water deficit affects plant and soil water status, plant growth, and ginsenoside contents in...
TRANSCRIPT

Water Deficit Affects Plant and Soil Water Status, Plant Growth,
and Ginsenoside Contents in American Ginseng
Jinwook Lee1,2*
and Kenneth W. Mudge1
1Department of Horticulture, Cornell University, Ithaca, NY 14853, U.S.A.
2USDA-ARS, Tree Fruit Research Laboratory, 1104 N. Western Ave., Wenatchee, WA 98801, U.S.A.
*Corresponding author: [email protected], [email protected]
Received July 4, 2013 / Revised September 4, 2013 / Accepted November 27, 2013
Korean Society for Horticultural Science and Springer 2013
Abstract. American ginseng (Panax quinquefolius L.) produces pharmacologically active secondary compounds known
as ginsenosides which have been shown to be influenced by both genetic and environmental factors. In a greenhouse
experiment, effects of water deficit on ginseng plant growth, predawn leaf water potential ( Leaf), soil water potential
( Soil), leaf abscisic acid (ABA) concentration, and root ginsenoside contents as well as photosynthesis-related
physiological responses were studied. Three-year-old seedlings, grown in 200 mL volume of plastic pots, were well
watered for 45 days prior to the initiation of water deficit treatments. Plants in the water deficit treatments were irrigated
every 10 or 20 days for the mild and severe water deficit treatments, respectively, while the control plants were watered
every 4 days. The experiment was terminated after 15, 6, and 3 dry down cycles (60 days) for the control, mild,
and severe water deficit treatments, respectively. As water deficit progressed, both Soil and Leaf decreased, but foliar
ABA concentration increased. Other physiological responses to water deficit, including transpiration rate, stomatal
conductance, and CO2 assimilation rate, were decreased. Water deficit decreased root growth, but unaffected shoot
growth. Foliar chlorophyll content was also decreased in the water deficit treatments. The contents of individual
ginsenosides Re, Rb1, Rc and Rd, and total ginsenosides were increased in the storage roots of water deficit-treated
plants as compared with well-watered controls. Rootlet fresh weight before transplanting (RFWBT) as a covariate had
a significant effect on the contents of ginsenoside Rb1, Rc, and Rb2. Overall, the results indicate that water deficit
could contribute not only to reducing plant performance but also increasing the levels of ABA and certain ginsenoisdes.
Additional key words: abscisic acid (ABA), drought stress, evapotranspiration, leaf water potential, Panax
quinquefolium L., soil water potential
Hort. Environ. Biotechnol. 54(6):475-483. 2013.
DOI 10.1007/s13580-013-0090-2
ISSN (print) : 2211-3452
ISSN (online) : 2211-3460
Research Report
Introduction
American ginseng (Panax quinquefolius L.), belonging to
Araliaceae family, has been widely produced in North
America for use in both the West and in Asia for medicinal
purposes which are associated with the pharmaceutically
active secondary constituents known as ginsenosides (Attele
et al., 1999). The compositions and contents of individual
ginsenosides are influenced by both genetic (population)
and environmental (location) factors (Lim et al., 2005).
Assinewe et al. (2003) also reported population difference in
ginsenoside accumulation among ten geographically isolated
wild populations. The compositions and contents of gin-
senosides were also affected by numerous environmental
factors, including understory light levels (Fournier et al.,
2003), soil mineral nutrients (Konsler et al., 1990; Lee and
Mudge, 2013; Li and Mazza, 1999), plant growth regulators
(Barbara et al., 2006), elevated temperatures (Jochum et al.,
2007), and water stress (Lim et al., 2006).
Intensive field production of ginseng involves using an
artificial shade and irrigation (Li, 1995). Woods cultivation
is an alternative production system involving natural forest
shade of deciduous hardwood tree species (Beyfuss, 1999).
Unlike field cultivation, irrigation is not practiced and ginseng
is in competition with trees for soil moisture. Therefore,
water deficit stress may have a significant impact on ginseng
growth and ginsenoside accumulation. Li and Berard (1998)
demonstrated that reduced soil moisture levels decreased
root fresh weight (FW) and dry weight (DW) and root
diameter. Under the condition of water deficit, the emergence
of shoots and subsequent root fresh weight of Asian ginseng
(Panax ginseng C.A. Meyer) were decreased (Mork et al.,

Jinwook Lee and Kenneth W. Mudge476
1981). Moreover, reduced soil moisture managed by elevating
ginseng beds in Asian ginseng adversely affected net photo-
synthetic activity and transpiration rate (Lee et al., 1982).
Lim et al. (2006) demonstrated that the moderate water
stress increased the contents of certain individual and total
ginsenosides, but did not affect plant growth of American
ginseng.
Water deficit effects on gas exchange and abscisic acid
(ABA) accumulation have been well documented for many
horticultural and agronomic crops (Clemente and Marler,
1996; Delfine et al., 2002; Ennahli and Earl, 2005; Wang et
al., 2004). When available water is limited, ABA concen-
tration is elevated causing stomatal closure (Alves and
Setter, 2000; Wang et al., 2004). Photosynthesis and tran-
spiration are reduced by water deficit in cassava (Manihot
esculenta Crantz) (Alves and Setter, 2000) and sweet potato
(Ipomoea batatas Lam.) (Haimeirong and Kubota, 2003)
which have storage root systems similar to ginseng. Fur-
thermore, reduced soil moisture causes reduced quantum
yield of PSII in sweet potato (Haimeirong and Kubota, 2003)
and correspondingly decreased sink capacity. During the
growing season, the elevated temperatures decreased net
photosynthetic rate and stomatal conductance of American
ginseng plants cultivated in a greenhouse (Jochum et al.,
2007). However, there are no such reports of the effect of
water deficit on CO2 assimilation and ABA accumulation in
American ginseng. Therefore, the objectives of this experiment
were to test the hypothesis that water deficit treatment would
reduce American ginseng growth, decrease photosynthetic
activity, while increasing ABA concentration, and thereby
influence the compositions and contents of individual and
total ginsenosides. This information will be useful to under-
standing the physiological responses of American ginseng
to water deficit.
Materials and Methods
Three-year-old American ginseng (Panax quinquefolius
L.) rootlets were grown on a bed under the natural shade of
deciduous trees at a local ginseng forest farm in upstate New
York, U.S.A., harvested in November 2004, then transported
and stored at 4°C for 12 weeks in Ithaca, New York, U.S.A.
A greenhouse potting mixture, consisting of 1: 2: 1; soil:
peat : perlite by volume, was fully air-dried for 10 days in a
greenhouse. Four hundred grams of the air-dried potting
mixture was added to 200 mL volume of plastic pots (13 cm
top diameter, 8.8 cm bottom diameter, and 11.5 cm depth)
in which ginseng rootlets were planted. Fresh weight of
each rootlet was recorded and then transplanted into the
plastic pots containing the greenhouse potting mixture. The
experiment was laid out in a completely randomized design
with 60 replications (individual pots) per treatment. All
plants were well watered every other day for the first 45
days before initiation of water deficit treatments. The water
deficit treatments consisted of 4, 10, and 20 days of dry
down interval for the control, mild, and severe water deficit
treatments, respectively. Experimental irrigation was repeated
for 15, 6, and 3 dry down cycles for the control, mild water
deficit, and severe water deficit treatments, respectively,
over a 60 days period. The potted rootlets were maintained
under 70% shade with a polypropylene shade cloth in a
greenhouse at 20 ± 2°C with a below-bench evaporative
cooling system. Lighting from 60 W incandescent bulbs was
progressively adjusted to approximate natural growing
season photoperiod.
Ginseng plants were harvested after 105 days of greenhouse
cultivation, approximating the length of the normal growing
season in the field. After measuring shoot growth charac-
teristics, including prong number, leaflet number, and sym-
podium height, plants were dug to harvest roots for the
evaluation of root growth characteristics and ginsenoside
contents. After that, shoot FW, leaf area, root length, root
diameter, and root FW were measured. Before transplanting
to 200 mL plastic pots at the beginning of the water deficit
experiment, rootlets were weighed for rootlet fresh weight
before transplanting (RFWBT), total root length (the sum of
storage root length and the longest fibrous root length), and
root diameter were measured. The same root growth parameters
were measured after roots were harvested at the end of the
water deficit experiment. The percentage change in each
root growth (RG) parameter based on root fresh weight was
calculated using the following formula:
Root Growth Rate (%) =(RGAH - RGBT)
× 100RGBT
where the AH and BT stand for after harvesting (end of
experiment) and before transplanting (beginning), respectively.
The harvested ginseng root samples were placed on a forced-
air food dehydrator (American Harvest Forced Air Food
Dehydrator FD 50/30, NESCO®, Milwaukee, Wisconsin,
U.S.A.) at 35°C for 7 days before measuring root DW, and
subsequent analysis of root ginsenoside contents. Leaf area
was measured with a leaf area meter (LI-3100, LI-COR, Inc.,
Lincoln, Nebraska, U.S.A.) and then shoot tissues were oven
(1321F SHEL LAB Forced Air Ovens, Sheldon Manufacturing,
Inc., Cornelius, OR, U.S.A.) dried at 70°C for 3 days for the
measurement of shoot DW. All plants from each treatment
were 24 plants (n = 24).
During the water deficit experiment, the volumetric soil
moisture content ( ) was recorded daily using a Theta probe®

Hort. Environ. Biotechnol. 54(6):475-483. 2013. 477
(Delta-T Devices Ltd., Cambridge, U.K.). Soil water potential
( Soil) was calculated from using the regression equation
between and Soil (n = 24) determined psychrometrically
as described by Reaves (2003). Predawn leaf water potential
( Leaf) was measured on the largest (central) leaflet of a
compound leaf with a pressure bomb (Soil Moisture Equip-
ment Co., Santa Barbara, CA, U.S.A.) on five individual
plants per treatment (n = 5). Daily evapotranspiration was
calculated by subtracting pot weight day by day from initial
pot weight (n = 24).
At the end of the water deficit experiment, leaf gas
exchange was measured on the biggest leaflet located in the
middle of a compound leaf chosen for sampling on 10 plants
per treatment (n = 10) using a portable steady state gas-
exchange system (CIRAS-I, PP Systems, Herts, U.K.) at
ambient CO2 (360 mol·mol-1
). The gas exchange measure-
ments were taken in random order to compensate for any
effects caused by sampling time from 1000 to 1100 HR.
Leaf temperature within the cuvette was controlled at 20 ±
0.5°C and photosynthetic photon flux density (PPFD) was
maintained at 550 ± 50 mol·m-2
·s-1
. After the measurement
of gas exchange, leaf chlorophyll was extracted with 80%
(v/v) acetone from 10 plants per treatment (n = 10), and
absorbance was measured at 663 nm and 645 nm. The
contents of chlorophyll a, b and a + b were calculated as
described by Arnon (1949).
For ABA analysis, at the end of the water deficit ex-
periment, a 1 cm diameter leaf disc was taken by a cork
borer, placed into 250 L ice-chilled solution of 80% (v/v)
ethanol, and then stored at -20°C freezer until ABA analysis.
The ABA analysis was performed by using enzyme-linked
immuno-sorbent assay (ELISA) as described by Alves and
Setter (2000) with the slight modification that ABA extracts
were finally incubated in 200 L of 0.9 M diethanolamine
(DEA) buffer with 0.2 g para-nitrophenylphosphate (PNPP)
for 150 min at room temperature before measurement.
The analysis of root ginsenoside contents followed Lee
and Mudge (2013). One hundred milligrams dried ground
ginseng root tissue which was screened with a 60 mesh was
extracted in 30 mL of 70% (v/v) MeOH. The MeOH extract
was vacuum-evaporated at 38°C with a rotary evaporator
(Buchi 011, BUCHI Analytical Inc., New Castle, DE, U.S.A.),
redissolved in 5 mL of 100% MeOH and dried with a rotary
evaporator (Buchi 011, BUCHI Analytical Inc., New Castle,
DE., U.S.A.). The residue was redissolved in 500 L of
16% acetonitrile before injection of 15 L into the HPLC.
The HPLC system for ginsenoside analysis was a Waters
2690 Separations Module HPLC with Waters 996 Photodiode
Array Detector at 203 nm. Empower Pro software (Build
1154, Waters Co., Milford, MA, U.S.A.) was used for the
solvent gradient and peak identification and integration. The
reversed phase C18 column (Varian HPLC Columns, Varian
Inc., Lake Forest, CA, U.S.A.) was used with a guard column
(Reversed Phase ChromSep Guard Column SS, Varian Inc.,
Lake Forest, CA, U.S.A.). A gradient of the eluents (A)
0.14% phosphate buffer and (B) 100% acetonitrile was used
as follows: 0-20 min, 84-82% A, 16-18% B; 20-60 min,
82-60% A, 18-40% B. The flow rate was 1.15 mL·min-1
. As
an internal standard, m-cresol (Sigma Chemical Co., St. Louis,
MO, U.S.A.) was added into each sample to confirm the
injection volume and retention time per injection. Individual
ginsenosides from the extracts were identified and quantified
by retention time and peak areas as compared with those of
authentic ginsenoside standards Rg1, Re, Rb1, Rc, Rb2, and
Rd (Indofine Chemical Co., Hillsborough, NJ, U.S.A.).
Each dependent variable (plant growth characteristics, leaf
chlorophyll content, gas exchange, ABA concentration, and
root ginsenosides) was statistically analyzed using the analysis
of variance (version 8.02; SAS Institute, Cary, NC, U.S.A.).
Rootlet fresh weight before transplanting (RFWBT) was used
as a covariate to evaluate variation among treatments. Fol-
lowing analysis of variance, mean separation was performed
by Duncan’s multiple range tests at 5% level.
Results
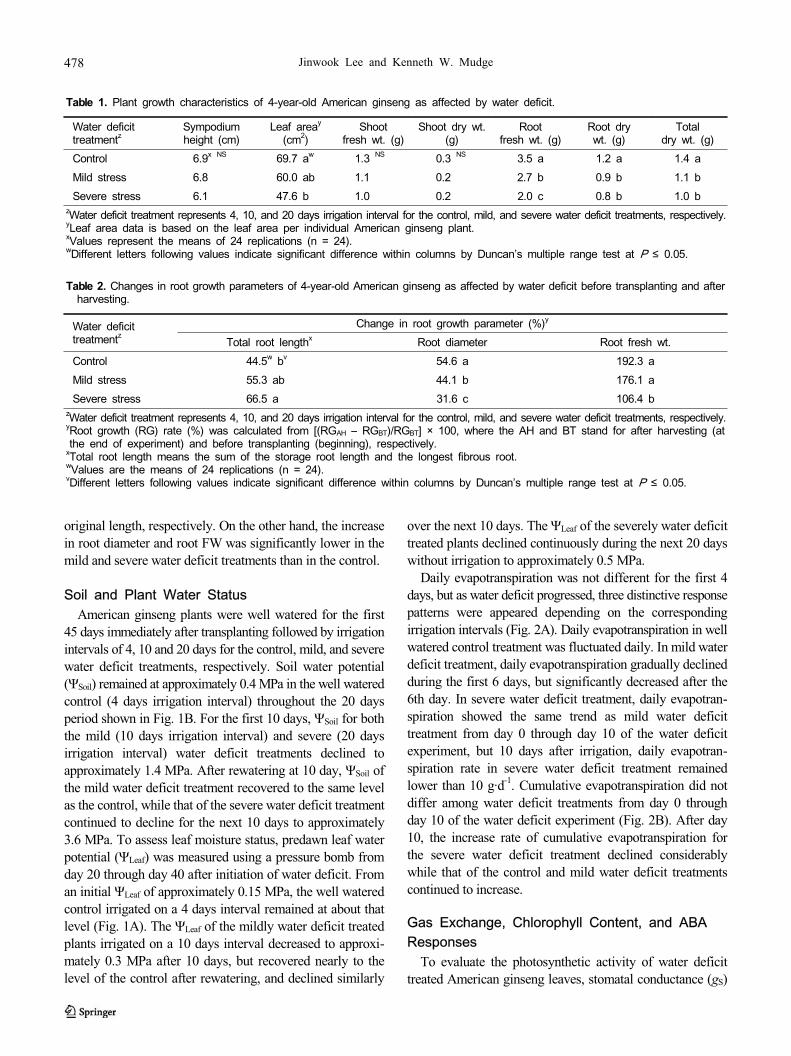
At the end of the water deficit experiment, shoot growth
was not significantly affected by water deficit treatment
except for leaf area per plant (Table 1). Leaf area in the mild
and severe water deficit treatments was 14 and 32% less
than that in the control, respectively. However, root FW and
DW were significantly decreased by water deficit. Compared
with the control, root DW was decreased by 24 and 35% for
mild and severe water deficit treatments, respectively. Fur-
thermore, mild and severe water deficit treatments reduced
total plant DW by 23 and 37%, respectively. To account for
the effects of water deficit treatments on ginseng root
growth, root growth parameters were measured immediately
before transplanting (beginning) and after harvesting (at the
end of the water deficit experiment) (Table 2). Total root
length in the control increased by only 45% over its original
length, whereas the mild and severe water deficit treated
ginseng root length increased by 55 and 67% over their
Jinwook Lee and Kenneth W. Mudge478
Table 1. Plant growth characteristics of 4-year-old American ginseng as affected by water deficit.
Table 2. Changes in root growth parameters of 4-year-old American ginseng as affected by water deficit before transplanting and after
harvesting.
original length, respectively. On the other hand, the increase
in root diameter and root FW was significantly lower in the
mild and severe water deficit treatments than in the control.
American ginseng plants were well watered for the first
45 days immediately after transplanting followed by irrigation
intervals of 4, 10 and 20 days for the control, mild, and severe
water deficit treatments, respectively. Soil water potential
( Soil) remained at approximately 0.4 MPa in the well watered
control (4 days irrigation interval) throughout the 20 days
period shown in Fig. 1B. For the first 10 days, Soil for both
the mild (10 days irrigation interval) and severe (20 days
irrigation interval) water deficit treatments declined to
approximately 1.4 MPa. After rewatering at 10 day, Soil of
the mild water deficit treatment recovered to the same level
as the control, while that of the severe water deficit treatment
continued to decline for the next 10 days to approximately
3.6 MPa. To assess leaf moisture status, predawn leaf water
potential ( Leaf) was measured using a pressure bomb from
day 20 through day 40 after initiation of water deficit. From
an initial Leaf of approximately 0.15 MPa, the well watered
control irrigated on a 4 days interval remained at about that
level (Fig. 1A). The Leaf of the mildly water deficit treated
plants irrigated on a 10 days interval decreased to approxi-
mately 0.3 MPa after 10 days, but recovered nearly to the
level of the control after rewatering, and declined similarly
over the next 10 days. The Leaf of the severely water deficit
treated plants declined continuously during the next 20 days
without irrigation to approximately 0.5 MPa.
Daily evapotranspiration was not different for the first 4
days, but as water deficit progressed, three distinctive response
patterns were appeared depending on the corresponding
irrigation intervals (Fig. 2A). Daily evapotranspiration in well
watered control treatment was fluctuated daily. In mild water
deficit treatment, daily evapotranspiration gradually declined
during the first 6 days, but significantly decreased after the
6th day. In severe water deficit treatment, daily evapotran-
spiration showed the same trend as mild water deficit
treatment from day 0 through day 10 of the water deficit
experiment, but 10 days after irrigation, daily evapotran-
spiration rate in severe water deficit treatment remained
lower than 10 g·d-1
. Cumulative evapotranspiration did not
differ among water deficit treatments from day 0 through
day 10 of the water deficit experiment (Fig. 2B). After day
10, the increase rate of cumulative evapotranspiration for
the severe water deficit treatment declined considerably
while that of the control and mild water deficit treatments
continued to increase.
To evaluate the photosynthetic activity of water deficit
treated American ginseng leaves, stomatal conductance (gS)

Hort. Environ. Biotechnol. 54(6):475-483. 2013. 479
A
B
Fig. 1. Leaf water potential ( Leaf, A) of American ginseng and soil
water potential ( Soil, B) in response to water deficit treatments
by irrigating every 4 (control), 10 (mild stress), and 20 (severe
stress) days. Measurements were taken from day 20 through
day 40 of the water deficit experiment corresponding to the 6th
to 10th dry down cycle for the control, 3rd to 4th dry down cycle
for the mild water deficit treatment, and 2nd dry down cycle for
the severe water deficit treatment. Vertical bars represent
standard errors of the means (n = 5 and 24, for Leaf, and Soil,
respectively). Error bars representing standard errors are shown
only if greater than the size of the data point symbol itself.
A
B
Fig. 2. Daily (A) and cumulative (B) evapotranspiration of American
ginseng in response to water deficit treatments by irrigating every
4 (control), 10 (mild stress), and 20 (severe stress) day interval.
Measurements were taken from day 20 through day 40 of the
water deficit experiment corresponding to the 6th to10th dry down
cycle for the control, 3rd to 4th dry down cycle for the mild water
deficit treatment, and 2nd dry down cycle for the severe water
deficit treatment, respectively. Vertical bars represent standard
errors of the means (n = 24). Error bars representing standard
errors are shown only if greater than the size of the data point
symbol itself. Nonlinear regression equations for cumulative
evapotranspiration (B) are y = 6.8869 + 35.4486x - 0.3098x2
(r2
= 0.9983, P < 0.0001) for the control (solid line), y = 15.7465
+ 32.0542x - 0.2789x2 (r
2= 0.9945, P < 0.0001) for the mild
water deficit treatment (short dash line), and y = -10.7949 +
53.2929x - 2.6419x2 + 0.0473x
3 (r
2= 0.9977, P < 0.0001) for
the severe water deficit treatment (dotted line).
and CO2 assimilation were measured at the end of water
deficit experiment. The gS and CO2 assimilation decreased
as water deficit progressed (Table 3). In severe water deficit
treatment, gS and CO2 assimilation was 5.6- and 3.1-fold
lower as compared with those of controlled plants, respect-
ively. In addition, leaf chlorophyll b and total chlorophyll
content (Chl a + b), which influence photosynthetic rate,
were significantly reduced by water deficit treatments, but
chlorophyll a content was not statistically different (Table
3). The ABA concentration increased with the progression
of water deficit (Table 3), and ABA response was inversely
responded to stomatal conductance and CO2 assimilation.
Individual ginsenosides Re, Rb1, Rc, and Rd, and total
ginsenoside contents increased with the progression of water
deficit (Table 4). However, individual ginsenosides Rg1 and
Rb2 were not affected by water deficit treatments. In addition,
individual ginsenosides Re and Rb1 made up more than
60% of total ginsenoside content. The content of total
ginsenosides, which are sum of six individual ginsenosides,
was increased in the severely water deficit treatment (1.9%)
as compared with the control (1.6%). Furthermore, individual
ginsenosides Rb1, Rc, and Rb2 were statistically influenced
by the covariate, the rootlet fresh weight before transplanting
(RFWBT). However, total ginsenoside was not affected by
the RFWBT, covariate, but only by water deficit treatments.
There was no statistical interaction effect on water deficit

Jinwook Lee and Kenneth W. Mudge480
Table 3. Stomatal conductance (gs), CO2 assimilation, contents of chlorophyll a, b and a + b, and ABA concentration of 4-year-old
American ginseng leaves as affected by water deficit.
Table 4. Contents of individual ginsenoside and total ginsenoside of 4-year-old American ginseng roots as affected by water deficit.
treatment and the covariate (RFWBT).
Discussion
Water deficit, one of the major abiotic stresses, can con-
tribute to changing the fundamental metabolism of higher
plants to cope with adverse environment. In a higher plant,
the secondary metabolism is up-regulated by water deficit
rather than the primary metabolism for the plant growth and
development, and/or maintenance. Therefore, the reduction
of plant growth and development, and horticultural and
agricultural crops yield caused by water deficit has been
well documented. Water deficit effects on American ginseng
have not previously been studied in detail. However, Lim et
al. (2006) reported that the moderate water stress did not
affect ginseng growth, but affected the compositions and
contents of certain individual root ginsenosides. In the
present study, a significant reduction of root growth in
response to reduced soil moisture was observed (Tables 1
and 2). This finding is partially consistent with that of Mork
et al. (1981) and Li and Berard (1998), who reported that
root growth and yield decreased with soil moisture levels in
Asian and American ginsengs, respectively. On the other
hand, they also presented that shoot growth was suppressed
by soil moisture content. In this study, however, shoot
growth in terms of shoot FW and DW was not significantly
affected by water deficit treatments, although leaf area was
statistically reduced (Table 1). This response can be explained
by the determinant growth and development pattern of
American ginseng plants, since the shoot is fully developed
and enlarged to full height and leaf size within several
weeks after emergence (Lim et al., 2006; Proctor and Bailey,
1987). In the present study and that of Lim et al. (2006),
water deficit treatment had no effect on shoot growth
because water deficit treatments were not imposed until 45
and 28 days after transplanting, respectively, after shoot
growth was completed. In addition, while shoot biomass
was not significantly affected by the elevated temperatures,
root biomass in three-year-old American ginseng plants
cultivated in a greenhouse was strongly reduced (Jochum et
al., 2007). On the other hand, Lee et al. (1982) and Li and
Berard (1998) imposed water deficit treatment immediately
after transplanting, before the completion of shoot emergence
and shoot elongation. Since root enlargement occurs after
the completion of shoot enlargement, until the end of the
growing season, it is reasonable that the water deficit

Hort. Environ. Biotechnol. 54(6):475-483. 2013. 481
treatment in this experiment should inhibit root growth but
not shoot growth. Therefore, during the stage of full devel-
opment and enlargement of shoot system after shoot emer-
gence, irrigation management for preventing water deficit
should play a significant role in the further growth and
development of ginseng storage root system for the better
quality and higher root yield. In addition, ginseng root
diameter and FW decreased with progressive water deficit,
while total root length, representing the sum of the storage
root length and the longest fibrous root length, increased
with increasing water deficit (Table 2). This result is in an
agreement with the results of Lee et al. (1982) and Li and
Berard (1998). Increased root elongation, rather than diameter
or fresh weight, in response to drought stress suggests that
more assimilates are directed toward root elongation, facilitating
uptake of soil moisture.
In the case of the severe water deficit treatment in this
study (20 days irrigation interval), the rate of increase of
cumulative evapotranspiration declined dramatically. The
transpiration of cassava under progressive water deficit was
dramatically reduced at the 3rd day after imposing water
deficit (Alves and Setter, 2000). With the decline of evap-
otranspiration, Leaf under severe water deficit treatment
gradually declined (Fig. 1A). The Leaf was approximately
0.3 MPa more negative under severe water deficit treatment
than in the control. This result is similar to the Leaf in
cassava affected by water stress (El-Sharkawy et al., 1992).
However, Leaf in the present study was significantly greater
(less negative) than that of previous study with American
ginseng (Lim et al., 2006). The longer water deficit progressed,
the more Leaf decreased. This is consistent with the behavior
of cassava as reported by El-Sharkawy et al. (1992).
The reduction of available internal moisture (decreased
Leaf) within ginseng plant in this study is consistent with
declining stomata conductance, CO2 assimilation and chloro-
phyll content (Table 3), and increasing ABA levels (Table
3), consequently resulting in decreased root fresh weight
and diameter (Table 1). This result is consistent with the
finding of Lee et al. (1982) that net photosynthesis rates in
Asian ginseng decreased with soil water content. The photo-
synthetic activity and stomatal conductance of sweet potato,
which has a storage root system like ginseng, was similarly
reduced in response to water deficit (Haimeirong and Kubota,
2003). Photosynthesis and stomatal conductance in sweet
potato were substantially greater than those in ginseng leaves,
but this is not unexpected since ginseng plants are well
adapted to low light environments under the dense shaded
canopy of a deciduous hardwood forest (Proctor and Bailey,
1987). As in soybean (Inamullah and Isoda, 2005), chlorophyll
content in this study was significantly decreased in response
to water deficit, indicating that the observed decline in CO2
assimilation might be related to the degradation of chlorophyll.
Furthermore, the elevated temperatures reduced light-saturated
net photosynthetic rates and stomatal conductance in three-
year-old American ginseng plants grown in a greenhouse
(Jochum et al., 2007). Total chlorophyll content was signifi-
cantly reduced by water deficit treatment as compared with
control. Although chlorophyll a was not statistically different
between the control and water deficit treatments, water
deficit treatment tended to reduce the level of chlorophyll a.
Thus, the reduction of total chlorophyll content was not only
come from the decline of chlorophyll b content, but also
partially derived from the decrease in chlorophyll a content.
Typically, water stress contributes to the reduction of total
chlorophyll, and chlorophyll a and b contents (Zhang et al.,
2011). It is understood that less contribution of chlorophyll
a content on total chlorophyll content might be originated
from physiological and ecological characteristics of American
ginseng as a shade loving plant (Proctor and Bailey, 1987).
Water deficit treatment likely reduced the chlorophyll a
content, but the statistical difference was not appeared. This
response might be resulted from the huge variation of
chlorophyll a content to water deficit in the individual plants.
Although the increase of ABA level in response to water
deficit has been well documented in many horticultural
crops (Alves and Setter, 2000; Bauerle et al., 2004, 2006;
Setter et al., 2001; Wang et al., 2004), ABA levels have not
been reported for American ginseng leaves. In this present
work, foliar ABA levels were 3.5 times higher than the
control by the end of severe water deficit (Table 3). The
increase in foliar ABA induced by water deficit has been
shown to reduce leaf area (Trejo et al., 1995; Zhang and
Davies, 1990) as reported in this study. In cassava which
has a storage root system similar to ginseng, a 5- to 7-fold
increase in ABA concentration was reported (Alves and
Setter, 2000). Although the pattern of ABA accumulation in
cassava and ginseng was similar, the absolute concentration
of foliar ABA was substantially higher in cassava.
Up-regulation of ginsenosides reported by Lim et al.
(2006) and this study appeared to be consistent with other
reports of up-regulation of secondary metabolites in response
to abiotic environmental stress as has been reported by
Bouchereau et al. (1996), Kirakosyan et al. (2003) and Liu
(2000). Environmental effects on the composition and accu-
mulation of root ginsenosides have been reported in numerous
papers (Fournier et al., 2003; Konsler et al., 1990; Li and
Mazza, 1999). Gypsum incorporation enhanced the contents
of individual and total ginsenosides with the increase in
gypsum level while plant growth decreased (Lee and Mudge,
2013). In addition, Lim et al. (2005) reported that the com-
position and contents of root ginsenosides are significantly
affected not only by environment (garden location) but also
by genotype (population). Jochum et al. (2007) reported that
during the growing season, the elevated temperatures con-

Jinwook Lee and Kenneth W. Mudge482
tributed to increasing concentrations of root ginsenosides
Rb1, Rc, and Re. Furthermore, in this study, water deficit
treatment enhanced the level of root ginsenosides Re, Rb1,
Rc, and Rd. Therefore, abiotic environmental stresses such
as elevated temperature and water deficit could affect not
only plant growth and development but also the response of
ginsenosides. On the other hand, the level of root ginsenoside
Rg1 and Rb2 was neither affected by water deficit treatment
nor by the elevated temperature (Jochum et al, 2007). It is
considered that these root ginsenosides as minor ginsenosides
might be relatively less influenced by abiotic environmental
perturbations.
In conclusion, severe water deficit reduced root growth
(fresh weight and diameter), but up-regulated certain individual
and total ginsenosides. Water deficit also induced decreased
Leaf, increased foliar ABA concentration, and finally reduced
CO2 assimilation rate. Consequently, American ginseng root
growth and yield were significantly reduced by water deficit.
The results of this study contribute to understanding the
physiological responses of American ginseng to water deficit
and its impact on ginsenoside accumulation. From the growers
stand point of maximizing root growth, a suitable irrigation
management program may apply to intensive ginseng production
system for the better quality and higher ginseng root pro-
duction. Commercially, use of irrigation may be justified
since the growers income from the sale of ginseng is based
on root weight, but this may be achieved at the expense of
ginsenoside content.
Acknowledgements: The authors thank Joe Lardner and
Dr. Wansang Lim (Department of Horticulture, Cornell
University) for their helpful assistance and Dr. Tim L. Setter
(Department of Crop and Soil Sciences, Cornell University)
for his technical assistance of the ABA measurement. Jinwook
Lee was supported by a graduate research assistantship in
the Department of Horticulture, Cornell University.
Literature Cited
Alves, A.A.C. and T.L. Setter. 2000. Response of cassava to water
deficit; leaf area growth and abscisic acid. Crop Sci. 40:131-137.
Arnon, D.I. 1949. Copper enzymes in isolated chloroplasts. Poly-
phenoloxidase in Beta vulgaris. Plant Physiol. 24:1-15.
Assinewe, V., B.R. Baum, D. Gagnon, and J.T. Arnason. 2003.
Phytochemistry of wild populations of Panax quinquefolius L.
(North American ginseng). J. Agric. Food Chem. 51:4549-4553.
Attele, A.S., J.A. Wua, and C. S. Yuan. 1999. Ginseng pharmacology:
Multiple constituents and multiple actions. Biochem. Pharmacol.
58:1685-1693.
Barbara, K., K. Ewa, K. Jerzy, and C. Aleksander. 2006. The effect
of growth regulators on quality parameters and ginsenosides
accumulation in Panax quinquefolium L. roots. Plant Growth
Regulat. 48:13-19.
Bauerle, W.L., W.W. Inman, and J.B. Dudley. 2006. Leaf abscisic
acid accumulation in response to substrate water content: Linking
leaf gas exchange regulation with leaf abscisic acid concentration.
J. Amer. Soc. Hort. Sci. 131:295-301.
Bauerle, W.L., T.H. Whitlow, T.L. Setter, and F.M. Vermeylen. 2004.
Abscisic acid synthesis in Acer rubrum L. leaves - A vapor-
pressure-deficit-mediated response. J. Amer. Soc. Hort. Sci.
129:182-187.
Beyfuss, R.L. 1999. American ginseng production in woodlots.
Agroforest. Notes 14:1-4.
Bouchereau, A., N. Clossais-Besnard, A. Bensaoud, L. Leport, and
M. Renard. 1996. Water stress effects on rapeseed quality. Eur.
J. Agron. 5:19-30.
Clemente, H.S. and T.E. Marler. 1996. Drought stress influences
gas-exchange responses of papaya leaves to rapid changes in
irradiance. J. Amer. Soc. Hort. Sci. 121:292-295.
Delfine, S., R. Tognetti, F. Loreto, and A. Alvino. 2002. Physiological
and growth responses to water stress in field-grown bell pepper
(Capsicum annuum L.). J. Hort. Sci. Biotechnol. 77:697-704.
El-Sharkawy, M.A., A.H. Del Pilar, and C. Hershey. 1992. Yield
stability of cassava during prolonged mid-season water stress. Exp.
Agric. 28:165-174.
Ennahli, S. and H.J. Earl. 2005. Physiological limitations to photo-
synthetic carbon assimilation in cotton under water stress. Crop
Sci. 45:2374-2382.
Fournier, A.R., J.T.A. Proctor, L. Gauthier, S. Khanizadeh, A. Belanger,
A. Gosselin, and M. Dorais. 2003. Understory light and root
ginsenosides in forest-grown Panax quinquefolius. Phytochemistry
63:777-782.
Jochum, G.M., K.W. Mudge, and R.B. Thomas. 2007. Elevated tem-
peratures increase leaf senescence and root secondary metabolite
concentrations in the understory herb Panax quinquefolius (Araliaceae).
Amer. J. Bot. 94:819-826.
Kirakosyan, A., E. Seymour, P.B. Kaufman, S. Warber, S. Bolling,
and S.C. Chang. 2003. Antioxidant capacity of polyphenolic extracts
from leaves of Crataegus laevigata and Crataegus monogyna
(Hawthorn) subjected to drought and cold stress. J. Agric. Food Chem.
51:3973-3976.
Konsler, T.R., S.W. Zito, J.E. Shelton, and E.J. Staba. 1990. Lime
and phosphorus effects on American ginseng. II. Root and leaf
ginsenoside content and their relationship. J. Amer. Soc. Hort. Sci.
115:575-580.
Lee, J. and K.W. Mudge. 2013. Gypsum effects on plant growth,
nutrients, ginsenosides, and their relationship in American ginseng.
Hort. Environ. Biotechnol. 54:228-235.
Lee, S.S., D.C. Yang, and Y.T. Kim. 1982. Effects of soil water
regimes on photosynthesis, growth and development of ginseng
plant (Panax ginseng C.A. Meyer). Kor. J. Crop Sci. 27:175-181.
Li, T.S.C. 1995. Asian and American ginseng - A review. Hort-
Technology 5:27-34.
Li, T.S.C. and R.G. Berard. 1998. Effects of soil moisture on the
growth of American ginseng (Panax quinquefolium L.). J. Ginseng
Res. 22:122-125.
Li, T.S.C. and G. Mazza. 1999. Correlations between leaf and soil
mineral concentrations and ginsenoside contents in American
ginseng. HortScience 34:85-87.
Lim, W., K.W. Mudge, and J.W. Lee. 2006. Effect of water stress
on ginsenoside production and growth of American ginseng.
HortTechnology 16:517-522.
Lim, W., K.W. Mudge, and F. Vermeylen. 2005. Effects of population,
age, and cultivation methods on ginsenoside content of American
ginseng (Panax quinquefolium). J. Agric. Food Chem. 53:8498-8505.
Liu, Z. 2000. Drought-induced in vivo synthesis of camptothecin in
Camptotheca acuminata seedlings. Physiol. Plant. 110:483-488.
Mork, S.K., S.Y. Son, and H. Park. 1981. Root and top growth of
Panax ginseng at various soil moisture regime. Kor. J. Crop Sci.
26:115-120.

Hort. Environ. Biotechnol. 54(6):475-483. 2013. 483
Proctor, J.T.A. and W.G. Bailey. 1987. Ginseng: Industry, botany,
and culture. Hort. Rev. 9:187-236.
Reaves, M.E., 2003. Gas exchange and water relations of red maple
Acer rubrum L. ecotypes and cultivars in response to drought.
Cornell University, Ithaca, NY.
Setter, T.L., B.A. Flannigan, and J. Melkonian. 2001. Loss of kernel
set due to water deficit and shade in maize: Carbohydrate supplies,
abscisic acid, and cytokinins. Crop Sci. 41:1530-1540.
Trejo, C.L., A.L. Clephan, and W.J. Davies. 1995. How do stomata
read abscisic acid signals? Plant Physiol. 109:803-811.
Wang, Z., B. Huang, S.A. Bonos, and W.A. Meyer. 2004. Abscisic
acid accumulation in relation to drought tolerance in Kentucky
bluegrass. HortScience 39:1133-1137.
Zhang, J. and W.J. Davies. 1990. Changes in the concentration of
ABA in xylem sap as a function of changing soil water status
can account for changes in leaf conductance and growth. Plant
Cell Environ. 13:277-285.
Zhang, Y.J., Z.K. Xie, Y.J. Wang, P.X. Su, L.P. An, and H. Gao.
2011. Effect of water stress on leaf photosynthesis, chlorophyll content,
and growth of oriental lily. Russian J. Plant Physiol. 58:844-850.