ultrastructural description of dientamoeba fragilis and a ... · ultrastructural description of...
TRANSCRIPT

Ultrastructural description of Dientamoeba fragilis and a new viral-like particle
By
GOURI RANI BANIK
A thesis submitted in fulfillment of the requirements for the
degree of Doctor of Philosophy
School of Medical and Molecular Biosciences
The i3 Institute,
Faculty of Science,
University of Technology, Sydney
Australia
2014

ii
Certificate of original authorship
This study was conducted in the School of Medical and Molecular Biosciences
and i3 institute, Faculty of Science, University of Technology, Sydney and in the
Microbiology Department, St. Vincent’s Hospital Sydney, under the supervision of
Professor John T. Ellis and Dr. Damien Stark.
I certify that the work in this thesis has not previously been submitted for a
degree nor has it been submitted as part of requirements for a degree except as fully
acknowledged within the text.
I also certify that the thesis has been written by me with editorial help from Prof.
John Ellis and Dr. Damien Stark as acknowledged in individual chapters. Any help that
I have received in my research work and the preparation of the thesis itself has been
acknowledged. Finally, I certify that all information sources and literatures used are
indicated in the thesis.
Gouri Rani Banik
March 2014

iii
Acknowledgements
This thesis is an outcome of my everyday work under the supervision of Prof.
John Ellis and Dr. Damien Stark. At the very beginning, I would like to gratefully
acknowledge both my supervisors for all their direction, patience, constructive
feedback, expert guidance and encouragement throughout my candidature which were
invaluable to my study. It would not be achievable without their continuous support.
Their valuable suggestions, contributions and supports are greatly appreciated.
Special thanks to Ms Debra Birch for her special guidance during my work at
Macquarie. I wish to thank her for valuable comment on relevant paper. I also would
like to thank Ms Nicole Vella and Dr Michael Johnson for their technical suggestions
during my work.
I wish to acknowledge all the team members in Professor Ellis’s lab for their
assistance in every way especially Joel Barratt, Varuni Munasinghe, Stephanie Fletcher
and Tamalee Roberts. I also thank Andrew Liew for his unconditional support all the
time. Also my other lab colleagues Heba, Jen and Atik for supporting me and make my
PhD life really enjoyable.
An individual thanks goes to Professor Steven Djordjevic and Dr. Matthew
Padula for their valuable suggestions during my protein work and Mr. Philip Lawrence,
Mr. Harry Simpson, Mr. Rowan Ikin and Dr. Ian Garthwaite for their everyday help in
the lab. Many thanks to Dr Lisa Sedger for her valuable suggestions.
I am grateful to ithree institute and University of Technology, Sydney for giving
me the opportunity to conduct my study and I do appreciate their financial support
during my study.
Last but not least my gratitude goes to my beloved husband Palash for his
everyday support and understanding whenever I was distressed. Thank you for your
never- ending faith in me. My appreciation goes to mom, dad and my brother for their
unconditional love, encouragement and endless support. My beloved daughter Anvi and
Aarna deserves a very special mention, who has sacrificed a lot so that I could achieve
my goal.

iv
Table of contents
Certificate of original authorship ........................................................... ii Acknowledgements .................................................................................. iii Table of contents ..................................................................................... iv List of Tables ......................................................................................... viii List of Figures .......................................................................................... ix Refereed publications arising from this thesis ...................................... x Conference proceedings .......................................................................... xi Abbreviations ......................................................................................... xiii Abstract ................................................................................................. xvii Chapter 1: Literature review on Dientamoeba fragilis and viruses
of parasitic protozoa .............................................................................. ..1
1.1 Literature review on Dientamoeba fragilis - introduction ....................................... 2
1.2 Taxonomy ................................................................................................................ 2
1.3 Morphology .............................................................................................................. 3
1.4 Update on life cycle and host distribution................................................................ 4
1.5 Transmission ............................................................................................................ 5
1.6 Genetic diversity ...................................................................................................... 7
1.7 Diagnostic methods .................................................................................................. 7
1.7.1 Fixative, staining and microscopy analysis ........................................................ 8
1.7.2 Different culture techniques ............................................................................... 8
1.7.3 Molecular diagnosis ........................................................................................... 9
1.8 Symptoms and treatment ........................................................................................ 10
1.9 Literature review on viruses of parasitic protozoa - introduction .......................... 11
1.10 Double-stranded RNA viruses of Trichomonas vaginalis ................................... 16

v
1.10.1 TVV, the first dsRNA virus of protozoa ........................................................ 16
1.10.2 Methods of purification and molecular identification of TVV ...................... 17
1.10.3 Trichomonas vaginalis virus sequencing and protein identification.............. 19
1.11 Viruses of Giardia lamblia .................................................................................. 21
1.11.1 Discovery of Giardiavirus ............................................................................. 21
1.12 Entamoeba histolytica virus- missing protozoan virus ........................................ 24
1.13 Virus-like RNA in Eimeria species...................................................................... 25
1.14 The RNA viruses of Leishmania .......................................................................... 28
1.15 Other viruses of protozoa ..................................................................................... 33
1.16 Virus as transfection vector .................................................................................. 34
1.17 Concluding remarks ............................................................................................. 35
1.18 Aims ..................................................................................................................... 36
Chapter 2 ................................................................................................. 38
Banik, G. R., Birch, D., Stark D., Ellis, J.T., 2011. A microscopic description
and ultrastructural characterisation of Dientamoeba fragilis: An emerging cause
of human enteric disease, International Journal for Parasitology, 42, 139-15.
Chapter 3: Electron microscopy characterisation of
Dientamoeba fragilis virus-like particles .............................................. 39
3.1 Introduction ............................................................................................................ 42
3.2 Materials and methods ........................................................................................... 43
3.2.1 Culture of Dientamoeba fragilis trophozoites.................................................. 43
3.2.2 Culture of Trichomonas vaginalis .................................................................... 44
3.2.3 Transmission electron microscopy ................................................................... 44

vi
3.2.4 Negative staining .............................................................................................. 44
3.2.5 VLPs purification ............................................................................................. 45
3.2.6 Isolation of dsRNA with phenol pH 8.0........................................................... 45
3.2.7 DNase and RNase sensitivity test .................................................................... 46
3.2.8 Acridine orange staining .................................................................................. 46
3.2.9 ImageJ analysis ................................................................................................ 47
3.3 Results .................................................................................................................... 47
3.3.1 Characterisation of VLPs ................................................................................. 47
3.3.2 Maturation pathway of D. fragilis virus-like particles ..................................... 50
3.3.3 Presence of VLPs in Trichomonas culture ....................................................... 57
3.3.4 Investigations on D. fragilis VLPs genome ..................................................... 57
3.4 Discussion .............................................................................................................. 60
Chapter 4: Evaluation of different extraction methods to
identify Dientamoeba fragilis viral nucleic acid .................................. 69
4.1 Introduction ............................................................................................................ 70
4.2 Materials and methods ........................................................................................... 71
4.2.1 Culture of Dientamoeba fragilis trophozoites.................................................. 71
4.2.2 Culture of Trichomonas vaginalis .................................................................... 72
4.2.3 Purification of Trichomonas virus ................................................................... 72
4.2.4 Dientamoeba fragilis cell storage .................................................................... 73
4.2.5 Extraction methods .......................................................................................... 73
4.2.5.1 Method 1 (isolation of dsRNA with phenol 8.0) .................................... 73
4.2.5.2 Method 2 (isolation of total nucleic acids) .............................................. 74
4.2.5.3 Method 3 (viral particle purification by CsCl density gradients and
extraction of RNA) .................................................................................. 75

vii
4.2.6 DNase and RNase sensitivity test .................................................................... 76
4.2.7 Rotavirus, adenovirus and norovirus testing .................................................... 76
4.3 Results .................................................................................................................... 77
4.4 Discussion .............................................................................................................. 82
Chapter 5 ................................................................................................. 85
Banik, G. R., Barratt, J. L. N., Marriott, D., Harkness J., Ellis, J. T. and Stark, D.,
2011. A case-controlled study of Dientamoeba fragilis infections in
children, Parasitology, 138, 819-823.
Chapter 6 ................................................................................................. 86
General discussion and future directions ..................................................................... 87
Chapter 7 ................................................................................................. 95
References .................................................................................................................... 96

viii
List of tables
Table 1.1 Comparison of several characteristics of protozoan viruses ....................... 12
Table 4.1 Trichomonas viral RNA isolated using three different extraction methods
...................................................................................................................... 78

ix
List of figures
Figure 1.1 Dientamoeba fragilis trophozoite stained with modified
iron-haematoxylin stain ................................................................................ 4
Figure 1.2 Life cycle of Dientamoeba fragilis………………………………………..5
Figure 1.3 Schematic diagram showing the replication strategy of double-stranded
RNA virus………………………………………………………………..14
Figure 1.4 Electron micrograph showing the appearance of VLPs from different
protozoan parasites ..................................................................................... 32
Figure 3.1 Transmission electron microscopy showing the presence of virus-like
particles in the perinuclear region of Dientamoeba fragilis
trophozoite................................................................................................. 48
Figure 3.2 Electron micrograph of the purified virus-like particles ............................ 49
Figure 3.3 The maturation pathway of VLPs in Dientamoeba fragilis ....................... 52
Figure 3.4 Gel electrophoresis of total nucleic acids from Trichomonas
vaginalis and Dientamoea fragilis trophozoites ......................................... 58
Figure 3.5 Acridine orange fluorescent staining of Dientamoeba fragilis trophozoite
(A-D) (isolate 2) and Trichomonas vaginalis B7268 isolate (E-H) ........... 59
Figure 3.6 Working model for Dientamoeba fragilis VLPs development .................. 67
Figure 4.1 Comparison of three different extraction methods to identify viral nucleic
acid from Trichomonas vaginalis trophozoites (B7268 isolate) ................ 79
Figure 4.2 Comparison of agarose gel electrophoresis of total nucleic acids isolated
from purified viral particles from Trichomonas vaginalis and
Dientamoeba fragilis growth media .......................................................... 81

x
Refereed publications arising from this thesis
1. Barratt, J.L., Banik, G. R., Harkness, J., Marriott, D., Ellis, J.T and Stark, D., 2010.
Newly defined conditions for the in vitro cultivation and cryopreservation of
Dientamoeba fragilis: new techniques set to fast track molecular studies on this
organism. Parasitology, 137: 1867-1878.
http://www.ncbi.nlm.nih.gov/pubmed/20609278
2. Banik, G. R., Barratt. J. L.N., Marriott, D., Harkness, J., Ellis, J. T., and Stark, D.,
2011. A case-controlled study of Dientamoeba fragilis infections in children,
Parasitology, 138: 819-823. http://www.ncbi.nlm.nih.gov/pubmed/21524324
3. Banik, G. R., Birch , D., Stark, D., Ellis, J. T., 2012. A microscopic description and
ultrastructural characterisation of Dientamoeba fragilis: An emerging cause of human
enteric disease. International Journal for Parasitology, 42: 139-153.
http://www.sciencedirect.com/science/article/pii/S0020751911002785
4. Banik, G. R., Birch, D., Stark, D., Ellis, J. T., 2013. Virus-like particles (VLPs) in
Dientamoeba fragilis: an ultrastructural study (Submitted for publication in the Journal
of Parasitology, November 2013).

xi
Conference proceedings
■ Banik, G. R., Barratt. J. L.N., Marriott, D., Harkness, J., Ellis, J. T., and Stark, D. A
case-controlled study of Dientamoeba fragilis infections in children, Poster
presentation, ICOPA, Melbourne, Australia, 15 -19th August, 2010.
■ Banik, G. R., Barratt. J. L.N., Marriott, D., Harkness, J., Ellis, J. T., and Stark, D. A
case-controlled study of Dientamoeba fragilis infections in children, Poster
presentation, 27th RNSH.UTS.USYD. Kolling Scientific Reserach Meeting, Sydney,
Australia, 9-10th November, 2010.
■ Banik, G. R., Birch, D., Stark, D., and Ellis, J. T. A microscopic description and
ultrastructural characterisation of Dientamoeba fragilis: An emerging cause of human
enteric diseases. Poster presentation, 28th RNSH.UTS.USYD. Kolling Scientific
Reserach Meeting, Sydney, Australia, 1-2nd November, 2011.
■ Banik, G. R., Birch, D., Stark, D., and Ellis, J. T. Electron microscopy
characterisation of Dientamoeba fragilis virus life cycle. Oral presentation, Australian
Society for Parasitology Annual Conference, Launceston, Tasmania, 2-5th July, 2012.
■ Banik, G. R., Birch, D., Stark, D., and Ellis, J. T. Electron microscopy
characterisation of Dientamoeba fragilis virus-like particles, Poster presentation, 29th
RNSH.UTS.USYD. Kolling Scientific Reserach Meeting, Sydney, Australia, 20 -21st
November, 2012.
■ Banik, G. R., Birch, D., Stark, D., and Ellis, J. T. Electron microscopy
characterisation of Dientamoeba fragilis virus-like particles, Oral and Poster
presentation, Gordon Research Seminar and Conference on Physical Virology,
Ventura, California, USA, 19-25th January, 2013.

xii
■ Banik, G. R., Birch, D., Stark, D., and Ellis, J. T. Electron microscopy
characterisation of Dientamoeba fragilis virus-like particles, Poster presentation, New
Horizons 2013, 30th Combined Health Science Conference, Kolling Building, Royal
North Shore Hospital, NSW, 18 -20th November, 2013.

xiii
Abbreviations Terms: Ax Axostyle
ATCC American Type Culture Collection
BB Basal Body
cDNA Complementary Deoxyribonucleic Acid Ch Chromatin Bodies
Co Costa
CP Capsid Protein
CsCl Caesium Chloride
DAPI 4', 6-diamidino-2-phenylindole
DFV Dientamoeba fragilis Virus
DNA Deoxyribonucleic Acid DNase Deoxyribonuclease
DIC Differential Interference Contrast
dsRNA Double Stranded Ribonucleic Acid
Dv Digestive Vacuole
ED Electron Dense
EDTA Ethylenediaminetetraacetic Acid
EGTA Ethylene Glycol Tetraacetic Acid
EM Electron Microscopy
ENV Eimeria necatrix Virus
ER Endoplasmic Reticulum
ESV Eimeria stiedae Virus
EtOH Ethyl Alcohol

xiv
Gc Golgi Complex
GFP Green Fluorescent Protein
GLV Giardia lamblia Virus
HCl Hydrochloric Acid
HIV Human Immunodeficiency Virus
ITS Internal Transcribed Spacer
LRV Leishmania RNA virus
MgCl2 Magnesium Chloride
My Myelin Sheath
Mt Microtubules
MTOC Microtubule Organizing Center
Nm Nuclear Membrane
NaCl Sodium Chloride
Np Nuclear Pore
ORF Open Reading Frame
OsO4 Osmium Tetroxide
PBS Phosphate Buffered Saline
PCR Polymerase Chain Reaction
PEG Polyethylene Glycol
Pf Parabasal Filament
Pm Plasmalemma
Ps Pseudopodia
PVA Polyvinyl Alcohol
RNA Ribonucleic Acid
RNase Ribonuclease

xv
RdRp RNA Dependent RNA Polymerase
Rs Rice Starch
RT-PCR Real-time Polymerase Chain Reaction
ssRNA Single Stranded Ribonucleic Acid
SAF Sodium Acetate -Acetic Acid- Formalin
ScV Saccharomyces cerevisiae Virus (ScV)
S.D. Standard Deviation
SDS Sodium Dodecyl Sulfate
SEM Scanning Electron Microscopy
ssRNA Single-Stranded Ribonucleic Acid
SSU rRNA Small Subunit Ribosomal RNA
sv Small Vacuole
TBE Tris Borate EDTA
TE Tris-EDTA
TEM Transmission Electron Microscopy
TM Tris- MgCl2
tRNA Transfer RNA
TVV Trichomonas vaginalis Virus
UTR Untranslated Region
VLP Virus-Like Particle
WSSV White Spot Shrimp Viruses
Units:
°C Degree Celsius
g Relative Centrifugal Force

xvi
h Hour
Kb Kilobase
KDa Kilo Daltons
Kg kilogram
M Molar
Mb Megabase
μM Micromolar
μm Micrometre
μg Microgram
μL Microlitre
mg Milligram
mL Millilitre
mM Millimolar
min Minute
ng Nanogram
nm Nanometer
U Unit

xvii
Abstract
Dientamoeba fragilis is a trichomonad protozoan found in the gastrointestinal
tract of humans and is implicated as a cause of diarrhoeal disease. Despite its
widespread occurrence and associated symptoms, very little is known about the biology
and pathogenicity of D. fragilis. Advances in cell biology of other trichomonads means
there is a need to advance knowledge on this neglected protozoan.
In this study, the morphological characteristics and ultrastructure of D. fragilis
were described in detail using different microscopy techniques. Scanning electron
microscopy, transmission electron microscopy, confocal and light microscopy were
used to characterise D. fragilis populations growing in xenic culture. Under the SEM,
two types of D. fragilis populations were identified based on cell surface structure:
smooth cells and ruffled cells. Typically D. fragilis has a spherical or oval shape with a
granular, vacuolated cytoplasm and some cells are amoeboid. Dientamoeba fragilis
exhibited different motile forms with visible pseudopodia. The organelles in D. fragilis
were analysed by transmission electron microscopy; the pelta, flagella, undulating
membrane or pseudocyst-like forms were not found. The presence of hydrogenosomes
in D. fragilis is described which has not been previously reported. The majority of cells
grown in culture were mononucleate while most cells in permanent stained faecal
smears were binucleate. Evidence is presented using confocal microscopy that the two
nuclei of D. fragilis are identical in DNA content. In addition, the discovery of a virus-
like particle is reported for the first time in D. fragilis. This study provides extensive
and new detail on the ultrastructure of D. fragilis that is an emerging cause of human
enteric disease.
Dientamoeba fragilis virus (virus-like particles or VLPs) was studied further: it
was approximately 33-40 nm in size and the most common shape was spherical. These
VLPs contain an inner dark core surrounded by an electron-lucent layer and an electron
dense capsid coat. Virus particles are found extensively in the perinuclear region of the
trophozoite, and especially around microtubules and in association with the Golgi
complex. Virus particles were also found in the vicinity of endoplasmic reticulum,
axostyle, and near to the parabasal filament but no VLPs were found in the nucleus.

xviii
Dientamoeba fragilis VLPs were also detectable in dying trophozoites present in in
vitro cultures. Whether viral load contributes to cell death is not yet known.
The identity of the D. fragilis viral genome was also studied. Several different
extraction methods were screened and three different methods were optimized to
identify dsRNA from Trichomonas vaginalis (B7268 isolate) which was used as a
positive control for the isolation of viral dsRNA. These optimized methods were
evaluated to identify D. fragilis viral genome. No viral RNA or dsRNA was isolated
from D. fragilis suggesting that unlike T. vaginalis, D. fragilis trophozoites do not
contain a dsRNA virus, or that the abundance of the virus was so low that it prevented
the identification of viral nucleic acid.
The epidemiology of D. fragilis has not been studied in detail and as a small side
project I investigated hospital records for infections of children. Consequently, a case-
controlled study was conducted to document the extent of D. fragilis infections in
children presenting to a major Sydney Hospital. Treatment options are also discussed.
In total, hospital data from 41 children were included in the study along with a control
group. Results showed that diarrhoea (71%) was found to be the most common
symptom followed by abdominal pain (29%). In addition, diarrhoea was statistically
more significant in children with D. fragilis infection compared to a control group. In
this study, the most common antimicrobial used for treatment was metronidazole
(n=41), with complete resolution of symptoms and clearance of parasite occurring in
85% of cases. Moreover, a treatment failure rate of 15% was identified in children
treated with metronidazole. These studies further suggest the pathogenic nature of D.
fragilis and it is recommended that all laboratories must routinely test for D. fragilis as
treatment which eliminates the parasite usually results in the resolution of symptoms.
In summary, this thesis has discussed many novel aspects on the biology of D.
fragilis and provide new knowledge on the cell biology of this protozoan and a new
protozoan virus.

1
Chapter 1: Literature review on Dientamoeba fragilis and
viruses of parasitic protozoa

2
1.1 Literature review on Dientamoeba fragilis- Introduction
Dientamoeba fragilis is a protozoan parasite commonly found in the
gastrointestinal tract of humans (Clark et al. 2014). It is associated with gastrointestinal
illness, mainly diarrhoea and abdominal pain in humans (Stark et al. 2010b; Barratt et
al. 2011a; Mumcuoğlu et al. 2013). Dientamoeba fragilis was first reported almost a
century ago but very little is known about it (Stark et al. 2006). It is now recognized as
a trichomonad within the Phylum Parabasalia (Barratt et al. 2011a; Banik et al. 2012).
Dientamoeba fragilis has been reported in throughout the world with higher prevalence
in developed countries (Stark et al. 2010b; Nagata et al. 2012a). Several studies
indicated that the incidence of this organism varies widely ranging from 5.2% to 52%
(Crotti et al. 2005; Stark et al. 2010b).
Over the last few decades, studies have been well conducted on its clinical
presentation though research on molecular, genetic and proteomic aspects are limited.
This review provides an up-to-date overview of this pathogenic parasite with more
emphasis on its morphology, life cycle, different diagnosis techniques and treatment of
the disease as these areas are related directly to my research.
1.2 Taxonomy This parasite has a long history of taxonomic reclassification (Hopkins 2006).
Since the discovery of this parasite, a number of studies have reported that the
predominant form of this parasite is binucleate (Jepps & Dobell 1918; Yang & Scholten
1977; Johnson et al. 2004). Initially, D. fragilis was included into subphylum Sarcodina
(Johnson et al. 2004). Camp et al. (1974) analysed the ultrastructure of D. fragilis by
Transmission Electron Microscopy (TEM) for the first time and indicated the similarity
with trichomonads (Stark et al. 2006).
Studies also analysed the complete SSU rDNA sequences of D. fragilis
comparing with several trichomonads and other eukaryotes (Silberman et al. 1996).
Delgado-Viscogliosi et al. (2000) showed that D. fragilis is closely related to
Histomonas meleagridis - an amoeboflagellate which was accepted as a trichomonad
flagellate. Subsequent studies have also provided further evidence based on sequence

3
analysis of SSU rDNA and confirmed that it is related to trichomonads (Gerbod et al.
2002; Ohkuma et al. 2005). Molecular phylogenetics also confirmed that D. fragilis
clustered with the trichomonads, but it lacks flagella (Gerbod et al. 2004; Kleina et al.
2004; Ohkuma et al. 2005; Lagacé-Wiens PR et al. 2006).
The current classification of Dientamoeba fragilis
(Source: http://en.wikipedia.org/wiki/Dientamoeba_fragilis)
Kingdom: Excavata Phylum: Metamonada Class: Parabasalia Order: Trichomonadida Family: Monocercomonadidae Genus: Dientamoeba Species: Dientamoeba fragilis
1.3 Morphology
Dientamoeba fragilis is a single-celled pleomorphic trophozoite, ranging from 5
to 15 μm in diameter (Stark et al. 2006). Light microscopic studies showed that D.
fragilis may contain one to four nuclei (Banik et al. 2012). In stained smears, a high
percentage of cells are binucleate and each nucleus has a large, fragmented, central
karyosome (Figure 1.1) (Sawangjaroen et al. 1993; Stark et al. 2006). However, the
nuclear structure is invisible in an unstained preparation (Stark et al. 2008). Banik et al.
(2012) reported that most of the cells grown in in vitro culture conditions were observed
as mononucleate compared with permanent stained faecal smears. The author also
suggested that the trophozoite from faecal samples do not present true morphology as
most of the cells are not viable in stained smears (Banik et al. 2012).
Although D. fragilis was first described a long time ago, there have been no
microscopic observations in the last 40 years time. Recently, the ultrastructures and
surface organizations of four isolates of D. fragilis trophozoite were described in details
by electron microscopy (Banik et al. 2012). In this study, the structure of
hydrogenosome and the basal body cytoskeleton including axostyle, costa were
observed in D. fragilis trophozoite for the first time (Banik et al. 2012). Another
concurrent study reported new cyst stages in D. fragilis and flagella components
observed only within the cyst (Munasinghe et al. 2013).

4
Figure 1.1 Dientamoeba fragilis trophozoite stained with a modified iron–haematoxylin
stain. Magnification in (A) =100 x. ( Unpublished data).
However, no motility was recovered if the cells were refrigerated. Some studies
showed that D. fragilis will be preserved and successfully revived after
cryopreservation maintaining the similar morphology (Sawangjaroen et al. 1993;
Barratt et al. 2010).
1.4 Update on life cycle and host distribution Although cyst stages of D. fragilis have been identified recently, the complete
life cycle of this parasite is yet to be definitely defined (Figure 1.2) (Clark et al. 2014).
There are few studies which investigated the host distribution as well as zoonotic
potential of D. fragilis (Johnson et al. 2004). A significant update for D. fragilis life
cycle and host distribution was reported by Stark et al. (2008). Dientamoeba fragilis has
a limited host range and humans are probably the definitive host of this parasite (Barratt
et al. 2011b; Cacciò et al. 2012).
Moreover, D. fragilis was reported in non-human primates such as pigs (Cacciò
et al. 2012), gorillas (Stark et al. 2008), swine (Crotti & D'Annibale 2007), baboons
(Myers & Kuntz 1968), sheep (Noble & Noble 1952) and macaques (Dobell 1940). All
of these reports were based on light microscopic analysis which needs to be confirmed
by molecular methods. More knowledge of the life cycle and host distributions of D.
fragilis would need analysis which leads to developing an animal model for
understanding of dientamoebiasis.

5
Figure 1.2 Life cycle of Dientamoeba fragilis.
(Source:
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=8242214
Barratt et al. 2011b).
(1) Humans are the most common host of Dientamoeba fragilis, although other primate
species such as gorillas and pigs may be the probable host (2) Dientamoeba moves to
the large intestine and divides by binary fission (3) Dientamoeba come into the
environment in the faeces (4) they infect food and water sources (5) the true role of
Enterobius vermicularis in the life cycle of D. fragilis is yet to be definitely defined.
1.5 Transmission
Even though many authors have proposed different opinions, the mode of
transmission of D. fragilis is still unknown (Clark et al. 2014). It is believed that the
transmission occurs via faecal-oral route (Barratt et al. 2011b). Most of the studies
mentioned that D. fragilis trophozoite is fragile and degenerates rapidly in the
environment (Johnson et al. 2004; Stark et al. 2006). The trophozoite stage of D.
fragilis survives in stool specimens typically from 6 to 48 hours (Johnson et al. 2004).
Some studies have tried to establish a pseudocystic, precystic or cystic stage of D.
fragilis but all of these studies were dismissed (Greenway 1928; Wenrich 1936, 1944;

6
Knoll & Howell 1946). Currently a cyst stages was observed in D. fragilis and
suggested that it facilitates the faecal-oral transmission of this protozoan (Munasinghe
et al. 2013).
Dientamoeba fragilis is morphologically similar to H. meleagridis which is
transmitted via a nematode (Graybill & Smith 1920). The possibility of transmission of
D. fragilis via the egg of an intestinal nematode was first suggested by Dobell (1940).
An attempt to infect a human volunteer and animals orally also failed (Dobell 1940).
During the middle of the nineteenth century, Dobell (1940) and Wenrich (1944)
suggested that the intermediate host might be the eggs of a nematode such as Trichuris
or Ascaris. Sukanahaketu (1977) reported some structures resembling D. fragilis, inside
the ova of Ascaris lumbricoides. For the first time, Burrows and Swerdlow (1956)
proposed that E. vermicularis, the human pinworm might be a vector of D. fragilis.
They examined 1518 appendices histologically and found 22 harbouring D. fragilis, 12
of which also contained adults or eggs of E. vermicularis. Other studies also reported
co-infection between D. fragilis and E. vermicularis (Yang & Scholten 1977; Ockert
1990).
Ockert (1975) also showed the association between D. fragilis and the eggs of E.
vermicularis. He infected himself with E. vermicularis eggs and consequently
developed both enterobiasis and dientamoebiasis. Subsequently, other studies reported
the co-infection between pinworm and D. fragilis in females (Ockert 1990) and children
(Girginkardesler et al. 2008). Due to these findings, it was suggested that D. fragilis
might be transmitted via E. vermicularis ova. Recent studies also suggested the
association between D. fragilis and E. vermicularis (Röser et al. 2013; Ögren et al.
2013).
On the contrary, some reports showed that there is no correlation between D.
fragilis and E. vermicularis. Kean and Malloch (1966) analysed 100 patients with D.
fragilis infections using sticky tape test and found them all negative for E. vermicularis.
Another study investigated 25 paediatric patients and no association was observed
between D. fragilis and E. vermicularis (Cuffari et al. 1998). In 2005, Stark et al.
performed a prospective study examining stool specimens from 6,750 patients and all
D. fragilis infected patients were tested for E. vermicularis, other helminths and

7
helminth ova (Stark et al. 2005a). No E. vermicularis ova were detected but they found
other protozoans which are usually transmitted via faecal–oral route (Stark et al. 2005a).
Furthermore, the same author reported recently that no correlation was observed
between helminths and D. fragilis (Stark et al. 2010b).
1.6 Genetic diversity To date, two possible genotypes of D. fragilis have been identified (Barratt et al.
2011a). Initially, the SSU rRNA gene was successfully amplified from D. fragilis in all
cases and examined using the control sample of D. fragilis strain Bi/PA (ATCC 30948)
(Silberman et al. 1996). For the first time, Johnson and Clark (2000) reported two
genetic entities of D. fragilis such as genotypes 1 and 2 (Johnson & Clark 2000; Stark et
al. 2006). Two other studies also supported their findings. Peek et al. (2004) examined
the genetic diversity of D. fragilis from 93 patients and 6 asymptomatic carriers by
polymerase chain reaction- restriction fragment length polymorphism (PCR-RFLP) and
found only genotype 1. In addition, in the same year, Windsor et al. (2004) also
performed PCR–RFLP of the SSU rRNA from 33 D. fragilis clinical isolates and
reported only the existence of genotype 1.
Moreover, Stark et al. (2005b) performed PCR-RFLP analysis on 50 D. fragilis
isolates to determine the genetic diversity and found only genotype 1 (Stark et al.
2005b). Genetic analysis between three D. fragilis housekeeping genes also showed
clear distinction between these two genotypes (Stensvold et al. 2013).
1.7 Diagnostic methods
The diagnosis of most intestinal protozoan infections by stool examination requires
the detection and identification of cysts or trophozoites. It is important to note that, the
cyst stage of D. fragilis was identified recently (Munasinghe et al. 2013). Earlier studies
were compromised by inadequate diagnosis or poor laboratory practices; as a result
there were low prevalences of D. fragilis in most of the literature (Spencer et al. 1982;
Yang & Scholten 1977).

8
1.7.1 Fixative, staining and microscopy analysis
Diagnosis by microscopy requires prompt fixation of clinical specimens as D.
fragilis trophozoites degrade rapidly in the environment (Stark et al. 2010b). In most
laboratories, permanent staining of faecal smears is recommended (Dobell 1940; Stark
et al. 2005b). Many different stain and fixative methods have been used successfully for
diagnosis of D. fragilis. These include polyvinyl alcohol (PVA) (Goldman & Brooke
1953), modified Schaudinn’s fixative (Scholten 1972), phenol-alcohol-formalin
(Burrows 1967) and sodium acetate- acetic acid-formalin (SAF) (Yang & Scholten
1977). Currently, most laboratories commonly use PVA and SAF fixatives, along with
iron–haematoxylin and trichrome stains (Johnson et al. 2004).
Studies have reported that permanent staining is time consuming and may not be
the right choice for diagnosis (Garcia 2002; Stark et al. 2011). Using these techniques,
D. fragilis may be difficult to differentiate from other non-pathogenic protozoa such as
Endolimax nana (Sawangjaroen et al. 1993). Moreover, it is possible to confuse the
morphology of D. fragilis with other single-celled protozoan like Blastocystis hominis.
1.7.2 Different culture techniques
There are quite a few studies which have investigated different culture
techniques for this parasite (Johnson et al. 2004; Stark et al. 2006). Usually culture is
more sensitive for diagnosis and needs less amounts of faeces than permanent stains
(Sawangjaroen et al. 1993; Windsor et al. 2003). Interestingly, Burg et al. (1938) found
that D. fragilis only survived in cultures which contain a large amount of bacteria, and a
scarcity of bacteria killed cultures within a day. Initially, Dobell (1940) used different
biphasic media inspissated by horse serum and egg slope to support D. fragilis growth
(Dobell 1940; Johnson et al. 2004). Other biphasic media were able to support the
growth of D. fragilis include Cleveland and Collier’s medium, modified Boeck and
Drbohlav’s (BD) medium and Robinson’s medium (Cleveland & Collier 1930; Windsor
et al. 2003; Rayan et al. 2007).
Over the past few years, research on D. fragilis has been significantly hindered
by the lack of an axenic culture system. Many authors attempted to produce an axenic
cuture for D. fragilis but most of the studies have failed (Johnson et al. 2004). Jacob

9
(1953) has tried in different ways to produce a monoculture of D. fragilis and eventually
found that Dientamoeba failed to grow in the presence of dead bacteria heated at 60-
65ºC for an hour. Jacob (1953) also tried to achieve an axenic culture by antibiotic
treatment. They added penicillin, streptomycin and in some cases sulphadiazine in the
culture medium while growing D. fragilis. They continued this experiment for at least
seven consecutive days and indicated that Dientamoeba can grow with Clostridium
perfringens alone (Jacob 1953). However, most of the attempts failed to maintain an
axenic culture of D. fragilis (Nagata et al. 2012a).
Barratt et al. (2010) conducted a potential study of in vitro culture conditions
that are able to support the long-term growth of D. fragilis trophozoites. Different types
of culture media like a modified BD medium, TYGM-9, Loeffler’s slope medium [a
modified Cleveland and Collier’s medium], Robinson’s medium, Medium 199,
Trichosel and a Tritrichomonas foetus medium were tested. Four media such as TYGM-
9 broth, Robinson’s medium, modified BD medium and Loeffler’s media were able to
support the in vitro growth of four clinical isolates of D. fragilis whereas
Tritrichomonas foetus media and Trichosel failed to maintain the growth of D. fragilis
(Barratt et al. 2010). Higher cell densities were obtained at 42ºC compared to 40ºC and
37ºC. The parasite propagated well under both microaerophilic (6% O2, 7.2% CO2,
3.6% H2, 83.3% N2) and anaerobic (0.2% O2, 9.9% CO2, 5% H2, 84.9% N2) conditions
rather than in the presence of atmospheric levels of oxygen. Furthermore, another
current study observed that Loeffler’s slope medium supplemented with EBSS (Earle's
Balanced Salt Solution) supported the highest growth of D. fragilis trophozoites
(Munasinghe et al. 2012).
1.7.3 Molecular diagnosis
Compare to other protozoans, there are few molecular diagnostic techniques
developed to detect D. fragilis from stool samples. For example, commercially available
monoclonal antibodies and enzyme immunoassays are available for the detection of
antigen in stools for Cryptosporidium parvum, Giardia intestinalis and E. histolytica
which is not available for D. fragilis, as there is no axenic culture system developed yet
for this pathogen (Stark et al. 2006). Molecular diagnostic techniques such as
conventional and real–time polymerase chain reaction (RT-PCR) targeting the 18S

10
rDNA have been developed (Peek et al. 2004; Stark et al. 2006; Verweij et al. 2007).
Initially, Peek et al. (2004) has reported the development of a PCR technique to detect
D. fragilis from human stool samples. It is possible to analyse D. fragilis sequences
directly from faecal specimens without culturing (Peek et al. 2004). However, in this
study the sensitivity of this PCR was not determined.
Stark et al. (2010a) suggested that real-time PCR was the most sensitive of all
diagnostic methods for the detection of D. fragilis. At first, Stark et al. (2005a)
developed the PCR assay which did not cross-react with other protozoan parasites. The
data showed a specificity of 100% and a sensitivity of 93.5%. In addition, the author
developed a 5' nuclease (TaqMan)-based real-time PCR assay, targeting the small
subunit rRNA gene (Stark et al. 2006). Recent studies also highlighted that nested PCR
and Multiplex Tandem Real-Time PCR (MT-PCR) could be a choice for rapid
diagnosis of D. fragilis from clinical samples (Stark et al 2011; Sarafraz et al. 2013).
1.8 Symptoms and treatment
There are plentiful reports from many different parts of the world to validate the
association of D. fragilis with clinical symptoms, typically diarrhoea, abdominal pain,
nausea, vomiting and fatigue (Stark et al. 2010b; Barratt et al. 2011a). Some studies
have also confirmed the link between this parasite and urticaria (Yang & Scholten
1977), biliary infections (Talis et al. 1971), pruritus (Spencer et al. 1982), allergic
colitis (Cuffari et al. 1998), irritable bowel syndrome (Borody et al. 2002), and
diarrhoea in people infected with human immunodeficiency virus (Lainson & Da Silva
1999). Several studies have shown that the most common gastrointestinal symptoms in
D. fragilis infected children are diarrhoea, abdominal pain, nausea, flatulence,
constipation as well as anorexia, fatigue and peripheral eosinophilia (Norberg et al.
2003; Banik et al. 2011; Schure et al. 2013). Cuffari et al. (1998) reported a case of a
female four-year old child who suffered with a three-year history of chronic diarrhoea.
Antimicrobials which are used to treat D. fragilis infections include iodoquinol
(diiodohydroxyquin) (Butler 1996), metronidazole (Preiss et al. 1990), paramomycin
(Vandenberg et al. 2007) and nitroimidazole derivatives such as secnidazole and
ornidazole (Kurt et al. 2008). However, most of the current studies are based only on

11
small sized case reports (Nagata et al. 2012b). Antimicrobial treatment that has been
successfully and commonly used for D. fragilis infection in children includes idoquinol
and metronidazole (Vandenberg et al. 2007; Banik et al. 2011). A recent study
conducted by Schure et al. (2013) indicated that clioquinol could be more effective than
metronidazole to treat D. fragilis infections in children. Barratt et al. (2013) also
reported that some dry plant extracts are not effective for eliminating D. fragilis growth
in in vitro culture.
1.9 Literature review on viruses of parasitic protozoa-introduction
At the end of the eighteenth century, viruses were described as ‘submicroscopic
infectious agents’ obtained from infected cell extracts (Mayer 1886; Enquist 2009).
Over the last few decades, the study of bacteriophages and viruses has provided
important knowledge on the control of gene expression, RNA processing, and
translation (Patterson 1990). Even though there is a long history of virus study in a wide
range of organisms, viruses in protozoa were not definitively identified until 1986
(Wang & Wang 1991). The impetus for research on them was then accelerated. These
protozoan viruses could be used as transforming vectors and thus play an important role
in developing studies in protozoan genetics where knowledge is relatively poor in
comparison to other microorganisms such as yeast or bacteria (Patterson 1990; Wang &
Wang 1991).
Viral infections have already been described in other protozoa such as
Plasmodium, Naegleria, Entamoeba, Leishmania, Endotrypanum, Trypanosoma,
Babesia, Blastocystis sp. and in Cryptosporidium (Garnham et al. 1962; Schuster 1969;
Diamond et al. 1972; Molyneux 1974; Croft et al. 1980; Molyneux & Heywood 1984;
Johnston et al. 1991; Teow et al. 1992; Khramtsov et al. 1997). Moreover, analysis of
numerous isolates revealed that one-half of Trichomonas vaginalis clinical isolates were
persistently infected with a double-stranded RNA (dsRNA) virus (Goodman et al.
2011a). Double-stranded RNA viruses were also found in several isolates of Giardia
(Wang & Wang 1986a) and Eimeria (Ellis & Revets 1990). The presence of virus-like
particles (VLP) is very common in parasitic protozoa, especially those that inhabit the
gut (Goodman et al. 2011a) (Table 1.1).

12
Table 1.1 Comparison of several characteristics of protozoan viruses.
aViruses for which full-length genome sequences have been reported to GenBank; acc. Accession
(RdRp = RNA-dependent RNA polymerase)
Properties Protozoan
viruses
Genbank acc. no.a
Nucleic acids (Kb)
Shape of virion
Virus particles
(diameter)
RdRp activity
Capsid protein (KDa)
Ref
Trichomonas
virus
U08999
dsRNA
(4.3 to 5.0 )
icosahedral
33-120 nm
non
segmented dsRNA genome
75-160
Benchimol, Chang & Alderete 2002b
Giardiavirus
AF525216
dsRNA (~ 7.0 )
icosahedral
33-36 nm
non segmented
dsRNA genome
95-190
Adam 2001
Entamoeba
virus
unknown
Unknown
icosahedral
or filaments
75-85 nm
Unknown
unknown
Adam 2001
Eimeriavirus
AF356189
dsRNA
(0.57–11.5 )
icosahedral
35-44 nm
dsRNA, unknown for some species
80-95
Han et al.
2011
Leishmania virus
NC002063.1
dsRNA/ssRNA
(~ 6.0 )
icosahedral (mostly
spherical)
32-55nm
dsRNA genome
80-180
Tarr et al.
1988

13
Typically, most virus-like particles (VLPs) of protozoa are RNA or dsRNA
viruses ranging from 30-200 nm in diameter and the size of their genome is 5-7 kb
(Wang & Wang 1991; Benchimol et al. 2002b). Double-stranded RNA viruses are
known in all major groups of organisms, from bacteria and fungi to animals and plants
(Dobos et al. 1979; Yoshikawa & Converse 1990; Wickner 1996; Fire et al. 1998).
Double-stranded RNA virus generally accompanies RNA virus replication which occurs
in the cytoplasm for all dsRNA viruses (Wickner 1993; Patton & Spencer 2000; Weber
et al. 2006). The life cycle of dsRNA viruses include some steps such as attachment,
penetration, uncoating, transcription, translation, assembly, and finally release from host
cells (Wickner 1993; Weber et al. 2006) (Figure 1.3).
It is reported that protozoan RNA viruses show several similarities and closely
correspond to dsRNA viruses of yeasts (Flegr et al. 1988; Kasprzak & Majewska 1995).
In addition, the viral genome is a non-segmented dsRNA, and viruses exhibit isometric
symmetry. The presence of dsRNA viruses or VLPs within T. vaginalis is associated
with expression of immunogenic proteins on the trichomonad surface. It also effects
protozoal phenotypes, upregulates certain proteins and causes disease pathogenesis
(Gerhold et al. 2009; Fraga et al. 2011; Malla et al. 2011).
Phylogenetic analyses revealed that the genera Trichomonasvirus, Giardiavirus,
and Leishmaniavirus clustered in the family Totiviridae (Goodman et al. 2011a, 2011b).
This family includes several viruses that infect either fungi or a number of medically
important protozoan parasites such as Trichomonas, Giardia and Leishmania. Briefly,
the family Totiviridae are characterised by : (1) icosahedral virions; (2) size ranging
between 30 and 40 nm in diameter; and (3) normally encapsidate monosegmented (i.e.,
nonsegmented) dsRNA genomes with overlapping open reading frames encoding a
capsid protein (CP) and an RNA-dependent RNA polymerase (RdRp) (Provenzano et
al. 1997; Ghabrial 2008; Goodman et al. 2011a; Parent et al. 2013). Recent genome
sequencing studies have confirmed that Trichomonas vaginalis viruses (TVVs) are
phylogenetically divergent from Giardia lamblia viruses (GLV) (Goodman et al.
2011a). It was also suggested that several TVV genomes showed homology to
monosegmented (i.e., nonsegmented) dsRNA viruses of the family Totiviridae (Tai &
Ip 1995; Su & Tai 1996; Bessarab et al. 2000).

14
Figure 1.3 Schematic diagram showing the replication strategy of double-stranded
RNA virus (Source: http://www.microbelibrary.org/images/rybicki/bigiii6.gif).
(1) Primary transcription; the synthesis of viral (+) strands from a dsRNA template
takes place within viral particles. Transcription is completed by the virus using an
RNA-dependent RNA polymerase (RdRp) and newly synthesized (+) sense RNA
extruded into cytoplasm. (2) Viral proteins are produced by translation of same (+)
sense RNA. (3) Assembly of (+) sense RNA occurs in cytoplasm and immature virions
are formed from viral proteins. (4) Once the new particles or cores have formed; dsRNA
is formed by the transcription of (+) sense RNA using viral RdRp. (5) Subsequently,
secondary transcription of dsRNA occurs. (6) Finally, progeny virions are found in
cytoplasm.

15
Among the protozoan viruses, the TVV-1 is the first virus reported with full
genome sequence and five full length genomic sequences for TVVs are now available in
Genbank to date (GenBank accession numbers: TVV1-1, U08999; TVV1-T5, U57898;
TVV1-IH2, DQ270032; TVV2-1, AF127178; TVV3-1, AF325840) (Goodman et al.
2011a). In a sequence comparison, putative RdRp of Trichomonas virus shows 20, 24,
and 25% sequence identity to the RdRp of Giardia lamblia virus (GLV),
Saccharomyces cerevisiae virus (ScV) and Leishmania RNA virus 1 (LRVl)
respectively (Tai & Ip 1995). When all four RdRp sequences from these viruses are
compared, extensive conserved regions are observed in the middle of RdRp sequences
(Tai & Ip 1995). Moreover, amino acid sequence alignment shows that capsid protein
sequences are very different among major protozoan parasites. Analysis of genome
sequences and RdRp sequences from TVV-T1 suggests that TVV-T1 is similar to
Saccharomyces cerevisiae virus L-A (ScV-L-A) and Leishmania RNA virus (Tai & Ip
1995) whereas it is only distantly related to G. lamblia virus (Tai & Ip 1995; Goodman
et al. 2011a). Gerhold et al. (2009) also reported that the identity between T. vaginalis
dsRNA viral sequences is variable suggesting that T. vaginalis isolates are infected by
several different dsRNA viruses (Benchimol et al. 2002b; Gerhold et al. 2009). Earlier
studies also indicated that the dsRNA genome of the T. vaginalis virus does not
hybridize to dsRNA viruses of other protozoa (Revets et al. 1989).
The phylogenetic relationship of TVV with other protozoan viruses has been
reviewed recently (Goodman et al. 2011a). They focused mainly on TVV and described
relatively little knowledge on other protozoan viruses. Due to recent advances in the cell
biology of these protozoan viruses, a revision is warranted. This review chapter covers
the general background of protozoan viruses with more emphasis in areas which
concern my research. I review the latest information on various purification methods,
morphological characterisation and molecular identification of viruses mainly from
protozoan parasites such as T. vaginalis, G. lamblia, E. histolytica, Eimeria spp., and
Leishmania. I also highlight the role of major capsid proteins identified from these
protozoan parasites. Furthermore, the effects of these viruses on parasite’s life cycle and
disease pathogenesis are also discussed.

16
1.10 Double-stranded RNA (dsRNA) viruses of Trichomonas vaginalis
1.10.1 TVV, the first dsRNA virus of protozoa
Trichomonas vaginalis virus (TVV) was the first virus described in a protozoan
parasite (Wang & Wang 1985, 1986b) (Figure 1.4 A) and biochemically characterised
(Wang & Wang 1986a; Wang et al. 1987; Khoshnan & Alderete 1993). A 5.5-kb
nucleic acid species was seen by electron microscopy and in hot phenol extracts of T.
vaginalis strain ATCC 30001 (Wang & Wang 1985). Rapid screening of various strains
of T. vaginalis for the dsRNA was performed. Of the 33 different strains of T. vaginalis
examined in their study, all contained similar dsRNA (Wang & Wang 1985). These
virus were refractory to enzymes such as DNase I, DNA polymerase I, or restriction
endonucleases that utilized DNA as substrates and readily degraded by treatment with
0.2M NaOH, ribonuclease (RNase) T1, or RNase A at room temperature, suggesting
that this new species is comprised of RNA (Wang & Wang 1985). Finally, the resulting
susceptibility to RNases, a DNA-like buoyant density and hyperchromicity concluded
that the nucleic acid species was dsRNA (Wang & Wang 1991).
In the last few years, several studies had provided information on the presence
and morphology of VLPs in T. vaginalis (Wang & Wang 1986a; Benchimol et al.
2002b; Benchimol 2004; Kim et al. 2007), Tritrichomonas foetus (Gomes Vancini &
Benchimol 2005), and Trichomonas gallinae (Gerhold et al. 2009) based on electron
microscopy. Various shapes such as filamentous, cylindrical, spherical and oblong-
shaped forms of VLPs were observed in T. vaginalis (Benchimol et al. 2002a, 2002b).
Trichomonas vaginalis virus was found at the cell periphery, close to the axostyle, and
inside the nucleus (Gomes Vancini & Benchimol 2005). The description of VLPs in T.
foetus was delayed due to its low density or latent conditions within the parasite. Virus-
like particles were detected in T. foetus only after the parasites were treated with
cytoskeleton-affecting drugs such as colchicine, vinblastine, taxol, nocodazole, and
griseofulvin (Gomes Vancini & Benchimol 2005) (Figure 1.4 B). They were detected
by electron microscopy and confirmed by immunofluorescence microscopy using
antibodies directed against viral proteins (Gomes Vancini & Benchimol 2005). Loss of
the viruses by some T. vaginalis isolates (such as T068-II) through daily passage over
long periods was also reported (Khoshnan & Alderete 1993; Wang et al. 1987). The

17
presence of a dsRNA virus was also described in Korean, Cuban, Tehran and different
South African T. vaginalis isolates (Weber et al. 2003; Fraga et al. 2005; Kim et al.
2007; Fraga et al. 2012; Heidary et al. 2013).
It has been suggested that the presence of dsRNA may be related to the
sensitivity of T. vaginalis toward the antiprotozoal agent metronidazole (Sobel et al.
1999). Two metronidazole-resistant strains of T. vaginalis, IR78 and 85, were
investigated, both strains showed only one band of bulk DNA in the CsCl density
gradient as well as by agarose gel electrophoresis. The trophozoites contained the
dsRNA at less than 0.1% of their bulk DNA (Muller & Gorrell 1983; Wang & Wang
1991). Recently, Malla et al. (2011) found that around 30 fresh T. vaginalis isolates
collected from both symptomatic and asymptomatic women harboured dsRNA virus
and were sensitive to metronidazole in vitro. In contrast, different results suggest that T.
vaginalis strain 375, which has no detectable dsRNA, has susceptibility to
metronidazole (Wang & Wang 1986b). Flegr et al. (1987) also reported that
metronidazole resistance does not correlate with the absence of dsRNA.
Evidence suggested that T. vaginalis infection with TVV may be associated with
attenuated cytopathogenicity, an acute host inflammatory response and hypervirulence
(Alderete et al. 1986; Provenzano et al. 1997; Kim et al. 2007; Goodman et al. 2011a).
Fraga et al. (2007) performed a study to determine the possible clinical significance of
dsRNA viral infection of T. vaginalis and found that the clinical signs were significantly
associated with TVV infection. It suggests that the T. vaginalis virus plays a vital role in
the pathogenic mechanisms of human trichomoniasis. Wendel et al. (2002) stated that
patients with virus-positive isolates were significantly older than patients with virus-
negative isolates and virus-positive isolates were more prevalent among women.
Consequently, it was also suggested that more studies are needed to correlate TVV-
infected T. vaginalis with pathogenicity in humans (Goodman et al. 2011a).
1.10.2 Methods of purification and molecular identification of TVV
A variety of purification techniques such as filtration, CsCl density-gradient
centrifugation, sucrose cushion and ultracentrifugation were used successfully to purify
viruses form different protozoa (Khoshnan & Alderete 1993; Kim et al. 2007; Gerhold

18
et al. 2009). Among them CsCl density gradients were used extensively to purify the
dsRNA virus from trichomonads (Wang & Wang 1986a, 1986b; Khoshnan & Alderete
1993). Nearly all studies followed similar purification methods to purify TVV from
different strains of T. vaginalis growing in axenic culture. Briefly, more than 105-107
trichomonad cells were suspended in TNM (50 mM Tris [pH 7.5], 150 mM NaCl, 5
mM MgCl2) buffer and lysed by sonication until more than 90% of the cells were lysed.
In some studies, the lysed trichomonads were also treated with proteinase K so that
dsRNAs could not be protected within viral capsids (Wang & Wang 1991; Khoshnan &
Alderete 1993). The lysates were centrifuged and virus-particles containing sediment
were prepared. Then they were equilibrated with CsCl (density between 1.35 -1.75g/ml)
and centrifuged at 100,000 x g for 24 h (Khoshnan & Alderete 1993). Some studies also
collected all the fractions after CsCl gradient and the absorbance of each fraction was
monitored with a spectrophotometer at 260 nm. Samples of individual fractions were
extracted with phenol and analysed by gel electrophoresis (Wang & Wang 1986b).
Fractions containing the viral RNA were further examined by negative staining and
electron microscopy for the presence of VLPs (Wang & Wang 1986b). The results
showed that the dsRNA remained stable in crude homogenates of T. vaginalis but all
other nucleic acids were found degraded (Wang & Wang 1986b, 1991).
Electron microscopic studies on Trichomonasvirus have shown that several
types of VLPs can simultaneously infect T. vaginalis (Benchimol et al. 2002a). The
number of dsRNA segments varied from 1 to 3 in different TVV isolates and the length
of those dsRNA segments varied from 4.3 to 7.0 kb (Khoshnan & Alderete 1993; Su &
Tai 1996; Kim et al. 2007).
The standard method to purify dsRNA from different TVV particle is phenol-
chloroform extraction (Wang & Wang 1986b; Khoshnan & Alderete 1993). Viral
genomes are precipitated with ethanol, separated via 0.8-1% agarose gel electrophoresis
and stained with ethidium bromide (Khoshnan & Alderete 1993; Kim et al. 2007; Malla
et al. 2011). Others also used isopropanol instead of EtOH (Flegr 1987). After isolating
the viral nucleic acid, its sensitivity to DNase and RNases was assessed (Khoshnan &
Alderete 1993).

19
1.10.3 Trichomonas vaginalis virus sequencing and protein identification
The Trichomonas vaginalis virus 1-1 (TVV1-1) was the first protozoan virus for
which full-length genome sequence data was reported (Tai & Ip 1995). Initially, Tai &
Ip (1995) cloned the dsRNA of TVV-T1 as a cDNA and obtained a contiguous 4647-bp
cDNA sequence. The 4.6-kb dsRNA of TVV-T1 encodes two overlapping genes, cap
and pol (Liu et al. 1998). Subsequently, Kim et al. (2007) described the presence of a
virus in a Korean T. vaginalis isolate [designated TVV INHA(IH)-2] for the first time
and verified its identity as a member of the Totiviridae family. They found that the
genomic dsRNA of TVV IH-2 was 4,647 bp in length and harbored 2 overlapping open
reading frames of the putative capsid protein and dsRNA dependent RdRp. Kim et al.
(2007) compared the sequence of RNA polymerase of Korean TVV IH-2 isolates with
other TVV isolates and revealed that the Korean isolate formed a compact group with
TVV 1-1 and 1-5 isolates. However, this Korean isolate was not identical to TVV 2-1, 3
and Giardiavirus. The Korean TVV IH-2 isolate also showed weak pathogenicity in the
mouse assay (Kim et al. 2007).
Gerhold et al. (2009) discovered a novel sequence for the RdRp gene of T.
vaginalis viruses (GenBank FJ997643). BLAST analysis revealed an 81-84%
nucleotide and 86-90% deduced amino acid identity to four T. vaginalis virus RdRp
partial sequences (GenBank accessions U08999.1, DQ270032.1, DQ528812.1, and
U57898.1). A pairwise comparison of the six TVV isolates with full length protein-
coding sequences confirmed the proposal of three distinct TVV species, with
interspecies amino acid sequence identities of ≤32 and ≤43% for CP and RdRp,
respectively (Goodman et al. 2011a). Totiviridae includes three different forms of
TVVs (TVV1, TVV2, and TVV3) (Bessarab et al. 2000; Bessarab & Tai 2001; Alderete
et al. 2003; Fraga et al. 2005). Newly identified T. vaginalis viruses TVV4 isolated
from strain TVV4-OC4 (renamed TVV4-1 as prototype of the new species) and TVV4-
OC5 were described recently (Goodman et al. 2011a, 2011b). Trichomonas vaginalis
virus 4 is more closely related to TVV3 than to TVV2 strains (Goodman et al. 2011b).
Moreover, three small and distinct satellite double-stranded RNAs (dsRNAs) denoted

20
s1, s1´, and s2 were subsequently described for another T. vaginalis dsRNA virus
(Khoshnan & Alderete 1995).
In summary, the Trichomonas vaginalis dsRNA genome encodes a CP (~ 70-
160 KDa) and a viral RdRp (~ 1429 aa, linear) in 2 overlapping open reading frames
(ORFs) (Bessarab et al. 2000; Bessarab et al. 2011). The CP ORF is expressed as an
independent protein, whereas the RdRp ORF is expressed only as part of a CP/RdRp
fusion protein following either a -1 or a +1 ribosomal frameshifting mechanism (Tai &
Ip 1995; Liu et al. 1998). This fusion protein is incorporated into viral particles at low
levels and permits these particles to mediate both transcription and replication
(Khoshnan & Alderete 1994; Bessarab et al. 2000; Goodman et al. 2011b). In the
Totiviridae family, similar overlapping cap-pol genomic structures have also been
identified in other protozoan dsRNA viruses such as G. lamblia viruses (Wang et al.
1993), and two related Leishmania RNA viruses (LRV1-1 and LRV1-4) (Stuart et al.
1992; Scheffter et al. 1994). Similar genomic organization was also found in fungal
dsRNA viruses such as Saccharomyces cerevisiae virus L-BC (Park et al. 1996) and
Helminthosporium victoriae 190S virus (Huang & Ghabrial 1996). In contrast, a
different overlapping cap-pol arrangement has been reported for Leishmaniavirus
LRV2-1 (Scheffter et al. 1995).
To date, five full length genomic sequences for T. vaginalis viruses are available
in Genbank and the protein-encoding portions of a sixth strain (Zhao et al. 2006) have
been deposited in Genbank (Goodman et al. 2011b). Goodman et al. (2011b) confirmed
the capacity for concurrent infections of T. vaginalis by showing the full-length cDNA
sequences of 3 distinct TVV strains that are present in the same isolate. Furthermore,
the full-length cDNA sequences of an additional 11 TVVs were obtained from four
other clinical isolates of T. vaginalis (Goodman et al. 2011b).
Despite the genomic complexity of TVV, the capsid proteins were identified in
most studies among the TVV isolates as 75-85 kDa in the SDS-PAGE. Liu et al. (1998)
detected a 75-kDa major protein (capsid protein) and a 160-kDa protein by anti-CAP
serum in a TVV-T1 sample. Although TVV was the first dsRNA virus to be identified
in pathogenic protozoa (Wang & Wang 1985), the protein compositions of its virions

21
were not clearly defined (Khoshnan & Alderete 1993; Tai & Ip 1995). This was
probably due to the difficulty in purifying a single viral species from a T. vaginalis
isolate for biochemical characterisation (Tai et al. 1995). Trichomonas vaginalis virus
free strains IR78 and CDC85 were infected by purified T. vaginalis virus but no dsRNA
were detected in the cell homogenates (Alderete et al. 1985). Subsequently, others
reported the failure of viral infection of T. vaginalis (Wang & Wang 1986b). Li et al.
(2012) developed a viral RNA-based transfection vector pTVV-EGFP/NEO which
incorporated green fluorescent protein gene (GFP) and the neomycin resistance gene
(NEO). These vectors can replace the complete gene encoding region of T. vaginalis
virus (Li et al. 2012). These authors established methods to transiently and stably
transfect the human pathogenic protist T. vaginalis.
1.11 Viruses of Giardia lamblia 1.11.1 Discovery of Giardiavirus
Soon after their previous work, Wang and Wang (1986a) discovered a second
dsRNA virus in another related anaerobic parasitic protozoan, G. lamblia (in the
Portland I isolate) designated as G. lamblia virus (GLV) (Figure 1.4 C). A 7.0-kb
nucleic acid was identified from Giardia trophozoites by agarose gel electrophoresis
which was susceptible toward alkali treatment, ribonuclease A and ribonuclease T1
(Wang & Wang 1986a, 1991). This observation was also confirmed by electron
microscopy which revealed a linear double-stranded structure with an average length of
1.5 micron (Wang & Wang 1986a). It was the first time a well-identified protozoan
virus was reported as highly infectious (De Jonckheere & Gordts 1987; Miller et al.
1988a, 1988b; Cao et al. 2009). Giardia lamblia virus (GLV) is now classified in the
family Totiviridae and assigned to the genus Giardiavirus (Goodman et al. 2011a).
The cell shape of GLV is usually icosahedral, non-enveloped with a diameter of
about 33 nm (Adam 2001). A rapid screening of 76 strains of G. lamblia examined the
presence or absence of any dsRNA in the crude nucleic acid extract; 28 contained the 7-
kb dsRNA, while 48 were virus-free (De Jonckheere & Gordts 1987; Miller et al.
1988b). Others reported that approximately 47% (of 38 isolates) of Giardia tested were
positive for the virus (De Jonckheere & Gordts 1987). Interestingly, the RNA virus was

22
found in Giardia strains isolated from humans in the U.S.A., England and from Poland
(Nash et al. 1987).
Another Giardiavirus of the family Totiviridae is Giardia canis virus (GCV).
Originally, it was isolated from the Giardia canis strain GCVNI infected by a double-
stranded RNA virus (Chen et al. 2006; Cao et al. 2009). Cao et al. (2009) showed that
virus free Giardia trophozoites could be infected by in vitro transcribed GCV
transcripts. In this study (Cao et al. 2009) the presence of a virus particle was detected
by electron microscopy in the cytoplasm as early as 24 h post transfection, the culture
medium at about 60 h and in nuclei at 72 h post transfection.
Giardia virus particles are typically localized on the plasma membrane and enter
into Giardia trophozoites via endocytosis (Tai et al. 1993). Giardia lamblia virus was
then translocated to the peripheral vacuole and finally spread to the cytoplasm (Wang &
Wang 1986a). Like other RNA viruses, GLV replicates in the cytoplasm during the
early phase of the infection but migrates to the nuclei toward the final stages of the cell
growth (Tai et al. 1993). Afterward, it may be discharged into the environment (Tai et
al. 1991) or mature virions are released by the infected cells into the culture medium
(Furfine et al. 1989; Sepp et al. 1994). Mature infectious Giardia viral particles are also
released from the host cell by budding or lysis of infected trophozoites (Lenstra et al.
1988; Cao et al. 2009). Comparatively, G. lamblia viruses were repeatedly found in the
nucleus more than other protozoan viruses (Miller et al. 1987; Miller et al. 1988b;
Wang et al. 1993). It is reported that GLV replicate in the nucleus of infected protozoan
cells and produce a fusion protein during replication (Wang & Wang 1986a; Wang et al.
1993). Interestingly, similar numbers of VLPs were found in both nuclei of Giardia
(Wang & Wang 1986a). In G. lamblia the presence of approximately 192 virus particles
in the left nucleus and 208 in the right nucleus has been documented (Wang & Wang
1986a). It was also documented that approximate 200 Giardia virus particles infect each
nucleus (Adam 2001).
Most of the studies separated and purified the GLV virions by CsCl density
gradient centrifugation following osmotic rupture of the trophozoites (Miller et al.
1988b; Tai et al. 1996). Initially, Wang and Wang (1986a) purified GLV by CsCl

23
density gradient centrifugation and dsRNA was extracted with phenol followed by
ethanol precipitation. Later, Miller et al. (1988b) also showed that GLV particles could
be isolated as dsRNA-enriched fractions by similar methods using infected G. lamblia
cell-free extracts and from the spent culture medium of infected cells. These authors
verified that these purified viruses could infect virus-free G. lamblia at a multiplicity of
infection (m.o.i.) of as low as 10 virus particles per cell (Miller et al. 1988b).
Giardia lamblia virus contains a linear, non-segmented dsRNA (approximate
size is 7.0-kb) with little homology to T. vaginalis virus dsRNA (Wang & Wang 1986a;
Adam 2001). The other characteristics of the GLV genome are: (1) it contains two
genes gag and pol: encoding the major capsid protein and a fusion of gag with the viral
RNA-dependent RNA polymerase (RdRp) respectively; (2) one of the two strands has a
free hydroxyl group at the 3' end; and (3) does not possess a poly (A) tail at its 3'
termini (Wang et al. 1988; Wang & Wang 1991). Like TVV and Leishmaniavirus,
Giardiavirus contains RdRp which is responsible for transcription and replication of the
viral genomes (Poch et al. 1989; Murphy & Kingsbury 1990; White & Wang 1990;
Köhler & Wang 1997). Studies dealing with molecular cloning and nucleotide sequence
analysis of cDNAs of Giardia dsRNA viruses also confirmed the existence of this
enzyme in the viral dsRNA genome (Miller et al. 1988b; Icho & Wickner 1989). A new
GLV was recently isolated from G. lamblia BJ isolate, whose dsRNA genome was
6273-bp and flanked by a 367-bp 5´ untranslated region (5´ UTR) and a 296-bp 3´ UTR
(Cao et al. 2009).
Giardia lamblia virus contains a 100-kDa major capsid protein and depends on a
cysteine protease for cleavage into the mature protein (Yu et al. 1995). A slightly
smaller capsid protein (95 kDa) was also identified, which differs significantly from the
100-kDa capsid protein (Tai et al. 1996). Studies found that 70% of virus positive G.
lamblia isolates contain virus receptors on the parasite cell surface (Sepp et al. 1994).
Additionally, two homologous dsRNAs were identified in G. lamblia from two distinct
virions; (1) GLVp100, whose capsid consists of p100 and p190; and (2) GLVp95,
whose capsid consists of a 95-kDa protein (p95) and a minor p190-equivalent protein
(Tai et al. 1996). Western blotting and peptide mapping experiments then confirmed
that p100 and p95 were closely related proteins but their amino acid sequences are quite

24
different (Tai et al. 1996). Giardiavirus protein is detectable inside or outside the cell
by immunofluorescent staining. Antisera against intact purified Giardiaviruses were
prepared in rabbits and used to analyse the expression of the viral proteins in the cells
transfected with viral RNA; positive signal was observed in virus-sensitive G. lamblia
WB strain but not in virus -resistant G. lamblia Ac and JH strains (Sepp et al. 1994).
Trichomonas vaginalis virus and GLV are distinct viruses in many aspects.
Evidence indicated that their RNAs do not cross-hybridize and also their capsid
polypeptides do not cross-react immunologically (Wang et al. 1988; Wang & Wang
1991). Studies also suggested that GLV can be isolated from the stationary-phase
growth of G. lamblia culture medium and used to infect virus free strains of G. lamblia
in vitro (Miller et al. 1988b). Thus it is readily different from TVV in this aspect.
Giardia lamblia trophozoites can harbor as many as 5 X 105 virus particles per cell and
these numbers are comparatively higher than other parasites and these virus particles do
not apparently affect their rate of growth (Miller et al. 1988b). However, T. vaginalis
and T. foetus cannot be infected by Giardiavirus via either viral infection or RNA
transfection (Sepp et al. 1994). Interestingly, both Giardia and Trichomonas viruses
overproduce an ssRNA which could act as a viral message and replicative intermediate
(Wang & Wang 1991).
1.12 Entamoeba histolytica virus- missing protozoan virus Unlike Giardia and Trichomonasvirus, Entamoebavirus has not been studied in
detail as it is uncommon (Wang & Wang 1991). Initially, Miller and Swartzwelder
(1960) observed virus-like particles within the perinuclear cytoplasm of Entamoeba
trophozoites. Subsequently, an amoebal virus ranging in size from 75-85 nm in diameter
was isolated from four different strains of Entamoeba histolytica (Diamond et al. 1972;
Mattern et al. 1972). Two different types of virus were described in their study: (1) the
first type replicated in the nucleus, lysed the nucleus and finally caused cell death; and
(2) the second type was seen only in the cytoplasm and was described as a typical
polyhedral (icosahedral) virus (Diamond et al. 1972). In fact, the second one consists of
DNA and is lytic to certain strains of E. histolytica (Hruska et al. 1973) (Figure 1.4 D).
After that, Mattern et al. (1974) described details of the ultrastructure of the polyhedral
virus V301 isolated from E. histolytica strain HB-301.

25
Wang and Wang (1991) stated that “Entamoeba VLPs could not be
differentiated from prokaryotic inclusions within the eukaryotic cell” (Lemke 1976;
Wang & Wang 1991). In 1977, the presence of a third type of beaded particle in the
nuclei of a few Entamoeba strains was reported (Mattern et al. 1977). None of the
above three VLPs of Entamoeba bear much resemblance to T. vaginalis virus (Wang &
Wang 1986b). Previously, rhabdovirus-like particles were also found in
electronmicrographs of thin section preparations of E. histolytica and E. invadens
(McCaul & Knight 1974; Bird & McCaul 1976). Olivier et al. (1984) found only a
filamentous (rhabdovirus-like) structure in E. histolytica and suggested that their
presence was not related to the pathogenicity of the amoeba.
Purification of Entamoebavirus from E. histolytica was attempted by differential
centrifugation. The purified virus pellet was membrane-free, biologically active but
showed no cytopathological effect in the host strain tested, HB-301 (Hruska et al.
1973). Mattern et al. (1977) used ultracentrifugation and CsCl density gradient
(isopycnic) centrifugation to purify beaded-like viruses from E. histolytica. However,
there is no clear molecular evidence to identify Entamoebavirus. Entamoeba histolytica
does not contain a dsRNA virus (Hruska et al. 1974, Mattern, et al. 1974).
1.13 Virus-like RNA in Eimeria species Over the last few years, virus-like particles have been described for different
species of Eimeria such as Eimeria stiedae (Revets et al. 1989), Eimeria necatrix (Ellis
& Revets 1990) and Eimeria nieschulzi (Sepp et al. 1991; Roditi et al. 1994). Initially,
Revets et al. (1989) isolated a 6.5 kb nucleic acid from E. stiedae sporozoites, which
was degradable by RNase treatment and it was dsRNA (Revets et al. 1989). Virus-like
RNAs were also detected in three species of avian Eimeria, i.e., Eimeria maxima (Ellis
& Revets 1990), Eimeria acervulina (Lee et al. 1996) and Eimeria brunetti (Lee et al.
1996). Eimeria stiedae virus (Revets et al. 1989) and E. nieschulzi virus (Roditi et al.
1994) were included in the family of Totiviridae. Icosahedral VLPs with a diameter of
35 nm in sporozoite lysates were identified in E. stiedae by electron microscopy (Revets
et al. 1989). Viruses with a diameter of approximately 39 nm in E. nieschulzi were
reported by Roditi et al. (1994). Eimeria necatrix VLPs were non-enveloped,
icosahedral and 42–44 nm in diameter (Lee et al. 1996) (Figure 1.4 E). Lately, Han et

26
al. (2011) identified E. tenella viruses (ETV) with a diameter of approximately 38 nm
from sporulated oocysts (Han et al. 2011).
Revets et al. (1989) suggested that RNA associated with the E. stiedae virus-like
particle strongly cross-hybridized to the GLV RNA and not to TVV RNA (Revets et al.
1989; Wang & Wang 1991). Even though Eimeria and Giardia infect different hosts
and replicate differently, these cross-hybridization experiments with GLV suggested
that they have sequences in common (Revets et al. 1989). In contrast, there was no
homology detected between TVV dsRNA and GLV dsRNA genome (Wang & Wang
1991).
Like other protozoans, a variety of purification methods were used to purify the
VLP from different species of Eimeria. Caesium chloride (CsCl) gradient density
centrifugation was used to purify E. stiedae (Revets et al. 1989), E. nieschulzi (Roditi et
al. 1994) and E. necatrix (del Cacho et al. 2001) viruses. Virus-like RNAs and VLPs
were not found in E. tenella with similar purification assays (Sepp et al. 1991; Lee, et
al. 1996; Shirley 2000). Sucrose density gradient centrifugation was also used to purify
Eimeria viruses (Han et al. 2011). The purified Eimeria viruses were stored and stable
at -220C in glycerol solution (Miller et al. 1988b; del Cacho et al. 2001).
For the first time, a 6.5-kb linear dsRNA measuring a relatively uniform length
of 1.63 μm (which is corresponding to a linear RNA of 5.5 kb) was identified by
electron microscopy from E. stiedae sporozoites (Revets et al. 1989). Ellis and Revets
(1990) then identified two viral-like RNAs in the oocysts and sporozoites of E. necatrix.
Most of the dsRNA isolated from different Eimeria viruses are linear but varied in size
among species (Ellis & Revets 1990). The size of these dsRNA was from 0.57–11.5 kb
and non-segmented (Han et al. 2011). A subgenomic viral dsRNA fragment of about
0.57 kb was also identified in E. nieschulzi virus (Han et al. 2011). In addition, the
presence of a 5.8 kb RNA in the oocysts and sporozoites of E. maxima was confirmed.
No dsRNA was found in the three different strains of E. tenella examined (Lee et al.
1996). Recently three extrachromosomal dsRNA segments of 1.4, 2.4 and 3.6 kb in size
were observed for E. tenella viruses (Han et al. 2011). Interestingly, E. nieschulzi was

27
persistently infected with five different VLP-RNAs of 0.57, 0.72, 5.0, 5.7 and 11.5 kb
(Roditi et al. 1994).
Han et al. (2011) detected RdRp activity in crude extracts of E. tenella
sporulated oocysts suggesting that it contains novel dsRNAs. Total nucleic acid of E.
tenella was isolated and analysed by agarose gel electrophoresis (Han et al. 2011).
RNA-dependent RNA polymerase activity for E. stiedae virus, E. acervulina virus and
E. brunetti virus remains unknown but was detectable for E. nieschulzi virus, E. maxima
virus, E. necatrix virus and other E. tenella viruses (Han et al. 2011). Overall, the
genome structures of E. nieschulzi viruses (ENV-1) are different from E. stiedae viruses
(ESV) (Roditi et al. 1994). The cDNA clone of ENV-l showed significant homology to
a region of the Leishmaniavirus (LRV 1) genome that encodes an RdRp (Lee &
Fernando 1999a). The presence of RNA polymerase associated with the dsRNA virus
was identified in the sporozoites and merozoites of both E. maxima and E. necatrix (Lee
& Fernando 1999a, 1999b).
Two E. necatrix viral proteins, RdRp (95 kDa) and the major capsid protein (80
kDa), were detected by immunoblot analysis using a polyclonal antibody raised against
the viral-like proteins (del Cacho et al. 2001). Both proteins were also identified in
Leishmaniaviruses (Ro et al. 1997). Capsid protein was found only in sporocysts and
sporozoites stages of the E. necatrix life cycle. Studies revealed that the synthesis of
viral capsid proteins occurred during the early events of sporulation in Eimeria life
cycle and were packaged into novel viruses during the late events (Wang & Stotish
1975; Pote et al. 1991; del Cacho et al. 2001). The putative coat proteins of E. stiedae
viruses and Leishmaniaviruses were shown to be 41% identical (Stuart et al. 1992,
1993).
There is no evidence that E. nieschulzi virus (ENV-1) particles are infectious to
host parasite (Roditi et al. 1994). Studies were unable to demonstrate the transmission
of the virus in vivo by coinfection of chickens with E. maxima either by co-cultivation
with lysates of E. nieschulzi or electroporation (Roditi et al. 1994). Moreover, chickens
were also infected with E. necatrix sporulated oocysts to determine the presence of both

28
viral particles and capsid proteins but no apparent infection was detected (del Cacho et
al. 2001).
1.14 The RNA viruses of Leishmania Phylogenetically, the genus Leishmaniavirus are classified in the family
Totiviridae and consists of Leishmania braziliensis virus, Leishmania guyanensis virus
(LRV 1–1 to LRV 1–12) and Leishmania major virus (LRV2–1) (Cadd et al. 1993;
Patterson et al. 1995; Scheffter et al. 1995; Zangger et al. 2013) (Figure 1.4 F). The
Leishmaniavirus was detected in cultured promastigotes of these isolates. In 1974,
Molyneux first identified VLPs in sandfly-stage promastigotes of eight separate strains
of Leishmania hertigi using TEM (Molyneux 1974). These VLPs were 55-60 nm in
diameter and found to be associated with induced tubules (Wang & Wang 1991).
Subsequently, an RNA virus was discovered in Leishmania braziliensis (strain
CUMC1-1A) (Tarr et al. 1988). Tarr et al. (1988) identified a multicopy RNA in
Leishmania braziliensis which they termed as LRI and sedimentation coefficient
analysis revealed that these RNA viruses were spherical and 32 nm in diameter. The
promastigote forms of 12 species of Leishmania were examined for the presence of
RNA virus and two of them were found to contain abundant ~ 6000 nucleotides by
agarose gel electrophoresis (Tarr et al. 1988). Widmer et al. (1989) was successful in
isolating an RNA virus by screening various strains of Leishmania for an RdRp activity.
Caesium chloride and sucrose density gradient centrifugation was used to
prepare purified virus of different Leishmaniavirus (Patterson 1990; Weeks et al. 1992;
Cadd & Patterson 1994). MacBeth and Patterson (1995) used caesium chloride purified
or EGTA-treated virus particles to determine the capsid endoribonuclease activity of
Leishmania RNA virus 1-4. In their study, cell lysates were fractionated on 10 to 40%
sucrose gradients and fractions containing viral dsRNAs were detected by RNA
cleavage assays (MacBeth & Patterson 1995). Subcellular fractionation also suggested
that Leishmania RNA viruses were localized in the cytoplasm (Tarr et al. 1988; Stuart
et al. 1992).
Up to now, the complete nucleotide sequences have been reported for
Leishmania RNA virus 1-1 (Stuart et al. 1992), Leishmania RNA virus 1-4 (Scheffter

29
et al. 1994) and Leishmania RNA virus 2-1 (Scheffter et al. 1995). Two types of LRV1
viral particles were found in L. guyanensis which contain double-stranded or single-
stranded RNA (Weeks et al. 1992). It was documented that the genomic organization of
LRV is similar to yeast LA virus and Giardiavirus (Fujimura & Wickner 1988; Wang et
al. 1993; Ro et al. 1997). In brief, the Leishmania viral genome is approximately 5 kb in
length and encodes two large open reading frames (ORFs) on the plus-strand. The first
large ORF is ORF2, which encodes the 82-kDa capsid protein (Diamond et al. 1989).
Open reading frame 3 (ORF3) is predicted to encode a 99-kDa protein containing
conserved RNA-dependent RNA polymerase motifs (Diamond et al. 1989). Initially,
RdRp activity was detected in L. braziliensis guyanensis (Widmer et al. 1989; White &
Wang 1990;). In Leishmaniavirus, the RdRp is expressed as a CP/RdRp fusion protein,
either as a consequence of ribosomal frameshifting (Diamond et al. 1989; Icho &
Wickner 1989) or as a direct fusion with CP (Kang et al. 2001). RNA-dependent RNA
polymerase activity was also employed to screen for viral infections in a number of
Leishmania isolates (Widmer et al. 1989; Scheffter et al. 1995;).
Studies also identified a short RNA transcript in Leishmania spp. and focused on
the nature of this RNA transcript. It could be a product in in vitro polymerase assay or a
by-product of natural virus infection. The precise role of RNA cleavage in the life cycle
of LRV (mainly in LRV1-4) was analysed (Ro & Patterson 2000). Their study
suggested that RNA cleavage alters the functionality of viral transcripts thus affecting
the efficiency of the translation of the viral gene products, in terms of either
enhancement or inhibition.
Similar genomic organization was found in LRV1-1 and LRV1-4. These viruses
have ORF2 (capsid protein) and ORF3 (polymerase) that encode a slippery translation
site and a potential RNA pseudoknot similar to those described for other
Leishmaniavirus (Ro et al. 1997). In addition, the 5´UTR of LRV1-4 transcripts contain
five conserved stem-loop structures (Scheffter et al. 1994; MacBeth 1996) which has
been analysed by ribonuclease mapping and site-specific mutagenesis (Ro & Patterson
2000). In fact, the highly conserved nucleotide level between LRV1-1 and LRV1-4 are
divergence in that LRV1-1 has one small ORF (ORFx) while LRV1-4 has two small
ORFs (ORF1 and ORFx) (Ro et al. 1997). Hybridization experiments also have shown

30
that 12 Leishmania isolates contain related viruses whose genomes have distinct regions
of sequence conservation and divergence (Guilbride et al. 1992; Ro et al. 1997).
Leishmania RNA virus 1 requires a translational frameshift to synthesize a Cap-
Pol fusion protein (Jacks et al. 1988a, 1988b). The LRV1-4 capsid gene encodes an
essential endoribonuclease activity which is responsible for single-site-specific cleavage
of its own viral RNA transcript (Ro & Patterson 2000). Cadd et al. (1993) used antisera
directed either against purified viral particles or baculovirus-expressed capsid protein
and finally detected an 180-kDa fusion-protein (Cadd et al. 1993; Cadd & Patterson
1994). Following studies also identified an immunologically reactive 180-kDa protein
which was presumed to be a capsid dimer. It is still unknown whether a Cap-Pol fusion
protein exists in vivo (Ro et al. 1997).
Gag-Pol-type fusion protein has also been found in Saccharomyces cerevisiae
virus and Giardiavirus (Wang et al. 1993) where the fusion protein is generated via a 21
base translational frameshift (Fujimura & Wickner 1988; Diamond et al. 1989).
Patterson (1993) raised polyclonal antiserum against the viral fraction of LRV1-4
infected cells and used it in western immunoblots to identify viral proteins from various
isolates. Two proteins, 82 and 180 kDa were detected which matched the predicted sizes
of a capsid and a capsid polymerase fusion protein, respectively (Cadd et al. 1993). Lee
et al. (1996) have provided in vitro evidence for an 11 base ribosomal frameshift during
translation of the overlapping gene sequences present on LRV1-4 transcripts.
Afterwards, Ro et al. (1997) identified RdRp which is associated with LRV1-4 VLPs.
The promastigotes of Leishmania hertigi in culture seemed unaffected by the
presence of VLPs even though viral replication had taken place within the cells (Croft &
Molyneux 1979; Wang & Wang 1991). Tarr et al. (1988) did not find any effect of this
potential virus LRI on parasite physiology or the disease caused by the parasite. A
significant reduction of viral number in liquid medium also has no effect on the
parasite’s viability (Widmer 1995).
Leishmaniavirus has been detected in Leishmania species causing human
leishmaniasis. Moreover, Leishmaniaviruses were found in human biopsy samples of

31
leishmaniasis prior to manipulation in culture (Saiz et al. 1998). A reverse transcription
assay was conducted to detect LRV1-1 in a leishmaniasis patient. In addition, a viral
marker may provide a valuable prognostic indicator for human leishmaniasis (Saiz et al.
1998).
Figure 1.4 Electron micrograph showing the appearance of VLPs from different
protozoan parasites. (A) TEM showing Trichomonas vaginalis viruses closely
associated with the Golgi complex (Benchimol 2004) (B) Tritrichomonas foetus viruses
were observed after cytoskeletal affecting drug treatment (Gomes Vancini & Benchimol
2005) (C) Negatively stained G. lamblia viruses obtained by CsCl density gradient
centrifugation (Miller et al. 1988b) (D) Electron micrograph showing the Entamoeba
virus V301 (Mattern et al. 1974) (E) Negatively stained Eimeria necatrix viruses (del
Cacho et al. 2001) (F) Negative staining showing purified Leishmaniavirus (Tarr et al.
1988) (Gc = Golgi complex, VLP= virus-like particle). All arrowheads point to VLPs.

32

33
1.15 Other viruses of protozoa
Viral-like particles have also been described in Babesia bovis (Johnston et al.
1991), Blastocystis sp. (Teow et al. 1992) and in Cryptosporidium parvum (Khramtsov
et al. 1997). Babesia bovis (isolates bovG1 and bovVl) carry a dsRNA virus (~5.5 kb)
that is quite similar to those described in other protozoa. Teow et al. (1992) first
identified the icosahedral VLPs in Blastocystis sp. (Teow et al. 1992). Virus particles
were observed in the cytoplasm of Blastocystis sp. (obtained from monkeys) and the
sizes of these VLPs were approximately 55-60 nm in diameter (Stenzel & Boreham
1991; Teow et al. 1992).
Khramtsov et al. (1997) discovered two linear extrachromosomal dsRNAs
within oocysts of Cryptosporidium parvum (north American isolates). Both dsRNA
genomes were found within the cytoplasm of sporozoites and were 1786 and 1374 kb
long, and encode one large open reading frame. Subsequently, VLPs with a diameter of
31 nm were copurified with RNA polymerase complexes and then polymerase studies
suggested that C. parvum harbors a putative dsRNA virus which separately encapsidate
the large and small RNA segments (Khramtsov & Upton 1998, 2000). The
morphological characteristics and replication mechanism of C. parvum viruses are
similar to those of the viruses of the family Partitiviridae (Khramtsov & Upton 2000).
Based on these criteria, the new genus Cryspovirus was elected for this family (Nibert et
al. 2009). The dsRNAs were considered as molecular markers for C. parvum because of
the widespread distribution, similarity between isolates and high host specificity
(Khramtsov & Upton 2000). They are also used for molecular genotyping of C. parvum
(Khramtsov et al. 2000). Kniel et al. (2004) suggested that C. parvum viral capsid
protein (CPV40) could be a target to detect C. parvum oocysts in water. In their study,
the authors produced a recombinant CPV40 in Escherichia coli, purified it by affinity
chromatography and then used it to prepare polyclonal rabbit sera specific for the viral
capsid protein. Anti-recombinant CPV40 serum recognized a 40 kDa and a 30 kDa
protein in C. parvum oocysts and they appeared to be localized to the apical end of the
parasite (Kniel et al. 2004). Two more viruses, Pepper cryptic virus 1 (PCV-1) and
Pepper cryptic virus 2 (PCV-2), from Cryptosporidium were characterised and share
limited sequence similarity in both genomic segments (Sabanadzovic & Valverde
2011). These two viruses are phylogenetically related and the genome of both viruses

34
consisted of two monocistronic dsRNAs encoding putative RdRp (dsRNA-1) and viral
capsid proteins (dsRNA-2).
Recently, DNA viruses were found with genomes greater than 300 kb and up to
1200 kb (Arslan et al. 2011; Colson et al. 2011a; Legendre et al. 2012). A number of
studies mentioned that the discovery of these viruses were unexpected as their
morphology, life-style and genomic structures are different from each other. These
viruses inhabit aquatic environments and infect bacteria, animals and protists; they are
large viruses (often called giruses) with large dsDNA genomes that encode up to 900
proteins and also many tRNAs (Colson & Raoult 2010; Van Etten et al. 2010; Colson et
al. 2011b). They also have more protein-encoding genes than many bacteria and
protozoan viruses. The well-recognized DNA viruses are: (1) Mimivirus, which infects
amoebae or marine protists and has a genome ~1.2 Mb (La Scola et al. 2008; Claverie
& Abergel 2009, 2010; Claverie et al. 2009); (2) Mamavirus which has an 18.3-kb
DNA (La Scola et al. 2008); (3) Marseillevirus, which also infects amoebae (Claverie &
Abergel 2009) ; (4) Phycodnaviruses, which infect algae and has genomes up to 560 kb;
(5) White spot shrimp viruses (WSSV) which have genomes up to 305 kb (Boyer et al.
2011); and (6) viruses that infect bacteria and have genomes up to 670 kb. Originally,
Mimivirus was thought to be a parasitic bacterium (Claverie & Abergel 2009, 2010).
Definitely, many large viruses are expected to be discovered and there is a need for
further research to be done on these large viruses.
1.16 Virus as transfection vector Viruses can be used as vectors for gene delivery and protein-based vaccine
production in plants and animals including protists. Viruses infect cells and transport
their genomes inside the cells (Dan et al. 2000; Hoffmann et al. 2000; Kurth et al.
2012).
The potential of creating a vector for the genetic transformation of G. lamblia
using its virus has been discussed in detail (Yu et al. 1995a, 1995b; Vanacova et al.
2003). Wang et al. (1993) isolated a single-stranded full-length copy of Giardia viral
RNA which can be electroporated into virus free G. lamblia cells to generate infectious
progeny virus particles (Furfine et al. 1989). Consequently, a transfection vector was
created and transferred into Giardia trophozoites via electroporation; the transcript

35
replicates inside the transfected cells in the presence of the wild type dsRNA virus (Yu
et al. 1995b). In order to transfect Giardia via electroporation, the Giardia dsRNA
viruses were elegantly engineered to introduce and express both exogenous and
endogenous genes (Yu et al. 1996a, 1996b; Vanacova et al. 2003). Liu et al. (2005)
developed a transient GLV-based RNA transfection system for G. lamblia which could
be used as a genetic tool for the study of possible intervention and pathogenesis.
Similarly, Giardia canis virus (GCV) was used to construct a stable transfection system
to transfect the trophozoites of G. canis (Chen et al. 2007; Liu et al. 2008). Thus GCV
could be useful tool for gene manipulation of G. canis.
Only a few studies have investigated whether overproduction of foreign mRNA
in GLV infected Giardia trophozoites can be exploited as a de facto gene knockout
system. It has been suggested that hammerhead ribozyme flanked by antisense RNA
against a specific mRNA of interest could be used for this purpose (Zaug et al. 1986;
Dan et al. 2000). The development of antisense RNA technology to inhibit specific gene
expression in Giardia is also described (Dan et al. 2000; Dan & Wang 2000). They also
checked a knock out of the PFOR ( Pyruvate-Ferredoxin Oxidoreductase) gene
expression in Giardia by cleaving its mRNA with a hammerhead ribozyme flanked by
PFOR antisense RNA replenished using a Giardia viral vector (Dan & Wang 2000).
1.17 Concluding remarks Gut protozoan parasites cause serious disease in humans and other animals and
are an increasing public health concern. To our knowledge, there are no recent studies
which reviewed the relationship of these existing protozoan viruses in detail. Numerous
studies show that gut protozoa are persistently infected by viruses that presumably have
co-evolved with the protozoa. Over the last decades, interesting new findings have
expanded studies to the anaerobic flagellates T. vaginalis, G. lamblia, the kinetoplastid
Leishmania and the sporozoan genus Eimeria. A wide variety of parasitic protozoa can
therefore harbor viruses. This current review provides an overview of gut protozoan
viruses; extra-intestinal Leishmaniaviruses was also included in this review because of
their significant availability and relationship with other protozoan viruses.

36
Although all of the RNA viruses found in gut protozoa have some interesting
similarities, their biological significance remains poorly understood. Previous studies
have shown that a number of protozoan viruses are similar to yeast dsRNA killer
viruses (Wang & Wang 1991). These dsRNA viruses are now known to be associated
with killer phenotypes in yeast genera (Tipper & Schmitt 1991). At present, it is not
clear whether the presence of protozoan viruses has any specific effect on the antigenic,
physiological and pathological properties of protozoa. It will be interesting to examine
the specific effect of these protozoan viruses on their host. Thus, more studies need to
investigate the viral replication, transcription and packaging in different protozoa in
detail. In addition, more sequence analysis of the viral genes from these protozoan
viruses in future studies will certainly provide some clues in relation to their effects in
human gut diseases.
1.18 Aims: The aims of this thesis were:
1) To investigate the morphology and ultrastructure of D. fragilis by electron
microscopy and analyse different nuclear conditions using confocal microscopy
(chapter 2).
2) During ultrastructural observation of D. fragilis by electron microscopy, virus-
like particles (VLPs) were discovered in cultured D. fragilis trophozoite. So
another aim was to describe, by transmission electron microscopy, the presence
and morphological details of the virus population found in different clinical
isolates of D. fragilis (chapter 3). The assembly pathway of VLPs in D. fragilis
was also described in detail.
3) Another objective was to compare different extraction methods to recover
dsRNA from T. vaginalis B7268 isolate for use as a positive control and to
evaluate these optimized methods to identify D. fragilis viral nucleic acid
(chapter 4).
4) The epidemiology of D. fragilis has not been studied in detail. While I was
studying the D. fragilis VLPs, I also conducted a case-controlled study to
determine the clinical signs associated with D. fragilis infection in children
presenting to a Sydney Hospital. I am adding the results in this thesis for
completeness of the study (chapter 5).

37
Dientamoeba fragilis was recognized as a significant human pathogen almost a
century ago (Jepps & Dobell 1918). Most of the microscopic observations of D. fragilis
date back to the early and mid-1900s and therefore may represent inappropriate
descriptions of this parasite. Understanding the cell biology of this protozoan in areas
such as surface structure, ultrastructure, subcellular organization and detailed
observations on nuclear characteristics need to be carried out. This study will be
important in providing a better knowledge on cell biology of this neglected parasite.

38
Chapter 2:
A microscopic description of Dientamoeba fragilis
Published as
Banik, G. R., Birch , D., Stark, D., Ellis, J. T., 2011.
A microscopic description and ultrastructural characterisation of
Dientamoeba fragilis: An emerging cause of human enteric
disease. International Journal for Parasitology, 42, 139-153.

A microscopic description and ultrastructural characterisation ofDientamoeba fragilis: An emerging cause of human enteric disease
Gouri R. Banik a,b,c, Debra Birch d, Damien Stark a,c, John T. Ellis b,c,⇑aDivision of Microbiology, SydPath, St. Vincent’s Hospital, Darlinghurst, AustraliabUniversity of Technology Sydney, i3 Institute, Broadway, AustraliacUniversity of Technology Sydney, School of Medical and Molecular Biosciences, Broadway, Australiad Faculty of Science, Macquarie University, Sydney, New South Wales, Australia
a r t i c l e i n f o
Article history:Received 6 August 2011Received in revised form 14 October 2011Accepted 17 October 2011Available online 26 November 2011
Keywords:Dientamoeba fragilisElectron microscopyUltrastructureTrichomonadDientamoebiasisVirus-like particle
a b s t r a c t
Dientamoeba fragilis is a pathogenic trichomonad found in the gastrointestinal tract of humans and isimplicated as a cause of diarrhoea. Despite its discovery over a century ago, there has been no recentthorough description of this parasite by microscopy. Scanning electron microscopy, transmission electronmicroscopy, confocal and light microscopy were therefore used to characterise D. fragilis populationsgrowing in xenic culture. Two different populations – smooth and ruffled cells – were identifiable byscanning electron microscopy. No flagella, pelta structures, undulating membrane or pseudocyst-likeforms were present. The organelles in D. fragilis were analysed by transmission electron microscopy; likeTrichomonas and Histomonas, D. fragilis contains hydrogenosomes that presumably represent the site ofanaerobic respiration. The nuclear morphology of D. fragilis trophozoites grown in vitro and trophozoitesfrom clinical isolates were also compared by confocal microscopy and light microscopy. The majority ofcells grown in culture were mononucleate while most cells in permanent stained faecal smears werebinucleate. The two nuclei of D. fragilis are morphologically indistinguishable and contain equivalentamounts of DNA as determined by DAPI staining. The approximate cell and nuclear volume of four iso-lates of D. fragilis were measured and shown to be comparable to other trichomonads. In addition, thediscovery of a virus-like particle is reported, to our knowledge for the first time in D. fragilis. This studytherefore provides extensive and novel details of the ultrastructure of a neglected protozoan parasite thatis an emerging cause of human disease.
� 2011 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Dientamoeba fragilis is a trichomonad protozoan that belongs tothe Parabasalia group (Gerbod et al., 2002; Ohkuma et al., 2005;Cepicka et al., 2010). It is found in the gastrointestinal tract of hu-mans where it is associated with chronic gastrointestinal diseaseand has worldwide distribution (Stark et al., 2010a,b; Barrattet al., 2011a). It is almost a century since D. fragilis was first de-scribed by Jepps and Dobell who initially classified it as an amoeba(Jepps and Dobell, 1918; Johnson et al., 2004). Studies related tomolecular phylogenetics (Delgado-Viscogliosi et al., 2000; Gerbodet al., 2001) and morphological analysis (Camp et al., 1974) thenconfirmed that D. fragilis was a trichomonad that has no flagellain the trophozoite stage of its life cycle. The complete life cycleof this parasite has not yet been fully determined and the tropho-zoite is the only known stage (Stark et al., 2008; Barratt et al.,2011b). Morphologically, D. fragilis is a single-celled pleiomorphic
trophozoite, typically ranging from 5 to 15 lm in diameter, thatcontains one to four nuclei (Sawangjaroen et al., 1993; Johnsonet al., 2004; Stark et al., 2006). The presence of five nuclei is alsodescribed (Dobell, 1940; Wenrich, 1944a; Moody and Fleck,1985). Characteristically high percentages of D. fragilis trophozo-ites are binucleate with a large, fragmented, central karyosomewithout peripheral chromatin observed previously in preparationsfrom stained faeces (Stark et al., 2006). It is not possible to see nu-clear structure in an unstained preparation (Windsor and Johnson,1999; Stark et al., 2008). Moreover, the nuclei of D. fragilis are notvisible in saline or iodine preparations, although food vacuoles orinclusions may be seen (Johnson et al., 2004). No cyst stages havebeen reported yet for this parasite (Windsor et al., 2003; Starket al., 2006; Barratt et al., 2011b).
One of the unusual features of D. fragilis is the presence of twonuclei (binucleate forms) (Johnson et al., 2004; Stark et al., 2010a).Previous studies reported that another protozoan parasite, Giardialamblia, also has two equal sized nuclei which are equivalent withrespect to the amount of DNA harboured in each and replicate atapproximately the same time (Wiesehahn et al., 1984; Kabnick
0020-7519/$36.00 � 2011 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.ijpara.2011.10.010
⇑ Corresponding author. Tel.: +61 (02) 95144161; fax: +61 (02) 95144003.E-mail address: [email protected] (J.T. Ellis).
International Journal for Parasitology 42 (2012) 139–153
Contents lists available at SciVerse ScienceDirect
International Journal for Parasitology
journal homepage: www.elsevier .com/locate / i jpara

and Peattie, 1990; Cerva and Nohynková, 1992; Svärd et al., 2003;Benchimol, 2004b). This organism represents one of the earliestdiverging lineages of eukaryotes (Adam, 2001; Svärd et al., 2003).There are other protozoans which have been studied in more de-tails with two or more nuclei. These include the ciliates, Tetrahy-mena and Paramecium (Southern, 1975; Prescott, 1994);Tetrahymena has one diploid micronucleus, and Paramecium hastwo equal, diploid micronuclei (Jahn and Klobutcher, 2002).Dientamoeba fragilis was reported as typically binucleate almost acentury ago (Jepps and Dobell, 1918; Craig, 1926) but the nucleihave not been studied with regard to DNA content or transcrip-tional activity. It is important to determine whether the two nucleiof D. fragilis are structurally and functionally identical as well as tounderstand the evolutionary, physiological and genetic signifi-cance of these two physically separate, but equal-sized, nuclei.
To our knowledge, only a few studies have described the ultra-structure of D. fragilis by transmission electron microscopy (TEM)(Camp et al., 1974; Ockert and Schneider, 1974; Silard et al.,1984; Silard and Burghelea, 1986). Camp et al. (1974) publisheda study of the binucleate stage of D. fragilis (strain Bi/pa) and theultrastructure of mononucleate stages (CE strain of D. fragilis) weredescribed by Silard et al. (1984).
New studies need to be undertaken due to recent advances inthe cell biology of trichomonads. For example, hydrogenosomesare known features of a trichomonad cell but they have yet to bedescribed for D. fragilis. To date, there are also no studies which re-port the scanning electron microscopy (SEM) characteristics of D.fragilis.
This study aimed to describe and characterise D. fragilis by dif-ferent microscopical methods in order to define cell size, shape,surface organisation and ultrastructure of different organellesincluding nucleus numbers. SEM, TEM, confocal and light micros-copy was used to examine these characteristics and the current re-sults are reported here. The ultrastructure of four isolates of D.fragilis is described. To our knowledge this study is the first SEManalysis of D. fragilis which contributes to the understanding ofthe surface biology of this parasite. This study provides new in-sights into the cell biology and many novel morphological aspectsof D. fragilis and thus allows better understanding of the main as-pects of its cell biology.
2. Materials and methods
2.1. Culture of D. fragilis trophozoites
Four D. fragilis isolates (isolates 1–4) used in this study werepreviously described (Barratt et al., 2010). All D. fragilis isolateswere grown anaerobically at 37 �C in tissue culture flasks withanaerobic or microaerophilic gas packs (Oxoid AnaeroGen™AN0035A (Oxoid, Australia)) in an anaerobic jar (BD GaspakTMEZ (BD, Australia)). Loeffler’s serum slopes containing heat-inacti-vated horse serum (Bovogen, Australia), glucose (Sigma, Australia)and nutrient broth (Sigma, Australia) in distilled water were pre-pared as previously described (Barratt et al., 2010). Fifteen ml ofLoeffler’s medium were poured into a 50 ml culture flask (Nunc™156367 (Thermo Fisher, Australia)), sloped and inspissated in an85 �C drying oven until the serum slopes solidified. Finally, themedia was completed by addition of 15 ml PBS (Sigma) and sup-plemented with 3–5 mg of sterilised rich starch (Sigma S7260).
2.2. Scanning electron microscopy
Cell suspensions of 1.0 � 106 trophozoites/ml were collectedafter 48 h of subculture and concentrated in 1.5 ml Eppendorftubes (100 ll) which were then centrifuged at 1,000g for 5 min.
The supernatant was removed and cell pellets were fixed in 5%(v/v) glutaraldehyde in PBS (0.1 M, pH 7.2) overnight at room tem-perature. The cells were then washed three times for 10 min eachwash with PBS. To prepare cells for SEM, coverslips (12 � 12 mm)were coated with 0.1% ethylene imine polymer solution (Fluka,Switzerland). Cell suspensions (20 ll) from each isolate wereadded to the coverslips and cells were allowed to adhere for10 min at room temperature. The adherent cells were then washedin buffer three times for 10 min each wash (0.1 M phosphate buf-fer, pH 7.2) and post-fixed in 1% osmium tetroxide (OsO4) in0.1 M PBS (pH 7.2) for 1 h. Following post-fixation, the sampleswere washed in the same buffer three times for 5 min each andthen dehydrated through a graded series of ethanol of 30%, 50%,70%, 90%, 95% and 100%, for 10 min each step. One set of sampleswas transferred to a critical point dryer (Emitech K850 (ProSciTech,Australia)) and the other set dehydrated using hexamethyldisilaz-ane (HMDS, Sigma). The coverslips were mounted on aluminumstubs and sputter coated with gold (Emitech K550). Observationswere carried out using a JEOL 6480 LA scanning electron micro-scope. All SEM materials were supplied from ProScitech, Australia.
2.3. Investigation of pseudocyst-like stages formation by SEM
Previous reports noted that cooling of cultures of trichomonadsfrom their normal growth temperature of 37 �C to below 16 �C(Granger et al., 2000), 4 �C for Histomonas (Zaragatzki et al.,2010) or 20 �C for Monocercomonas sp. (Borges et al., 2007) cantrigger pseudocyst formation. To investigate the effects of temper-ature on the life cycle of D. fragilis, trophozoites were incubated at4, 16 and 20 �C for 48 h. The initial trophozoite density was1.0 � 106 trophozoites/ml and the trophozoites were counted in ahemocytometer every 24 h. All experiments were performed intriplicate. Controls were incubated under normal in vitro cultiva-tion at 37 �C. Post-incubation cells were fixed and processed forSEM as above.
2.4. Transmission electron microscopy
Cell suspensions (1.0 � 106 trophozoites/ml) from four D. fragi-lis isolates were collected after 48 h of subculture into 1.5 mlEppendorf tubes (100 ll) and were then centrifuged at 1,000g for5 min. The supernatant was removed and cell pellets were fixedin 5% (v/v) glutaraldehyde in PBS (0.1 M, pH 7.2), overnight at roomtemperature. Fixed cells were centrifuged at 500,000g for 5 min,the fixative was removed and fresh buffer added. The buffer wash-ing step was repeated three times for 15 min each wash. Cell pel-lets were embedded in 1% low melt agarose (J.T. Baker inc.,Phillipsburg, USA). The agar blocks were cut into 1 mm cubes,transferred to glass vials and post-fixed in 1% osmium tetroxide(OsO4) in 0.1 M PBS for 1 h at room temperature. Following post-fixation, the samples were washed in distilled water three timesfor 5 min each and immersed in 2% aqueous uranyl acetate for30 min. The pellets were dehydrated through a graded series ofethanols (50–100%), infiltrated and subsequently embedded inLondon White Resin (L.R. White). Semi-thin and ultra-thin sectionswere cut using a Reichert ultramicrotome (Ultracut, Leica Micro-systems, Germany). Semi-thin sections (1 lm thickness) werestained with 1% methylene blue and observed by transmitted lightmicroscopy (Olympus BH2). Ultra-thin sections (70 nm) weremounted onto Pioloform coated, 300 mesh, thin-bar copper grids,stained with saturated aqueous uranyl acetate (7.7%) for 30 minand Reynold’s lead citrate for 4 min. Ultrathin sections were exam-ined using a Philips CM10 transmission electron microscope (Eind-hoven, the Netherlands). All TEM materials were supplied fromProScitech, Australia.
140 G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153

2.5. Nuclei, cell cytoplasm and cell membrane staining of Dientamoebafragilis
2.5.1. Nuclear staining with DAPIDientamoeba fragilis trophozoites in the logarithmic phase of
growth and exhibiting more than 95% viability were collected bycentrifugation at 1,000g for 5 min. The cell pellets were washed inPBS and initial trophozoite density was 1.0 � 106 trophozoites/ml(as counted in a hemocytometer). For DAPI staining, all D. fragilisisolates were fixed in sodium acetate acetic acid formalin (SAF)at a 1:1 ratio for 3–4 h. Fixed cells were washed with isotonic sal-ine (0.9% NaCl) two times at 500g for 5 min. After washing, the cellswere centrifuged at 1,000g for 5 min and resuspended in a mini-mum amount of saline. A drop of the cell suspension was appliedto a microscope slide which was then mixed with a drop of 0.5%albumin (Mayer’s Albumin solution, Fronine, Australia 2604029)and spread over the slide to create a smear. The slides were al-lowed to air dry for approximately 3–4 h to adhere the cells ontothe slides. The fixed cells were stained with 1 lg/ml DAPI (Invitro-gen, Australia) in the dark for 15 min at room temperature. Thecells were rinsed carefully with PBS to remove unbound dye. Ex-cess liquid was removed from the slide by gently blotting aroundthe sample with an absorbent tissue. A glass coverslip was placedon the slide and the edges sealed with wax or nail polish. DAPIstained smears were then viewed under a Laser Scanning ConfocalFluorescence Microscope (Nikon A1).
2.5.2. Nile Red stainingCultured trophozoites were fixed in SAF solution and micro-
scopic slides were processed for DAPI staining as above. Stock solu-tions of Nile Red (9-diethylamino-5H-benzo[a]phenoxazine-5-one)(Sigma) (1,000 lg/ml) were prepared in acetone and stored pro-tected from light. Nile Red was added directly to the trophozoitepreparation at l:100 dilution. Initially, cells were stained with DAPIand then immediately stained with Nile Red. The slides were incu-bated at room temperature for a minimum of 10–15 min. Excessdye was removed by brief rinsing in PBS. A coverslip was addedand the samples were examined using a Laser Scanning ConfocalFluorescence Microscope (Nikon A1). The suspension medium didnot contain serum or albumin since this was found to act as a sinkto draw Nile Red out of the cells.
2.5.3. Permanent stained smearsHuman stool specimens were fixed in SAF solution and stained
with a modified iron–haematoxylin stain (Fronine, Australia)according to the manufacturer’s recommendations. All stainedsmears were examined by oil-immersion microscopy at 1000�magnification. Photographs were taken using Olympus DP70 photomicroscope connected to a camera (Leica DC 300F) at magnifica-tion 100� Differential Interference Contrast (DIC). Cell diameterswere measured using Image Pro Express Software.
2.5.4. Photometry and data analysisAll SEM and TEM images were analysed and measured quantita-
tively by using ImageJ Software (1.44). The confocal fluorescencemicroscope equippedwith afilter forDAPI analysis (laser: excitation400 nm and emission 450–475 nm) was used. Cells were photo-graphed atmagnification 100�DICwith the pinhole open.Mononu-cleate and binucleate cells were viewed in a three-dimensional (3D)focal plane (X–Y and Y–Z) using Z- Stack optical sectioning duringscanning by confocal microscopy.
The DAPI fluorescence emitted from each nucleus was quanti-tated photometrically as previously described (Kabnick andPeattie, 1990). The dimension (length and width) of each cell wasmeasured by using 3D measurement tools in the NIS ViewerSoftware.
2.5.5. Estimation of cell and nucleus volumeCell dimensions as well as approximate cell volume were mea-
sured for all four isolates of D. fragilis. All four D. fragilis isolateswere attached to microscope slides that were fixed with SAF solu-tion (as described for DAPI staining method) and observed using acombination of DIC illumination and fluorescence of DAPI-stainedtrophozoites in a confocal microscope (Nikon A1). Photographswere taken of 50 non-dividing cells of each D. fragilis isolate. Thedimensions of cells and nuclei were made using NIS Viewer Soft-ware. Approximate cell and nucleus volumes (V) were calculatedaccording to the formula V = 3/4pab2 where ‘‘a’’ is the cell length,and ‘‘b’’ is the cell width as previously described (Zubácová et al.,2008). S.D. of cell length and width of 50 non-dividing cells fromeach isolate were determined.
3. Results
3.1. Characterisation of cell surfaces of D. fragilis
The external structures of four isolates of D. fragilis studied bySEM are described. Firstly, the cell shapes and sizes of four clinicalisolates of D. fragilis trophozoites (Fig. 1) were examined. By SEM,the cell shape of the four isolates of D. fragilis varied from sphericalto ovoidal, sometimes amoeboid ranging in size from approxi-mately 4–10 lm. There were no flagella, undulating membrane oraxostyle–pelta structures. Other typical characteristics of D. fragilisobserved were bacteria adherent to D. fragilis surfaces and phago-cytosis of bacteria and rice starch (Fig. 2).
3.2. Phagocytic activity of D. fragilis
It was observed by SEM that trophozoites in fresh culturesexhibited phagocytosis (Fig. 2). The attachment of bacteria to thecell surface of the parasite was observed frequently (Fig. 2C). Dient-amoeba fragilis constantly changes its shape and engulfs bacteriaand rice starch available from the culture medium. It also exhibitsan amoeboid morphology during the internalisation of food parti-cles. A variety of motile forms of D. fragilis were observed. Mostlytwo forms of phagocytosis were observed: a ‘sinking’ process with-out any apparent participation of plasma membrane extensions(Fig. 2A and B), and the classical phagocytosis where pseudopodiawere extended by amoeboid movement toward the target foodparticles (Fig. 2D). Interestingly, D. fragilis was also able to phago-cytose even when undergoing the division process (Fig. 3C and D).
3.3. Two different populations of D. fragilis
Under the SEM, two types of D. fragilis populations were classi-fied based on cell surface structure: smooth cells and ruffled cells(Fig. 1E and F). To examine the relative numbers of both cell types,a time interval experiment was performed by collecting cells at dif-ferent time points of culture (6, 12, 24, 32, 48 and 72 h). Ruffledcells were more common than smooth cells. During the time inter-val, from 6 to 48 h, 90% of the cells were ruffled while at 72 h, 30%of cells appeared to be smooth.
3.4. Growth stages of D. fragilis
In addition, different stages of development and activities of D.fragilis trophozoites were observed during this experiment (Fig. 3).Phagocytosis was the most common phenomenum observed inmost of the cells at 6 and 12 h. A number of trophozoites dividedby binary fission which occurred by the simple constriction ofthe cell body (Fig. 3C and D). Ninety-eight percent of trophozoiteswere considered to have finished their binary fission between 32
G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153 141

Fig. 1. Scanning electron micrograph showing the external surface structures of four isolates of Dientamoeba fragilis. (A) Isolate 1, (B) Isolate 2, (C) Isolate 3 and (D) Isolate 4.Two different populations of Dientamoeba fragilis exist; (E) smooth cell and (F) ruffled cell.
Fig. 2. Scanning electron micrograph showing two forms of phagocytosis of Dientamoeba fragilis. (A and B) Formation of a phagocytic cup (arrow) and moving to foodparticles without extension of the plasma membrane. (C and D) Attachment of bacteria to the parasite occurs via the whole cell surface with pseudopodia which engulfbacteria. (b, bacteria; Rs, rice starch).
142 G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153

and 48 h. Due to the lack of nutrient in the cell culture medium at72 h, trophozoites were probably approaching stationary phase atthis time point (Fig. 3F).
3.5. Pseudocyst-like formation
This experimental system was used for the study of rapid andreversible formation of pseudocyst-like stages in culture in re-sponse to unfavourable culture conditions. To induce encystation,D. fragilis was cultivated for 48 h at either 4, 16 or 20 �C and thepopulations analysed by SEM. The comparison showed variablesize differences between trophozoites grown under the differentconditions. Nearly all of the cells appeared non-viable, damagedand smaller in size than normal cells grown at 37 �C. The cell wallsurface appeared to be non-intact under these adverse conditions.It was concluded that pseudocysts were not present in thesecultures.
3.6. Transmission electron microscopy: ultrastructural observations onfour isolates of D. fragilis
The morphological characteristics, such as cell shape, size andultrastructure of different organelles including nuclei numbers offour isolates of D. fragilis collected from in vitro cultures wereexamined (Fig. 4A–C) (Table 3). Usually D. fragilis has a sphericalor oval shape with a granular, vacuolated cytoplasm and some cells
are amoeboid. Dientamoeba fragilis exhibited different motile formswith visible pseudopodia (Fig. 4D). The cell size varied from 3 to15 lm and the nucleus diameter from 0.8 to 3.5 lm in all four iso-lates. The ultrastructure of 100 cells (25 cells from each isolate)was examined in detail and more than 80% of cells from each iso-late (isolate 1: mononucleate cells 96%, binucleate 4%; isolate 2:mononucleate cells 84%, binucleate 16%; isolate 3: mononucleatecells 92%, binucleate 8%, isolate 4: mononucleate cells 92%, binu-cleate 8%) were mononucleate. A small number of trinucleateand anucleate cells were also observed.
3.6.1. NucleusThe size of the nucleus in mononucleated trophozoites varied
from 0.86 to 3.52 lm and in binucleated cells from 1.12 to2.06 lm. A number of mononucleated cells appeared to have divid-ing nuclei and nucleoli (Fig. 5C and D) and were larger than binu-cleated nuclei. The nucleus in non-dividing mononucleate andbinucleate cells was spherical but some oval or irregularly-shapednuclei were observed. The nuclear envelope consisted of a doublemembrane containing numerous nuclear pores (Fig. 5A, arrow).The nuclear matrix was filamentous and contained electron densematerial.
The nucleus of D. fragilis is normally seen in the central region ofthe cell. Both mononucleate and binucleate trophozoites have frag-mented nuclei and usually contain two to eight chromatin bodies(Fig. 5A) without peripheral chromatin. Chromatin bodies were
Fig. 3. Scanning electron micrograph showing the different stages of Dientamoeba fragilis in culture at different time intervals. (A and B) Cells at early stages of culture (6–12 h) showing phagocytosis and engulfing rice starch (Rs) and bacteria (b). (C and D) Trophozoites dividing by binary fission; the arrow indicates that it can stretch like abridge. (E and F) At 48 and 72 h of cultures cells are rounded.
G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153 143

found scattered throughout the nucleus and varied in size. Thenucleolus was mainly located in the periphery of the nucleus, notat the polar center, and appeared as a rounded structure. It con-sisted of a dense fibrillar component and was not surrounded bya membrane. There appears to be only one nucleolus per nucleus(excluding the dividing stages) (Fig. 5C).
The mononucleate form of D. fragilis is the predominant stageobserved in in vitro cultures. A number of cells appeared to bedividing by binary fission (Fig. 5B). The mechanism of this divisionwas the simple constriction of the cell body. Nuclear division was
observed only in mononucleated trophozoites. The extranuclearspindle was found between the nuclei in binucleate cells, emanat-ing from the polar complex adjacent to one of the nuclei (Fig. 5B, ar-row). It extended between the two nuclei, each nucleus surroundedby an envelope with nuclear pores. The spindle microtubulesoriginated in pairs and non-periodic structures. Approximately20–30 microtubules were often seen as clusters and occasionallyin ordered arrays. Different types of microtubules were observedthroughout the cell: pole-to-pole, pole-to-nucleus and pole-to-cytosol microtubules. The spindle microtubules were assembled
Fig. 4. Transmission electron micrograph showing the fine ultrastructure of Dientamoeba fragilis trophozoite (A) mononucleate (B) binucleate (C) anucleate (D) amoeboid –like with pseudopodia. (B, bacteria; Ch, chromatin bodies; Dv, digestve vacuole; G, glycogen; H, hydrogenosomes; L, lysosome; My, myelin sheath; N, nucleus; Pf, parabasalfilament; Ps, pseudopodia; Rs, rice starch).
Fig. 5. Transmission electron micrograph showing the nuclear structure of a Dientamoeba fragilis trophozoite. (A) Nuclear structure. Nm, nuclear membrane; Ch, chromatinbodies; Np, nuclear pore (arrow). (B) Formation of an extranuclear spindle (arrow) during nuclear division. (C) Arrow indicates binary fission. (D) Nucleolus dividing; Nu,nucleolus.
144 G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153

from a attractophore (V-shaped center) which was located under-neath the basal bodies (Fig. 8A). No kinetosomes or centriole-likeorganelles were found in the polar complexes or elsewhere in theorganism.
3.6.2. The Golgi complexA well-developed Golgi complex was found in all isolates of D.
fragilis (Fig. 6A). The Golgi complex was observed as a vesicularstructure, generally in the perinuclear area and in close proximityto the endoplasmic reticulum (ER). Golgi vesicles were also seen inthe area between branching bundles of microtubules. The Golgi ofD. fragilis had the following characteristics: it was a single and veryprominent structure, approximately 450 nm long and it had 7–10cisternae. The Golgi complex appeared to be better developed inthe mononucleated cells. On the other hand, in binucleated cells,the Golgi complex did not appear in an organised form and wasmostly fragmented. Numerous circular vesicles could be seen inproximity to the Golgi complex.
3.6.3. Endoplasmic Reticulum (ER)Both smooth and roughERswerewell-developed in themononu-
cleate and binucleate D. fragilis trophozoite (Fig. 6B and C). It wasclearly observed in all four isolates. Rough ER was more frequentlyfound in the cytoplasm of mononucleated cells. The ER was alsoclearly seen around the nucleus and sometimes closely associatedwith the hydrogenosomes, food vacuoles and microtubules.
3.6.4. HydrogenosomesTEM showed the presence of hydrogenosomes inD. fragilis, while
mitochondria and peroxisomes were not observed. Hydrogeno-somes are double layered membrane-bound electron-dense organ-elles located in the cytoplasm of D. fragilis. These were found in allfour D. fragilis isolates used in this study, sizes ranging from 0.12to 0.83 lmand the number varying from5 to 15 (Fig. 6D and E). Thisorganelle was spherical or oval shaped or sometimes slightly elon-gated in structure. The membrane of the hydrogenosome wassmooth, while the matrix of the hydrogenosome was homoge-neously granular. Hydrogenosomes with a variety of electron den-sities were present. They possessed a peripheral vesicle, althoughit is clear that the shape of these organelles largely depends onthe plane of section. They were usually associated with cytoplasmicinclusions and with digestive vacuoles. Close proximity, and evencontinuity, between the ER and hydrogenosomes was observed.
3.6.5. Digestive vacuole (DV)/food vacuoleDigestive or food vacuoles (DVs) were commonly found
throughout the cytoplasm and could contain bacteria, rice starchand myelin configurations (Fig. 7A and B). Their size varied from0.59 to 4.2 lm. The numbers of DVs were seen to vary from 1 to10 in all four isolates. Rice starch was comparatively small and ob-served throughout the cells. A number of digestive vacuoles con-tained rice starch and bacteria, clearly recognisable in the earlystages of digestion. Dientamoeba fragilis feeds by phagocytosis
Fig. 6. Organelles of Dientamoeba fragilis trophozoite (A) Golgi complex; (B) Smooth endoplasmic reticulum (ER); (C) rough ER attaching ribosomes on its surfaces; (D)hydrogenosomes in cytoplasm; (E) double layered membrane-bound electron dense hydrogenosome at higher magnification; (F) lysosome located near cell membrane. (Dv,digestive vacuole; L, lysosome; Gc, Golgi complex; N, nucleus; r, ribosome; m, cell membrane; H, hydrogenosome; ED, electron dense materials).
G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153 145

(Fig. 7C and E), and waste products are released from the DVs byexocytosis (Fig. 7F). It was also observed that Dientamoeba tropho-zoites were bound by a double layered cell membrane (Fig. 8F).This membrane did not give rise to the cell mouth (micropore,cytostome) but appeared to form pinocytotic vesicles and smallfood vacuoles, which engorged material from the culture media.
3.6.6. LysosomesLysosomes were present in different sizes (0.50–2.0 lm) mainly
in the posterior region of the cell in close proximity to the cellmembrane (Fig. 6F). They were also seen in close proximity toDVs and other cytoplasmic organelles. The internalised bacteriaand rice starch were digested in lysosomes.
3.6.7. Cytoplasm and other cytoplasmic inclusionsThe cytoplasm contained large deposits of glycogen, electron
dense materials and ribosomes and was surrounded by a doublelayered cell membrane. During phagocytosis, an invagination ap-peared in the cytoplasm and gradually its edges were drawn to-gether (Fig. 7C). Glycogen granules of different sizes weredistributed throughout the cytoplasm (Fig. 4A). Rough ER was lessfrequently identifiable and not concentrated in the perinuclear re-gion. Electron dense materials (Fig. 8F) were scattered throughoutthe cytoplasm. The central zone of some cells was completely filledwith the electron dense materials.
3.6.8. The basal body structure: axostyle-costaThe basal body structure of Dientamoeba contains an axostyle
and a costa. The pelta structures or flagella were not found. Multi-ple ribbons of microtubules formed the axostyle, which ran fromthe basal bodies to the cell tip (Fig. 8A, arrow) and outlined the axisof the cell. The axostyle was usually found in the anterior region ofthe cell near the cell membrane. The axostyle was also observed inthe middle of cells adjacent to the nucleus. The anterior portion ofthe axostyle was wider than the posterior region. It was also asso-ciated with other organelles such as ER, hydrogenosomes and sig-moid filaments. It is noteworthy to mention that the completestructure of the axostyle was very seldom seen. The appearanceof the axostyle under TEM was dependent on the plane of section.The costa, another basal body cytoskeletal structure, is periodicand proteinaceous and formed by a microtubular sheet. The costawas frequently found near the cell surface and in the middle regionof the cell (Fig. 8D). It comprised a complex array of filaments andoften made contact with a network of other filaments present inthe cytoplasm.
3.6.9. Parabasal filamentThe parabasal filament (PF) is another component of the basal
body cytoskeleton and was found in all isolates of D. fragilis. It ap-peared as a microfibrillar strand linked to the basal bodies. Thisstructure was seen as singular (Fig. 8E) or in a cluster (Fig. 8B),
Fig. 7. Digestive vacuole of Dientamoeba fragilis trophozoite. (A) Presence of digestive vacuoles (Dv); (B) containing rod-shaped bacterium (B) and myelin sheath (My) indigestive vacuole; (C) phagocytosis (arrow) of D. fragilis, engulfing bacterium; (D) formation of phagosome (Ph); (E) engulfing rice starch; (F) exocytosis phenomenon (arrow).(b, bacteria; c, cytoplasm; Rs, rice starch; Pm, plasmalemma).
146 G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153

close to the nucleus and Golgi complex and extended laterally tothe external surface of the atractophores. Clusters of the parabasalapparatus were frequently observed in all isolates, often underly-ing a V-shaped structure (attractophore) (Fig. 8A). The PF was com-posed of approximately 30–40 hair-like segments. Occasionally, itwas composed of two bundles; one bundle appeared at some dis-tance from the nucleus, whereas the other was juxtanuclear andwas often seen in a groove of the nuclear envelope.
3.6.10. PhagocytosisA number of trophozoites showed irregularly protruding pseu-
dopodia by TEM. An amoeboid form of D. fragiliswas frequently ob-served (Fig. 4D). Dientamoeba fragilis frequently changed its shapewhen engulfing bacteria or rice starch (Fig. 7C and E). TEM obser-vations clearly showed that phagocytosis was initiated by thebinding of bacteria or rice starch on the surface of the D. fragilis cell.These interactions triggered cytoskeletal changes that led to pseu-dopodial extensions, engulfing bacteria or rice starch. Finally,phagosomes were formed (Fig. 7D) which allowed digestion ofthe food particles. A convoluted plasmalemma (Fig. 7E, arrow)was occasionally observed, probably associated with the earlystages of phagocytosis forming a ‘phagocytic cup’ around bacteriaor rice starch.
3.6.11. Virus-like particle (VLP) in Dientamoeba fragilisVirus-like particles (VLPs) were seen frequently inside the cyto-
plasm of trophozoites by TEM during ultrastructural analysis(Fig. 9). VLP sizes varied between 40 and 200 nm. Their most com-mon shape was spherical, enclosing a dense core, a middle elec-tron-lucent layer and an outer coat.
3.7. Confocal and light microscopy observations
Dientamoeba fragilis cells and nuclei from in vitro cultured con-ditions were studied by confocal microscopy and in permanentlystained faecal smears by light microscopy. In most experiments,cells were fixed onto microscope slides. This provided a constant,known orientation of the nuclei in all cells in these experiments.
3.7.1. The two nuclei contain equal amounts of DNATo determine whether the two nuclei of D. fragilis contained
equivalent amounts of DNA, D. fragilis trophozoites were stainedwith DAPI, an agent that binds specifically and strongly to DNAindependent of its sequence. When DAPI becomes intercalatedwith nuclear DNA, a bright blue fluorescence is emitted (Fig. 11).The fluorescence emitted from each nucleus was quantitatedphotometrically by readings made through a microscopic aperture.
Fig. 8. Basal body structures of Dientamoeba fragilis. (A) multiple ribbons of microtubules forming the axostyle (arrow), which runs from the basal bodies to the cell tip; (B)closer view of attractophore (V-shaped structure) and axostyle (Ax); (C) basal body (BB) structures; (D) costa (Co) structure in the cytoplasm; (E) presence of parabasalfilament (Pf) in close proximity to the nucleus; (F) double layered D. fragilis cell membrane (m) (c, cytoplasm; ED, electron dense materials; Mt, microtubules; N, nucleus; H,hydrogenosome).
G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153 147

The nuclei of all four isolates of D. fragilis were examined. Therewas no apparent difference in the DNA content of nuclei in cellscontaining two nuclei i.e. DNA content of both nuclei is equal(Table 1).
3.7.2. Comparison of D. fragilis nuclei from in vitro culture andpermanent stained smears
Sixty percent of trophozoites in stained faecal smears werebinucleate, 30% were mononucleate and 9% were anucleate(Fig. 10). In contrast, 70% of in vitro cultured D. fragilis trophozoiteswere mononucleate, 29% were binucleate and 1% were anucleate asdetermined by confocal microscopy. However, one cell containingthree nuclei was found in stained smears (Fig. 10C) and very fewtrophozoites with three nuclei were observed in cultures (datanot shown). The cells containing three nuclei observed in culturesseemed to be a dividing form. For this reason trophozoites with
this nuclei number were not included in the data analysis. Lightand confocal microscopical observation showed that D. fragilis tro-phozoites appeared to have a wide variation in shapes and sizes.From stained smears, D. fragilis was from 4 to 16 lm in size. Thesize range of cultured D. fragilis trophozoites was from 5 to25 lm and the nuclei were from 0.5 to 3.5 lm for all four isolates.Dientamoeba fragilis usually appeared rounded but was also ovalshaped, ameboid or rod shaped and phagocytic stages were alsoseen (Fig. 13D). Dientamoeba fragilis in cultured conditions movedby using leaf-like pseudopodia which were irregularly lobed. Inaddition, typical fragmented nuclei were found in stained smears.
3.7.3. Comparison of mononucleate and binucleate D. fragilistrophozoites at different time intervals of cultures
The ratio of mononucleate and binucleate cell numbers duringdifferent growth stages in culture media was determined(Fig. 12). A total of 500 cells from isolate 1 were stained with DAPIand analysed at different time intervals (6, 12, 24, 48 and 72 h) byconfocal microscopy. Approximately 100 non-dividing DAPI-stained cells were counted at each time point. Eighty three percentof cells were found as mononucleate and 17% of the trophozoiteswere binucleate after 6 h. The number of binucleate trophozoitesincreased at 24 and 48 h time points of in vitro growth. Duringcounting, dividing cells were omitted. It appeared that D. fragilisnuclei divided non-synchronously during binary fission. At 6 and12 h trophozoites were generally smaller in size (5–10 lm).
3.7.4. Cell and nuclear volume of D. fragilisThe cell and nuclear dimensions of D. fragilis were obtained
from Nile Red-stained D. fragilis and DAPI-stained nuclei(Fig. 13A–C). Finally, the approximate volumes of D. fragilis cellsas well as nuclei were calculated (Table 2). These steps were re-peated for each of the four isolates of D. fragilis. There was no sig-nificant difference between the cell and nucleus volume betweenthe four isolates.
4. Discussion
To our knowledge, this is the first electron microscopy study in40 years to examine the ultrastructure of D. fragilis. Even thoughD. fragilis is recognised as a significant human pathogen (Starket al., 2010b; Barratt et al., 2011a), most of the microscopic obser-vations of D. fragilis date back to the early and mid-1900s (Johnsonet al., 2004) and therefore may represent inappropriate descrip-tions of this parasite. Compared with other common trichomonadssuch as Trichomonas vaginalis (Benchimol et al., 2002; Benchimol,2004a; Chen et al., 2004), Tritrichomonas foetus (Benchimol et al.,1993, 1996; Benchimol, 2000), Trichomonas gallinae (Mehlhornet al., 2009) and Histomonas meleagridis (Mielewczik et al., 2008;Munsch et al., 2009; Zaragatzki et al., 2010) very little informationis available about the biology of this organism. The present studydeals with four different D. fragilis isolates which were cultivatedin vitro. Different microscopical methods were performed to char-acterise their surface structures, ultrastructure and detailed obser-vations on nuclear characteristics of this neglected protozoan.
During recent years, several studies provided information onthe morphology of other protozoan parasites based on SEM. Trich-omonas vaginalis (Ovcinnikov et al., 1975; Kurnatowska and Hajdu-kiewicz, 1977; Warton and Honigberg, 1979; Benchimol, 2004a), T.foetus (Warton and Honigberg, 1979), Pentatrichomonas hominis(Warton and Honigberg, 1979) and Trichomitus batrachorum(Honigberg et al., 1972), Hypotrichomonas acosta (Warton andHonigberg, 1979) have all been studied in detail. Much has beenlearned about the structure of T. gallinae, first by light microscopy(Stabler, 1941, 1954; Abraham and Honigberg, 1964; Honigberg,
Fig. 9. Transmission electron micrograph showing the presence of virus-likeparticles (VLPs) in the perinuclear region (arrow) of a Dientamoeba fragilistrophozoite. VLP is budding through the Golgi complex. (N, nucleus; Gc, Golgicomplex).
Fig. 10. Iron–haematoxylin stained smears of a Dientamoeba fragilis trophozoite(clinical samples). (A) Mononucleate; (B) binucleate; (C) three nuclei; (D) anucleate.Images were taken at magnification 100X Differential Interference Contrast. Thescale bar is 10 lm. (N, nucleus).
148 G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153
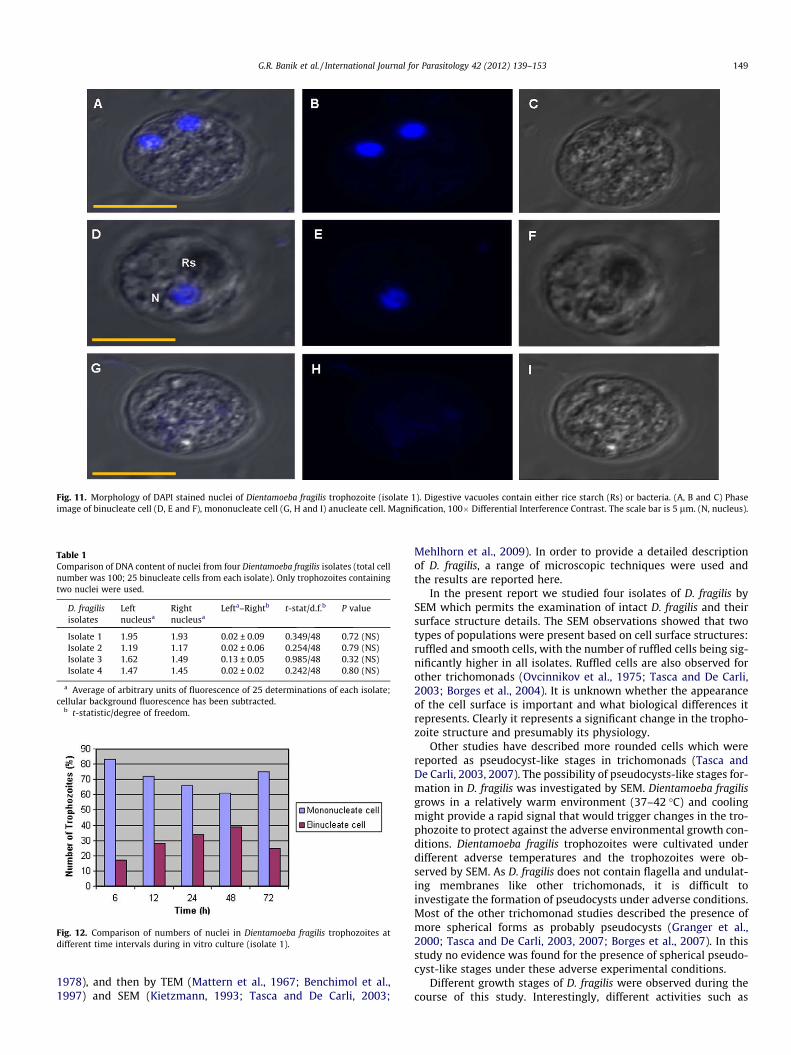
1978), and then by TEM (Mattern et al., 1967; Benchimol et al.,1997) and SEM (Kietzmann, 1993; Tasca and De Carli, 2003;
Mehlhorn et al., 2009). In order to provide a detailed descriptionof D. fragilis, a range of microscopic techniques were used andthe results are reported here.
In the present report we studied four isolates of D. fragilis bySEM which permits the examination of intact D. fragilis and theirsurface structure details. The SEM observations showed that twotypes of populations were present based on cell surface structures:ruffled and smooth cells, with the number of ruffled cells being sig-nificantly higher in all isolates. Ruffled cells are also observed forother trichomonads (Ovcinnikov et al., 1975; Tasca and De Carli,2003; Borges et al., 2004). It is unknown whether the appearanceof the cell surface is important and what biological differences itrepresents. Clearly it represents a significant change in the tropho-zoite structure and presumably its physiology.
Other studies have described more rounded cells which werereported as pseudocyst-like stages in trichomonads (Tasca andDe Carli, 2003, 2007). The possibility of pseudocysts-like stages for-mation in D. fragilis was investigated by SEM. Dientamoeba fragilisgrows in a relatively warm environment (37–42 �C) and coolingmight provide a rapid signal that would trigger changes in the tro-phozoite to protect against the adverse environmental growth con-ditions. Dientamoeba fragilis trophozoites were cultivated underdifferent adverse temperatures and the trophozoites were ob-served by SEM. As D. fragilis does not contain flagella and undulat-ing membranes like other trichomonads, it is difficult toinvestigate the formation of pseudocysts under adverse conditions.Most of the other trichomonad studies described the presence ofmore spherical forms as probably pseudocysts (Granger et al.,2000; Tasca and De Carli, 2003, 2007; Borges et al., 2007). In thisstudy no evidence was found for the presence of spherical pseudo-cyst-like stages under these adverse experimental conditions.
Different growth stages of D. fragilis were observed during thecourse of this study. Interestingly, different activities such as
Table 1Comparison of DNA content of nuclei from four Dientamoeba fragilis isolates (total cellnumber was 100; 25 binucleate cells from each isolate). Only trophozoites containingtwo nuclei were used.
D. fragilisisolates
Leftnucleusa
Rightnucleusa
Lefta–Rightb t-stat/d.f.b P value
Isolate 1 1.95 1.93 0.02 ± 0.09 0.349/48 0.72 (NS)Isolate 2 1.19 1.17 0.02 ± 0.06 0.254/48 0.79 (NS)Isolate 3 1.62 1.49 0.13 ± 0.05 0.985/48 0.32 (NS)Isolate 4 1.47 1.45 0.02 ± 0.02 0.242/48 0.80 (NS)
a Average of arbitrary units of fluorescence of 25 determinations of each isolate;cellular background fluorescence has been subtracted.
b t-statistic/degree of freedom.
Fig. 12. Comparison of numbers of nuclei in Dientamoeba fragilis trophozoites atdifferent time intervals during in vitro culture (isolate 1).
Fig. 11. Morphology of DAPI stained nuclei of Dientamoeba fragilis trophozoite (isolate 1). Digestive vacuoles contain either rice starch (Rs) or bacteria. (A, B and C) Phaseimage of binucleate cell (D, E and F), mononucleate cell (G, H and I) anucleate cell. Magnification, 100� Differential Interference Contrast. The scale bar is 5 lm. (N, nucleus).
G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153 149

amoeboid movement, phagocytosis and bacterial adhesion to tro-phozoite surfaces were also observed clearly. No flagellar struc-tures, undulating membrane or axostyle were found by SEM.Even though phagocytosis is an essential phenomenon in D. fragilis,the details of the phagocytic activity of this parasite have not yet
been elucidated. In this study, the in vitro capacity of D. fragilisto phagocytose (bacteria and rice starch) was observed. Two formsof phagocytosis were observed for D. fragilis in this study whichwere a ‘sinking’ process without any apparent participation ofplasma membrane extensions and the classical phagocytosis
Table 2Comparison of cell dimensions and approximate volumes of four isolates of Dientamoeba fragilis (total cell number was 200; 50 non-dividing cells from each isolate).
D. fragilis isolates Cell dimensions Cell volume V b
= 3/4pab2 (lm3)Nucleus dimensions Nucleus volume V b = 3/4pab2 (lm3)
Length a (lm) Width a (lm) Length a (lm) Width a (lm)Mean ± SD Mean ± SD Mean ± SD Mean ± SD
Isolate 1 11.91 ± 2.33 6.00 ± 1.25 1.0 1.8 ± 0.52 0.97 ± 0.33 0.003Isolate 2 11.78 ± 2.79 5.88 ± 1.39 0.9 2.28 ± 0.74 1.12 ± 0.32 0.006Isolate 3 12.07 ± 2.93 6.01 ± 1.45 1.0 1.50 ± 0.38 0.73 ± 0.22 0.001Isolate 4 12.69 ± 3.99 7.40 ± 3.03 1.6 2.09 ± 0.61 1.11 ± 0.49 0.006
a Average of 50 trophozoites ± S.D.b The cell volume was calculated according to the formula V = 3/4pab2; ‘‘a’’ is the cell length; ‘‘b’’ is the cell width. (p = 3.14).
Table 3Ultrastructural characteristics of the organelles of Dientamoeba fragilis.
Name Characteristics Sizes
Nucleus One, two or three; mostly spherical; located in the central region of cell; double membrane containing numerousnuclear pores; usually containing two to eight chromatin bodies without peripheral chromatin
0.86–3.52 lm
Golgi complex Vesicular structure generally located in the perinuclear area; 7–10 cisternae About 450 nm longEndoplasmic reticulum Smooth and rough endoplasmic reticulums were observed; found around the nucleus About 500 nm longHydrogenosomes Spherical or oval shaped; double layered membrane; located in the cytoplasm; 5–15 per cell 0.12–0.83 lmDigestive vacuoles Found throughout the cytoplasm; 1–10 per cell 0.59–4.2 lmLysosomes Located mainly in the posterior region of the cell in close proximity to the cell membrane 0.50–2 lmBasal body componentsAxostyle Microtubular ribbon; usually found in the anterior region of the cellCosta Proteinaceous structureParabasal filament Located near to nucleus; singular or in a cluster form; formed by 30–40 hairy-like segments
Fig. 13. Nile Red stained Dientamoeba fragilis trophozoite and DAPI stained nuclei showing the cell volume. (A) Nile Red stained mononucleate; (B) binucleate; (C) anucleate;(D) D. fragilis exhibiting phagocytosis. Magnification, 100� Differential Interference Contrast. The scale bar is 3 lm. (Rs, rice starch; N, nucleus).
150 G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153

where pseudopodia were extended by amoeboid movement. Thephagocytic process may occur during the division of parasite.
By electron microscopy, D. fragilis usually has a spherical or ovalshape, although some are amoeboid with a finely granular cyto-plasm. Trophozoites with one, two or three nuclei were quite com-mon. The nuclear structure of D. fragilis more closely resemblesthat of trichomonads (Benchimol, 2004a) rather than that of Ent-amoeba spp. (Ludvík and Shipstone, 1970; Jhingan et al., 2009) orHistomonas spp. (Zaragatzki et al., 2010). The cell division occurredby binary fission i.e. simple constriction of the cell body (Fig. 3Cand D) as described previously (Johnson et al., 2004). Nuclear divi-sion was found only in mononucleated trophozoites as previouslyreported (Wenrich, 1939; Dobell, 1940). In some cells chromatinbodies were not prominent; perhaps such nuclei are approachingthe interphase stages. Previous studies have reported the presenceof microtubules in the nucleus which apparently penetratethrough the nuclear membrane into the nucleoplasm forming atransnuclear spindle (Camp et al., 1974; Silard et al., 1984). Thesewere not observed in this present study.
A prominent Golgi complex was observed in mononucleateorganisms in all isolates. In the binucleate cell the Golgi complexwas mostly fragmented as reported earlier (Silard et al., 1984). Itis believed that the Golgi complex in trichomonads may be in-volved in the processing of adhesion proteins that mediate theinteraction of cells with host cells (Alderete and Garza, 1988;Arroyo et al., 1992; Shaia et al., 1998). Both smooth and rough ERswere also clearly seen around the nucleus and sometimes closelyassociated with the hydrogenosomes, DVs and microtubules.
Hydrogenosomes were observed in all four isolates and thenumber varied between 5 and 15 per trophozoite. These organelleswere previously termed ‘‘microbody-like’’ inclusions for D. fragilis(Johnson et al., 2004) and were presumed to be homologous tothe paraxostylar granules of trichomonads (Camp et al., 1974).These inclusions were subsequently recognised as being hydro-genosomes (Müller, 1975). In trichomonads they are found closeto the axostyle and costa (Benchimol and Souza, 1983; Benchimol,2000). The matrix of the hydrogenosome is homogeneously granu-lar, similar to other trichomonads (Benchimol, 2004a).
Hydrogenosomes are an unusual organelle found in severaltrichomonad species and other protists living in oxygen poor oranoxic environments (Benchimol et al., 1993; Benchimol andEngelke, 2003). They do not have mitochondria nor peroxisomes(Benchimol, 2004a; Carlton et al., 2007). The hydrogenosomecontains enzymes that probably participate in the metabolismof pyruvate and is the site of formation of ATP and molecularhydrogen (Benchimol, 2009; Shiflett and Johnson, 2010; Stairset al., 2011). The function of hydrogenosomes in D. fragilis are likelyto be similar and related to energy production.
Basal body structures, namely the axostyle, costa and PFs wereobserved in D. fragilis by TEM. Previous phylogenetic analyses ofparabasalids observed that both the mastigont and the pelta-axo-styler complex were completely lost in D. fragilis and its inclusionin the Parabasalia was based on the presence of hydrogenosomes(Silberman et al., 1996; Gerbod et al., 2001; Cepicka et al., 2010).The axostyle is usually seen along the longitudinal axis of the cellin trichomonads by TEM (Benchimol, 2004a). The appearance ofaxostyle tips as ‘‘sticking out’’ from the posterior end of the cellwas also observed in trichomonads by SEM (Borges et al., 2004)but such arrangements for axostyle were not observed in D. fragilis.
No flagella was observed by SEM and TEM showing that D.fragilis has permanently lost its flagella. Similar to trichomonads(Benchimol, 2004a) and Histomonas (Schuster, 1968; Honigbergand Bennett, 1971) the parabasal apparatus of D. fragilis consistsof periodic filaments that often appeared V-shaped in section(Camp et al., 1974). Clusters of the parabasal apparatus were alsoobserved in some cells in this study. Camp et al. (1974) reported
that this PF and the overlying Golgi complex corresponded to thetrichomonad parabasal apparatus. It was described that the polarcomplex is mainly a paired non-periodic element found in a divid-ing organism (Camp et al., 1974). It contains microtubule struc-tures with an affiliation with spindle microtubules. Similarobservations were made in this study.
The presence of VLPs is very common in other parasitic protozoabut there was no previous evidence for their existance in D. fragilis.Most VLPs of protozoa are double-stranded RNA (dsRNA) virusesranging from 30 to 200 nm in diameter and the size of their genomeis 5–7 kb (Wang andWang, 1991a,b; Benchimol et al., 2002). dsRNAviruses are also found in several isolates of T. vaginalis (Wang andWang, 1985,1986a; Benchimol, 2004a), Giardia (Wang and Wang,1986b), Leishmania (Tarr et al., 1988) and Eimeria (Revets et al.,1989; Ellis and Revets, 1990). Kasprzak andMajewska (1995) statedthat all of the RNA viruses observed in parasitic protozoa showedseveral similarities and did not considerably differ from the virusesfound in simple eukaryotic cells; they closely correspond to thedsRNA viruses of yeast (Kasprzak and Majewska, 1995). It is re-ported that the presence of VLPs within T. vaginalis is associatedwith expression of immunogenic proteins on the trichomonad sur-face, variations in protozoal phenotypes, and upregulation of cer-tain proteins, including known virulence factors and with diseasepathogenesis (Gerhold et al., 2009; Fraga et al., 2011; Malla et al.,2011). The identification of a VLP in D. fragilis in this study, forthe first time, extends the distribution of VLPs to another inhabitantof the human gut.
Almost all previous studies dealing with microscopic observa-tions of D. fragilis described that the characteristics of D. fragilisvaried greatly according to the samples taken whether they befrom cultures or faecal smears (Craig, 1926; Wenrich, 1939,1944a; Dobell, 1940; Johnson et al., 2004). The present study in-cluded a comparative analysis of nucleus numbers by both lightand confocal microscopy of faecal smears and cultures. The confo-cal microscopy showed that the cultivated trophozoite (reachingapproximately 25 lm in size) appeared more or less spherical orshowed amoeba-like morphology. Much larger sizes of trophozoitewere found in cultured trophozoites compared with clinical sam-ples. This finding supports previous observations (Silard et al.,1979). The diameter of the nuclei of D. fragilis ranged from 0.5 to3.5 lm in cultured trophozoites from all four isolates which is sim-ilar to that reported previously (Johnson et al., 2004). In perma-nently stained smears, 60% of trophozoites were binucleatewhereas 70% of in vitro cultured trophozoites were mononucleate.This observation is also consistent with previous studies (Jepps andDobell, 1918; Wenrich, 1944b; Windsor et al., 2003; Johnson et al.,2004; Stark et al., 2006). Dobell (1940) reported that the morphol-ogy of D. fragilis as seen in stools discharged from the human bodydiffers considerably from that observable in flourishing cultures.This is partly because most specimens in faeces are not viableand therefore do not present a true picture of the structure of thisorganism (Dobell, 1940).
The cytoplasm of D. fragilis appeared granular, vacuolated anddensely filled with mainly food vacuoles containing either ricestarch or bacteria and this has been observed before (Dobell,1940; Johnson et al., 2004). In both stained smears and in vitro cul-tures, one of the granules (typically with starch) was larger thanthe others. Similarly, numerous food vacuoles are present in H.meleagridis (Munsch et al., 2009). In the nucleus, the chromatinmaterial accumulates around the periphery and the nucleolus iscentrally situated. The nucleus divides non-synchronously withina cell and cell division is not always associated with synchronousreplication of the DNA within these nuclei. Cell division also occursnon-synchronously in G. lamblia (Kabnick and Peattie, 1990). A pre-vious study showed that in stained smears, using iron–haematox-ylin, the nuclear membrane is delicate and does not possess any
G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153 151

peripheral chromatin (Stark et al., 2006). It is noteworthy that thestructure of D. fragilis also varies greatly, as observed in stainedpreparations, with the degree of differentiation of the stain, andunless the preparations are well differentiated, the typical appear-ance of the karyosome is entirely lost.
An unusual feature of D. fragilis is the presence of two nuclei inthe same trophozoite. Dientamoeba fragilis is not alone in this re-spect, since others like Giardia also possess two nuclei. The two nu-clei of D. fragilis trophozoites were shown to be equivalent withrespect to the amount of DNA contained in each. The approximatecell volume and nuclear volume of four isolates of D. fragilis werealso obtained and the findings are comparable to Trichomonasspp. (Zubácová et al., 2008). The changes observed during cell cul-ture of the number of nuclei present in trophozoites indicate astage containing two nuclei is probably an intermediary of celldivision.
To conclude, despite its widespread occurrence and associatedsymptoms, remarkably little is known about the biology and path-ogenicity of D. fragilis. For the first time, the characteristics andultrastructure of D. fragilis are described here in detail. 3D surfacestructures observed by SEM and ultrastructural detailed observa-tions by TEM showed that there were no significant differencesbetween these four isolates except for possibly the presence ofVLPs which require further investigation. This study was only pos-sible because of the recent major advances made in the culture ofD. fragilis (Barratt et al., 2010).
Acknowledgements
This study was supported by funds from University of Technol-ogy, Sydney, Australia and the Australian Research Council. Allelectron microscopy was conducted at the Macquarie University,Australia, microscopy unit. The authors thank Ms. Nicole Vella fortechnical assistance. We also wish to express our appreciation toDr. Michael Johnson for his support with confocal microscopyand Mr. Andrew Liew for providing access to the image software.
References
Abraham, R., Honigberg, B.M., 1964. Structure of Trichomonas gallinae (Rivolta). J.Parasitol. 50, 608–619.
Adam, R.D., 2001. Biology of Giardia lamblia. Clin. Microbiol. Rev. 14, 447–475.Alderete, J.F., Garza, G.E., 1988. Identification and properties of Trichomonas
vaginalis proteins involved in cytadherence. Infect. Immun. 56, 28–33.Arroyo, R., Engbring, J., Alderete, J.F., 1992. Molecular basis of host epithelial cell
recognition by Trichomonas vaginalis. Mol. Microbiol. 6, 853–862.Barratt, J.L., Banik, G.R., Harkness, J., Marriott, D., Ellis, J.T., Stark, D., 2010. Newly
defined conditions for the in vitro cultivation and cryopreservation ofDientamoeba fragilis: new techniques set to fast track molecular studies onthis organism. Parasitology 137, 1867–1878.
Barratt, J.L.N., Harkness, J., Marriott, D., Ellis, J.T., Stark, D., 2011a. A review ofDientamoeba fragilis carriage in humans: several reasons why this organismshould be considered in the diagnosis of gastrointestinal illness. Gut Microb. 2,3–12.
Barratt, J.L., Harkness, J., Marriott, D., Ellis, J.T., Stark, D., 2011b. The ambiguous lifeof Dientamoeba fragilis: the need to investigate current hypotheses ontransmission. Parasitology 138, 557–572.
Benchimol, M., D Souza, W., 1983. Fine structure and cytochemistry of thehydrogenosome of Tritrichomonas foetus. J. Protozool. 30, 422–425.
Benchimol, M., Kachar, B., de Souza, W., 1993. The structural organization of thepathogenic protozoan Tritrichomonas foetus as seen in replicas of quick frozen,freeze-fractured and deep etched cells. Biol. Cell. 77, 289–295.
Benchimol, M., Almeida, J.C., de Souza, W., 1996. Further studies on the organizationof the hydrogenosome in Tritrichomonas foetus. Tissue Cell. 28, 287–299.
Benchimol, M., Leal, D., Mattos, A., Diniz, J.A.P., 1997. Fine structure of Trichomonasgallinae. Biocell. 21, 47–58.
Benchimol, M., 2000. Ultrastructural characterization of the isolatedhydrogenosome in Tritrichomonas foetus. Tissue Cell. 32, 518–526.
Benchimol, M., Monteiro, S., Chang, T.H., Alderete, J.F., 2002. Virus in Trichomonas –an ultrastructural study. Parasitol. Int. 51, 293–298.
Benchimol, M., Engelke, F., 2003. Hydrogenosome behavior during the cell cycle inTritrichomonas foetus. Biol Cell. 95, 283–293.
Benchimol, M., 2004a. Trichomonads under microscopy. Microsc. Microanal. 10,528–550.
Benchimol, M., 2004b. Mitosis in Giardia lamblia: multiple modes of cytokinesis.Protist. 155, 33–44.
Benchimol, M., 2009. Hydrogenosomes under microscopy. Tissue Cell. 41, 151–168.Borges, F.P., Gottardi, B., Stuepp, C., Larré, A.B., de Brum Vieira, P., Tasca, T., De Carli,
G.A., 2007. Morphological aspects of Monocercomonas sp. and investigation onprobable pseudocysts occurrence. Parasitol. Res. 101, 1503–1509.
Borges, F.P., Wiltuschnig, R.C., Tasca, T., De Carli, G.A., 2004. Scanning electronmicroscopy study of Tritrichomonas augusta. Parasitol. Res. 94, 158–161.
Camp, R.R., Mattern, C.F., Honigberg, B.M., 1974. Study of Dientamoeba fragilis Jepps& Dobell. I. Electronmicroscopic observations of the binucleate stages. II.Taxonomic position and revision of the genus. J. Protozool. 21, 69–82.
Carlton, J.M., Hirt, R.P., Silva, J.C., Delcher, A.L., Schatz, M., Zhao, Q., Wortman, J.R.,Bidwell, S.L., Alsmark, U.C.M., Besteiro, S.b., Sicheritz-Ponten, T., Noel, C.J.,Dacks, J.B., Foster, P.G., Simillion, C., Van de Peer, Y., Miranda-Saavedra, D.,Barton, G.J., Westrop, G.D., Müller, S., Dessi, D., Fiori, P.L., Ren, Q., Paulsen, I.,Zhang, H., Bastida-Corcuera, F.D., Simoes-Barbosa, A., Brown, M.T., Hayes, R.D.,Mukherjee, M., Okumura, C.Y., Schneider, R., Smith, A.J., Vanacova, S., Villalvazo,M., Haas, B.J., Pertea, M., Feldblyum, T.V., Utterback, T.R., Shu, C.-L., Osoegawa,K., de Jong, P.J., Hrdy, I., Horvathova, L., Zubacova, Z., Dolezal, P., Malik, S.-B.,Logsdon, J.M., Henze, K., Gupta, A., Wang, C.C., Dunne, R.L., Upcroft, J.A., Upcroft,P., White, O., Salzberg, S.L., Tang, P., Chiu, C.-H., Lee, Y.-S., Embley, T.M., Coombs,G.H., Mottram, J.C., Tachezy, J., Fraser-Liggett, C.M., Johnson, P.J., 2007. Draftgenome sequence of the sexually transmitted pathogen Trichomonas vaginalis.Science 315, 207–212.
Cepicka, I., Hampl, V., Kulda, J., 2010. Critical taxonomic revision of Parabasalidswith description of one new genus and three new species. Protist. 161, 400–433.
Cerva, L., Nohynková, E., 1992. A light microscopic study of the course of cellulardivision of Giardia intestinalis trophozoites grown in vitro. Folia. Parasitol. 39,97–104.
Chen, W., Chen, J., Zhong, X., Lin, X., Chen, L., 2004. Electron microscopic study onTrichomonas vaginalis adhering to and phagocytizing male genitourinaryepithelial cells. Zhonghua Nan Ke Xue 10, 86–89.
Craig, C.F., 1926. The nuclear structure of Dientamoeba fragilis. J. Parasitol. 13, 137–140.
Delgado-Viscogliosi, P., Viscogliosi, E., Gerbod, D., Kulda, J., Sogin, M.L., Edgcomb,V.P., 2000. Molecular phylogeny of parabasalids based on small subunit rRNAsequences, with emphasis on the Trichomonadinae subfamily. J. Eukaryot.Microbiol. 47, 70–75.
Dobell, C., 1940. Researches on the intestinal protozoa of monkeys and man. X. Thelife history of Dientamoeba fragilis: observations, experiments and speculations.Parasitology 32, 417–461.
Ellis, J., Revets, H., 1990. Eimeria species which infect the chicken contain virus-likeRNA molecules. Parasitology 101, 163–169.
Fraga, J., Rojas, L., Sariego, I., Fernández-Calienes, A., 2011. Double-stranded RNAviral infection of Trichomonas vaginalis and correlation with geneticpolymorphism of isolates. Exp. Parasitol. 127, 593–599.
Gerbod, D., Edgcomb, V.P., Noël, C., Zenner, L., Wintjens, R., Delgado-Viscogliosi, P.,Holder, M.E., Sogin, M.L., Viscogliosi, E., 2001. Phylogenetic position of thetrichomonad parasite of turkeys, Histomonas meleagridis (Smith) Tyzzer,inferred from small subunit rRNA sequence. J. Eukaryot. Microbiol. 48,498–504.
Gerbod, D., Noël, C., Dolan, M.F., Edgcomb, V.P., Kitade, O., Noda, S., Dufernez, F.,Ohkuma, M., Kudo, T., Capron, M., Sogin, M.L., Viscogliosi, E., 2002. Molecularphylogeny of parabasalids inferred from small subunit rRNA sequences, withemphasis on the Devescovinidae and Calonymphidae (Trichomonadea). Mol.Phylogenet. Evol. 25, 545–556.
Gerhold, R.W., Allison, A.B., Sellers, H., Linnemann, E., Chang, T.H., Alderete, J.F.,2009. Examination for double-stranded RNA viruses in Trichomonas gallinae andidentification of a novel sequence of a Trichomonas vaginalis virus. Parasitol. Res.105, 775–779.
Granger, B.L., Warwood, S.J., Benchimol, M., De Souza, W., 2000. Transientinvagination of flagella by Tritrichomonas foetus. Parasitol. Res. 86, 699–709.
Honigberg, B.M., Bennett, C.J., 1971. Lightmicroscopic observations on structure anddivision of Histomonas meleagridis (Smith). J. Protozool. 18, 687–697.
Honigberg, B.M., Daniel, W.A., Mattern, C.F., 1972. Fine Structure of Trichomitusbatrachorum (Perty). J. Protozool. 19, 446–453.
Honigberg, B.M., 1978. Trichomonads of veterinary importance. In: Kreier, J.P. (Ed.),Parasitic Protozoa. Academic Press, New York, pp. 163–273.
Jahn, C.L., Klobutcher, L.A., 2002. Genome remodeling in ciliated protozoa. Annu.Rev. Microbiol. 56, 489–520.
Jepps, M.W., Dobell, C., 1918. Dientamoeba fragilis n.g., n. sp., new intestinal amoebafrom man. Parasitology 10, 352–367.
Jhingan, G.D., Panigrahi, S.K., Bhattacharya, A., Bhattacharya, S., 2009. The nucleolusin Entamoeba histolytica and Entamoeba invadens is located at the nuclearperiphery. Mol. Biochem. Parasitol. 167, 72–80.
Johnson, E.H., Windsor, J.J., Clark, C.G., 2004. Emerging from obscurity: biological,clinical, and diagnostic aspects of Dientamoeba fragilis. Clin. Microbiol. Rev. 17,553–570.
Kabnick, K.S., Peattie, D.A., 1990. In situ analyses reveal that the two nuclei ofGiardia lamblia are equivalent. J. Cell. Sci. 95, 353–360.
Kasprzak, W., Majewska, A.C., 1995. Viruses of parasitic protozoa. Wiad Parazytol.41, 131–137.
Kietzmann Jr., G.E., 1993. Relationships of Trichomonas gallinae to the palatal-esophageal junction of ring doves (Streptopelia risoria) as revealed by scanningelectron microscopy. J. Parasitol. 79, 408–415.
152 G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153

Kurnatowska, A., Hajdukiewicz, G., 1977. Trichomonas vaginalis variability in axeniccultures evaluated in scanning electron microscope. Wiad. Parazytol 23, 481–487.
Ludvík, J., Shipstone, A.C., 1970. The ultrastructure of Entamoeba histolytica. Bull.World Health Organ. 43, 301–308.
Malla, N., Kaul, P., Sehgal, R., Gupta, I., 2011. The presence of dsRNA virus inTrichomonas vaginalis isolates from symptomatic and asymptomatic Indianwomen and its correlation with in vitro metronidazole sensitivity. Indian J.Med. Microbiol. 29, 152–157.
Mattern, C.F., Honigberg, B.M., Daniel, W.A., 1967. The mastigont system ofTrichomonas gallinae (Rivolta) as revealed by electron microscopy. J. Protozool.14, 320–339.
Mehlhorn, H., Al-Quraishy, S., Aziza, A., Hess, M., 2009. Fine structure of the birdparasites Trichomonas gallinae and Tetratrichomonas gallinarum from cultures.Parasitol. Res. 105, 751–756.
Mielewczik, M., Mehlhorn, H., Al-Quraishy, S., Grabensteiner, E., Hess, M., 2008.Transmission electron microscopic studies of stages of Histomonas meleagridisfrom clonal cultures. Parasitol. Res. 103, 745–750.
Moody, A.H., Fleck, S.L., 1985. Versatile Field’s stain. J. Clin. Pathol. 38, 842–843.Müller, M., 1975. Biochemistry of protozoan microbodies: peroxisomes, alpha-
glycerophosphate oxidase bodies, hydrogenosomes. Annu. Rev. Microbiol. 29,467–483.
Munsch, M., Lotfi, A., Hafez, H.M., Al-Quraishy, S., Mehlhorn, H., 2009. Light andtransmission electron microscopic studies on trophozoites and cyst-like stagesof Histomonas meleagridis from cultures. Parasitol. Res. 104, 683–689.
Ockert, G., Schneider, W., 1974. Electron microscopic findings in Dientamoebafragilis. Z. Gesamte Hyg. 20, 555–557.
Ohkuma, M., Iida, T., Ohtoko, K., Yuzawa, H., Noda, S., Viscogliosi, E., Kudo, T., 2005.Molecular phylogeny of parabasalids inferred from small subunit rRNAsequences, with emphasis on the Hypermastigea. Mol. Phylogenet. Evol. 35,646–655.
Ovcinnikov, N.M., Delektorskij, V.V., Turanova, E.N., Yashkova, G.N., 1975. Furtherstudies of Trichomonas vaginalis with transmission and scanning electronmicroscopy. Br. J. Vener Dis. 51, 357–375.
Prescott, D.M., 1994. The DNA of ciliated protozoa. Microbiol. Rev. 58, 233–267.Revets, H., Dekegel, D., Deleersnijder, W., De Jonckheere, J., Peeters, J., Leysen, E.,
Hamers, R., 1989. Identification of virus-like particles in Eimeria stiedae. Mol.Biochem. Parasitol. 36, 209–215.
Sawangjaroen, N., Luke, R., Prociv, P., 1993. Diagnosis by faecal culture ofDientamoeba fragilis infections in Australian patients with diarrhoea. Trans. RSoc. Trop. Med. Hyg. 87, 163–165.
Schuster, F.L., 1968. Ultrastructure of Histomonas meleagridis (Smith) Tyzzer, aParasitic Amebo-Flagellate. J. Parasitol. 54, 725–737.
Shaia, C.I., Voyich, J., Gillis, S.J., Singh, B.N., Burgess, D.E., 1998. Purification andexpression of the Tf190 adhesin in Tritrichomonas foetus. Infect. Immun. 66,1100–1105.
Shiflett, A.M., Johnson, P.J., 2010. Mitochondrion-related organelles in eukaryoticprotists. Annu. Rev. Microbiol. 64, 409–429.
Silard, R., Colea, A., Panaitescu, D., Florescu, P., Roman, N., 1979. Studies onDientamoeba fragilis in Romania. I. Dientamoeba fragilis isolated from clinicalcases. Problems of diagnosis, incidence, clinical aspects. Arch. Roum. Pathol.Exp. Microbiol. 38, 359–372.
Silard, R., Burghelea, B., Panaitescu, D., Burcos, V., 1984. Ultrastructure ofDientamoeba fragilis: a study of the mononucleated stage. Arch. Roum. Pathol.Exp. Microbiol. 43, 87–101.
Silard, R., Burghelea, B., 1986. Endosymbionts in Dientamoeba fragilistrophozoites resistant to antiprotozoal drugs. Arch. Roum. Pathol. Exp.Microbiol. 45, 65–74.
Silberman, J.D., Clark, C.G., Sogin, M.L., 1996. Dientamoeba fragilis shares a recentcommon evolutionary history with the trichomonads. Mol. Biochem. Parasitol.76, 311–314.
Southern, E.M., 1975. Detection of specific sequences among DNA fragmentsseparated by gel electrophoresis. J. Mol. Biol. 98, 503–517.
Stabler, R.M., 1941. The morphology of Trichomonas gallinae (= columbae). J.Morphol. 69, 501–515.
Stabler, R.M., 1954. Trichomonas gallinae: a review. Exp. Parasitol. 3, 368–402.Stairs, C.W., Roger, A.J., Hampl, V., 2011. Eukaryotic pyruvate formate lyase and its
activating enzyme were acquired laterally from a firmicute. Mol. Biol. Evol. 28,2087–2099.
Stark, D., Phillips, O., Peckett, D., Munro, U., Marriott, D., Harkness, J., Ellis, J., 2008.Gorillas are a host for Dientamoeba fragilis: an update on the life cycle and hostdistribution. Vet. Parasitol. 151, 21–26.
Stark, D., Barratt, J., Roberts, T., Marriot, D., Harkness, J., Ellis, J., 2010a. Comparisonof microscopy, two xenic culture techniques, conventional and real-time PCRfor the detection of Dientamoeba fragilis in clinical stool samples. Eur. J. Clin.Microbiol. Infect. Dis. 29, 411–416.
Stark, D., Barratt, J., Roberts, T., Marriott, D., Harkness, J., Ellis, J., 2010b. A review ofthe clinical presentation of dientamoebiasis. Am. J. Trop. Med. Hyg. 82, 614–619.
Stark, D.J., Beebe, N., Marriott, D., Ellis, J.T., Harkness, J., 2006. Dientamoebiasis:clinical importance and recent advances. Trends Parasitol. 22, 92–96.
Svärd, S.G., Hagblom, P., Palm, J.E., 2003. Giardia lamblia – a model organism foreukaryotic cell differentiation. FEMS Microbiol. Lett. 218, 3–7.
Tarr, P.I., Aline Jr., R.F., Smiley, B.L., Scholler, J., Keithly, J., Stuart, K., 1988. LR1: acandidate RNA virus of Leishmania. Proc. Natl. Acad. Sci. U S A 85, 9572–9575.
Tasca, T., De Carli, G.A., 2003. Scanning electron microscopy study of Trichomonasgallinae. Vet. Parasitol. 118, 37–42.
Tasca, T., De Carli, G.A., 2007. Morphological study of Tetratrichomonas didelphidisisolated from opossum Lutreolina crassicaudata by scanning electronmicroscopy. Parasitol. Res. 100, 1385–1388.
Wang, A.L., Wang, C.C., 1986a. The double-stranded RNA in Trichomonas vaginalismay originate from virus-like particles. Proc. Natl. Acad. Sci. U S A 83, 7956–7960.
Wang, A.L., Wang, C.C., 1986b. Discovery of a specific double-stranded RNA virus inGiardia lamblia. Mol. Biochem. Parasitol. 21, 269–276.
Wang, A.L., Wang, C.C., 1991a. Viruses of the protozoa. Annu. Rev. Microbiol. 45,251–263.
Wang, A.L., Wang, C.C., 1991b. Viruses of parasitic protozoa. Parasitol. Today 7, 76–80.
Wang, A.L., Wang, C.C., 1985. A linear double-stranded RNA in Trichomonasvaginalis. J. Biol. Chem. 260, 3697–3702.
Warton, A., Honigberg, B.M., 1979. Structure of trichomonads as revealed byscanning electron microscopy. J. Protozool. 26, 56–62.
Wenrich, D.H., 1939. Studies on Dientamoeba fragilis (protozoa). III. Binary fissionwith special reference to nuclear division. J. Parasitol. 25, 43–45.
Wenrich, D.H., 1944a. Nuclear structure and nuclear division in Dientamoeba fragilis(Protozoa). J. Morphol. 74, 467–491.
Wenrich, D.H., 1944b. Studies on Dientamoeba fragilis (protozoa). IV. Furtherobservations, with an outline of present-day knowledge of this species. J.Parasitol. 30, 322–337.
Wiesehahn, G.P., Jarroll, E.L., Lindmark, D.G., Meyer, E.A., Hallick, L.M., 1984. Giardialamblia: autoradiographic analysis of nuclear replication. Exp. Parasitol. 58, 94–100.
Windsor, J.J., Johnson, E.H., 1999. Dientamoeba fragilis: the unflagellated humanflagellate. Br. J. Biomed. Sci. 56, 293–306.
Windsor, J.J., Macfarlane, L., Hughes-Thapa, G., Jones, S.K., Whiteside, T.M., 2003.Detection of Dientamoeba fragilis by culture. Br. J. Biomed. Sci. 60, 79–83.
Zaragatzki, E., Hess, M., Grabensteiner, E., Abdel-Ghaffar, F., Al-Rasheid, K.A.,Mehlhorn, H., 2010. Light and transmission electron microscopic studies on theencystation of Histomonas meleagridis. Parasitol. Res. 106, 977–983.
Zubácová, Z., Cimburek, Z., Tachezy, J., 2008. Comparative analysis of trichomonadgenome sizes and karyotypes. Mol. Biochem. Parasitol. 161, 49–54.
G.R. Banik et al. / International Journal for Parasitology 42 (2012) 139–153 153

39
Chapter 3:
Electron microscopy characterisation of Dientamoeba fragilis
virus-like particles
(Submitted for publication in the Journal of Parasitology,
November 2013)

40
Virus-Like Particles in Dientamoeba fragilis: an ultrastructural study
(Running title: Characterisation of D. fragilis virus life cycle)
Gouri R. Banik a, b, Debra Birch c, Damien Stark a,d , John T. Ellis a,b,
aUniversity of Technology Sydney, School of Medical and Molecular Biosciences,
Broadway, Australia
bUniversity of Technology Sydney, i3 Institute, Broadway, Australia
cFaculty of Science, Macquarie University, Sydney, New South Wales, Australia
dDivision of Microbiology, SydPath, St. Vincent's Hospital, Darlinghurst, Australia
# Corresponding author: Professor J.T. Ellis, Ph: 61-2-9514-4161; Fax: 61-2-9514-
8206; Email: [email protected]

41
ABSTRACT
Dientamoeba fragilis is a pathogenic trichomonad parasite found in the gastrointestinal
tract of humans and is implicated as a cause of diarrhoeal disease. Previous studies led
to the discovery of virus-like particles (VLPs) in D. fragilis. The objective of this study
was to describe, by transmission electron microscopy, the presence and morphological
details of the virus population found in different clinical isolates of D. fragilis growing
in xenic culture. These heterogeneous virus populations comprise different sizes ranging
from 33-40 nm and most common shape was spherical. These VLPs contain an inner
dark core surrounded by an electron-lucent layer and an electron-dense capsid coat.
Dientamoeba fragilis VLPs are mainly found outside the nucleus in the perinuclear
region of the trophozoite. In addition, virus particles were observed to be associated
with the Golgibody in the perinuclear region while other VLPs were found in the
vicinity of the parasite plasma membrane. These VLPs attach and penetrate into D.
fragilis by endocytosis. Internally, VLPs are commonly seen in lysosomes which may
be where viral uncoating occurs. Virus-like particles were abundant in the growth media
of stationary-phase cultures of D. fragilis. Virus-like particles were found in growth
media of four isolates of D. fragilis. Dientamoeba fragilis VLPs are detectable in dying
trophozoites present in in vitro cultures. Whether viral load contributes to cell death is
unknown. The identity of the viral genome was also studied. Unlike Trichomonas
vaginalis, D. fragilis trophozoites do not contain an abundant viral dsRNA. This is the
first study to describe in detail the ultrastructural characteristics of a Dientamoeba
fragilis virus (DFV) and its mode of maturation in different cultured isolates of D.
fragilis.
Key words: Ultrastructure; Virus- like particles; Trichomonad; dsRNA.

42
3.1 Introduction
Dientamoeba fragilis is a trichomonad parasite of the human gastrointestinal
track with the propensity to cause chronic gastrointestinal disease (Barratt et al. 2011a).
Numerous studies now report that D. fragilis has a worldwide distribution with a
prevalence ranging from 5.2% to 52% (Stark et al. 2010b; Banik et al. 2011).
Dientamoeba fragilis has a limited host range with Homo sapiens being the most
common host although gorillas, pigs and sheep may also be potential hosts (Barratt et
al. 2011b; Cacciò et al. 2012). The complete life cycle of this parasite has not yet been
fully determined and the trophozoite is the only well described stage (Barratt et al.
2011b). The transmission of D. fragilis is still an enigma although many studies have
provided different opinions since its discovery (Barratt et al. 2011b; Röser et al. 2013).
Morphologically, D. fragilis is single-celled pleomorphic trophozoite typically ranging
from 5 to 15 μm in diameter (Stark et al. 2006). One of the unusual features of D.
fragilis is the presence of two nuclei (Banik et al. 2012). It is not possible to see nuclear
structure in an unstained preparation (Johnson et al. 2004).
The presence of virus-like particles (VLPs) are very common in parasitic
protozoa, especially those that inhabit the gut (Wang & Wang 1991; Goodman et al.
2011a) but there was no previous report of their existence in D. fragilis. Recently the
ultrastructure of D. fragilis was described by electron microscopy, including the
discovery of VLPs in cultured trophozoites (Banik et al. 2012). Almost all previous
reports of VLPs in protozoa derive from electron micrographs of thin sections (Wang &
Wang 1991; Goodman et al. 2011a). The VLPs of protozoa are either RNA or dsRNA
viruses ranging in size from 30-200 nm in diameter (Wang & Wang 1991; Benchimol et
al. 2002). These viruses all have genomes between 5-7 kb (Wang & Wang 1991). For
example, protozoan viruses can be found in Trichomonas (Benchimol 2004; Benchimol
et al. 2002b), Giardia (Wang & Wang 1986a), Eimeria (del Cacho et al. 2001),
Entamoeba (Mattern et al. 1974) and Leishmania (Tarr et al. 1988). It was also reported
that all of the RNA viruses detected in parasitic protozoa have similar characteristics
and were closely related to dsRNA viruses of yeast (Kasprzak & Majewska 1995).
Genome sequencing and phylogenetic analyses of several virus genomes from T.

43
vaginalis has demonstrated that they have homology to nonsegmented dsRNA viruses
of the family Totiviridae (Bessarab et al. 2011; Goodman et al. 2011b).
Several studies reported that the presence of dsRNA viruses within T. vaginalis is
associated with expression of immunogenic proteins (such as p270) on the trichomonad
surface, variations in protozoal phenotypes, upregulation of certain proteins including
altered cysteine proteinase expression profiles known as parasite virulence factors and
disease pathogenesis (Goodman et al. 2011a, 2011b; Malla et al. 2011). Goodman et al.
(2011a) postulated that TVV modulation of T. vaginalis proteinase expression increased
its survival in its host.
The main objective of this study was to investigate the presence of VLPs in
different isolates (isolates 1- 4) of D. fragilis and characterise their development in
trophozoites. Transmission electron microscopy of thin sections made from trophozoites
and negative staining of VLPs in growth media was performed to determine biological
characterisitics. The virus maturation pathway in D. fragilis trophozoite was also
described in detail. The identity of these VLP genomes was investigated by the
application of methods to purify dsRNA. No abundant dsRNA was detected suggesting
the VLPs may not be a dsRNA virus. In contrast, acridine orange staining indicated the
presence of a dsRNA in the cytoplasm of D. fragilis. This study of VLPs in different
isolates of D. fragilis, for the first time, extends the distribution of VLPs to another
inhabitant of the human gut and thus provides new knowledge on the cell biology of this
protozoan and a new protozoan virus.
3.2 Materials and methods
3.2.1 Culture of Dientamoeba fragilis trophozoites
Four D. fragilis isolates (isolates 1-4) used in this study were previously
described (Nagata et al. 2012a). All D. fragilis isolates were grown anaerobically at
37°C in tissue culture flasks with anaerobic or microaerophilic gas packs (Oxoid
AnaeroGenTM AN0035A (Oxoid, Australia)) in an anaerobic jar (BD GaspakTM EZ
(BD, Australia)). Loeffler’s serum slopes containing heat-inactivated horse serum
(Bovogen, Australia), glucose (Sigma, Australia) and nutrient broth (Sigma, Australia)

44
in distilled water were prepared as previously described (Barratt et al. 2010). Fifteen ml
of Loeffler’s medium were poured into a 50 ml culture flask (NuncTM 156367 (Thermo
Fisher, Australia)), sloped and inspissated in an 85°C drying oven until the serum slopes
solidified. Finally, the media was completed by addition of 15 ml PBS (Sigma) and
supplemented with 3-5 mg of sterilized rich starch (Sigma S7260).
3.2.2 Culture of Trichomonas vaginalis
Trichomonas vaginalis (isolate B7268) was kindly provided by Dr Jacqui
Upcroft (Queensland Institute for Medical Research). Trichomonas vaginalis was
grown in axenic culture using Diamond’s TYM media supplemented with 10% heat-
inactivated horse serum (González-Lázaro et al. 2005). Cultures (10 ml) were
maintained at 37°C in 15 ml or 50 ml conical polypropylene tubes on a ~15o slant. The
growth curves of T. vaginalis trophozoites were determined and subcultured at an initial
density of 1× 106 trophozoites/ ml.
3.2.3 Transmission electron microscopy
Cell suspensions of 1.0 x 106 D. fragilis trophozoites/ml were collected after 48
h of subculture and concentrated in 1.5 ml Eppendorf tubes (100 μl) which were then
centrifuged at 1,000 x g for 5 min. The supernatant was removed and cell pellets were
fixed in 3% (v/v) glutaraldehyde in PBS (0.1 M, pH 7.2) overnight at room temperature. The cells were then washed three times with PBS for 10 min each. Cells were prepared
for TEM as described by Banik et al. (2012).
3.2.4 Negative staining
Dientamoeba fragilis growth media (after 56 h of subculture) from all four
isolates were adsorbed onto Pioloform coated, 300 mesh, thin bar copper grids and fixed
with 3% glutaraldehyde in PBS for 5 min. The grids were washed 4 times for 5 min
each with PBS. Finally, all grids were washed with distilled water and negatively
stained with 2% uranyl acetate for 2 min. Grids were examined using a Philips CM10
transmission electron microscope.

45
3.2.5 VLPs purification
Approximately 5 x 106 D. fragilis trophozoites (isolate 1, after 72h of culture)
were pelleted by centrifugation at 1000 x g for 10 min. The supernatant (growth media)
was collected and filtered through a 0.2 μm filter (Minisart®, Sigma) and then
recentrifuged in an SW40 rotor (WX Ultra 80, Thermo Scientific, Australia) at 130,000
x g for 3h. After ultracentrifugation, pellets containing VLPs were further examined by
negative staining and electron microscopy as described above.
3.2.6 Isolation of dsRNA with phenol pH 8.0
Cultures of T. vaginalis (after 3 days) and D. fragilis (isolate 2, after 5 days)
were screened for presence of viral dsRNA (Flegr 1987). All steps of isolation were
performed at room temperature. Trichomonas (5 x 106 cells) were harvested by
centrifugation at 1000 x g for 10 min and D. fragilis (10 x 106 cells) were centrifuged at
1000 x g for 5 min. Dientamoeba trophozoites were lysed with sterile dH20 for about 15
min, vortexed vigorously for 5 min and checked for lysis under a light microscope. The
supernatant (growth media) was also collected after the first centrifugation and filtered
through a 0.2 μm filter (Minisart®, Sigma) and then recentrifuged in an SW40 rotor
(WX Ultra 80, Thermo Scientific, Australia) at 130,000 x g for 3 h to pellet the VLP
particles. Pellet material was added to the D. fragilis cell lysates (to increase numbers of
VLPs in the preparation) after which the lysate was again vortexed vigorously for 3
min. Finally, one volume of phenol pH 8.0 was added and the tube was tightly capped
and vigorously vortexed for 5 min. The phenol and aqueous phases were separated by
centrifugation at 13,000 x g for 20 min. The aqueous phase was re-extracted with 8
volumes of chloroform-isoamyl alcohol (v/v 24:1). After recentrifugation at the same
speed, the upper aqueous phase was transferred to a new tube. 0.75 volumes of
isopropanol were then added, and nucleic acids were precipitated from the mixture by
three cycles of freezing (in dry ice) and thawing (10 min each time). The precipitated
nucleic acid was collected by centrifugation at 13,000 x g for 20 min at 40C, washed
with 70% ethanol, dried and dissolved in TE buffer (10 mM Tris, 1 mM EDTA, pH
8.0).

46
3.2.7 DNase and RNase sensitivity test
To determine the identity of any nucleic acid purified from the VLPs, total
nucleic acids were incubated in appropriate enzymes prior to electrophoresis. Digestion
with DNase was carried out by incubation of total nucleic acids with 1 U RNase-free
DNase (Promega, Australia) for 1 h at 370C. The reaction was terminated by adding 1 U
stop solution in 10 mM EGTA (pH 8.0) (Promega, Australia) and incubated at 65°C for
10 minutes to inactivate the DNase. Digestion with RNase A was carried out by
incubation of total nucleic acids with 1 μg of RNase A (Sigma, Australia) for 1 h at 37 0C. Total nucleic acids were electrophoresed on a 0.8% agarose gel (80V/cm, 3 hours)
in 1x TBE buffer and stained with gel red.
3.2.8 Acridine orange staining
Dientamoeba fragilis isolate-2 and T. vaginalis (B7268) in the late logarithmic
phase of growth were collected and initial trophozoite density was 1.0×106
trophozoites/ml (as counted in a hemocytometer). Dientamoeba fragilis were
centrifuged at 1000 x g for 5 min and T. vaginalis were harvested by centrifugation at
1000 x g for 10 min. The cell pellets from both parasites were washed by PBS. Both
preparations were fixed in 4% (v/v) paraformaldehyde for 1 h and washed with PBS (4
times for 3 min). To prepare D. fragilis cells for acridine orange staining, coverslips (22
mm X 22 mm) were coated with 0.1% ethylene imine polymer solution (Fluka,
Switzerland). Cell suspensions (100 μl) of D. fragilis were added to the coverslips and
cells were allowed to adhere for 10 min at room temperature. The cells were then
washed 2 times with PBS for 3 min each. Finally, the adherent cells were stained in
0.01% acridine orange (AO, BDH, Chemical Ltd, England) in 0.067M phosphate buffer
solution (pH 6) for 5-10 min. Excess dye was removed by brief rinsing in PBS. The
coverslips were mounted onto microscopic slides and the edges sealed with nail polish.
Same procedures were followed to stain T. vaginalis (B7268 isolate). Slides were then
viewed under a Laser Scanning Confocal Fluorescence Microscope (Fluoview 300,
Olympus, Japan), using a Plan Apo 60X oil immersion objective, NA 1.42. An argon
ion laser (488 nm) was used for excitation of the acridine orange; the DNA emission
signal was collected at 515 nm (interference filter 510/530) and RNA emission signal
collected at 654 nm (longpass filter 610 nm). Optical slices of multidimensional images
were collected and analysed from constructed, extended focus images.

47
3.2.9 ImageJ analysis
A total of 100 VLPs were measured using ImageJ (1.46) program.
3.3 Results 3.3.1 Characterisation of VLPs
Virus-like particles were seen frequently inside the cytoplasm of D. fragilis cells
by TEM during ultrastructural analysis (Figure 3.1). Virus-like particles were observed
in all four clinical isolates (isolates 1- 4) of D. fragilis (data not shown). The sizes of
these VLPs were approximately 33- 40 nm. Their most common shape in thin sections
was spherical and formed heterogeneous populations. These VLPs contain: (1) an inner
dark core which encloses viral nucleic acid; and (2) an electron-dense capsid coat.
Furthermore, clusters of the VLPs were also observed in the cytoplasm (Figure 3.1).

48
Figure 3.1 Transmission electron microscopy showing the presence of virus-like
particles in the perinuclear region of Dientamoeba fragilis trophozoite. (All arrowheads
indicate VLP; N= nucleus, Nm= nuclear membrane, Np= nuclear pore, Mt=
microtubules).

49
Purified VLPs were obtained from trophozoite growth media filtered through a
0.2 μm filter and then concentrated by ultracentrifugation (Figure 3.2A). After negative
staining, the presence of VLPs in D. fragilis growth medium was confirmed (Figure
3.2B). Virus-like particles were found in growth media of all four isolates (data not
shown). Ultrastructural analysis showed that the viral envelope is covered by
capsomeres (data not shown). Comparatively, it appeared that VLPs were observed
more commonly in isolate 1 and isolate 4. In addition, the capsid structure of D. fragilis
VLP is compared with a polyhedral shape of viral capsid (Figure 3.2 C and D).
Figure 3.2 Electron micrograph of the purified virus-like particles. (A) Growth media
fractions (isolate 1) were collected from D. fragilis cultures and purified through 0.2 μm
filter. Virus-like particle showing an inner dark core which encloses the viral nucleic
acid. (Arrow indicate VLPs). (B) Virus-like particles in D. fragilis growth media. (C-D)
Comparison of D. fragilis VLPs structure with a polyhedral shape of viral capsid. (Figure C: Icosahedron shape of a viral capsid;
Source:http://www.sandforsk.se/articles/Virus%20structure,1-7,%20in%20spherical%20space.pdf).

50
3.3.2 Maturation pathway of D. fragilis virus-like particles
The maturation pathway of VLPs in D. fragilis was determined by TEM. Virus-like
particles were found outside the cells in growth medium (Figure 3.3A1). Virus-like
particles were also seen in coated pit-like structures formed on the plasma membrane
(Figure 3.3A2). Virus-like particles were seen attached to the parasite plasma membrane
and appear to enter trophozoites through endocytosis/internalization (Figure 3.3B).
Virus particles were also found internally in the vicinity of the plasma membrane
(Figure 3.3C) and in small vacuoles (Figure 3.3D). Virus-like particles were commonly
seen in these vacuoles before transferring into lysosomes. Subsequently, the VLP
particles appear to be rapidly transferred into lysosomes where viral uncoating occurs
(Figure 3.3E and F). Differences between microtubules structure and VLPs are shown
(Figure 3.3G). The synthesis of virions commences in the vicinity of the microtubules
(Figure 3.3G). Coated (electron-dense coating) and uncoated microtubules are found.
Coated microtubules are seen in the perinuclear region in the vicinity of the microtubule
organizing centre (Figure 3.3H and I). Accumulation of virus particles occurs in the
perinuclear region of cytoplasm. During VLPs maturation, microtubules appear to be
binding sites for the VLP genomes. Association occurs between microtubules and virus
particles for virus maturation. After association between microtubules and immature
viral material, virus progeny particles were found embedded within these coated
filaments (Figure 3.3J). Virus particles were also seen in close association with the
Golgi complex (Figure 3.3K). The Golgi complex showed dense particles closely
associated and adjacent to the Golgi cisternae. Progeny virus particles are seen
distributed over the cytoplasm (Figure 3.3L). Virus particles were also found in the
vicinity of endoplasmic reticulum (Figure 3.3M), axostyle and parabasal filament
(Figure 3.3N). A number of cells which contained a large number of VLPs appeared to
be lysed. Dientamoeba fragilis VLPs are observed in dying trophozoites present in in
vitro cultures (Figure 3.3O and P). In these cells, the nuclear structure is ruptured and
the cytoplasm is starting to break down and other organelles appear to be degenerating.
Probably VLPs are released from cells by cell lysis or exocytosis (data not shown). No
virus-like particles were found in nucleus.

51
Figure 3.3 The maturation pathway of VLPs in Dientamoeba fragilis. (A1) VLP in the
vicinity of the parasite plasma membrane (arrowheads). (A2) VLPs (arrowheads) are
seen in coated pit-like structure. (B) Virus-like particles appear to enter trophozoites
through endocytosis. (C) VLPs (arrowheads) seen inside D. fragilis cell.
(D) Subsequently, the virions were found in a small vacuole. (E, F) The VLPs are seen
in lysosomes. (G) Virions (arrowheads) are found in the perinuclear region around the
microtubule organizing region. (H, I, J) Immature VLPs (small arrow) associated with
microtubules and mature VLPs (arrowheads) were observed. (K) Golgi complex
associated with VLPs. (L) Progeny VLPs in cell. (M, N) Virus-like particles were also
found in the vicinity of endoplasmic reticulum and parabasal filament. (O, P) VLPs in
dying trophozoite. b= bacteria, CP= coated pit, ER= endoplasmic reticulum, Gc= Golgi
complex, H= hydrogenosomes, N= nucleus, Nm= nuclear membrane, Mt=
microtubules, L= lysosome, VLP= virus-like particles, SV= small vacuole, Pf=
parabasal filament, Pm= plasma membrane. (All arrowheads indicate VLPs).

52

53

54

55

56

57
3.3.3 Presence of VLPs in Trichomonas culture
Virus-like particles were found free in the cytoplasm of T. vaginalis cells
(B7268 isolate) by TEM (data not shown) which were used as a positive control. Virus-
like particles were also found in Trichomonas growth media (data not shown). Growth
media were collected at 2 to 5 days of subculture of T. vaginalis, negatively stained and
analysed by TEM. Virus-like particles were found by negative staining at 42 h to 96 h in
Trichomonas culture.
3.3.4 Investigations on D. fragilis VLPs genome
Total nucleic acids were extracted from T. vaginalis by a well-established
method commonly used to purify dsRNA from viruses of protozoa (Flegr, 1987). When
total nucleic acid samples from T. vaginalis were treated with DNase or RNase A prior
to electrophoresis, a 4 kb nucleic acid was DNase resistant and digested with RNase A
(Figure 3.4A, Lanes 1, 2, 3). This represents the dsRNA of T. vaginalis virus. In
contrast, no similar dsRNA bands were identified from D. fragilis. DNA of 1.7 kb, 2.8
kb and 4 kb were identified (Figure 3.4B, Lanes 1 and 3).

58
Figure 3.4 Gel-electrophoresis of total nucleic acids from T. vaginalis and D. fragilis
trophozoites. (A) Lane M: Molecular marker (1 kb ladder, HyperLadder™ I
BIOLINE); untreated total nucleic acid from T. vaginalis shown in lane 1 and nucleic
acid treated with DNase or RNase A is displayed in lanes 2 and 3, respectively (B)
Untreated total nucleic acid from D. fragilis trophozoites shown in lane 1 and nucleic
acid treated with DNase or RNase A is displayed in lanes 2 and 3, respectively. a, b and
c represent 4.0, 2.8 and 1.7 kb of the nucleic acids from D. fragilis xenic culture. No
obvious dsRNA is detectable from D. fragilis.
Acridine orange (AO) staining was also performed to investigate the nature of
the nucleic acid core of these VLPs. After staining the D. fragilis trophozoites, DNA
from both nuclei fluoresces light-green (Figure 3.5B) and the RNA is red (Figure 3.5C).
In addition, a yellowish-green color in the cytoplasm was observed suggesting the
presence of dsRNA in the cytoplasm (Figure 3.5D). Similar yellowish-green color was
also observed indicating the presence of dsRNA in the cytoplasm for T. vaginalis
(Figure 3.5H).

59
Figure 3.5 Acridine orange fluorescent staining of Dientamoeba fragilis trophozoite
(A-D) (isolate 2) and Trichomonas vaginalis B7268 isolate (E-H). (A) Differential
interference contrast (DIC) images (B) DNA from both nuclei fluorescence light-green
(C) presence of RNA fluorescence red (D) yellow-green color indicates probable
presence of dsRNA. Similar dsRNA signals were observed in Trichomonas. (E)
Differential interference contrast images (F) DNA from Trichomonas nuclei
fluorescence light-green (G) presence of RNA fluorescence red (H) yellow-green color
indicates probable presence of dsRNA. (N= nucleus).

60
3.4 Discussion
Analysis of thin sections of parasites and negatively stained samples by TEM
was performed to observe the presence of VLPs in cells and in growth medium from all
four D. fragilis isolates. Most previous studies used these techniques to identify VLPs in
different parasites (Wang & Wang 1991; Benchimol 2004). Much has been learned
about the presence of VLPs in T. vaginalis (Wang & Wang 1991). Numerous T.
vaginalis clinical isolates are persistently infected with double-stranded RNA viruses
(Goodman et al. 2011b). Similarly T. foetus (Gomes Vancini & Benchimol 2005) and
Trichomonas gallinae (Gerhold et al. 2009) contain VLPs. The presence of a dsRNA
virus was also described in Korean, Cuban, South African and Tehran T. vaginalis
isolates (Weber et al. 2003; Kim et al. 2007; Fraga et al. 2012; Heidary et al. 2013).
In the present work, the presence of VLPs was examined in different clinical
isolates (isolates 1-4) of D. fragilis for the first time. Very little information is available
about the biology of D. fragilis (Barratt et al. 2011b). Recently the surface structure and
ultrastructural characteristics of D. fragilis were described in great detail using electron
microscopy (Banik et al. 2012). This study provided extensive and novel details of
ultrastructure on four D. fragilis clinical isolates (isolates 1-4) growing in xenic culture
and the discovery of their VLPs was also reported. As part of a continuing study on D.
fragilis cell biology, VLPs were observed in all isolates of D. fragilis studied by TEM.
Virus-like particles were also observed in the growth medium in which D. fragilis was
cultured. In order to provide a comparison with other trichomonads, a culture of T.
vaginalis isolate B7268 was used as a control and VLPs were found in this isolate by
TEM.
Virus-like particles of D. fragilis vary in size between 33 to 40 nm in diameter.
Their most common shape observed in thin sections was spherical, enclosing a dense
core which encloses the viral nucleic acid and an outer capsid coat. These spherical
forms were observed in all four isolates. These results are similar with few studies of T.
vaginalis where heterogeneous populations of VLPs were described (Khoshnan &
Alderete 1993). Heterogeneous populations of VLPs are also found in Giardia (Cao et
al. 2009). Various shapes such as filamentous, cylindrical, spherical including oblong-
shaped forms of VLPs were observed in T. vaginalis (Benchimol et al. 2002).

61
Heterogeneous virus populations were also observed in T. foetus, mainly after the
parasites were treated with different cytoskeletal-affecting drugs (such as colchicine,
vinblastine, taxol, nocodazole, and griseofulvin) or drugs inducing cell death, such as
lactacystin and hydrogen peroxide (Gomes Vancini & Benchimol 2005). Benchimol et
al. (2002a) showed that T. vaginalis could be a reservoir for several different dsRNA
viruses simultaneously. In contrast, cDNA analysis of various trichomonads has shown
that they may be infected by multiple distinct but related non-segmented dsRNA viruses
(Tai & Ip 1995).
The presence of VLPs were observed more commonly in isolate 1 and isolate 4
in comparison to isolate 2 and isolate 3. The low abundance of VLP’s observed in
isolate 2 and 3 may have been due to their low density or latent conditions within these
isolates. Other studies have also reported that the virus can be lost during batch
cultivation (Benchimol et al. 2002a). In addition, these current ultrastructural
observations of VLPs for D. fragilis trophozoite were also largely dependent on the
plane of sectioning during TEM analysis. Some authors also claimed that they were not
able to find virus particles in T. vaginalis under electron microscopy (Wang & Wang
1986b).
Several maturation stages of D. fragilis virus particles were identified in this
study. The VLPs were found commonly within the cytoplasm of D. fragilis. The
maturation pathway of D. fragilis VLP is similar to other animal RNA viruses (Dalton
& Haguenau 1973) and dsRNA viruses described in T. vaginalis (Benchimol 2004) and
also in G. lamblia (Tai et al. 1993). It was observed that VLPs entered D. fragilis cells
by a process similar to endocytosis. Some VLPs were also observed in coated pit-like
structures during endocytosis. Similar phenomena were observed for Trichomonas virus
(Benchimol et al. 2002). Inside the D. fragilis trophozoite, virus particles were
commonly seen in the vicinity of the parasite plasma membrane which are not in
vacuole. The VLPs are often found in small vacuoles which are likely to be the product
of endocytosis. At the early events of VLPs maturation pathway, the VLPs were found
frequently in lysosomes. Trichomonas vaginalis viruses were also found in vacuoles
(Benchimol et al. 2002). The D. fragilis VLPs were also observed in the vicinity of
axostyle and parabasal filament. Trichomonas vaginalis virus is also found at the cell

62
periphery and close to the axostyle (Gomes Vancini & Benchimol 2005). It was
reported that Giardia lamblia virus (GLV) particles were initially found on plasma
membrane, translocated to the peripheral vesicles and then spread to the cytoplasm
(Wang & Wang 1986a; Benchimol et al. 2002).
No VLPs were found in D. fragilis nucleus whereas VLPs were observed in the
nucleus of Trichomonas and Giardia (Wang & Wang 1986a; Gomes Vancini &
Benchimol 2005). Both G. lamblia virus and Giardia canis virus were also found in the
nucleus (Cao et al. 2009). Interestingly, when both nuclei of the same G. lamblia
trophozoite were examined, they were found to contain similar numbers of the VLP
(Wang & Wang 1986a). Interestingly, VLPs were also observed in Trichomonas growth
media by TEM which is different from other studies on Trichomonasvirus (Goodman et
al. 2011a).
Synthesis of D. fragilis VLPs apparently commenced in the vicinity of
microtubules which were found frequently in the cytoplasm. It was found that
accumulation and assembly of D. fragilis VLPs occurs in the perinuclear region around
the microtubule organizing region. Studies reported that the replication of many viruses
occurred near the microtubule organizing center (MTOC) (Wileman 2006). A recent
study showed that the replication of Norovirus (a single-stranded RNA virus) is
associated with the MTOC (Hyde et al. 2012). Other RNA viruses also used the host
MTOC to complete their replication cycle (Netherton et al. 2007). The assembly of
capsids of these viruses occurred near the host MTOC (Life et al. 2008). The maturation
pathway for D. fragilis VLPs are similar to a recent study which proposed that the
cytoskeleton in trichomonads plays a role in the processing of virus maturation or
release (Gomes Vancini & Benchimol 2005; Vieira et al. 2008). Previous studies also
provided evidence that host cytoskeletal elements have important roles in the entry,
replication, and egress of HIV viruses (Smith & Enquist 2002). Moreover, studies
suggested that many viruses interact with cytoskeletal elements, and these interactions
are significant to complete the virus infectious cycle (Sodeik 2000; Smith & Enquist
2002). Importantly, this maturation pathway in D. fragilis is similar to RNA viruses
found in trichomonads (Benchimol et al. 2002).

63
In this study, VLPs were frequently observed in dying D. fragilis trophozoites
present in in vitro culture. Most of the cells containing clusters of viruses appeared to be
lysing. But whether viral load contributes to cell death is unknown. In cultures, lysis is
probably the main mechanism by which VLPs are released into the culture medium.
Any cytopathogenic effect of these viruses on D. fragilis cell biology is yet unknown.
Most of the recent studies indicate that TVV (Trichomonas vaginalis virus), GLV
(Giardia lamblia virus) and ENV (Eimeria necatrix virus) infections are generally
noncytopathogenic (Goodman et al. 2011a). It suggests a stable, persistent infection is
occurring in the protozoan cell (Goodman et al. 2011a). Trichomonas vaginalis strains
IR78 and CDC85 (known to contain no virus-like particles) were infected by purified T.
vaginalis virus but no dsRNA was detected in the cell homogenates (Alderete et al.
1985; Alderete et al. 1987). Consequently, several authors suggested the failure of viral
infection of T. vaginalis (Wang & Wang 1986b). Studies suggested that GLV can be
isolated from the stationary-phase growth of G. lamblia culture medium and used to
infect virus free strains of G. lamblia in vitro (Miller et al. 1988a, 1988b). It was
reported that GLV-infected G. lamblia trophozoites can harbor ~5 X 105 GLV per cell
and no cytopathogenic effects were observed on their rate of growth (Miller, Wang &
Wang 1988b). Olivier et al. (1984) found rhabdovirus-like structures in Entamoeba
histolytica and suggested that their presence was not related to the pathogenicity of the
amoeba. Moreover, the virus pellets purified from Entamoeba which were membrane-
free and showed no cytopathological effect in the host strain tested (Hruska et al. 1973).
There is no evidence that ENV 1 particles are infectious. Studies were unable to
demonstrate the transmission of the virus in vivo by coinfection of chickens with
Eimeria maxima either by co-cultivation with lysates of Eimeria nieschulzi or by
electroporation (Roditi et al. 1994). Chickens were also infected with E. necatrix
sporulated oocysts to determine the presence of both viral particles and capsid proteins
(del Cacho et al. 2001). No apparent infection of chicken cells was found suggesting
that E. necatrix viruses do not have extracellular routes of infection. Croft & Molyneux
(1979) found that promastigotes of Leishmania hertigi in culture seemed unaffected by
the presence of VLPs even though viral replication had taken place within the cells.
Many protozoan viruses are dsRNA viruses that were first identified by gel
electrophoresis (Pindak et al. 1989; Roditi et al. 1994; Ro & Patterson 2000; Goodman

64
et al. 2011a). Virus-like particles were observed in all four clinical isolates (isolates 1-4)
of D. fragilis by TEM and these viruses are morphologically similar to other protozoan
viruses (Banik et al. 2012). Based on TEM and acridine orange staining results we
therefore hypothesized that these D. fragilis VLPs are dsRNA- like viruses. To identify
viral nucleic acid from D. fragilis trophozoites, well established extraction methods
were used in this study. To validate the extraction method, T. vaginalis was used as a
control and dsRNA was obtained as per previous studies (Flegr 1987). It was also
reported in Flegr’s study that acid phenol (pH 4.4) gave the highest yields of dsRNA
from T. vaginalis but no difference was observed in our evaluations (data not shown).
No dsRNA was isolated from D. fragilis. There are several explanations to consider
from these results. Firstly, these VLP populations in D. fragilis may not be a dsRNA
virus. An alternative explanation is that these VLPs exist in very low abundance, and
may not be detectable using the cell numbers studied here. It is noteworthy to mention
that very low numbers of VLP (~ 3-4) were observed in each D. fragilis cell during
ultrastructural analysis. Interestingly, it was also reported that the difficulty in
identifying viruses in trichomonads could be due to the relatively low numbers present,
estimated at between 280 and 1350 particles per cell (Wang & Wang 1986b). In T.
foetus, VLPs were invisible in routine culture so the virions could be at low density or
not mature enough to detect (Gomes Vancini & Benchimol 2005).
Unfortunately D. fragilis grows to only low cell densities in culture making the
generation of large numbers difficult. Therefore, our next steps will be directed towards
obtaining more cells and investigating other available cultured isolates. If these are
unsuccessful, then we will investigate the presence of a DNA virus, similar to those
reported for some other protozoa (Van Etten et al. 2010; Legendre et al. 2012).
Recently several DNA viruses were discovered with genomes sizes greater than
300 kb to 1200 kb (Van Etten et al. 2010; Arslan et al. 2011; Van Etten 2011a,b). Some
of the well recognized DNA viruses are: (1) Mimivirus, which infects amoebae or
marine protists and has a genome ~1.2 Mb (La Scola et al. 2008; Claverie et al. 2009;
Claverie & Abergel 2009, 2010); (2) Mamavirus has an 18.3-kb DNA (La Scola et al.
2008); (3) Marseillevirus, which also infects amoebae (Claverie & Abergel 2009); (4)
Phycodnaviruses, which infects algae and has genomes up to ~560 kb; (5) White spot

65
shrimp viruses (WSSV) which have genomes up to ~305 kb (Boyer et al. 2011); and (6)
viruses that infect bacteria and has genomes up to ~670 kb. Initially, Mimivirus was
thought to be parasitic bacterium (La Scola et al. 2008; Claverie & Abergel 2009,
2010). These viruses inhabit aquatic environments and infect bacteria, animals and
protists (Van Etten 2011a); they are large viruses (often called giruses) with large
dsDNA genomes that encode up to 900 proteins and also many tRNAs (Colson &
Raoult 2010; Colson et al. 2011b; Van Etten et al. 2010). They also have more protein-
encoding genes than many bacteria and protozoan viruses. Obviously many large
viruses are expected to be discovered and there is a need for further research to be done
on these large viruses.
Indirect evidence for the presence of dsRNA in D. fragilis was obtained by
acridine orange (AO) staining. Acridine orange is a metachromatic intercalator and it
can differentiate double- from single-stranded nucleic acid (Lauretti et al. 2003).
Acridine orange has been used previously to study other parasites such as Entamoeba
histolytica (Hruska et al. 1973; Mattern et al. 1977), Blastocystis sp. (Suresh et al.
1994), Plasmodium (Hansen et al. 1970), Pneumocystis carinii (Thomson & Smith
1982) and T. vaginalis (Cevahir et al. 2002). It was previously shown that the AO
staining properties of viruses are quite distinctive; DNA is green, RNA is flaming red-
orange and dsRNA is yellow-green providing an accurate and relatively simple
procedure for determining the nature of the nucleic acid core of viruses (Dutta 1970).
When D. fragilis trophozoites were stained with AO, a yellow-green color was observed
in the cytoplasm which indicates the presence of double stranded RNA in the
cytoplasm. This suggests the D. fragilis VLPs may contain dsRNA.
Although most of the previous research on VLPs is based on electron microscopic
studies, more recent studies of viruses in protozoa have included molecular studies
(Goodman et al. 2011a; Han et al. 2011). Evidence suggests that the expression of a
virulence factor in T. vaginalis has been correlated with the presence of an endogenous
dsRNA virus (Goodman et al. 2011a). Interestingly, virus infection has also been
correlated with differential expression of many cellular proteins in T. vaginalis (Liu, et
al. 1998). The prevalence of TVV in different T. vaginalis clinical isolates was also
reported previously. It was reported before that infection rates as high as 82 and 75%

66
were found in clinical isolates from South Africa (Weber et al. 2003) and Baltimore
(Wendel et al. 2002) respectively. In addition, the prevalence was 55% from Cuba
(Fraga et al. 2005), 50% from USA (Snipes et al. 2000), and 44% from various other
geographic locations (Vanacova et al. 1997). In each of the listed studies, TVVs were
detected by gel electrophoresis of nucleic acids (Goodman et al. 2011a). Goodman et al.
(2011a) suggested that the improved diagnostics of TVV infection include
immunodetection methods with TVV-specific antibodies and nucleic acid microarrays
to detect TVV RNAs (Baptista et al. 2007).
To conclude, an increasing number of studies have been carried out to elucidate the
life cycle of D. fragilis and the epidemiology of disease, but there is a paucity of
knowledge on the mechanism of disease pathogenesis for this parasite species. The
identification and description of VLPs in D. fragilis is novel and represents a new
potential species of a protozoan virus that tentatively should be called Dientamoeba
fragilis virus (DFV) in keeping with the nomenclature for other viruses found in
protozoa. We propose a model which shows the development of VLPs in D. fragilis
trophozoites (Figure 3.6). Given the potential role of these VLPs in pathogenesis,
identification of viral genes and virulence factors need further investigation. The
description of virus in different clinical isolates of D. fragilis in this study will provide a
significant contribution in the molecular identification and phylogenetic analysis of
these newly identified protozoan viruses. More studies to determine the effect of DFV
on the pathogenesis and virulence of D. fragilis are also needed, as well as studies to
determine the role they play in the overall biology of the parasite and the host–parasite
interrelationship.

67
Figure 3.6 Working model for Dientamoeba fragilis VLPs development. Virus particles (indicated as VLP in figure) are found in the culture media and they
enter the cells by endocytosis/internalization, after which they are located in small
vacuoles and then they are transferred into lysosomes where viral uncoating occur.
Assembly of VLPs occur around the microtubule in the perinuclear region. Probably
VLPs are released from cell by cell lysis or exocytosis. (b= bacteria, VLP= Virus -like
particle, Sv= small vacuole, L= Lysosome, Mt= Microtubules).

68
Acknowledgements
This study was supported by funds from University of Technology, Sydney,
Australia and the Australian Research Council. All electron microscopy was conducted
at the Microscopy Unit, Faculty of Science, Macquarie University. This work was
presented at the Australia Society of Parasitology Annual Conference (on 3rd July
2012), Country Club, Launceston, Tasmania and the Gordon Research Seminar and
Conference on Physical Virology (on 20th of January, 2013) at Ventura, California,
USA.

69
Chapter 4:
Evaluation of different extraction methods to identify
Dientamoeba fragilis viral nucleic acid

70
4.1 Introduction
Dientamoeba fragilis is a trichomonad protozoan parasite that belongs to the
Parabasalia group (Gerbod et al. 2002; Cepicka et al. 2010; Stark et al. 2010b). It was
first described in the scientific literature in 1918 by Jepps and Dobell. The complete life
cycle of this parasite is still unknown and the trophozoite is the only well described
stage (Barratt et al. 2011b; Clark et al. 2014). Despite its widespread occurrence
remarkably little is known about the biology of D. fragilis. Recently the morphology of
this parasite was described by electron microscopy (Banik et al. 2012). Dientamoeba
fragilis is single-celled and ranges in size from 5 to 15 μm in diameter (Stark et al.
2006). It has two nuclei and contains equivalent amounts of DNA as determined by
DAPI staining (Banik et al. 2012).
Evidence suggests that the presence of virus-like particles (VLPs) are very
common in parasitic protozoa such as T. vaginalis (Goodman et al. 2011a, 2011b), T.
foetus (Gomes Vancini & Benchimol 2005), T. gallinae (Gerhold et al. 2009), G.
lamblia (Adam 2001), G. canis (Cao et al. 2009), Eimeria (del Cacho et al. 2001) and
Leishmania (Ro & Patterson 2000). The discovery of D. fragilis VLPs was reported
recently for the first time (Banik et al. 2012). Dientamoeba fragilis VLPs were
frequently seen outside the nucleus in the perinuclear region of the trophozoite
cytoplasm and closely associated with the Golgi complex (Banik et al. 2013,
unpublished, chapter 3). Virus particles were found in all four isolates (isolates 1-4) of
D. fragilis by TEM. These virus populations were also found in growth media in which
D. fragilis was cultured. In addition, the maturation pathway of D. fragilis VLPs in D.
fragilis trophozoite appears to be similar to that found in T. vaginalis virus (Banik et al.
2013, unpublished, chapter 3).
Trichomonas vaginalis virus was the first protozoan virus described in the
literature and biochemically characterised (Wang & Wang 1985, 1986b; Wang et al.
1987; Khoshnan & Alderete 1993). In the last few years, several studies had provided
detailed information on the presence and morphology of VLPs in T. vaginalis
(Benchimol et al. 2002a; Benchimol 2004; Kim et al. 2007), T. foetus (Gomes Vancini
& Benchimol 2005), and T. gallinae (Gerhold et al. 2009) based on electron
microscopy. Molecular studies on Trichomonasvirus have shown that several types of

71
VLPs can simultaneously infect T. vaginalis (Bessarab et al. 2000; Benchimol et al.
2002a). In these studies, rapid screening for the presence of double-stranded RNA
(dsRNA) was performed in various T. vaginalis isolates and the results suggested that
nearly all contained similar dsRNA. The number of dsRNA segments present varied
from 1 to 3 in different TVV isolates, and the length of those dsRNA segments varied
from 3.0 to 7.0 kb (Wang et al. 1987; Khoshnan & Alderete 1993; Su & Tai 1996).
Evidence also indicated that the dsRNA in T. vaginalis may be derived from virus-like
particles (Wang & Wang 1986b).
The most common method to purify dsRNA from different TVV particles was
phenol-chloroform extraction (Khoshnan & Alderete 1993; Kim et al. 2007). In most of
these studies, viral genomes were extracted with phenol-chloroform, precipitated with
ethanol/isopropanol and separated via 0.8-1% agarose gel electrophoresis (Wang et al.
1987; Khoshnan & Alderete 1993; Kim et al. 2007; Malla et al. 2011). After identifying
the viral nucleic acid, its sensitivity to DNase and RNase was assessed in order to
determine the nature of the nucleic acid present (Khoshnan & Alderete 1993).
Previously, acridine orange staining was used to provide evidence for the
presence of dsRNA in D. fragilis trophozoites and it suggests that the D. fragilis VLPs
may contain dsRNA (Banik et al. 2013, unpublished, chapter 3). The aim of this present
study was to select a suitable method to identify viral nucleic acid from the T. vaginalis
B7268 isolate (used as a positive control) and evaluate these optimized methods to
identify the D. fragilis viral genome. To determine the nature of the D. fragilis viral
genome, three extraction methods were evaluated and optimized. No obvious dsRNA
was detected in this study, suggesting that D. fragilis trophozoites do not contain an
abundant viral dsRNA like other trichomonads.
4.2 Materials and methods 4.2.1 Culture of D. fragilis trophozoites
Dientamoeba fragilis isolate E (isolate 2) was used in this study and grown
anaerobically at 37°C in Loeffler’s media with anaerobic or microaerophilic gas packs
(Oxoid AnaeroGenTM AN0035A (Oxoid, Australia)) in an anaerobic jar (BD GaspakTM
EZ (BD, Australia)). Loeffler’s serum slopes containing heat-inactivated horse serum

72
(Bovogen, Australia), glucose (Sigma, Australia) and nutrient broth (Sigma, Australia)
in distilled water were prepared as previously described (Barratt et al. 2010). Fifteen ml
of Loeffler’s medium were poured into a 50 ml culture flask (NuncTM 156367 (Thermo
Fisher, Australia)), sloped and inspissated in an 85°C drying oven until the serum slopes
solidified. Finally, the media was completed by addition of 15 ml PBS (Sigma) and
supplemented with 3-5 mg of sterilized rich starch (Sigma S7260).
In addition, D. fragilis isolate 2 was also grown for about 14 days to increase the
amount of virus in culture. After 4 days of subculture, rice starch, fresh PBS and
supernatant containing bacteria (from 2 days old culture flask) were added very gently.
Subsequently, at 8 days and 12 days after subculture the same amount of nutrients were
supplied. At 14 days, the cells were counted by haemocytometer and 1X 105/ ml was
found.
4.2.2 Culture of Trichomonas vaginalis
Trichomonas vaginalis (B7268 isolate) was kindly provided by Dr Jacqui
Upcroft (Queensland Institute for Medical Research). Trichomonas vaginalis was
grown in axenic culture using Diamonds’s TYM media supplemented with 10% heat-
inactivated horse serum (González-Lázaro et al. 2005). Cultures (~10 ml) were
maintained at 37°C in 15 ml or 50 ml conical polypropylene tubes on a ~15o slant. The
growth curves of T. vaginalis trophozoites were determined and subcultured at an initial
density of 1× 106 trophozoites/ ml. For CsCl density gradient methods, Trichomonas
cells (~ 200 ml) were cultured overnight in a 500 ml glass beaker and maintained
anaerobically at 37°C.
4.2.3 Purification of Trichomonas virus
Approximately 2x109 of logarithmic-phase-growing trichomonads (after 48 h of
culture) were pelleted by centrifugation at 1000 x g for 10 min and supernatant was
discarded. The cell pellet was washed three times with PBS. Trichomonads were lysed
with TNM buffer (50 mM Tris pH 7.5, 150 mM NaCl, 5 mM MgCl2) and sonicated at
about 1~2 min until more than 90% cells were lysed. The cell lysates were cleared by
centrifugation at 10,787g for 20 min (SORVALL(R) SUPER T-21, Kendro Laboratory,

73
Australia). After the centrifugation, the cell pellets were discarded and supernatant
containing virus was mixed with NaCl (0.5M) and PEG 6000 (to 10%) by gently
shaking. Trichomonas virus was precipitated overnight and recovered by centrifugation
at 10,787g for 20 min. The virus pellet was collected and dissolved in TM buffer (10
mM Tris HCl pH7.5, 2 mM MgCl2). Virus was further purified using stepped caesium
chloride (CsCl) density gradients. The virus suspension was layered over steps of ~ 1.3,
1.5, and 1.7g CsCl/ml in TM buffer and spun for 2 h at 40C in a swing out rotor (Sorvall (R) TH-641 rotor, WX Ultra 80, Thermo Scientific, Australia) at 119,952g. The virus
fraction, found at the interface of the 1.3 and 1.5 steps, was carefully removed using a
transfer pipette and dialysed overnight against 10 mM Tris (pH 7.5) and 2 mM MgCl2 at
40C.
4.2.4 Dientamoeba fragilis cell storage
Dientamoeba fragilis trophozoites were collected after every subculture and also
from 14 days culture for about two months. Cells were counted using a haemocytometer
and pelleted at 1000 x g for 5 min. The cell pellet was mixed with TYM media
supplemented with 50% glycerol and stored at -200C until the desired numbers of D.
fragilis trophozoites were obtained. Before purification, stored cells were dialysed
overnight against 10 mM Tris-HCl (pH 7.5) and 2 mM MgCl2 at 40C.
4.2.5 Extraction methods
All extraction methods were performed in triplicate. Total RNA was measured using the
Nanodrop. The absorbance is measured at 260 and 280 nm.
4.2.5.1 Method 1 (isolation of dsRNA with phenol pH 8.0)
Cultures of T. vaginalis B7268 (after 48 h) were screened for presence of viral
dsRNA as described previously (Flegr 1987). All steps of isolation were performed at
room temperature. Trichomonas (5 x 106 cells) were harvested by centrifugation at 1000
x g for 10 min and washed three times with PBS. One volume of phenol pH 8.0 was
added to cell pellets and the tube was tightly capped and vigorously shaken for 5 min.
The phenol and aqueous phases were separated by centrifugation at 13,000 x g for 20
min. The aqueous phase was re-extracted with 8 volumes of chloroform-isoamyl

74
alcohol (v/v 24:1). After recentrifugation at the same speed, the upper aqueous phase
was transferred to a new tube. 0.75 volumes of isopropanol were then added, and
nucleic acids were precipitated from the mixture by three cycles of freezing (in dry ice)
and thawing (10 min each time). The precipitated nucleic acid was collected by
centrifugation at 13,000 x g for 20 min at 40C, washed with 70% ethanol, dried and
dissolved in TE buffer (10 mM Tris, 1 mM EDTA, pH 8.0).
Dientamoeba fragilis cultures (isolate 2, after 72 h) were also screened for
presence of viral dsRNA using method 1. All steps of isolation were performed at room
temperature. Dientamoeba fragilis (1 x 107 cells) were pelleted at 1000 x g for 5 min
and washed three times with PBS. Dientamoeba trophozoites were lysed with sterile
dH20 for about 15 min, vortexed vigorously for 5 min and checked with light
microscope until more than 90% cells were lysed. The supernatant (growth media) was
also collected after the first centrifugation and filtered through a 0.2 μm filter
(Minisart®, Sigma) and then recentrifuged in an SW40 rotor (WX Ultra 80, Thermo
Scientific, Australia) at 130,000 x g for 3 h to pellet the VLP particles. Pellet material
was added to the D. fragilis cell lysates (to increase numbers of VLPs in the
preparation) and the lysate was again vortexed vigorously for 3 min. Finally, viral
nucleic acid was precipitated using method 1 as described for Trichomonas.
4.2.5.2 Method 2 (isolation of total nucleic acids)
Approximately 5 x 106 of logarithmic-phase-growing trichomonads (B7268
isolate, after 72 h of culture) were pelleted by centrifugation at 1000 x g for 10 min and
washed three times with PBS. The supernatant was discarded and Trichomonas cell
pellets were lysed in 1% SDS, 10mM Tris (pH 8.0), 10mM EDTA containing 100
μg/ml proteinase K for 2 h at 55°C. Total nucleic acids were extracted with phenol-
chloroform and precipitated with 100% ethanol in 0.2 M NaCl and dissolved in TE
buffer (10 mM Tris, 1 mM EDTA, pH 8.0).
Approximately 1 x 107 D. fragilis cells were centrifuged at 1000 x g for 5 min.
Cell pellets were collected and washed three times with PBS. Cell pellets were lysed in
1% SDS, 10mM Tris (pH 8.0), 10mM EDTA containing 100 μg/ml proteinase K for 2 h
at 55°C. Total nucleic acids were extracted with phenol-chloroform and precipitated

75
with 100% ethanol in 0.2 M NaCl and dissolved in TE buffer (10 mM Tris, 1 mM
EDTA, pH 8.0).
4.2.5.3 Method 3 (viral particle purification by CsCl density gradients and
extraction of RNA)
Approximately 2 x 109 Trichomonas cells were pelleted by centrifugation at
1000 x g for 10 min and washed three times with PBS. Trichomonas virus was purified
through CsCl density step gradients as described above (section 4.2.3). Trichomonas
virus fractions (~ 500 μl to 1 ml) were collected after purification. Total nucleic acids
were extracted using phenol (pH 8.0)/ chloroform from these viral fractions and
precipitated with isopropanol (as described in method 1).
To evaluate CsCl density gradient methods to purify D. fragilis VLPs, large
volume of growth media (~ 400 ml) was collected after 4 days of subculture. Media
were cleared by centrifugation two times at 10,787g for 20 min (SORVALL(R) SUPER
T-21, Kendro Laboratory, Australia). The supernatant was collected and filtered through
a 0.2 μm filter. After filtration, supernatants containing viruses were mixed with NaCl
(0.5M) and PEG 6000 (to 10%) by gently shaking. Virus was precipitated overnight and
recovered by centrifugation at 10,787g for 20 min. A tiny amount of virus pellet was
obtained and dissolved in TM buffer (10mM Tris HCl pH 7.5, 2 mM MgCl2). Virus was
further purified using CsCl density gradients. The virus suspension was layered over
steps of ~ 1.3, 1.5, and 1.7g CsCl/ ml in TM buffer and spun for 2 h at 40C in a swing
out rotor (Sorvall (R) TH-641 rotor, WX Ultra 80, Thermo Scientific, Australia) at
119,952g. A light band was observed and these probable virus fractions (~ 500 μl) were
carefully removed using a transfer pipette and dialysed overnight against 10mM Tris-
HCl pH 7.5, 2mM MgCl2 at 40C. Subsequently, total nucleic acids were extracted using
phenol (pH 8.0)/ chloroform and precipitated with isopropanol (as described in method
1).
Furthermore, greater than 2x109 D. fragilis trophozoites were collected from
glycerol stocks (described in section 4.2.4) and dialysed overnight against 10 mM Tris
(pH 7.5) and 2 mM MgCl2 at 40C. After dialysis, cells were dissolved in TM buffer and
sonicated at high speed. The cell lysates were cleared by centrifugation two times at
10,787g for 20 min. The pellets were discarded and supernatant containing viruses were

76
mixed with NaCl (0.5M) and PEG 6000 (to 10%) by gently shaking. Virus particles
were precipitated overnight and recovered by centrifugation at 10,787g for 20 min. A
very small virus pellet was collected and dissolved in TM buffer. Virus particle was
further purified using caesium chloride (CsCl) density gradients. The cell suspension
was layered over steps of ~ 1.3, 1.5, and 1.7g CsCl/ ml in TM buffer and spun for 2 h at
40C in a swing out rotor at 119,952g. After ultracentrifugation, the virus band was
carefully removed using a transfer pipette and dialysed overnight against 10 mM Tris
(pH 7.5) and 2 mM MgCl2 at 40C. Subsequently, viral nucleic acids were extracted
using phenol (pH 8.0)/ chloroform and precipitated with isopropanol (as described in
method 1).
4.2.6 DNase and RNase sensitivity test (for all three extraction methods)
To determine the identity of any nucleic acid purified from the VLPs, total
nucleic acids were incubated in appropriate enzymes prior to electrophoresis. The
sensitivity of nucleic acids extracted from VLP fractions to DNase and RNase were
tested for methods 1, 2 and 3. Digestion with DNase was carried out by incubation of
total nucleic acids with 1 U RNase-free DNase (Promega, Australia) for 1 h at 370C.
The reaction was terminated by adding 1 U stop solution in 10 mM EGTA (pH 8.0)
(Promega, Australia) and incubated at 65°C for 10 minutes to inactivate the DNase.
Digestion with RNase A was carried out by incubation of total nucleic acids with 1 μg
of RNase A (Sigma, Australia) for 1 h at 370C. Total nucleic acids were electrophoresed
on a 0.8% agarose gel (80V/cm, 3 hours) in 1x TBE buffer and stained with gel red.
4.2.7 Rotavirus, adenovirus and norovirus testing
The presence of any rotavirus, adenovirus and norovirus in D. fragilis cultures
(isolate 2) were tested. Four different viral samples were prepared from D. fragilis in
vitro culture (isolate 2) such that: (1) D. fragilis cultures were collected at 4 days of
subculture; (2) growth media were collected at 4 days of subculture; (3) growth media
were filtered through a 0.2 μm filter, centrifuged in an SW40 rotor (WX Ultra 80,
Thermo Scientific, Australia) at 130,000 x g for 3 h and pellets containing VLPs were
collected; (4) approximately 1x107 D. fragilis trophozoites were lysed using lysis buffer
(1% SDS, 10mM Tris (pH 8.0), 10mM EDTA containing 100 μg/ ml proteinase K) for
2 h at 55°C. All four samples were checked for the presence of rotavirus, adenovirus

77
and norovirus antigen using RIDA(R) QUICK Rotavirus /Adenovirus Combi (N1003)
and RIDA(R) QUICK Norovirus (N1403) according to the manufacturer’s
recommendations.
4.3 Results Three different extraction methods were optimized to recover viral nucleic acid
from T. vaginalis B7268 isolate. In method 1 (isolation of dsRNA with phenol pH 8.0),
no centrifugation steps were mentioned in the referred paper; therefore all centrifugation
steps were optimized during this method (see section 4.2.5.1). When total nucleic acid
samples from T. vaginalis were treated with DNase or RNase A prior to electrophoresis,
a nucleic acid of approximate size 4 kb was obtained that was DNase resistant and
digested with RNase A (Figure 4.1 A, method 1; lanes 1, 2, 3 ). This 4 kb band
represents the dsRNA of T. vaginalis virus that is well described in the literature (Flegr
1987).
In method 2 (isolation of total nucleic acids), a different lysis buffer was used to
prepare the Trichomonas virus. Total nucleic acids were extracted from T. vaginalis
(B7268 isolate) by phenol-chloroform and treated with DNase or RNase A prior to
electrophoresis. The 4 kb band was identified again and found to be DNase resistant and
digested with RNase A (Figure 4.1 B, method 2; lanes 4, 5, 6). The dsRNA did not
contain any genomic DNA contamination.
In method 3 (viral particle purification by CsCl density gradients and extraction of
RNA), Trichomonas virus were initially purified through CsCl density gradients and the
same 4 kb band was found in agarose gel electrophoresis after nucleic acid extraction
(Figure 4.1 C, method 3; lane 7). Trichomonas viral RNA obtained from these three
methods are shown in Table 4.1.

78
Table 4.1 Trichomonas viral RNA isolated using three different extraction methods.
Total sample
(μl)
Total RNA
(ng)
± SE A260/A280
Method 1 15 30365.45 ± 14439 1.96
Method 2 15 761.95 ± 265 1.87
Method 3 15 12967.45 ± 8441 2.03
(± SE, Standard error calculated from three replicates)

79
Figure 4.1 Comparison of three different extraction methods to identify viral nucleic
acid from T. vaginalis trophozoites (B7268 isolate). (A) Lane M: Molecular marker (1
kb ladder, HyperLadder™ I BIOLINE); untreated total nucleic acid from T. vaginalis
identified using method 1 shown in lane 1 and nucleic acid treated with DNase or
RNase A is displayed in lanes 2 and 3, respectively. (B) Untreated total nucleic acid
from T. vaginalis trophozoites using method 2 shown in lane 4 and nucleic acid treated
with DNase or RNase A is displayed in lanes 5 and 6, respectively. (C) A similar 4 kb
dsRNA band was found by CsCl density gradient (method 3) as shown in lane 7.

80
No viral dsRNA bands were identified from D. fragilis in vitro cultures and its
growth media using these three extraction methods. But DNA of 1.7 kb, 2.8 kb and 4 kb
were consistently observed by method 1 (Figure 3.4B).
To purify D. fragilis VLPs by CsCl density step gradients, large volume of
growth media and D. fragilis trophozoites were collected and analysed using method 3.
After ultracentrifugation, a light band was visible equal to 1.5% CsCl density from both
growth media and cells. There was no other visible band observed in the gradient. These
probable viral bands were collected and total nucleic acids were extracted by phenol-
chloroform (method 1). However, no viral RNA was obtained from growth media or D.
fragilis trophozoites. When total nucleic acid were isolated from growth media by
method 3 and treated with DNase or RNase A prior to electrophoresis, DNA of 4 kb
obtained (Figure 4.2 B, lanes 4, 5, 6). No significant band was observed from D. fragilis
cells by method 3 (data not shown).

81
Figure 4.2 Comparison of agarose gel electrophoresis of total nucleic acids isolated
from purified viral particles from Trichomonas vaginalis and Dientamoeba fragilis
growth media. (A) Lane M: Molecular marker; Trichomonas viruses were purified by
CsCl density gradient and untreated total nucleic acid shown in lane 1. Nucleic acid
treated with DNase or RNase A is displayed in lanes 2 and 3, respectively. (B) D.
fragilis VLPs were initially purified from growth media by CsCl density gradients and
untreated total nucleic acid is shown in lane 4; nucleic acid treated with DNase or
RNase A is displayed in lanes 5 and 6, respectively.
No rotavirus, adenovirus or noroviruses were found in D. fragilis cultures or
growth media in which D. fragilis was cultured.

82
4.4 Discussion
The present study aimed to identify the viral nucleic acids from Dientamoeba
fragilis VLP in cultured trophozoites (isolate 2) and its growth media. Initially, a
number of different extraction methods were screened to isolate RNA or dsRNA from
D. fragilis virus-like particles. To identify viral nucleic acid from D. fragilis in vitro
culture, total nucleic acids were extracted from D. fragilis trophozoites and analysed by
agarose gel electrophoresis as previously described for other species of protozoa (Wang
& Wang 1986a, 1986b; Tarr et al. 1988; Revets et al. 1989; Khoshnan & Alderete
1993; Han et al. 2011). Finally, three different extraction methods were optimized using
T. vaginalis B7268 isolate used as a positive control.
Virus-like particles were observed inside the cytoplasm of D. fragilis
trophozoites by TEM (Banik et al. 2012). Nearly all VLPs of protozoa are dsRNA
viruses ranging in diameter from 30-200 nm and the size of their genome is 5-7 kb
(Wang & Wang 1991; Benchimol et al. 2002a; Goodman et al. 2011a, 2011b).
Dientamoeba fragilis viruses are morphologically similar to other protozoan viruses
(Banik et al. 2013, unpublished, chapter 3). Therefore we hypothesized that D. fragilis
VLPs may contain dsRNA.
Two T. vaginalis isolates, T1 and B7268 isolates, were analysed to recognize the
presence of VLPs by TEM (data not shown). Virus-like particles were found only in
B7268 isolate. To validate the viral RNA extraction methods, B7268 isolate was used as
a positive control in this study. Dientamoeba fragilis cells and growth media were
collected at late logarithmic-phase to perform all extractions. Most of the Trichomonas
virus purification and dsRNA extractions were performed directly from late
logarithmic-phase or stationary phase cultures of T. vaginalis (Khoshnan & Alderete
1993; Kim et al. 2007). Using method 1, viral dsRNA was constantly obtained from
Trichomonas B7268 isolate as per previous studies (Flegr 1987). Acid phenol (pH 4.4)
gave the highest yields of dsRNA from T. vaginalis in Flegr’s study but no difference
was observed in our evaluations (data not shown). In contrast, no similar dsRNA was
found from D. fragilis cells and its growth media using this method. In addition, virus
was purified from growth media by 0.2 μm filtration and concentrated by
ultracentrifugation. A similar process was used to pellet Giardiavirus from growth

83
media (Miller et al. 1988b). Moreover, D. fragilis cell lysates were treated with
proteinase K (method 2) so that dsRNAs could not be protected within viral capsids.
Similar treatments were suggested for Trichomonas viruses (Wang & Wang 1991).
In this study, different fractionation methods were used to purify virus and
extract nucleic acids in them. Further, different types of lysis buffer as well as
sonication were used to digest D. fragilis trophozoites to increase the virus titre in cell
lysates. Dientamoeba fragilis virus is present in very low abundance in cultured cells
and in growth media as observed by TEM (Banik et al. 2013, unpublished, chapter 3). It
is noteworthy to mention that Dientamoeba trophozoites have complex vacuolar
systems that contain a variety of vesicles (Banik et al. 2013, unpublished, chapter 3).
Also D. fragilis uses rice starch in their culture as food source. During evaluation of
different extraction methods, carbohydrates were copurified with nucleic acids (data not
shown). As D. fragilis isolate 2 is isolated from clinical sample, the presence of any
rotavirus, adenovirus and norovirus in D. fragilis cultures and growth media were
tested. However, no rotavirus, adenovirus and norovirus were found in D. fragilis
cultures or growth media.
To determine whether D. fragilis VLPs can be purified from large volumes of
growth media, media were filtered and subject to CsCl density gradient centrifugation.
A very light band was observed in CsCl gradients. However, only DNA was obtained
from this probable viral band and no viral RNA was obtained in gel-electrophoresis.
Dientamoeba fragilis cells were also stored in glycerol solution to get large number of
trophozoites. But, no viral RNA was obtained. Several studies have used stored
protozoan cells or purified protozoan viruses in glycerol solution for study (Miller, et al.
1988a; del Cacho et al. 2001) and this seemed a logical way to accumulate large
numbers of D. fragilis trophzoites for study.
One major DNA band of 1.7 kb size and two faint staining bands of 2.8 kb and 4
kb were found running below the genomic DNA for D. fragilis using method 1. This 1.7
kb band obtained from D. fragilis trophozoites and growth media fraction was
consistently found during the optimization of different isolation methods. However, this
band was digested with DNase and RNase resistant showing that the 1.7 kb band was
DNA. In addition, DNA of 4 kb was also isolated from growth media by method 3. It is
unclear the relevance of these bands at this moment in time.

84
Martin et al. (2006) reported that more than 20 extractions were performed to
isolate dsRNA from virus infected blueberry plants and the dsRNA virus isolation was
very inconsistent. Moreover, most of the published protocols were unsuitable to isolate
these viruses (Choi & Randles 1997; Benthack et al. 2005; Martin et al. 2006). Even
though VLPs were purified via ultracentrifugation from T. gallinae they were not
observed by TEM (Gerhold et al. 2009). RNA was extracted from 12 T. gallinae
isolates but no dsRNA segments were visualized in agarose gels (Gerhold et al. 2009).
Even though the ultrastructure of the polyhedral virus V301 (isolated from Entamoeba
histolytica strain HB-301) was discovered a long time ago, there is still no molecular
evidence for these viruses yet (Hruska et al. 1973; Mattern et al. 1974; Mattern et al.
1979; Ackers & Mirelman 2006).
To conclude, no viral RNA or dsRNA was isolated from D. fragilis in vitro
culture (isolate 2). The possible explanation of these results could be due to firstly, the
VLP in D. fragilis may not be a dsRNA virus. In addition, VLP exists in very low
abundance, and simply may not be detectable using the cell numbers studied here. It
was previously reported that the difficulty in identifying viruses in trichomonads could
be due to the relatively low numbers present (Wang & Wang 1986b; Champney et al.
1995). Unfortunately, D. fragilis grows to only low cell densities in culture, making the
generation of large numbers difficult. Consequently, our next steps will be directed
towards obtaining more cells and analyzing other cultured isolates. However, the
presence of large DNA viruses in D. fragilis in vitro cultures also needs to be
investigated similar to those reported for some other protozoa (Colson & Raoult 2010;
Colson et al. 2011b; Legendre et al. 2012). We believe this current study will
considerably assist to identify D. fragilis viral nucleic acid in coming future.

85
Chapter 5:
Dientamoeba fragilis infection in children
Published as
G. R. Banik, J. L. N. Barratt, D. Marriott, J. Harkness, J. T. Ellis and D.
Stark, 2011. A case-controlled study of Dientamoeba fragilis infections in
children, Parasitology, 138, 819-823.

A case-controlled study of Dientamoeba fragilis infectionsin children
G. R. BANIK1,2,3, J. L. N. BARRATT1,2,3, D. MARRIOTT1,3, J. HARKNESS1,3,J. T. ELLIS2,3 and D. STARK1,3*1Division of Microbiology, SydPath, St Vincent’s Hospital, Darlinghurst, Australia2University of Technology Sydney, i3 Institute, Broadway, Australia3University of Technology Sydney, School of Medical and Molecular Biosciences, Broadway, Australia
(Received 12 February 2011; revised 17 February 2011; accepted 17 February 2011; first published online 27 April 2011)
SUMMARY
Dientamoeba fragilis is a pathogenic protozoan parasite that is implicated as a cause of human diarrhoea. A case-controlledstudy was conducted to determine the clinical signs associated with D. fragilis infection in children presenting to a SydneyHospital. Treatment options are also discussed. Stool specimens were collected from children aged 15 years or younger andanalysed for the presence of D. fragilis. In total, 41 children were included in the study along with a control group.Laboratory diagnosis was performed by microscopy of permanently stained, fixed faecal smears and by real-time PCR.Gastrointestinal symptoms were present in 40/41 (98%) of these children with dientamoebiasis, with diarrhoea (71%) andabdominal pain (29%) the most common clinical signs. Chronic gastrointestinal symptoms were present in 2% of cases. Themost common anti-microbial used for treatment was metronidazole (n=41), with complete resolution of symptoms andclearance of parasite occurring in 85% of cases. A treatment failure rate occurred in 15% of those treated with metronidazole.Follow-up treatment comprised of an additional course of metronidazole or iodoquinol was needed in order to achievecomplete resolution of infection and symptoms in this group. This study demonstrates the pathogenic potential ofD. fragilisin children and as such it is recommended that all laboratories must routinely test for this organism and treat if detected.
Key words: Dientamoeba fragilis, children, gastrointestinal symptoms, metronidazole, diarrhoea.
INTRODUCTION
Dientamoeba fragilis is a trichomonad-like protozoanparasite with a worldwide distribution commonlyfound in the gastrointestinal tract of humans (Starket al. 2006; Cepicka et al. 2010; Stark et al. 2010a,b;Barratt et al. 2011a). Despite widespread belief to thecontrary, numerous reports document thatD. fragilisis a common cause of gastrointestinal disease in bothdeveloped and developing regions of the world andhas the propensity to exist as a chronic infection withassociated clinical signs of disease (Grendon et al.1991, 1995; Dickinson et al. 2002; Norberg et al.2003; Stark et al. 2006, 2010b; Barratt et al. 2011a). Itis found in all patient groups studied so far includingthe general population, travellers and HIV-infectedindividuals (Stark et al. 2005, 2007a,b; Barratt et al.2011a).Several studies have reported that children are
susceptible to infection with D. fragilis and presentwith clinical symptoms at higher rates than adults(Preiss et al. 1991; Ayadi and Bahri, 1999; Crotti et al.2005). Consequently,D. fragilis should be consideredin the differential diagnosis of gastrointestinal
infections in children (Spencer et al. 1979;Keystone et al. 1984; Preiss et al. 1990; Stark et al.2009) but it is generally ignored as a cause ofdisease. Clinical symptoms reported include acuteand chronic diarrhoea, lower abdominal pain, nausea,flatulence and constipation (Spencer et al. 1983;Cuffari et al. 1998). Transmission of D. fragilis isbelieved to be via the faecal-oral route, but themechanism and whether it includes a helminth or acyst-like stage, are still unclear (Stark et al. 2006;Barratt et al. 2011b).Here we review existing knowledge on dientamoe-
biasis in children. A case-controlled study was alsoconducted to document the extent of D. fragilisinfections in children presenting to a major SydneyHospital.
MATERIALS AND METHODS
The study was performed at St Vincent’s Hospital,Sydney.Laboratory and clinical records ofD. fragilis-infected children were collected from August 2004to July 2010. The following criteria were used forinclusion into this study: children were aged 15 yearsor less, full clinical history, confirmed laboratorydiagnosis ofD. fragilis as the sole pathogen, treatmenthistory, follow-up stool samples were analysed toevaluate treatment regimes, clinical follow up and
* Corresponding author: Department of Microbiology,St Vincent’sHospital, Darlinghurst 2010, NSW,Australia.Tel: +61 2 8382 9196. Fax: +61 2 8382 2989 E-mail:[email protected]
819
Parasitology (2011), 138, 819–823. © Cambridge University Press 2011doi:10.1017/S0031182011000448

bacteriological cultures for enteric pathogens andvirological screening for the presence of rotavirusand enteric adenoviruses was performed. A total of41 children were included in the study along witha control group. A control group of children(established using the same criteria) free frominfection of D. fragilis and confirmed by PCR (asdescribed below), were included in the study. Wherepossible control subjects were age and sex matchedwith D. fragilis-infected children. However, this wasnot possible for 5 children. In the majority of thesecases (90%, 37/41) only a single stool sample wascollected and analysed.
Microbiological analysis
Laboratory diagnosis was performed by microscopyof permanently stained fixed faecal smears and byreal-time PCR as previously described (Stark et al.2010a). Bacterial cultures were performed usingstandard microbiological techniques to rule out thefollowing infections; Salmonella spp., Shigella spp.,Campylobacter spp., Yersinia enterocolitica, Vibriospp., Plesiomonas spp., Aeromonas and Clostridiumdifficile. Virology testing was performed by an im-munochromatographic screening test (Adeno/RotaSTAT-PAK; Chembio Diagnostic Systems Inc.,Sydney) for the detection of adenovirus and rotavirusantigen in faeces according to the manufacturer’srecommendations.
Statistical methods
Differences between means of gastrointestinal symp-toms of control subjects andD. fragilis-infected caseswere evaluated by using a Chi-Square (χ2) Test ofAssociation.
RESULTS
In total, 41 children were identified from laboratoryand hospital records as meeting the inclusion criteriafor the study. All 41 children had laboratoryconfirmed D. fragilis infection along with clinicalnotes detailing symptoms, anti-microbial treatmentregime and follow-up stool samples to check forparasite clearance following treatment. The studygroup ranged in age from 1 to 15 years with 88%(n=36) being less than 10 years of age (Fig. 1). Noneof the children were immunosuppressed. No bac-terial pathogens were isolated from the stools of the41 D. fragilis-infected children. All children’s stoolswere also negative for enteric adenoviruses androtaviruses. The control group was comprised of41 children, in which no D. fragilis was detected bymicroscopy and PCR. No bacterial or viral pathogenswere detected in this group.
Initially, 47 D. fragilis-infected children wereidentified from records; among them 10were infectedwith other enteric parasites; 4 with Blastocystishominis, 2 with Cryptosporidium spp. and B. hominis,2 with Endolimax nana, 1 with Entamoeba coli, and1 with Enteromonas hominis. As Cryptosporidium spp.is considered to be pathogenic and B. hominisis potentially pathogenic and capable of causinggastrointestinal symptoms (Yakoob et al. 2010),these 6 children were excluded from the data dealingwith symptoms and treatment. Of the children withD. fragilis infection, 98% (40/41) presented with atleast 1 or more gastrointestinal symptoms. Diarrhoea(71%) was found to be the most common symptomfollowed by abdominal pain (29%) (Table 1).Persistent or chronic infection was reported in 1child.
When compared to the control group diarrhoeawas significantly higher (P<0·002) in childrenwith dientamoebiasis. Other enteric protozoa werepresent in 2% (1/41) of control children. Originally,44 control patients were identified; however, 3 wereremoved because they harboured B. hominis and hadsymptoms of diarrhoea.
In total, 41 children were treated with metronida-zole and 35/41 (85%) reported complete resolutionof symptoms. Follow-up stool samples collectedbetween 1 and 4 weeks following treatment werecollected from these children and all showed clear-ance of the parasite. However, in 6 children, gastro-intestinal symptoms did not resolve following initialmetronidazole treatment. Four of those children weresubsequently treated with a repeat course of metro-nidazole for either 10 or 14 days and the other2 children were treated with iodoquinol. Followingthis additional anti-microbial therapy the childrenreported improved clinical symptoms and follow-upstool samples collected 2 weeks post-treatment werenegative for D. fragilis.
DISCUSSION
This study, conducted over a 6-year period, high-lights the association of D. fragilis with clinical signs
Fig. 1. Age of children with dientamoebiasis, (M, Male;F, Female).
820G. R. Banik and others

of disease: 98% of the children infected withD. fragilis studied presented with gastrointestinalsymptoms. Diarrhoea was found in 29/41 (71%) ofthe children and abdominal pain in 12/41 (29%). Thecontrol group represents a group of symptomaticchildren free of D. fragilis infection. Diarrhoea was,however, more common in children with D. fragilisinfection compared to the control group.There are other reports from various parts of the
world that also describe an association betweenD. fragilis infection in children and various clinicalsymptoms, most commonly diarrhoea and ab-dominal pain. A large study comprising over 43029children first reported a correlation betweenD. fragilis infection and symptoms of diarrhoea,abdominal pain and loose stools (Yang and Scholten,1977). In that study, chronic infections were found in2% of children. Spencer et al. (1983) reported a studyfrom 104 children, in which diarrhoea and abdominalpain were the most common symptoms in thosewithD. fragilis infection. Preiss et al. (1990) reportedthat among 102 children, 7 had acute diarrhoea, 39had relapsing diarrhoea, 7 had bloody stools and 29had abdominal pain. A retrospective study of 87Swedish children diagnosed with D. fragilis foundthe majority of children had symptoms of diarrhoea,abdominal pain and flatus (Norberg et al. 2003).Previous reports have also highlighted the propensityof the parasite to cause prolonged infection (Starket al. 2005; Crotti and D’Annibale, 2007) withchronic infections reported in the literature to lastas long as 2 years (Wenrich, 1944).Dientamoeba fragilis is a commonly encountered
enteric protozoan parasite in children that should beconsidered in any differential diagnosis of gastroin-testinal disease. Several studies have shownD. fragilisto be more prevalent than Giardia intestinalis inpaediatric populations (Preiss et al. 1990; Crotti and
D’Annibale, 2007; Rayan et al. 2007). One serologicalstudy carried out in children reported a D. fragilisseroprevalence of 91% (Chan et al. 1996). Whencompared to the seroprevalence of Giardia andCryptosporidium this study suggests that childhoodcontact with Dientamoeba is common (Chan et al.1996). Yang and Scholten (1977) found D. fragilisin 4·2% of individuals who submitted stools forparasitological examination during 1970–1974 inOntario, Canada. Infections were found to be morecommon in children, with nearly half of the infec-tions occurring in individuals under 20 years of age(Yang and Scholten, 1977). A recent study from theNetherlands found thatD. fragilis infection was mostcommon in children aged between 5 and 14 years(de Wit et al. 2001).Interestingly, an association was observed between
D. fragilis infection and carriage of other entericprotozoa normally transmitted via the faecal-oralroute. This association has previously been observed(Stark et al. 2010b) and suggests that transmission ofD. fragilis also occurs in the same way (by the faecal-oral route). There was no evidence for the presence ofhelminths in this study, which have previously beensuggested to be a host for D. fragilis (Stark et al.2006). Recent studies also suggest that helminthssuch as Enterobius vermicularis appear to play no rolein transmission of D. fragilis (Stark et al. 2010b,Barratt et al. 2011b).Anti-microbial treatment most commonly used for
treatment of D. fragilis infection in children includesmetronidazole and idoquinol (diidohydroxyquin)(Stark et al. 2010b). Therapy with metronidazolewas effective for most of the children in this study.Forty-one children were administered the drug,with the duration of treatment varying from 5 to10 days. Of the children treated with metronidazole85% (35/41) resulted in clearance of D. fragilis as
Table 1. Summary of results from children with Dientamoeba fragilis infection
D. fragilis-infectedchildren (n=41)
Control groupchildren (n=41)
Age range 1–15 1–15Sex – male 25 20
female 16 21Female/male ratio 1/1·6 1/0·9Other enteric protozoa present 2/41 (5%) 1/41 (2%)Clinical signs shown by childrenDiarrhoea 29/41 (71%) (P<0·002)* 14/41 (34%)Abdominal pain/discomfort 12/41 (29%) (NS) 9/41 (22%)Chronic diarrhoea (>2 weeks) 1/41 (2%) (NS) 0/41 (0%)Loose/abnormal stools 1/41 (2%) (NS) 2/41 (5%)Vomiting 1/41(2%) (NS) 2/41(5%)Cramps/constipation 0/41(0%) 1/41(2%)TreatmentMetronidazole 41/41Metronidazole treatment failures 6/41 (15%)
* P40·05, (NS, not significant).
821Dientamoeba fragilis infections in children

determined by analyses of follow-up stools andcomplete resolution of gastrointestinal symptoms.However, 6/41 (15%) children who underwentmetronidazole treatment failed to clear the infectionparasitologically or clinically. There was no corre-lation between the dose received, the duration oftreatment and treatment failure associated withmetronidazole use. These 6 children underwentfurther treatment, 4 were put on a repeat course ofmetronidazole for 2 weeks duration and the other2 children were treated with iodoquinol. On follow-up examination all children reported marked clinicalimprovement and clearance of parasite from stoolsamples. Iodoquinol has been widely used to treatD. fragilis infections (Butler, 1996). In a recentstudy, 27/33 children were successfully treated withclioquinol, a member of the same drug family asiodoquinol (Bosman et al. 2004).
There are varying reports on the efficacy ofmetronidazole treatment for D. fragilis infections inchildren. Spencer et al. (1979) reported that therapywith metronidazole was effective in 35 children.Similarly, in New Zealand, metronidazole wassuccessfully used in the treatment of dientamoebiasisin 3 children (Oxner et al. 1987). Preiss et al. (1990)studied 123 paediatric children with D. fragilisinfections. They found metronidazole to be effective,with 70% of children eliminating the parasite andsymptoms after a single treatment. A second treat-ment was required for 21 other children with anotherdrug. Ten children were treated a third time in orderto eliminateD. fragilis and accompanying abdominalcomplaints. They recommended a 10-day treatmentwith metronidazole for D. fragilis infections. Cuffariet al. (1998) showed that metronidazole was effectivein the treatment of 5 children. A study from Swedenincluded 32 children whom were treated withmetronidazole. The drug was given at various dosesfor various lengths of time, and they found that only4 children responded to the metronidazole treatment(Norberg et al. 2003). Recently, it was also docu-mented that 2 children presented with D. fragilisinfection over prolonged periods (Stark et al. 2009).In that study, metronidazole was used initially, butsubsequently treatment was done with paramomycinfor 10 days to effect clearance of the parasite. At thispoint in time, it is not clear why some cases ofdientamoebiasis do not respond to metronidazoletreatment; such observations may be the result ofmetronidazole resistance or failure to comply withmedication (Stark et al. 2010b).
In conclusion, this study serves to highlight thefact that children infected with D. fragilis typicallyhave diarrhoea. Studies such as those reported here,and elsewhere, strongly implicate D. fragilis as acommon cause of gastrointestinal disease in childrenand other patient populations (Stark et al. 2006,2010b; Barratt et al. 2011a). Dientamoeba fragiliswas not detected in the control group. We therefore
recommend that all laboratories must routinely testfor D. fragilis because treatment that eliminates theparasite usually results in the resolution of symp-toms. It is essential that a correct clinical andlaboratory diagnosis is made in order that treatmentcan be initiated.
ACKNOWLEDGMENTS
We acknowledge the help of the staff at St Vincent’sHospital Microbiology Department in the collection andprocessing of stool samples positive for D. fragilis. Part ofthis work was presented at theXIIth International Congressof Parasitology (ICOPA 2010), Melbourne, Australia(16 August, 2010). This research was supported by agrant from the St Vincent’s Hospital, Darlinghurst,Australia, University of Technology, Sydney andAustralian Research Council. Ms S. Fletcher providedadvice on statistics.
REFERENCES
Ayadi, A. and Bahri, I. (1999). Dientamoeba fragilis: pathogenic flagellate?Bulletin de la Société de Pathologie Exotique 92, 299–301.Barratt, J. L. N., Harkness, J., Marriott, D., Ellis, J. T. and Stark, D.(2011a). A review of Dientamoeba fragilis carriage in man: several reasonswhy this organism should be considered in the diagnosis of gastrointestinalillness. Gut Microbes 2, 3–12.Barratt, J. L. N., Harkness, J., Marriott, D., Ellis, J. T. and Stark, D.(2011b). The ambiguous life of Dientamoeba fragilis: the need to investigatecurrent hypotheses on transmission. Parasitology 138, 557–572.Bosman, D. K., Benninga,M. A., vanDe Berg, P., Kooijman, G. C. andvan Gool, T. (2004). Dientamoeba fragilis: possibly an important cause ofpersistent abdominal pain in children. Nederlands Tijdschrift VoorGeneeskunde 148, 575–579.Butler, W. P. (1996). Dientamoeba fragilis. An unusual intestinal pathogen.Digestive Diseases and Sciences 41, 1811–1813.Cepicka, I., Hampl, V. and Kulda, J. (2010). Critical taxonomic revisionof Parabasalids with description of one new genus and three new species.Protist 161, 400–433.Chan, F., Stewart, N., Guan, M., Robb, I., Fuite, L., Chan, I., Diaz-Mitoma, F., King, J., MacDonald, N. and Mackenzie, A. (1996).Prevalence ofDientamoeba fragilis antibodies in children and recognition of a39 kDa immunodominant protein antigen of the organism. EuropeanJournal of Clinical Microbiology & Infectious Diseases 15, 950–954.Crotti, D. and D’annibale, M. L. (2007). Intestinal infections caused byDientamoeba fragilis and Giardia duodenalis in our experience. RecentiProgressi in Medicina 98, 361–366.Crotti, D., D’annibale, M. L., Fonzo, G., Lalle, M., Caccio, S.M. andPozio, E. (2005). Dientamoeba fragilis is more prevalent than Giardiaduodenalis in children and adults attending a day care centre in Central Italy.Parasite 12, 165–170.Cuffari, C., Oligny, L. and Seidman, E. G. (1998). Dientamoeba fragilismasquerading as allergic colitis. Journal of Pediatric Gastroenterology andNutrition 26, 16–20.De Wit, M. A., Koopmans, M. P., Kortbeek, L.M., vanLeeuwen, N. J., Vinjé, J. and van Duynhoven., Y. T. (2001). Etiologyof gastroenteritis in sentinel general practices in the netherlands. ClinicalInfectious Diseases 33, 280–288.Dickinson, E. C., Cohen, M. A. and Schlenker, M. K. (2002).Dientamoeba fragilis: a significant pathogen. The American Journal ofEmergency Medicine 20, 62–63.Grendon, J. H., Digiacomo, R. F. and Frost, F. J. (1991). Dientamoebafragilis detection methods and prevalence: a survey of state public healthlaboratories. Public Health Reports 106, 322–325.Grendon, J. H., Digiacomo, R. F. and Frost, F. J. (1995). Descriptivefeatures of Dientamoeba fragilis infections. The Journal of Tropical Medicineand Hygiene 98, 309–315.Keystone, J. S., Yang, J., Grisdale, D., Harrington, M., Pillon, L.and Andreychuk, R. (1984). Intestinal parasites in metropolitan Torontoday-care centres. Canadian Medical Association Journal 131, 733–735.
822G. R. Banik and others

Norberg, A., Nord, C. E. and Evengård, B. (2003). Dientamoebafragilis – a protozoal infection which may cause severe bowel distress.Clinical Microbiology and Infection 9, 65–68.Oxner, R. B., Paltridge, G. P., Chapman, B. A., Cook, H. B. andSheppard, P. F. (1987). Dientamoeba fragilis: a bowel pathogen? The NewZealand Medical Journal 100, 64–65.Preiss, U., Ockert, G., Brömme, S., and Otto, A. (1990). Dientamoebafragilis infection, a cause of gastrointestinal symptoms in childhood.Klinische Pädiatrie 202, 120–123.Preiss, U., Ockert, G., Broemme, S. and Otto, A. (1991). On the clinicalimportance of Dientamoeba fragilis infections in childhood. Journal ofHygiene, Epidemiology, Microbiology, and Immunology 35, 27–34.Rayan, H. Z., Ismail, O. A. and El Gayar, E. K. (2007). Prevalence andclinical features of Dientamoeba fragilis infections in patients suspected tohave intestinal parasitic infection. Journal of the Egyptian Society ofParasitology 37, 599–608.Spencer, M. J., Garcia, L. S. and Chapin, M. R. (1979). Dientamoebafragilis. An intestinal pathogen in children? American Journal of Diseases ofChildren 133, 390–393.Spencer, M. J., Millet, V. E., Garcia, L. S., Rhee, L. andMasterson, L.(1983). Parasitic infections in a pediatric population. Pediatric InfectiousDiseases 2, 110–113.Stark, D., Barratt, J., Ellis, J., Harkness, J. and Marriott, D. (2009).‘Repeated Dientamoeba fragilis infections: a case report of two families fromSydney, Australia’. Infectious Disease Reports 1, 7–9.Stark, D., Barratt, J., Roberts, T.,Marriott, D.,Harkness, J. and Ellis, J.(2010a). Comparison of microscopy, two xenic culture techniques,conventional and real-time PCR for the detection of Dientamoeba fragilisin clinical stool samples. European Journal of Clinical Microbiology &Infectious Diseases 29, 411–416.
Stark, D., Barratt, J., Roberts, T., Marriott, D., Harkness, J. andEllis, J. (2010b). A review of the clinical presentation ofdientamoebiasis. The American Journal of Tropical Medicine and Hygiene82, 614–619.Stark, D., Beebe, N., Marriott, D., Ellis, J. and Harkness, J. (2005).Prospective study of the prevalence, genotyping, and clinical relevance ofDientamoeba fragilis infections in an Australian population. Journal ofClinical Microbiology 43, 2718–2723.Stark, D. J., Beebe, N., Marriott, D., Ellis, J. T. andHarkness, J. (2006).Dientamoebiasis: clinical importance and recent advances. Trends inParasitology 22, 92–96.Stark, D., Beebe, N., Marriott, D., Ellis, J. and Harkness, J. (2007a).Dientamoeba fragilis as a cause of travelers’ diarrhea: report of seven cases.Journal of Travel Medicine 14, 72–73.Stark, D., Fotedar, R., Van Hal, S., Beebe, N., Marriott, D., Ellis, J. T.and Harkness, J. (2007b). Prevalence of enteric protozoa in humanimmunodeficiency virus (HIV)-positive and HIV-negative men who havesex with men from Sydney, Australia. The American Journal of TropicalMedicine and Hygiene 76, 549–552.Wenrich, D. H. (1944). Studies on Dientamoeba fragilis (protozoa). IV.Further observations, with an outline of present-day knowledge of thisspecies. The Journal of Parasitology 30, 322–338.Yakoob, J., Jafri, W., Beg, M. A., Abbas, Z., Naz, S., Islam, M. andKhan, R. (2010). Blastocystis hominis and Dientamoeba fragilis in patientsfulfilling irritable bowel syndrome criteria. Parasitology Research 107,679–684.Yang, J. and Scholten, T. (1977). Dientamoeba fragilis: a reviewwith notes on its epidemiology, pathogenicity, mode of transmission,and diagnosis. The American Journal of Tropical Medicine and Hygiene 26,16–22.
823Dientamoeba fragilis infections in children

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

86
Chapter 6:
General discussion
And
Future directions

87
General discussion and future directions Dientamoeba fragilis is a pathogenic protozoan parasite found in the
gastrointestinal tract of humans with the propensity to cause chronic gastrointestinal
disease (Stark et al. 2010b, 2012). Dientamoeba fragilis was first reported almost a
century ago but very little is known about it, so many aspects of this protozoan are still
poorly understood (Johnson et al. 2004; Barratt et al. 2011a). Initially, D. fragilis was
considered as an amoeba, but subsequent molecular studies and phylogenetic analysis of
the small subunit ribosomal RNA (SSU rRNA) gene sequences, it is now recognized as
a trichomonad within the Phylum Parabasalia (Delgado-Viscogliosi et al. 2000; Peek,
et al. 2004; Stark et al. 2006). Numerous reports now document that D. fragilis is a
common cause of gastrointestinal disease in both developed and developing regions of
the world and has been found in several different patient populations including adults,
children and the immunosuppressed including those with HIV (Dickinson et al. 2002;
Girginkardesler et al. 2003; Stark et al. 2010b). The prevalence of this organism varies
widely and in faecal specimens from patients with diarrhoea is often more prevalent
than Giardia intestinalis (Crotti et al. 2005). Humans are probably the definitive host of
this parasite even though D. fragilis trophozoites have been reported in nonhuman
primates including macaques, baboons, gorillas and pigs (Barratt et al. 2011b; Cacciò et
al. 2012; Stark et al. 2012).
It is now obvious that there is more clinical evidence for D. fragilis as a
pathogen but little research on its biology has occurred in the last 40 years.
Improvement of cell biological techniques and recent advances in our knowledge of
other trichomonads led to this investigation on the cell biology of D. fragilis. The main
aim of this thesis was to investigate the structure and function of the D. fragilis
trophozoite. An EM study of the D. fragilis trophozoite led to a redescription of the
trophozoite structure and morphology. In addition, the presence of VLPs was identified.
This study therefore makes an excellent contribution to new knowledge of this lesser
known parasite.
In this study, I investigated the morphological characteristics of D. fragilis in
details by different microscopy methods (chapter 2) (Banik et al. 2012).
Morphologically, D. fragilis is single-celled; typically ranging from 5 to 15 μm in

88
diameter and it has two nuclei. Electron microscopy, fluorescence and light microscopy
were used to characterise D. fragilis populations growing in xenic culture. The
distinctiveness of external cell surface structures were examined by scanning electron
microscopy (SEM) and two different populations- smooth and ruffled cells were
observed. Scanning electron microscopy was also used to investigate possible
pseudocyst-like stages in this parasite’s life cycle. Temperature cooling experiments
were performed in an attempt to trigger pseudocyst formation. However, in this study it
was shown that there was no sign of flagella, pelta structure, undulating membrane and
pseudocyst-like forms in this protozoan’s life cycle. Moreover, the ultrastructure of four
isolates (isolates 1-4) of D. fragilis was described. Most of the organelles of D. fragilis
were observed by TEM. The presence of hydrogenosomes in D. fragilis is also
described which has not been previously reported.
The nuclear morphology of D. fragilis trophozoites grown in vitro and
trophozoites from clinical isolates were also compared by confocal microscopy and
light microscopy in our study. It was observed that the majority of cells grown in
culture were mononucleate while most cells in permanent stained faecal smears were
binucleate. Interestingly, it was also found that the two nuclei of D. fragilis are
morphologically indistinguishable and contain equivalent amounts of DNA.
Furthermore, the approximate cell and nuclear volume of four isolates of D. fragilis
were measured, which is comparable to other trichomonads.
Despite its discovery over a century ago, there has been no recent thorough
description of this parasite by microscopy methods. Recently, major advances were
made in in vitro culturing and cryopreservation of these four isolates of D. fragilis
(Barratt et al. 2010). In this thesis, I described the cell size, shape, surface organization,
ultrastructure of different organelles including nuclei numbers of different isolates of D.
fragilis (Banik et al. 2012). This study is the first SEM analysis of D. fragilis which
contributes to the understanding of the surface biology of this parasite. Ultrastructural
studies suggested that there is no phenotypic variation between four isolates (isolates 1-
4). This study revealed the presence of some common cytoskeletal organelles of
parabasalids such as costa and basal body in D. fragilis trophozoite. Dientamoeba
fragilis was reported as an anaerobic protozoan a long time ago but I showed for the

89
first time the anaerobic nature of the parasite by the absence of mitochondria (lacking
aerobic Kreb's cycle) and the presence of hydrogenosomes which is common in other
microaerophilic and anaerobes such as Trichomonas and Histomonas.
A tangential cut of the TEM block could lead us to misidentify the correct
number of nuclei, so approaches using confocal microscopy facilities showing the three
dimensional images of D. fragilis. The approximate cell volume and nucleus volume of
D. fragilis was observed for the first time which helps to estimate the genome sizes of
D. fragilis. This study provides details of the ultrastructure of this neglected protozoan
and helps explain phenomena associated with cell division, genetics and the proteome.
It is also important to determine whether the two nuclei of D. fragilis are structurally
and functionally identical as well as to understand the evolutionary, physiological and
genetic significance of these two physically separate, but equal-sized, nuclei (Banik et
al. 2012). In addition, the discovery of a virus-like particle (VLP) in D. fragilis
trophozoite is also reported for the first time (Banik et al. 2012).
An up to date review of all the related literature to present on gut protozoan
viruses and trichomonad protozoan viruses is presented in chapter 1. I described their
discovery, morphology details, different purification methods of VLPs, genome and
proteome and finally their effect on parasites disease. Definitely, this review provides
broad knowledge on gut protozoan viruses.
My third aim was to investigate the presence and morphological details of the D.
fragilis VLPs in all four isolates (chapter 3). Virus–like particles were frequently found
in four isolates of D. fragilis and the growth media in which these four isolates were
cultured. Virus particles were heterogeneous in size and approximately 33- 40 nm in
diameter. Their most common shape was spherical and contained an inner dark core
surrounded by electron dense capsid coat. These VLPs have a similar structure to a
polyhedral capsid structure (as seen in chapter 3). I speculate these VLPs attach to the
trophozoite and penetrate into D. fragilis by a similar mechanism to endocytosis. It is
observed that VLPs are maintained within vacuoles mainly in lysosomes during several
passages. The D. fragilis VLPs were extensively found outside the nucleus around
microtubules in the perinuclear region of the trophozoite. Coating of microtubules was

90
also seen in the perinuclear region around the microtubule organizing region. Also,
accumulation of VLPs occurs in the perinuclear region of cytoplasm. Dientamoeba
fragilis VLPs were also commonly found in association with the Golgi complex. During
these ultrastructural observations, VLPs were frequently seen distributed throughout the
cytoplasm. Interestingly, clusters of VLPs were observed in dying trophozoites compare
to healthy cells present in in vitro cultures. The structures of these cells appeared to
rupture and cytoplasm was starting to break down. But it is still unknown whether viral
load contributes to cell death or not. However, in culture this is probably the main
mechanism by which VLPs are released into the culture medium.
No VLPs were reported before in Dientamoeba fragilis. But the presence of
virus is very common in parasitic protozoa especially those found in the gut such as
Giardia (Wang & Wang 1986a), Eimeria (Ellis & Revets 1990), Cryptosporidium
(Khramtsov et al. 1997) and Blastocystis (Teow et al. 1992) as well as other protozoa
such as Trichomonas (Benchimol et al. 2002b; Benchimol 2004) and Leishmania (Tarr
et al. 1988). All of them contain VLPs containing dsRNA (Goodman et al. 2011a;
Wang & Wang 1991). Our study was the first study to describe in detail the
ultrastructural characteristics of a Dientamoeba fragilis virus (DFV) and its mode of
maturation in different cultured isolates of D. fragilis (Banik et al. 2013, unpublishd,
chapter 3). This study extends the distribution of VLPs to another inhabitant of the
human gut and thus provides new knowledge on the cell biology of this protozoan and a
new protozoan virus.
The final aim was to purify D. fragilis virus and investigate the identity of its
virus genome (chapter 4). Initially, D. fragilis virus were purified from growth media
filtered through a 0.2 μm filter and concentrated by ultracentrifugation. Purified VLPs
were observed by negative staining and all contained an intact capsid. Dientamoeba
fragilis viruses are consistent with the size and shape of other protozoan viruses.
Moreover, acridine orange staining suggested the presence of dsRNA in D. fragilis
trophozoite (as seen in chapter 3). Therefore we hypothesized that D. fragilis may
contain dsRNA. Acridine orange staining was also used to detect the viral nature of
another gut protozoan Entamoeba histolytica (Hruska et al. 1973; Mattern et al. 1977).

91
To identify D. fragilis viral RNA, I evaluated different extraction methods
which were commonly used to identify viral RNA from other trichomonads. To
evaluate these extraction methods, T. vaginalis B7268 isolate was used as a positive
control and dsRNA was detected. The presence of VLPs were also detected in this
Trichomonas isolate by TEM (described in chapter 3). Trichomonas virus was also
purified by CsCl density gradient centrifugation and dsRNA was identified by gel
electrophoresis.
However, despite long intensive efforts to identify the D. fragilis viral genome,
this was not achieved. Also despite vigorous validation of dsRNA extraction methods,
no viral-like RNA was purified from D. fragilis cells and growth media. I have analysed
all possible purification techniques such as filtration, differential centrifugation,
ultracentrifugation and gradient methods used to purify other protozoan virus. In
addition, different fractionation methods including DNase treatment to concentrate
RNA viruses and cellulose treatment to bind dsRNA were performed. There are several
explanation for these results such as the numbers of cells used for virus purification. It
is possible to grow large amounts of T. vaginalis cells in an overnight culture and these
cultures do not contain bacteria. In contrast, D. fragilis grows only at low cell densities
in culture making the generation of large numbers difficult. Moreover, the ability to
maintain Dientamoeba in in vitro culture for prolonged periods of time at high cell
densities is extremely difficult and cultures contain different species of bacteria (Barratt
et al. 2010).
Furthermore, only low numbers of viral particles were found in D. fragilis
trophozoites during ultrastructural analysis which were comparatively very low
compared to Trichomonas virus. Evidence suggested that the difficulty in identifying
viruses in trichomonads could simply be due to the relatively low numbers present
(Wang & Wang 1991; Gomes Vancini & Benchimol 2005). In Giardiavirus research, it
has been reported that at least 104 viral particle were needed to infect virus free Giardia
or to do any experiment like immunostaining analysis (Sepp et al. 1994). I suspect the
reason I was unable to identify the D. fragilis VLPs genome is due to low viral
abundance in the D. fragilis trophozoite and the number of D. fragilis trophozoites that
can be produced by culture. To overcome this problem a large number of cells (greater

92
than 109) and large volume of growth media were used in an attempt to purify VLP
from D. fragilis VLPs. No viral nucleic acid was observed in agarose gel
electrophoresis. Consequently it was concluded from these evaluations of different
methods that unlike T. vaginalis, D. fragilis trophozoites do not contain an abundant
viral dsRNA. As all D. fragilis isolates (isolates1-4) used in this study were isolated
from clinical sources, the presence of rotavirus, norovirus and adenovirus were tested
(chapter 4). However, no rotavirus, adenovirus or noroviruses were found in D. fragilis
cultures or growth media.
Importantly, another gut protozoan virus, Entamoebavirus was first discovered
in 1960 by electron microscopy but still no molecular evidence is available (Hruska, et
al. 1974; Mattern et al. 1974). Some authors did not find any VLPs in Trichomonas by
EM (Wang & Wang 1986b). In fact it was challenging to purify most of the protozoan
viruses because of their abundance in cells, latent condition, loss of viruses after several
passages and viability issues during storage (Benchimol et al. 2002a). There are also a
few studies reported that when virus free T. vaginalis strains (such as IR78 and CDC85)
were infected by purified VLPs, no dsRNA were detected by molecular identification
(Alderete et al. 1985; Wang & Wang 1986b; Alderete et al. 1987). In T. gallinae, VLPs
were purified via ultracentrifugation but not observed by TEM or no dsRNA segments
were visualized in agarose gels (Gerhold et al. 2009). The point to be made by these
studies is that purification and detection of VLPs from protozoa is difficult and not
solved easily.
All D. fragilis isolates (isolates 1-4) used in this study harboured VLPs detected
by TEM. A difference between Trichomonasvirus, Giardiavirus and D. fragilis VLPs is
that D. fragilis VLP was only found in the cytoplasm whereas TVV and GLV are also
found in the nucleus. Also in T. foetus, VLPs were detected only after the trichomonads
were treated with cytoskeleton-affecting chemicals including colchicine, vinblastine,
taxol, nocodazole, and griseofulvin (Gomes Vancini & Benchimol 2005). This is one
approach to consider in the future as a method that would assist in the purification of
VLPs from D. fragilis trophozoites.

93
The epidemiology of dientamoebiasis is still severely under investigated and as a
small side project I investigated hospital records for infections of children. Several
studies have reported that children may be susceptible to infection with D. fragilis and
present with clinical symptoms at higher rates than adults (Preiss et al. 1991; Ayadi &
Bahri 1999; Crotti et al. 2005). Consequently, D. fragilis infection has been suggested
to be common in children (Preiss et al. 1991). Studies also have shown D. fragilis to be
more prevalent than Giardia intestinalis in paediatric populations (Crotti & D'Annibale
2007; Rayan et al. 2007). Therefore, existing knowledge on dientamoebiasis in children
was reviewed in this study. In addition, the pathogenic potential of D. fragilis was
examined in a case-controlled study of dientamoebiasis in children presenting to a
Sydney Hospital (chapter 5).
A case-controlled study on D. fragilis infected children aged 15 years or
younger was performed at St. Vincent’s Hospital, Sydney. A total of 41 children were
included in the study along with a control group. A total of 41 children were identified
from laboratory and hospital records as meeting the inclusion criteria for the study and
had laboratory confirmed D. fragilis infection along with clinical notes detailing
symptoms, antimicrobial treatment regime and follow up stool samples to check for
parasite clearance following treatment. A control group of children (established using
the same criteria), free from infection of D. fragilis that was confirmed by PCR was
included in this study. During this study, laboratory diagnosis was performed by
microscopy of permanently stained fixed faecal smears and by real-time PCR as
described previously (Stark et al. 2010a).
Results showed that gastrointestinal symptoms were present in 40/41 (98%) of
these children with dientamoebiasis. Diarrhoea (71%) was found to be the most
common symptom followed by abdominal pain (29%). In this study, the difference
between infected and uninfected was statistically significant. Moreover, chronic
gastrointestinal symptoms were present in 2% of cases. The most common
antimicrobial used for treatment was metronidazole (n=41), with complete resolution of
symptoms and clearance of parasite occurring in 85% of cases. Also a treatment failure
rate of 15% was identified in children treated with metronidazole. In this group, follow-

94
up treatment comprised of an additional course of metronidazole or iodoquinol was
needed in order to achieve complete resolution of infection and symptoms.
This study highlights the pathogenic potential of D. fragilis and strongly
implicates it as a common cause of gastrointestinal disease in children. Dientamoeba
fragilis is a commonly encountered enteric protozoan parasite in children that should be
considered in any differential diagnosis of gastrointestinal disease. There was a marked
increase in the number of gastrointestinal symptoms of the D. fragilis infected cohort
when compared to the D. fragilis negative cohort. It is strongly recommended that all
laboratories must routinely test for D. fragilis as treatment which eliminates the parasite
usually results in the resolution of symptoms.
Overall, this thesis focused on investigating various cell biological aspects for
this neglected parasite. This extensive investigation based on different microscopy and
molecular methods uncovered many novel morphological aspects of this protozoan.
This study will make an excellent contribution to future research on protozoan biology,
especially for examining lesser known parasites. Furthermore, very little is known
regarding different protozoan viruses yet. The discovery of D. fragilis viruses and
description of its development within the cell has exposed many new areas to study
such as the effect of these viruses on parasite’s diseases pathogenesis, potential role as a
vector for the genetic transformation and association in host-parasite relationship. An
exciting period of research is ahead for upcoming researchers in this arena.

95
Chapter 7:
References
(For chapter 1, chapter 3, chapter 4 and chapter 6)

96
References
Ackers, J.P. & Mirelman, D. 2006, 'Progress in research on Entamoeba histolytica
pathogenesis', Current Opinion in Microbiology, vol. 9, no. 4, pp. 367-73.
Adam, R.D. 2001, 'Biology of Giardia lamblia', Clinical Microbiology Review, vol. 14,
no. 3, pp. 447-75.
Alderete, J.F., Demĕs, P., Gombosová, A., Valent, M., Yánoska, A., Fabusová, H.,
Kasmala, L., Garza, G.E. & Metcalfe, E.C. 1987, 'Phenotypes and protein-
epitope phenotypic variation among fresh isolates of Trichomonas vaginalis ',
Infection and Immunity, vol. 55, no. 5, pp. 1037-41.
Alderete, J.F., Kasmala, L., Metcalfe, E. & Garza, G.E. 1986, 'Phenotypic variation and
diversity among Trichomonas vaginalis isolates and correlation of phenotype
with trichomonal virulence determinants', Infection and Immunity, vol. 53, no. 2,
pp. 285-93.
Alderete, J.F., Suprun-Brown, L., Kasmala, L., Smith, J. & Spence, M. 1985,
'Heterogeneity of Trichomonas vaginalis and discrimination among trichomonal
isolates and subpopulations with sera of patients and experimentally infected
mice', Infection and Immunity, vol. 49, no. 3, pp. 463-8.
Alderete, J.F., Wendel, K.A., Rompalo, A.M., Erbelding, E.J., Benchimol, M. & Chang,
T.H. 2003, 'Trichomonas vaginalis: evaluating capsid proteins of dsRNA viruses
and the dsRNA virus within patients attending a sexually transmitted disease
clinic', Experimental Parasitology, vol. 103, no. 1-2, pp. 44-50.
Arslan, D., Legendre, M., Seltzer, V., Abergel, C. & Claverie, J.M. 2011, 'Distant
Mimivirus relative with a larger genome highlights the fundamental features of
Megaviridae', Proceedings of the National Academy of Sciences, vol. 108, no.
42, pp. 17486-91.
Ayadi, A. & Bahri, I. 1999, 'Dientamoeba fragilis: pathogenic flagellate?', Bulletin de la
Société de pathologie exotique, vol. 92, no. 5, pp. 299-301.
Banik, G.R., Barratt, J.L.N., Marriott, D., Harkness, J., Ellis, J.T. & Stark, D. 2011, 'A
case-controlled study of Dientamoeba fragilis infections in children',
Parasitology, vol. 138, no. 07, pp. 819-23.
Banik, G.R., Birch, D., Stark, D. & Ellis, J.T. 2012, 'A microscopic description and
ultrastructural characterisation of Dientamoeba fragilis: An emerging cause of

97
human enteric disease', International Journal for Parasitology, vol. 42, no. 2,
pp. 139-53.
Banik, G.R., Birch, D., Stark, D. & Ellis, J.T. 2013, 'Virus-like particles in
Dientamoeba fragilis: an ultrastructural study', Unpublished. (Presented at
Gordon research seminar and conference on physical virology, Ventura,
California, USA, 19-25th, 2013).
Baptista, C.S., Wu, X. & Munroe, D.J. 2007, 'International patent WO/2007/130519,
Viral nucleic acid microarray and method of use.', World Intellectual Property
Organization, Geneva, Switzerland.
Barratt, J.L.N., Banik, G.R., Harkness, J., Marriott, D., Ellis, J. T& Stark, D. 2010,
'Newly defined conditions for the in vitro cultivation and cryopreservation of
Dientamoeba fragilis: new techniques set to fast track molecular studies on this
organism', Parasitology, vol. 137, no. 13, pp. 1867-78.
Barratt, J.L.N., Harkness, J., Marriott, D., Ellis, J.T. & Stark, D. 2011a, 'A review of
Dientamoeba fragilis carriage in humans: several reasons why this organism
should be considered in the diagnosis of gastrointestinal illness.', Gut Microbes,
vol. 2, no. 1, pp. 3-12.
Barratt, J.L., Harkness, J., Marriott, D., Ellis, J.T. & Stark, D. 2011b, 'The ambiguous
life of Dientamoeba fragilis: the need to investigate current hypotheses on
transmission', Parasitology, vol. 138, no. 05, pp. 557-72.
Barratt, J., Ellis, J., Harkness, J., Marriott, D. & Stark, D. 2013, ‘Evaluation of the
in vitro Antiprotozoal activity of various dry plant extracts against
Dientamoeba fragilis’, Journal of Infectious Diseases and Therapy, vol. 1,
no. 03, pp. 111.
Benchimol, M. 2004, 'Trichomonads under microscopy', Microscopy and
Microanalysis, vol. 10, no. 5, pp. 528-50.
Benchimol, M., Chang, T.H. & Alderete, J.F. 2002a, 'Trichomonas vaginalis:
observation of coexistence of multiple viruses in the same isolate', FEMS
Microbiology Letters, vol. 215, no. 2, pp. 197-201.
Benchimol, M., Chang, T.H. & Alderete, J.F. 2002b, 'Visualization of new virus-like-
particles in Trichomonas vaginalis', Tissue and Cell, vol. 34, no. 6, pp. 406-15.

98
Benchimol, M., Monteiro, S.P., Chang, T.H. & Alderete, J.F. 2002,'Virus in
Trichomonas—an ultrastructural study', Parasitology International, vol. 51, no.
3, pp. 293-8.
Benthack, W., Mielke, N., Büttner, C. & Mühlbach, H.P. 2005, 'Double-stranded RNA
pattern and partial sequence data indicate plant virus infection associated with
the ringspot disease of European mountain ash (Sorbus aucuparia L.)', Archives
of Virology, vol. 150, no. 1, pp. 37-52.
Bessarab, I.N., Nakajima, R., Liu, H.W. & Tai, J.H. 2011, 'Identification and
characterization of a type III Trichomonas vaginalis virus in the protozoan
pathogen Trichomonas vaginalis', Archives of Virology, vol. 156, no. 2, pp. 285-
94.
Bessarab, I.N., Liu, H.W., Ip, C.F. & Tai, J.H. 2000, 'The complete cDNA sequence of
a type II Trichomonas vaginalis virus', Virology, vol. 267, no. 2, pp. 350-9.
Bessarab, I.N. & Tai, J.H. 2001, 'Trichomonas vaginalis virus 3 capsid protein (cap) and
RNA-dependent RNA polymerase (pol) genes, complete cds.', Genbank,
Accession No. AF325840.
Bird, R.G. & McCaul, T.F. 1976, 'The rhabdoviruses of Entamoeba histolytica and
Entamoeba invadens', Annal of Tropical Medicine and Parasitology, vol. 70, no.
1, pp. 81-93.
Bird, R.G., McCaul, T.F. & Knight, R. 1974, 'Rhabdo-virus-like particles of Entamoeba
histolytica', Transactions of the Royal Society of Tropical Medicine and
Hygiene, vol. 68, no. 1, pp. 2.
Borody, T.J., Warren, E.F., Wettstein, A., Robertson, A., Recabarren, P., Fontella, A.,
Herdman, K. & Surace, R. 2002, 'Eradication of Dientamoeba fragilis can
resolve IBS-like symptoms' Jounal of Gastroenterology Hepatology, vol. 17.
Boyer, M., Azza, S., Barrassi, L., Klose, T., Campocasso, A., Pagnier, I.,
Fournous, G., Borg, A., Robert, C. & Zhang, X., Desnues, C.,
Henrissat,B., Rossmann, M.G., La Scola, B. & Raoult, D.
2011, 'Mimivirus shows dramatic genome reduction after intraamoebal
culture', Proceedings of the National Academy of Sciences of the United
States of America, vol. 108, no. 25, pp. 10296-301.

99
Burrows, R.B. & Swerdlow, M.A. 1956, 'Enterobius vermicularis as a probable vector
of Dientamoeba fragilis', The American Journal of Tropical Medicine and
Hygiene, vol. 5, no. 2, pp. 258-65.
Burrows, R.B. 1967, 'A new fixative and technics for the diagnosis of intestinal
parasites', Technical bulletin of the Registry of Medical Technologists, vol. 37,
pp. 208-12.
Butler, W.P. 1996, 'Dientamoeba fragilis. An unusual intestinal pathogen', Digestive
Diseases and Sciences, vol. 41, no. 9, pp. 1811-13.
Cacciò, S.M., Sannella, A.R., Manuali, E., Tosini, F., Sensi, M., Crotti, D. & Pozio, E.
2012, 'Pigs as natural hosts of Dientamoeba fragilis genotypes found in
Humans', Emerging Infectious Diseases, vol. 18, no. 5, pp. 838-41.
Cadd, T.L., Keenan, M.C. & Patterson, J.L. 1993, 'Detection of Leishmania RNA virus
1 proteins', Journal of Virology, vol. 67, no. 9, pp. 5647-50.
Cadd, T.L. & Patterson, J.L. 1994, 'Synthesis of viruslike particles by expression of the
putative capsid protein of Leishmania RNA virus in a recombinant baculovirus
expression system', Journal of Virology, vol. 68, no. 1, pp. 358-65.
Camp, R.R., Mattern, C.F. T & Honigberg, B.M. 1974, 'Study of Dientamoeba fragilis
Jepps & Dobell. I. Electronmicroscopic observations of the binucleate stages. II.
Taxonomic position and revision of the genus', The Journal of Protozoology,
vol. 21, no. 1, pp. 69-82.
Cao, L., Gong, P., Li, J., Zhang, X., Zou, X., Tuo, W., Liu, Q., Wang, Q., Zhang, G.,
Chen, L., Li, L. & Su, L. 2009, 'Giardia canis: Ultrastructural analysis of G.
canis trophozoites transfected with full length G. canis virus cDNA transcripts',
Experimental Parasitology, vol. 123, no. 3, pp. 212-7.
Cepicka, I., Hampl, V. & Kulda, J. 2010, 'Critical taxonomic revision of Parabasalids
with description of one new genus and three new species', Protist, vol. 161, no.
3, pp. 400-33.
Cevahir, N., Kaleli, I. & Kaleli, B. 2002, 'Evaluation of direct microscopic examination,
acridine orange staining and culture methods for studies of Trichomonas
vaginalis in vaginal discharge specimens', Mikrobiyoloji Bülteni, vol. 36, no. 3-
4, pp. 329-35.

100
Champney, W.S., Curtis, S.K. & Samuels, R. 1995, 'Cytopathology and release of an
RNA virus from a strain of Trichomonas vaginalis', International Journal for
Parasitology, vol. 25, no. 12, pp. 1463-71.
Chen, L., Li, J., Zhang, X., Liu, Q., Yin, J., Yao, L., Zhao, Y. & Cao, L. 2007,
'Inhibition of krr1 gene expression in Giardia canis by a virus-mediated
hammerhead ribozyme', Veterinary Parasitology, vol. 143, no. 1, pp. 14-20.
Chen, L.F., Li, J.H., Zhang, X.C., Liu, Q., Zhao, Y.P. & Cao, L.L. 2006, 'Establishment
of in vitro cultivation of Giardia canis trophozoites infected with Giardia canis
virus', Zhongguo ji Sheng Chong Xue Yu ji Sheng Chong Bing Za Zhi, vol. 24,
no. 4, pp. 261-5.
Choi, Y. G. & Randles, J.W. 1997, 'Microgranular cellulose improves dsRNA recovery
from plant nucleic acid extracts', Biotechniques, vol. 23, no. 4, pp. 610-1.
Clark, C. G., Röser, D. & Stensvold, C. R. 2014, 'Transmission of Dientamoeba
fragilis: pinworm or cysts? ', Trends in parasitology, vol.30, no. 3, pp. 136-40.
Claverie, J.M. & Abergel, C. 2009, 'Mimivirus and its virophage', Annual Review of
Genetics, vol. 43, no. 1, pp. 49-66.
Claverie, J.M. & Abergel, C. 2010, 'Mimivirus: the emerging paradox of quasi-
autonomous viruses', Trends in Genetics, vol. 26, no. 10, pp. 431-7.
Claverie, J.M., Grzela, R., Lartigue, A., Bernadac, A., Nitsche, S., Vacelet, J., Ogata, H.
& Abergel, C. 2009, 'Mimivirus and Mimiviridae: giant viruses with an
increasing number of potential hosts, including corals and sponges', Journal of
Invertebrate Pathology, vol. 101, no. 3, pp. 172-80.
Cleveland, L.R. & Collier, J. 1930, 'Various improvements in the cultivation of
Entamoeba histolytica', American Journal of Hygiene, vol. 12, no. 3, pp. 606-13.
Colson, P. & Raoult, D. 2010, 'Gene repertoire of amoeba-associated giant viruses',
Intervirology, vol. 53, no. 5, pp. 330-43.
Colson, P., Gimenez, G., Boyer, M., Fournous, G. & Raoult, D. 2011a, 'The giant
Cafeteria roenbergensis virus that infects a widespread marine phagocytic protist
is a new member of the fourth domain of Life', PLoS ONE, vol. 6, no. 4, pp.
e18935.
Colson, P., Yutin, N., Shabalina, S.A., Robert, C., Fournous, G., La Scola, B., Raoult,
D. & Koonin, E.V. 2011b, 'Viruses with more than 1,000 genes: Mamavirus, a

101
new Acanthamoeba polyphaga mimivirus strain, and reannotation of Mimivirus
genes', Genome Biology and Evolution, vol. 3, pp. 737-42.
Croft , S.L., Chance, M.L. & Gardener, P.J. 1980, 'Ultrastructural and biochemical
characterization of stocks of Endotrypanum', Annals of Tropical Medicine and
Parasitology, vol. 74, no. 6, pp. 585-9.
Croft, S.L. & Molyneux, D.H. 1979, 'Studies on the ultrastructure, virus-like particles
and infectivity of Leishmania hertigi', Annals of Tropical Medicine and
Parasitology, vol. 73, no. 3, pp. 213-26.
Crotti, D. & D'Annibale, M.L. 2007, 'Intestinal infections caused by Dientamoeba
fragilis and Giardia duodenalis in our experience', Recenti Progressi in
Medicina, vol. 98, no. 6, pp. 361-6.
Crotti, D., D'Annibale, M.L., Fonzo, G., Lalle, M., Caccio, S.M. & Pozio, E. 2005,
'Dientamoeba fragilis is more prevalent than Giardia duodenalis in children and
adults attending a day care centre in Central Italy', Parasite, vol. 12, no. 2, pp.
165-70.
Cuffari, C., Oligny, L. & Seidman, E.G. 1998, 'Dientamoeba fragilis masquerading as
allergic colitis', Journal of Pediatric Gastroenterology and Nutrition, vol. 26,
no. 1, pp. 16-20.
Dalton, A.J. & Haguenau, F. 1973, ‘Ultrastructure of animal viruses and
bacteriophages’, An Atlas, vol. 5.
Dan, M., Wang, A.L. & Wang, C.C. 2000, 'Inhibition of pyruvate-ferredoxin
oxidoreductase gene expression in Giardia lamblia by a virus-mediated
hammerhead ribozyme', Molecular Microbiology, vol. 36, no. 2, pp. 447-56.
Dan, M. & Wang, C.C. 2000, 'Role of alcohol dehydrogenase E (ADHE) in the energy
metabolism of Giardia lamblia', Molecular and Biochemical Parasitology, vol.
109, no. 1, pp. 25-36.
De Jonckheere, J.F. & Gordts, B. 1987, 'Occurrence and transfection of a Giardia virus',
Molecular and Biochemical Parasitology, vol. 23, no. 1, pp. 85-9.
del Cacho, E., Gallego, M., Montes, C., Lopez-Bernad, F., Quilez, J. & Sanchez-Acedo,
C. 2001, 'Eimeria necatrix virus: intracellular localisation of viral particles and
proteins', International Journal for Parasitology, vol. 31, no. 11, pp. 1269-74.
Delgado-Viscogliosi, P., Viscogliosi, E., Gerbod, D., Kulda, J., Sogin, M.L. &
Edgcomb, V.P. 2000, 'Molecular phylogeny of parabasalids based on small

102
subunit rRNA sequences, with emphasis on the Trichomonadinae subfamily',
The Journal of Eukaryotic Microbiology., vol. 47, no. 1, pp. 70-5.
Diamond, L.S., Mattern, C.F.T. & Bartgis, I.L. 1972, 'Viruses of Entamoeba histolytica.
I. Identification of transmissible virus-like agents', Journal of Virology, vol. 9,
no. 2, pp. 326-41.
Diamond, M.E., Dowhanick, J.J., Nemeroff, M.E., Pietras, D.F., Tu, C.L. & Bruenn,
J.A. 1989, 'Overlapping genes in a yeast double-stranded RNA virus', Journal of
Virology, vol. 63, no. 9, pp. 3983-90.
Dickinson, E.C., Cohen, M.A. & Schlenker, M.K. 2002, 'Dientamoeba fragilis: a
significant pathogen', The American Journal of Emergency Medicine, vol. 20,
no. 1, pp. 62-3.
Dobell, C. 1940, 'Researches on the intestinal protozoa of monkeys and man. X. The life
history of Dientamoeba fragilis: observations, experiments and speculations',
Parasitology, vol. 32, pp. 417-461.
Dobos, P., Hill, B. J., Hallett, R., Kells, D. T., Becht, H. & Teninges, D. 1979,
'Biophysical and biochemical characterization of five animal viruses with
bisegmented double-stranded RNA genomes', Journal of Virology, vol. 32, no.
2, pp. 593-605.
Dutta, G.P. 1970, 'Cytochemical significance of acridine orange staining of human
plasmodia with some comments on double-stranded DNA', Histochemistry and
Cell Biology, vol. 24, no. 1, pp. 29-32.
Ellis, J. & Revets, H. 1990, 'Eimeria species which infect the chicken contain virus-like
RNA molecules', Parasitology, vol. 101, no. 2, pp. 163-9.
Enquist, L.W. 2009, 'Virology in the 21st Century', Journal of Virology, vol. 83, no. 11,
pp. 5296-308.
Fire, A., Xu, S., Montgomery, M.K., Kostas, S.A., Driver, S.E. & Mello, C.C. 1998,
'Potent and specific genetic interference by double-stranded RNA in
Caenorhabditis elegans', Nature, vol. 391, no. 6669, pp. 806-11.
Flegr, J. 1987, 'A rapid method for isolation of double stranded RNA', Preparative
Biochemistry, vol. 17, no. 4, pp. 423-33.
Flegr, J., Cerkasov, J., Kulda, J., Tachezy, J. & Stokrova, J. 1987, 'The dsRNA of
Trichomonas vaginalis is associated with virus-like particles and does not

103
correlate with metronidazole resistance ', Folia Microbiologica (Praha), vol. 32,
no. 4, pp. 345-8.
Flegr, J., Cerkasov, J. & Stokrova, J. 1988, 'Multiple populations of double -stranded
RNA in two virus- harbouring strains of Trichomonas vaginalis', Folia
Microbiologica (Praha), vol. 33, no. 6, pp. 462-5.
Fraga, J., Rojas, L., Sariego, I. & Fernandez-Calienes, A. 2005, 'Double - stranded RNA
viral infection in Cuban Trichomonas vaginalis isolates', Brazilian Journal of
Infectious Diseases, vol. 9, no. 6, pp. 521-4.
Fraga,J., Rojas, L., Sariego, I. & Fernandez-Calienes, A. 2012, 'Genetic characterization
of three Cuban Trichomonas vaginalis virus. Phylogeny of Totiviridae family',
Infection, Genetics and Evolution, vol. 12, no. 1, pp. 113-20.
Fraga, J., Rojas, L., Sariego, I. & Fernández-Calienes, A. 2011, 'Double-stranded RNA
viral infection of Trichomonas vaginalis and correlation with genetic
polymorphism of isolates', Experimental Parasitology, vol. 127, no. 2, pp. 593-
9.
Fraga, J., Rojas, L., Sarieg,o I., Fernandez-Calienes, A. & FA, N. 2007, 'Double-
stranded RNA viral infection of Trichomonas vaginalis and association with
clinical presentation', Acta Protozoologica, vol. 46, pp. 93-8.
Fujimura, T. & Wickner, R.B. 1988, 'Replicase of L-A virus-like particles of
Saccharomyces cerevisiae. In vitro conversion of exogenous L-A and M1
single-stranded RNAs to double-stranded form', The Journal of Biological
Chemistry, vol. 263, no. 1, pp. 454-60.
Furfine, E.S., White, T.C., Wang, A.L. & Wang, C.C. 1989, 'A single-stranded RNA
copy of the Giardia lamblia virus double-stranded RNA genome is present in
the infected Giardia lamblia', Nucleic Acids Research, vol. 17, no. 18, pp. 7453-
67.
Garnham, P.C., Bird, R.G. & Baker, J.R. 1962, 'Electron microscope studies of motile
stages of malaria parasites: III. The ookinetes of Haemamoeba and
Plasmodium', Transactions of the Royal Society of Tropical Medicine and
Hygiene, vol. 56, no. 2, pp. 116-20.
Garcia, L.S. 2002, 'Laboratory methods for diagnosis of parasitic infection,' In B. A.
Forbes, D. F. Sahm, and A. S. Weissfeld (ed.), Bailey and Scott’s Diagnostic
Microbiology, 11th ed. C. V. Mosby, St. Louis, Mo., pp. 776-861.

104
Gerbod, D., Edgcomb, V.P., Noel, C., Zenner, L., Wintjens, R., Delgado-Viscogliosi,
P., Holder, M.E., Sogin, M.L. & Viscogliosi, E. 2001, 'Phylogenetic position of
the trichomonad parasite of turkeys, Histomonas meleagridis (Smith) Tyzzer,
inferred from small subunit rRNA sequence', The Journal of Eukaryotic
Microbiology, vol. 48, no. 4, pp. 498-504.
Gerbod, D., Noël, C., Dolan, M.F., Edgcomb, V.P., Kitade, O., Noda, S., Dufernez, F.,
Ohkuma, M., Kudo, T., Capron, M., Sogin, M.L. & Viscogliosi, E. 2002,
'Molecular phylogeny of parabasalids inferred from small subunit rRNA
sequences, with emphasis on the Devescovinidae and Calonymphidae
(Trichomonadea)', Molecular Phylogenetics and Evolution, vol. 25, no. 3, pp.
545-56.
Gerbod, D., Sanders, E., Moriya, S., Noël, C., Takasu, H., Fast, N.M., Delgado-
Viscogliosi, P., Ohkuma, M., Kudo, T., Capron, M., Palmer, J.D., Keeling, P.J.
& Viscogliosi, E. 2004, 'Molecular phylogenies of Parabasalia inferred from
four protein genes and comparison with rRNA trees', Molecular Phylogenetics
and Evolution, vol. 31, no. 2, pp. 572-80.
Gerhold, R.W., Allison, A.B., Sellers, H., Linnemann, E., Chang, T.H. & Alderete, J.F.
2009, 'Examination for double-stranded RNA viruses in Trichomonas gallinae
and identification of a novel sequence of a Trichomonas vaginalis virus',
Parasitology Research, vol. 105, no. 3, pp. 775-9.
Ghabrial, S.A. 2008, ‘Totiviruses, In Mahy B. W. J., Van Regenmortel M. H. V.,
editors. (ed.), Encyclopedia of Virology (Elsevier, Oxford, United Kingdom),
vol. 5, no. 3, pp. 163-74.
Girginkardesler, N., Coskun, S., Cuneyt Balcioglu, I., Ertan, P. & Ok, U.Z. 2003,
'Dientamoeba fragilis, a neglected cause of diarrhea, successfully treated with
secnidazole', Clinical Microbiology and Infection, vol. 9, no. 2, pp. 110-3.
Girginkardesler, N., Kurt, Ö., Kilimcioglu, A.A. & Ok, Ü.Z. 2008, 'Transmission of
Dientamoeba fragilis: evaluation of the role of Enterobius vermicularis',
Parasitology International, vol. 57, no. 1, pp. 72-75.
Gomes Vancini, R. & Benchimol, M. 2005, 'Appearance of virus-like particles in
Tritrichomonas foetus after drug treatment', Tissue and Cell, vol. 37, no. 4, pp.
317-23.

105
González-Lázaro, M., González-Robles, A., Hernández-Gutiérrez, R. & Arroyo, R.
2005, 'HeLa cell nucleus, a source of thymidine for Trichomonas vaginalis
growing in vitro', The International Journal of Biochemistry & Cell Biology,
vol. 37, no. 1, pp. 166-76.
Goldman, M. & Brooke, M.M. 1953, 'Protozoans in stools unpreserved and preserved in
PVA fixative.', Public Health Reports, vol. 68, no. 7, pp. 703-6.
Goodman, R. P., Ghabrial, S. A, Fichorova, R. N. & Nibert, M. L. 2011a,
'Trichomonasvirus: a new genus of protozoan viruses in the family Totiviridae',
Archives of Virology, vol. 156, no. 1, pp. 171-9.
Goodman, R.P., Freret, T.S., Kula, T., Geller, A.M., Talkington, M.W., Tang-
Fernandez, V., Suciu, O., Demidenko, A.A., Ghabrial, S.A., Beach, D.H., Singh,
B.N., Fichorova, R.N. & Nibert, M.L. 2011b, 'Clinical isolates of Trichomonas
vaginalis concurrently infected by strains of up to four Trichomonasvirus
species (Family Totiviridae)', Journal of Virology, vol. 85, no. 9, pp. 4258-70.
Graybill, H.W. & Smith, T. 1920, 'Production of fatal blackhead in turkeys by feeding
embryonated eggs of Heterakis Papillosa.’ The Journal of Experimental
Medicine, vol. 31, no. 5, pp. 647-55.
Greenway, D. 1928, 'Dientamoeba fragilis en la Argentina. Arch. Argent. Enferm. ',
Aparato Dig. (Buenos Aires) vol. 3, pp. 897.
Guilbride, L., Myler, P.J. & Stuart, K. 1992, 'Distribution and sequence divergence of
LRV1 viruses among different Leishmania species', Molecular and Biochemical
Parasitology, vol. 54, no. 1, pp. 101-4.
Han, Q., Li, J., Gong, P., Gai, J., Li, S. & Zhang, X. 2011, 'Virus-like particles in
Eimeria tenella are associated with multiple RNA segments', Experimental
Parasitology, vol. 127, no. 3, pp. 646-50.
Hansen, D.W., Hunter, D.T., Richards, D.F. & Allred, L. 1970, 'Acridine orange in the
staining of blood parasites', Journal of Parasitology vol. 56, no. 2, pp. 386-87.
Heidary, S., Bandehpour, M., Valadkhani, Z., Seyyed–Tabaee, S. J., Haghighi, A.,
Abadi, A. R., & Kazemi, B., 2013, ‘Double-Stranded RNA viral infection
In Tehran Trichomonas vaginalis isolates’, Iranian Journal of Parasitology,
vol. 8, no. 1, pp. 60-64.

106
Hoffmann, E., Neumann, G., Kawaoka, Y., Hobom, G. & Webster, R.G. 2000, 'A DNA
transfection system for generation of influenza A virus from eight plasmids',
Proceedings of the National Academy of Sciences, vol. 97, no. 11, pp. 6108-13.
Hopkins, J. 2006, ' Microbiology newsletter ', vol. 25, no. 18.
Hruska, J.F., Mattern, C.F., Diamond, L.S. & Keister, D.B. 1973, 'Viruses of
Entamoeba histolytica. 3. Properties of the polyhedral virus of the HB-301
Strain', Journal of Virology, vol. 11, no. 1, pp. 129-36.
Hruska, J.F., Mattern, C.F. T & Diamond, L.S. 1974, 'Viruses of Entamoeba histolytica
IV. Studies on the nucleic acids of the filamentous and polyhedral viruses',
Journal of Virology, vol. 13, no. 1, pp. 205-210.
Huang, S. & Ghabrial, S.A. 1996, 'Organization and expression of the double-stranded
RNA genome of Helminthosporium victoriae 190S virus, a totivirus infecting a
plant pathogenic filamentous fungus', Proceedings of the National Academy of
Sciences of the United States of America, vol. 93, no. 22, pp. 12541-6.
Hyde, J.L., Gillespie, L.K. & Mackenzie, J.M. 2012, 'Mouse norovirus 1 utilizes the
cytoskeleton network to establish localization of the replication complex
proximal to the microtubule organizing center', Journal of Virology, vol. 86, no.
8, pp. 4110-22.
Icho, T. & Wickner, R.B. 1989, 'The double-stranded RNA genome of yeast virus L-A
encodes its own putative RNA polymerase by fusing two open reading frames',
Journal of Biological Chemistry, vol. 264, no. 12, pp. 6716-23.
Jacob, L. 1953, 'The cultivation of Dientamoeba fragilis', Annals of the
New York Academy of Sciences, vol. 56, no. 5, pp. 1057-61.
Jacks, T., Madhani, H.D., Masiarz, F.R. & Varmus, H.E. 1988a, 'Signals for ribosomal
frameshifting in the Rous sarcoma virus gag-pol region.', Cell, vol. 55, no. 3, pp.
447-58.
Jacks, T., Power, M.D., Masiarz, F.R., Luciw, P.A., Barr, P.J. & Varmus, H.E. 1988b,
'Characterization of ribosomal frameshifting in HIV-1 gag-pol expression',
Nature, vol. 331, no. 6153, pp. 280-3.
Jepps, M.W. & Dobell, C. 1918, 'Dientamoeba fragilis n.g., n. sp., a new intestinal
amoeba from man.', Parasitology, vol. 10, no. 3, pp. 352-67.

107
Johnston, R.C., Farias, N.A., Gonzales, J.C., Dewes, H., Masuda, A., Termignoni, C.,
Amako, K. & Ozaki, L.S. 1991, 'A putative RNA virus in Babesia bovis',
Molecular and Biochemical Parasitology, vol. 45, no. 1, pp. 155-8.
Johnson, J.A. & Clark, C.G. 2000, 'Cryptic genetic diversity in Dientamoeba fragilis',
Journal of Clinical Microbiology, vol. 38, no. 12, pp. 4653-54.
Johnson, E.H., Windsor, J.J. & Clark, C.G. 2004, 'Emerging from obscurity: biological,
clinical, and diagnostic aspects of Dientamoeba fragilis', Clinical Microbiology
Reviews, vol. 17, no. 3, pp. 553-70.
Kang, J, Wu, J., Bruenn, J.A. & Park, C. 2001, 'The H1 double-stranded RNA genome
of Ustilago maydis virus-H1 encodes a polyprotein that contains structural
motifs for capsid polypeptide, papain-like protease, and RNA-dependent RNA
polymerase', Virus Research, vol. 76, no. 2, pp. 183-9.
Kasprzak, W. & Majewska, A.C. 1995, '[Viruses of parasitic protozoa]', Wiadomości
Parazytologiczne, vol. 41, no. 2, pp. 131-7.
Kean, B.H. & Malloch, C.L. 1966, 'The neglected ameba: dientamoeba fragilis. A
report of 100 "pure" infections', The American Journal of Digestive Diseases,
vol. 11, no. 9, pp. 735-46.
Khoshnan, A. & Alderete, J.F. 1993, 'Multiple double-stranded RNA segments are
associated with virus particles infecting Trichomonas vaginalis', Journal of
Virology, vol. 67, no. 12, pp. 6950-5.
Khoshnan, A. & Alderete, J.F. 1994, 'Trichomonas vaginalis with a double-stranded
RNA virus has upregulated levels of phenotypically variable immunogen
mRNA', Journal of Virology, vol. 68, no. 6, pp. 4035-8.
Khoshnan, A. & Alderete, J.F. 1995, 'Characterization of double-stranded RNA
satellites associated with the Trichomonas vaginalis virus', Journal of Virology,
vol. 69, no. 11, pp. 6892-7.
Khramtsov, N.V., Chung, P.A., Dykstra, C.C., Griffiths, J.K., Morgan, U.M.,
Arrowood, M.J. & Upton, S.J. 2000, 'Presence of double-stranded RNAs in
human and calf isolates of Cryptosporidium parvum', Journal of Parasitology,
vol. 86, no. 2, pp. 275-82.
Khramtsov, N.V. & Upton, S.J. 1998, 'High-temperature inducible cell-free
transcription and replication of double-stranded RNAs within the parasitic
protozoan Cryptosporidium parvum', Virology, vol. 245, no. 2, pp. 331-7.

108
Khramtsov, N.V. & Upton, S.J. 2000, 'Association of RNA polymerase complexes of
the parasitic protozoan Cryptosporidium parvum with virus-like particles:
heterogeneous system', Journal of Virology, vol. 74, no. 13, pp. 5788-95.
Khramtsov, N.V., Woods, K.M., Nesterenko, M.V., Dykstra, C.C. & Upton, S.J. 1997,
'Virus-like, double-stranded RNAs in the parasitic protozoan Cryptosporidium
parvum', Molecular Microbiology, vol. 26, no. 2, pp. 289-300.
Kim, J.W., Chung, P.-R., Hwang, M.-K. & Choi, E.Y. 2007, 'Double-stranded RNA
virus in Korean Isolate IH-2 of Trichomonas vaginalis', The Korean Journal of
Parasitology, vol. 45, no. 2, pp. 87-94.
Kleina, P., Bettim-Bandinelli, J., Bonatto, S.L., Benchimol, M. & Bogo, M.R. 2004,
'Molecular phylogeny of Trichomonadidae family inferred from ITS-1, 5.8S
rRNA and ITS-2 sequences', International Journal for Parasitology, vol. 34, no.
8, pp. 963-70.
Knoll, E.W. & Howell, K. M. 1946, 'Studies on Dientamoeba fragilis. Its incidence and
possible pathogenicity', Medical Woman’s Journal, vol. 53, no. 11, pp. 40-6.
Kniel, K.E., Higgins, J.A., Trout, J.M., Fayer, R. & Jenkins, M.C. 2004,
'Characterization and potential use of a Cryptosporidium parvum virus (CPV)
antigen for detecting C. parvum oocysts', Journal of Microbiological Methods,
vol. 58, no. 2, pp. 189-95.
Köhler, S. & Wang, C.C. 1997, 'Site-specific binding of polymerase-containing
particles of the Giardia lamblia double-stranded RNA virus to the viral plus-
strand RNA'', Research in Virology, vol. 148, no. 5, pp. 311-21.
Kurt, O., Girginkardesler, N., Balcioglu, I.C., Ozbilgin, A. & Ok, U.Z. 2008, 'A
comparison of metronidazole and single-dose ornidazole for the treatment of
dientamoebiasis', Clinical Microbiology and Infection, vol. 14, no. 6, pp. 601-4.
Kurth, E.G., Peremyslov, V.V., Prokhnevsky, A.I., Kasschau, K.D., Miller, M.,
Carrington, J.C. & Dolja, V.V. 2012, 'Virus-derived gene expression and RNA
interference vector for grapevine', Journal of Virology, vol. 86, no. 11, pp. 6002-
9.
Lainson, R. & Da Silva, B.A. 1999, 'Intestinal parasites of some diarrhoeic HIV-
seropositive individuals in North Brazil with particular reference to Isospora
belli Wenyon, 1923 and Dientamoeba fragilis Jepps & Dobell, 1918.', Memórias
do Instituto Oswaldo Cruz, vol. 94, no. 5, pp. 611-13.

109
Lagacé-Wiens, P.R., VanCaeseele, P.G. & Koschik, C. 2006, 'Dientamoeba fragilis: an
emerging role in intestinal disease', Canadian Medical Association Journal, vol.
175, no. 5, pp. 468-69.
La Scola, B., Desnues, C., Pagnier, I., Robert, C., Barrassi, L., Fournous, G., Merchat,
M., Suzan-Monti, M., Forterre, P., Koonin, E. & Raoult, D. 2008, 'The
virophage as a unique parasite of the giant mimivirus', Nature, vol. 455, no.
7209, pp. 100-4.
Lauretti, F., Lucas de Melo, F., Benati, F.J., de Mello Volotão, E., Santos, N., Linhares,
R.E. & Nozawa, C. 2003, 'Use of acridine orange staining for the detection of
rotavirus RNA in polyacrylamide gels', Journal of Virological Methods, vol.
114, no. 1, pp. 29-35.
Lee, S. & Fernando, M.A. 1999a, 'RNA-dependent RNA polymerase activity associated
with Eimeria necatrix virus particles containing either double-stranded or single-
stranded RNA. ', The Journal of Parasitology, vol. 85, no. 2, pp. 208-12.
Lee, S. & Fernando, M.A. 1999b, 'RNA-dependent RNA polymerase activity associated
with virus-like dsRNA in Eimeria maxima and Eimeria necatrix of the domestic
fowl', Parasitology Research, vol. 85, no. 1, pp. 25-9.
Lee, S., Fernando, M.A. & Nagy, E. 1996, 'dsRNA associated with virus-like particles
in Eimeria spp. of the domestic fowl', Parasitology Research, vol. 82, no. 6, pp.
518-23.
Lee, S.E., Suh, J.M., Scheffter, S., Patterson, J.L. & Chung, I.K. 1996, 'Identification of
a ribosomal frameshift in Leishmania RNA Virus 1–4', Journal of Biochemistry,
vol. 120, no. 1, pp. 22-5.
Legendre, M., Arslan, D., Abergel, C. & Claverie, J.M. 2012, 'Genomics of Megavirus
and the elusive fourth domain of life', Communicative & Integrative Biology,
vol. 5, no. 1, pp. 102-6.
Lemke, P.A. 1976, 'Viruses of eukaryotic microorganisms', Annual Review of
Microbiology, vol. 30, pp. 105-45.
Lenstra, R., Samso, A., Andrieu, B., Le Bras, J. & Galibert, F. 1988, 'Virus like particles
containing knob-associated histidine-rich protein are secreted into the culture
medium of Plasmodium falciparum in vitro cultures', Biochemical and
Biophysical Research Communications, vol. 151, no. 2, pp. 749-57.

110
Li, W., Ding, H., Zhang, X., Cao, L., Li, J., Gong, P., Li, H., Zhang, G., Li, S. & Zhang,
X. 2012, 'The viral RNA-based transfection of enhanced green fluorescent
protein (EGFP) in the parasitic protozoan Trichomonas vaginalis', Parasitology
Research, vol. 110, no. 3, pp. 1305-10.
Life, R. B., Lee, E. G., Eastman, S.W. & Linial M. L. 2008, 'Mutations in the amino
terminus of foamy virus gag disrupt morphology and infectivity but do not target
assembly', Journal of Virology, vol. 82, no. 13, pp. 6109-6119.
Liu, C., Li, J., Zhang, X., Liu, Q., Liu, H., Gong, P., Zhang, G., Yao, L. & Zhang, X.
2008, 'Stable expression of green fluorescent protein mediated by GCV in
Giardia canis', Parasitology International, vol. 57, no. 3, pp. 320-4.
Liu, H.W., Chu, Y.D. & Tai, J.H. 1998, 'Characterization of Trichomonas vaginalis
virus proteins in the pathogenic protozoan T. vaginalis', Archives of Virology,
vol. 143, no. 5, pp. 963-70.
Liu, Q., Zhang, X., Li, J., Ying, J., Chen, L., Zhao, Y., Wei, F. & Wu, T. 2005, 'Giardia
lamblia: stable expression of green fluorescent protein mediated by giardiavirus',
Experimental Parasitology, vol. 109, no. 3, pp. 181-7.
MacBeth, K.J. 1996, 'Identification and characterization of an endoribonuclease activity
of Leishmaniavirus capsids', Ph.D. thesis. Harvard University Cambridge,
Mass.
MacBeth, K.J. & Patterson, J.L. 1995, 'The short transcript of Leishmania RNA virus is
generated by RNA cleavage', Journal of Virology, vol. 69, no. 6, pp. 3458-64.
Malla, N., Kaul, P., Sehgal, R. & Gupta, I. 2011, 'The presence of dsRNA virus in
Trichomonas vaginalis isolates from symptomatic and asymptomatic Indian
women and its correlation with in vitro metronidazole sensitivity', Indian
Journal of Medical Microbiology, vol. 29, no. 2, pp. 152-7.
Martin, R., Tzanetakis, I.E., Sweeney, M., Wegnener, L.A. 2006, 'A virus associated
with blueberry fruit drop disease', Acta Horticulturae, vol 715, pp. 497-501.
Mattern, C.F., Keister, D.B., Daniel, W.A., Diamond, L.S. & Kontonis, A.D. 1977,
'Viruses of Entamoeba histolytica. VII. Novel beaded virus', Journal of
Virology, vol. 23, no. 3, pp. 685-91.
Mattern, C.F.T., Diamond, L.S. & Daniel, W. A. 1972, 'Viruses of Entamoeba
histolytica II. Morphogenesis of the polyhedral particle (ABRM2→HK-9)→HB-

111
301 and the filamentous agent (ABRM)2→HK-9', Journal of Virology, vol. 9,
no. 2, pp. 342-58.
Mattern, C.F.T., Hruska, J.F. & Diamond, L.S. 1974, 'Viruses of Entamoeba histolytica.
V. Ultrastructure of the polyhedral virus V301.', Journal of Viroogy, vol. 13, no.
1, pp. 247-49.
Mattern, C.F., Keister, D.B. & Diamond, L.S. 1979, 'Experimental amebiasis.
IV. Amebal viruses and the virulence of Entamoeba histolytica’,
The American Journal of Tropical Medicine and Hygiene, vol. 28, no. 4, pp.
653-7.
Mayer, A. 1886, 'Die Landwirtschaftliche Versuchs-stationen ', Über die
Mosaikkrankheit des Tabaks (in German), vol. 32, pp. 451-67.
Miller, J.H. & Swartzwelder, J.C. 1960, 'Virus-like particles in an Entamoeba
histolytica trophozoite', Journal of Parasitology, vol. 46, no. 4, pp. 523-4.
Miller, R.L., Nelson, D.J., LaFon, S.W., Miller, W.H. & Krenitsky, T.A. 1987,
'Antigiardial activity of guanine arabinoside. Mechanism studies', Biochemical
Pharmacology, vol. 36, no. 15, pp. 2519-25.
Miller, R.L., Wang, A.L. & Wang, C.C. 1988a, 'Identification of Giardia lamblia
isolates susceptible and resistant to infection by the double-stranded RNA virus',
Experimental Parasitology, vol. 66, no. 1, pp. 118-23.
Miller, R.L., Wang, A.L. & Wang, C.C. 1988b, 'Purification and characterization of the
Giardia lamblia double-stranded RNA virus', Molecular and Biochemical
Parasitology, vol. 28, no. 3, pp. 189-95.
Molyneux, D.H. 1974, 'Virus-like particles in Leishmania parasites. ', Nature, vol. 249,
pp. 588-9.
Molyneux, D.H. & Heywood, P. 1984, 'Evidence for the incorporation of virus-like
particles into Trypanosoma', Parasitology Research, vol. 70, no. 4, pp. 553-6.
Müller, M. & Gorrell, T.E. 1983, ‘Metabolism and metronidazole uptake in
Trichomonas vaginalis isolates with different metronidazole susceptibilities’.
Antimicrobial Agents and Chemotherapy, vol. 24, no. 5, pp. 667-73.
Murphy, F.A. & Kingsbury, D.W. 1990, 'Virus taxonomy ', In fields virology, 2nd. Edn.
(BN Fields et al. Eds.) Raven Press, New York, pp 9-35.
Mumcuoğlu, I., Coşkun, F. A., Aksu, N., Pürnak, T. & Güngör, C. 2013. ‘[Role of
Dientamoeba fragilis and Blastocystis spp. in Irritable Bowel Syndrome]’,

112
Turkiye parazitolojii dergisi / Turkiye Parazitoloji Dernegi =
Acta parasitologica Turcica / Turkish Society for Parasitology, vol. 37, no. 2, pp.73-77.
Munasinghe, V. S., Stark, D. & Ellis, J. T. 2012. ‘New advances in the in-vitro
culture of Dientamoeba fragilis’, Parasitology, vol.139, no. 7, pp. 864-69.
Munasinghe, V. S., Vella, N. G. F., Ellis, J. T., Windsor, P. A. & Stark, D. 2013.
‘Cyst formation and faecal–oral transmission of Dientamoeba fragilis –
the missing link in the life cycle of an emerging pathogen’, International Journal for Parasitology, vol. 43, no. 11, pp. 879-83.
Myers, B.J. & Kuntz, R.E. 1968, 'Intestinal protozoa of the baboon Papio doguera
Pucheran, 1856', The Journal of Protozoology, vol. 15, no. 2, pp. 363-5.
Nagata, N., Marriott, D., Harkness, J., Ellis, J.T. & Stark, D. 2012a, 'In vitro
susceptibility testing of Dientamoeba fragilis', Antimicrobial Agents and
Chemotherapy, vol. 56, no. 1, pp. 487-94.
Nagata, N., Marriott, D., Harkness, J., Ellis, J. T. & Stark, D. 2012b, ‘Current treatment
options for Dientamoeba fragilis infections’, International Journal
for Parasitology: Drugs and Drug Resistance, vol. 2, pp. 204-15.
Nash, T.E., Herrington, D.A., Losonsky, G.A. & Levine, M.M. 1987, 'Experimental
human infections with Giardia lamblia', Journal of Infectious Diseases, vol.
156, no. 6, pp. 974-84.
Netherton, C., Moffat, K., Brooks, E. & Wileman, T. 2007, 'A guide to viral inclusions,
membrane rearrangements, factories, and viroplasm produced during virus
replication', Advances in Virus Research, vol. 70, pp. 101-82.
Nibert, M.L., Woods, K.M., Upton, S.J. & Ghabrial, S.A. 2009, 'Cryspovirus: a new
genus of protozoan viruses in the family Partitiviridae', Archives of Virology, vol
154, no. 12, pp. 1959-65.
Noble, G.A. & Noble, E.R. 1952, 'Entamoeba in farm animals', Journal of Parasitology,
vol. 38, no. 6, pp. 571-95.
Norberg, A., Nord, C.E. & Evengard, B. 2003, 'Dientamoeba fragilis--a protozoal
infection which may cause severe bowel distress', Clinical Microbiology and
Infection, vol. 9, no. 1, pp. 65-68.

113
Ockert, G. 1990, 'Symptomatology, pathology, epidemiology, and diagnosis of
Dientamoeba fragilis', In B. M. Honigberg (ed.), Trichomonads Parasitic in
Humans. Springer Publications, New York, N.Y., pp. 394-410.
Ockert, G. 1975, '[Epidemiology of Dientamoeba fragilis Jepps and Dobell, 1918. 3.
Further studies on Enterobius transmission through eggs]', Journal of Hygiene,
Epidemiology, Microbiology, and Immunology, vol. 19, no. 1, pp. 17-21.
Ögren, J., Dienus, O., Löfgren, S., Iveroth, P., & Matussek A., 2013, ‘Dientamoeba
Fragilis DNA detection in Enterobius vermicularis eggs’, Pathogens
and Disease, vol.69, no.2 pp. 157-8.
Ohkuma, M., Iida, T., Ohtoko, K., Yuzawa, H., Noda, S., Viscogliosi, E. & Kudo, T.
2005, 'Molecular phylogeny of parabasalids inferred from small subunit rRNA
sequences, with emphasis on the Hypermastigea', Molecular Phylogenetics and
Evolution, vol. 35, no. 3, pp. 646-55.
Olivier, L.R., Lecatsas, M., Jackson, T.F.H. G., Gathiram, V., Simjee, A.E. & Prozesky,
O.W. 1984, 'Viruses and pathogenicity of Entamoeba histolytica', The Lancet,
vol. 324, no. 8401, pp. 528.
Park, C.M., Lopinski, J.D., Masuda, J., Tzeng, T.H. & Bruenn, J.A. 1996, 'A second
double-stranded RNA virus from yeast', Virology, vol. 216, no. 2, pp. 451-4.
Parent, K. N., Takagi, Y., Cardone, G., Olson, N. H., Ericsson, M., Yang, M., Lee, Y.,
Asara, J. M., Fichorova, R. N., Baker, T. S. & Nibert, M. L. 2013, ‘Structure of
a protozoan virus from the human genitourinary parasite Trichomonas
vaginalis’, mBio, vol.4, no. 2, pp. 1-11.
Patterson, J.L. 1990, 'Viruses of protozoan parasites', Experimental Parasitology, vol.
70, no 1. pp. 111-13.
Patterson, J.L. 1993, 'The current status of Leishmania RNA virus I', Parasitology
Today, vol. 9, no. 4, pp. 135-6.
Patterson, J.L., Ghabrial, S.A., Bruenn, J.A., Buck, K.W., Wickner, R.B., Stuart, K.D.,
Wang, A.L. & Wang, C. C. 1995, 'Virus taxonomy: sixth report of the
International committee on taxonomy of viruses', Arcives of Virology, (Suppl.
10), pp. 245-51.
Patton, J.T. & Spencer, E. 2000, 'Genome replication and packaging of segmented
double-stranded RNA viruses', Virology, vol. 277, no. 2, pp. 217-25.

114
Peek, R., Reedeker, F.R. & Van Gool, T. 2004, 'Direct amplification and genotyping of
Dientamoeba fragilis from human stool specimens', Journal of Clinical
Microbiology, vol. 42, no. 2, pp. 631-5.
Pindak, F.F., Pindak, M.M. de, Hyde, B.M. & Gardner Jr., W.A. 1989, 'Acquisition and
retention of viruses by Trichomonas vaginalis.', Genitourinary Medicine, vol.
65, no. 6, pp. 366-71.
Poch,O., Sauvaget, I., Delarue, M. & Tordo, N. 1989, 'Identification of four conserved
motifs among the RNA-dependent polymerase encoding elements', The EMBO
Journal, vol. 8, no. 12, pp. 3867-74.
Pote, L.M., Ainsworth, A.J., Brown, J.E. & Haney, J.A. 1991, 'Characterization of
proteins in sporulated and unsporulated Eimeria maxima oocysts ', American
Journal of Veterinary Research, vol. 52, no. 1, pp. 72-4.
Preiss, U., Ockert, G., Broemme, S. & Otto, A. 1991, 'On the clinical importance of
Dientamoeba fragilis infections in childhood', Journal of Hygiene,
Epidemiology, Microbiology, and Immunology, vol. 35, no. 1, pp. 27-34.
Preiss, U., Ockert, G., Brömme, S. & Otto, A. 1990, 'Dientamoeba fragilis infection, a
cause of gastrointestinal symptoms in childhood', Klinische Pädiatrie, vol. 202,
no. 2, pp. 120-3.
Provenzano, D., Khoshnan, A. & Alderete, J.F. 1997, 'Involvement of dsRNA virus in
the protein composition and growth kinetics of host Trichomonas vaginalis',
Archives of Virology, vol. 142, no. 5, pp. 939-52.
Rayan, H.Z., Ismail, O.A. & El Gayar, E.K. 2007, 'Prevalence and clinical features of
Dientamoeba fragilis infections in patients suspected to have intestinal parasitic
infection', Journal of the Egyptian Society of Parasitology, vol. 37, no. 2, pp.
599-608.
Revets, H., Dekegel, D., Deleersnijder, W., De Jonckheere, J., Peeters, J., Leysen, E. &
Hamers, R. 1989, 'Identification of virus-like particles in Eimeria stiedae',
Molecular and Biochemical Parasitology, vol. 36, no. 3, pp. 209-15.
Ro, Y.T. & Patterson, J.L. 2000, 'Identification of the minimal essential RNA sequences
responsible for site-specific targeting of the Leishmania RNA Virus 1-4 capsid
endoribonuclease', Journal of Virology, vol. 74, no. 1, pp. 130-8.

115
Ro, Y.T., Scheffter, S.M. & Patterson, J.L. 1997, 'Specific in vitro cleavage of a
Leishmania virus capsid-RNA-dependent RNA polymerase polyprotein by a
host cysteine-like protease', Journal of Virology, vol. 71, no. 12, pp. 8983-90.
Roditi, I., Wyler, T., Smith, N. & Braun, R. 1994, 'Virus-like particles in Eimeria
nieschulzi are associated with multiple RNA segments', Molecular and
Biochemical Parasitology, vol. 63, no. 2, pp. 275-82.
Röser, D., Nejsum, P., Carlsgart, A.J., Nielsen, H.V. & Stensvold, C.R. 2013, 'DNA of
Dientamoeba fragilis detected within surface-sterilized eggs of Enterobius
vermicularis', Experimental Parasitology, vol. 133, no. 1, pp. 57-61.
Sabanadzovic, S. & Valverde, R. A. 2011, 'Properties and detection of two
cryptoviruses from pepper (Capsicum annuum)', Virus Genes, vol. 43, no. 2, pp.
307-12.
Saiz, M., Llanos-Cuentas, A., Echevarria, J., Roncal, N., Cruz, M., Muniz, M.T., Lucas,
C., Wirth, D.F., Scheffter, S., Magill, A.J. & Patterson, J.L. 1998, 'Short report:
detection of Leishmaniavirus in human biopsy samples of leishmaniasis from
Peru', The American journal of tropical medicine and hygiene, vol. 58, no. 2, pp.
192-4.
Sarafraz, S., Farajnia, S., Jamali, J., Khodabakhsh, F. & Khanipour, F. 2013,
‘Detection of Dientamoeba fragilis among diarrheal patients referred to
Tabriz health care centers by nested PCR’, Tropical Biomedicine, vol.30,
no. 1, pp. 113-8.
Sawangjaroen, N., Luke, R. & Prociv, P. 1993, 'Diagnosis by faecal culture of
Dientamoeba fragilis infections in Australian patients with diarrhoea',
Transactions of the Royal Society of Tropical Medicine and Hygiene, vol. 87,
no. 2, pp. 163-65.
Scholten, T. 1972, 'An improved technique for the recovery of intestinal protozoa.'
Journal of Parasitology, vol. 58, pp. 633-34.
Schure, J.M., de Vries, M., Weel, J.F.,, van Roon, E.N. & Faber, T.E. 2013,
‘Symptoms and treatment of Dientamoeba fragilis infection in children,
a retrospective study’ The Pediatric Infectious Disease Journal, vol. 32,
no. 4, pp. 148-50.

116
Scheffter, S., Widmer, G. & Patterson, J.L. 1994, 'Complete sequence of Leishmania
RNA Virus 1-4 and identification of conserved sequences', Virology, vol. 199,
no. 2, pp. 479-83.
Scheffter, S.M., Ro, Y.T., Chung, I.K. & Patterson, J.L. 1995, 'The complete sequence
of Leishmania RNA Virus LRV2-1, a virus of an Old World parasite strain',
Virology, vol. 212, no. 1, pp. 84-90.
Schuster, F.L. 1969, 'Intranuclear virus-like bodies in the amoeboflagellate Naegleria
gruberi ', The Journal of Eukaryotic Microbiology, vol. 16, no.4, pp 724-27.
Sepp, T., Entzeroth, R., Mertsching, J., Hofschneider, P.H. & Kandolf, R. 1991, 'Novel
ribonucleic acid species in Eimeria nieschulzi are associated with RNA-
dependent RNA polymerase activity', Parasitology Research, vol. 77, no. 7, pp.
581-4.
Sepp, T., Wang, A.L. & Wang, C.C. 1994, 'Giardiavirus-resistant Giardia lamblia lacks
a virus receptor on the cell membrane surface', Journal of Virology, vol. 68, no.
3, pp. 1426-31.
Shirley, M.W. 2000, 'The genome of Eimeria spp., with special reference to Eimeria
tenella—a coccidium from the chicken', International Journal for Parasitology,
vol. 30, no. 4, pp. 485-93.
Silberman, J.D., Clark, C.G. & Sogin, M.L. 1996, 'Dientamoeba fragilis shares a recent
common evolutionary history with the trichomonads', Molecular and
Biochemical Parasitology, vol. 76, no. 1-2, pp. 311-14.
Smith, G.A. & Enquist, L.W. 2002, 'Break ins and break outs: viral interactions with the
cytoskeleton of Mammalian cells', Annual Review of Cell and Developmental
Biology, vol. 18, no. 1, pp. 135-61.
Snipes, L.J., Gamard, P.M., Narcisi, E.M., Beard, C.B., Lehmann, T. & Secor, W.E.
2000, 'Molecular epidemiology of metronidazole resistance in a population of
Trichomonas vaginalis clinical isolates', Journal of Clinical Microbiology, vol.
38, no. 8, pp. 3004-9.
Sobel, J.D., Nagappan, V. & Nyirjesy, P. 1999, 'Metronidazole-resistant vaginal
Trichomoniasis -an emerging problem', The New England Journal of Medicine,
vol. 341, no. 4, pp. 292-3.
Sodeik, B. 2000, 'Mechanisms of viral transport in the cytoplasm', Trends in
Microbiology, vol. 8, no. 10, pp. 465-72.

117
Spencer, M.J., Chapin, M.R. & Garcia, L.S. 1982, 'Dientamoeba fragilis: a
gastrointestinal protozoan infection in adults', The American Journal of
Gastroenterology, vol. 77, no. 8, pp. 565-69.
Stark, D., Beebe, N., Marriott, D., Ellis, J. & Harkness, J. 2005a, 'Detection of
Dientamoeba fragilis in fresh stool specimens using PCR', International Journal
for Parasitology, vol. 35, no. 1, pp. 57-62.
Stark, D., Beebe, N., Marriott, D., Ellis, J. & Harkness, J. 2005b, 'Prospective study of
the prevalence, genotyping, and clinical relevance of Dientamoeba fragilis
infections in an Australian population', Journal of Clinical Microbiology, vol.
43, no. 6, pp. 2718-23.
Stark, D., Phillips, O., Peckett, D., Munro, U., Marriott, D., Harkness, J. & Ellis, J.
2008, 'Gorillas are a host for Dientamoeba fragilis: an update on the life cycle
and host distribution', Veterinary Parasitology, vol. 151, no. 1, pp. 21-26.
Stark, D., Barratt, J., Roberts, T., Marriott, D., Harkness, J. & Ellis, J. 2010a,
'Comparison of microscopy, two xenic culture techniques, conventional and
real-time PCR for the detection of Dientamoeba fragilis in clinical stool
samples', European Journal of Clinical Microbiology & Infectious Diseases,
vol. 29, no. 4, pp. 411-6.
Stark, D., Barratt, J., Roberts, T., Marriott, D., Harkness, J. & Ellis, J. 2010b, 'A review
of the clinical presentation of dientamoebiasis', The American Journal of
Tropical Medicine and Hygiene, vol. 82, no. 4, pp. 614-9.
Stark, D., Al-qassab, S. E., Barratt, J. L. N., Stanley, K., Roberts, T.,
Marriott, D., Harkness, J. & Ellis, J. T. 2011. Evaluation of Multiplex
Tandem Real-Time PCR for Detection of Cryptosporidium spp.,
Dientamoeba fragilis, Entamoeba histolytica, and Giardia intestinalis
in Clinical Stool Samples. Journal of Clinical Microbiology, vol. 49, no. 1,
pp. 257-262.
Stark, D., Roberts, T., Marriott, D., Harkness, J. & Ellis, J.T. 2012, 'Detection and
transmission of Dientamoeba fragilis from environmental and household
samples', The American Journal of Tropical Medicine and Hygiene, vol. 86, no.
2, pp. 233-6.
Stark, D.J., Beebe, N., Marriott, D., Ellis, J.T. & Harkness, J. 2006, 'Dientamoebiasis:
clinical importance and recent advances', Trends in Parasitology, vol. 22, no. 2,
pp. 92-6.

118
Stenzel, D.J. & Boreham, P.F. 1991, 'A cyst-like stage of Blastocystis hominis',
International Journal for Parasitology, vol. 21, no. 5, pp. 613-5.
Stuart , K.D., Weeks, R., Guilbride, L. & Myler, P.J. 1992, 'Molecular organization of
Leishmania RNA virus 1', Proceedings of the National Academy of Sciences of
the United States of America, vol. 89, no. 18, pp. 8596-600.
Stuart, K.D., Weeks, R., Guilbride, L. & Myler, P.J. 1993, 'Nucleotide sequence
and particle organsiation of the LRV 1 RNA virus of Leishmania’, Current
Trends in Leishmania Research', Council of Scientific and Industrial
Research, India, pp. 173-86.
Stensvold, C. R., Clark, C. G. & Röser, D. 2013, Limited intra-genetic diversity in Dientamoeba fragilis housekeeping genes. Infection, Genetics and
Evolution, vol. 18,pp. 284-6.
Sukanahaketu, S. 1977, 'The presence of Dientamoeba fragilis in the Ascaris
lumbricoides ova: the first report from Thailand', Journal of the Medical
Association of Thailand, vol. 60, no. 6, pp. 256-58.
Su, H.M. & Tai, J.H. 1996, 'Genomic organization and sequence conservation in type I
Trichomonas vaginalis viruses', Virology, vol. 222, no. 2, pp. 470-3.
Suresh, K., Ng, G.C., Ho, L.C., Yap, E.H. & Singh, M. 1994, 'Differentiation of the
various stages of Blastocystis hominis by acridine orange staining', International
Journal for Parasitology, vol. 24, no. 4, pp. 605-6.
Tai, J.H., Chang, S.C., Chou, C.F. & Ong, S.J. 1996, 'Separation and characterization of
two related giardiaviruses in the parasitic protozoan Giardia lamblia', Virology,
vol. 216, no. 1, pp. 124-32.
Tai, J.H., Chang, S.C., Ip, C.F. & Ong, S.J. 1995, 'Identification of a satellite double-
stranded RNA in the parasitic protozoan Trichomonas vaginalis infected with T.
vaginalis virus T1', Virology, vol. 208, no. 1, pp. 189-96.
Tai, J.H. & Ip, C.F. 1995, 'The cDNA sequence of Trichomonas vaginalis virus-T1
double-stranded RNA', Virology, vol. 206, no. 1, pp. 773-6.
Tai, J.H., Wang, A.L., Ong, S.J., Lai, K.S., Lo, C. & Wang, C.C. 1991, 'The course of
giardiavirus infection in the Giardia lamblia trophozoites', Experimental
Parasitology, vol. 73, no. 4, pp. 413-23.

119
Tai, J.H., Ong, S.J., Chang, S.C. & Su, H.M. 1993, 'Giardiavirus enters Giardia lamblia
WB trophozoite via endocytosis', Experimental Parasitology, vol. 76, no. 2, pp.
165-74.
Talis, B., Stein, B. & Lengy, J. 1971, 'Dientamoeba fragilis in human feces and bile',
Israel Journal of Medical Sciences, vol. 7, no. 9, pp. 1063-69.
Tarr, P.I., Aline, RF, J.r., Smiley, B.L. , Scholler, J., Keithly, J., Stuart, K . 1988, 'LR1:
a candidate RNA virus of Leishmania', Proceedings of the National Academy of
Sciences of the United States of America., vol. 85, no. 24, pp. 9572-5.
Teow, W.L., Ho, L.C., Ng, G.C., Chan, Y.C., Yap, E.H., Chan, P.P., Howe, J., Zaman,
V. & Singh, M. 1992, 'Virus-like particles in a Blastocystis species from the sea-
snake, Lapemis hardwickii', International Journal for Parasitology, vol. 22, no.
7, pp. 1029-32.
Thomson Jr., R.B. & Smith, T.F. 1982, 'Acridine orange staining of Pneumocystis
carinii', Journal of Clinical Microbiology, vol. 16, no. 1, pp. 191-2.
Tipper, D.J. & Schmitt, M.J. 1991, 'Yeast dsRNA viruses: replication and killer
phenotypes', Molecular Microbiology, vol. 5, no. 10, pp. 2331-8.
Uhlenbeck, O.C. 1987, 'A small catalytic oligoribonucleotide', Nature, vol. 328, no.
6131, pp. 596-600.
Van Etten, J.L. 2011a, 'Another really, really big virus', Viruses, vol. 3, no. 1, pp. 32-46.
Van Etten, J. L 2011b, 'The recent discovery of really, really big viruses is changing
views about the nature of viruses and the history of life', American Scientist ,vol.
99, no. 4, pp. 304.
Van Etten, J.L., Lane, L.C. & Dunigan, D.D. 2010, 'DNA viruses: The really big ones
(giruses)', Annual Review of Microbiology, vol. 64, pp. 83-99.
Vanacova, S., Liston, D.R., Tachezy, J. & Johnson, P.J. 2003, 'Molecular biology of the
amitochondriate parasites, Giardia intestinalis, Entamoeba histolytica and
Trichomonas vaginalis', International Journal for Parasitology, vol. 33, no. 3,
pp. 235-55.
Vanácová, S., Tachezy, J., Kulda, J. & Flegr, J. 1997, 'Characterization of trichomonad
species and strains by PCR fingerprinting', Journal of Eukaryotic Microbiology,
vol. 44, no. 6, pp. 545-52.

120
Vandenberg, O., Souayah, H., Mouchet, F., Dediste, A. & van Gool, T. 2007,
'Treatment of Dientamoeba fragilis infection with paromomycin', The Pediatric
Infectious Disease Journal, vol. 26, no. 1, pp. 88-90.
Verweij, J.J., Mulder, B., Poell, B., van Middelkoop, D., Brienen, E.A. & van Lieshout,
L. 2007, 'Real-time PCR for the detection of Dientamoeba fragilis in fecal
samples', Molecular and Cellular Probes, vol. 21, no. 5-6, pp. 400-4.
Vieira, P.B., Borges, F.P., Gottardi, B., Stuepp, C., Larré, A.B., Tasca, T. & De Carli,
G.A. 2008, 'Analysis of microtubule cytoskeleton distribution using a
fluorescent taxoid in two trichomonadid protozoa: Trichomonas gallinae and
Tritrichomonas foetus', Experimental Parasitology, vol. 119, no. 1, pp. 186-91.
Wang, A.L., Miller, R.L. & Wang, C.C. 1988, 'Antibodies to the Giardia lamblia
double-stranded RNA virus major protein can block the viral infection',
Molecular and Biochemical Parasitology, vol. 30, no. 3, pp. 225-32.
Wang, A.L. & Wang, C.C. 1985 'A linear double-stranded RNA in Trichomonas
vaginalis', Journal of Biological Chemistry, vol. 260, no. 6, pp. 3697-702.
Wang, A.L. & Wang, C.C. 1986a, 'Discovery of a specific double-stranded RNA virus
in Giardia lamblia', Molecular and Biochemical Parasitology, vol. 21, no. 3, pp.
269-76.
Wang, A.L. & Wang, C.C. 1986b, 'The double-stranded RNA in Trichomonas vaginalis
may originate from virus-like particles', Proceedings of the National Academy of
Sciences of the United States of America, vol. 83, no. 20, pp. 7956-60.
Wang, A.L. & Wang, C.C. 1991, 'Viruses of the Protozoa', Annual Review of
Microbiology, vol. 45, no. 1, pp. 251-63.
Wang, A., Wang, C.C. & Alderete, J.F. 1987, 'Trichomonas vaginalis phenotypic
variation occurs only among trichomonads infected with the double-stranded
RNA virus', The Journal of Experimental Medicine, vol. 166, no. 1, pp. 142-50.
Wang, A.L., Yang, H.M., Shen, K.A. & Wang, C.C. 1993, 'Giardiavirus double-
stranded RNA genome encodes a capsid polypeptide and a gag-pol-like fusion
protein by a translation frameshift', Proceedings of the National Academy of
Sciences of the United States of America, vol. 90, no. 18, pp. 8595-9.
Wang, C.C. & Stotish, R.L. 1975, 'Changes of nucleic acids and proteins in the oocysts
of Eimeria tenella during sporulation', Journal of Eukaryotic Microbiology, vol.
22, no. 3, pp. 438-43.

121
Weber, B., Mapeka, T.M., Maahlo, M.A. & Hoosen, A.A. 2003, 'Double stranded RNA
virus in South African Trichomonas vaginalis isolates', Journal of Clinical
Pathology, vol. 56, no. 7, pp. 542-3.
Weber, F., Wagner, V., Rasmussen, S.B., Hartmann, R. & Paludan, S.R. 2006, 'Double-
stranded RNA is produced by positive-strand RNA viruses and DNA viruses but
not in detectable amounts by negative-strand RNA Viruses', Journal of Virology,
vol. 80, no. 10, pp. 5059-64.
Weeks, R., Aline Jr, R.F., Myler, P.J. & Stuart, K. 1992, 'LRV1 viral particles in
Leishmania guyanensis contain double-stranded or single-stranded RNA',
Journal of Virology, vol. 66, no. 3, pp. 1389-93.
Wendel, K.A., Rompalo, A.M., Erbelding, E.J., Chang, T.H. & Alderete, J.F. 2002,
'Double-Stranded RNA viral infection of Trichomonas vaginalis infecting
patients attending a sexually transmitted diseases clinic', Journal of Infectious
Diseases, vol. 186, no. 4, pp. 558-61.
Wenrich, D.H. 1936, 'Studies on Dientamoeba fragilis (protozoa). I. Observations with
special reference to nuclear structure.’ Journal of Parasitology, vol. 22, no. 1,
pp. 76-83.
Wenrich, D.H. 1944, 'Studies on Dientamoeba fragilis (protozoa). IV. Further
observations, with an outline of present-day knowledge of this species', Journal
of Parasitology, vol. 30, no. 5, pp. 322-37.
White, T.C. & Wang, C.C. 1990, 'RNA dependent RNA polymerase activity associated
with the double-stranded RNA virus of Giardia lamblia', Nucleic Acids
Research, vol. 18, no. 3, pp. 553-9.
Wickner, R.B. 1993, 'Double-stranded RNA virus replication and packaging', Journal of
Biological Chemistry, vol. 268, no. 6, pp. 3797-800.
Wickner, R.B. 1996, 'Double-stranded RNA viruses of Saccharomyces cerevisiae',
Microbiological reviews, vol. 60, no. 1, p. 250-65.
Widmer, G. 1995, 'Suppression of Leishmania RNA virus replication by capsid protein
overexpression', Journal of Virology, vol. 69, no. 7, pp. 4122-6.
Widmer, G., Comeau, A.M., Furlong, D.B., Wirth, D.F. & Patterson, J.L. 1989,
'Characterization of a RNA virus from the parasite Leishmania', Proceedings of
the National Academy of Sciences of the United States of America, vol. 86, no.
15, pp. 5979-82.

122
Wileman, T. 2006, 'Aggresomes and autophagy generate sites for virus replication',
Science, vol. 312, no. 5775, pp. 875-8.
Windsor, J.J. & Johnson, E.H. 1999, 'Dientamoeba fragilis: the unflagellated human
flagellate', British Journal of Biomedical Science, vol. 56, no. 4, pp. 293-306.
Windsor, J.J., Macfarlane, L., Hughes-Thapa, G., Jones, S.K. & Whiteside, T.M. 2003,
'Detection of Dientamoeba fragilis by culture', British Journal of Biomedical
Science, vol. 60, no. 2, pp. 79-83.
Windsor, J.J., Clark, C.G. & Macfarlane, L. 2004, 'Molecular typing of Dientamoeba
fragilis', British Journal of Biomedical Science, vol. 61, no. 3, pp. 153-155.
Yang, J. & Scholten, T. 1977, 'Dientamoeba fragilis: a review with notes on its
epidemiology, pathogenicity, mode of transmission, and diagnosis', The
American Journal of Tropical Medicine and Hygiene, vol. 26, no. 1, pp. 16-22.
Yoshikawa, N. & Converse, R.H 1990, 'Strawberry pallidosis disease: distinctive
dsRNA species associated with latent infections in indicators and in diseased
strawberry cultivars', Phytopathology, vol. 80, no. 6. pp. 543-548.
Yu, D., Wang, C.C. & Wang, A.L. 1995a, 'Maturation of giardiavirus capsid protein
involves posttranslational proteolytic processing by a cysteine protease', Journal
of Virology, vol. 69, no. 5, pp. 2825-30.
Yu, D.C., Wang, A.L., Wu, C.H. & Wang, C.C. 1995b, 'Virus-mediated expression of
firefly luciferase in the parasitic protozoan Giardia lamblia', Molecular and
Cellular Biology, vol. 15, no. 9, pp. 4867-72.
Yu, D.C., Wang, A.L. & Wang, C.C. 1996a, 'Stable coexpression of a drug-resistance
gene and a heterologous gene in an ancient parasitic protozoan Giardia lamblia',
Molecular and Biochemical Parasitology, vol. 83, no. 1, pp. 81-91.
Yu, D.C., Wang, A.L. & Wang, C.C. 1996b, 'Amplification, expression, and packaging
of a foreign gene by giardiavirus in Giardia lamblia', Journal of Virology, vol.
70, no. 12, pp. 8752-7.
Zangger, H., Ronet, C., Desponds, C., kuhlmann, F. M., Robinson, J.,
Hartley, M.-A., Prevel, F., Castiglioni, P., Pratlong, F., Bastien, P.,
Müller, N., Parmentier, I., Saravia, N. G., Beverley, S. M. & Fasel, N. 2013,
‘Detection of Leishmania RNA virus in Leishmania parasites, PLoS
Neglected Tropical Diseases, vol. 7, no. 1, pp. 2006.

123
Zaug, A.J., Been, M.D. & Cech, T.R. 1986, 'The Tetrahymena ribozyme acts like an
RNA restriction endonuclease', Nature, vol. 324, no. 6096, pp. 429-33.
Zhao, Y.P., Zhang, X.C., Chen, L.F., Li, J.H., Yin, J.G., Liu, Q. & Gong, P.T. 2006,
'Cloning and sequence analysis of a partial gene of Trichomonas vaginalis
dsRNA virus', Zhongguo ji sheng chong xue yu ji sheng chong bing za zhi, vol.
24, no. 5, pp. 389-90.