universidade federal rural de pernambuco … · leonardo brasil mendes ... ufpe/cav recife...
TRANSCRIPT

UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO
DEPARTAMENTO DE BIOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA
LEONARDO BRASIL MENDES
INFLUÊNCIA DA SAZONALIDADE CLIMÁTICA E ESPAÇO VERTI CAL SOBRE
O BANCO DE SEMENTES DO SOLO DE UMA ÁREA DE CAATINGA :
PARALELOS ENTRE ÁREAS PRESERVADA E ANTROPIZADA
RECIFE
FEVEREIRO/2012

LEONARDO BRASIL MENDES
INFLUÊNCIA DA SAZONALIDADE CLIMÁTICA E ESPAÇO VERTI CAL SOBRE
O BANCO DE SEMENTES DO SOLO DE UMA ÁREA DE CAATINGA :
PARALELOS ENTRE ÁREAS PRESERVADA E ANTROPIZADA
Dissertação apresentada ao Programa de Pós-
Graduação em Ecologia da Universidade Federal
Rural de Pernambuco, como parte dos requisitos
para obtenção do título de mestre em Ecologia.
Orientadora: Profª. Drª. Elcida de Lima Araújo – UFRPE Co-orientadores: Prof. Dr. Ulysses Paulino de Albuquerque – UFRPE Prof. Dr. Kleber Andrade da Silva – UFPE/CAV
RECIFE
FEVEREIRO/2012

Ficha catalográfica
M538i Mendes, Leonardo Brasil Influência da sazonalidade climática e espaço vertical sobre o banco de sementes do solo de uma área de caatinga: paralelos entre áreas preservada e antropizada / Leonardo Brasil Mendes. – Recife, 2012. 83 f. : il. Orientadora: Elcida de Lima Araújo. Dissertação (Mestrado em Ecologia) – Universidade Federal Rural de Pernambuco, Departamento de Biologia, Recife, 2012. Inclui referências, anexo e apêndice.
1. Serrapilheira 2. Regeneração natural 3. Campo abandonado 4. Antropização 5. Estação I. Araújo, Elcida de Lima, orientadora II. Título CDD 574.5

ii
LEONARDO BRASIL MENDES
Influência da sazonalidade climática e espaço vertical sobre o banco de sementes do solo
de uma área de caatinga: paralelos entre áreas preservada e antropizada
Dissertação defendida no dia 15 de fevereiro de 2012 e aprovada pela banca examinadora:
Orientadora:
_____________________________________________ Profª. Drª. Elcida de Lima Araújo – UFRPE
Examinadores:
___________________________________________ Profª. Drª. Cibele Cardoso de Castro – UFRPE
___________________________________________ Dr. Marcelo Alves Ramos – UFRPE
___________________________________________ Profª. Drª. Margareth Ferreira Sales – UFRPE
RECIFE
FEVEREIRO/2012

iii
Aos amigos e colegas que fiz ao longo desses dois anos
de curso e laboratório, em especial aos professores e
orientadores, pois todos contribuíram na minha formação como
ecólogo, inclusive, repassando conhecimentos e experiências
que transcenderam a minha formação acadêmica e adentraram
na pessoal.
Dedico.

iv
AGRADECIMENTOS
Esta dissertação é fruto de trabalho árduo e de um desafio interdisciplinar.
As pessoas e instituições aqui citadas, ao longo da formação desta humilde obra,
deixaram de alguma forma suas marcas no resultado final obtido.
A Deus, pela vida, pela saúde e pelas felicidades que tenho até hoje.
A minha orientadora, Profª. Drª. Elcida de Lima Araújo, que, com compreensão e
paciência, aceitou este desafio interdisciplinar de orientar um bacharel em Direito,
contribuindo na minha formação acadêmica e profissional, ao formar o único advogado do
Brasil especializado em ecologia. Muito obrigado Profª Elcida!
Ao co-orientador Dr. Ulysses Paulino de Albuquerque, pelas críticas e conselhos que
contribuíram na construção desta obra, também pelas excelentes aulas no PPGE e PPGB.
Ao também co-orientador Dr. Kleber Andrade da Silva pela atenção, dedicação e
paciência, além das críticas e contribuições para esta obra.
Ao Instituto Agronômico de Pesquisas Agropecuárias – IPA, unidade de Caruaru,
pelo apoio logístico, receptividade e amizade dos seus funcionários.
A todos os mestres do Programa de Pós-Graduação em Ecologia – PPGE/UFRPE, que
me serviram de fonte e são os formadores do meu conhecimento sobre meio ambiente.
Aos herbários Professor Vasconcelos Sobrinho (PEUFR) da Universidade Federal
Rural de Pernambuco e Dárdano de Andrade Lima, da Empresa Pernambucana de Pesquisa
Agropecuária (IPA) pela disponibilização do material botânico.
A toda a turma do LEVEN: Kleber, Josiene, Clarissa, Juliana, Elifábia, Danielle,
Priscila, Renata, Diego, Izabelle, Thiago, Vanessa, Hermano, Michelle e Aline, pela grande
amizade e apoio na conclusão deste trabalho. Muito obrigado LEVENanos! Em especial
agradeço a Kleber e a Dani, por todo o apoio, dedicação, amizade e ensinamentos sobre banco
de sementes, vocês foram incríveis comigo.
Aos meus eternos amigos do Colégio Vera Cruz: João, Pablo, Alexandre, Marcelo,
Gustavo, Rodrigo, Gaspar, Ricardo e ao restante da turma.
À minha família: Auxiliadora, Viviane, Gabriela, Hélio, Heitor e a todos os tios, pela
franqueza e carinho sempre abundantes.
In memore, ao meu querido pai, Arremar e a minha querida avó, Zuleide.

v
SUMÁRIO
AGRADECIMENTOS iv
LISTA DE TABELAS vi
LISTA DE FIGURAS vii
RESUMO ix
ABSTRACT x
1. INTRODUÇÃO 11
2. FUNDAMENTAÇÃO TEÓRICA 13
2.1. Influência da sazonalidade sobre a dinâmica do banco de sementes 13
2.2. Dinâmica do banco de sementes em ambientes secos 14
2.3. Variação do banco de sementes no espaço vertical 19
2.4. . Importância do banco de sementes de campos abandonados 23
3. REFERÊNCIAS BIBLIOGRÁFICAS 27
Artigo a ser enviado ao periódico Journal of Arid Environments 38
Variação espaço-temporal do banco de sementes em uma região semiárida no Nordeste do Brasil: paralelos entre áreas preservada e antropizada
39
Resumo 40
1. Introdução 41
2. Material e métodos 43
2.1. Área de estudo 43
2.2. Amostragem do banco de sementes 45
2.3. Análises do banco de sementes 46
3. Resultados 47
3.1. Riqueza e composição florística do banco de sementes do campo 47
3.2. Densidade do banco de sementes do campo 48
3.3. Diferenças entre o banco de sementes do campo e do remanescente florestal 49
4. Discussão 49
4.1. Riqueza e composição florística do banco de sementes 50
4.2. Densidade do banco de sementes do campo 52
4.3. Diferenças entre o banco de sementes do campo e do remanescente florestal 55
Agradecimentos 57
Referências bibliográficas 58
APÊNDICE 1 66
ANEXOS 73

vi
LISTA DE TABELAS
Manuscrito:
Tabela 1. Variação espaço-temporal na riqueza e densidade média (± desvio padrão) do
banco de sementes de uma área de agricultura abandonada em uma região semiárida no
Nordeste do Brasil. Valores em negrito denotam diferença significativa pelo teste de Tukey.
...................................................................................................................................................69
Tabela 2. Variação na riqueza de espécies e na densidade de sementes (GLM) por estação (chuva e seca), distribuição das sementes no espaço vertical - DEF (serrapilheira e solo) e interações................................................................................................................................70
Tabela 3. Riqueza média de espécies e número médio de plântulas germinadas das amostras
de serrapilheira (SR) e de solo (SL) (0-5cm de profundidade) do Campo e do Remanescente
florestal (RF), em uma região semi-árida no Nordeste do Brasil. Valores de P em negrito
denotam diferenças significativas (P < 0,05), utilizando o teste t uma amostra.......................71

vii
LISTA DE FIGURAS
Manuscrito:
Figura 1. Desenho esquemático da plotagem das parcelas amostrais no campo abandonado
próximo a um remanescente de floresta seca na Estação Experimental do Instituto
Agronômico de Pernambuco (IPA), município de Caruaru-PE. No detalhe da parcela, 1ª
coleta na estação seca e 2ª coleta na estação chuvosa, ambas em subparcelas.......................72

ix
RESUMO
Estudos sobre o banco de sementes permitem avaliar a renovação das plantas em florestas e a
conservação da diversidade vegetal nos habitats, assim como estimar as comunidades vegetais
futuras em uma área. No semiárido do nordeste brasileiro, durante um ano, são previsíveis as
estações chuvosa e seca as quais influenciam as espécies da vegetação caatinga na fenologia,
no ciclo de vida e, consequentemente, na dinâmica do banco de sementes cuja estocagem no
solo pode ocorrer desde as camadas mais rasas até as mais profundas ou sobre ele
(serrapilheira) em dada área e momento. Porém, a vegetação que se estabelece em áreas de
agricultura abandonada modificam as características do banco de sementes em comparação
com a floresta existente no passado. Em campos abandonados, o banco de sementes pode dar
início à regeneração das comunidades vegetais, e depois vem a fazer parte de um processo
continuo de regeneração. Por isso, as características do banco de sementes foram verificadas
em um campo abandonado durante as estações chuvosa e seca, e comparadas com outro
estudo sobre o banco de sementes da floresta seca remanescente que também utilizou o
método de emergência de plântulas. Em uma região do semiárido do Nordeste do Brasil, parte
de um fragmento de floresta seca foi convertido em área agrícola que após o abandono há 17
anos já apresenta uma cobertura vegetal, ficando evidente a regeneração das comunidades de
plantas na área. Neste campo abandonado foram coletadas 105 amostras de solo e de
serrapilheira nos finais das estações seca e chuvosa, totalizando 420 amostras. Verificou-se
que o banco de sementes foi dominado por espécies herbáceas e a similaridade florística entre
estações foi de 57% e entre serrapilheira e solo foi de 70%. Não houve diferença significativa
na variação sazonal da riqueza e da densidade de sementes do solo entre estações. Por outro
lado, a riqueza e a densidade de sementes foram significativamente maiores no remanescente
florestal do que no campo. Apesar de não ter havido variação sazonal nos totais do banco do
solo, isoladamente na serrapilheira foram registradas variações na riqueza e na densidade de
sementes, e isoladamente na camada rasa de solo foram registradas variações na riqueza,
sugerindo que o banco de sementes do campo não é sazonalmente constante. As diferenças
encontradas entre a serrapilheira e o solo também vêm sendo registradas em outras florestas
secas e é uma tendência que vem se repetindo sucessivamente no remanescente florestal.
Evidencia-se que em 17 anos de regeneração natural o campo recuperou parte da composição
florística, riqueza de espécies, densidade do banco de sementes do solo, mas estas
características ainda são inferiores às registradas no remanescente florestal.
Palavras chave: serrapilheira, regeneração natural, campo abandonado, antropização, estação.

x
ABSTRACT
Studies on seed banks allow us to evaluate forest renewal processes and plant diversity
conservation in different habitats, as well as to estimate what plant communities may establish
themselves in the forest in the future. In the semi-arid region of northeastern Brazil,
throughout the year it is possible to predict the rainy and dry seasons, which influence
caatinga plant species’ phenology, life cycle and, consequently, seed banks’ dynamics. The
seed bank, in turn, may be stored in the soil’s shallowest or deeper layers, or above the soil
(leaf litter), at a given area and time. Nevertheless, the vegetation that establishes itself in
abandoned agricultural areas modifies the seed bank’s characteristics when compared to the
forest that existed in the same area in the past. In abandoned fields the seed bank may: initiate
(or not) the regeneration of plant communities; take years to set off regeneration; or initiate it
right after the area’s abandonment. Thus, seed bank characteristics were verified in an
abandoned field during the rainy and dry seasons, and compared with another study about the
seed bank of an adjacent dry forest remnant that also used the plantule emergence method.
The locale studied is situated in Northeast Brazil’s semi-arid region and is part of a dry forest
fragment that was converted into an agricultural area; 17 years after abandonment it already
has a vegetation cover and the regeneration of the area’s plant community is evident. One-
hundred and five soil and leaf litter samples were collected from this abandoned field at the
end of the dry and rainy seasons – a total of 420 samples. The seed bank was dominated by
herbaceous species and the floristic similarity was of 57% between seasons and of 70%
between the leaf litter and the soil. There were no significant differences in the seasonal
variations of soil seed richness and density between seasons. Conversely, seed richness and
density was significantly higher in the forest remnant than in the abandoned field. Although
there was no seasonal variation in the soil’s seed bank totals, variations in seed richness and
density were recorded isolatedly in the leaf litter and in the soil’s shallow layer, which
suggests that the field’s seed bank is not seasonally constant. The differences found between
the leaf litter and the soil have also been recorded in other dry forests and is a trend that has
been repeating itself successively at the forest remnant studied. It is evident that in 17 years of
natural regeneration the field has recovered part of its floristic composition, species richness
and soil seed bank density, yet the values of such variables are still lower than those recorded
in the forest remnant.
Keywords: leaf litter, natural regeneration, abandoned field, anthropization, season.

11
1. INTRODUÇÃO
A dinâmica do banco de sementes do solo tem importância vital para regeneração das
comunidades vegetais em áreas antropizadas (GARWOOD, 1989; VIEIRA e SCARIOT,
2006), pois o início da regeneração depende da viabilidade e germinação do estoque de
sementes armazenadas no solo (VIEIRA e SCARIOT, 2006).
Porém, o número de trabalhos sobre as características do banco de sementes da
caatinga é reduzido e concentrado em áreas de estudo sem relato de interferência antrópica
(COSTA e ARAÚJO, 2003; PESSOA, 2007; LOBO, 2008; SANTOS, 2010; BARBOSA,
2008; SILVA, 2009). Estes estudos têm indicado que: a densidade de sementes diminui com o
aumento da profundidade no solo, acompanhando a tendência de outras florestas secas do
mundo (COFFIN e LAUENROTH, 1989; FACELLI et al., 2005; WILLIAMS et al., 2005); a
densidade é maior no solo do que na serapilheira (mas vide COSTA e ARAÚJO, 2003); a
composição florística do banco de sementes não é similar entre estações chuvosa e seca; e as
diferenças na composição de espécies e densidade de sementes do banco do solo entre as
estações climáticas podem variar entre os anos.
As florestas tropicais secas, originalmente, ocupavam 42% da vegetação tropical do
mundo (MURPHY e LUGO, 1995), porém, atualmente, correspondem a cerca de 24% dessa
vegetação (KHURANA e SINGH, 2001; VIEIRA e SCARIOT, 2006). Apesar de ser um
ecossistema tropical terrestre ameaçado por ações antrópicas que convertem rapidamente a
floresta em áreas agrícolas (MURPHY e LUGO, 1995), as florestas secas ainda ocupam
grande extensão dos continentes Americano, Africano, Asiático e a Oceania (KHURANA e
SINGH, 2001; VIEIRA e SCARIOT, 2006).
No nordeste do Brasil, a fitofisionomia de caatinga é a principal represente das
florestas tropicais secas (SAMPAIO, 1995), apresentando marcada sazonalidade climática,
com 6 a 9 meses secos por ano e totais pluviométricos anuais variando de 380 a 1.100mm
(SAMPAIO, 1995; ARAÚJO et al., 2005a; ARAÚJO et al., 2007); elevada riqueza de
espécies (herbáceas e lenhosas), comportamento fenológico influenciado pelas estações
climáticas (MACHADO et al., 1997; GRIZ e MACHADO, 2001), marcantes diferenças
sazonais na chuva de semente (ARAÚJO et al., 2005b) e na dinâmica do banco de sementes
do solo (COSTA e ARAÚJO, 2003; SILVA, 2009).

12
Esse tipo de vegetação seca vem sendo modificada por ações antrópicas
(CASTELLETTI et al., 2003). O corte raso, a queima e o cultivo itinerante tem sido as
principais práticas executadas no semiárido do nordeste brasileiro para converter florestas em
áreas agrícolas (KAUFFMAN et al., 1993; SAMPAIO et al., 1993; PEREIRA et al., 2003;
MAMEDE e ARAÚJO, 2008), e estas ações antrópicas causam um grande impacto sobre o
banco de sementes do solo, representando uma séria ameaça à conservação de espécies da
vegetação caatinga (MAMEDE e ARAUJO, 2008).
Porém, como alguns campos são abandonadas devido ao modo itinerante de cultivo
tradicionalmente praticado no semiárido do nordeste brasileiro (MAMEDE e ARAÚJO,
2008), as comunidades vegetais podem começar a se regenerar naturalmente formando
“campos abandonados sucessionais” (RICKFLES, 2009), exibindo mudanças na composição
de espécies e diferenças na estrutura da comunidade (PEREIRA et al., 2003), indicando que
os processos ecológicos e os fatores que influenciam a regeneração são alterados.
Apesar da maioria destes estudos mostrarem que a sazonalidade climática é um fator
de influência na disponibilidade de sementes para a renovação do estoque no banco do solo,
praticamente nada se sabe sobre o que ocorre com essa influência em um campo abandonado,
pois o único estudo realizado em uma floresta de caatinga suprimida (MAMEDE e ARAÚJO,
2008) analisou apenas o efeito do corte e da queima sobre o banco de sementes. Além disso,
não se sabe se o maior estoque de sementes é no solo ou na serrapilheira em um campo
abandonado, dificultando uma discussão sobre: os impactos das ações humanas na resiliência
das comunidades vegetais; as diferenças entre campos abandonados e florestas secas maduras;
e o papel do banco de sementes na regeneração natural em campos abandonados.
A partir da premissa de que a sazonalidade climática é um fator influenciador da
dinâmica do banco de sementes de florestas secas e que as ações antrópicas causam mudanças
nas características do banco de sementes do solo de áreas florestais, a hipótese deste estudo é
que a sazonalidade climática é um fator influenciador da dinâmica do banco de sementes
também em uma área de cultivo abandonada, porém a composição florística, a riqueza e a
densidade de sementes do banco solo são diferentes do banco de sementes da floresta tropical
seca remanescente. O objetivo deste trabalho foi avaliar as características do banco de
sementes de um campo abandonado e compará-lo com o estudo de Silva (2009) realizado em
uma floresta tropical seca circunvizinha a este campo, onde foram constatadas variações
sazonais no banco de sementes.

13
2. FUNDAMENTAÇÃO TEÓRICA
2.1. Influência da sazonalidade sobre a dinâmica do banco de sementes
Variação sazonal é a mudança de estações climáticas durante uma escala de tempo
previsível (TOWNSEND et al., 2006). Durante um ano, de acordo com cada ambiente, as
estações podem variar: entre primavera, verão, outono e inverno (TOWNSEND et al., 2006;
RICKLEFS, 2009) ou entre estações chuvosa e seca (REICHMAN, 1984; PRICE e JOYNER,
1997; ARAÚJO et al., 2007) ou manter-se constante (TOWNSEND et al., 2006).
Cada estação climática apresenta uma série de condições ambientais fazendo com que
os organismos se adaptem para sobreviver (TOWNSEND et al., 2006). Por isso, algumas
populações vegetais com uma história de vida anual estão adaptadas a sobreviver e completar
o ciclo de vida em determinada estação do ano, morrendo nas estações subseqüentes
(TOWNSEND et al., 2006), mas perpetuando a espécie ao suportar as condições estacionais
adversas na forma de sementes no banco do solo e germinando na próxima estação favorável
(RAUNKIAER, 1934; COSTA et al., 2007).
Por outro lado, algumas populações vegetais sobrevivem durante o ano todo, mas com
o comportamento influenciado pela sazonalidade, permitindo relacionar a estação climática ao
período de dispersão de sementes, consequentemente, à entrada de sementes recém
produzidas no banco do solo (MACHADO et al., 1997; GRIZ e MACHADO, 2001; LIMA et
al., 2007).
Desta forma, em florestas tropicais de sazonalidade marcante, a dispersão dos
propágulos não é contínua (GARWOOD, 1989). Por isso, as médias do banco de sementes
flutuam durante o ano, havendo variação entre estações climáticas (GARWOOD, 1989;
SIMPSON, et al., 1989; ALMEIDA-CORTEZ, 2004).
A sazonalidade climática também está relacionada à dinâmica de saída das sementes
do banco do solo, uma vez que os fenômenos que ocorrem durante uma estação podem
potencializar ou amenizar a ação da água e do vento na dispersão secundária, assim como a
presença e ausência de animais dispersores ou predadores de sementes (WALCK et al., 2005).
Por isso, estações com climas amenos podem oferecer condições e recursos requeridos
pelas sementes (nutrientes, luminosidade, temperatura e, principalmente, água) possibilitando
a germinação (BAKER, 1989; ROBERTS e SIMPSON, 1989; ALMEIDA-CORTEZ, 2004).

14
E, também, podem atrair predadores e proliferar patógenos que podem conduzir à morte
fisiológica das sementes (LABOURIAU, 1983; BASKIN e BASKIN, 1989).
Enquanto estações com climas extremos ou estressantes podem estender o período de
dormência até a semente atingir a senescência (LABOURIAU, 1983; BASKIN e BASKIN,
1989). Outros eventos podem atuar tanto na saída como entrada de sementes, como a remoção
que está ligada aos processos de dispersão secundária que podem ocorrer através do vento, da
água ou do transporte por animais (SIMPSON, et al., 1989; ALMEIDA-CORTEZ, 2004), que
podem distribuir ou concentrar sementes num espaço em dado momento (THOMPSON e
GRIME, 1979; ALMEIDA-CORTEZ, 2004), exigindo uma compreensão apurada da atuação
dos dispersores atuantes em uma área para entender a dinâmica do banco de sementes na
mesma.
Portanto, existe uma grande influência da variação sazonal sobre a densidade e a
riqueza de espécies de semente do banco do solo no tempo (THOMPSON e GRIME, 1979).
Mas durante um ano, os fenômenos característicos de uma estação climática podem ser mais
influentes ou menos influentes quando comparado entre anos (SILVA, 2009). Assim como o
período de duração de cada estação pode ser mais longo ou curto (TOWNSEND et al., 2006,
RICKLEFS, 2009).
2.2. Dinâmica do banco de sementes em ambientes secos
As sementes viáveis existentes no solo desde a superfície até as camadas mais
profundas formam o banco de sementes de um determinado espaço no tempo (ALMEIDA-
CORTEZ, 2004), representando o armazenamento da memória evolutiva das plantas
(HARPER, 1977), e permitindo estimar as comunidades vegetais futuras. No entanto, a
dinâmica do banco de sementes é controlada pela entrada e saída de sementes no solo
(ALMEIDA-CORTEZ, 2004), que varia no tempo e no espaço (THOMPSON e GRIME,
1979; ALMEIDA-CORTEZ, 2004).
Em florestas tropicais, a densidade e a riqueza de sementes estocadas no solo podem
estar relacionadas com a época de dispersão e emergência de plântulas (GARWOOD, 1989;
SIMPSON, et al., 1989), o mesmo ocorre em florestas secas, como a caatinga (ALMEIDA-
CORTEZ, 2004; BARBOSA, 2008; SILVA, 2009). Estas por sua vez seriam influenciadas
pela variação pluviométrica entre as estações climáticas de chuva e de seca (GARWOOD,
1989; CABALLERO et al., 2005; ALMEIDA-CORTEZ, 2004; BARBOSA, 2008; SILVA,

15
2009), mas o banco de sementes também pode ser reflexo da intensidade de eventos
pluviométricos de anos passados (SILVA, 2009).
Desta forma, em ambientes tropicais secos, os eventos pluviométricos geralmente se
concentram em determinada época do ano, ficando seco o restante dos meses. Por isso, as
populações vegetais adaptaram o ciclo de vida e o comportamento fenológico aproveitando ao
máximo a disponibilidade de água, havendo assim variação sazonal também no banco de
sementes.
Kellerman e Van Rooyen (2007) investigaram a riqueza e a densidade do banco de
sementes do solo em cinco vegetações diferentes localizadas na África do Sul, algumas áreas
são consideradas úmidas e outras secas, mas todas possuem estações climáticas definidas.
Como os autores estudaram áreas com variações sazonais que ocorrem em tempos diferentes
no calendário, adotaram como critério a época em que ocorre a dispersão de sementes e a
época em que ocorre a emergência de plântulas nas áreas estudadas, independentemente da
data em que ocorrem esses eventos no calendário. Das cinco áreas estudadas, apenas uma não
apresentou variação sazonal significativa, sendo maior a densidade e a riqueza de sementes no
período após a dispersão e menor no período após a emergência de plântulas.
Caballero et al. (2005) estudaram a riqueza, a composição e a densidade do banco de
sementes em uma área de floresta seca na Espanha, realizando coletas sazonais. A primeira
coleta realizada logo após o período de dispersão da maioria das sementes, mas antes do
surgimento das plântulas, apresentou a maior riqueza, 69 espécies. Enquanto a segunda coleta
realizada no período de emergência de plântulas, mas antes da dispersão das sementes
apresentou uma riqueza menor, apenas 42 espécies. Já a densidade de sementes no banco foi
três vezes maior na primeira amostragem do que na segunda. Com relação à composição
florística, a sazonalidade climática foi considerada um efeito secundário, diante de outras
variáveis analisadas, como o relevo do terreno.
Os valores discrepantes da densidade de sementes no banco do solo foram atribuídos à
dispersão tardia e a estratégia do banco de sementes persistente (THOMPSON e GRIME,
1979 apud CABALLERO et al., 2005). Desta forma, os resultados obtidos sugerem uma
interferência da sazonalidade sobre os processos de dispersão, germinação e morte fisiológica
das sementes, consequentemente sobre a dinâmica do banco. A correlação entre a densidade
de sementes no banco do solo e a concentração das precipitações em estações climáticas
distintas (chuvosa e seca) também vem sendo relatada em outros trabalhos realizados em

16
áreas secas (REICHMAN, 1984; PRICE e REICHMAN, 1987; KEMP, 1989; PRICE e
JOYNER, 1997).
Traba et al. (2006) fizeram coletas sazonais do solo em uma área de pasto e em outra
de floresta, registrando que a densidade de sementes do banco diminuiu significativamente
durante o verão nas duas áreas. Também no verão, a riqueza de espécies diminuiu na floresta
e a composição florística variou entre áreas.
Da análise dos resultados, os autores relataram que o verão é seco e a coleta sazonal de
verão foi realizada após a dispersão de sementes e antes da emergência de plântulas, ou seja,
período mais propenso a uma densidade elevada no banco de sementes, porém, a predação de
sementes por formigas é intensa nas áreas durante o verão (TRABA, 2000). Relatam ainda
que as formigas carreiam as sementes para seus ninhos durante o verão (TRABA, 2000), para
terem estoque de recursos durante as estações frias, concluindo que a predação pós-dispersão
interfere na densidade de sementes nas camadas de solo mais superficiais. Constata-se que o
forrageamento das formigas também é um fator influenciado pela sazonalidade climática,
assim como a fenologia e a dispersão de propágulos, pois a predação de sementes por
formigas ocorre durante o verão, tornando assim previsível estes processos ecológicos.
Ao norte do Colorado - EUA, Coffin e Lauenroth (1989) investigaram as variações
entre anos do banco de sementes sobre a influência da sazonalidade e da textura do solo,
registrando diferenças significativas entre estações, enquanto as diferenças entre anos não
foram significativas. A variação na temperatura e na precipitação da área de estudo ao longo
dos anos é o aspecto sazonal que deve ter interferido na dinâmica do banco de sementes
segundo os autores, concluindo que os indícios apontam para a existência de um banco
transitório de sementes maior do que o banco persistente.
Desta forma, o final de cada estação climática concentra as maiores variações do
banco de sementes, pois a transição entre estações gera impulsos nas plantas e sementes,
influenciando na dispersão e no período de emergência de plântulas (BERTILLER e ALOIA,
1997; GHERMANDI, 1997; MAYOR et al., 1999; LÓPEZ, 2003; MAYOR et al., 2003;
FLORES e DEZZEO et al., 2005; FACELLI et al., 2005; WILLIAMS et al., 2005; WANG et
al., 2005; SHEN et al., 2007; NING et al., 2007). E, como as espécies podem adotar
estratégias diferentes diante da previsibilidade das estações climáticas (BASKIN e BASKIN,
1989; WALCK et al., 2005), a composição florística do banco de sementes pode variar entre
estações.

17
Neste sentido, o banco de sementes da caatinga costuma variar entre estações
(PESSOA, 2007; LOBO, 2008; BARBOSA, 2008; SILVA, 2009; SANTOS, 2010), sendo a
estação chuvosa o período de maior emergência de plântulas. A estação seca é mais severa,
quando ocorre a da caducifólia da maioria das espécies lenhosas e elevada mortalidade,
sobretudo de plantas (BARBOSA et al., 1989; MACHADO et al.,1997; BARBOSA et al.,
2003).
Pessoa (2007), na encosta de uma área de caatinga preservada de Serra Talhada-PE,
realizou cinco coletas de solo na parte superior (convexa) e outras cinco coletas na parte
inferior da encosta (côncava) de três sulcos e de três entressulcos (áreas entre os sulcos), em
duas estações do ano, totalizando 120 amostras e constatou que só existe diferença na
densidade de sementes do banco do solo nas condições de sulco e entressulco durante a
estação chuvosa, mas que no conjunto o quantitativo de sementes no banco do solo difere
entre estações climáticas. No total, a autora, registrou 5.225 sementes, sendo 4.217 na estação
chuvosa e 1.008 sementes na seca, com 79% das espécies herbáceas e 21% lenhosas.
Lobo (2008) avaliou o tempo de permanência das sementes viáveis no banco do solo
de uma área de caatinga, assim como investigou os mecanismos de dormência das sementes,
identificando bancos transitórios e permanentes. A longevidade das sementes no banco foi
determinada e as espécies que refletem tardiamente a influência das variações climáticas na
dinâmica populacional foram identificadas. Para tanto, Lobo (2008) simulou em laboratório
uma estação chuvosa seguida de uma estação seca durante a aplicação do método de
emergência de plântulas. Antes da simulação de seca registrou na serrapilheira uma densidade
de sementes de 49,5 sem.m-2, e na camada do solo de 0-5 cm registrou uma densidade de
310,9 sem.m-2. Depois da simulação de seca registrou uma densidade de 14,5 sem.m-2 na
serrapilheira e de 284,04 sem.m-2 no solo de 0-5 cm de profundidade.
Lobo (2008) observou que a maioria das espécies apresentou banco de sementes
transitório ou com tendência a ser transitório no solo, possivelmente porque a maioria das
espécies da caatinga são herbáceas e terófitas (ARAÚJO et al., 2005a; REIS et al., 2006),
porém, não foi observado um banco de sementes claramente permanente. Apenas algumas
espécies, que apresentaram tendência a possuir banco permanente, foram vistas, indicando
que as sementes de algumas espécies podem permanecer dormentes no solo durante a estação
chuvosa e precisam suportar uma estação seca, para germinarem na estação favorável
seguinte. Com isso, Lobo (2008) evidenciou que o monitoramento das amostras do solo por

18
um período de apenas seis meses subestima a densidade real de sementes do solo, pois 45,3%
da densidade de sementes presentes no banco do solo foram registradas na segunda simulação
de chuva.
Barbosa (2008) analisou a variação sazonal do banco de sementes do solo em uma
mata ciliar coletando 36 amostras de solo de 50 x 50 cm de largura e 5 cm de profundidade
distribuídas em 1,02ha, identificando as espécies e contabilizando-as através do método de
emergência de plântulas. Assim sendo, 165 espécies representaram a flora do banco, sendo
122 espécies no final da estação seca e 124 no final da estação chuvosa, já a densidade foi
maior no final da estação seca (887 sementes/m2) do que no final da estação chuvosa (616
sementes/m2).
Silva (2009) estabeleceu 105 parcelas de 20x20cm em uma área preservada da
caatinga e coletou amostras nos finais das estações chuvosa e seca, durante três anos
consecutivos, na serrapilheira e na camada de 0-5 cm de profundidade do solo. O autor
observou que a similaridade florística interanual do banco de sementes de uma área de
caatinga era menor que 50%; que a densidade de sementes do banco solo é maior na
profundidade de 0-5 cm que na serrapilheira; e a densidade de sementes pode ser maior tanto
na estação chuvosa quanto na seca ou manter-se constante durante o ano.
Santos (2010) estabeleceu 105 parcelas de 20x20 cm em três microhabitats (ciliar,
rochoso e plano) e duas camadas (solo superficial com 5 cm de profundidade e a serrapilheira)
de uma área de caatinga preservada, e fez uma amostragem em cada estação climática (seca e
chuvosa) e constatou que a densidade de sementes no banco do solo foi significativamente
superior na estação chuvosa quando comparado à estação seca e que existem diferenças nas
condições de microhabitats no quantitativo de sementes estocado no banco.
Por outro lado, após a dispersão, as sementes podem formar no solo um banco
persistente, permanecendo estocadas por um período até germinarem (GARWOOD, 1989). A
literatura geralmente utiliza um período de cinco anos para propor uma classificação (entre
longa e curta duração) para o banco de sementes persistente (BAKKER et al., 1996).
O banco de sementes persistente é uma estratégia que pode ter um importante papel
para a regeneração natural em florestas tropicais (GARWOOD, 1989), pois a dormência
possibilita que uma área, após a interferência de ações antrópicas, ainda possua sementes
armazenadas no solo que possam iniciar a regeneração natural das comunidades vegetais,
independentemente da chuva de sementes alóctones.

19
Thompson e Grime (1979) propuseram a diferença entre banco de sementes transitório
e persistente através de uma linha do tempo de um ano entre a entrada das sementes no solo e
a germinação, permanecendo as sementes transitórias até um ano estocadas no solo e as
persistentes por mais de um ano. Porém, Walck et al. (2005) propuseram um conceito
ecológico-evolutivo que consiste na contagem de estações em vez de anos ou meses.
Segundo Walck et al (2005), o banco transitório seria composto por sementes que
permanecessem viáveis da entrada no solo até a segunda estação chuvosa, a qual é favorável a
germinação. Já o banco de sementes persistente mantém sementes viáveis no solo por tempo
indeterminado.
O conceito proposto por Walck et al. (2005) complementam os conceitos de estudos
anteriores (por exemplo: BASKIN e BASKIN, 1989) sobre variações sazonais que ocorrem
em florestas tropicais secas, pois um ano possui as estações seca e chuvosa bem definidas
(MURPHY e LUGO, 1995; SAMPAIO, 1995), onde a chuvosa é a estação favorável a
germinação, enquanto a seca é desfavorável (COSTA e ARAÚJO, 2003), podendo as
sementes ficarem estocadas no solo até a chegada da estação chuvosa (WALCK et al., 2005).
Por isso, o banco de sementes persistente é reflexo de eventos pluviométricos
ocorridos há pelo menos dois anos, enquanto o banco de sementes transitório é reflexo da
última estação chuvosa.
Geralmente as sementes do banco transitório germinam na primeira estação chuvosa
pós-dispersão (WALCK et al., 2005), mas chuvas erráticas podem ocorrer dentro da estação
seca que precede a estação chuvosa pós dispersão e induzir a germinação. Contudo, devido à
escassez de água nesta estação seca, a mortalidade das plântulas é elevada (ALMEIDA-
CORTEZ, 2004; ARAÚJO et al., 2005a; ARAÚJO et al., 2005b; LIMA et al., 2007; SILVA,
2009). Desta forma, a quantidade de ocorrência de chuvas eventuais durante a estação seca
pode influenciar a densidade do banco de sementes do final da estação seca, reduzindo o
estoque de sementes para renovação das populações.
2.3. Variação do banco de sementes no espaço vertical.
Quanto à influência da variação sazonal sobre a densidade e a riqueza de semente do
banco do solo no espaço (THOMPSON e GRIME, 1979), a dinâmica do banco de sementes
estaria mais relacionada aos processos de dispersão primária e secundária, cuja distribuição no
solo destes propágulos ocorreria verticalmente e horizontalmente (SIMPSON et al., 1989).

20
No entanto, os estudos sobre a distribuição no espaço vertical do banco de semente já
aponta como tendência uma maior densidade nas camadas mais rasas e redução do banco de
sementes com o aumento da profundidade (COFFIN e LAUENROTH, 1989; WILLIAMS et
al., 2005). Luzuriaga et al. (2005) também constatou esta tendência ao estudar perfis de solo
num campo abandonado de uma região subúmida da Espanha, concordando com a literatura
que relata esta tendência ao atribuir os resultados à morte fisiológica das sementes que não
resistem às condições impostas pelas camadas de solo mais profundas e à incapacidade de
emergir em camadas profundas.
Em ambientes secos, o vento, a germinação, a predação de sementes, a variação
topográfica da superfície do solo e o acúmulo de serrapilheira são influenciadores da
distribuição (vertical e horizontal) de sementes no solo (GOODALL et al., 1972;
REICHMAN, 1984), assim como são responsáveis pela dinâmica de entrada e saída das
sementes no banco do solo (SIMPSON et al., 1989).
A síndrome de dispersão anemocórica tende a ser maior em florestas tropicais secas,
enquanto em áreas úmidas a síndrome de dispersão zoocórica é mais freqüente (GENTRY,
1983; WILLSON et al., 1989). Frutos com estruturas aladas permitem que a dispersão pelos
ventos fortes seja longa (HOWE e SMALLWOOD, 1982).
Ao se desprenderem da planta as sementes, geralmente, atingem primeiro a
serrapilheira e depois são enterradas no solo. Desta forma, os ventos podem atuar na dinâmica
do espaço vertical, remexendo serrapilheira e dispersando as sementes para o solo.
Esta síndrome de dispersão anemocórica secundária pode ser mais intensa durante a
estação seca, quando o dossel está aberto, permitindo que rajadas de vento penetrem na
vegetação e atinja diretamente a serrapilheira.
A germinação é uma forma de saída das sementes do banco do solo (SIMPSON et al.,
1989; ALMEIDA-CORTEZ, 2004), podendo ocorrer assim que atingem a serrapilheira ou o
solo (GARWOOD, 1989). Porém, mesmo ocorrendo eventos favoráveis à germinação,
algumas sementes podem permanecer dormentes no solo por um período e depois germinarem
(GARWOOD, 1989; ARAÚJO e FERRAZ, 2003), sendo mais difícil que o banco de
sementes persista na serrapilheira devido à maior exposição. Porém, quando a semente é
estocada numa camada profunda do solo, a germinação não ocorre (COFFIN e
LAUENROTH, 1989; FACELLI et al., 2005; WILLIAMS et al., 2005).

21
Nos desertos, os animais consumidores de sementes selecionam as áreas de forragem
de acordo com o tamanho e o teor de nutrientes das mesmas (GUO et al., 1998). Geralmente,
roedores e aves preferem sementes maiores, no entanto, formigas têm preferência pelo
carreamento de sementes pequenas (SAMSON et al., 1992; GUO et al., 1995; TRABA,
2000). Mas como na caatinga, as espécies herbáceas possuem sementes pequenas (MAMEDE
e ARAÚJO, 2008) que dominam o banco do solo (COSTA e ARAÚJO, 2003; PESSOA,
2007; LOBO, 2008; SANTOS, 2010; BARBOSA, 2008; SILVA, 2009), provavelmente as
formigas tem maior atuação na distribuição vertical.
Na distribuição vertical, a atividade de cavar pode promover a entrada de propágulos
nas camadas mais profundas do solo (CHAMBERS e MACMAHON, 1994), assim como, as
saídas de sementes armazenadas no solo para a superfície, aumentando a possibilidade de
germinação (CHAMBERS e MACMAHON, 1994).
As condições do solo, tais como o tamanho dos grãos, estrutura, textura e composição
química, também podem influenciar na distribuição vertical do armazenamento das sementes
ao facilitar ou dificultar a penetração das sementes no solo, a escavação dos vetores, a
germinação e a dormência (HARPER, 1977; SILVERTOWN, 1981; COFFIN e
LAUENROTH, 1989; CHAMBERS e MACMAHON, 1994).
As variações na topografia exercem sua influência na dinâmica do banco de sementes
ao reter as sementes e a matéria orgânica em saliências no solo formadas por trechos com
afloramentos rochosos, arenosos, declives e áreas inundáveis (CABALLERO et al., 2003;
ARAÚJO et al., 2005a; MA et al., 2006; PESSOA, 2007). Enquanto, as estruturas
topográficas mais elevadas, como o topo dos afloramentos rochosos e os aclives,
desfavorecem o acúmulo de sementes e serrapilheira (CABALLERO et al., 2003; ARAÚJO et
al., 2005a; MA et al., 2006; PESSOA, 2007).
Uma característica marcante em florestas secas é a caducifólia durante a estiagem,
momento em que a maioria das espécies lenhosas da caatinga perde as folhas (BARBOSA et
al., 1989; MACHADO et al.,1997; BARBOSA et al., 2003), aumentando o acúmulo de
serrapilheira sobre o solo durante a estação seca, do que na estação chuvosa quando a
disponibilidade de água acelera a decomposição da serrapilheira, reduzindo-a (REGINA,
2001; AUSTIN e VIVANCO, 2006; VOURLITIS et al., 2007).
Esta característica influencia a distribuição vertical, pois os propágulos geralmente
atingem primeiro a serrapilheira antes de chegarem ao solo, podendo germinar nela mesmo.

22
Além disso, o maior acúmulo durante a estação seca aumenta o espaço físico disponível para
a estocagem de sementes, podendo causar variações sazonais no banco de sementes.
Por isso, alguns estudos realizados em ambientes secos isolaram o banco de sementes
da serrapilheira e do solo, encontrando variações na riqueza e na densidade de sementes entre
a serrapilheira e o solo, no tempo e no espaço vertical (COSTA e ARAÚJO, 2003; YU et al.,
2008; SILVA, 2009; SANTOS et al., 2010).
No estudo sobre banco de sementes do solo numa área de caatinga, Costa e Araújo
(2003) registraram uma densidade total do banco de 807 sem.m-2. A densidade na
serrapilheira de 352 sem.m-2, nas amostras de 0-5cm de profundidade de 304 sem.m-2 e nas
amostras de 5-10cm de 108 sem.m-2. Constataram que nas amostras de solo de 5-10 cm de
profundidade, a germinação induzida pelo método de emergência de plântulas cessou na
quinta semana do experimento.
Pessoa (2007) analisou a influência do tipo de erosão sobre o banco de sementes,
amostrando os sulcos e entressulcos de uma encosta na caatinga que sofre erosão pelo
escoamento pluvial. Nos sulcos registrou uma média de 58.900 sementes por amostra,
enquanto nos entressulcos a média foi 82.567 sementes por amostra. A maior concentração de
sementes no espaço vertical mais alto (entressulco) do que no mais baixo (sulco) foi atribuído
ao tipo de erosão observada, que tinha um escoamento de lâmina rasa e com pouca
capacidade de transporte, o que justificou a maior densidade encontrada nos entressulcos,
demonstrando que alguns eventos podem influenciar mais do que o efeito da gravidade na
distribuição vertical do banco de sementes.
Lobo (2008) simulou as estações chuvosa e seca em laboratório para investigar a
dormência das sementes no solo de uma caatinga. A densidade nas duas estações simuladas
foi maior no solo do que na serrapilheira.
Silva (2009) investigou diferenças na riqueza e na densidade de sementes do banco
entre o solo na profundidade de 0-5 cm e na serrapilheira e entre estações climáticas de uma
área de caatinga. Durante os três anos de estudo verificou que a riqueza e a densidade de
sementes foram mais elevadas no solo do que na serrapilheira.
Mamede e Araújo (2008) estudaram os efeitos do corte e da queima da vegetação
sobre o banco de sementes numa área de caatinga. Durante a estação seca, foi realizado o
desmatamento da área de estudo e logo após coletaram amostras de solo na profundidade de
0-5 cm e de serrapilheira. Ainda na estação seca, após a indução de uma queimada, foi

23
realizada a coleta apenas do solo a 0-5 cm de profundidade, porque toda a serrapilheira foi
reduzida a cinzas. Registraram temperaturas máximas no solo letais para o tecido vegetal até 2
cm de profundidade. A queimada reduziu significativamente a densidade do banco de
sementes em todas as amostras de solo. A serrapilheira e os materiais nela estocados foram
reduzidos a cinzas, já o banco de sementes do solo isolado da serrapilheira teve uma redução
em torno 80%, havendo assim, uma maior exposição à letalidade do fogo sobre a superfície
do solo e suas camadas mais rasas. Estes resultados mostram que a riqueza e a densidade de
sementes tende a ser maior no solo do que na serrapilheira, apesar de Costa e Araújo (2003)
terem registrado resultado contrário.
Mas ainda não se sabe se a densidade, a riqueza de espécies e a composição florística
do banco de sementes do solo, também variam verticalmente no espaço em uma área de
caatinga em sucessão ecológica secundária.
2.4. Importância do banco de sementes de campos abandonados.
Um dos ecossistemas mais ameaçados do mundo são as florestas tropicais secas
(JANZEN, 1997). No Brasil, a caatinga já sofreu e vem sofrendo elevados níveis de
destruição causados principalmente pela agricultura, limitando a ocorrência de espécies de
interesse econômico a fragmentos da vegetação (MMA/SBF, 2005). Além disso,
comunidades tradicionais sertanejas praticam uma agricultura itinerante que consiste em
suprimir vegetações, plantar e posteriormente abandonar os terrenos (MAMEDE e ARAÚJO,
2008).
No entanto, estas comunidades vegetais suprimidas podem se regenerar naturalmente
apresentando uma fitofisionomia diferente da inicial (VIEIRA e SCARIOT, 2006). Estas
áreas antropizadas são chamadas de “campos abandonados”, onde pode se iniciar uma
sucessão ecológica secundária (RICKLEFS, 2009)
Neste sentido, o Centro do Sensoriamento Remoto do Instituto Brasileiro do Meio
Ambiente e dos Recursos Naturais Renováveis (CSR-IBAMA) constatou no ano de 2002 que
apenas 55,7% da área do semiárido brasileiro dominada pela fitofisionomia caatinga
encontravam-se com a cobertura vegetal nativa, o que correspondia a 443.121 km² de
vegetação. Ao realizar novo monitoramento em 2008, os dados indicaram que 16.576 km²

24
desta área foram desmatados, ou seja, 2% durante o período entre 2002 e 2008, estabelecendo
uma média de desmatamento de 2.763 km² (0,33%) ao ano (CSR/IBAMA, 2010).
Diante deste cenário de degradação do ecossistema, a política pública de gestão de
fragmentos do Ministério do Meio Ambiente brasileiro – MMA/SBF (2005) tem
recomendado a restauração ambiental do entorno dos fragmentos após interferências
antrópicas, como cultivo ou extração de madeira. A recomendação ministerial é que as áreas
com condições climáticas favoráveis ao desenvolvimento de vegetação nativa, próximas a
florestas remanescentes e em que a degradação do solo não tenha sido extrema, sejam
isoladas. A ideia é promover a restauração ambiental aproveitando os processos naturais de
sucessão vegetal, como a chuva e o banco de sementes. Este procedimento é bastante
vantajoso porque o custo de implantação pode se reduzir ao isolamento da área antropizada
(MMA/SBF, 2005).
Mitchell et al. (2000), confirmam que a restauração de florestas pode usar técnicas que
aproveitam o processo sucessional. Algumas destas técnicas são classificadas como não-
intervencionistas (MITCHELL et al., 2000) cujas características básicas são: combate a
agentes degradantes, impactos reduzidos e dependência de remanescentes próximos ou de um
banco de sementes do solo de grande longevidades.
Porém, a entrada das sementes dispersas no solo não assegura a germinação
(THOMPSON e GRIME, 1979; SIMPSON et al., 1989), pois, antes da germinação, as
sementes podem ser predadas ou estocadas em camadas profundas do solo, ou mesmo serem
afetadas por agentes patogênicos (GARWOOD, 1989; ALMEIDA-CORTEZ, 2004). Os
processos regenerativos precisam ter continuidade, renovando as comunidades vegetais
através da germinação das sementes e do estabelecimento de plântulas (GARWOOD, 1989).
Então, para entender a regeneração natural da vegetação da caatinga, é necessário
compreender as interações bióticas, abióticas e antrópicas que atuam sobre esse tipo de
vegetação. Só que, até o momento, existem poucos trabalhos sobre a influência das ações
antrópicas na dinâmica do banco de sementes na caatinga.
O banco de sementes já foi investigado em florestas secas no semiárido do Brasil, em
Pernambuco existem quatro estudos (PESSOA, 2007; LOBO, 2008; SANTOS, 2010; SILVA,
2009), no Ceará dois (COSTA e ARAÚJO, 2003; MAMEDE e ARAÚJO, 2008) e na Paraíba
um (BARBOSA, 2008), mas as ações antrópicas não foram avaliadas na vegetação. Porém, os
efeitos de ações antrópicas, como o corte e a queima, sobre o banco de sementes do solo na

25
caatinga já foram estudados (MAMEDE e ARAÚJO, 2008). Mas, aspectos sobre o papel do
banco de sementes na caatinga como a regeneração natural e o tempo de resiliência após uma
ação antrópica continuam obscuros, existindo uma grande lacuna no conhecimento quanto aos
processos de sucessão ecológica desde o abandono de uma área de cultivo até a formação de
uma floresta de caatinga.
Apesar de poucos estudos, algumas características do banco de sementes da caatinga
vêm sendo relacionados a aspectos regenerativos. Por exemplo, todos os estudos na caatinga
constataram um domínio de sementes herbáceas no banco do solo (COSTA e ARAÚJO,
2003; PESSOA, 2007; LOBO, 2008; MAMEDE e ARAÚJO, 2008; BARBOSA, 2008;
SILVA, 2009; SANTOS, 2010). E segundo Almeida-Cortez (2004), em florestas tropicais, o
predomínio de sementes de espécies herbáceas no solo pode auxiliar na regeneração natural,
pois o estrato herbáceo pode cobrir rapidamente o solo após uma perturbação protegendo-o de
processos erosivos causados pela chuva.
Bertiller e Aloia (1997) acreditam que cada espécie de semente pode apresentar
estratégias características no banco do solo, mas isso pode variar de acordo com o habitat. Por
isso, estudaram a dinâmica do banco de sementes no semiárido da Patagônia em três áreas
perturbadas por pastagens. Dentre os resultados, três tipos de estratégias do banco de
sementes foram identificados: um tipo de transição composto por gramíneas perenes, um tipo
intermediário composto por gramíneas anuais e um tipo persistente de não-graminóide perene.
As autoras concluem que as espécies mais persistentes no banco do solo são, provavelmente,
mais bem sucedidas na colonização de habitats perturbados pelo pastoreio do que os bancos
não persistentes. Fundamentadas nestas conclusões, sugerem que as estratégias de gestão e
conservação de áreas antropizadas devem ser orientadas considerando estas gramíneas
perenes.
No norte de Espanha, numa área de clima temperado sub-mediterrâneo e sub-úmido,
Luzuriaga, et al., (2005) observaram alterações na composição florística, riqueza e densidade
de sementes na distribuição vertical no banco do solo. Um campo de pastagem sofreu uma
perturbação com uma aração mecânica (uso de aradores) profunda e uma homogeneização
completa do solo de uma pastagem perene. Uma das linhas de investigação dos autores era
comparar a contribuição para a regeneração natural da chuva de sementes com o banco de
sementes, então parcelas com solo esterilizado (eliminação do banco de sementes) e não
esterilizado foram plotadas na área de estudo. Após cinco anos de abandono, o solo utilizado

26
no experimento foi recolhido para análise comparativa com a vegetação estabelecida, que
também foi amostrada. Constatou-se que, no início da sucessão secundária, o banco de
sementes possui um papel importante na regeneração natural, enquanto a chuva de sementes
assume um papel mais discreto, pois a vegetação que se estabeleceu no local teve elevada
similaridade com a composição do banco de sementes representado pelas amostras não
esterilizadas. O estudo contribuiu para entendimento da função do banco de sementes na
dinâmica da vegetação durante a sucessão secundária após uma antropização.
Reubens et al. (2007) estudaram a contribuição do banco de sementes do solo para a
regeneração de florestas tropicais secas na Etiópia e constataram que a composição florística
do banco de sementes e a vegetação estabelecida foram relacionadas em quatro florestas
preservadas e em quatro florestas antropizadas por pastagens, corte seletivo e cultivos
itinerante e permanente. Segundo os autores, a vegetação, a composição florística do banco de
sementes, as características químicas do solo e a degradação ambiental estão fortemente
relacionadas.
A diferença mais marcante entre o banco de sementes das florestas secas preservadas e
antropizadas é o número de espécies lenhosas que é maior nas florestas preservadas. Segundo
Reubens et al., (2007), o banco de sementes contribui pouco para a regeneração das florestas
antropizadas estudadas, porque o componente lenhoso foi substituído por um domínio de
espécies herbáceas, com predomínio de gramíneas.
Vieira e Scariot (2006) analisaram a regeneração natural para indicar estratégias de
manejo e restauração de florestas tropicais secas. Segundo os autores, o final da estação seca
concentra a maior abundância de sementes no solo, sendo o melhor período para realizar
coletas de serrapilheira e solo para a indução do banco de sementes em áreas degradadas.
Afirmam que incêndios precisam ser evitados, pois, além de destruir a vegetação, afetam a
densidade e a riqueza do banco de sementes do solo, comprometendo a capacidade de
resiliência das comunidades vegetais nas áreas atingidas pelo fogo.
Sendo assim, as perturbações sofridas causam mudanças no ambiente, de tal forma que
a regeneração natural nas áreas antropizadas possa ser diferenciada das áreas preservadas, que
não sofrem perturbações antrópicas (RICKFLES, 2009).
Logo, fatores que influenciam na dinâmica do banco de sementes como o solo, a
formação da camada de serrapilheira, as interações dos predadores com as sementes, dentre

27
outros, podem estar modificados também, mas não se sabe ainda quais as diferenças na
dinâmica do banco de sementes do solo entre áreas antropizadas e preservadas de caatinga.
Desta forma, a ausência de variação na riqueza e na densidade de sementes no banco
solo entre serrapilheira e solo de uma área antropizada pode indicar que os processos
ecológicos se encontram alterados. Assim como a ausência de variações no banco do solo
entre as áreas preservada e antropizada podem sugerir que a sucessão vegetal seja a mesma.
REFERÊNCIA BIBLIOGRAFIA
ALMEIDA-CORTEZ, J. S. Dispersão e banco de sementes. In: A.G. FERREIRA; F.
BORGHETTI (Orgs.). Germinação: do básico ao aplicado. Porto Alegre: Editora Artmed,
2004. p. 225-235.
ARAÚJO E. L., SILVA K. A., FERRAZ E. M. N., SAMPAIO E. V. S. B., SILVA S. I.
Diversidade de herbáceas em microhabitats rochoso, plano e ciliar em uma área de Caatinga,
Caruaru- PE. Acta Botanica Brasilica, 2005a. v. 19, p. 285-294.
ARAÚJO, E. L.; CASTRO, C. C.; ALBUQUERQUE, U. P. Dynamics of Brazilian Caatinga -
A Review Concerning the Plants, Environment and People. Functional Ecosystems and
Communities, 2007. v. 1, p. 15-28.
ARAÚJO, E. L.; FERRAZ, E. M. N. Processos ecológicos mantenedores da diversidade
vegetal na caatinga: estado atual do conhecimento. In: CLAUDINO SALES, V. (Org.)
Ecossistemas brasileiros: manejo e conservação. Fortaleza: Expressão Gráfica. 2003.
ARAÚJO, E. L.; MARTINS, F. R.; SANTOS, A. M. Establishment and death of two dry
tropical forest woody species in dry and rainy seasons in northeastern Brazil. In: Nogueira,
RJMC; Araújo, EL; Willadino, LG; Cavalcante, UMT, eds. Estresses ambientais: danos e
benefícios em plantas. Recife: MXM Gráfica e Editora, 2005b. p.76- 91.
AUSTIN, A. T.; VIVANCO, L. Plant litter decomposition in a semi-arid ecosystem
controlled by photodegradation. Nature, 2006. v. 442, n. 3, p. 555-558.

28
BAKER, H. G. Some aspects of the natural history of seed banks. In: LECK M. A.; T. V.
PARKER; R. L. A. F. SIMPSON (Eds.). Ecology of soil seed banks. London: Academic
Press, 1989. p. 9-21.
BAKKER, J. P.; POSCHLOD, P.; STRYKSTRA, R. J.; BEKKER, R. M.; THOMPSON, K.
Seed banks and seed dispersal: important topics in restoration ecology. Acta Botanica
Neerlandica, 1996. v. 45, p. 461–490.
BARBOSA, D. C. A.; ALVES, J. L. H.; PRAZERES, S. M.; PAIVA, A. M. A. Dados
fenológicos de 10 espécies arbóreas em uma área de Caatinga (Alagoinha - PE). Acta
Botanica Brasilica, 1989. v 3, p 109-117.
BARBOSA, D. C. A.; BARBOSA, M. C.; LIMA, L. C. M. Fenologia de espécies lenhosas da
caatinga. In: Leal, I.F.; Tabarelli, M.; Silva, J. M. C.; eds. Ecologia e conservação da
caatinga. Recife: Editora Universitária da UFPE, 2003. p. 657-693.
BARBOSA, F. M. Estudo do potencial de regeneração natural:uma análise da chuva de
sementes, banco de sementes e do estrato regenerante da vegetação ciliar na Bacia
Hidrográfica do Rio Taperoá, semiárido paraibano, Brasil. 95f. Tese (Doutorado em
Ecologia e Recursos Naturais) - Universidade Federal de São Carlos. 2008.
BASKIN, C.C.; BASKIN, J. M. Seeds, ecology, biogeography, and evolution of dormancy
and germination. New York: Academic Press. 1989.
BERTILLER, M. N. B.; ALOIA D. A. Seed bank strategies in Patagonian semi-arid
grasslands in relation to their management and conservation. Biodiversity and
Conservation,1997. v 6, n 4, p. 639-650.
CABALLERO, I.; OLANO, J. M.; LOIDI, J.; ESCUDERO, A. Seed bank structure along a
semi-arid gypsum gradient in central Spain. Journal of Environments, 2003. v. 55, p. 287-
299.

29
CABALLERO, I.; OLANO, J. M.; LUZURIAGA, A. L.; ESCUDERO, A. Spatial coherence
between seasonal seed banks in a semi-arid gypsum community: density changes but structure
does not. Seed Science Research. 2005, v. 15, p. 153-160.
CASTELLETTI, C. H. M. Quanto ainda resta da caatinga? Uma estimativa preliminar. In:
LEAL, I. R.; TABARELLI, M.; SILVA, J. M. C. Ecologia e Conservação da Caatinga.
Recife, Ed. Universitária da UFPE, 2003. p.719-734.
CENTRO DE SENSORIAMENTO REMOTO DO INSTITUTO BRASILEIRO DO MEIO
AMBIENTE E DOS RECURSOS NATURAIS RENOVÁVEIS (CSR/IBAMA).
Monitoramento do bioma Caatinga 2002 a 2008, monitoramento do desmatamento nos
biomas brasileiros por satélite: acordo de cooperação técnica MMA/IBAMA. Brasília,
2010. Disponível em: <http://www.mma.gov.br/portalbio>. Acesso em: 21 abr. 2010.
CHAMBERS, J. C.; MACMAHON, J. A. A day in the life of a seed: movements and fates of
seeds and their implications for natural and managed systems. Annual Review of Ecology
and Systematics, 1994. v. 25, p. 263-292.
COFFIN, P. D; LAUENROTH, K. W. Spatial and temporal variation in the seed bank of a
semiarid grassland. Journal of Botany, 1989. v. 76, p. 53-58.
COSTA, R. C.; ARAÚJO, F. S. Densidade, germinação e flora do banco de sementes do solo
no final da estação seca, em uma área de Caatinga, Quixadá, CE. Acta Botanica Brasilica,
2003. v. 17, n. 2. p. 259-264.
COSTA, R. C.; ARAÚJO, F. S.; LIMAVERDE, L. W. Flora and life-form spectrum in an
area of deciduous thorn woodland (caatinga) in northeastern, Brazil. Journal of Arid
Environments, 2007. v. 68, n. 2. p. 237–247.
FACELLI, J. M.; CHESSON, P.; BARNES, N. Differences in seed biology of annual plants
in arid lands: a key ingredient of the storage effect. Ecology, 2005. v. 86, n. 11, p. 2998-3006.

30
FLORES, S.; DEZZEO, N. Variaciones temporales en cantidad de semillas en el suelo y en
lluvia de semillas en un gradiente bosque-sabana en la gran sabana, Venezuela. Interciencia,
2005. v. 30, n. 1, p. 39-43.
GARWOOD, N.C. Tropical soil seed banks: a review. In: LECK, M.A.; PARKER, V.T.;
SIMPSON, R.L. (Ed). Ecology of soil seed banks. San Diego: Academic Press, 1989. p.149-
209.
GENTRY, A. H. Dispersal ecology and diversity in neotropical forest communities.
Sonderbaende des Naturwissenschaftlichen Vereis Hamburg, 1983. v. 7, p. 303-311.
GHERMANDI, L. Seasonal patterns in the seed bank of a grassland in north-western,
Patagonia. Journal of arid environments, 1997. v. 35, p. 215-224.
GOODALL, D. W.; CHILDS, S.; WIEBE, H. H. Methodological and Validation Study of
Seed Reserves in Desert Soils. US/IBP Desert Biome Research Memorandum 72–8. Utah
State University, Logan, 1972.
GRIZ, I.M.S., MACHADO I. C. S. Fruiting phenology and seed dispersal syndromes in
caatinga, a tropical dry forest in the Northeast of Brazil. Journal of Tropical Ecology, 2001.
v. 17. p. 303-321.
GUO, Q.; THOMPSON, D. B.; VALONE, T. J.; BROWN, J. H. The effects of vertebrate
granivores and folivores on plant community structure in the Chihuahuan Desert. Oikos,
1995. v. 72, p. 251-259.
GUO, Q; RUNDEL, P.W. GOODALL, D.W. Horizontal and vertical distribution of desert
seed banks: patterns, causes, and implications. Journal of Arid Environments . 1998, v. 38,
p. 465-478.

31
HARPER, J. L. Population biology of plants. New York, NY, USA: Academic Press. 1977,
892 p.
HOWE, H. F; SMALLWOOD, J. Ecology of seed dispersal. Annual Review of Ecology and
Systematics. Palo Alto. 1982, v. 13, p. 201-228.
JANZEN, D. H. Causes and consequences of biodiversity loss: liquidation of natural capital
and biodiversity resource development in Costa Rica. In Biodiversity and human health, eds.
F. Grifo and J. Rosenthal, Island Press. Washington, D. C. 1997, p. 302-311.
KAUFFMAN, J. B.; SANFORD JR., R. L.; CUMMINGS, D. L.; SALCEDO, I. H.;
SAMPAIO, E. V. S. B. Biomass and nutrients dynamics associated with slash fires in
neotropical dry forests. Ecology. 1993, v. 74, p.140–151.
KELLERMAN, M. J. S.; VAN ROOYEN, M. W. Seasonal variation in soil seed bank size
and species composition of selected habitat types in Maputaland, South Africa. Bothalia.
2007, v. 37, n. 2, p. 249-258.
KEMP, P. R. Seed banks and vegetation processes in deserts. In: M.A. LECK; V.T.
PARKER; R.L. SIMPSON, (Eds). Ecology of soil seed banks. New York: Academic Press.
1989, p. 257-280.
KHURANA, E; SINGH, J. S. Ecology of seed and seedling growth for conservation and
restoration of tropical dry forest: a review. Environmental conservation, 2001. v. 28. p 39-
52.
LABOURIAU, L. G. A germinação das sementes. Washington: OEA, 1983. 174p.
LIMA, E. M.; ARAÚJO, E. L.; FERRAZ, E. M. N.; SAMPAIO, E. V. S. B.; SILVA, K. A.;
PIMENTEL, R. M. M. Fenologia e dinâmica de duas populações herbáceas da Caatinga.
Revista de Geografia. 2007, v. 24, p. 124-141.

32
LOBO, P. C. A. Análise do banco de sementes em uma área de caatinga-PE após
simulação de seca. 34 f. Monografia (Graduação em biologia), Universidade de Pernambuco,
Recife, 2008.
LÓPEZ, R. P. Soil seed bank in the semi-arid Prepuna of Bolivia. Plant Ecology. 2003, v.
168, p. 85-92.
LUZURIAGA, A. L., ESCUDERO, A., OLANO, J. M., LOIDI, J. Regenerative role of seed
banks following an intense soil disturbance. Acta Oecologica, 2005. n 27. p. 57–66.
MA, J. Y.; REN J.; WANG, G.; CHEN, F. H. Influence of different microhabitats and stand
age on viable soil seed banks of sand-stabilising species. South African Journal of Botany.
2006, v. 72, p.46-50.
MACHADO, I. S.; SANTOS, L. M.; SAMPAIO, E. V. S. B. Phenology of Caatinga species
at Serra Talhada, PE, northeastern Brazil. Biotropica. 1997, v. 29, p. 57-68.
MAMEDE, M. A.; ARAÚJO F. S. Effects of slash and burn practices on a soil seed bank of
Caatinga vegetation in Northeastern Brazil. Journal of Arid Environments . 2008, v. 72, p.
458-470.
MAYOR, M. D.; BÓO, M. R.; PELÁEZ, V. D.; ELÍA, R. O. Seazonal variation of the seed
bank of Medicago minima and Erodium cicutarium as related to grazing history and presence
of shrubs in central Argentina. Journal of Arid Environments . 1999. v. 43, p. 205-212.
MAYOR, M. D.; BÓO, M. R.; PELÁEZ, V. D.; ELÍA, R. O. Seasonal variation of the soil
seed bank of grasses in central Argentina as related to grazing and shrub cover. Journal of
Arid Environments . 2003, v. 53, p. 467-477.
MINISTÉRIO DO MEIO AMBIENTE – Secretaria de biodiversidade e floresta.
Fragmentação de ecossistemas: causa e efeitos sobre a biodiversidade e recomendações

33
de políticas públicas. In: Denise Marçal Rambaldi, Daniela America Suáres de Oliveira
(Org.). Brasília: 2ª edição, 510 p. 2005.
MITCHELL, R. J.; NEEL, W. L.; HIERS, J. K.; COLE, F. T.; ATKINSON, J. B. A model
management plan for conservation easements in longleaf pine-dominated landscapes.
Newton, GA: Joseph W. Jones Ecological Research Center. 2000, 24 p.
MURPHY, P. G.; LUGO, A. E. Dry forests of Central America and Caribbean islands. In
Seasonally dry tropical forests (S.H. Bullock, H. Mooney & E. Medina, eds.). Cambridge
University Press, New York. 1995, p. 9-34.
NING, L. G. U. F.; CHANGYAN, T. Characteristics and dynamics of the soil seed bank at
the north edge of Taklimakan Desert. Science in China Series D: Earth Sciences. 2007, v.
50, sup. 1, p. 122-127.
PEREIRA, I. M.; ANDRADE, L. A.; SAMPAIO, E. V. S. B.; BARBOSA, M. R. V., Use-
history effects on structure and flora of Caatinga. Biotropica. 2003, v. 35, n. 2, p 154–165.
PESSOA, L. M. Variação espacial e sazonal do banco de sementes do solo em uma área
de Caatinga Serra Talhada, PE. 45f. Dissertação (Mestrado em Botânica) – Universidade
Federal Rural de Pernambuco, Recife, 2007.
PRICE, M. V.; JOYNER, J. W. What resources are available for desert granivores: Seed rain
or soil seed bank? Ecology. 1997, v 78, p. 764–773.
PRICE, M. V.; REICHMAN, O. J. Distribution of seeds in Sonoran desert soils – implications
for heteromyid rodent foraging. Ecology. 1987, v 68, p. 1797–1811.
RAUNKIAER, C. The life forms of plants and statiscal plant geography. London: Oxford
University Press. 1934, 620p.

34
REGINA, I. S. Litter fall, decomposition and nutrient release in three semi-arid forests of the
Duero basin, Spain. Forestry. 2001, v. 74, n. 4. p. 347-358.
REICHMAN, O. J. Spatial and temporal variation of seed distributions in Sonoran desert
soils. Journal of Biogeography. 1984, v. 11, p. 1–11.
REIS, A. M.; ARAÚJO, E. L.; FERRAZ, E. M. N.; MOURA, A. N. Inter-annual variations in
the floristic and population structure of an herbaceous community of “Caatinga” vegetation in
Pernambuco, Brazil. Revista Brasileira de Botânica. 2006, v. 29, n. 3, p. 497- 508.
REUBENS, B.; HEYN, M.; GEBREHIWOT, K.; HERMY, M.; MUYS, B. Persistent Soil
Seed Banks for Natural Rehabilitation of Dry Tropical Forests in Northern Ethiopia.
Tropicultura . 2007, v. 25, n. 4, p. 204-214.
RICKLEFS, R. E. A economia da natureza. 5.ed. Rio de Janeiro: Guanabara Koogan S.A.
2009.
ROBERTS, H. A; SIMPSON, R. L. Seed banks: general concepts and methodological issues.
In: M.A. LECK; T.V. PARKER; R.L.A.F. SIMPSON (Ed.) Ecology of soil seed banks. New
York. 1989, p. 3-7.
SAMPAIO, E. V. S. B.; SALCEDO, I. H.; KAUFFMAN, J. B. Effect of different fire
severities on coppicing of caatinga vegetation in Serra Talhada, PE, Brazil. Biotropica. 1993,
v. 25 n. 4, p. 452-460.
SAMPAIO, E. V. S. B. Overview of the Brazilian Caatinga. In: Bullock, S.; Mooney, H.A. e
Medina, E., (Ed). Seasonally dry tropical forests. Cambridge: University Press. 1995, p. 35-
58.
SAMSON, D. A.; PHILIPPI, T. E.; DAVIDSON, D. W. Granivory and competition as
determinants of annual plant diversity in the Chihuahuan Desert. Oikos. 1992, v. 165, p. 61-
80.

35
SANTOS, D. M. Variação espacial na dinâmica do banco de sementes em uma área de
caatinga em Pernambuco durante três anos consecutivos. 72 f. Dissertação (Mestrado em
Botânica) – Universidade Federal Rural de Pernambuco. Recife. 2010.
SANTOS, D. M.; SILVA, K. A.; SANTOS, J. M. F. F.; LOPES, C. G. R.; PIMENTEL,
R.M.M.; ARAÚJO, E.L. Variação espaço-temporal do banco de sementes em uma área de
floresta tropical seca (caatinga) – Pernambuco. Revista de Geografia. , v. 27, n.1, p. 234-
253.
SHEN, Y.; LIU, W.; CAO, M.; LI, Y. Seasonal variation in density and species richness of
soil seed-banks in karst forests and degraded vegetation in central Yunnan, SW China. Seed
Science Research, 2007. v. 17, p. 99–107.
SILVA, K. A. Banco de sementes (lenhosas e herbáceas) e dinâmica de quatro
populações herbáceas em uma área da caatinga em Pernambuco. 132 f. Tese (Doutorado
em Botânica) - Universidade Federal Rural de Pernambuco. Recife. 2009.
SILVERTOWN, J. W. Seed size, life span and germination date as co-adapted features of
plant life history. American Naturalist. 1981, v. 118, p. 860-864.
SIMPSON, R. L.; LECK, M. A.; PARKER, V. T. Seed banks: General concepts and
methodological issues. In Leck, M.A.; Parker, V.T. & Simpson, R.L.: Ecology of soil seed
banks. Academic Press Inc. 1989, p 3-8.
THOMPSON, K.; GRIME, J. P. Seasonal variation in the seed bank of herbaceous species in
ten contrasting habitats. Journal of Ecology. 1979, v. 67, p. 893-921.
TOWNSEND, C. R.; BEGON, M.; HARPER, J. L. Fundamentos em ecologia. 2ª Ed. Porto
Alegre: Artmed, 2006.

36
TRABA, D. J. Uso ganadero y diversidad de pastizales. Relaciones con la disponibilidad
y el movimiento de propágulos. 266f. Tese (Doutorado em Ecologia) - Universidad
Autónoma de Madrid. Madrid. 2000.
TRABA, J.; AZCÁRATE, F. M.; PECO, B. The fate of seeds in Mediterranean soil seed bank
in relation to their traits. Journal of vegetation science. 2006, v. 17, p. 5-10.
VIEIRA, D. L. M; SCARIOT, A. Principles of natural regeneration of tropical dry forests for
regeneration. Restoration ecology, 2006. v. 14. p. 11-20.
VOURLITIS, G. L.; ZORBA, G.; PASQUINI, S. C.; MUSTARD. R.. Carbon and nitrogen
storage in soil and litter of southern Californian semi-arid shrublands. Journal of Arid
Environments. 2007, v. 70, p. 164-173.
WALCK, J. L.; BASKIN, J. M., BASKIN, C. C.; HIDAYATI, S. N. Defining transient and
persistent seed banks in species with pronounced seasonal dormancy and germination
patterns. Seed Science Research. 2005, v. 15, p. 189-196.
WANG, R. Z. Demographic variation and biomass allocation of Agropyron cristatum grown
on steppe and dune sites in the Hunshandake Desert, North China. Grass and Forage
Science. 2005, v. 60, p. 99-102.
WILLIAMS, P. R.; CONGDON, R. A; GRICE, A. C; CLARKE, P. J. Germinable soil seed
banks in a tropical savanna: seasonal dynamics and effects of fire. Austral Ecology. 2005. v.
30, p. 79–90.
WILLSON, M. F.; IRVINE, A. K.; WALSH, N. G. Vertebrate dispersal syndromes in some
Australian and New Zealand plant communities, with geographic comparisons. Biotropica.
1989, v. 21, p. 133-147.

37
YU, S.; BELL, D.; STERNBERG, M.; KUTIEL, P. The effect of microhabitats on vegetation
and its relationships with seedlings and soil seed bank in a Mediterranean coastal sand dune
community. Journal of Arid Environments . 2008, v.72, p. 2040-2053.

38
Artigo a ser enviado ao periódico Journal of Arid E nvironments

39
Variação espaço-temporal do banco de sementes em uma região semiárida no Nordeste
do Brasil: paralelos entre áreas preservada e antropizada
Leonardo Brasil Mendes1*; Kleber Andrade da Silva2; Danielle Melo dos Santos1;
Ulysses Paulino de Albuquerque1; Elcida de Lima Araújo1
1. Universidade Federal Rural de Pernambuco, Departamento de Biologia, Programa de
Pós-Graduação em Ecologia, Dois Irmãos, 52171-900, Recife-PE, Brasil.
*Autor para correspondência: Tel.: +558133206308; fax: +558133206360/ Endereço de
email: [email protected]
2. Universidade Federal de Pernambuco, Centro Acadêmico de Vitória, Rua do Alto do
Reservatório s/n, Bela Vista CEP: 55608-680, Vitória de Santo Antão-PE, Brasil.

40
RESUMO
Variações sazonais podem influenciar a composição florística, a riqueza e a densidade
de sementes do banco do solo, por isso, estas características foram verificadas em um
campo abandonado durante as estações chuvosa e seca, e comparadas com o banco de
sementes de uma floresta seca remanescente mais madura investigado em outro estudo.
O campo abandonado localiza-se no semiárido do Nordeste do Brasil, era uma área de
floresta seca que foi convertida em área agrícola, mas logo após foi abandonada e vem
se regenerando há 17 anos. No campo foram coletadas 105 amostras de solo e de
serrapilheira nos finais das estações secas e chuvosas, totalizando 420 amostras. O
banco de sementes foi dominado por espécies herbáceas e a similaridade florística entre
estações foi de 57%, e entre serrapilheira e solo foi de 70%. A variação sazonal da
riqueza e da densidade de sementes do solo entre estações não foi significativa. Por
outro lado, a riqueza e a densidade de sementes foram significativamente maiores no
remanescente florestal do que no campo. Apesar de não ter havido variação sazonal nos
totais do banco do solo, isoladamente na serrapilheira foram registradas variações na
riqueza e na densidade de sementes, e isoladamente na camada rasa de solo foram
registradas variações na riqueza, sugerindo que o banco de sementes do campo não é
sazonalmente constante. As diferenças encontradas entre a serrapilheira e o solo
também vêm sendo registradas em outras florestas secas e é uma tendência que vem se
repetindo sucessivamente no remanescente florestal. Evidencia-se que em 17 anos de
regeneração natural o campo recuperou parte da composição florística, riqueza de
espécies, densidade do banco de sementes do solo, mas estas características ainda são
inferiores às registradas no remanescente florestal.
Palavras chaves: floresta tropical seca, regeneração natural, campo abandonado.

41
1. Introdução
A sazonalidade climática exerce influência sobre a riqueza e a densidade de
sementes estocadas no banco do solo (López, 2003; Williams et al., 2005; Silva, 2009),
pois em florestas tropicais sazonais, com marcante período de estiagem, a dispersão dos
propágulos não mantém a mesma intensidade durante todos os meses do ano (Garwood,
1989). Consequentemente, a riqueza e a densidade médias do banco de sementes do
solo podem flutuar entre estações climáticas e entre anos, em função dos totais de
precipitação e da forma de distribuição das chuvas de cada ano (Garwood, 1989;
Simpson et al., 1989; Almeida-Cortez, 2004; Silva, 2009; Amorim et al., 2009). Logo,
existe uma dinâmica no banco de semente do solo que deve ser minuciosamente
avaliada, devido à importância da mesma para a renovação das florestas, conservação da
diversidade vegetal dos habitats e para a estimativa de comunidades futuras que possam
se estabelecer na floresta.
Apesar da importância do banco de sementes, o conhecimento existente sobre o
mesmo é insipiente, pois a maioria dos estudos foi realizada em áreas sem histórico de
supressão de vegetação (Guo et al., 1998; Costa e Araújo, 2003; Facelli et al., 2005;
López, 2003; Williams et al., 2005; Santos et al., 2010). Estes estudos mostram que
existem áreas onde a riqueza e densidade de sementes do banco do solo podem ser
maiores na estação chuvosa (Facelli et al., 2005), maiores na estação seca (Williams et
al., 2005) e áreas onde não existe diferença entre estações climáticas (López, 2003).
Quanto ao espaço vertical, geralmente a serrapilheira abriga poucas sementes e grande
parte delas está armazenada nas camadas superficiais do solo (Guo et al., 1998). No
entanto, estudo realizado por Costa e Araújo (2003) registrou maior densidade de

42
sementes na serrapilheira do que no solo. Portanto, o banco de sementes de áreas
consideradas como preservadas apresenta marcante heterogeneidade espaço-temporal.
Na região nordeste do Brasil, a floresta tropical seca é predominante e passa por
período sazonal de estiagem de 5 a 6 meses (Sampaio, 1995; Barbosa et al., 2003;
Amorim et al., 2009). As áreas preservadas desse tipo de vegetação vêm sendo
reduzidas continuamente para o estabelecimento de atividades agrícolas, mas muitas das
áreas utilizadas na agricultura são, posteriormente, abandonadas (Vieira e Scariot, 2006;
Andrade et al., 2007).
O cerceamento da atividade antrópica permite que as áreas de cultivo se
regenerem naturalmente após abandono (Sampaio, 1993; Pereira et al., 2003; Andrade
et al., 2007; Souza, 2010; Lopes et al., 2012), demonstrando que as comunidades
vegetais possuem capacidade de recuperação natural à perturbações artificiais, mas
pouco se é conhecido sobre o papel do banco de sementes nesse processo ou se a
dinâmica do mesmo é modificada pela ação antrópica.
No semiárido do Brasil, a maioria dos estudos sobre banco de sementes de
florestas secas foi realizada em áreas não perturbadas (Costa e Araújo, 2003; Silva,
2009; Santos et al., 2010), os quais evidenciam alguns padrões: 1) a riqueza e a
densidade de sementes no banco do solo diminui com o aumento da profundidade no
solo, acompanhando a tendência de outras florestas secas do mundo (Coffin e
Lauenroth, 1989; Williams et al., 2005); 2) a serrapilheira abriga parte das sementes que
chega ao solo, mas se decompõe muito rapidamente (Santos et al., 2010), o que
também ocorre em outras regiões semiáridas (Austin e Vianco, 2006). Logo, a
densidade de sementes no solo fica mais elevada na camada superficial do mesmo
(Silva, 2009; Santos et al., 2010) e 3. Existem diferenças sazonais na riqueza de

43
espécies, na densidade de sementes e na composição florística armazenada no solo
(Silva, 2009; Santos et al., 2010).
Este estudo questiona se estes padrões são confirmados para áreas de agricultura
abandonadas. Portanto, o objetivo deste estudo é responder as seguintes perguntas: 1)
Existem diferenças sazonais na composição florística, na riqueza e na densidade de
sementes do banco do solo em uma área de agricultura abandonada? 2) Existem
diferenças na composição florística, na riqueza e na densidade de sementes do banco do
solo entre a camada mais rasa do solo e a serrapilheira? 3) As variações na riqueza de
espécies e na densidade de sementes do banco do solo podem ser explicadas pela
sazonalidade climática e pelo espaço vertical (serrapilheira e solo)? 4. As características
do banco de semente do solo (composição florística, riqueza e a densidade de sementes)
de uma área de agricultura abandonada divergem das características registradas no
banco de sementes de um fragmento de floresta circunvizinho [que nuca sofreu corte
raso e já estudado por Silva (2009)]? Espera-se que existam diferenças sazonais nas
características do banco de sementes do solo, com maior riqueza e densidade no solo na
do que serrapilheira, assim como na estação seca do que na estação chuvosa. Em adição,
que apesar de áreas antropizadas exibirem potencial de regeneração, as comunidades
vegetais futuras apresentem menor riqueza e densidade no banco de sementes em
relação a um fragmento de floresta circunvizinho, assim como ocorra diferenças na
composição de espécies.
2. Material e métodos
2.1. Área de Estudo

44
O estudo foi realizado no Instituto Agronômico de Pernambuco - IPA (Estação
Experimental José Nilson de Melo) localizado em Caruaru, Pernambuco, Brasil (8o 14’
S e 35o 55’ W, 537m de altitude), em uma área de floresta tropical seca conhecida
localmente como “caatinga”, que é drenada pelo Riacho Olaria.
O clima local é estacional com a estação seca geralmente ocorrendo entre
setembro a fevereiro enquanto a estação chuvosa se concentra nos meses restantes.
Estes fatores climáticos locais atuam de forma semelhante aos relatados por Caballero et
al. (2005), Traba et al. (2006) e Walck et al. (2005) em outros ambientes secos,
permitindo classificar as estações em favoráveis e desfavoráveis à dispersão de
sementes, à germinação e ao estabelecimento de plântulas.
Neste estudo, a disponibilidade hídrica torna a estação chuvosa favorável à
germinação e ao estabelecimento de plântulas, enquanto a estiagem severa da estação
seca inibe a germinação e seleciona as plântulas mais resistentes a ausência de água. A
precipitação média anual é 694 mm, a temperatura média anual é 22,7o C e o solo
classificado como Podzólico Amarelo Eutrófico (Alcoforado-Filho et al., 2003; Silva,
2009). Neste estudo, o total de chuva na estação chuvosa (abril a agosto de 2010) foi de
646,2 mm e na estação seca (setembro de 2009 a março 2010) foi de 166,8 mm, com
total anual de 813 mm.
Na área de estudo, inicialmente existia uma mancha de floresta tropical seca
ocupando 190 hectares até o IPA ser constituído em 1935 com a finalidade de realizar
pesquisas voltadas para agricultura e pecuária, reduzindo a área florestal a um
fragmento remanescente de 30 hectares que vem sendo mantido preservado há mais de
45 anos, que será chamado de “remanescente florestal”. O remanescente florestal exibe
dossel fechado durante a estação chuvosa, populações arbóreas de grande porte

45
(Alcoforado-Filho et al., 2003; Lima, 2011; Lopes et al., 2012) e similaridade entre a
vegetação e o banco de sementes (Silva, 2009; Santos et al., 2010).
No entorno deste fragmento, a uma distância de aproximadamente cinco metros,
localiza-se a área selecionada para este estudo, uma extensão do remanescente florestal
que sofreu corte raso (sem queima) para o plantio de Opuntia fícus-indica (L.) Mill.
(palma gigante). Durante o cultivo, não foi utilizado adubo e nem sistema de irrigação.
Há 17 anos o campo de agricultura de O. fícus-indica foi abandonado e, atualmente,
exibe uma vegetação em fase inicial de sucessão com poucos indivíduos arbóreos
adultos, dossel mal formado, vegetação rala e dominada por arbustos e herbáceas, ou
seja, com estrutura diferente do remanescente florestal (Souza, 2010; Lima, 2011;
Lopes et al., 2012). Esta área será chamada de “campo” (Lopes et al., 2012).
2.2. Amostragem do banco de sementes
No campo existe um trecho de 1 ha onde foram instalados cinco transectos
paralelos entre si e inter-espaçados em 3 m (Figura 1). Em cada transecto, foram
alocadas 21 parcelas contíguas e fixas de 10x10 m, totalizando 105 parcelas.
No final das estações chuvosa e seca de 2010 foram realizadas as coletas de solo
no centro de cada parcela de 10x10 m, em uma subparcela móvel de 20x20 cm,
considerando dois níveis de distribuição vertical das sementes: 1) serrapilheira; 2)
camada de solo de até 5 cm de profundidade. No total, foram coletadas 210 amostras
(105 de serrapilheira e 105 de solo) em cada estação (chuvosa e seca), totalizando 420
amostras.
As amostras foram levadas a casa de vegetação e acomodadas em bandejas de
isopor, irrigadas diariamente e mantidas em temperatura ambiente durante seis meses,

46
sendo as plântulas emergidas removidas após a identificação (Christoffoleti e Caetano,
1998). Para assegurar à inocorrência de entrada de sementes do meio externo a casa de
vegetação, bandejas com solo esterilizado por autoclave foram distribuídas
sistematicamente por entre as bandejas com amostras de solo do experimento. Durante o
monitoramento da emergência não houve registro de plântulas no solo esterilizado,
comprovando a ausência de contaminação.
O método de emergência de plântulas desconsidera a dormência das sementes no
solo amostrado (Baskin e Baskin, 1989; Thompson et al., 2003; Walck et al., 2005).
Desta forma, o método estimou todos os hábitos de plantas, mas pode subestimar o
quantitativo de espécies lenhosas, assim como ocorre nos trabalhos de referência sobre
banco de sementes que utilizaram este método.
A identificação das espécies foi feita através de comparações com exsicatas
acondicionadas nos herbários Prof. Vasconcelos Sobrinho (PEUFR) e Dárdano de
Andrade Lima (IPA), com auxílio de bibliografia especializada e, quando necessário, o
material foi encaminhado para especialistas. As plantas não identificadas foram
classificadas por morfoespécie. A densidade de sementes foi expressa em sementes por
metro quadrado estimada a partir do quantitativo de plântulas emergidas nas amostras
(Baskin e Baskin, 1989).
2.3. Análises do banco de sementes
No campo, semelhanças na composição florística do banco de sementes entre
estações chuvosa e seca e entre serrapilheira e solo foram calculadas utilizando-se o
índice de similaridade florística de Sorensen (Krebs, 1989). O mesmo procedimento foi
adotado para verificar as semelhanças na composição florística do banco de sementes

47
entre o campo e os dados do remanescente florestal levantados por Silva (2009). Para
facilitar a discussão, os índices foram expressos na forma de porcentagem, sendo os
valores obtidos entre as escalas 0-33%, 34-66% e 67-100%, considerados baixo, médio
e alto, respectivamente.
No campo, para verificar se: 1) As variações na riqueza de espécies e na
densidade de sementes do banco do solo podem ser explicadas pela sazonalidade
climática e pelo espaço vertical (serrapilheira e solo); 2) Existe diferença na riqueza e na
densidade média de sementes entre o solo e a serrapilheira e entre as estações chuvosa e
seca; foi realizada uma análise GLM, com teste de Tukey a posteriori (Zar, 1996). As
análises foram realizadas pelo programa Statistica 7.0. Diferenças na riqueza e na
densidade médias do banco de sementes entre o campo e o remanescente florestal foram
verificadas pelo teste “t” uma amostra (Zar, 1996). As análises foram realizadas pelo
programa BioEstat 5.0.
3. Resultados
3.1. Riqueza e composição florística do banco de sementes do campo
Um total de 47 espécies, distribuídas em 24 famílias, foi registrado no banco de
sementes. Das 47 espécies, três foram identificadas ao nível de gênero, sete ao nível de
família e três apenas como morfoespécie, devido à morte prematura das plântulas.
Excluindo as morfoespécies, 80% foram herbáceas, 4% árvores e 16% arbustos.
Somente uma planta sem semente (a pteridófita Selaginella sucata) foi registrada neste
estudo, ocorrendo durante a estação seca. Do total de espécies registradas, 23 teve
ocorrência apenas na estação seca, 11 com ocorrência apenas na estação chuvosa e 14

48
espécies comuns às duas estações. A similaridade florística entre a estação chuvosa e
seca foi classificada como média (57%). No espaço vertical, considerando as duas
estações, oito ocorreram apenas as serrapilheira, 14 apenas no solo e 26 foram comuns,
sendo considerada alta (70%) a similaridade florística entre serapilheira e solo. Apenas
as espécies Delilia biflora, Phaseolus peduncularis, Panicum trichoides, Poaceae sp1,
Poaceae sp2 e Pilea hyalina ocorreram nas duas estações climáticas, na serrapilheira e
no solo (Apêndice 1).
A riqueza média do banco de sementes foi significativamente maior no solo do
que na serrapilheira tanto na estação seca quanto na estação chuvosa. Na serrapilheira, a
riqueza média do banco de sementes registrada na estação seca foi significativamente
maior do que a registrada na estação chuvosa. No solo, a riqueza média do banco de
sementes foi significativamente maior na estação chuvosa do que na seca (Tabela 1). A
riqueza média sazonal do banco de sementes (serrapilheira + solo) foi semelhante entre
as estações chuvosa e seca. A variação na riqueza de espécies do banco de sementes foi
significativamente relacionada com a profundidade (que explicou 36% da riqueza do
banco de sementes) e a interação entre estação e profundidade (que explicou 4%)
(Tabela 2).
3.2. Densidade do banco de sementes do campo
Um total de 1277 (365 na serrapilheira e 912 no solo) e 1038 (101 na
serrapilheira e 937 no solo) plântulas emergiram do banco de sementes nas amostras das
estações seca e chuvosa, respectivamente. Estes valores permitem estimar a densidade
do banco de sementes da estação seca para 304 sem.m-2 (87 sem.m-2, na serrapilheira e

49
217 sem.m-2, no solo) e da estação chuvosa para 247 sem.m-2 (24 sem.m-2, na
serrapilheira e 223 sem.m-2, no solo). A densidade média anual foi de 276 sem.m-2.
A densidade média do banco de sementes foi significativamente maior no solo
do que na serrapilheira tanto na estação seca quanto na estação chuvosa. A densidade
média do banco de sementes registrada tanto na serrapilheira quanto no solo foi
semelhante entre as estações chuvosa e seca (Tabela 1). A densidade média sazonal do
banco de sementes (serrapilheira + solo) também foi semelhante entre as estações
chuvosa e seca. A variação na densidade do banco de sementes foi significativamente
relacionada apenas com a profundidade, que explicou 16% da densidade do banco de
sementes (Tabela 2).
3.3. Diferenças entre o banco de sementes do campo e do remanescente florestal
Considerando o conjunto florístico do banco de sementes do campo e do
remanescente florestal, 12 espécies foram registradas exclusivamente no campo, 43
apenas no remanescente florestal e 36 foram comuns às duas áreas. A similaridade
florística entre o campo e o remanescente florestal foi classificada como média (53%).
A riqueza média anual do banco de sementes foi significativamente maior no
remanescente florestal do que no campo (Tabela 3). Considerando a estação chuvosa e
seca isoladamente, apenas na estação chuvosa a riqueza média do banco de sementes foi
significativamente maior no remanescente florestal do que no campo (Tabela 3). A
densidade média de sementes no remanescente florestal foi significativamente maior do
que a do campo tanto no tempo quanto no espaço vertical (Tabela 3).
4. Discussão

50
4.1. Riqueza e composição florística do banco de sementes do campo
A riqueza de espécies registrada neste estudo (47) está dentro da faixa de
variação da riqueza (9 a 117 espécies) registrada em outros estudos sobre o banco de
sementes de florestas secas (Ning et al., 2007; Traba et al., 2006; Silva, 2009). O
elevado percentual de espécies herbáceas (80%) encontradas neste estudo é semelhante
ao registrado em outros ambientes áridos e semiáridos (López, 2003; Reubens et al.,
2007) e em outros estudos realizados em áreas de floresta seca no Brasil consideradas
como preservadas (Costa e Araújo, 2003; Silva, 2009; Santos et al., 2010).
A similaridade florística do banco de sementes do solo em florestas secas tem
variado de 22 a 53% entre estações climáticas (Ghermandi, 1997; López, 2003;
Caballero et al., 2005). Apesar de um pouco superior ao registrado em outros estudos, a
semelhança entre estações chuvosa e seca (57%) não é alta e não exclui a existência de
variação temporal na composição do banco de sementes. A similaridade florística alta
entre serrapilheira e solo (70%) indica que a maioria das espécies ocorre nos dois níveis
de distribuição espacial durante o ano, sugerindo que as sementes chegam primeiro à
serrapilheira e depois são dispersas para o solo.
No espaço vertical, a elevada riqueza do banco de sementes registrada no solo
(0-5 cm) do que na serrapilheira, tanto na estação seca quanto na estação chuvosa, é
semelhante ao registrado em outros estudos realizados em florestas secas (Yu et al.,
2008; Silva, 2009; Santos et al., 2010). A vegetação do campo possui dossel mal
formado e clareiras, o que reduz o acúmulo de serrapilheira sobre o solo e,
consequentemente, reduz também o espaço físico disponível para armazenagem,

51
tornando mais forte a tendência de uma maior variedade de sementes serem estocadas
no solo do que na serrapilheira.
Neste estudo, foi registrado que o espaço vertical (36%) e sua interação com a
sazonalidade climática (4%) foram variáveis significativas na determinação da riqueza
do banco de sementes do campo. O poder de explicação destas duas variáveis (40%)
pode ser considerado elevado, pois outras variáveis não mensuradas neste estudo (Por
exemplo: variação no espaço horizontal, predação, patógenos etc.) podem também
influenciar a riqueza de espécies do banco de sementes de ambientes secos. Em adição,
o fato de o espaço vertical ter explicado 36% da variação na riqueza de espécies mostra
que esta variável tem grande influência sobre o banco de sementes de áreas de
agricultura abandonadas e pode ser considerado como preditor da riqueza de espécies
destes ambientes.
Considerando a serrapilheira e o solo isoladamente, houve variação sazonal na
riqueza média de espécies. Enquanto na serrapilheira a riqueza foi maior na estação
seca, no solo, o oposto foi registrado. Em ambientes estacionais de vegetação seca, esta
tendência pode ser explicada pela caducifólia durante a estiagem que aumenta o
acúmulo de serrapilheira sobre o solo (Barbosa et al., 2003), aumentando,
consequentemente, a retenção de uma maior variedade de sementes na serrapilheira,
onde podem germinar sem atingir o solo (Almeida-Cortez, 2004). Durante a estação
chuvosa, a camada de serrapilheira tende a ser mais fina (Barbosa et al., 2003; Austin e
Vivanco, 2006), portanto a retenção de sementes é menor. No solo, a maior riqueza
média na estação chuvosa do que da estação seca pode ser explicada pela menor
retenção de sementes na serrapilheira (na estação chuvosa a camada de serrapilheira é
mais fina e as sementes podem cair diretamente no solo) e pela maior disponibilidade de
água, que acelera a decomposição da biomassa da serrapilheira acumulada durante a

52
estiagem nutrindo o solo e favorecendo a emergência de plântulas (Barbosa et al., 2003;
Austin e Vivanco, 2006; Almeida-Cortez, 2004)
Os estudos sobre sazonalidade climática do banco de sementes têm apontado que
a riqueza de espécies pode variar de acordo com a pluviosidade (Pugnaire e Lazaro,
2000) determinando períodos favoráveis e desaforáveis à emergência de plântulas,
podendo a riqueza do banco de sementes ser: 1) maior na época desfavorável à
emergência de plântulas (Williams et al., 2005); 2) maior na época favorável à
emergência de plântulas (Caballero et al., 2005; Kellerman e Van Rooyen, 2007); 3)
semelhante entre estas duas épocas (Silva, 2009). Desta forma, a serrapilheira
acompanhou a primeira tendência e o solo a segunda tendência encontrada em outras
florestas secas. Considerando a riqueza média de espécies em cada estação
(serrapilheira + solo) não foi registrada a variação sazonal significativa seguindo,
portanto, a terceira tendência.
Isto sugere que apenas um ano não é suficiente para encontrar um padrão de
variação sazonal para a riqueza do banco de sementes de campos de agricultura
abandonados, pois pode haver algum fator atenuador dos efeitos da sazonalidade que
ainda não foi identificado, assim como ocorre no remanescente florestal.
4.2. Densidade do banco de sementes do campo
A densidade média de sementes do banco do solo está dentro da faixa de
variação (50 a 42.000 sem.m2) registrada em outros estudos sobre o banco de sementes
de florestas secas (Pugnaire e Lazaro, 2000; Williams et al., 2005; Silva, 2009). A maior
densidade de sementes no solo do que na serrapilheira é uma tendência que vem se
repetindo no remanescente florestal há três anos (Silva, 2009; Santos et al., 2010),

53
também se repetiu após ações antrópicas em outra floresta seca no Brasil (Mamede e
Araújo, 2008), porém Costa e Araújo (2003) registraram o contrário.
Neste estudo, foi registrado que apenas o espaço vertical (16%) foi uma variável
significativa na determinação da densidade do banco de sementes do campo. O fato de o
espaço vertical ter explicado 16% da variação na densidade de sementes mostra que esta
variável também tem grande influência sobre o banco de sementes de áreas de
agricultura abandonadas e, assim como registrado para a riqueza, pode ser considerado
como preditor da densidade de sementes desses ambientes.
A maioria dos estudos realizados em florestas secas corrobora a tendência de
que, em ambientes secos, o quantitativo de sementes do banco do solo tende a ser maior
nas camadas mais rasas do solo do que em camadas mais profundas (Coffin e
Lauenroth, 1989; Williams et al., 2005). Inclusive, quando a serrapilheira é isolada das
camadas de solo, vem sendo registrado uma maior armazenagem de sementes nas
camadas mais rasas do que na serrapilheira (Yu et al., 2008; Silva, 2009; Santos et al.,
2010). Porém, em comparação com ambientes úmidos, a serrapilheira acumula pouco
em ambientes secos (Austin e Vivanco, 2006), podendo ser uma das causas para o baixo
acúmulo. A maior exposição das sementes a predadores e ao carregamento também
podem explicar o maior acúmulo das sementes nas camadas rasas do solo do que na
serrapilheira (Guo et al., 1998; Traba, 2000; Traba et al., 2006).
Em ambientes secos, a anemocoria é a principal síndrome de dispersão (Gentry,
1983; Willson et al., 1989) e a abertura do dossel provocada pela caducifólia durante a
estação seca (Barbosa et al., 2003) permite que rajadas de vento atinjam diretamente a
superfície do solo, potencializando a dispersão secundária anemocórica, uma vez que
quase não existe folhagem na vegetação para frear a velocidade do vento. Na estação
chuvosa, o dossel fechado freia a ação do vento, a disponibilidade hídrica induz a

54
germinação das sementes (Labouriau, 1983; Baskin e Baskin, 1989), a umidade
aumenta a disponibilidade de recursos, atraindo animais predadores de sementes (Guo et
al., 1998; Traba, 2000; Traba et al., 2006) e propagando patógenos, que podem conduzir
a morte fisiológica das sementes (Labouriau, 1983; Baskin e Baskin, 1989). Todos estes
eventos favorecem a saída das sementes da serrapilheira, reduzindo o estoque.
Além disso, as ações antrópicas podem reduzir o acúmulo de serrapilheira e o
quantitativo de sementes estocadas, aumentando a diferença entre o armazenamento de
sementes no solo e na serrapilheira (Mamede e Araújo, 2008). O campo também
acompanhou esta tendência, até porque o tempo em regeneração não foi suficiente para
recompor as comunidades arbóreas, colonizar todas as clareiras e fechar o dossel
(Souza, 2010; Lopes et al., 2012) reduzindo assim, a biomassa que vai formar a
serrapilheira. Consequentemente, o espaço físico reduzido para a estocagem de semente,
torna ainda mais forte esta tendência no campo.
Estudos sobre variação sazonal na densidade do banco de sementes também
apontam as três tendências citadas anteriormente para a riqueza: 1) maior densidade na
estação chuvosa (Facelli et al., 2005); 2) maior densidade na estação seca (Williams et
al., 2005); 3) ausência de diferença na densidade entre estações chuvosa e seca (López,
2003; Silva, 2009). Como não foi registrada variação sazonal significativa na densidade
de sementes tanto considerando a serrapilheira e o solo isoladamente quanto em
conjunto (serrapilheira + solo), o campo acompanhou a terceira tendência. A ausência
de variação sazonal na densidade do banco de sementes pode estar relacionada à
ocorrência sazonal de espécies e ao comportamento fenológico (Onaindia e Amezaga,
2000; Vieira e Scariot, 2006; Silva, 2009), a intensidade degradante da ação antrópica
(Vieira e Scariot, 2006; Mamede e Araújo, 2008) e a capacidade de resistência e

55
resiliência das populações vegetais às ações antrópicas (Vieira e Scariot, 2006; Reubens
et al., 2007).
4.3. Diferenças entre o banco de sementes do campo e do remanescente florestal
Estudos sobre chuva de sementes e sucessão florestal realizados no campo
apontam que o remanescente florestal atua como fonte de propágulos para a regeneração
natural do campo (Souza, 2010; Lopes et al., 2012). A semelhança média (53%) entre a
composição florística do banco de sementes do campo e do remanescente florestal
evidencia a influência do remanescente florestal sobre a formação do banco de sementes
do solo no campo. Apesar de haver influência da chuva de sementes do remanescente
florestal (Souza, 2010) e das sementes poderem resistir à perturbações antrópicas e à
estiagem (Almeida-Cortez, 2004), provavelmente, quando a agricultura no campo era
ativa, o banco de sementes era dominado artificialmente pela Opuntia fícus-indica (L.)
Mill.
Atualmente, passados 17 anos desde o abandono, o banco de sementes do campo
não apresenta mais vestígios de Opuntia fícus-indica (L.) Mill. (Apêndice 1),
apresentando: 1) a riqueza de espécies inferior ao registrado no remanescente florestal
(Tabela 3). Estudos realizados em outros ambientes secos vêm registrando que de fato,
ocorrem modificações no banco de sementes do solo após ações antrópicas (Onaindia e
Amezaga, 2000; Reubens et al., 2007; Mamede e Araújo, 2008), como por exemplo, a
redução na riqueza de espécies e mudanças na composição florística no banco do solo
(Mamede e Araújo, 2008; Vieira e Scariot, 2006), podendo inclusive reduzir uma
comunidade vegetal a uma única população resistente às perturbações (Vieira e Scariot,
2006); 2) a mesma proporção de espécies herbáceas do remanescente florestal (80%). O

56
predomínio de sementes de espécies herbáceas no solo do campo e do remanescente
florestal pode auxiliar na regeneração natural de ambos, pois o estrato herbáceo pode
cobrir rapidamente o solo após uma perturbação protegendo-o de processos erosivos
causados pela chuva e inibindo o carreamento de sementes por hidrocoria para outras
áreas (Almeida-Cortez, 2004; Reubens et al., 2007). Isto sugere que algumas estratégias
de renovação das populações vegetais já podem ser evidenciadas no campo, assim como
ocorrem no remanescente florestal.
O fato de a densidade média ter sido menor no campo do que no remanescente
florestal confirma a hipótese de que a densidade do banco de sementes de áreas que
sofreram corte raso e encontra-se em fase de sucessão inicial difere da densidade do
banco de sementes de fragmentos de floresta seca mais maduros. O solo protege as
sementes e aumenta a longevidade através de estratégias de dormência, tornando assim
o banco do solo menos sensível a eventos recentes, mesmo que a anemocoria seja
intensa. Estas características favorecem o acumulo de sementes que tende a ser maior
em florestas mais maduras (Reubens et al., 2007), podendo explicar a maior densidade
de sementes do solo no remanescente florestal do que no campo.
Relacionando os resultados obtidos no presente estudo com o estudo de Silva
(2009), verifica-se que o autor não registrou variação sazonal significativa na densidade
do banco de sementes no primeiro ano de análise no remanescente florestal, apesar de
ter registrado nos dois anos seguintes, inclusive apontando fatores atenuadores da
influência da sazonalidade. Além disso, outros estudos recentes realizados no campo
constataram que as comunidades vegetais estão se regenerando e re-estabelecendo os
processos ecológicos (Souza, 2010; Lima, 2011; Lopes et al., 2012).
Diante disso, pode-se afirmar que o banco de sementes do remanescente florestal
não é sazonalmente constante, podendo haver variações na riqueza de espécies e

57
densidade de sementes no solo. Porém, para poder afirmar que há variação sazonal na
densidade de sementes do banco do solo no campo é necessário admitir novas hipóteses,
como: 1º o campo estudado pode apresentar anos com variações sazonais na densidade
de sementes do banco do solo e anos sem essas variações, assim como ocorre no
remanescente florestal; 2º após 17 anos em regeneração, o campo apresenta fatores
atenuadores dos efeitos da sazonalidade climática sobre banco de sementes. Estas
hipóteses podem ser testadas através de estudos interanuais sobre a variação sazonal do
banco de sementes em campos abandonados.
Então, resultados deste estudo demonstram que o banco de sementes do solo no
campo abandonado exibiu potencial de regeneração estando em estagio inicial de
sucessão ecológica, mas as comunidades vegetais atuais apresentam menor riqueza e
densidade no banco de sementes do que no remanescente florestal, assim como,
ocorrem diferenças na composição florística do banco de sementes do solo entre o
campo e o remanescente florestal. Em adição, a supressão da vegetação pode modificar
as características da estrutura das comunidades do banco do solo e que 17 anos
contínuos de abandono não são suficientes para que a riqueza, a composição florística e
a densidade do banco de sementes do campo sejam semelhantes à do banco de sementes
de um fragmento de floresta seca mais maduro. Diante do exposto, o tempo de
renovação da riqueza, da composição florística e da densidade do banco de sementes
situado em áreas de floresta seca antropizadas continua sendo desconhecido.
Agradecimentos
Os autores agradecem ao Instituto Agronômico de Pesquisa Agropecuária (IPA) pela
logística e permissão de trabalhar em sua propriedade; a Universidade Federal Rural de

58
Pernambuco e ao Programa de Pós-Graduação em Ecologia; aos pesquisadores do
Laboratório de Ecologia Vegetal e Ecossistemas Nordestinos (LEVEN) pelo auxílio na
elaboração deste trabalho; ao CNPq pela bolsa de produtividade dos pesquisadores e
apoio financeiro ao projeto, processo 477239/2009-9; e à FACEPE pela bolsa de estudo
concedida.
Referência Bibliografia
Alcoforado-Filho, F.G., Sampaio, E.V.S.B., Rodal, M.J.N., 2003. Florística
fitossociologia de um remanescente de vegetação caducifólia espinhosa arbórea em
Caruaru. Acta Botânica Brasileira. 17, 287-303.
Almeida-Cortez, J.S., 2004. Dispersão e banco de sementes. In: Ferreira, A.G.,
Borghetti, F. (Orgs.). Germinação: do básico ao aplicado. Editora Artmed, Porto Alegre,
pp. 225-235.
Amorim, I.L., Samapaio, E.V.S.B., Araujo, E.L., 2009. Fenologia de Espécies Lenhosas
da caatinga do Seridó, RN. Revista Árvore 33, 491 - 499.
Andrade, L.A.; Oliveira, F.X.; Neves, C.M.L.; Félix, L.P., 2007. Análise da vegetação
sucessional em campos abandonados no Agreste Paraibano. Agrária (Recife. Online) 2,
135-142.
Austin, A.T., Vivanco, L., 2006. Plant litter decomposition in a semi-arid ecosystem
controlled by photodegradation. Nature. 442 (3), 555-558.

59
Barbosa, D.C.A, Barbosa, M.C., Lima, L.C.M., 2003. Fenologia de espécies lenhosas da
caatinga, in: Leal, I.F., Tabarelli, M., Silva, J.M.C. (Eds.), Ecologia e conservação da
caatinga. Editora Universitária da UFPE, Recife, pp. 657-693.
Baskin, C.C., Baskin, J.M., 1989. Seeds, ecology, biogeography, and evolution of
dormancy and germination, in: Leck, A.M., Parker, T.V., Simpson, L.R. (Eds.), Ecology
of soil seed banks. Academic Press, New York, pp. 53-67.
Caballero, I.; Olano, J.M.; Luzuriaga, A.L.; Escudero, A., 2005. Spatial coherence
between seasonal seed banks in a semi-arid gypsum community: density changes but
structure does not. Seed Science Research 15, 153-160.
Christoffoleti, P.J., Caetano, R.S.X., 1998. Soil seed bank. Scientia Agricola, 55, 74-78.
Coffin, P.D., Lauenroth, K.W., 1989. Spatial and temporal variation in the seed bank of
a semiarid grassland. Journal of Botany, 76, 53-58.
Costa, R.C., Araújo, F.S., 2003. Densidade, germinação e flora do banco de sementes
do solo no final da estação seca, em uma área de caatinga, Quixadá, CE. Acta Botanica
Brasilica, 17, 259-264.
Facelli, J.M, Chesson, P, Barnes, N., 2005. Differences in seed biology of annual plants
in arid lands: a key ingredient of the storage effect. Ecology 86(11), 2998-3006.

60
Garwood, N.C., 1989. Tropical soil seed banks in: Leck, M.A., Parker, T.V., Simpson.
R.L.A.F., (Eds) Ecology of soil seed banks. New York. pp. 149-209.
Gentry, A.H. 1983. Dispersal ecology and diversity in neotropical forest communities.
Sonderbaende des Naturwissenschaftlichen Vereis Hamburg 7, 303-311.
Ghermandi, L. 1997. Seasonal patterns in the seed bank of a grassland in north-western,
Patagonia. Journal of arid environments. 35, 215-224.
Guo, Q., Rundel, P.W., Goodall, D.W., 1998. Horizontal and vertical distribution of
desert seed banks: patterns, causes, and implications. Journal of Arid Environments, 38,
465-478.
Kellerman, M.J.S., Van Rooyen, M.W., 2007. Seasonal variation in soil seed bank size
and species composition of selected habitat types in Maputaland, South Africa. Bothalia
37(2): 249-258.
Krebs, C.J. 1989. Ecological methodology. Harper & Row Publishers, New York.
Labouriau, L.G. 1983. A germinação das sementes. OEA, Washington.
Lima, E.N. 2011. Influência do componente herbáceo da caatinga na regeneração
natural de plantas lenhosas em uma área de vegetação preservada e uma área de
agricultura abandonada. http://www.pgb.ufrpe.br/doctos (acessado em 12/01/2012).

61
López, R.P. 2003. Soil seed bank in the semi-arid Prepuna of Bolivia. Plant Ecology.
168, 85-92.
Lopes, C. G. R., Ferraz, E. M. N., Castro, C. C., Lima, E. N., Santos, J. M. F. F., Santos,
D. M., Araújo, E. L., 2012. Forest succession and distance from preserved patches in
the Brazilian semiarid region. Forest Ecology and Management 271, 115-123.
Mamede, M.A., Araújo F.S, 2008. Effects of slash and burn practices on a soil seed
bank of Caatinga vegetation in Northeastern Brazil. Journal of Arid Environmentsv. 72,
458-470.
Ning, L, Gu, F, Changyan, T., 2007. Characteristics and dynamics of the soil seed bank
at the north edge of Taklimakan Desert. Science in China Series D: Earth Sciences
50(1): 122-127.
Onaindia, M., Amezaga, I., 2000. Seasonal variation in the seed banks of native
woodland and coniferous plantations in Northern Spain. Forest Ecology and
Management. 126, 163-172.
Pereira, I. M., Andrade, L. A., Sampaio, E.V.S.B., Barbosa, M. R. V., 2003. Use-history
effects on structure and flora of caatinga. Biotropica, 35 (2), 154-165.
Pugnaire, F.I., Lazaró, R., 2000. Seed bank and understorey species composition in a
semi-arid environment: the effect of shrub age and rainfall. Annals of Botany 86, 807-
813.

62
Reubens B., Heyn M., Gebrehiwot K., Hermy M., Muys B, 2007. Persistent Soil Seed
Banks for Natural Rehabilitation of Dry Tropical Forests in Northern Ethiopia.
Tropicultura. 25 (4), 204-214.
Sampaio, E.V.S.B., Salcedo, I.H., Kauffman, J.B., 1993. Effect of different fire
severities on coppicing of caatinga vegetation in Serra Talhada, PE, Brazil. Biotropica.
25 (4), 452-460.
Sampaio, E.V.S.B. 1995. Overview of the Brazilian Caatinga. In: Bullock, S, Mooney,
H.A., Medina, E. (Eds). Seasonally dry tropical forests. University Press, Cambridge.
pp. 35-58.
Santos, D. M., Silva, K. A., Santos, J. M. F.F., Lopes, C.G.R., Pimentel, R.M.M.,
Araújo, E.L., 2010. Variação espaço-temporal do banco de sementes em uma área de
floresta tropical seca (caatinga) – Pernambuco. Revista de Geografia. 27 (1), 234-253.
Silva, K.A. 2009. Banco de sementes (lenhosas e herbáceas) e dinâmica de quatro
populações herbáceas em uma área da caatinga em Pernambuco.
http://www.pgb.ufrpe.br/doctos (acessado em 12/01/2012).
Simpson, R.L., Leck, M.A., Parker, V.T., 1989. Seed banks: General concepts and
methodological issues, in: Leck, M.A., Parker, V.T. & Simpson, R.L. (Eds). Ecology of
soil seed banks. Academic Press Inc, pp. 3-8.

63
Souza, J.T. 2010. Chuva de sementes em área abandonada após cultivo próximo a um
fragmento preservado de Caatinga em Pernambuco. http://www.pgb.ufrpe.br/doctos
(acessado em 12/01/2012).
Thompson, K., Ceriani, R.M., Bakker, J.P., Bekker, R.M., 2003. Are seed dormancy
and persistence in soil related? Seed Science Research, 13, 97-100.
Traba, J., Azcárate, F.M., Peco, B, 2006. The fate of seeds in Mediterranean soil seed
bank in relation to their traits. Journal of vegetation science. 17, 5-10.
Traba, D. J. 2000. Uso ganadero y diversidad de pastizales. Relaciones con la
disponibilidad y el movimiento de propágulos. http://metalib-uam.greendata.es
(acessado em 12/01/2012).
Vieira, D.L.M, Scariot, A, 2006. Principles of natural regeneration of tropical dry
forests for regeneration. Restoration ecology. 14, 11-20.
Walck, J.L., Baskin, J.M., Baskin, C.C., Hidayati, S.N, 2005. Defining transient and
persistent seed banks in species with pronounced seasonal dormancy and germination
patterns. Seed Science Research. 15, 189-196.
Williams, P.R., Congdon, R.A., Grice, A.C., Clarke, P.J., 2005. Germinable soil seed
banks in a tropical savanna: seasonal dynamics and effects of fire. Austral Ecology. 30,
79-90.

64
Willson, M.F., Irvine, A.K., Walsh, N.G, 1989. Vertebrate dispersal syndromes in some
Australian and New Zealand plant communities, with geographic comparisons.
Biotropica. 21, 133-147.
Yu, S., Bell, D., Sternberg, M., Kutiel, P., 2008. The effect of microhabitats on
vegetation and its relationships with seedlings and soil seed bank in a Mediterranean
coastal sand dune community. Journal of Arid Environments. 72, 2040-2053.
Zar, J.H. 1996. Bioestatistical analysis. 3º ed. New Jersey: Prentice Hall.
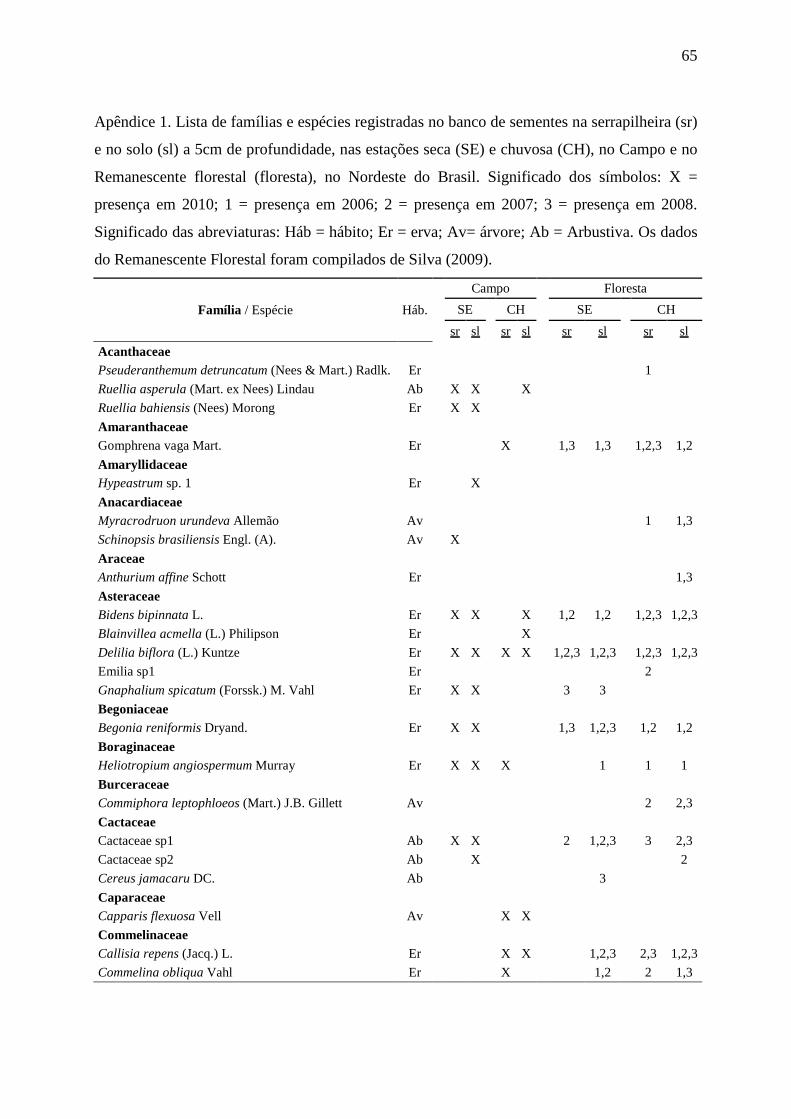
65
Apêndice 1. Lista de famílias e espécies registradas no banco de sementes na serrapilheira (sr)
e no solo (sl) a 5cm de profundidade, nas estações seca (SE) e chuvosa (CH), no Campo e no
Remanescente florestal (floresta), no Nordeste do Brasil. Significado dos símbolos: X =
presença em 2010; 1 = presença em 2006; 2 = presença em 2007; 3 = presença em 2008.
Significado das abreviaturas: Háb = hábito; Er = erva; Av= árvore; Ab = Arbustiva. Os dados
do Remanescente Florestal foram compilados de Silva (2009).
Família / Espécie Háb.
Campo Floresta
SE CH SE CH
sr sl sr sl sr sl sr sl
Acanthaceae
Pseuderanthemum detruncatum (Nees & Mart.) Radlk. Er 1
Ruellia asperula (Mart. ex Nees) Lindau Ab X X X
Ruellia bahiensis (Nees) Morong Er X X
Amaranthaceae
Gomphrena vaga Mart. Er X 1,3 1,3 1,2,3 1,2
Amaryllidaceae
Hypeastrum sp. 1 Er X
Anacardiaceae
Myracrodruon urundeva Allemão Av 1 1,3
Schinopsis brasiliensis Engl. (A). Av X
Araceae
Anthurium affine Schott Er 1,3
Asteraceae
Bidens bipinnata L. Er X X X 1,2 1,2 1,2,3 1,2,3
Blainvillea acmella (L.) Philipson Er X
Delilia biflora (L.) Kuntze Er X X X X 1,2,3 1,2,3 1,2,3 1,2,3
Emilia sp1 Er 2
Gnaphalium spicatum (Forssk.) M. Vahl Er X X 3 3
Begoniaceae
Begonia reniformis Dryand. Er X X 1,3 1,2,3 1,2 1,2
Boraginaceae
Heliotropium angiospermum Murray Er X X X 1 1 1
Burceraceae
Commiphora leptophloeos (Mart.) J.B. Gillett Av 2 2,3
Cactaceae
Cactaceae sp1 Ab X X 2 1,2,3 3 2,3
Cactaceae sp2 Ab X 2
Cereus jamacaru DC. Ab 3
Caparaceae
Capparis flexuosa Vell Av X X
Commelinaceae
Callisia repens (Jacq.) L. Er X X 1,2,3 2,3 1,2,3
Commelina obliqua Vahl Er X 1,2 2 1,3
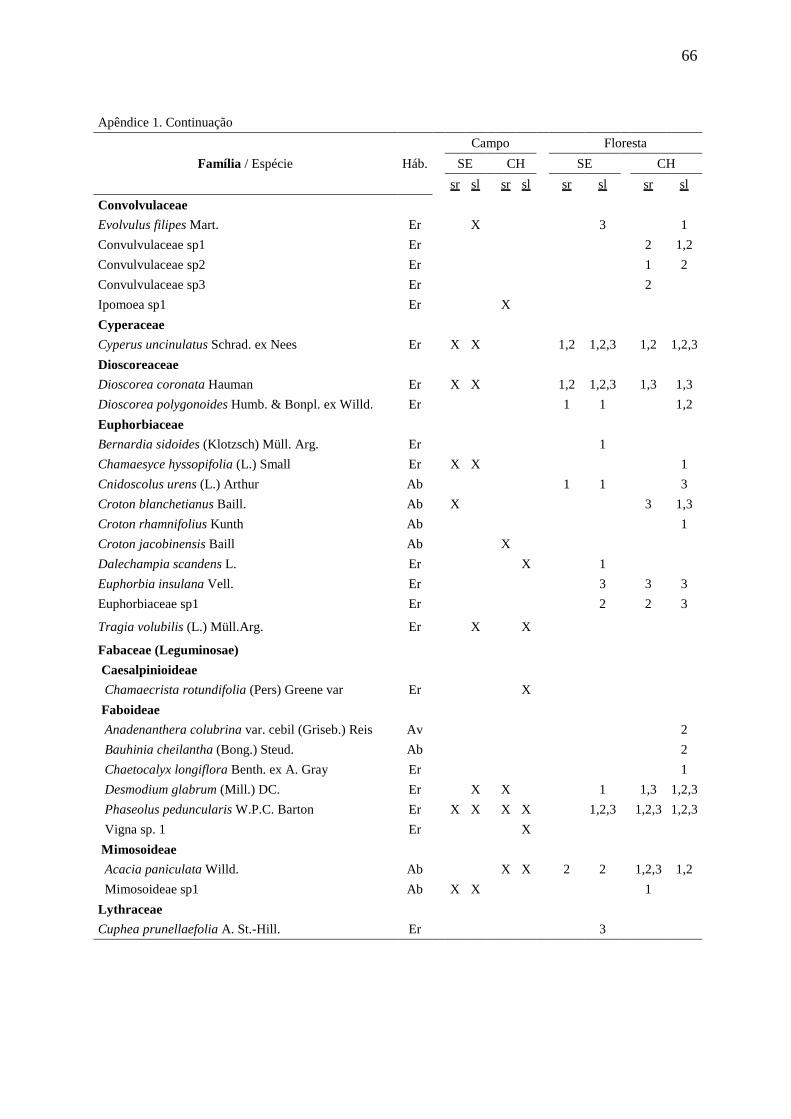
66
Apêndice 1. Continuação
Campo Floresta
Família / Espécie Háb. SE CH SE CH
sr sl sr sl sr sl sr sl
Convolvulaceae
Evolvulus filipes Mart. Er X 3 1
Convulvulaceae sp1 Er 2 1,2
Convulvulaceae sp2 Er 1 2
Convulvulaceae sp3 Er 2
Ipomoea sp1 Er X
Cyperaceae
Cyperus uncinulatus Schrad. ex Nees Er X X 1,2 1,2,3 1,2 1,2,3
Dioscoreaceae
Dioscorea coronata Hauman Er X X 1,2 1,2,3 1,3 1,3
Dioscorea polygonoides Humb. & Bonpl. ex Willd. Er 1 1 1,2
Euphorbiaceae
Bernardia sidoides (Klotzsch) Müll. Arg. Er 1
Chamaesyce hyssopifolia (L.) Small Er X X 1
Cnidoscolus urens (L.) Arthur Ab 1 1 3
Croton blanchetianus Baill. Ab X 3 1,3
Croton rhamnifolius Kunth Ab 1
Croton jacobinensis Baill Ab X
Dalechampia scandens L. Er X 1
Euphorbia insulana Vell. Er 3 3 3
Euphorbiaceae sp1 Er 2 2 3
Tragia volubilis (L.) Müll.Arg. Er X X
Fabaceae (Leguminosae)
Caesalpinioideae
Chamaecrista rotundifolia (Pers) Greene var Er X
Faboideae
Anadenanthera colubrina var. cebil (Griseb.) Reis Av 2
Bauhinia cheilantha (Bong.) Steud. Ab 2
Chaetocalyx longiflora Benth. ex A. Gray Er 1
Desmodium glabrum (Mill.) DC. Er X X 1 1,3 1,2,3
Phaseolus peduncularis W.P.C. Barton Er X X X X 1,2,3 1,2,3 1,2,3
Vigna sp. 1 Er X
Mimosoideae
Acacia paniculata Willd. Ab X X 2 2 1,2,3 1,2
Mimosoideae sp1 Ab X X 1
Lythraceae
Cuphea prunellaefolia A. St.-Hill. Er 3
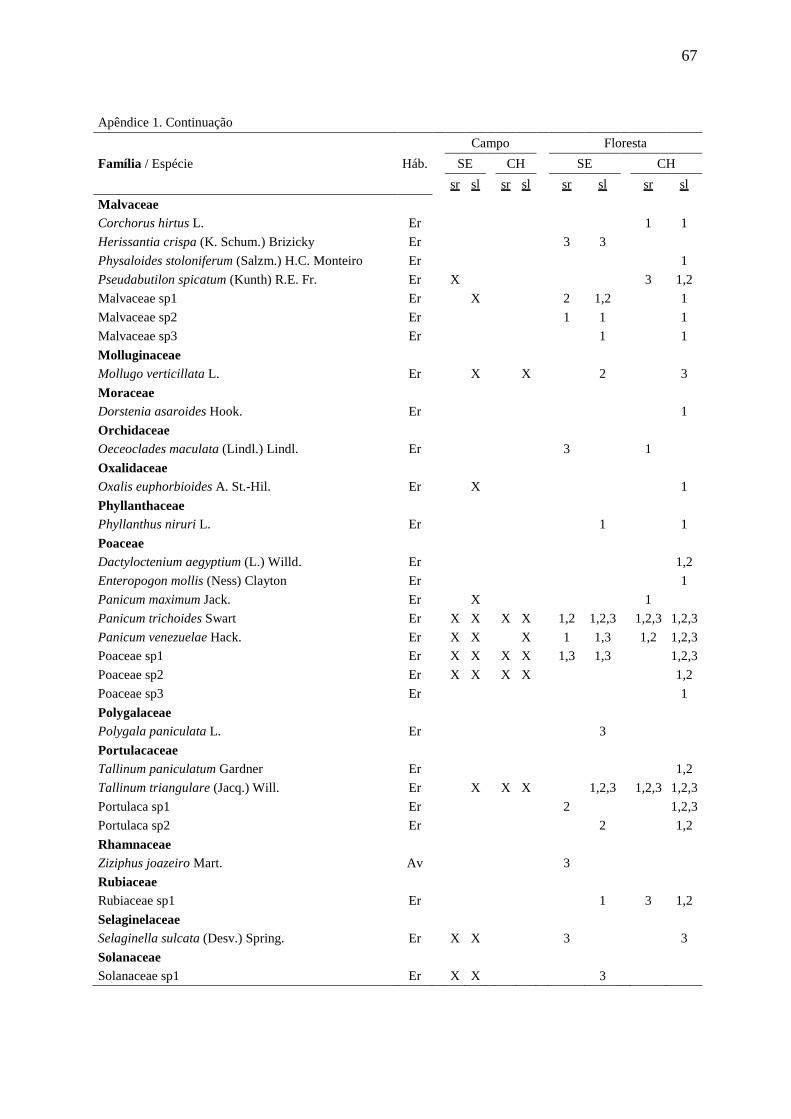
67
Apêndice 1. Continuação
Campo Floresta
Família / Espécie Háb. SE CH SE CH
sr sl sr sl sr sl sr sl
Malvaceae
Corchorus hirtus L. Er 1 1
Herissantia crispa (K. Schum.) Brizicky Er 3 3
Physaloides stoloniferum (Salzm.) H.C. Monteiro Er 1
Pseudabutilon spicatum (Kunth) R.E. Fr. Er X 3 1,2
Malvaceae sp1 Er X 2 1,2 1
Malvaceae sp2 Er 1 1 1
Malvaceae sp3 Er 1 1
Molluginaceae
Mollugo verticillata L. Er X X 2 3
Moraceae
Dorstenia asaroides Hook. Er 1
Orchidaceae
Oeceoclades maculata (Lindl.) Lindl. Er 3 1
Oxalidaceae
Oxalis euphorbioides A. St.-Hil. Er X 1
Phyllanthaceae
Phyllanthus niruri L. Er 1 1
Poaceae
Dactyloctenium aegyptium (L.) Willd. Er 1,2
Enteropogon mollis (Ness) Clayton Er 1
Panicum maximum Jack. Er X 1
Panicum trichoides Swart Er X X X X 1,2 1,2,3 1,2,3 1,2,3
Panicum venezuelae Hack. Er X X X 1 1,3 1,2 1,2,3
Poaceae sp1 Er X X X X 1,3 1,3 1,2,3
Poaceae sp2 Er X X X X 1,2
Poaceae sp3 Er 1
Polygalaceae
Polygala paniculata L. Er 3
Portulacaceae
Tallinum paniculatum Gardner Er 1,2
Tallinum triangulare (Jacq.) Will. Er X X X 1,2,3 1,2,3 1,2,3
Portulaca sp1 Er 2 1,2,3
Portulaca sp2 Er 2 1,2
Rhamnaceae
Ziziphus joazeiro Mart. Av 3
Rubiaceae
Rubiaceae sp1 Er 1 3 1,2
Selaginelaceae
Selaginella sulcata (Desv.) Spring. Er X X 3 3
Solanaceae
Solanaceae sp1 Er X X 3
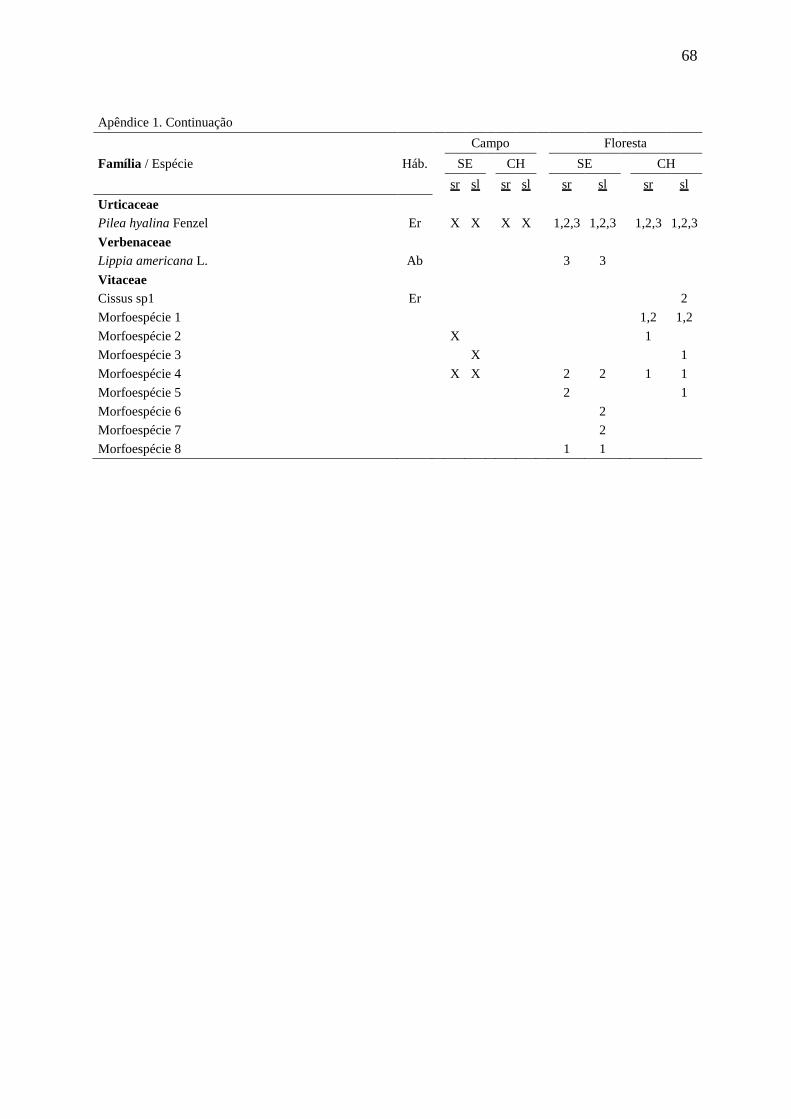
68
Apêndice 1. Continuação
Campo Floresta
Família / Espécie Háb. SE CH SE CH
sr sl sr sl sr sl sr sl
Urticaceae
Pilea hyalina Fenzel Er X X X X 1,2,3 1,2,3 1,2,3 1,2,3
Verbenaceae
Lippia americana L. Ab 3 3
Vitaceae
Cissus sp1 Er 2
Morfoespécie 1 1,2 1,2
Morfoespécie 2 X 1
Morfoespécie 3 X 1
Morfoespécie 4 X X 2 2 1 1
Morfoespécie 5 2 1
Morfoespécie 6 2
Morfoespécie 7 2
Morfoespécie 8 1 1
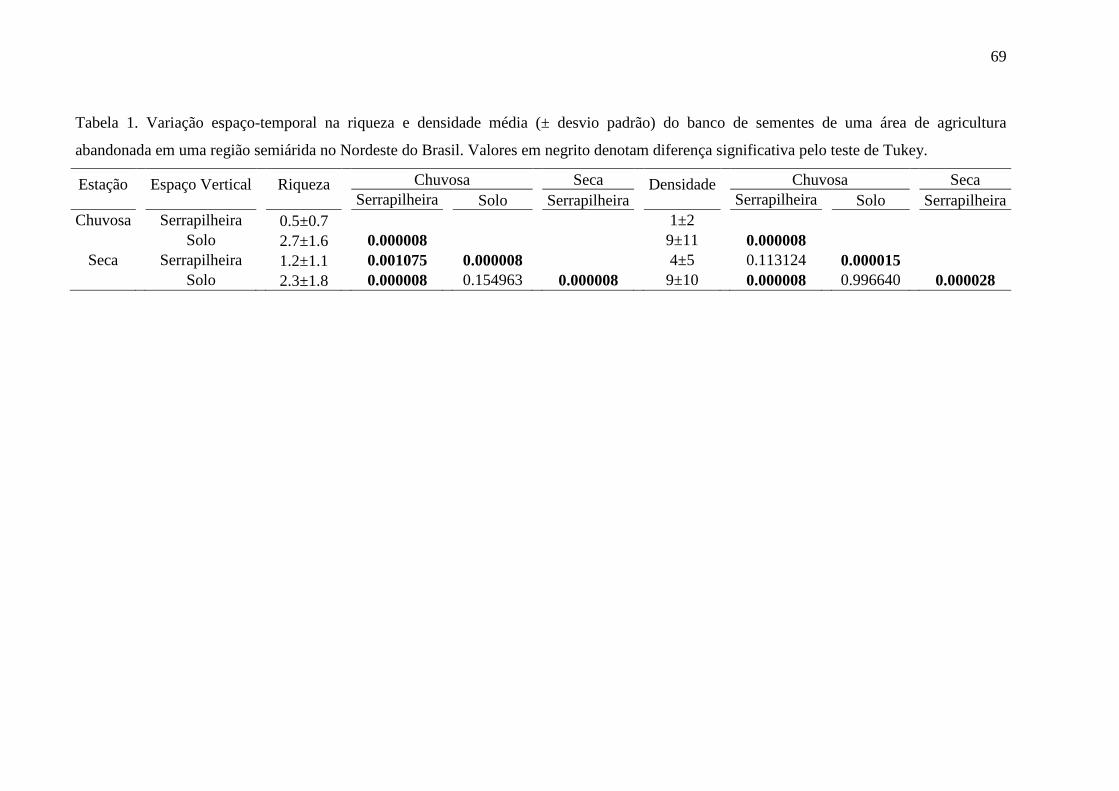
69
Tabela 1. Variação espaço-temporal na riqueza e densidade média (± desvio padrão) do banco de sementes de uma área de agricultura
abandonada em uma região semiárida no Nordeste do Brasil. Valores em negrito denotam diferença significativa pelo teste de Tukey.
Estação Espaço Vertical Riqueza Chuvosa Seca Densidade Chuvosa Seca Serrapilheira Solo Serrapilheira Serrapilheira Solo Serrapilheira
Chuvosa Serrapilheira 0.5±0.7
1±2
Solo 2.7±1.6 0.000008
9±11 0.000008
Seca Serrapilheira 1.2±1.1 0.001075 0.000008
4±5 0.113124 0.000015
Solo 2.3±1.8 0.000008 0.154963 0.000008
9±10 0.000008 0.996640 0.000028
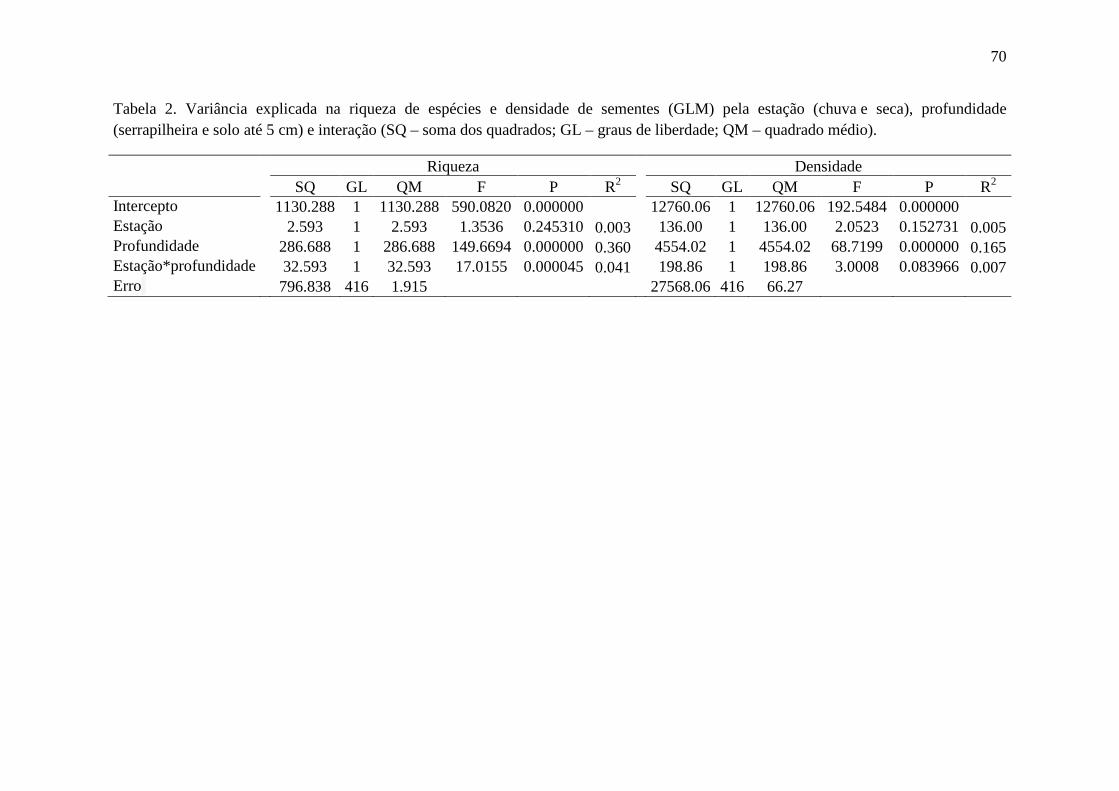
70
Tabela 2. Variância explicada na riqueza de espécies e densidade de sementes (GLM) pela estação (chuva e seca), profundidade (serrapilheira e solo até 5 cm) e interação (SQ – soma dos quadrados; GL – graus de liberdade; QM – quadrado médio).
Riqueza Densidade SQ GL QM F P R2 SQ GL QM F P R2
Intercepto 1130.288 1 1130.288 590.0820 0.000000 12760.06 1 12760.06 192.5484 0.000000 Estação 2.593 1 2.593 1.3536 0.245310 0.003 136.00 1 136.00 2.0523 0.152731 0.005 Profundidade 286.688 1 286.688 149.6694 0.000000 0.360 4554.02 1 4554.02 68.7199 0.000000 0.165 Estação*profundidade 32.593 1 32.593 17.0155 0.000045 0.041 198.86 1 198.86 3.0008 0.083966 0.007 Erro 796.838 416 1.915
27568.06 416 66.27
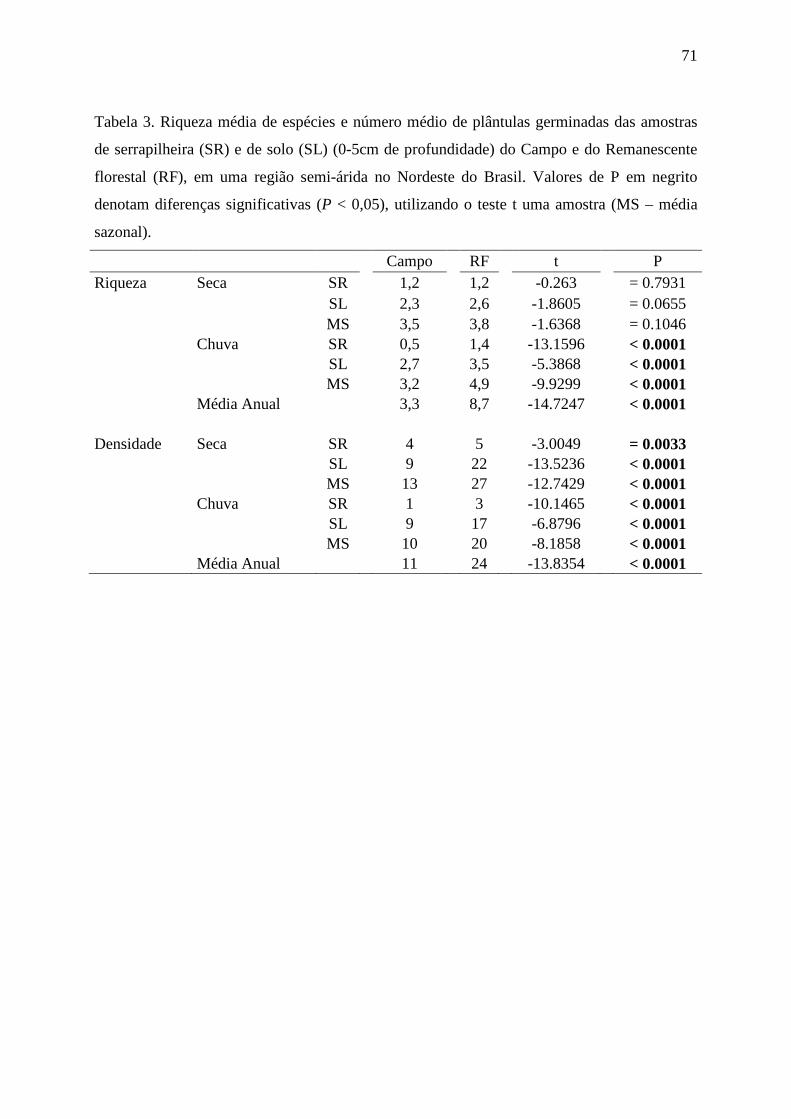
71
Tabela 3. Riqueza média de espécies e número médio de plântulas germinadas das amostras
de serrapilheira (SR) e de solo (SL) (0-5cm de profundidade) do Campo e do Remanescente
florestal (RF), em uma região semi-árida no Nordeste do Brasil. Valores de P em negrito
denotam diferenças significativas (P < 0,05), utilizando o teste t uma amostra (MS – média
sazonal).
Campo RF t P Riqueza Seca SR 1,2 1,2 -0.263 = 0.7931
SL 2,3 2,6 -1.8605 = 0.0655 MS 3,5 3,8 -1.6368 = 0.1046 Chuva SR 0,5 1,4 -13.1596 < 0.0001 SL 2,7 3,5 -5.3868 < 0.0001 MS 3,2 4,9 -9.9299 < 0.0001 Média Anual 3,3 8,7 -14.7247 < 0.0001
Densidade Seca SR 4 5 -3.0049 = 0.0033
SL 9 22 -13.5236 < 0.0001 MS 13 27 -12.7429 < 0.0001 Chuva SR 1 3 -10.1465 < 0.0001 SL 9 17 -6.8796 < 0.0001 MS 10 20 -8.1858 < 0.0001 Média Anual 11 24 -13.8354 < 0.0001
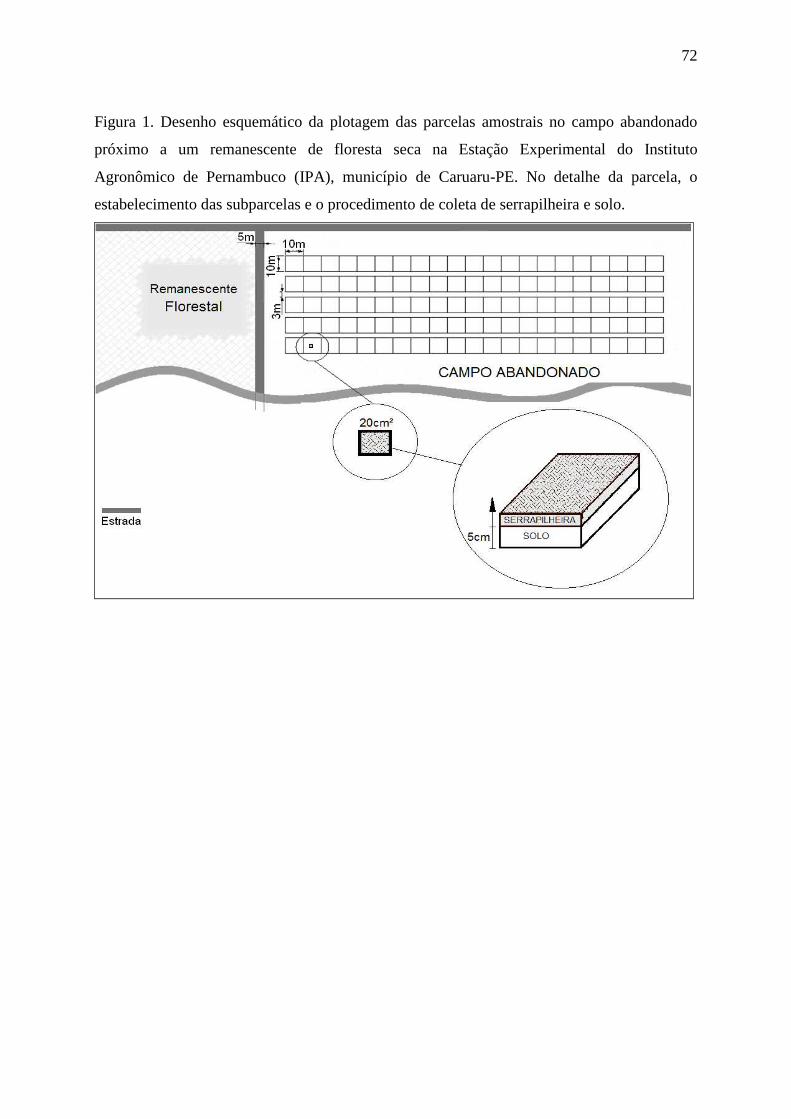
72
Figura 1. Desenho esquemático da plotagem das parcelas amostrais no campo abandonado
próximo a um remanescente de floresta seca na Estação Experimental do Instituto
Agronômico de Pernambuco (IPA), município de Caruaru-PE. No detalhe da parcela, o
estabelecimento das subparcelas e o procedimento de coleta de serrapilheira e solo.

73
ANEXOS

74

75

76

77

78

79

80

81

82

83