vegetation responses to mid-holocene extreme drought events and subsequent long-term drought on the...
TRANSCRIPT

Vl
YC
a
ARRA
KDVFSST
1
(tu2dtdhhdtoovttm
0h
Agricultural and Forest Meteorology 178–179 (2013) 3–9
Contents lists available at SciVerse ScienceDirect
Agricultural and Forest Meteorology
journa l homepage: www.e lsev ier .com/ locate /agr formet
egetation responses to mid-Holocene extreme drought events and subsequentong-term drought on the southeastern Inner Mongolian Plateau, China
i Yin, Hongyan Liu ∗, Guo Liu, Qian Hao, Hongya Wangollege of Urban and Environmental Sciences and MOE Laboratory for Earth Surface Processes, Peking University, Beijing 100871, China
r t i c l e i n f o
rticle history:eceived 10 May 2012eceived in revised form 12 October 2012ccepted 15 October 2012
eywords:roughtegetation responsesorest resilienceemi-arid regionoil coarsening
a b s t r a c t
Ecological responses to climate change are strongly regulated by long-term processes, such as changesin species composition and carbon dynamics in soil; therefore, understanding and predicting these pro-cesses require long time scale studies. Based on high-resolution lake sediment records of the Angili NuurLake from the forest-steppe ecotone at the southeastern margin of the Asian Gobi, we found three pro-nounced drought events during the mid-Holocene (∼7.0–6.3 ka BP, BP means “before present”, wherepresent means 1950 and ka = 1000 years) and a subsequent graduate drying trend. During the extremedrought events, lake aquatic system showed strong desiccation and instant salty marsh expansion;whereas regional forest showed strong resilience. During the following long-term drying at millennialscale, broadleaf forest, coniferous forest and steppe successively dominated in this region, in 6.3–4.4 kaBP, 4.0–2.7 ka BP and since 2.7 ka BP, respectively; but the changing dominance between forest and steppe
he Holocene did not match the climate change rate as reconstructed in previous studies, suggesting a lagged and non-proportional change of vegetation type to the range of climate change. The resilience of forest impliesa strong buffering effect of biotic and abiotic factors other than climate, probably explained by diversemicro-environments of this mountainous region that allowed forest to survive during the drought spells,and also by gradual soil coarsening that retarded soil field capacity change and nutrient loss. However,
g tre
under accumulative dryin. Introduction
In the context of recent global warming and drought intensifyingSolomon et al., 2008; Dai, 2011), the understanding of terres-rial ecosystem-climate interactions under rapid climate change isndoubtedly crucial to both scientific research and public (IPCC,007). Drought-related ecological degradation, including forestieback (Breshears et al., 2005; Allen et al., 2010), grassland deser-ification (Schlesinger et al., 1990; Yang et al., 2005), wetlandegradation and lake desiccation (Zhao et al., 2011; Lake, 2011)ave been widely reported, especially in the semi-arid regions;owever, these studies have mostly been restricted to annual toecadal scales, which might fail to cover the climate change rangehat current global change is likely to bring (Held, 2012). More-ver, the relative long lifespan of trees and long-term processesf forest regeneration and succession made it difficult to discussegetation responses to accumulative climate change on this rela-
ively short time scale (Peng et al., 2011). Therefore, it is importanto use palaeo-ecological records, where past droughts have beenore severe and persistent than those of instrument-recorded his-
∗ Corresponding author.E-mail address: [email protected] (H. Liu).
168-1923/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.agrformet.2012.10.005
nd, forest was in general replaced by steppe around 2.0 ka BP.© 2012 Elsevier B.V. All rights reserved.
tory (Laird et al., 1996; Clark et al., 2002; Chen et al., 2003a, b; Cooket al., 2010; Nelson et al., 2011), to study vegetation responses toclimate change over a longer time scale.
Forest and steppe exhibit mosaic distribution at the southernmargin of the Asian Gobi desert and surrounding steppe region(Fig. 1), where lies the drought limit of temperate forest distribu-tion range and water availability greatly controls vegetation growth(Walter and Breckle, 2002; Wu, 1980). Meanwhile, it is also themargin of Pacific monsoon influence, with highly fluctuated inter-annual precipitation associated with changing intensity of Pacificmonsoon (Clift and Plumb, 2008). Vegetation dynamics in thisregion is hence sensitive to climate change, especially drought (Piaoet al., 2003); but how vegetation type will respond to long-termclimate change remains unclear due to complicated interactionsamong vegetation, climate and soil, including processes in carbonand nitrogen cycling, albedo and snow cover change, dust emis-sion and transportation, as well as aerosol concentration change(Cox et al., 2000; Ridgwell and Watson, 2002; Rotenberg and Yakir,2010).
A number of studies have reported the middle Holocene drought
spells in these region (Chen et al., 2003a, b; Xiao et al., 2006, 2009;Chen et al., 2003a, b, 2008a), as well as soil development and sandaccumulation cycles (Lu et al., 2005) and dust flux changes (Miaoet al., 2007; An et al., 2011) during the Holocene. These drought
4 Y. Yin et al. / Agricultural and Forest M
Fig. 1. Study area with sampling sites on a biome map with hills shaded; yellowcircle represents sediment core site, Anguli Nuur; orange dots represent topsoil sam-ples, including cited sites in the compiled database; purple dots represent pairedlca
scHsdwetaobebm
2
2
∼(slsocusTa
dfttGav
cwmossi
ake surface sediment-surrounding topsoil sample sites. The inset in the upper leftorner illustrates modern MAT (mean annual temperature, color) and MAP (meannnual precipitation, isolines).
pells provided us an opportunity to study a severe arid period andorresponding vegetation responses on a millennium time scale.owever, these studies mainly worked on environment recon-
truction, while vegetation responses to climate change are seldomiscussed from the ecological point of view. In this study, based onell dated high-resolution lake sediment records, we found several
xtreme drought events in the mid-Holocene and subsequent long-erm drought trend in the mid to late Holocene. Thus, we aimedt exploring following questions: (1) is this mid-Holocene droughtnly site-specific or of regional importance and what was the possi-le reason? (2) how vegetation responded to rapid extreme droughtvents and long-term drying trend, respectively? and (3) how otheriotic or abiotic factors have influenced vegetation response to theid-Holocene droughts?
. Materials and methods
.1. Data preparation and field sampling
Sediment cores retrieved from Anguli Nuur Lake (∼41◦20′N,114◦20′E; altitude, 1315 m.a.s.l.; mean annual precipitation
MAP), 380 mm; mean annual temperature (MAT), 3.1 ◦C) wereelected for analysis in this study (Fig. 1, yellow dot). This inlandake lies in the woodland-steppe ecotone of north China, at theouthern border of the Asian Gobi, where is also the marginal areaf the Pacific monsoon influence (Wu, 1980). The 1138 cm An-Sore was drilled near the lowest position of the desiccated lake bedsing a HZ-100Y drilling rig in 2005, and a second 1023 cm parallelupplementary core (An-A) was drilled in 2007 (Liu et al., 2010).he sediment cores were divided into 1 cm subsamples for furthernalysis.
Topsoil samples from 174 sites across the vegetation type gra-ient (desert, desert steppe, typical steppe, meadow steppe andorest) were collected for pollen analysis; in addition, a regionalopsoil pollen database was established to expand the environmen-al range of topsoil pollen from Liu et al. (1999), Xu et al. (2007) andunin et al. (1999). In total, 461 topsoil pollen samples were used tonalyze the relationship between surface pollen composition andegetation (Fig. 1, orange dots).
Lake surface sediments from 45 natural lakes (Fig. 1, purple dots)overing a large annual precipitation range (from 100 to 800 mm)ere sampled over water by a sediment sampler to grab the upper-ost 5 cm; meanwhile, corresponding topsoil (uppermost 5 cm)
f dominant vegetation types in the lake shore region were alsoampled to validate the usage of topsoil pollen-vegetation relation-hips in sediments and to analyze the indicative chemical elementsn the sediments. MAP and MAT of these 45 sample sites were
eteorology 178–179 (2013) 3–9
interpolated from those of meteorological stations in this regionprovided by the Chinese National Meteorological Center. Ariditywas calculated as the inverse of the Thornthwaite humidity index(Thornthwaite, 1948).
2.2. Laboratory analysis
Next, sediment core samples (at 5 cm intervals) and all top-soil and lake surface samples were analyzed in the laboratory. Thechronology of the sediment core was defined by integrated resultsof AMS 14C dating and 210Pb and 137Cs activity analysis; all datesin this study were calibrated to calendar years (Fig. 3, left column,Age-depth, see details in Yin et al., 2011). Soluble salt and mainion contents were analyzed by the ion titration method. Chemi-cal element components were analyzed by ARL ADVANT-XP X-rayFluorescence Sequential Spectrometer (XRF). Grain size distribu-tions were determined by Malvern Mastersizer 2000 after standardpretreatment (Yin et al., 2011). Total organic carbon (TOC) andTotal nitrogen (TN) were analyzed by colormetric detection andhigh temperature oxidation (Elementar Vario EL), respectively, afterremoval of roots and other macro organic remains and grinding topass through a 0.149 mm soil sieve. Pollen analysis, including three-step pretreatment with HCl, NaOH and coarse material sieving,an extraction procedure by centrifugal with heavy liquid solution(KI + HI + Zn) of specific gravity around 1.9, and an after-treatmentwith acetolysis and fine sieving were conducted for all sedimentcore, topsoil and lake surface samples. Approximately 300–400 ter-restrial carpophyte pollen grains were counted and total pollen sum(not including aquatic plant, fern and algae) was used for pollenpercentage calculation.
2.3. Statistical analysis
The main statistical techniques applied in this paper wereRedundancy Analysis (RDA, Anderson and Willis, 2003), used foranalyzing modern indexes–climate relationships, and ConstrainedClustering (Bennett, 1996) for classifying profile pollen composi-tion. These analyses were performed in R using the vegan 2.0 andrioja 0.7-3 packages.
3. Results
3.1. Indication of pollen taxon and chemical elements in thesemi-arid region
Thirty-two families or genera of pollen grains were identified intotal. RDA results for topsoil and lake surface pollen showed dif-ferent clusters of pollen composition across vegetation types andillustrated the corresponding indication of labeled families/genera(Fig. 2a). The influences of environmental variables, including meanannual precipitation (MAP), mean annual temperature (MAT) andaltitude (ALT) are statistically significant (p < 0.005), suggestingthat differences in pollen composition effectively indicate environ-mental changes. Each pollen taxon showed similar characteristicswhen conducting RDA analysis in topsoil and lake surface pollendatasets separately, suggesting that the indication of topsoil andlake sediment pollen are generally in good agreement. Therefore, itis reasonable to use a topsoil pollen–climate relationship to inter-pret lake sediment pollen composition. In general, arboreal pollenindicates a humid environment, of which Quercus and Carpinusindicate warm conditions, while Larix and Betula indicate cool con-
ditions; on the contrary, Chenopodiaceae, Ephedra and Nitraria areindicators of a dry climate.RDA results of chemical elements showed that chemical ele-ment components in both soil and lake sediment differed across
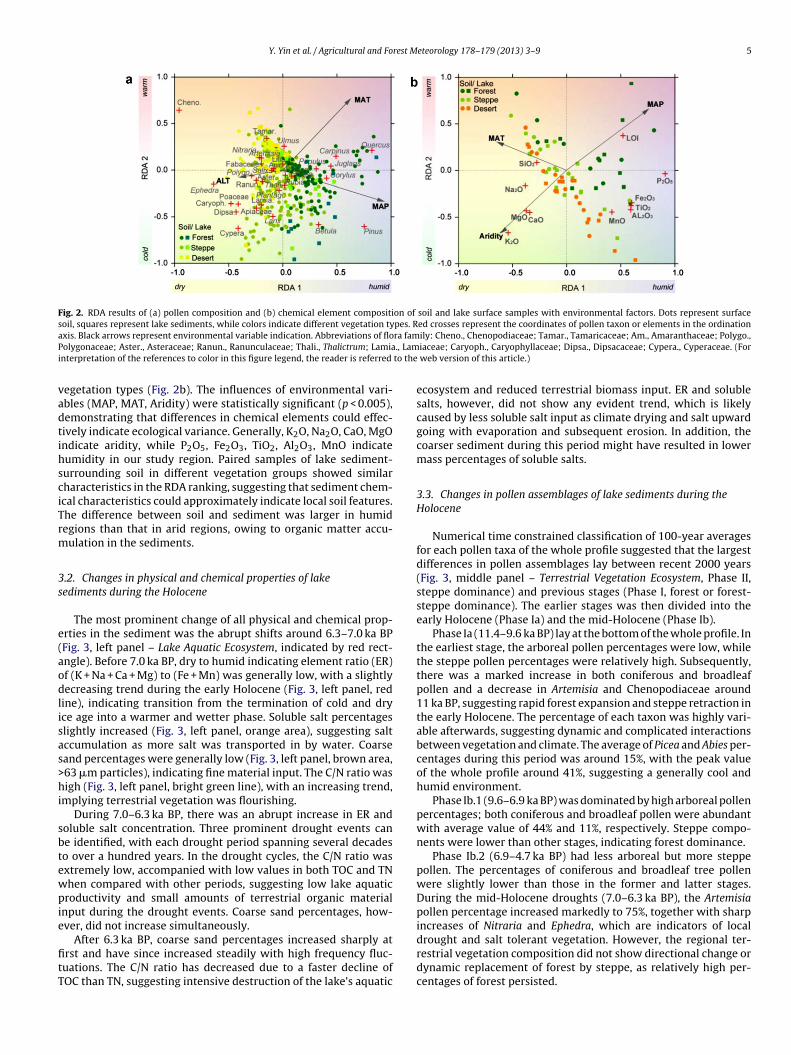
Y. Yin et al. / Agricultural and Forest Meteorology 178–179 (2013) 3–9 5
Fig. 2. RDA results of (a) pollen composition and (b) chemical element composition of soil and lake surface samples with environmental factors. Dots represent surfacesoil, squares represent lake sediments, while colors indicate different vegetation types. Red crosses represent the coordinates of pollen taxon or elements in the ordinationa ra famP ., Lami to the
vadtihsciTrm
3s
e(aodlisas>hi
sbtewpie
fitT
xis. Black arrows represent environmental variable indication. Abbreviations of floolygonaceae; Aster., Asteraceae; Ranun., Ranunculaceae; Thali., Thalictrum; Lamianterpretation of the references to color in this figure legend, the reader is referred
egetation types (Fig. 2b). The influences of environmental vari-bles (MAP, MAT, Aridity) were statistically significant (p < 0.005),emonstrating that differences in chemical elements could effec-ively indicate ecological variance. Generally, K2O, Na2O, CaO, MgOndicate aridity, while P2O5, Fe2O3, TiO2, Al2O3, MnO indicateumidity in our study region. Paired samples of lake sediment-urrounding soil in different vegetation groups showed similarharacteristics in the RDA ranking, suggesting that sediment chem-cal characteristics could approximately indicate local soil features.he difference between soil and sediment was larger in humidegions than that in arid regions, owing to organic matter accu-ulation in the sediments.
.2. Changes in physical and chemical properties of lakeediments during the Holocene
The most prominent change of all physical and chemical prop-rties in the sediment was the abrupt shifts around 6.3–7.0 ka BPFig. 3, left panel – Lake Aquatic Ecosystem, indicated by red rect-ngle). Before 7.0 ka BP, dry to humid indicating element ratio (ER)f (K + Na + Ca + Mg) to (Fe + Mn) was generally low, with a slightlyecreasing trend during the early Holocene (Fig. 3, left panel, red
ine), indicating transition from the termination of cold and dryce age into a warmer and wetter phase. Soluble salt percentageslightly increased (Fig. 3, left panel, orange area), suggesting saltccumulation as more salt was transported in by water. Coarseand percentages were generally low (Fig. 3, left panel, brown area,63 �m particles), indicating fine material input. The C/N ratio wasigh (Fig. 3, left panel, bright green line), with an increasing trend,
mplying terrestrial vegetation was flourishing.During 7.0–6.3 ka BP, there was an abrupt increase in ER and
oluble salt concentration. Three prominent drought events cane identified, with each drought period spanning several decadeso over a hundred years. In the drought cycles, the C/N ratio wasxtremely low, accompanied with low values in both TOC and TNhen compared with other periods, suggesting low lake aquaticroductivity and small amounts of terrestrial organic material
nput during the drought events. Coarse sand percentages, how-ver, did not increase simultaneously.
After 6.3 ka BP, coarse sand percentages increased sharply atrst and have since increased steadily with high frequency fluc-uations. The C/N ratio has decreased due to a faster decline ofOC than TN, suggesting intensive destruction of the lake’s aquatic
ily: Cheno., Chenopodiaceae; Tamar., Tamaricaceae; Am., Amaranthaceae; Polygo.,iaceae; Caryoph., Caryophyllaceae; Dipsa., Dipsacaceae; Cypera., Cyperaceae. (Forweb version of this article.)
ecosystem and reduced terrestrial biomass input. ER and solublesalts, however, did not show any evident trend, which is likelycaused by less soluble salt input as climate drying and salt upwardgoing with evaporation and subsequent erosion. In addition, thecoarser sediment during this period might have resulted in lowermass percentages of soluble salts.
3.3. Changes in pollen assemblages of lake sediments during theHolocene
Numerical time constrained classification of 100-year averagesfor each pollen taxa of the whole profile suggested that the largestdifferences in pollen assemblages lay between recent 2000 years(Fig. 3, middle panel – Terrestrial Vegetation Ecosystem, Phase II,steppe dominance) and previous stages (Phase I, forest or forest-steppe dominance). The earlier stages was then divided into theearly Holocene (Phase Ia) and the mid-Holocene (Phase Ib).
Phase Ia (11.4–9.6 ka BP) lay at the bottom of the whole profile. Inthe earliest stage, the arboreal pollen percentages were low, whilethe steppe pollen percentages were relatively high. Subsequently,there was a marked increase in both coniferous and broadleafpollen and a decrease in Artemisia and Chenopodiaceae around11 ka BP, suggesting rapid forest expansion and steppe retraction inthe early Holocene. The percentage of each taxon was highly vari-able afterwards, suggesting dynamic and complicated interactionsbetween vegetation and climate. The average of Picea and Abies per-centages during this period was around 15%, with the peak valueof the whole profile around 41%, suggesting a generally cool andhumid environment.
Phase Ib.1 (9.6–6.9 ka BP) was dominated by high arboreal pollenpercentages; both coniferous and broadleaf pollen were abundantwith average value of 44% and 11%, respectively. Steppe compo-nents were lower than other stages, indicating forest dominance.
Phase Ib.2 (6.9–4.7 ka BP) had less arboreal but more steppepollen. The percentages of coniferous and broadleaf tree pollenwere slightly lower than those in the former and latter stages.During the mid-Holocene droughts (7.0–6.3 ka BP), the Artemisiapollen percentage increased markedly to 75%, together with sharpincreases of Nitraria and Ephedra, which are indicators of local
drought and salt tolerant vegetation. However, the regional ter-restrial vegetation composition did not show directional change ordynamic replacement of forest by steppe, as relatively high per-centages of forest persisted.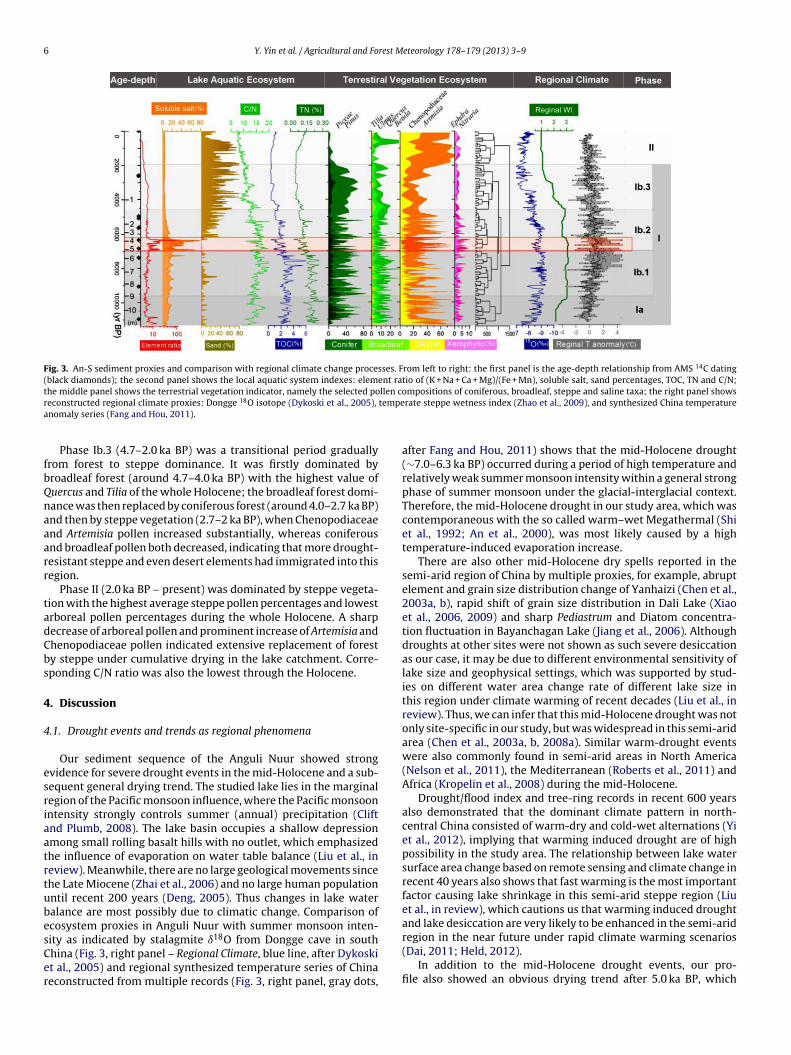
6 Y. Yin et al. / Agricultural and Forest Meteorology 178–179 (2013) 3–9
Fig. 3. An-S sediment proxies and comparison with regional climate change processes. From left to right: the first panel is the age-depth relationship from AMS 14C dating(black diamonds); the second panel shows the local aquatic system indexes: element ratio of (K + Na + Ca + Mg)/(Fe + Mn), soluble salt, sand percentages, TOC, TN and C/N;t llen cr tempea
fbQnaaarr
tadCbs
4
4
esriaatrtubesCer
he middle panel shows the terrestrial vegetation indicator, namely the selected poeconstructed regional climate proxies: Dongge 18O isotope (Dykoski et al., 2005),nomaly series (Fang and Hou, 2011).
Phase Ib.3 (4.7–2.0 ka BP) was a transitional period graduallyrom forest to steppe dominance. It was firstly dominated byroadleaf forest (around 4.7–4.0 ka BP) with the highest value ofuercus and Tilia of the whole Holocene; the broadleaf forest domi-ance was then replaced by coniferous forest (around 4.0–2.7 ka BP)nd then by steppe vegetation (2.7–2 ka BP), when Chenopodiaceaend Artemisia pollen increased substantially, whereas coniferousnd broadleaf pollen both decreased, indicating that more drought-esistant steppe and even desert elements had immigrated into thisegion.
Phase II (2.0 ka BP – present) was dominated by steppe vegeta-ion with the highest average steppe pollen percentages and lowestrboreal pollen percentages during the whole Holocene. A sharpecrease of arboreal pollen and prominent increase of Artemisia andhenopodiaceae pollen indicated extensive replacement of foresty steppe under cumulative drying in the lake catchment. Corre-ponding C/N ratio was also the lowest through the Holocene.
. Discussion
.1. Drought events and trends as regional phenomena
Our sediment sequence of the Anguli Nuur showed strongvidence for severe drought events in the mid-Holocene and a sub-equent general drying trend. The studied lake lies in the marginalegion of the Pacific monsoon influence, where the Pacific monsoonntensity strongly controls summer (annual) precipitation (Cliftnd Plumb, 2008). The lake basin occupies a shallow depressionmong small rolling basalt hills with no outlet, which emphasizedhe influence of evaporation on water table balance (Liu et al., ineview). Meanwhile, there are no large geological movements sincehe Late Miocene (Zhai et al., 2006) and no large human populationntil recent 200 years (Deng, 2005). Thus changes in lake wateralance are most possibly due to climatic change. Comparison ofcosystem proxies in Anguli Nuur with summer monsoon inten-
ity as indicated by stalagmite ı18O from Dongge cave in southhina (Fig. 3, right panel – Regional Climate, blue line, after Dykoskit al., 2005) and regional synthesized temperature series of Chinaeconstructed from multiple records (Fig. 3, right panel, gray dots,ompositions of coniferous, broadleaf, steppe and saline taxa; the right panel showsrate steppe wetness index (Zhao et al., 2009), and synthesized China temperature
after Fang and Hou, 2011) shows that the mid-Holocene drought(∼7.0–6.3 ka BP) occurred during a period of high temperature andrelatively weak summer monsoon intensity within a general strongphase of summer monsoon under the glacial-interglacial context.Therefore, the mid-Holocene drought in our study area, which wascontemporaneous with the so called warm–wet Megathermal (Shiet al., 1992; An et al., 2000), was most likely caused by a hightemperature-induced evaporation increase.
There are also other mid-Holocene dry spells reported in thesemi-arid region of China by multiple proxies, for example, abruptelement and grain size distribution change of Yanhaizi (Chen et al.,2003a, b), rapid shift of grain size distribution in Dali Lake (Xiaoet al., 2006, 2009) and sharp Pediastrum and Diatom concentra-tion fluctuation in Bayanchagan Lake (Jiang et al., 2006). Althoughdroughts at other sites were not shown as such severe desiccationas our case, it may be due to different environmental sensitivity oflake size and geophysical settings, which was supported by stud-ies on different water area change rate of different lake size inthis region under climate warming of recent decades (Liu et al., inreview). Thus, we can infer that this mid-Holocene drought was notonly site-specific in our study, but was widespread in this semi-aridarea (Chen et al., 2003a, b, 2008a). Similar warm-drought eventswere also commonly found in semi-arid areas in North America(Nelson et al., 2011), the Mediterranean (Roberts et al., 2011) andAfrica (Kropelin et al., 2008) during the mid-Holocene.
Drought/flood index and tree-ring records in recent 600 yearsalso demonstrated that the dominant climate pattern in north-central China consisted of warm-dry and cold-wet alternations (Yiet al., 2012), implying that warming induced drought are of highpossibility in the study area. The relationship between lake watersurface area change based on remote sensing and climate change inrecent 40 years also shows that fast warming is the most importantfactor causing lake shrinkage in this semi-arid steppe region (Liuet al., in review), which cautions us that warming induced droughtand lake desiccation are very likely to be enhanced in the semi-arid
region in the near future under rapid climate warming scenarios(Dai, 2011; Held, 2012).In addition to the mid-Holocene drought events, our pro-file also showed an obvious drying trend after 5.0 ka BP, which

rest M
cefiasDiiaLcdo
4d
rrcttmcltwtatptic1r
orespptwiplt2avdweinIdtthce
Y. Yin et al. / Agricultural and Fo
oincided well with other profiles in this semi-arid steppe region,.g. reconstructed wetness index (WI) from multiple pollen pro-les in temperate steppe region (Fig. 3, right panel, green line)fter Zhao et al. (2009), and was possibly influenced by decreasedummer monsoon intensity as indicated by stalagmite ı18O fromongge cave in south China (Dykoski et al., 2005) and Sanbao cave
n central China (Wang et al., 2008). This widespread reductionn monsoon intensity has been widely discussed and generallyccepted as driven by decreased summer insolation (Berger andoutre, 1991; Wang et al., 2005, 2008). The stronger regional syn-hronization of this drying trend than that of the mid-Holocenerought suggests that longer-term droughts have more influencen regional climate pattern than short-term drought events.
.2. Differentiated responses of local and regional vegetation torought events
The lake aquatic system (hydrology and wetland vegetation) andegional terrestrial vegetation showed different sensitivities andesilience to the mid-Holocene drought events. Salt concentration,hemical components, TOC and TN, as indicators of lake aquatic sys-em showed instant and marked responses to droughts, whereaserrestrial vegetation, as indicated by pollen assemblages, showed
ore progressive change. The dramatic changes in lake soluble saltoncentration and sediment element components showed that theake water has been greatly condensed and that the aquatic sys-em underwent abrupt change (Fig. 3, left panel). These changesere very likely caused by evaporation greatly exceeding precipi-
ation, when considering the geological background of this regions discussed above. Meanwhile, the C/N ratio has decreased con-inuously, with faster decrease in TOC than TN, suggesting that theroductivity of the lake aquatic system and terrestrial organic mat-er input has greatly decreased (Woodward et al., 2012). Abruptncreases and subsequent sharp decreases in algae Pediastrum con-entration (7.4–6.3 kyr BP) in the Bayanchagan Lake, which is only20 km away from our study site, also support the inferred fastesponse of the lake aquatic system (Jiang et al., 2006).
In contrast to the lake system, regional vegetation indicated byur profile did not show such abrupt and directional trend of foresteplacement by steppe during the mid-Holocene extreme droughtvents. During the drought spells, arboreal pollen decreasedlightly, accompanied with an increase in the steppe speciesercentages and the local drought and saline tolerant species com-osition, Ephedra and Nitraria. When interpreting percentage data,he prominent increase of pollen taxa from relatively small patchesith low productivity and inferior transportation could strongly
ndicate local vegetation responses (Minckley et al., 2012). Thisrovides reliable evidence that, during the drought events the
ocal saline vegetation expanded promptly, which is very similaro modern drought impacts on lakeshore wetlands (Zhao et al.,011). However, after the drought events, unlike sand percent-ges and the C/N ratio, arboreal pollen percentages increased againery rapidly after the drought, showing the resilience of forest torought impacts. A similar progressive vegetation pattern changeas also found in other profiles in this region (Li et al., 2004; Jiang
t al., 2006; Chen et al., 2008a; Zhao et al., 2009). Regional wetnessndex (WI) from synthesized pollen results did not show promi-ent decline during the drought events either (Fig. 3, right panel).
n addition, sand percentages in the sediment was low during therought events but increased afterwards, implying that the terres-rial vegetation cover might have sustained relatively high during
his extreme drought event and higher vegetation coverage mightave prevented soil from erosion (Munson et al., 2011). It also indi-ates that the terrestrial vegetation change might have lagged soilrosion and aeolian activity.eteorology 178–179 (2013) 3–9 7
4.3. Forest resilience to fast climate change
Regional vegetation, mainly forest, showed strong resilience todrought disturbances in the mid-Holocene drought events, andquickly recovered in the following period, e.g. increase in broadleafpollen in late Ib.2 (6.3–4.7 ka BP) and increase in coniferous pollenin Ib.3 (4.7–2.0 ka BP), during which period precipitation was rela-tively low as summer monsoon intensity generally declined (Fig. 3).Wetness Index (WI) of temperate steppe region also declinedmarkedly since 4.7 ka BP; however, final replacement of forest bysteppe in our study area did not occur until recent 2.0 ka BP. Thislagged response of vegetation development implies that vegetationand climate condition may not always be in equilibrium (Webb,1986) and other biotic or abiotic factors also strongly influencevegetation development except for climate changes (Davis, 1986).Modern studies suggest that vegetation responses to drought aredirectly affected by biotic factors (McDowell et al., 2008; Jacksonet al., 2009; Scherrer et al., 2011), and additionally by soil moisturethat is determined not only by precipitation, but also by topog-raphy resultant water redistribution, evapotranspiration and soiltexture (Liu et al., 2008; Katul and Siqueira, 2010). In addition, winderosion is severe in this semi-arid region, particularly during dryperiods, strongly influencing vegetation growth and soil condition(Shi et al., 2004; Zhang et al., 2003). Therefore, the strong resilienceof forest to drought stress in our study area is likely due to twoprocesses: diverse micro-environment resulted from mountainoustopography and lagged slow-process soil condition change.
Firstly, considering the effect of topographical heterogeneity,the woodland-steppe ecotone occupied in the rolling mountain-ous region (Fig. 1), where topographical settings have greatlyredistributed precipitation, and also insolation intensity, evapo-transpiration, as well as wind speed, and soil condition (Walterand Breckle, 2002). Forest patches existed nowadays under MAP ofless than 400 mm on the northern steep slopes, where soil moisturecould support forest survival (He et al., 2008). Thus under droughtstress, when forest in some parts declined, there still could havebeen suitable microenvironment which allowed forest to surviveand to reproduce; accordingly, when climate ameliorated, forestwas then able to re-establish very rapidly, avoiding long-term dis-persal or even extinction with active seed bank (Chen et al., 2008b;Tian et al., 2009; Svenning et al., 2008). These diverse niches pro-vided by topographical heterogeneity permitted species to endureextreme climatic conditions, and recover when the climate becomesuitable again.
Secondly, considering the long-term soil process, sand percent-ages were low during the mid-Holocene drought events, however,increased markedly afterwards, implying that the terrestrial veg-etation cover might have been relatively high during this extremedrought event and have thus alleviated soil erosion and aeolianactivities (Munson et al., 2011). It also indicates that the slow pro-cess of soil erosion and resultant soil coarsening and nutrient lossmight have retarded vegetation decline (Liu et al., 2008), and actedas a buffering effect. In return, soil condition of the late Holocene,with coarser texture and lower nutrients as suggested by grain sizeand partly by C/N, might have further caused vegetation deteriora-tion as demonstrated by modern processes (Bakker et al., 2005; Liet al., 2009).
There has been wide debate regarding whether extant specieshave coped successfully with the extremely rapid climatic changes(Hof et al., 2011; Parducci et al., 2012), for example local tem-perature changes of up to 4 ◦C per year near the end of the lastglacial period (14.7 ka BP) in the Late Quaternary (Steffensen et al.,
2008). These successes are perhaps due to the phenotypic vari-ability of populations and their ability to survive in microclimaticpockets in a heterogeneous landscape (Botkin et al., 2007). How-ever, many central European trees failed to survive the last glacial
8 rest M
mtmwnv(cerbtcmstetpeslpild
5
lMoarartsasorpcemetHr
A
ogC
R
A
Y. Yin et al. / Agricultural and Fo
aximum (LGM) (Svenning, 2003): the flat plain of west and cen-ral Europe, lacking refuge, may be a possible reason. In contrary,
any tree species survived the LGM in Northern China; refugesere maintained across the range of forest distribution in bothorth and south during the LGM rather than that the species sur-ived only in the south and subsequently dispersed northwardsRen and Beug, 2002; Chen et al., 2008b; Tian et al., 2009). We thusame up with the hypothesis that topography and resultant micro-nvironment diversity are important for species’ survival underapid climate change by offering diverse micro-environment anduffering climate change. This hypothesis was primarily tested byhe sediment results of the Anguli Nuur Lake, where vegetation overomplicated regional topological settings was more resilient to theid-Holocene drought, and the diverse topographical settings and
oil conditions may have provided a buffering effect for vegeta-ion change and allowed species to retreat or encroach according tonvironmental condition. However, it also needs to be stressed that,hough forest may have strong resilience in mountainous areas,rominent forest dieback may also occur under cumulative droughtffect, like the case after 2.0 ka BP. Soil coarsening induced by ero-ion may in return have accelerated vegetation decline as nutrientoss and soil field capacity decrease (Li et al., 2009). Therefore, forestatches in the forest-steppe ecotone, which is at the drought lim-
ts of forest survival, together with much coarsened soil texture, isikely to be more threatened by drought events in current patchyistribution than the mid-Holocene forest dominant condition.
. Conclusions
From our study of palaeo-ecological records in the Anguli Nuurake, we found severe drought events during the mid-Holocene
egathermal and subsequent gradual drying trend. These kindsf warm-induced droughts are likely to have regional significancend are of highly possibility to be enhanced in the semi-aridegion under future rapid climate warming. Lake aquatic systemnd terrestrial forest vegetation showed different sensitivities inesponse to drought events: lake aquatic system and wetland vege-ation showed profound changes instantly, whereas regional foresthowed strong resilience. During the following long-term dryingt millennial scale, broadleaf forest, coniferous forest and steppeuccessively dominated in this region, but the final replacementf forest by steppe did not match the climate change rate aseconstructed in previous studies, suggesting a lagged and non-roportional change of vegetation type to the range of climatehange. This strong resilience of forest implied a strong bufferingffect of other biotic and abiotic factors, probably including diverseicro-environments of this mountainous region that allowed for-
st to survive during the droughts, as well as gradual soil coarseninghat retarded soil water carrying capacity change and nutrient loss.owever, under accumulative drying trend, forest was eventually
eplaced by steppe in 2.0 ka BP.
cknowledgments
This study was granted by National Natural Science Foundationf China (No. 41071124, 31021001, 41011120251). We thank Jian-ling Zhu and Ji Ren for their assistance in field work, and Davehandler for editing the text.
eferences
llen, C., Macalady, A., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M.,Kitzberger, T., Rigling, A., Breshears, D., Hogg, E., 2010. A global overview ofdrought and heat-induced tree mortality reveals emerging climate change risksfor forests. For. Ecol. Manage. 259, 660–684.
eteorology 178–179 (2013) 3–9
An, C.B., Zhao, J., Tao, S., Lv, Y., Dong, W., Li, H., Jin, M., Wang, Z., 2011. Dust variationrecorded by lacustrine sediments from arid Central Asia since ∼15 cal ka BP andits implication for atmospheric circulation. Quat. Res. 75, 566–573.
An, Z., Porter, S.C., Kutzbach, J.E., Wu, X., Wang, S., Liu, X., Li, X., Zhou, W., 2000.Asynchronous Holocene optimum of the East Asian monsoon. Quat. Sci. Rev. 19,743–762.
Anderson, M.J., Willis, T.J., 2003. Canonical analysis of principal coordinates: a usefulmethod of constrained ordination for ecology. Ecology 84, 511–525.
Bakker, M.M., Govers, G., Kosmas, C., Vanacker, C., Oost, K.V., Rounsevell, M., 2005.Soil erosion as a driver of land-use change. Agric. Ecosyst. Environ. 105, 467–481.
Bennett, K., 1996. Determination of the number of zones in a biostratigraphicsequence. New Phytol. 132, 155–170.
Berger, A., Loutre, M., 1991. Insolation values for the climate of the last 10 millionyears. Quat. Sci. Rev. 10, 297–317.
Botkin, D.B., Saxe, H., Araujo, M.B., Betts, R., Bradshaw, R.H.W., Cedhagen, T., Chesson,P., Dawson, T.P., Etterson, J.R., Faith, D.P., 2007. Forecasting the effects of globalwarming on biodiversity. Bioscience 57, 227–236.
Breshears, D., Cobb, N., Rich, P., Price, K., Allen, C., Balice, R., Romme, W., Kas-tens, J., Floyd, M., Belnap, J., 2005. Regional vegetation die-off in responseto global-change-type drought. Proc. Natl. Acad. Sci. U.S.A. 102, 15144–15148.
Chen, C., Lan, H., Lou, J., Chen, Y., 2003a. The dry Holocene Megathermal in InnerMongolia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 193, 181–200.
Chen, F., Wu, W., Holmes, J., Madsen, D., Zhu, Y., Jin, M., Oviatt, C., 2003b. A mid-Holocene drought interval as evidenced by lake desiccation in the AlashanPlateau, Inner Mongolia China. Chin. Sci. Bull. 48, 1401–1410.
Chen, F., Yu, Z., Yang, M., Ito, E., Wang, S., Madsen, D., Huang, X., Zhao, Y., Sato,T., Birks, H., 2008a. Holocene moisture evolution in arid central Asia and itsout-of-phase relationship with Asian monsoon history. Quat. Sci. Rev. 27, 351–364.
Chen, K., Abbott, R.J., Milne, R.I., Tian, X., Liu, J., 2008b. Phylogeography of Pinus tab-ulaeformis Carr. (Pinaceae), a dominant species of coniferous forest in northernChina. Mol. Ecol. 17, 4276–4288.
Clark, J.S., Grimm, E.C., Donovan, J.J., Fritz, S.C., Engstrom, D.R., Almendinger, J.E.,2002. Drought cycles and landscape responses to past aridity on prairies of thenorthern great plains, USA. Ecology 83, 595–601.
Clift, P.D., Plumb, R.A., 2008. The Asian Monsoon: Causes, History and Effects. Cam-bridge University Press, Cambridge.
Cook, E.R., Anchukaitis, K.J., Buckley, B.M., D’Arrigo, R.D., Jacoby, G.C., Wright, W.E.,2010. Asian monsoon failure and megadrought during the last millennium. Sci-ence 328, 486–489.
Cox, P.M., Betts, R.A., Jones, C.D., Spall, S.A., Totterdell, I.J., 2000. Acceleration of globalwarming due to carbon-cycle feedbacks in a coupled climate model. Nature 408,184–187.
Dai, A., 2011. Drought under global warming: a review. Clim. Change 2, 45–65.Davis, M.B., 1986. Climatic instability, time lags and community disequilibrium. In:
Diamond, J., Case, T.J. (Eds.), Community Ecology. Harper and Row Publisher.Deng, H., 2005. From Natural to Cultural Landscape: Evolution of Man–Land Rela-
tionship in the Farming-pastoral Ecotone of North China from a HistoricalGeography Perspective. Commercial Press, Beijing (in Chinese).
Dykoski, C., Edwards, R., Cheng, H., Yuan, D., Cai, Y., Zhang, M., Lin, Y., Qing, J., An, Z.,Revenaugh, J., 2005. A high-resolution, absolute-dated Holocene and deglacialAsian monsoon record from Dongge Cave, China. Earth Planet. Sci. Lett. 233,71–86.
Fang, X., Hou, G., 2011. Synthetically reconstructed Holocene temperature changein China. Sci. Geogr. Sin. 31, 385–393 (in Chinese with English abstract).
Gunin, P.D., Vostokova, E.A., Dorofeyuk, N.I., Tarasov, P.E., Black, C.C., 1999. Vegeta-tion Dynamics of Mongolia. Kluwer Academic Publishers.
He, S., Liu, H., Ren, J., Yin, Y., 2008. Landform–climate–vegetation patterns andcountermeasures for vegetation rehabilitation of forest-steppe ecotone onsoutheastern Inner Mongolia plateau. Sci. Geogr. Sin. 28, 253–258 (in Chinesewith English abstract).
Held, I., 2012. Climate science: constraints on the high end. Nat. Geosci. 5 (4),236–237.
Hof, C., Levinsky, I., Araujo, M.B., Rahbek, C., 2011. Rethinking species’ ability to copewith rapid climate change. Glob. Change Biol. 17, 2987–2990.
IPCC, 2007. Climate Change 2007: Synthesis Report. Contribution of Working GroupsI, II and III to the Fourth Assessment Report of the Intergovernmental Panel onClimate Change. IPCC, Geneva, Switzerland.
Jackson, S.T., Betancourt, J.L., Booth, R.K., Gray, S.T., 2009. Ecology and the ratchetof events: Climate variability, niche dimensions, and species distributions. Proc.Natl. Acad. Sci. U.S.A. 106, 19685–19692.
Jiang, W., Guo, Z., Sun, X., Wu, H., Chu, G., Yuan, B., Hatté, C., Guiot, J., 2006.Reconstruction of climate and vegetation changes of Lake Bayanchagan (InnerMongolia): Holocene variability of the East Asian monsoon. Quat. Res. 65,411–420.
Katul, G.G., Siqueira, M.B., 2010. Biotic and abiotic factors act in coordination toamplify hydraulic redistribution and lift. New Phytol. 187, 3–6.
Kropelin, S., Verschuren, D., Lézine, A., Eggermont, H., Cocquyt, C., Francus, P., Cazet,J., Fagot, M., Rumes, B., Russell, J., 2008. Climate-driven ecosystem succession inthe Sahara: the past 6000 years. Science 320, 765–768.
Laird, K.R., Fritz, S.C., Maasch, K.A., Cumming, B.F., 1996. Greater drought intensityand frequency before AD 1200 in the Northern Great Plains, USA. Nature 384,552–554.
Lake, P.S., 2011. Drought and Aquatic Ecosystems: Effects and Responses. WileyBlackwell.

rest M
L
L
L
L
L
L
L
M
M
M
M
N
P
P
P
R
R
R
R
S
S
S
Y. Yin et al. / Agricultural and Fo
i, J., Okin, G.S., Epstein, H.E., 2009. Effects of enhanced wind erosion on surfacesoil texture and characteristics of windblown sediments. J. Geophys. Res. 114,G02003.
i, X.Q., Zhou, J., Shen, J., Weng, C.Y., Zhao, H.L., Sun, Q.L., 2004. Vegetation historyand climatic variations during the last 14 ka BP inferred from a pollen record atDaihai Lake, north-central China. Rev. Palaeobot. Palynol. 132, 195–205.
iu, H., Cui, H., Pott, R., Speier, M., 1999. The surface pollen of the woodland-steppeecotone in southeastern Inner Mongolia, China. Rev. Palaeobot. Palynol. 105,237–250.
iu, H., Yin, Y., Tian, Y., Ren, J., Wang, H., 2008. Climatic and anthropogenic controlsof topsoil features in the semi-arid East Asian steppe. Geophys. Res. Lett. 35,L04401.
iu, H., Yin, Y., Zhu, J., Zhao, F., Wang, H., 2010. How did the forest respond to Holoceneclimate drying at the forest-steppe ecotone in northern China? Quat. Int. 227,46–52.
iu, H., Piao, S., Yin, Y., Zhao, F., Zhang, Y., Long-term unusual climate warming inthe semi-arid region of China threatens endangered cranes. Sci. Rep., in review.
u, H., Miao, X., Zhou, Y., Mason, J., Swinehart, J., Zhang, J., Zhou, L., Yi, S., 2005. Latequaternary aeolian activity in the Mu Us and Otindag dune fields (north China)and lagged response to insolation forcing. Geophys. Res. Lett. 32, L21716.
cDowell, N., Pockman, W., Allen, C., Breshears, D., Cobb, N., Kolb, T., Plaut, J., Sperry,J., West, A., Williams, D., 2008. Mechanisms of plant survival and mortality dur-ing drought: why do some plants survive while others succumb to drought?New Phytol. 178, 719–739.
iao, X., Mason, J.A., m Swinehart, J.B., Loope, D.B., Hanson, P.R., Goble, R.J., Liu, X.,2007. A 10,000 year record of dune activity, dust storms, and severe drought inthe central Great Plains. Geology 35, 119–122.
inckley, T.A., Booth, R.K., Jackson, S.T., 2012. Response of arboreal pollen abun-dance to late-Holocene drought events in the Upper Midwest, USA. Holocene22, 531–539.
unson, S.M., Belnap, J., Okin, G.S., 2011. Responses of wind erosion to climate-induced vegetation changes on the Colorado Plateau. Proc. Natl. Acad. Sci. U.S.A.108, 3854–3859.
elson, D.B., Abbott, M.B., Steinman, B., Polissar, P.J., Stansell, N.D., Ortiz, J.D., Rosen-meier, M.F., Finney, B.P., Riedel, J., 2011. Drought variability in the PacificNorthwest from a 6,000-yr lake sediment record. Proc. Natl. Acad. Sci. U.S.A.108, 3870–3875.
arducci, L., Jøgensen, T., Tollefsrud, Elverland, E., Alm, T., Fontana, S.L., Bennett,K., Haile, J., Matetovici, I., Suyama, Y., 2012. Glacial survival of boreal trees innorthern Scandinavia. Science 335, 1083–1086.
eng, C., Guiot, J., Wu, H., Jiang, H., Luo, Y., 2011. Integrating models with data inecology and palaeoecology: advances towards a model—data fusion approach.Ecol. Lett. 14, 522–536.
iao, S., Fang, J., Zhou, L., Guo, Q., Henderson, M., Ji, W., Li, Y., Tao, S., 2003. Interan-nual variations of monthly and seasonal normalized difference vegetation index(NDVI) in China from 1982 to 1999. J. Geophys. Res. 108, 4401.
en, G., Beug, H.J., 2002. Mapping Holocene pollen data and vegetation of China.Quat. Sci. Rev. 21, 1395–1422.
idgwell, A.J., Watson, A.J., 2002. Feedback between aeolian dust, climate, and atmo-spheric CO2 in glacial time. Paleoceanography 17, 1059–1069.
oberts, N., Brayshaw, D., Kuzucuoglu, C., Perez, R., Sadori, L., 2011. The mid-Holocene climatic transition in the Mediterranean: causes and consequences.Holocene 21, 3–13.
otenberg, E., Yakir, D., 2010. Contribution of semi-arid forests to the climate system.Science 327, 451–454.
cherrer, D., Bader, M.K.-F., Körner, C., 2011. Drought-sensitivity ranking of decid-uous tree species based on thermal imaging of forest canopies. Agric. For.Meteorol. 151, 1632–1640.
chlesinger, W.H., Reynolds, J.F., Cunningham, G.L., Huenneke, L.F., Jarrell, W.M., Vir-ginia, R.A., Whitford, W.G., 1990. Biological feedbacks in global desertification.Science 247, 1043–1048.
hi, P., Yan, P., Yuan, Y., Nearing, M.A., 2004. Wind erosion research in China: past,present and future. Prog. Phys. Geogr. 28, 366–385.
eteorology 178–179 (2013) 3–9 9
Shi, Y., Kong, Z., Wang, S., Tang, L., Wang, F., Yao, S., Zhao, X., Zhang, P., Shi, S., 1992.Basic feature of climates and environments during the Holocene Megathermalin China. Sci. China (Ser. B) 35, 1300–1308.
Solomon, S., Plattner, G.K., Knutti, R., Friedlingstein, P., 2008. Irreversible climatechange due to carbon dioxide emissions. Proc. Natl. Acad. Sci. U.S.A. 106,1704–1709.
Steffensen, J.P., Andersen, K.K., Bigler, M., Clausen, H.B., Dahl-Jensen, D., Fischer,H., Goto-Azuma, K., Hansson, M., Johnsen, S.J., Jouzel, J., 2008. High-resolutionGreenland ice core data show abrupt climate change happens in few years.Science 321, 680–684.
Svenning, J.C., 2003. Deterministic Plio-Pleistocene extinctions in the European cool-temperate tree flora. Ecol. Lett. 6, 646–653.
Svenning, J.C., Normand, S., Skov, F., 2008. Postglacial dispersal limitation ofwidespread forest plant species in nemoral Europe. Ecography 31, 316–326.
Thornthwaite, C.W., 1948. An approach toward a rational classification of climate.Geogr. Rev. 38, 55–94.
Tian, B., Liu, R., Wang, L., Qiu, Q., Chen, K., Liu, J., 2009. Phylogeographic analyses sug-gest that a deciduous species (Ostryopsis davidiana Decne., Betulaceae) survivedin northern China during the Last Glacial Maximum. J. Biogeogr. 36, 2148–2155.
Walter, H., Breckle, S.W., 2002. Walter’s Vegetation of the Earth: The EcologicalSystems of the Geo-biosphere. Springer Verlag.
Wang, Y., Cheng, H., Edwards, R., He, Y., Kong, X., An, Z., Wu, J., Kelly, M., Dykoski,C., Li, X., 2005. The Holocene Asian monsoon: links to solar changes and NorthAtlantic climate. Science 308, 854–857.
Wang, Y., Cheng, H., Edwards, R., Kong, X., Shao, X., Chen, S., Wu, J., Jiang, X., Wang,X., An, Z., 2008. Millennial- and orbital-scale changes in the East Asian monsoonover the past 224,000 years. Nature 451, 1090–1093.
Webb, T., 1986. Is vegetation in equilibrium with climate? How to interpret late-Quaternary pollen data. Plant Ecol. 67, 75–91.
Woodward, C., Potito, A., Beilman, D., 2012. Carbon and nitrogen stable isotoperatios in surface sediments from lakes of western Ireland: implications for infer-ring past lake productivity and nitrogen loading. J. Paleolimnol. 47, 167–184 (inChinese).
Wu, Z. (Ed.), 1980. Vegetation of China. Science Press, Beijing (in Chinese).Xiao, J., Chang, Z., Si, B., Qin, X., Itoh, S., Lomtatidze, Z., 2009. Partitioning of the grain-
size components of Dali Lake core sediments: evidence for lake-level changesduring the Holocene. J. Paleolimnol. 42, 249–260.
Xiao, J., Wu, J., Si, B., Liang, W., Nakamura, T., Liu, B., Inouchi, Y., 2006. Holocene cli-mate changes in the monsoon/arid transition reflected by carbon concentrationin Daihai Lake of Inner Mongolia. Holocene 16, 551–560.
Xu, Q., Li, Y., Yang, X., Zheng, Z., 2007. Quantitative relationship between pollen andvegetation in northern China. Sci. China (Ser. B) 50, 582–599.
Yang, X., Zhang, K., Jia, B., Ci, L., 2005. Desertification assessment in China: anoverview. J. Arid Environ. 63, 517–531.
Yi, L., Yu, H., Ge, J., Lai, Z., Xu, X., Qin, L., Peng, S., 2012. Reconstructions of annualsummer precipitation and temperature in north-central China since 1470 ADbased on drought/flood index and tree-ring records. Clim. Change 110, 469–498.
Yin, Y., Liu, H., He, S., Zhao, F., Zhu, J., Wang, H., Liu, G., Wu, X., 2011. Patterns of localand regional grain size distribution and their application to Holocene climatereconstruction in semi-arid Inner Mongolia, China. Palaeogeogr. Palaeoclimatol.Palaeoecol. 307, 168–176.
Zhai, Q., Guo, Z., Li, Y., Li, R., 2006. Annually laminated lake sediments and environ-mental changes in Bashang Plateau, North China. Palaeogeogr. Palaeoclimatol.Palaeoecol. 241, 95–102.
Zhang, X.Y., Gong, S., Zhao, T., Arimoto, R., Wang, Y., Zhou, Z., 2003. Sources of Asiandust and role of climate change versus desertification in Asian dust emission.Geophys. Res. Lett. 30, 2272.
Zhao, F., Liu, H., Yin, Y., Hu, G., Wu, X., 2011. Vegetation succession prevents dry lakebeds from becoming dust sources in the semi-arid steppe region of China. EarthSurf. Proc. Landforms 36, 864–871.
Zhao, Y., Yu, Z., Chen, F., Zhang, J., Yang, B., 2009. Vegetation response to Holocene cli-mate change in monsoon-influenced region of China. Earth Sci. Rev. 97, 242–256.