transforming growth factor-b1 induces desmoplasia in an...
TRANSCRIPT

[CANCER RESEARCH 61, 550–555, January 15, 2001]
Transforming Growth Factor- b1 Induces Desmoplasia in an ExperimentalModel of Human Pancreatic Carcinoma1
Matthias Lohr, 2 Christian Schmidt, Jorg Ringel, Mario Kluth, Petra Muller, Horst Nizze, and Ralf JesnowskiDivision of Gastroenterology, Departments of Medicine [M. L., C. S., J. R., M. K., P. M., R. J.] and Pathology [H. N.], University of Rostock, D-18055 Rostock, Germany
ABSTRACT
Proliferation of fibrotic tissue (desmoplasia) is one of the hallmarks ofseveral epithelial tumors including pancreatic adenocarcinoma. This tis-sue reaction may be deleterious or advantageous to the host or tumor. Ina systematic analysis, we identified two growth factors expressed byhuman pancreatic carcinoma cells that are positively correlated with theability to induce fibroblast proliferation both in vitro and in vivo, i.e.,transforming growth factor (TGF)- b1 and fibroblast growth factor-2.Here we demonstrate that the overexpression of TGF-b1 induced up-regulation of matrix proteins and growth factors in the TGFb1-trans-fected pancreatic tumor cells. Furthermore, transfection of PANC-1 cellsinduces the same change in fibroblasts in either cocultivation experimentsor when they are grown in conditioned medium from TGF-b1-transfectedPANC-1 cells. TGF-b1-transfected pancreatic tumor cells induced a richstroma after orthotopical transplantation in the nude mouse pancreas.The transfer of a single growth factor, TGF-b1, conveys the ability toinduce a fibroblast response similar to that seen in desmoplasia in humanpancreatic adenocarcinoma. This effect cannot only be attributed to directeffects of TGF-b1 but also results from the up-regulation of several otherfactors including collagen type I, connective tissue growth factor, andplatelet-derived growth factor.
INTRODUCTION
Desmoplasia is a characteristic feature of the growth of somecarcinomas (1). To date, it is not clear whether this process is amechanism to protect the tumor from the host or represents a defensemechanism by the host (2), although there are hints that this stroma isbeneficial for the tumor (3). To tackle desmoplasia therapeutically byeither supporting or suppressing this development, it becomes neces-sary to study the etiology and to attribute this feature to either thetumor cells themselves or the host. Desmoplasia is of particularpredominance in ductal adenocarcinomas of the pancreas exhibiting astrong stromal reaction (4). Therefore, pancreatic carcinoma has be-come a model system to study the interrelation of epithelial tumorcells, matrices, fibroblasts, and growth factors (5–8).
Desmoplastic tissue consists of fibroblasts, as the main cellularcomponent, and extracellular matrix proteins (9). The pancreatic tu-mor cells themselves are able to produce ECM3 proteins (10–13) andinteract with ECM by expressing functionally active integrins (6, 14, 15).
To test the hypothesis of desmoplasia induction by a tumor-derivedgrowth factor, we conducted a deductive analysis correlating theability to induce desmoplasia with the expression of certain growth
factors. Furthermore, we reasoned that the overexpression of such agrowth factor,e.g., TGF-b1 in a pancreatic tumor cell line knownneither to induce desmoplasia nor to express substantial amounts ofTGF-b1 and FGF-2, should result in the gain of the ability to inducefibroblast growth and in an induction of desmoplasia in a xenograftednude mouse model by virtue of direct and indirect effects of TGF-b1.
MATERIALS AND METHODS
Cell Culture and Transfection. AsPC-1, BxPC-3, Capan-1, and PANC-1cells, all from American Type Culture Collection, were cultivated in DMEMwith GlutaMAX I (Life Technologies, Inc.) supplemented with 10% heat-inactivated FCS and antibiotics (100 units/ml penicillin, 100mg/ml strepto-mycinsulfate, and 250 ng/ml amphotericin B; Life Technologies, Inc.; Ref. 10).Mature human recombinant TGF-b1 was purchased from R&D Systems.Full-length cDNA of TGF-b1(16) was cut out of pRK5b1E (BamHI) andcloned into the pcDNA3 vector (Invitrogen) under the control of a cytomeg-alovirus promoter. PANC-1 cells were transfected with this construct or withthe empty pcDNA3 plasmid (mock) by calcium phosphate coprecipitation inN,N-bis(2-hydroxyethyl)-2-aminoethanesulfonic acid buffered saline usingstandard protocols as described (17). This plasmid also codes for theneoresistance gene, enabling selection of transfectants with the antibiotic G418(Sigma; 400mg/ml). Resistant clones were expanded, and expression of thetransfected cDNA was confirmed by Northern blot, Western blot, and ELISA(R&D).
Northern Blot and RT-PCR. Subconfluent layers of PANC-1/TGF-b1cells, mock transfected, untransfected PANC-1 cells, and AsPC-1 and BxPC-3cells were lysed in ice-cold guanidine thiocyanate. RNA preparation wasperformed as described (18). Tenmg of total RNA were subjected to standardformamide gel electrophoresis as described. Gels were blotted to nylon mem-branes (Qiagen) and hybridized with cDNA probes for TGF-b1 (EcoRI/HindIII digest of pcDNA3/TGF-b1), type I collagen (pHCAL1U; Refs. 10 and19), PDGF (Amersham), FGF-2,(20), and CTGF (21) using the nonradioactiveDig labeling kit (Boehringer Mannheim, Mannheim, Germany). In addition,RT-PCR was performed using published primers for TGF-b1, PDGF-A, typeI collagen, and GAPDH. The primers were as follows: TGF-b1 (22), sense59-CAG AAA TAC AGC AAC AAT TCC TGG-39 and antisense 59-TTGCAG TGT GTT ATC CCT GCT GTC-39(190-bp product); PDGF-A (23),sense 59CAG TCA GAT CCA CAG CAT CC-39and antisense 59-AAT GACCGT CCT GGT CTT GC-39(200-bp product); collagen type I (23), sense59-ACG TGA TCT GTG ACG AGA CC-39and antisense 59-AGC AAA GTTTCC TCC GAG GC-39(250-bp product); and GAPDH (24), sense 59-ACCACA GTC CAT GCC ATC AC-39and antisense 59-TCC ACC ACC CTGTTG CTG TA-39 (450-bp product). PCR conditions were the following:denaturing for 30 s at 94°C; annealing for 60 s at 60°C (TGF-b1) or at 64°C(collagen, GAPDH, and PDGF); and extension for 60 s at 72°C. AmplifiedDNA was sampled after 21, 24, 27, and 30 cycles, and the resulting PCRproducts for TGF-b1, collagen, and PDGF-A were loaded in the same gelpockets as the GAPDH amplificate.
Reverse Slot Blot.Expression of genes of several growth factors, recep-tors, and genes of ECM proteins was investigated by reverse slot blot. For thispurpose, plasmid DNA corresponding to 1mg of cDNA insert was blotted ontoa nylon membrane (Qiagen) by use of a slot blot apparatus (Schleicher &Schuell). Hybridization was performed according to standard procedures witha probe obtained by Dig labeling (Boehringer Mannheim) of 7.5mg of totalRNA in a reverse transcription reaction (25, 26). Hybrids were detected usingthe chemiluminescent Dig detection system (Boehringer Mannheim) accordingto the manufacturer’s instructions.
Cocultivation. PANC-1/TGFb1 cells (53 104) were seeded onto Tran-swell inserts (Costar) and were cocultivated with fibroblasts (53 104 cells/
Received 1/28/00; accepted 11/14/00.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 Supported by Grant Lo 431/6 from the Deutsche Forschungsgemeinschaft as part ofthe special topic program “Matrix in Biology and Medicine” (to M. L.). C. S. acknowl-edges the support of the Bundesministerium fur Bildung und Forschung.
2 To whom requests for reprints should be addressed, at Sektion Molekulare Gastro-enterologie, Medizinische Klinik IV, Fakulta¨t fur Klinische Medizin Mannheim, Univer-sitat Heidelberg, Theodor Kutzer Ufer 1-3, D-68135 Mannheim, Germany. Phone: 49-621-383-2900; Fax: 49-381-383-1986; E-mail: [email protected].
3 The abbreviations used are: ECM, extracellular matrix; TGF, transforming growthfactor; RT-PCR, reverse transcription-PCR; PDGF, platelet-derived growth factor; CTGF,connective tissue growth factor; GAPDH, glyceraldehyde-3-phosphate dehydrogenase;Dig, digoxygenin; FGF, fibroblast growth factor; Erk, extracellular signal-regulatedkinase.
550
on July 3, 2019. © 2001 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
well) seeded in six-well tissue culture plates. After 7 days of incubation inDMEM/1% FCS, the cells were trypsinized and counted (trypan blue exclusiontest). Controls were cocultivated of fibroblasts with fibroblasts, PANC-1,PANC-1 mock transfected, and BxPC-1 cells (American Type Culture Col-
lection). From respective parallel experiments, conditioned media (see below)and RNA (see above) were prepared for subsequent analysis.
Conditioned Media. PANC-1/TGFb1 cells were seeded in DMEM/10%FCS. After 2 days of incubation, cells were washed three times with PBS (pH
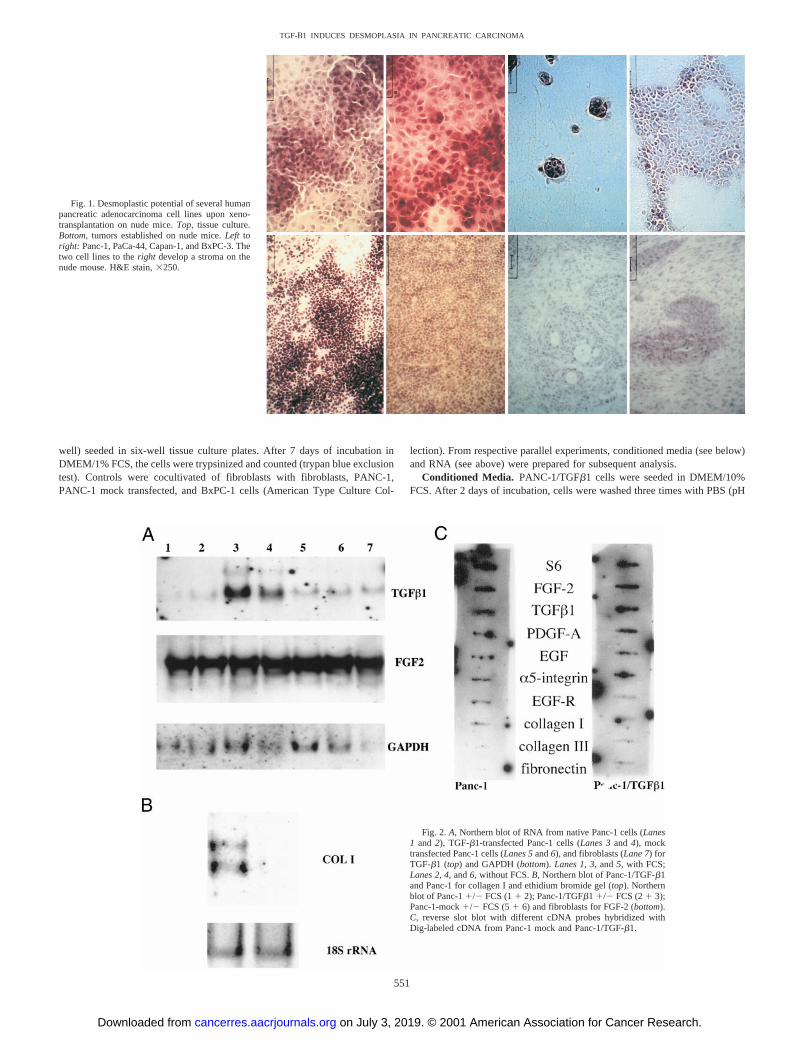
Fig. 1. Desmoplastic potential of several humanpancreatic adenocarcinoma cell lines upon xeno-transplantation on nude mice.Top, tissue culture.Bottom, tumors established on nude mice.Left toright: Panc-1, PaCa-44, Capan-1, and BxPC-3. Thetwo cell lines to theright develop a stroma on thenude mouse. H&E stain,3250.
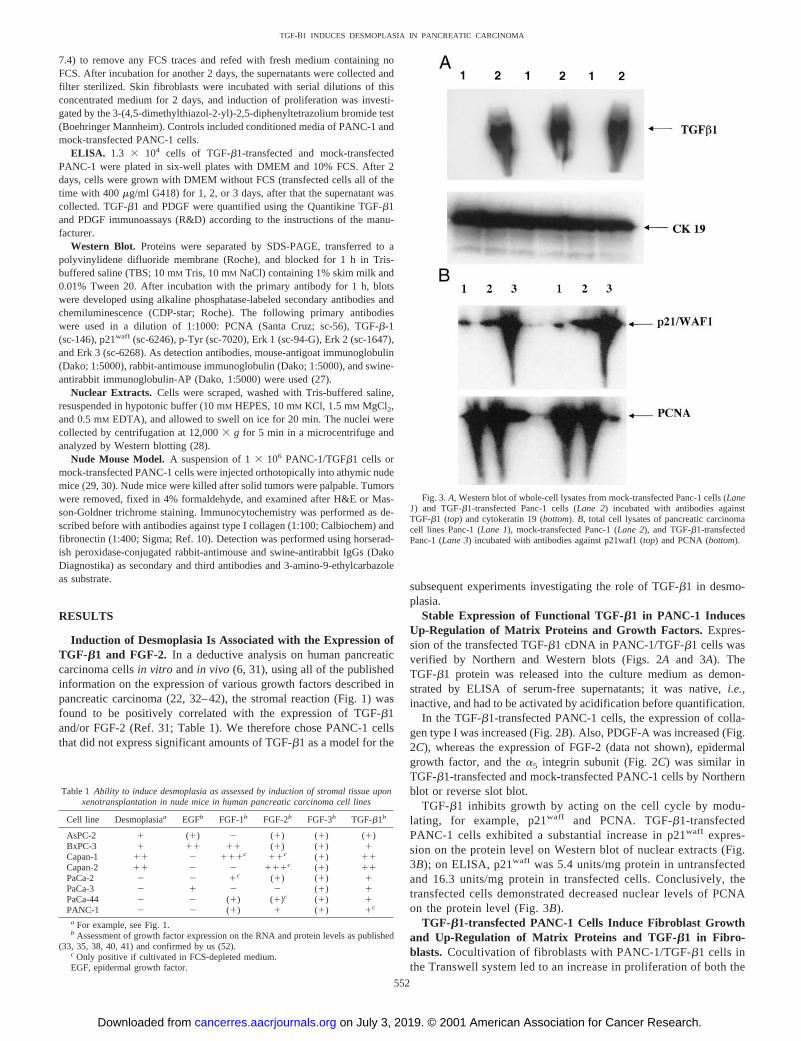
Fig. 2.A, Northern blot of RNA from native Panc-1 cells (Lanes1 and 2), TGF-b1-transfected Panc-1 cells (Lanes 3and 4), mocktransfected Panc-1 cells (Lanes 5and6), and fibroblasts (Lane 7) forTGF-b1 (top) and GAPDH (bottom).Lanes 1, 3,and5, with FCS;Lanes 2, 4,and6, without FCS.B, Northern blot of Panc-1/TGF-b1and Panc-1 for collagen I and ethidium bromide gel (top). Northernblot of Panc-11/2 FCS (11 2); Panc-1/TGFb1 1/2 FCS (21 3);Panc-1-mock1/2 FCS (51 6) and fibroblasts for FGF-2 (bottom).C, reverse slot blot with different cDNA probes hybridized withDig-labeled cDNA from Panc-1 mock and Panc-1/TGF-b1.
551
TGF-B1 INDUCES DESMOPLASIA IN PANCREATIC CARCINOMA
on July 3, 2019. © 2001 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
7.4) to remove any FCS traces and refed with fresh medium containing noFCS. After incubation for another 2 days, the supernatants were collected andfilter sterilized. Skin fibroblasts were incubated with serial dilutions of thisconcentrated medium for 2 days, and induction of proliferation was investi-gated by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide test(Boehringer Mannheim). Controls included conditioned media of PANC-1 andmock-transfected PANC-1 cells.
ELISA. 1.3 3 104 cells of TGF-b1-transfected and mock-transfectedPANC-1 were plated in six-well plates with DMEM and 10% FCS. After 2days, cells were grown with DMEM without FCS (transfected cells all of thetime with 400mg/ml G418) for 1, 2, or 3 days, after that the supernatant wascollected. TGF-b1 and PDGF were quantified using the Quantikine TGF-b1and PDGF immunoassays (R&D) according to the instructions of the manu-facturer.
Western Blot. Proteins were separated by SDS-PAGE, transferred to apolyvinylidene difluoride membrane (Roche), and blocked for 1 h in Tris-buffered saline (TBS; 10 mM Tris, 10 mM NaCl) containing 1% skim milk and0.01% Tween 20. After incubation with the primary antibody for 1 h, blotswere developed using alkaline phosphatase-labeled secondary antibodies andchemiluminescence (CDP-star; Roche). The following primary antibodieswere used in a dilution of 1:1000: PCNA (Santa Cruz; sc-56), TGF-b-1(sc-146), p21wafI (sc-6246), p-Tyr (sc-7020), Erk 1 (sc-94-G), Erk 2 (sc-1647),and Erk 3 (sc-6268). As detection antibodies, mouse-antigoat immunoglobulin(Dako; 1:5000), rabbit-antimouse immunoglobulin (Dako; 1:5000), and swine-antirabbit immunoglobulin-AP (Dako, 1:5000) were used (27).
Nuclear Extracts. Cells were scraped, washed with Tris-buffered saline,resuspended in hypotonic buffer (10 mM HEPES, 10 mM KCl, 1.5 mM MgCl2,and 0.5 mM EDTA), and allowed to swell on ice for 20 min. The nuclei werecollected by centrifugation at 12,0003 g for 5 min in a microcentrifuge andanalyzed by Western blotting (28).
Nude Mouse Model. A suspension of 13 106 PANC-1/TGFb1 cells ormock-transfected PANC-1 cells were injected orthotopically into athymic nudemice (29, 30). Nude mice were killed after solid tumors were palpable. Tumorswere removed, fixed in 4% formaldehyde, and examined after H&E or Mas-son-Goldner trichrome staining. Immunocytochemistry was performed as de-scribed before with antibodies against type I collagen (1:100; Calbiochem) andfibronectin (1:400; Sigma; Ref. 10). Detection was performed using horserad-ish peroxidase-conjugated rabbit-antimouse and swine-antirabbit IgGs (DakoDiagnostika) as secondary and third antibodies and 3-amino-9-ethylcarbazoleas substrate.
RESULTS
Induction of Desmoplasia Is Associated with the Expression ofTGF-b1 and FGF-2. In a deductive analysis on human pancreaticcarcinoma cellsin vitro andin vivo (6, 31), using all of the publishedinformation on the expression of various growth factors described inpancreatic carcinoma (22, 32–42), the stromal reaction (Fig. 1) wasfound to be positively correlated with the expression of TGF-b1and/or FGF-2 (Ref. 31; Table 1). We therefore chose PANC-1 cellsthat did not express significant amounts of TGF-b1 as a model for the
subsequent experiments investigating the role of TGF-b1 in desmo-plasia.
Stable Expression of Functional TGF-b1 in PANC-1 InducesUp-Regulation of Matrix Proteins and Growth Factors. Expres-sion of the transfected TGF-b1 cDNA in PANC-1/TGF-b1 cells wasverified by Northern and Western blots (Figs. 2Aand 3A). TheTGF-b1 protein was released into the culture medium as demon-strated by ELISA of serum-free supernatants; it was native,i.e.,inactive, and had to be activated by acidification before quantification.
In the TGF-b1-transfected PANC-1 cells, the expression of colla-gen type I was increased (Fig. 2B). Also, PDGF-A was increased (Fig.2C), whereas the expression of FGF-2 (data not shown), epidermalgrowth factor, and thea5 integrin subunit (Fig. 2C) was similar inTGF-b1-transfected and mock-transfected PANC-1 cells by Northernblot or reverse slot blot.
TGF-b1 inhibits growth by acting on the cell cycle by modu-lating, for example, p21wafI and PCNA. TGF-b1-transfectedPANC-1 cells exhibited a substantial increase in p21wafI expres-sion on the protein level on Western blot of nuclear extracts (Fig.3B); on ELISA, p21wafI was 5.4 units/mg protein in untransfectedand 16.3 units/mg protein in transfected cells. Conclusively, thetransfected cells demonstrated decreased nuclear levels of PCNAon the protein level (Fig. 3B).
TGF-b1-transfected PANC-1 Cells Induce Fibroblast Growthand Up-Regulation of Matrix Proteins and TGF-b1 in Fibro-blasts. Cocultivation of fibroblasts with PANC-1/TGF-b1 cells inthe Transwell system led to an increase in proliferation of both the
Table 1 Ability to induce desmoplasia as assessed by induction of stromal tissue uponxenotransplantation in nude mice in human pancreatic carcinoma cell lines
Cell line Desmoplasiaa EGFb FGF-1b FGF-2b FGF-3b TGF-b1b
AsPC-2 1 (1) 2 (1) (1) (1)BxPC-3 1 11 11 (1) (1) 1Capan-1 11 2 111c 11c (1) 11Capan-2 11 2 2 111c (1) 11PaCa-2 2 2 1c (1) (1) 1PaCa-3 2 1 2 2 (1) 1PaCa-44 2 2 (1) (1)c (1) 1PANC-1 2 2 (1) 1 (1) 1c
a For example, see Fig. 1.b Assessment of growth factor expression on the RNA and protein levels as published
(33, 35, 38, 40, 41) and confirmed by us (52).c Only positive if cultivated in FCS-depleted medium.EGF, epidermal growth factor.
Fig. 3.A, Western blot of whole-cell lysates from mock-transfected Panc-1 cells (Lane1) and TGF-b1-transfected Panc-1 cells (Lane 2) incubated with antibodies againstTGF-b1 (top) and cytokeratin 19 (bottom).B, total cell lysates of pancreatic carcinomacell lines Panc-1 (Lane 1), mock-transfected Panc-1 (Lane 2), and TGF-b1-transfectedPanc-1 (Lane 3) incubated with antibodies against p21waf1 (top) and PCNA (bottom).
552
TGF-B1 INDUCES DESMOPLASIA IN PANCREATIC CARCINOMA
on July 3, 2019. © 2001 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

fibroblasts and the tumor cells (Fig. 4A), whereas cocultivationwith mock-transfected PANC-1 cells did not exhibit this effect. Onthe RNA level, induction of collagen type I in the fibroblasts couldbe demonstrated after incubation with conditioned media fromTGF-b1-transfected PANC-1 cells (Fig. 4B); moreover, an up-regulation of TGF-b1 expression could be demonstrated by RT-PCR under this conditions (Fig. 4B). Although CTGF expressionremained unchanged in the PANC-1 cells after TGF-b1 transfec-tion (data not shown), a significant increase in CTGF mRNA wasdetectable in fibroblasts after cocultivation with TGF-b1-trans-fected PANC-1 cells (Fig. 4B).
Incubation of fibroblasts in conditioned media of TGF-b1-trans-fected PANC-1 cells resulted in more pronounced tyrosine phos-phorylation of proteins in fibroblasts than incubation with super-natants from mock-transfected and untransfected PANC-1 cells
(Fig. 5, top). Furthermore, mitogen-activated protein kinases wereactivated as indicated by a mobility shift of Erk 1/2 (Fig. 5,bottom). As mentioned with the tyrosine phosphorylation, the mostpronounced phosphorylation of Erk 1/2 and 3 could be demon-strated after incubation with supernatants of PANC-1/TGF-b1(Fig. 5, bottom). Here, an increase in the activated,i.e., phospho-rylated, kinases was evident.
TGF-b1-transfected PANC-1 Cells Induce Desmoplasia withIncrease in Matrix Proteins in Vivo. PANC-1/TGF-b1 trans-fected cells and mock-transfected cells were injected orthotopi-cally into the nude mouse pancreas. Tumors were harvested after 2months. The tumors grown from TGF-b1-transfected cells demon-strated an increased desmoplasia surrounding the tumor cells ascompared with the mock-transfected cells (Fig. 6). This was evi-dent both on the tumor margin toward the normal mouse pancreasas well as within the tumor. In addition, collagen type I andfibronectin could be detected in increased amounts surrounding thetumor cells (Fig. 6).
DISCUSSION
The desmoplastic reaction is one of the morphological hallmarks ofseveral human tumors (1) originating from solid epithelial glands,such as pancreatic adenocarcinoma, that sets it apart from otherepithelial tumors. Beside the description and static expression analysisof potential factors, no detailed analysis has been performed to dissectthis phenomenon. The pancreatic tumor cells themselves producematrix proteins (10) and express a variety of integrins (6, 15). Fur-
Fig. 4. A, cocultivation of mock-transfected Panc-1 cells and TGF-b1-transfectedPanc-1 cells with fibroblasts in the TransWell system. Cultivation of the tumor cells on topin the insert with the fibroblasts in the bottom well orvice versais shown. Theouter ofthe four columnsin each set represent the baseline of tumor cells (left/light gray) andfibroblasts (right/white) grown without cocultivation. Theinner columnsrepresent the cellcounts for tumor cells (dark gray) and fibroblasts (black) under cocultivation for 3 days.COL I, collagen I.B, Northern blot of RNA from fibroblasts cultivated with and withoutFCS (Lanes 1and2); or with conditioned media from Panc-1 (Lane 3); Panc-1/TGFb1(Lane 4) and Panc-1-mock (Lane 5) hybridized for collagen I and 18S rRNA (loadingcontrol. Middle: RT-PCR products for TGF-b1 and GAPDH (control) of fibroblastsincubated in DMEM1 FCS (Lane 1); or in conditioned media from Panc-1-mock for 1or 3d (Lanes 2and3) and Panc-1/TGF-b1 for 1 or 3 days (Lanes 4and5) after 27 (left)and 30 (right) cycles.Bottom,Northern blot for CTGF in fibroblasts after cultivation alone(F) or after cocultivation with mock-transfected (FM) and TGF-b1-transfected (FT)PANC-1 cells (loading control, 18S rRNA).
Fig. 5. Induction of tyrosine phosphorylation in fibroblasts after incubation in super-natants from Panc-1, Panc-1 mock, and Panc-1/TGF-b1; Lane 1,control (plain DMEMmedium; top). Activation of mitogen-activated protein kinases Erk 1 and Erk 2 afterincubation in supernatants (bottom).P, the activated, hence phosphorylated, kinase.
553
TGF-B1 INDUCES DESMOPLASIA IN PANCREATIC CARCINOMA
on July 3, 2019. © 2001 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

thermore, the expression of growth factors and their receptors hasbeen demonstrated conclusively, however, mostly related to a dem-onstration of the autocrine growth-promoting effect (35, 36, 43).
To test our hypothesis of a positive correlation of stroma inductionand TGF-b1 expression, we successfully transfected the tumor cellline PANC-1 with a TGF-b1 expression vector.
For TGF-b1, it has been suggested that the major regulatory stepcontrolling TGF-b1 activity takes place extracellularly. The same wastrue for the transfected PANC-1 cells; TGF-b1 was released into theculture medium in a latent,i.e., not activated, state. Recently, it wasdemonstrated that latent TGF-b1 can bind to and be activated by theavb6 integrin (44). These integrin subunits are also expressed bypancreatic carcinomas (15),i.e., the PANC-1 cells (6). Thus, activa-tion of the released TGF-b1 may be accomplished in this way.Expression of TGF-b1 resulted in an up-regulation of the matrixproteins collagen type I and fibronectin in the tumor cells themselves.Furthermore, PDGF expression was increased in the transfected cells.This altered gene expression resulted in several paracrine effects onfibroblasts in cocultivation experiments. We could demonstrate anincrease in collagen type I synthesis in the fibroblasts after stimulationwith supernatants from TGF-b1-transfected PANC-1 cells. Similarly,the activation of collagen type VII regulatory elements by TGF hasbeen described recently (45). The fibroblasts themselves producedmore TGF-b1 upon stimulation (cocultivation or conditioned media)by the TGF-b1-transfected PANC-1 cells. This is supported by theobservation that in pancreatic carcinoma tissue, TGF-b1 is mostpredominant in the stroma (46). Furthermore, collagen type I, the mostpredominant basal membrane matrix protein in pancreatic carcinoma
(10), is also up-regulated, both in the tumor cells themselves and inthe fibroblasts upon cocultivation. This up-regulation, however, mayonly be in part attributed to TGF-b1 itself; it could also be the resultof the up-regulation of PDGF-A that has been shown to be a cofactorin TGF-b1-induced collagen type I stimulation (23). In the fibroblasts,after cocultivation with TGF-b1-transfected PANC-1 cells,CTGF,one of the index TGF-b1 response genes (47), was increased. Inhibi-tion of CTGF abrogated the TGF-b1-induced collagen gene up-regulation, confirming the pivotal role of this growth factor (48). Asa result of these alterations in gene expression mentioned above, thetransfection of TGF-b1 in the pancreatic tumor cell line PANC-1 ledto a gain of stromal tissue after orthotopic transplantation in the nudemouse when compared with mock-transfected PANC-1 cells.
The influence of the matrix on signal transduction has long beenunder debate (49, 50). We have shown that a single growth factor,TGF-b1, is capable of conferring the desmoplastic potential to tumorcells not capable of these features. Some of the effects may beattributed to a direct effect of TGF-b1, whereas others,e.g., theup-regulation of collagen type I (51), may be the result of indirecteffects of TGF-b1 intimately associated with the signal transductionpathway involved in TGF-b1 activities.
ACKNOWLEDGMENTS
We thank Roland M. Schmid for assistance in subcloning the TGF-b1plasmid and Thomas Gress (both of University of Ulm, Ulm, Germany) forsupplying us with the CTGF plasmid.
Fig. 6. Orthotopic tumors after intrapancreaticinjection of TGF-b1-transfected PANC-1 cells (B,D, andF) and mock-transfected PANC-1 cells (A,C, andE) into the nude mouse pancreas.A andB,Masson-Goldner trichrome staining. Immunocyto-chemistry for collagen type I (Cand D) and fi-bronectin (EandF) is shown.
554
TGF-B1 INDUCES DESMOPLASIA IN PANCREATIC CARCINOMA
on July 3, 2019. © 2001 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

REFERENCES
1. van den Hooff, A. Connective tissue changes in cancer. Int. Rev. Connect. TissueRes.,10: 395–432, 1983.
2. Dvorak, H. F. Tumors: wounds that do not heal. Similarities between tumor stromageneration and wound healing. N. Engl. J. Med.,315: 1650–1659, 1986.
3. Sethi, T., Rintoul, R. C., Moore, S. M., MacKinnon, A. C., Salter, D., Choo, C.,Chilvers, E. R., Dransfield, I., Donnelly, S. C., Strieter, R., and Haslett, C. Extracel-lular matrix proteins protect small cell lung cancer cells against apoptosis: a mech-anism for small cell lung cancer growth and drug resistancein vivo. Nat. Med.,5:662–668, 1999.
4. Kloppel, G., Lingenthal, G., Bulow, M. V., and Kern, H. F. Histological and finestructural features of pancreatic ductal adenocarcinomas in relation to growth andprognosis: studies in xenografted tumours and clinico-histopathological correlation ina series of 75 cases. Histopathol. (Oxf.),9: 841–856, 1985.
5. Longnecker, D. S., Jamieson, J. D., and Asch, H. L. Regulation of growth anddifferentiation in pancreatic cancer. Pancreas,4: 256–275, 1989.
6. Lohr, M., Trautmann, B., Peters, S., Zauner, I., Meier, A., Kloppel, G., Liebe, S., andKreuser, E. D. Expression and function of receptors for extracellular matrix proteinsin human ductal adenocarcinomas of the pancreas. Pancreas,12: 248–259, 1996.
7. Ingber, D. E., Madri, J. A., and Jamieson, J. D. Neoplastic disorganization ofpancreatic epithelial cell-cell relations. Am. J. Pathol.,121: 248–260, 1985.
8. Hall, P. A., and Lemoine, N. R. Models for pancreatic cancer.In: N. R. Lemoine andN. A. Wright (eds.), Cancer Surveys, Vol. 16, The Molecular Pathology of Cancer,pp. 135–155. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory, 1993.
9. Liotta, L. A., Rao, C. N., and Barsky, S. H. Tumor cell interaction with theextracellular matrix.In: L. A. Liotta (ed.), The Role of Extracellular Matrix inDevelopment, pp. 357–371. New York: Alan R. Liss, Inc., 1984.
10. Lohr, M., Trautmann, B., Gottler, M., Peters, S., Zauner, I., Maillet, B., and Kloppel,G. Human ductal adenocarcinomas of the pancreas express extracellular matrixproteins. Br. J. Cancer,69: 144–151, 1994.
11. Haberern-Blood, C., Liotta, L. A., Rao, C. N., and Kupchik, H. Z. Laminin expressionby human pancreatic carcinoma cells in the nude mouse and in culture. J. Natl. CancerInst., 79: 891–898, 1987.
12. Haglund, C., Roberts, P. J., Nordling, S., and Ekblom, P. Expression of laminin inpancreatic neoplasms and in chronic pancreatitis. Am. J. Surg. Pathol.,8: 669–676,1984.
13. Tani, T., Lumme, A., Linnala, A., Kivilaakso, E., Kiviluoto, T., Burgeson, R. E.,Kangas, L., Leivo, I., and Virtanen, I. Pancreatic carcinomas deposit laminin-5,preferably adhere to laminin-5, and migrate on the newly deposited basement mem-brane. Am. J. Pathol.,151: 1289–1302, 1997.
14. Weinel, R. J., Rosendahl, A., Neumann, K., Chaloupka, B., Erb, D., Rothmund, M.,and Santoso, S. Expression and function of VLA-a2, -a3, -a5, and a6-integrinreceptors in pancreatic carcinoma. Int. J. Cancer,52: 827–833, 1992.
15. Hall, P. A., Coates, P., Lemoine, N. R., and Horton, M. A. Characterization of integrinchains in normal and neoplastic human pancreas. J. Pathol.,165: 33–41, 1991.
16. Wrann, M., Bodmer, S., Martin, R. D., Siepl, C., Hofer-Warbinek, R., Frei, K., Hofer,E., and Fontana, A. T cell suppressor factor from human glioblastoma cells is a 12.5Kd protein closely related to transforming growth factor-b. EMBO J.,6: 1633–1636,1987.
17. Jesnowski, R., Liebe, S., and Lohr, M. Increasing the transfection efficacy andsubsequent long-term culture of resting human pancreatic duct epithelial cells. Pan-creas,17: 262–265, 1998.
18. Sparmann, G., Jaschke, A., Muller, P., Lohr, M., Liebe, S., and Emmrich, J. Tissuehomogenization as a key step extracting RNA from human and rat pancreatic tissue.Biotechniques,22: 408–412, 1997.
19. Makela, J. K., Raassina, M., and Vuorio, E. Human proa1(I) collagen: cDNAsequence for the C-propeptide domain. Nucleic Acids Res.,16: 349, 1988.
20. Abraham, J. A., Mergia, A., Whang, J. L., Tumolo, A., Friedman, J., Hjerrild, K. A.,Gospodarowicz, D., and Fiddes, J. C. Nucleotide sequence of a bovine clone encodingthe angiogenic protein, basic fibroblast growth factor. Science (Washington DC),545: 545–548, 1986.
21. Wenger, C., Ellenrieder, V., Alber, B., Lacher, U., Menke, A., Hameister, H., Wilda,M., Iwamura, T., Beger, H. G., Adler, G., and Gress, T. M. Expression and differ-ential regulation of connective tissue growth factor in pancreatic cancer cells. Onco-gene,18: 1078–1080, 1999.
22. van Laethem, J. L., Resibois, A., Rickaert, F., Deviere, J., Gelin, M., Cremer, M., andRobberecht, P. Different expression of transforming growth factorb1 in pancreaticductal adenocarcinoma and cystic neoplasms. Pancreas,15: 41–47, 1997.
23. Halloran, B. G., So, B. J., and Baxter, B. T. Platelet-derived growth factor is acofactor in the induction of 1a(I) procollagen expression by transforming growthfactor-b 1 in smooth muscle cells. J. Vasc. Surg.,23: 767–773, 1996.
24. Ning, Y., Roschke, A., Christian, S., Lesser, J., Sutcliffe, J. S., and Ledbetter, D. H.Identification of a novel paternally expressed transcript adjacent to snRPN in thePrader-Willi syndrome critical region. Genome Res.,6: 742–746, 1996.
25. von Heimendahl, G., Gebhardt, E., and Dingermann, T. Proto-oncogene activation insurgical specimens of rectal carcinoma. Anticancer Res.,8: 805–812, 1988.
26. Jesnowski, R., Muller, P., Schareck, W., Liebe, S., and Lohr, M. Immortalizedpancreatic duct cellsin vitro and in vivo. Ann. NY Acad. Sci.,880: 50–65, 1999.
27. Schmidt, C., Pommerencke, H., Durr, F., Nebe, B., and Rychly, J. Mechanicalstressing of integrin receptors induces enhanced tyrosine phosphorylation of cytoskel-etally anchored proteins. J. Biol. Chem.,273: 5081–5085, 1998.
28. Tilbrook, P. A., Bittorf, T., Callus, B., Busfield, S. J., Spadaccini, A., Ali, M., Ingley,E., and Klinken, S. P. Regulation of the erythropoietin receptor and involvement ofJAK2 in differentiation of J2E erythroid cells. Cell Growth Differ.,7: 511–520, 1996.
29. Fu, X., Guadagni, F., and Hoffman, R. M. A metastatic nude-mouse model of humanpancreatic cancer constructed orthotopically with histologically intact patient speci-mens. Proc. Natl. Acad. Sci. USA,89: 5645–5649, 1992.
30. Reyes, G., Villanueva, A., Garcia, C., Sancho, F. J., Piulats, J., Lluıs, F., and Capella´,G. Orthotopic xenografts of human pancreatic carcinomas acquire genetic aberrationsduring dissemination in nude mice. Cancer Res.,56: 5713–5719, 1996.
31. Lohr, M. Stromal-epithelial interactions in pancreatic carcinomas.In: N. R. Lemoineand J. Neoptolemos (eds.), Cell Biology of Pancreatic Cancer, pp. 18–35. London:Blackwell Science, 1996.
32. Korc, M., Meltzer, P., and Trent, J. Enhanced expression of epidermal growth factorreceptor correlates with alterations of chromosome 7 in human pancreatic cancer.Proc. Natl. Acad. Sci. USA,83: 5141–5144, 1986.
33. Leung, H. Y., Hughes, C. M., Kloppel, G., Williamsons, R. C. N., and Lemoine, N. R.Localization of expression of fibroblast growth factors and their receptors in pancre-atic adenocarcinoma byin situ hybridization. Int. J. Oncol.,4: 1219–1223, 1994.
34. Estival, A., Clerc, P., Vayesse, N., Tam, J. P., and Clemente, F. Decreased expressionof transforming growth factora during differentiation of human pancreatic cancercells. Gastroenterology,103: 1851–1859, 1992.
35. Leung, H. Y., Gullick, W. J., and Lemoine, N. R. Expression and functional activityof fibroblast growth factors and their receptors in human pancreatic cancer. Int. J.Cancer,59: 667–675, 1994.
36. Kornmann, M., Beger, H. G., and Korc, M. Role of fibroblast growth factors and theirreceptors in pancreatic cancer and chronic pancreatitis. Pancreas,17: 169–175, 1998.
37. Friess, H., Yamanaka, Y., Buchler, M., Ebert, M., Beger, H. G., Gold, L. I., and Korc,M. Enhanced expression of transforming growth factorb isoforms in pancreaticcancer correlates with decreased survival. Gastroenterology,105: 1846–1856, 1993.
38. Barton, C., Hall, P. A., Hughes, C. M., Gullick, W. J., and Lemoine, N. R. Trans-forming growth factora and epidermal growth factor in human pancreatic cancer.J. Pathol.,163: 111–116, 1991.
39. Beauchamp, R. D., Lyons, R. M., Yang, E. Y., Coffey, R. J., and Moses, H. L.Expression of and response to growth regulatory peptides by two human pancreaticcarcinoma cell lines. Pancreas,5: 369–380, 1990.
40. Kalthoff, H., Roeder, C., Humburg, I., Thiele, H. G., Greten, H., and Schmiegel, W.Modulation of platelet-derived growth factor A- and B-chain/c-sis mRNA by tumornecrosis factor and other agents in adenocarcinoma cells. Oncogene,6: 1015–1021,1991.
41. Kalthoff, H., Roeder, C., Gieseking, J., Humburg, I., and Schmiegel, W. Inverseregulation of human ERBB2 and epidermal growth factor receptors by tumor necrosisfactor a. Proc. Natl. Acad. Sci. USA,90: 8972–8976, 1993.
42. Schmiegel, W., Roeder, C., Schmielau, J., Rodeck, U., and Kalthoff, H. Tumornecrosis factora and the epithelial growth factor receptor in human pancreatic cancercells. Proc. Natl. Acad. Sci. USA,90: 863–867, 1993.
43. Lemoine, N. R., Leung, H. Y., Barton, C. M., Hughes, C. M., Kloppel, G., andGullick, W. J. Autocrine growth control of pancreatic cancer. Int. J. Pancreatol.,14:69–70, 1993.
44. Munger, J. S., Huang, X., Kawakatsu, H., Griffiths, M. J. D., Dalton, S. L., Wu, J.,Pittet, J. F., Kaminski, N., Garat, C., Matthay, M. A., Rifkin, D. B., and Sheppard, D.The integrinavb6 binds and activates latent TGF-b 1: a mechanism for regulatingpulmonary inflammation and fibrosis. Cell,96: 319–328, 1999.
45. Rodeck, U., Nishiyama, T., and Mauviel, A. Independent regulation of growth andSMAD-mediated transcription by transforming growth factorb in human melanomacells. Cancer Res.,59: 547–550, 1999.
46. Satoh, K., Shimosegawa, T., Hirota, M., Koizumi, M., and Toyota, T. Expression oftransforming growth factorb1 (TGFb1) and its receptors in pancreatic duct cellcarcinoma and in chronic pancreatitis. Pancreas,16: 468–474, 1998.
47. Geng, M. M., Ellenrieder, V., Wallrapp, C., Muller-Pilasch, F., Sommer, G., Adler,G., and Gress, T. M. Use of representational difference analysis to study the effect ofTGFb on the expression profile of a pancreatic cancer cell line. Genes ChromosomesCancer,26: 70–79, 1999.
48. Duncan, M. R., Frazier, K. S., Abramson, S., Williams, S., Klapper, H., Huang, X.,and Grotendorst, G. R. Connective tissue growth factor mediates transforming growthfactor b-induced collagen synthesis: down-regulation by cAMP. FASEB J.,13:1774–1786, 1999.
49. Bissell, M. J., Hall, H. G., and Parry, G. How does the extracellular matrix direct geneexpression? J. Theor. Biol.,99: 31–68, 1982.
50. Bissell, M. J., and Barcellos-Hoff, M. H. The influence of extracellular matrix ongene expression: is structure the message? J. Cell Sci. (Suppl.),8: 327–343, 1987.
51. Sparmann, G., Merkord, J., Jaschke, A., Nizze, H., Jonas, L., Lohr, M., Liebe, S., andEmmrich, J. Pancreatic fibrosis in experimental pancreatitis induced by dibutylindichloride. Gastroenterology,112: 1664–1672, 1997.
52. Kalthoff, H., Lohr, M., Roeder, C., and Schmiegel, W. Das Pankreaskarzinom.Zellbiologie, Matrixproteine und Wachstumsregulation.In: G. Adler, U. R. Folsch, J.Mossner, and M. V. Singer (eds.), Erkrankungen des exkretorischen Pankreas, Ed. 2,pp. 385–404. Jena: Fischer, 1995.
555
TGF-B1 INDUCES DESMOPLASIA IN PANCREATIC CARCINOMA
on July 3, 2019. © 2001 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

2001;61:550-555. Cancer Res Matthias Löhr, Christian Schmidt, Jörg Ringel, et al. Experimental Model of Human Pancreatic Carcinoma
1 Induces Desmoplasia in anβTransforming Growth Factor-
Updated version
http://cancerres.aacrjournals.org/content/61/2/550
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/61/2/550.full#ref-list-1
This article cites 46 articles, 9 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/61/2/550.full#related-urls
This article has been cited by 31 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/61/2/550To request permission to re-use all or part of this article, use this link
on July 3, 2019. © 2001 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from