinfmal wiederholt.
Aus WeiDkohl wird ein urn 0 C lagerstabiles Acetontrockenpraparat hergestellt, das 16 h bei 4 "C mit 0,5 M Natriumacetatlbsung extrahien wird. Der durch Abpressen und Zentrifugieren isolierte Extrakt wird dreimal durch Ammoniumsulfat (30 bis 70 0" Sattigung) gefallt.
Die entsalzten pflanzlichen PE-Praparate werden in 0.025 M Acetatpuffer (pH 6,O) an Sephadex G 100 weiter gereinigt. dann dialysiert und gefriergetrocknet.
Bestimniung der Prktinesteraseuktivirat
Die Aktivitat wird bei 30 -C und einem pH-Wert von 4.5 (PE aus .4. niger) bzw. 7.5 (pflanzliche PE) in einer 0.5 Galakturonan enthaltenden Substratldsung in Gegenwart von 0,15 M Natriumchlorid und 0.0025 M Natriumoxalat nach der pH-stat-Methode (Titration mit 0,05 M Natronlauge) ermittelt. Eiiie Pektinesterase- Einheit ist der Emsatz \on 1 pmol Estermethoxylgruppen pro min und mg Enzympraparat innerhalb der linearen Reakt ionsphase.
Best irnni irnp (lei k inet ischen Ptrrtrniet er
Zur Ermittlung drr kinetischen Parameter wird die Enzymaktivitat in Gegenwart einer Substratkonzen- tration zwischen 0.031 und 0,257, Galakturonan bei 30 C und bci pH-Werten von 4,5 (PE aus A . niger) bzw. 7.5 (Orangen-PE) bestimmt. Die Auswertung erfolgt nach LINEWEAVER-BURK. Der K,,-Wert wird in mrnol Galakturonsdure-Einheiten angegeben.
Aktivitat von Pektinesterasen 509
Ergebnisse.
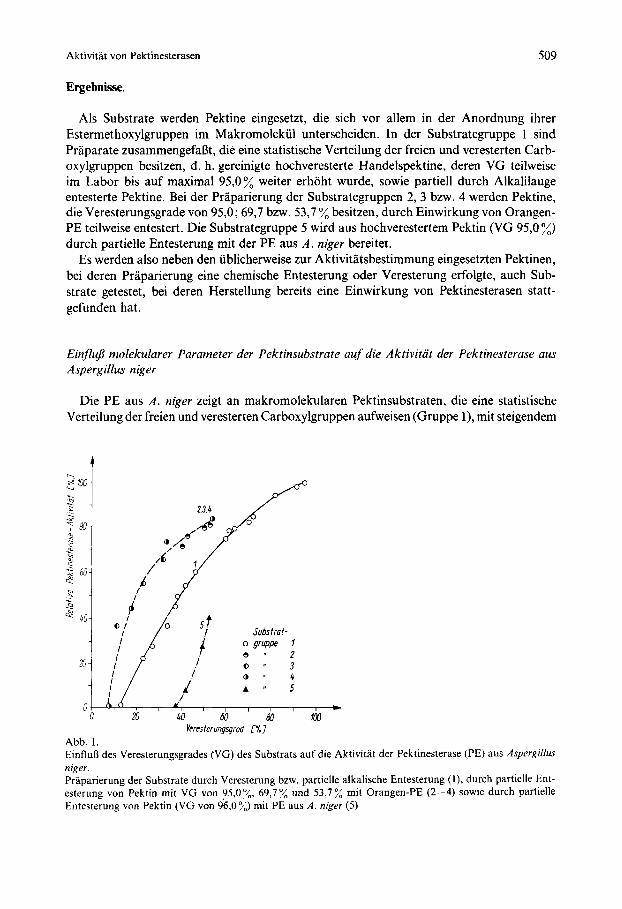
Als Substrate werden Pektine eingesetzt, die sich vor allem in der Anordnung ihrer Estermethoxylgruppen im Makromolekul unterscheiden. In der Substrategruppe 1 sind Praparate zusammengefafit, die eine statistische Verteilung der freien und veresterten Carb- oxylgruppen besitzen, d. h. gereinigte hochveresterte Handelspektine, deren VG teilweise im Labor bis auf maximal 95,0% weiter erhoht wurde, sowie partiell durch Alkalilauge entesterte Pektine. Bei der Praparierung der Substrategruppen 2, 3 bzw. 4 werden Pektine, die Veresterungsgrade von 95,O; 69,7 bzw. 53,7 % besitzen, durch Einwirkung von Orangen- PE teilweise entestert. Die Substrategruppe 5 wird aus hochverestertem Pektin (VG 95,O %) durch partielle Entesterung rnit der PE aus A . niger bereitet.
Es werden also neben den iiblichenveise zur Aktivitatsbestimmung eingesetzten Pektinen, bei deren Praparierung eine chemische Entesterung oder Veresterung erfolgte, auch Sub- strate getestet, bei deren Herstellung bereits eine Einwirkung von Pektinesterasen statt- gefunden hat.
EinjluJ molekularer Parameter der Pektinsubstrate auf die Aktivitat der Pektinesterase aus Aspergillus niger
Die PE aus A . niger zeigt an makromolekularen Pektinsubstraten, die eine statistische Verteilung der freien und veresterten Carboxylgruppen aufweisen (Gruppe l), mit steigendem
Veresterungsgrod PI*] Abb. 1. EinfluD des Veresterungsgrades CVG) des Substrats auf die Aktivitit der Pektinesterase (PE) aus Aspergillus nigrr. Praparierung der Substrate durch Veresterung bzw. partielle alkalische Entesterung ( I ) , durch partielle Ent- esterung von Pektin mit VG von 95,0%, 69,7% und 53,7% mit Orangen-PE (2-4) sowie durch partielle Entesterung von Pektin (VG vm 9S,O%) rnit PE aus A . niger ( 5 )

510 DONGOWSKI~BOCK
Veresterungsgrad im untersuchten Bereich von 12,8 bis 95,O :,, eine zunehmende Aktivitat (Abb. I ) . Dieses Enzym bevorzugt die am hochsten veresterten Pektine als Substrat und zeigt an ihnen auch die langste Reaktionsphase nullter Ordnung.
An Pektinsubstraten, bei deren Praparierung bereits eine partielle Entesterung dureh PE aus Orangen erfolgte, ergibt die Pektinesterase aus A . niger eine hohere Aktivitat als an den Substraten der Gruppe 1 (Abb. I). Der VG der Ausgangspektine (95,O bis 53,773 vor dieser Einwirkung des pflanzlichen Enzyms hat offensichtlich keinen EinfluB auf die ermittelte Aktivitat. Es wird eine gemeinsame Kurve fur die Gruppen 2 bis 4 erhalten. In diese ordnet sich auch ein handelsiibliches Citruspektin ein. Wahrend seiner Gewinnung ist offensichtlich neben der saurehydrolytischen auch eine enzymatische Entesterung durch fruchteigene PE eingetreten. Andererseits werden an den Pektinen der Gruppe 5 unerwartet niedrige Aktivitaten mit der PE aus A . niger gemessen. Diese Substrate wurden bereits bei ihrer Praparierung mit dem gleichen Enzym behandelt.
Die PE-Aktivitat wird von der Molmasse des Pektins nicht beeinflufit, wenn die Viskosi- tatszahl [q] oberhalb von 100 ml/g liegt. Pektine von niedrigerem Molekulargewicht werden jedoch langsamer entestert. Dieser Zusammenhang gilt unabhangig vom VG des Substrats (Tab. I ) . Die Messungen wurden an Pektinen der Gruppe 1 durchgefuhrt.
EinjluJ molekulurer Parameter der Pektinsuhstrate auf die .4ktivitar der Pektinesterusen aus Orangen, Tomaten und We@iohl
Die untersuchten pflanzlichen PE-Praparate entfalten an makromolekularen Pektinen, die eine statistische Verteilung der Estermethoxylgruppen aufweisen (Gruppe I) , ihre hochste Aktivitat im mittleren VG-Bereich (Abb. 2 ) . Bei Veresterungsgraden unterhalb von etwa 25 7;. aber auch oberhalb von 60 "" wird in unterschiedlichem AusmaR eine geringere Aktivi- tat ermittelt. So ist die WeiDkohl-PE bei Veresterungsgraden zwischen 45 und 55 % maximal aktiv (Kurve IV). Beim PE-Praparat aus Orangen nimmt die Aktivitat zunachst mit fallen- dem VG bis etwa 45 "/, kontinuierlich zu und bleibt dann bis zu einem Veresterungsgrad von etwa 25 ' lo weitgehend konstant (Kurve 11). Das PE-Praparat aus Tomaten (Kurve 111) besitzt einen relativ breiten optimalen VG-Bereich (25 bis 55 9.:). In jedem Fall verringert sich jedoch die Reaktionsphase nullter Ordnung mit abnehmendem Veresterungsgrad.
An Substraten, bei deren Praparierung bereits eine partielle enzymatische Entesterung erfolgte, werden im Vergleich zu Pektinen der Gruppe 1 veranderte Aktivitaten bei der Be- stimmung der Orangen-PE erhalten (Abb. 3). Eine derartige Vorbehandlung des Pektins durch das gleiche pflanzliche Enzym (Gruppe 2 bis 4) bewirkt, daD wesentlich niedrigere Aktivitaten ermittelt werden. Dieses trifft besonders bei Vorliegen einer weitgehend idealen blockweisen Anordnung der freien Carboxylgruppen im Pektinmolekiil zu (Gruppe 2).
An Substraten der Gruppe 5 zeigt die Orangen-PE dagegen eine hohere Reaktionsge- schwindigkeit als an den partiell alkalisch entesterten Pektinen (Gruppe l), obwohl in beiden Fallen eine statistische Anordnung der freien und veresterten Carboxylgruppen vorliegt.
Die Molmasse des Substrats hat in einem weiten Bereich ebenfalls keinen EinfluD auf die Aktivitiit der untersuchten pflanzlichen Pektinesterasepriiparate (Tab. 1). Bekanntlich konnen besonders hochmolekulare, niederveresterte Pektine schwer loslich und salzempfind- lich sein. In diesem Fall ist ein partieller Abbau, z. B. durch Mechanolyse, zu empfehlen. Es muD jedoch gewahrleistet sein. daD die Viskositatszahl [,7] dieses abgebauten Pektins noch oberhalb von 100 ml/g liegt.

Aktivitat von Pektinesterasen 51 1
I
I k r' i i i
0 20 10 w 80 IW Veresterungsgrad P/J '
Abb. 2. Aktivitat von Pektinesterasepraparaten aus Aspergillus niger (I), Orangen (II), Tomaten (111) und WeiDkohl (IV) in Abhangigkeit vom Veresterungsgrad hochmolekularer Pektinsubstrate rnit sta- tistischer Verteilung der Estermethoxylgruppen (Substrategruppe 1)
EinfluP von Molmasse und Veresterungsgrad der Pektinsubstrate auf die kinetischen Parameter der Pektinesterasen aus Aspergillus niger und Orangen
An hochmolekularen Substraten rnit statistischer Verteilung der Estermethoxylgruppen (Gruppe 1) erniedrigt sich im untersuchten Bereich der V,,,-Wert mit fallendem Vereste- rungsgrad (95,O bis 37,779 bei der PE aus A. niger, wahrend er bei der Orangen-PE an- steigt (Tab. 2, Substrate A, B und D). Fur den K,,,-Wert wird ein entgegengesetzter Zusam- menhang ermittelt. Diese Versuche zeigen ebenfalls, daB die Pektinesterase aus Schirnmel- pilzen hochveresterte und das pflanzliche Enzym mittelveresterte Pektine als Substrat be- vorzugen. Am stark abgebauten Pektin C, das nur eine Viskositatszahl [q] von 34 ml/g besitzt, wird fur das PE-Praparat aus Orangen im Unterschied zum makromolekularen Substrat B bei vergleichbarem VG ein kleinerer V,,,,,- und ein groBerer K,-Wert bestimmt.
Die Substrate D, E und F, deren Veresterungsgrad urn 40% liegt, wurden durch eine vorhergehende partielle Entesterung eines zu 95 % veresterten Pektins rnit Natronlauge sowie mit den PE aus Orangen oder A. niger hergestellt. Der jeweils hochste K,-Wert wird fur die Pektinesterasen aus A . niger bzw. Orangen an den Pektinsubstraten F bzw: E ge- messen.

DONGQWSKI/BOCK
Abb. 3. EinfluB des Veresterungsgrades (VG) des Substrats auf die Aktivitat der Pektinesterase (PE) aus Orangen. Praparierung der Substrate durch Veresterung bm. partielle alkali- sche Entesterung ( I ) , durch partielle Entesterung von Pektin mit VG von 95,00;, 69,7% und 53,73; mit Orangen-PE (2-4) sowie durch partielle Entesterung von Pektin (VG von 95,0%) mit PE aus A . niger (5)
/
P I I
9' / I I I I A
P 2/
d Substrat-
o qruppe 1 e " 2 0 " 3 a 4 A " 5
o i , , , 1 1 I I I I I c 0 28 $0 60 Bo I!%
Veresterungsgra;? !%I
Tabelle 1 EinfluD der Viskositltszahl [q] der Pektinsubstrate (Praparate der Gruppe 1) auf die Aktivitat der Pektinesterasen (PE) aus Aspergillus niger und hoheren Pflanzen
Pektinsubstrat PE-Einheiten [pmol/rnin mg] des Praparates aus
Veresterungsgrad [VI 4 niger Orangen Tomaten WeiBkohl [3J [ml gl
69,l
59,7
33.9
500 34 I 94 25
394 259 98 34 17
30 I 160 130 105 76 19
24 1 24 1 24 I 194
219 219 215 203 I75
118 I18 118 I18 115 97
6,13 6,13 6.05 5,98
6,68 6,68 6.68 6,30 6.35
8,98 8,95 9,OO 9,OO 8.80 8,575
9,30 9,30 9,30 9.10
10,35 10.35 9,80 9,35 9.15
1 l,05 11.10 11,05 11.05 10.80 10,20
3.68 3,68 3,63 3,45
3.88 3,88 3,88 3,75 3.68
3,88 3.88 4,OO 3.90 3,75 3,50

Aktivitat von Pektinesterasen 513
Tabelle 2 EinfluD molekularer Substratparameter auf die kinetischen Wirkungsparameter der Pektin- esterasen (PE) aus Aspergillus niger und Orangen
Pektinsubstrat A . riiger-PE Orangen-PE
Nr. Gruppe Vereste- Viskositats- V,,, Km Vmax Km rungsgrad zahl [q] [pmol/min . mg] [mmol] [pmol/min . mg] [mmol] [ %I [ m k l
~
A 1 95,O 250 33,3 5,9 5,4 I 5,16 B 1 62.0 369 31,9 10,8 6.84 1,78 C I 59,7 34 - - 6,29 1,91 D 1 37.7 125 26,2 30,O 8,68 1,47 E 2 40.7 394 27,8 21,6 4,65 4,69 F 5 37,5 298 2,3 71,O 10,48 2,03
Diskussion der Ergebnisse
Die molekularen Parameter des Substrats beeinflussen nicht nur die Aktivitat der de- polymerisierenden pektinolytischen Enzyme [ 12; 16 ; 171, sondern auch der Pektinesterasen. Hierbei zeigen sich zusatzlich bei PE verschiedener Herkunft neben Gemeinsarnkeiten, z. B. in bezug auf die Molmasse des Substrats, einige charakteristische Unterschiede, die vor allem vom Veresterungsgrad und der Art der Verteilung der freien und veresterten Carboxylgruppen im Pektinmolekiil hervorgerufen werden.
Fur die untersuchten pflanzlichen PE-Praparate wird neben den bekannten pH- und Temperaturoptima auch ein optimaler Bereich des VG gefunden. In jedem Fall werden hier die maximalen Aktivitaten an mittelveresterten Pektinen bestimmt. Eine Ursache fur die unterschiedlichen Aktivitatsprofile diirfte die Zusammensetzung der multiplen Enzym- formen sein. In Enzympraparaten aus Citrusfriichten konnten beispielsweise bis zu 12 PE-Formen mit teilweise unterschiedlichen Eigenschaften isoliert werden [6]. Zur Ausbil- dung des Enzym-Substrat-Komplexes sind bei pflanzlichen PE freie Carboxylgruppen erforderlich [2; 41.
Diese Voraussetzung erfiillen mittelveresterte Pektine besser als hochveresterte. Da andererseits Pektinsaure die pflanzliche PE hemmt [6], verlangsamt sich unterhalb eines VG von 25 % wegen des zunehmenden ,,Pektinsaurecharakters" des Substrats die Reaktions- geschwindigkeit stark und kommt bei einem Veresterungsgrad um 10 % praktisch zum Stillstand [2].
Bei verschiedenen mikrobiellen Pektinesterasen, z. B. aus Coniothyrium dipodiella [ 181 und Corticium rolfsii [19], wurde in ahnlicher Weise wie bei der hier untersuchten PE aus A . niger an Substraten mit statistischer Verteilung der Estermethoxylgruppen eine zuneh- mende Aktivitat mit steigendem Veresterungsgrad ermittelt. LIFSHITS u. a. [20; 211 fanden eine lineare Beziehung zwischen der PE-Aktivitat und dem VG. Diese Messungen wurden allerdings in Gegenwart von Polygalakturonase und nicht bei konstantem pH-Wert durchge- fuhrt. Der EinfluB des VG auf die Aktivitat deutet bei der PE aus A . niger im Vergleich zu den pflanzlichen Enzymen [6; 71 auf einen andersartigen Entesterungsmechanismus. Nach ISHII u. a. [8; 221 sowie BARON u. a. [9] sind mit Schimmelpilz-PE bereitete niederveresterte Pektine in bezug auf die Verteilung der veresterten und freien Carboxylgruppen homogen.

In gleicher Weise wirkt die hier untersuchte PE aus A . niger [lo; 23; 241. Pektinesterasen aus hoheren Pflanzen und Schimmelpilzen unterscheiden sich also nicht nur im bevorzugten Substrat. sondern auch in den entstehenden Reaktionsprodukten.
Die durchgefuhrten Aktivitatsbestimmungen deuten auch darauf hin, dal3 bei der par- tiellen Entesterung durch Alkalilauge und Schimmelpilz-PE keine vollstandig identische Anordnung der Estermethoxylgruppen im Pektinsubstrat erhalten wird.
Kiirzlich wurde gezeigt. dal3 PE aus Tomaten bzw. Aspergi//us./oeiidus als kleinstes Sub- strat veresterte Penta- bzw. Digalakturonsaure angreifen. Im oligomeren Bereich erhoht sich die Reaktionsgeschwindigkeit rnit dem Polymerisationsgrad [25]. Bereits SOLMS u. a. [2] wiesen darauf hin, dal3 ein zuvor rnit Orangen-PE partiell entestertes Pektin ein schlech- teres Substrat fur dasselbe pflanzliche Enzym ist als ein auf alkalischem Wege teilweise entestertes Pektin. Unsere Versuche zeigen dariiber hinaus, dal3 eine Vorbehandlung des Substrats mit der PE aus A . rzigrr die Aktivitit der Orangen-PE deutlich fordert. Offen- sichtlich entstehen durch die Einwirkung dieser mikrobiellen PE freie Carboxylgruppen an Positionen, die fur eine Ausbildung des Enzym-Substrat-Komplexes bei den pflanzlichen Pektinesterasen besonders giinstig sind.
Die aufgezeigten Wirkunterschiede der Pektinesterasen spielen eine Rolle bei verscbie- denen physiologischen und phytopathogenen Vorgangen, z. B. wahrend des Wachstums, der Reifung oder des mikrobiellen Befalls. aber auch bei der Herstellung fliissiger Produkte aus Obst und Gemiise [26; 271.
Es sollte beachtet werden, daB selbst hochveresterte handelsiibliche Pektine wahrend ihrer Produktion bei unsachgemaoer Technologie (2. B. bei der Fruchtsaftgewinnung oder bei der Zwischenlagerung der Trester bis zum Trocknen) durchaus rnit den pflanzeneigenen undioder mit mikrobiellen PE in Kontakt kommen konnen. Bei Einsatz derartiger Pektine als Substrat werden veranderte Pektinesteraseaktivitaten trotz gleichen Veresterungsgrades gemessen.
Die Beeinflussung der PE-Aktivitat durch die molekularen Parameter des Pektins er- schwert die Standardisierung des Analyseverfahrens. Offensichtlich gibt es kein Substrat, das fur alle Pektinesterasen in gleicher Weise geeignet ist. Der Einsatz des jeweils optimalen Substrats bei der Aktivitatsbestimmung wird nicht fur jedes Enzympraparat realisierbar sein. Zur Abschatzung von PE-Aktivitaten ist es deshalb erforderlich, auch Angaben uber den Veresterungsgrad und die Praparierungsweise der als Substrat eingesetzten Pektine zu machen. Fur den Routinebetrieb zur Aktivitatsbestimmung von Pektinesterasen kann ein hochmolekulares, 60 bis 65 7; verestertes Pektinsubstrat rnit statistischer Verteilung der Estermethoxylgruppen empfohlen werden.
Summary
G . DONCOWSKI and W. BOCK: On the effects of molecular parameters of pectinsubstrates on the activities of pectinesterases from Aspergdlta niger and from higher plants
The activities of the pectinesterases depend to a great extent upon the degree of esterification and upon the distribution of the free and esterified carboxyl groups in the macromolecular pectin substrate. On partially alkaline de-esterified pectin substrates. the activity of the pectinesterase from A . niger increases with the in- crease in degree of esterification, whereas pectinesterase preparations from oranges, tomatoes and white cabbage show optimal activity ranges at degrees of esterification of 30 ”/, 50 % and 40 7;. respectively. On the contrary, the effect of the molecular mass of the pectin substrate, measured as viscosity number [ q ] , is relatively

Aktivitat von Pektinesterasen 515
weak within a wide range. Pectin substrates with a blockwise distribution of the ester methoxyl groups, which is produced by the previous influence of vegetable pectinesterase, are more slowly (more rapidly) split by the pectinesterase from oranges (from A. niger) than the partially alkaline de-esterified pectins. Previous partial de-esterification of the pectin substrates by the pectinesterase from A. niger exerts just the opposite effect on the activities of the pectinesterase preparations studied.
r. flOHrOBCKM €I B. ~ O K K : BnHaHkie MOJIeKynRpHbIX napaMeTpOB IIeKTHHOBbIX Cy6CTpaTOB Ha aKTHBHOCTb n e ~ ~ ~ ~ 3 c ~ e p a 3 1(3 Aspergillus niger A 113 BbIcmix pacTeHuii
Literatur
[ I ] LINEWEAVER, H.. und E. F. JANSEN, Adv. Enzymol. 11. 267 (1951). [2] SOLMS, J., und H. DEUEL, Helv. Chim. Acta 38, 321 (1955). [3] LEE, M., und J. D. MACMILLAN, Biochemistry 7,4005 (1968). [4] REXOVA-BENKOVA. t., und 0. MARKOVI~ . Adv. Carbohydr. Chem. Biochem. 33. 323 (1976). [5] MARKOVI~ , 0.. Collect. Czech. Chem. Commun. 39, 908 (1974). [6] VERSTEEG, C., Diss., Landbouwhogeschool Wageningen/Niederlande (1979). [7] KOHN, R., I. FURDA und Z. KOPEC, Collect. Czech. Chem. Commun. 33, 264 (1968). [8] ISHII, S . , K. KITO. S. SUGIYAMA und H. SUGIMOTO, J. Food Sci. 44, 611 (1979). [9] BARON, A,, F. ROMBOUTS, J. DRILLEAU und W. PILNIK, Lebensm.-Wiss. Technol. 13,330 (1980).
[lo] DONGOWSKI, G., und W. BOCK, Nahrung 24, K 5 (1981). [ I l l KOHN, R., 0. MARKOVIC und E. MACHOVA, Collect. Czech. Chem. Commun. 48, 790 (1983). [I21 DONcowsKi, G., H. ANGER und W. BOCK, Nahrung 27, 817 (1983). [I31 BOCK, W., H. ANGER. R. KOHN. A. MALOVIKOVA, G. DONGOWSKI und R. FRIEBE, Angew. makromol.
[I41 KRAUSE M.. und W. BOCK, Ernahrungsforschung 18, I 1 1 (1973). [IS] BERTH, G., H. ANGER und F. LINOW, Nahrung 21,939 (1977). [I61 PILNIK, W., F. M. RoMBouTsund A. G. J . VORAGEN, Chem. Mikrobiol. Technol. Lebensm. 2,122(1973). [I71 DONGOWSKI, G., W. BOCK, 0. MARKOVIC und A. SLEZARIK. Nahrung 24, 619 (1980). [I81 ENDO, A,, Agric. biol. Chem. 28. 757 (1964). [I91 YOSHIHARA, O., T. MATSUO und A. KAJI, Agric. biol. Chem. 41, 2335 (1977). [20] LIFSHITS, D. B., D. E. PLOTKINA, D. S. SHMUILOVICH und M. G. DYSHKANT, Prikl. Biochim. Mikro-
[21] PLOTKINA, D. E., D. B. LIFSHITS. D. S. SHMUILOVICH und M. G. DYSHKANT, Prikl. Biochim. Mikrobiol.
Chem. 64, 133 (1977).
biol. 5, 491 (1969).
5, 623 (1969).

516 DONGOWSKI/BOCK
[22] ISHII, S.. K. KIHO, S . SUGIYAMA und H. SUGIMOTO, DT-OS 28 43 351 vom 4. 10. 1978, ert. 27. 3. 1980. [23] KOHN, R., G. DONGOWSKI und W. BOCK, Nahrung 29 (1985) (im Druck). [24] ANGER, H.. und G. DONGOWSKI, Nahrung 28, 199 (1984). [25] MARKOVI~: , O., E. MACHOVA und A. SLEZARIK, Carbohydr. Res. 116, 105 (1983). [26] PRESSEY, R . , ACS Symposium Series 47, 172 (1977). [27] ROMBOUTS, F. M.. und W. PILNIK, Pectic Enzymes. In: Economic Microbiology, Vol. 5, S. 227, Academic
Press. New York und London 1980.
Fur die gewissenhafte experimentelie Mitarbeit danken wir Frau ERIKA SCHNEIDER
Dr. G. DONGOWSKI und Dr. W. BOCK. Zentralinstitut fur Ernahrung, DDR-1505 Bergholz-Rehbriicke, Arthur-Scheunert-Allee 114- 116.
Eingegangen 27. 7. 1983