tracers of past ocean circulation - georgia institute of...
TRANSCRIPT

Tre
8.16 Tracers of Past Ocean CirculationJ Lynch-Stieglitz, Georgia Institute of Technology, Atlanta, GA, USATM Marchitto, University of Colorado, Boulder, CO, USA
ã 2014 Elsevier Ltd. All rights reserved.
This article is a revision of the previous edition article by J. Lynch-Stieglitz, volume 6, pp. 433–451, © 2003, Elsevier Ltd.
8.16.1 Introduction 4358.16.2 Nutrient Water Mass Tracers 4368.16.2.1 Carbon Isotopes 4368.16.2.1.1 Controls on d13C of oceanic carbon 4368.16.2.1.2 Carbon isotope ratios in benthic foraminifera 4378.16.2.2 Cadmium in Benthic Foraminifera 4388.16.2.2.1 Cd in seawater 4388.16.2.2.2 Cd/Ca in foraminifera 4398.16.2.3 Ba/Ca 4408.16.2.4 Zn/Ca 4408.16.2.5 Silicon Isotopes 4408.16.3 Conservative Water Mass Tracers 4408.16.3.1 Mg/Ca in Benthic Foraminifera 4408.16.3.2 Pore Water Chemistry 4408.16.3.3 Oxygen Isotopes in Benthic Foraminifera 4418.16.3.4 Clumped Isotopes 4418.16.3.5 Carbon Isotope Air–Sea Exchange Signature 4418.16.4 Neodymium Isotope Ratios 4428.16.5 Circulation Rate Tracers 4428.16.5.1 Radiocarbon 4428.16.5.2 231Pa/230Th Ratio 4438.16.5.3 Geostrophic Shear Estimates from d18O in Benthic Foraminifera 4438.16.6 Nongeochemical Tracers of Past Ocean Circulation 4438.16.7 Ocean Circulation during the LGM 4448.16.8 Conclusions 448References 448
8.16.1 Introduction
Information about how the ocean circulated during the past is
useful in understanding changes in ocean and atmospheric
chemistry, changes in the fluxes of heat and freshwater be-
tween the ocean and the atmosphere, and changes in global
wind patterns. The circulation of surface waters in the ocean
leaves an imprint on sea surface temperature and is also inex-
tricably linked to the patterns of oceanic productivity. Much
valuable information about past ocean circulation has been
inferred from reconstructions of surface ocean temperature and
productivity, which are covered in separate chapters. Here, the
focus is on geochemical tracers that are used to infer the flow
patterns and mixing of subsurface water masses.
Several decades ago, it was realized that the chemistry of the
shells of benthic foraminifera (carbon isotope and Cd/Ca
ratios) carries an imprint of the nutrient content of deep water
masses (Boyle, 1981; Broecker, 1982; Shackleton, 1977).
This led rapidly to the recognition that water masses in the
Atlantic Ocean were arrayed differently during the Last Glacial
Maximum (LGM) than they are today and the hypothesis that
the glacial arrangement reflected a diminished contribution of
atise on Geochemistry 2nd Edition http://dx.doi.org/10.1016/B978-0-08-095975
low-nutrient North Atlantic Deep Water (NADW; Boyle and
Keigwin, 1982; Curry and Lohmann, 1982). More detailed
spatial reconstructions indicated a shallow nutrient-depleted
water mass overlying a more nutrient-rich water mass in the
glacial Atlantic. These findings spurred advances not only in
geochemistry but also in oceanography and climatology, as
workers in these fields attempted to simulate the inferred glacial
circulation patterns and assess the vulnerability of the modern
ocean circulation to changes observed for the last ice age.
While the nutrient distributions in the glacial Atlantic
Ocean were consistent with a diminished flow of NADW,
they also could have reflected an increase in inflow from the
South Atlantic and/or a shallower yet undiminished deep
water mass. Clearly, tracers capable of giving information on
deep water flow rate, rather than nutrient content alone, were
needed. Differences between surface water (measured on
planktonic foraminifera) and deepwater (measured on coexist-
ing benthic foraminifera) radiocarbon concentrations pro-
vided the first rate constraint (Broecker et al., 1988;
Shackleton et al., 1988). Reduced amounts of Pa relative to
the more particle-reactive Th in the glacial Atlantic suggested
that deep water was exported from the Atlantic during glacial
-7.00616-1 435
436 Tracers of Past Ocean Circulation
times (Yu et al., 1996). Density gradients in upper waters
reflect the vertical shear in oceanic transports, especially at
intermediate depths where these gradients are largest (Lynch-
Stieglitz et al., 1999b).
Many of these tracers of paleo-ocean flow have been ap-
plied to all of the ocean basins and have been extended in time
throughout the Neogene. Despite this progress, a consistent
picture of the circulation of the ocean during even the last ice
age is yet to emerge. While circulation tracers suggest a rearran-
gement of water masses in the Atlantic, there is still consider-
able disagreement about the water masses and circulation in
the rest of the world ocean. Some of this uncertainty arises
from still insufficient data coverage but some is the result of
conflicting information from the various deep circulation
tracers. In this chapter, the methods used to reconstruct
past ocean circulation, which will illuminate the source of
some of this conflicting information, are examined in more
detail.
8.16.2 Nutrient Water Mass Tracers
8.16.2.1 Carbon Isotopes
Measuring the carbon isotope ratio in calcite tests of bottom-
dwelling (benthic) foraminifera is perhaps the most
widespread method for reconstructing the distribution and
properties of deep-sea water masses. Carbon isotope ratios in
foraminifera reflect that ratio in seawater. In turn, the carbon
isotope ratio in the deep sea is primarily controlled by the
regeneration of 13C-poor organic material and has a distribu-
tion much like a nutrient such as phosphate in the modern
ocean. While a nutrient tracer reflects the increased contribu-
tion from the regeneration of organic material as the water
mass ages, it also carries a signature of the initial nutrient
NADW
1.5
1.0
0.5
0.0
−0.51.0
PO4 (μ
d 13Ca
d 13Cas =0
δ13C
(‰ P
DB
)
Air–
sea
exch
angeBiology
1.5
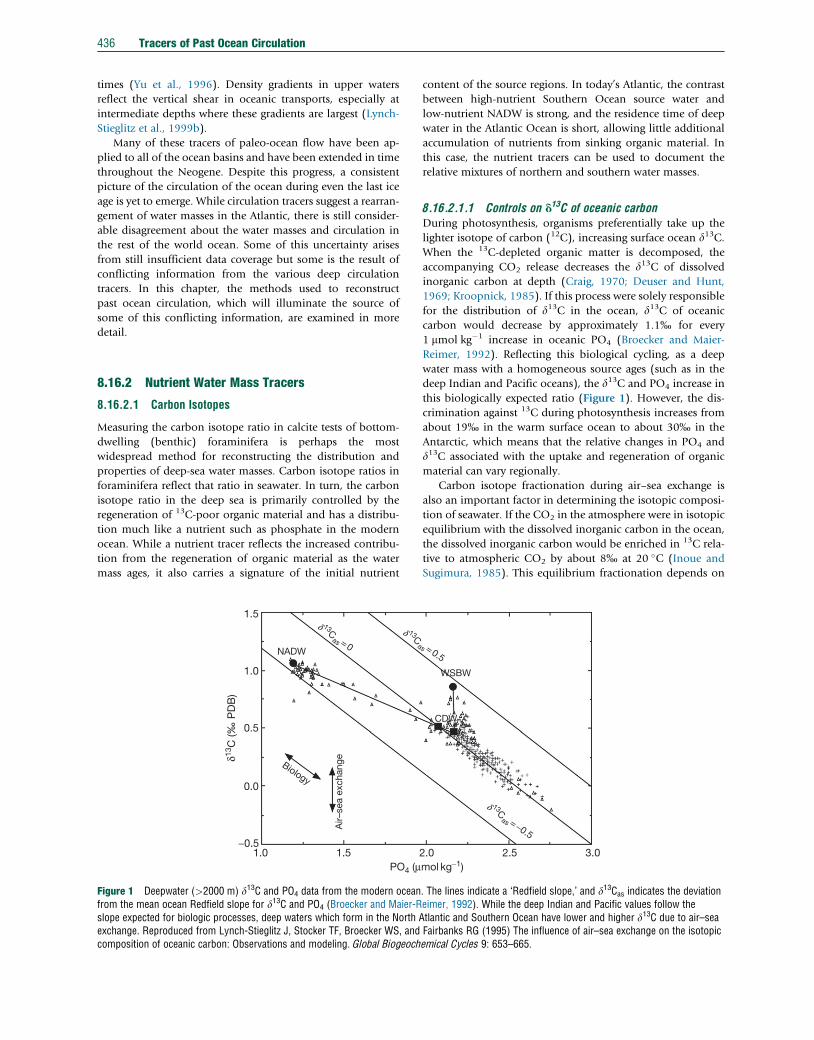
Figure 1 Deepwater (>2000 m) d13C and PO4 data from the modern oceanfrom the mean ocean Redfield slope for d13C and PO4 (Broecker and Maier-Rslope expected for biologic processes, deep waters which form in the North Aexchange. Reproduced from Lynch-Stieglitz J, Stocker TF, Broecker WS, andcomposition of oceanic carbon: Observations and modeling. Global Biogeoch
content of the source regions. In today’s Atlantic, the contrast
between high-nutrient Southern Ocean source water and
low-nutrient NADW is strong, and the residence time of deep
water in the Atlantic Ocean is short, allowing little additional
accumulation of nutrients from sinking organic material. In
this case, the nutrient tracers can be used to document the
relative mixtures of northern and southern water masses.
8.16.2.1.1 Controls on d13C of oceanic carbonDuring photosynthesis, organisms preferentially take up the
lighter isotope of carbon (12C), increasing surface ocean d13C.When the 13C-depleted organic matter is decomposed, the
accompanying CO2 release decreases the d13C of dissolved
inorganic carbon at depth (Craig, 1970; Deuser and Hunt,
1969; Kroopnick, 1985). If this process were solely responsible
for the distribution of d13C in the ocean, d13C of oceanic
carbon would decrease by approximately 1.1% for every
1 mmol kg�1 increase in oceanic PO4 (Broecker and Maier-
Reimer, 1992). Reflecting this biological cycling, as a deep
water mass with a homogeneous source ages (such as in the
deep Indian and Pacific oceans), the d13C and PO4 increase in
this biologically expected ratio (Figure 1). However, the dis-
crimination against 13C during photosynthesis increases from
about 19% in the warm surface ocean to about 30% in the
Antarctic, which means that the relative changes in PO4 and
d13C associated with the uptake and regeneration of organic
material can vary regionally.
Carbon isotope fractionation during air–sea exchange is
also an important factor in determining the isotopic composi-
tion of seawater. If the CO2 in the atmosphere were in isotopic
equilibrium with the dissolved inorganic carbon in the ocean,
the dissolved inorganic carbon would be enriched in 13C rela-
tive to atmospheric CO2 by about 8% at 20 �C (Inoue and
Sugimura, 1985). This equilibrium fractionation depends on
WSBW
CDW
2.5mol kg−1)
d 13Cas =−0.5
s =0.5
2.0 3.0
. The lines indicate a ‘Redfield slope,’ and d13Cas indicates the deviationeimer, 1992). While the deep Indian and Pacific values follow thetlantic and Southern Ocean have lower and higher d13C due to air–seaFairbanks RG (1995) The influence of air–sea exchange on the isotopicemical Cycles 9: 653–665.
−0.40−0.40 1.20
Estimated seawater d13C (‰ PDB)
d13C
rec
onst
ruct
ed fr
om fo
ram
inife
ra (‰
PD
B)
0.800.400.00 1.60
1.20
0.80
0.40
0.00
1.60
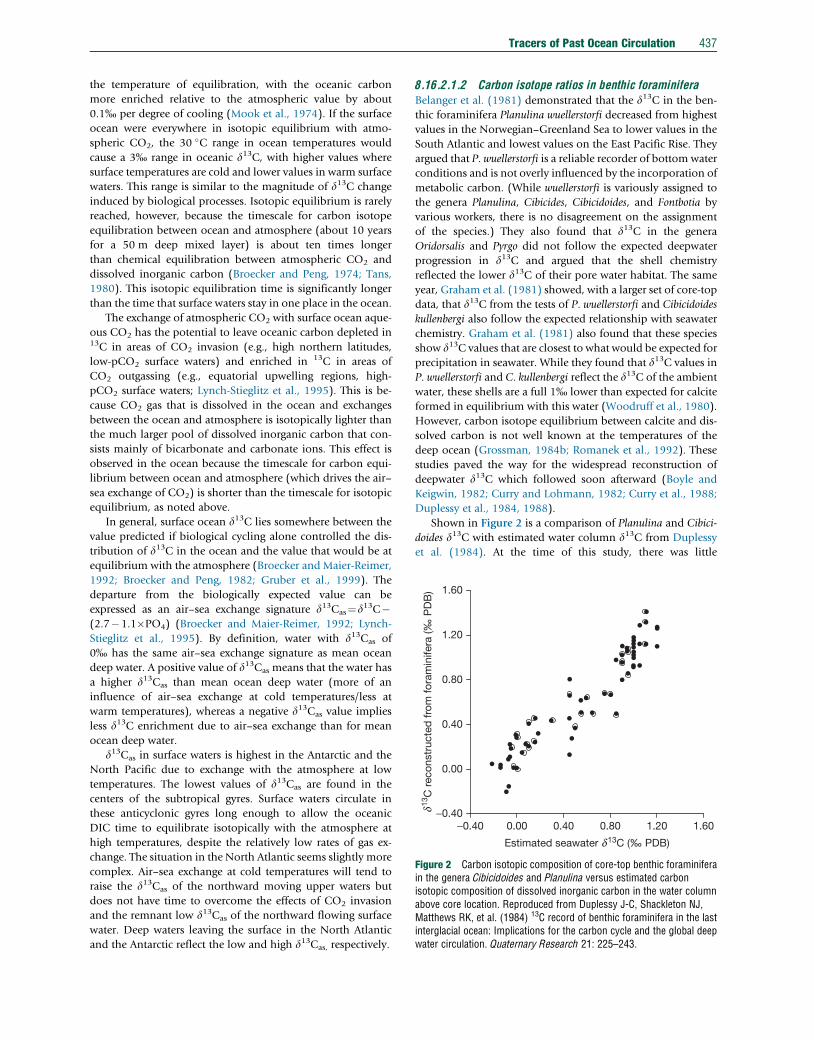
Figure 2 Carbon isotopic composition of core-top benthic foraminiferain the genera Cibicidoides and Planulina versus estimated carbonisotopic composition of dissolved inorganic carbon in the water columnabove core location. Reproduced from Duplessy J-C, Shackleton NJ,Matthews RK, et al. (1984) 13C record of benthic foraminifera in the lastinterglacial ocean: Implications for the carbon cycle and the global deepwater circulation. Quaternary Research 21: 225–243.
Tracers of Past Ocean Circulation 437
the temperature of equilibration, with the oceanic carbon
more enriched relative to the atmospheric value by about
0.1% per degree of cooling (Mook et al., 1974). If the surface
ocean were everywhere in isotopic equilibrium with atmo-
spheric CO2, the 30 �C range in ocean temperatures would
cause a 3% range in oceanic d13C, with higher values where
surface temperatures are cold and lower values in warm surface
waters. This range is similar to the magnitude of d13C change
induced by biological processes. Isotopic equilibrium is rarely
reached, however, because the timescale for carbon isotope
equilibration between ocean and atmosphere (about 10 years
for a 50 m deep mixed layer) is about ten times longer
than chemical equilibration between atmospheric CO2 and
dissolved inorganic carbon (Broecker and Peng, 1974; Tans,
1980). This isotopic equilibration time is significantly longer
than the time that surface waters stay in one place in the ocean.
The exchange of atmospheric CO2 with surface ocean aque-
ous CO2 has the potential to leave oceanic carbon depleted in13C in areas of CO2 invasion (e.g., high northern latitudes,
low-pCO2 surface waters) and enriched in 13C in areas of
CO2 outgassing (e.g., equatorial upwelling regions, high-
pCO2 surface waters; Lynch-Stieglitz et al., 1995). This is be-
cause CO2 gas that is dissolved in the ocean and exchanges
between the ocean and atmosphere is isotopically lighter than
the much larger pool of dissolved inorganic carbon that con-
sists mainly of bicarbonate and carbonate ions. This effect is
observed in the ocean because the timescale for carbon equi-
librium between ocean and atmosphere (which drives the air–
sea exchange of CO2) is shorter than the timescale for isotopic
equilibrium, as noted above.
In general, surface ocean d13C lies somewhere between the
value predicted if biological cycling alone controlled the dis-
tribution of d13C in the ocean and the value that would be at
equilibrium with the atmosphere (Broecker and Maier-Reimer,
1992; Broecker and Peng, 1982; Gruber et al., 1999). The
departure from the biologically expected value can be
expressed as an air–sea exchange signature d13Cas¼d13C�(2.7�1.1�PO4) (Broecker and Maier-Reimer, 1992; Lynch-
Stieglitz et al., 1995). By definition, water with d13Cas of
0% has the same air–sea exchange signature as mean ocean
deep water. A positive value of d13Cas means that the water has
a higher d13Cas than mean ocean deep water (more of an
influence of air–sea exchange at cold temperatures/less at
warm temperatures), whereas a negative d13Cas value implies
less d13C enrichment due to air–sea exchange than for mean
ocean deep water.
d13Cas in surface waters is highest in the Antarctic and the
North Pacific due to exchange with the atmosphere at low
temperatures. The lowest values of d13Cas are found in the
centers of the subtropical gyres. Surface waters circulate in
these anticyclonic gyres long enough to allow the oceanic
DIC time to equilibrate isotopically with the atmosphere at
high temperatures, despite the relatively low rates of gas ex-
change. The situation in the North Atlantic seems slightly more
complex. Air–sea exchange at cold temperatures will tend to
raise the d13Cas of the northward moving upper waters but
does not have time to overcome the effects of CO2 invasion
and the remnant low d13Cas of the northward flowing surface
water. Deep waters leaving the surface in the North Atlantic
and the Antarctic reflect the low and high d13Cas, respectively.
8.16.2.1.2 Carbon isotope ratios in benthic foraminiferaBelanger et al. (1981) demonstrated that the d13C in the ben-
thic foraminifera Planulina wuellerstorfi decreased from highest
values in the Norwegian–Greenland Sea to lower values in the
South Atlantic and lowest values on the East Pacific Rise. They
argued that P. wuellerstorfi is a reliable recorder of bottom water
conditions and is not overly influenced by the incorporation of
metabolic carbon. (While wuellerstorfi is variously assigned to
the genera Planulina, Cibicides, Cibicidoides, and Fontbotia by
various workers, there is no disagreement on the assignment
of the species.) They also found that d13C in the genera
Oridorsalis and Pyrgo did not follow the expected deepwater
progression in d13C and argued that the shell chemistry
reflected the lower d13C of their pore water habitat. The same
year, Graham et al. (1981) showed, with a larger set of core-top
data, that d13C from the tests of P. wuellerstorfi and Cibicidoides
kullenbergi also follow the expected relationship with seawater
chemistry. Graham et al. (1981) also found that these species
show d13C values that are closest to what would be expected for
precipitation in seawater. While they found that d13C values in
P. wuellerstorfi and C. kullenbergi reflect the d13C of the ambient
water, these shells are a full 1% lower than expected for calcite
formed in equilibrium with this water (Woodruff et al., 1980).
However, carbon isotope equilibrium between calcite and dis-
solved carbon is not well known at the temperatures of the
deep ocean (Grossman, 1984b; Romanek et al., 1992). These
studies paved the way for the widespread reconstruction of
deepwater d13C which followed soon afterward (Boyle and
Keigwin, 1982; Curry and Lohmann, 1982; Curry et al., 1988;
Duplessy et al., 1984, 1988).
Shown in Figure 2 is a comparison of Planulina and Cibici-
doides d13C with estimated water column d13C from Duplessy
et al. (1984). At the time of this study, there was little

438 Tracers of Past Ocean Circulation
high-quality water column d13C data with which to compare
the core-top values. In subsequent years, however, more exten-
sive and improved datasets have confirmed that, on average,
Planulina and Cibicidoides d13C reflect water column d13C with
little or no offset. Some of the scatter in Figure 2 may be
attributed to errors in estimated oceanic d13C or core tops
which are not Holocene in age. However, the more recent
datasets also show that there can be significant offsets between
the foraminifera and water column values at some locations,
even when living foraminifera are compared to water column
d13C collected at the same location.
Perhaps it is not unexpected that benthic foraminifera do
not always cleanly record the isotopic composition of bottom
water. The fact that all of the benthic foraminifera have d13Cthat is significantly lighter than what would be expected
for equilibrium with bottom water provides the first clue that
additional processes may be at work. As postulated by Belanger
et al. (1981), some foraminifera may live in the pore waters of
the sediment, where d13C of dissolved inorganic carbon is
lower than in the overlying water column. Even so, the carbon
isotopic composition of the calcite skeleton of most calcitic
organisms does not reflect the composition of the ambient
waters (Wefer and Berger, 1991) but is dominated by physio-
logical or ‘vital’ effects which usually involve the incorporation
of isotopically light metabolic carbon or kinetic fractionation
during the transport of carbon to the calcification site or during
calcification itself. The isotopic ratios of planktonic foraminif-
era are also dominated by these vital effects and are, in general,
not in equilibrium with seawater (e.g., Spero and Lea, 1996;
Spero et al., 1997).
A more detailed look at how benthic foraminifera record
bottom water d13C has been made through careful analysis of
stained (live or recently living) benthic foraminifera collected
from surface sediment, combined with measurements of
both water column and pore water d13C (Grossman, 1984a,b;
Mackensen and Douglas, 1989; Mackensen et al., 1993;
McCorkle et al., 1990, 1997; Rathburn et al., 1996; Tachikawa
and Elderfield, 2002). These studies confirm earlier notions
that different species show different microhabitat preferences,
with some species (including most Cibicidoides and Planulina)
living on or near the sediment surface and others (including
Uvigerina) living within the sediment. The deeper-dwelling
species show depleted d13C values relative to bottom waters
that, in general, reflect the magnitude of the pore water de-
pletions. These studies suggest that this microhabitat effect is
the dominant control on benthic foraminiferal d13C and that
variations in the vital effects between species are small. While
the number of Cibicidoides analyzed has been small, the Cibici-
doides species tend to have the smallest isotopic depletion
relative to bottom waters.
However, the isotopic depletion even for Cibicidoides can be
significant, especially in areas where productivity, and thus the
supply of organic material to the sediment, is high. Mackensen
et al. (1993) showed that living and dead specimens of
P. wuellerstorfi showed substantial depletions relative to bot-
tom waters, particularly beneath high-productivity fronts in
the Southern Ocean. While P. wuellerstorfi has been reported
to live above the sediment surface (Lutze and Thiel, 1989),
Mackensen et al. (1993) postulate that the foraminifera are
calcifying within the organic fluff layer that can be found in
high-productivity environments, with their low d13C value
reflecting either the low-d13C environment within the fluff
layer or high growth rates of the foraminifera within these
environments. Subsequent studies, however, have shown that
the relationship between d13C and overlying productivity can
be complex (Corliss et al., 2006; Eberwein and Mackensen,
2006; Zarriess and Mackensen, 2011).
These relationships have been generally confirmed with
downcore d13C studies. These studies show that the d13Cfrom Uvigerina (infaunal) is always more negative than for
Cibicidoides and that this offset is variable in space and time,
consistent with the microhabitat model. While many early
isotopic studies were performed using Uvigerina because
it was thought to best reflect oxygen isotope equilibrium,
most deepwater d13C reconstructions now use Cibicidoides
or Planulina species. It is broadly acknowledged that these
records, particularly in high-productivity areas, might be influ-
enced by a microhabitat effect. However, by avoiding high-
productivity areas and using supplemental information about
changes in bottom water characteristics and overlying produc-
tivity, this method has continued to yield regionally coherent
and sensible patterns of deepwater d13C.
8.16.2.2 Cadmium in Benthic Foraminifera
The use of Cd/Ca measurements in the tests of benthic forami-
nifera for deepwater nutrient reconstructions was developed in
parallel with the carbon isotope method. Dissolved Cd con-
centrations in seawater follow a nutrient-like distribution,
while Ca concentrations mainly reflect variations in salinity.
Benthic foraminifera incorporate Cd and Ca into their shells in
proportion to the ratio of these elements in seawater, which
allows for the reconstruction of deepwater Cd (and thus mac-
ronutrient) distributions.
8.16.2.2.1 Cd in seawaterCd in seawater has a nutrient-like profile, with depleted values
in warm surface waters and the most enriched concentrations
in intermediate and deep waters (Boyle et al., 1976). Many
high-quality Cd measurements in seawater over the last
20 years have confirmed this nutrient-like behavior, with Cd
showing a tight correlation with phosphorus in seawater.
While it is still not clear whether most Cd is taken up as an
essential micronutrient or inadvertently, the cycling within the
ocean is dominated by incorporation into organic tissue in the
surface ocean and regeneration of elemental Cd as this material
is respired below the surface. The relationship with phospho-
rus is not linear, with a lower slope at lower phosphorus
concentrations (Figure 3). This curvature may be due to pref-
erential uptake of Cd over P by phytoplankton, though not at a
fixed (‘Redfield’) ratio (Elderfield and Rickaby, 2000). As dis-
solved phosphorus is drawn down in the sunlit surface ocean,
the Cd/P ratio of seawater decreases. When high-Cd/P organic
matter is remineralized below the surface, the Cd/P of seawater
climbs again, and the net result is a curve whose shape depends
on the effective fractionation factor between organic matter
Cd/P and seawater Cd/P. Since different deep waters have
undergone different extents of remineralization, and may be
mixtures of water masses with different preformed Cd/P ratios,
there are real differences in the Cd/P relationships in different

0.0
1.0
0.8
0.6
0.4
0.2
1.2
0.0 1.0
Estimated seawater Cd (nmol kg−1)
Cd
rec
onst
ruct
ed fr
om fo
ram
inife
ra (n
mol
kg−1
)
0.80.60.40.2 1.2
Figure 4 Seawater Cd reconstructed from foraminifera using adepth-dependent empirical distribution coefficient versus estimatedbottom water Cd. Data are from Boyle (1988, 1992).
1.2
1
0.8
0.6
0.4
Cd
SW
(nm
olkg
−1)
0.2
00.0 2.41.60.8
PSW (μmol kg−1)3.2
1.4
Figure 3 Seawater Cd measurements from the modern ocean plottedversus PO4. The curvature in this relationship is modeled by Elderfieldand Rickaby (2000) (solid line) as resulting from the preferential uptakeof Cd into marine organic matter. Reproduced from Elderfield H andRickaby REM (2000) Oceanic Cd/P ratio and nutrient utilization in theglacial Southern Ocean. Nature 405: 305–310.
Tracers of Past Ocean Circulation 439
ocean regions (Elderfield and Rickaby, 2000), which may of
course change through time. Because Cd is also removed from
seawater via CdS precipitation in suboxic sediments, the global
seawater Cd inventory, and thus the global Cd/P relationship,
can change as the extent of suboxic sediments changes
(Rosenthal et al., 1995a,b; van Geen et al., 1995). However,
the residence time for Cd in seawater is sufficiently long, and
the expansion of deep-sea suboxia sufficiently small, that the
Cd concentration during the last glacial period is unlikely to
have been lower than today by more than 5% (Boyle and
Rosenthal, 1996). This allows us to make meaningful infer-
ences about the cycling of major nutrients via reconstructions
of the past Cd concentration in seawater.
8.16.2.2.2 Cd/Ca in foraminiferaBoyle (1981) showed that with careful attention to cleaning,
benthic foraminiferal shells reflect the concentration of some
trace metals (including Cd) in seawater, with a preferential
incorporation of Cd over Ca into foraminifera. The empirical
distribution coefficient (D, ratio of Cd/Ca in foraminifera to
Cd/Ca in seawater) was shown to be 2.9, based primarily on
deep-sea core tops (Boyle, 1988). More core-topmeasurements
at all water depths have subsequently shown that this distribu-
tion coefficient increases with depth, starting at �1.3 above
1 km and increasing to a maximum of �2.9 below 3 km
(Boyle, 1992). Recent measurements suggest that the shallow
water value may be as high as 1.7 and is not discernibly
temperature dependent (Bryan and Marchitto, 2010; March-
itto, 2004). Once the depth-dependent distribution coefficient
is applied, the reconstructed Cd from foraminiferal tests shows
a linear relationship to Cd estimated for the overlying water
(Figure 4). Boyle et al. (1995) showed that the aragonitic
foraminifer Hoeglundina elegans has a D of 1 that shows
no depth dependence. McCorkle et al. (1995) observed an
unexpected decrease in the apparent distribution coefficient
of Cd and other trace metals with depth between 2.5 and
4 km in the deep Pacific, which suggested dissolution-driven
loss of Cd from the foraminiferal calcite after burial. Elderfield
et al. (1996) favored the idea that the lower D in deep Pacific
waters is a calcification response to the low carbonate satura-
tion state, rather than representing postdepositional dis-
solution, a possibility acknowledged by McCorkle et al.
(1995) and supported by laboratory dissolution experiments
(Marchitto et al., 2000). There has been no systematic attempt
to correct paleoceanographic Cd reconstructions for this
apparent low carbonate saturation state effect, though Zn/Ca
(Marchitto et al., 2000) and B/Ca (Yu and Elderfield, 2007)
hold some promise in this regard.
To the extent that the d13C of infaunal benthic foraminifera
reflects the low d13C of pore waters, it would be expected that
infaunal Cd/Ca also reflects the interstitial rather than the
bottom water concentrations. However, while interstitial d13Cvalues decrease with increasing depth in the sediment due to
the regeneration of isotopically light organic matter, the distri-
bution of Cd in pore water is more complex. In shallow pore
waters, Cd increases due to regeneration of organic material
but in suboxic sediments Cd decreases due to precipitation of
CdS (McCorkle and Klinkhammer, 1991; Rosenthal et al.,
1995a,b). As a consequence, we would not necessarily expect
a systematic relationship between the Cd/Ca of epifaunal
versus infaunal foraminifera as there is for d13C. Tachikawaand Elderfield (2002) showed that for three sites in the North
Atlantic, pore waters were 2–4 times enriched in Cd relative to
bottom waters. They found that P. wuellerstorfi, whose d13Cvalues suggested that they calcified in bottom waters at the
sediment surface, had a D of 3–4, consistent with culture
studies (Havach et al., 2001). The infaunal species showed

440 Tracers of Past Ocean Circulation
a lower D of about 1 with respect to the pore water Cd con-
centrations. The net effect is that the Cd/Ca in P. wuellerstorfi
and the infaunal species is quite similar, consistent with previ-
ous observations across a range of conditions (e.g., Boyle,
1992). Further, Tachikawa and Elderfield argue that the
depth-dependent D for the infaunal Uvigerina may actually
reflect a trend in the enrichment of pore water Cd relative to
bottom water Cd, though this cannot explain the depth depen-
dence of D in the epifaunal P. wuellerstorfi (Boyle, 1992).
Regardless of the details surrounding infaunal D values, it is
expected that Cd/Ca in P. wuellerstorfi or other surface dwelling
foraminifera should give a more direct measure of bottom
water Cd. As with d13C, careful interpretation aided by consid-
eration of supplemental information about changes in bottom
water characteristics has yielded regionally coherent and sensi-
ble patterns of deepwater Cd (e.g., Boyle, 1992; Marchitto and
Broecker, 2006).
8.16.2.3 Ba/Ca
Like Cd, dissolved Ba in a broad sense cycles like a nutrient,
depleted in surface waters and higher in deep waters, but the
surface depletion is moderate, and the regeneration of Ba
occurs deeper than for labile nutrients like phosphorus. This
pattern is due to the removal of Ba by barite precipitation in
decaying organic matter and subsequent barite dissolution in
the deep sea. This process coincidently results in a close corre-
lation between Ba and alkalinity in today’s ocean (Lea, 1993).
Lea and Boyle (1989) showed that the barium content in
benthic foraminiferal shells is controlled by the barium con-
centration in bottom waters, with a D of �0.4. Ba/Ca in
foraminifera has been used, like Cd/Ca, as a paleotracer of
water masses (e.g., Lea and Boyle, 1990) and appears to suffer
from the same carbonate saturation state-linked effect as
Cd/Ca (McCorkle et al., 1995).
8.16.2.4 Zn/Ca
Zn is a necessary micronutrient for phytoplankton and espe-
cially diatom growth. This requirement results in a very strong
correlation between oceanic dissolved Zn and Si concentrations
(Bruland et al., 1978). Benthic foraminiferal Zn/Ca is controlled
by bottom water dissolved Zn concentration, with a maximum
D in P. wuellerstorfi and Uvigerina of �9 (Marchitto et al., 2000)
and a value closer to 22 in C. pachyderma (Bryan and Marchitto,
2010). Because of the close relationship between Si and Zn,
Zn/Ca in foraminifera is particularly useful for tracing the
Si-rich bottom water that originates in the Southern Ocean
(Marchitto et al., 2002). Like Cd/Ca and Ba/Ca, the apparent
distribution coefficient for Zn decreases in waters which are
more undersaturated with respect to calcium carbonate. Because
they respond to saturation state differently, Cd/Ca and Zn/Ca
can theoretically be used together to assess both the nutrient
content and carbonate ion concentrations in deep water masses
(Marchitto et al., 2000, 2002).
8.16.2.5 Silicon Isotopes
A promising new proxy for the reconstruction of deepwater
silicic acid concentrations is the isotopic composition of Si in
sponge spicules. Hendry et al. (2010) have shown that the
fractionation of Si isotopes increases with ambient Si(OH)4concentration. This effect is thought to be related to the growth
rate of the sponge spicules, which should respond to the
Si(OH)4 concentration.
8.16.3 Conservative Water Mass Tracers
In general, nutrient-type tracers have been used to infer the
sources and mixing of deep water masses in the past, despite
the complication that the nutrient content of deep water de-
pends both on the nutrient content at its source and the
addition of nutrients via regeneration of organic matter as the
water mass ages. The utility of this method is primarily due to
the large and readily measured contrasts in the nutrient-related
tracers. Headway is now being made in the use of conservative
tracers that reflect the mixing of source waters alone (such as
temperature and salinity in the modern ocean).
8.16.3.1 Mg/Ca in Benthic Foraminifera
Mg/Ca ratios in benthic foraminifera reflect the calcification
temperature (e.g., Martin et al., 2002; Rathburn and
deDeckker, 1997; Rosenthal et al., 1997b). In principle, tem-
perature can be used as a conservative tracer for subsurface
water masses, but at present, the uncertainties in converting
Mg/Ca to temperature overwhelm the deep water temperature
gradients that distinguish water masses of different origins. In
particular, the saturation state of bottom water appears to exert
a strong secondary influence on Mg/Ca, as it does for the other
trace metal ratios (Elderfield et al., 2006). Emerging research is
targeting additional ratios like Mg/Li (Bryan and Marchitto,
2008; Lear et al., 2010) and infaunal taxa like Uvigerina
(Elderfield et al., 2010) in the hope of isolating or minimizing
the carbonate ion effect so that Mg/Ca could be used to assess
the contributions to intermediate water masses, where the
temperature signals are more easily distinguished.
8.16.3.2 Pore Water Chemistry
The d18O of seawater is set in the short term by processes which
occur at the ocean surface (evaporation, precipitation, sea ice
formation and melt; Craig and Gordon, 1965), so it can be
used as a conservative tracer in the subsurface ocean. The d18Oof glacial-age water has been reconstructed by measuring the
d18O of pore water in deep-sea sediment (e.g., Schrag and
dePaolo, 1993; Schrag et al., 2002). The maximum contribu-
tion of high d18O from the LGM appears at a depth of 20–60 m
within the sediments. The actual d18O of the glacial-age deep
water must be reconstructed by modeling the diffusion of
water within the sediments. Estimates for the bottom water
temperature (also a conservative tracer) at the LGM can then be
made by using the benthic foraminiferal d18O along with
information from pore waters about the d18O of the water in
which it calcified. Similarly, salinity can be reconstructed using
pore water [Cl] (Adkins and Schrag, 2001). While the number
of locations with such data is limited, Adkins et al. (2002)
show the promise of using pore water d18O and [Cl] to
Tracers of Past Ocean Circulation 441
reconstruct the properties of deep water masses. As more data
are collected, these tracers will also be useful for reconstructing
the sources and mixing of deep water masses.
8.16.3.3 Oxygen Isotopes in Benthic Foraminifera
The d18O of foraminiferal calcite reflects the d18O of the water
in which it calcified and the temperature of calcification. Since
both temperature and d18O of seawater are imprinted on a
water mass at the surface, the d18O of foraminifera is a conser-
vative tracer that can be used to determine the sources and
mixing of water masses in the deep ocean. The use of d18O as a
conservative tracer does not require the separation of the tem-
perature and d18O/salinity components of the foraminiferal
d18O signal. Despite the fact that oxygen isotope measure-
ments are routinely gathered simultaneously with carbon iso-
topes, the use of d18O as a tracer for ocean circulation has
lagged behind the use of d13C. Zahn and Mix (1991) showed
that gradients in deep water foraminiferal d18O are small due
to the fact that NADW is warmer than Antarctic Bottom Water
(which would lead to lower foraminiferal d18O), while d18O of
NADW is higher (counteracting the temperature effect). They
also demonstrated that there are significant issues with inter-
laboratory and interspecies calibration that interfere with the
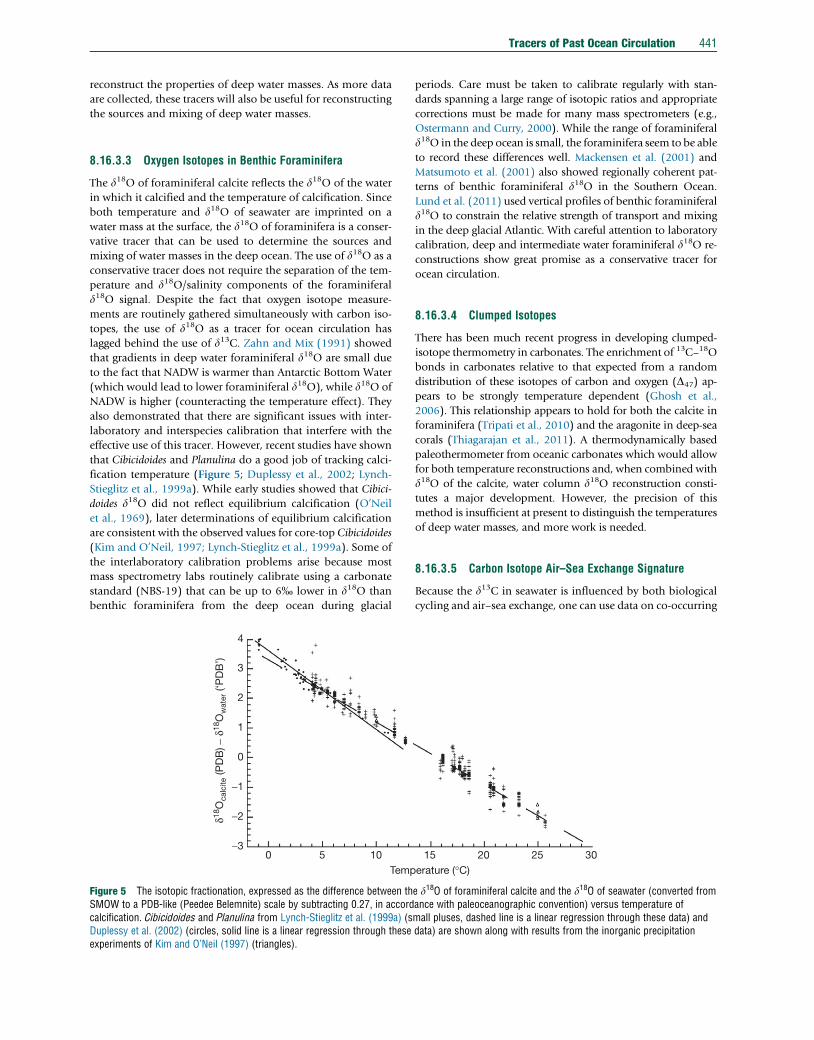
effective use of this tracer. However, recent studies have shown
that Cibicidoides and Planulina do a good job of tracking calci-
fication temperature (Figure 5; Duplessy et al., 2002; Lynch-
Stieglitz et al., 1999a). While early studies showed that Cibici-
doides d18O did not reflect equilibrium calcification (O’Neil
et al., 1969), later determinations of equilibrium calcification
are consistent with the observed values for core-top Cibicidoides
(Kim and O’Neil, 1997; Lynch-Stieglitz et al., 1999a). Some of
the interlaboratory calibration problems arise because most
mass spectrometry labs routinely calibrate using a carbonate
standard (NBS-19) that can be up to 6% lower in d18O than
benthic foraminifera from the deep ocean during glacial
3
2
1
0
0Temp
δ18O
calc
ite (P
DB
) − δ
18O
wat
er (‘
PD
B’)
105
−1
−2
−3
4
Figure 5 The isotopic fractionation, expressed as the difference between thSMOW to a PDB-like (Peedee Belemnite) scale by subtracting 0.27, in accordcalcification. Cibicidoides and Planulina from Lynch-Stieglitz et al. (1999a) (sDuplessy et al. (2002) (circles, solid line is a linear regression through theseexperiments of Kim and O’Neil (1997) (triangles).
periods. Care must be taken to calibrate regularly with stan-
dards spanning a large range of isotopic ratios and appropriate
corrections must be made for many mass spectrometers (e.g.,
Ostermann and Curry, 2000). While the range of foraminiferal
d18O in the deep ocean is small, the foraminifera seem to be able
to record these differences well. Mackensen et al. (2001) and
Matsumoto et al. (2001) also showed regionally coherent pat-
terns of benthic foraminiferal d18O in the Southern Ocean.
Lund et al. (2011) used vertical profiles of benthic foraminiferal
d18O to constrain the relative strength of transport and mixing
in the deep glacial Atlantic. With careful attention to laboratory
calibration, deep and intermediate water foraminiferal d18O re-
constructions show great promise as a conservative tracer for
ocean circulation.
8.16.3.4 Clumped Isotopes
There has been much recent progress in developing clumped-
isotope thermometry in carbonates. The enrichment of 13C–18O
bonds in carbonates relative to that expected from a random
distribution of these isotopes of carbon and oxygen (D47) ap-
pears to be strongly temperature dependent (Ghosh et al.,
2006). This relationship appears to hold for both the calcite in
foraminifera (Tripati et al., 2010) and the aragonite in deep-sea
corals (Thiagarajan et al., 2011). A thermodynamically based
paleothermometer from oceanic carbonates which would allow
for both temperature reconstructions and, when combined with
d18O of the calcite, water column d18O reconstruction consti-
tutes a major development. However, the precision of this
method is insufficient at present to distinguish the temperatures
of deep water masses, and more work is needed.
8.16.3.5 Carbon Isotope Air–Sea Exchange Signature
Because the d13C in seawater is influenced by both biological
cycling and air–sea exchange, one can use data on co-occurring
252015erature (°C)
30
e d18O of foraminiferal calcite and the d18O of seawater (converted fromance with paleoceanographic convention) versus temperature ofmall pluses, dashed line is a linear regression through these data) anddata) are shown along with results from the inorganic precipitation

442 Tracers of Past Ocean Circulation
nutrients (which are influenced only by biological cycling) to
isolate d13C differences that arise from air–sea exchange
(Lynch-Stieglitz and Fairbanks, 1994; Oppo and Horowitz,
2000; Rickaby and Elderfield, 2005; Zahn and Stuber, 2002).
Because the air–sea exchange signature is reset only at the
surface, it is a conservative tracer for ocean circulation. Today,
newly formed NADW and Weddell Sea Bottom Water have
d13C air–sea exchange signatures that differ by 0.8% (Lynch-
Stieglitz et al., 1995). However, the patterns in the air–sea
exchange signature are only as good as the contributing d13Cand Cd/Ca data, and areas where either dissolution or produc-
tivity overprints may be a problem must be avoided, limiting
the use of this tracer.
8.16.4 Neodymium Isotope Ratios
Deep water masses of the ocean have distinctive neodymium
isotope ratios that are recorded in manganese nodules
(Albarede and Goldstein, 1992; Albarede et al., 1997), dis-
persed manganese–iron oxides in deep-sea sediment (e.g.,
Gutjahr et al., 2007; Rutberg et al., 2000), and iron oxides on
fish teeth (Martin and Haley, 2000) and in foraminifera (e.g.,
Klevenz et al., 2008; Vance et al., 2004). Seawater is thought to
acquire its Nd isotope signal at the ocean boundaries, via
surface inputs (dust and riverine) and exchange with sediment
(Frank, 2002; Goldstein and Hemming, 2003; Lacan and
Jeandel, 2005). Away from the boundaries, especially in the
deep Atlantic where the residence time of water is short
compared to the residence time of Nd, the isotopes of this
element can serve as a quasiconservative tracer (Piepgras and
Wasserburg, 1980). Nd isotope ratios in NADW are low,
reflecting the addition of Nd from continentally derived detri-
tal material, which is predominantly old, whereas Nd isotope
ratios in the North Pacific are high, reflecting input from recent
volcanic activity. In regions where there is active mixing be-
tween these water masses (the Atlantic Ocean and the Southern
Ocean), both seawater and the Fe–Mn oxyhydroxides that
precipitate from it have intermediate values, as is the case for
the nutrient and conservative water mass tracers.
8.16.5 Circulation Rate Tracers
The water mass tracers discussed above can be used to assess the
sources of subsurface water masses and the mixture of waters
from different sources. However, this information can be used
only to assess the relative rates of renewal of these deep water
masses from the various source regions (e.g., Legrand and
Wunsch, 1995). There are several geochemical techniques that
can be used to assess the rates of ocean circulation.
8.16.5.1 Radiocarbon
Carbon-14 (radiocarbon) is produced in the atmosphere by
cosmic rays and decays with a half-life of 5730 years. Natural
radiocarbon levels in the ocean are highest at the surface where
they partially equilibrate with atmosphere. Deepwater concen-
trations are highest in newly formed NADW and decrease to
the lowest values in the deep Pacific where the deep waters
have been out of contact with the atmosphere for longest
(Stuiver et al., 1983). Benthic foraminifera record the radiocar-
bon concentrations of the deep water in which they grew, but
the 14C in the foraminifera decays over time, with >99% gone
after 40000 years. In order to reconstruct the seawater radio-
carbon content at the time the foraminifera were living, radio-
carbon measurements can be made on benthic and planktonic
foraminifera from the same interval in the core (Broecker et al.,
1988, 1990; Shackleton et al., 1988) in order to determine the
age of the benthic foraminifera. However, the radiocarbon
content of the planktonic foraminifera will reflect not only
their age but also the initial radiocarbon content of the near-
surface water in which they grew. The initial radiocarbon in
surface and near-surface waters can be significantly less than
expected for equilibrium with the atmosphere due to the rela-
tively long time required for carbon isotopic equilibrium be-
tween the surface ocean and atmosphere. One must also
account for the fact that the radiocarbon content in the atmo-
sphere changes with time, a change which impacts the initial
radiocarbon content of surface waters as well. This means that
deep waters at a given locationmay have initially formed under
a different atmospheric radiocarbon burden than that recorded
by codeposited planktonic foraminifera (Adkins and Boyle,
1997).
Also complicating the use of concurrent benthic and
planktonic radiocarbon concentrations to assess water mass
radiocarbon is the necessary assumption that benthic and
planktonic foraminifera from the same depth interval are
actually the same age. If the peak abundances of planktonic
and benthic foraminifera occur at different depths in a core,
bioturbation can introduce a spread in the planktonic–benthic
ages in cores with low accumulation rates, which is unrelated
to changes in bottom water age (Broecker et al., 1999; Peng
et al., 1977). While these problems can be overcome by using
high-sedimentation-rate cores (e.g., Keigwin and Schlegel,
2002), the widespread use of this technique has been limited
primarily by the identification of suitable cores with high
enough foraminiferal abundances for accurate radiocarbon
determinations. Sediments with high accumulation rates tend
either to have low foraminiferal abundances (due to dilution
by terrigenous material) or to be in high-productivity regions
of the ocean where surface waters are expected to be far from
equilibrium with the atmosphere and variable through time.
Another approach to constraining the age required to ex-
tract seawater 14C content from benthic foraminifera is to use
stratigraphic correlation. Numerous cores with high accum-
ulation rates record seawater characteristics such as surface
temperature or oxygen content that vary in concert with either
Greenland or Antarctic air temperatures, as recorded by ice
cores. Assuming the oceanic changes are nearly synchronous
with the polar records, the ice core chronologies (which are
independent of 14C) can be applied to the sediments and
therefore to the benthic foraminifera (Skinner and Shackleton,
2004). This approach is mainly limited by the quality of the
correlation and the assumption of synchroneity.
A third way to measure the deepwater radiocarbon ventila-
tion age in the past is by using deep-dwelling benthic corals
(Adkins et al., 1998; Goldstein et al., 2001; Mangini et al.,
1998). Here, the age of the coral can be independently deter-
mined using uranium series dating, allowing the radiocarbon

Tracers of Past Ocean Circulation 443
content of the coral to be used to determine the radiocarbon
content of deep water at the time the coral grew. This method is
unaffected by bioturbation but is currently limited by the
availability of deep-sea benthic corals.
Changes in the ventilation of the ocean can also be assessed
by looking at how the 14C content of the atmosphere has
changed through time (e.g., Hughen et al., 1998). The atmo-
spheric 14C content is controlled both by the rate of 14C pro-
duction in the atmosphere from cosmic ray bombardment and
the rate at which 14C is transferred into the ocean. While a
decrease in the ventilation of the ocean causes a buildup of
radiocarbon in the atmosphere, one must also account for
changes in production (believed to be dominated by changes
in geomagnetic field strength) before interpreting the atmo-
spheric radiocarbon changes in terms of ocean circulation
(Hughen et al., 1998; Lal and Charles, 2007).
8.16.5.2 231Pa/230Th Ratio
Both protactinium and thorium are produced in the water
column and scavenged by particles onto the seafloor. Because
protactinium is scavenged less efficiently by marine particles
(Anderson et al., 1983), much of the protactinium produced
in the Atlantic Ocean is transported to the Southern Ocean by
deep waters before it is scavenged onto the seafloor. This
produces a low (below production) 231Pa/230Th ratio in
Atlantic Ocean sediments and a high 231Pa/230Th ratio in
Southern Ocean sediments (Yu et al., 1996). Reconstruction
of the 231Pa/230Th ratio suggests that this pattern of Pa export
persisted in glacial times, and a low deepwater residence time
for the LGM Atlantic has been inferred (Yu et al., 1996). Time
series of Pa/Th from the North Atlantic have been used to infer
changes in circulation over the deglaciation (Gherardi et al.,
2009; McManus et al., 2004).
However, the 231Pa/230Th ratio in sediments depends on
the complex interplay between deep water movement and the
differential scavenging of the two elements throughout the
ocean basin (Marchal et al., 2001; Siddall et al., 2007). The
scavenging of Pa and Th depends both on the composition of
the particles and the particle flux. Biogenic opal fractionates Th
from Pa much less effectively than other particle types; as a
consequence, ratios in Southern Ocean sediments will be very
sensitive to particle composition (Luo and Ku, 1999; Yu et al.,
2001). While the Atlantic 231Pa/230Th fluxes are low and today
predominantly reflect the short residence time of deep waters
in this basin, in the Pacific ocean, 231Pa/230Th ratios are most
sensitive to particle flux, with higher ratios where the scaveng-
ing is most intense at the boundaries (Yu et al., 2001). Robust
paleocirculation reconstructions from 231Pa/230Th ratios will,
in general, require detailed knowledge of particle fluxes and
composition (Gil et al., 2009; Lippold et al., 2009) as well as
good spatial coverage from a number of sediment cores (Burke
et al., 2011; Siddall et al., 2007).
8.16.5.3 Geostrophic Shear Estimates from d18O in BenthicForaminifera
Physical oceanographers routinely use the distribution of
density in the ocean to help quantify ocean circulation. A
balance between the pressure gradient force and Coriolis
term (geostrophic balance) is assumed. Velocity can then be
calculated from density information using either a measured or
assumed velocity at a reference level or by combining the
geostrophic balance with other constraints, such as conserva-
tion of tracer flow across a section. This approach has also been
applied to paleoceanography by using vertical density profiles
reconstructed from the oxygen isotopic composition of fora-
miniferal calcite (Lynch-Stieglitz et al., 1999a).
The d18O from calcite tests of benthic foraminifera preserved
in ocean sediments can be used to estimate upper ocean density
because both the d18O of calcite (d18Ocalcite) and seawater den-
sity increase with increasing salinity or decreasing temperature
(Lynch-Stieglitz et al., 1999a). For times in the geologic past, the
ability to reconstruct density from d18Ocalcite is most limited by
knowledge of the relationships between d18Owater and salinity as
well as the relationships between temperature and salinity. How-
ever, it is the gradients in density that are used for geostrophic
calculations, and the reconstructed density gradients are less
affected by these limitations than the density itself.
This technique has been used to reconstruct the flow of the
Gulf Stream in the Florida Straits by reconstructing vertical
density profiles on either side of this current (Lund et al.,
2006; Lynch-Stieglitz et al., 1999b, 2009, 2011). Perhaps more
useful for reconstruction of the large-scale ocean circulation
is the fact that the density difference between the eastern and
western margins of the ocean basins reflects the strength of
the geostrophic overturning circulation (Lynch-Stieglitz, 2001;
Lynch-Stieglitz et al., 2006).
At present, the vertical density structure for the past ocean
can be reconstructed using benthic foraminifera only where the
seafloor intersects the upper water column, at ocean margins,
islands, and shallow seamounts. Although planktonic forami-
nifera calcify at various depths within the upper water column,
we have no way to reconstruct quantitatively the depth at which
they calcified. However, the oxygen isotopic composition of
deep-dwelling planktonic foraminifera can be used to recon-
struct the spatial pattern and direction of upper ocean flows
(Bostock et al., 2006; LeGrande and Lynch-Stieglitz, 2007;
Matsumoto and Lynch-Stieglitz, 2003; Ortiz et al., 1997).
8.16.6 Nongeochemical Tracers of Past OceanCirculation
While this is a geochemistry volume, any review of tracers of past
ocean circulationwould be incomplete withoutmention of some
of the sedimentological andpaleontological approaches to recon-
structing past ocean flow. Sediment grain size has been used
to reconstruct current intensity since the 1970s (Ledbetter and
Johnson, 1976). More recent grain size studies emphasize the
mean size of the ‘sortable silt’ fraction (>10 mm), which is least
likely to display cohesive behavior (McCave et al., 1995). Stron-
ger currents show larger sortable silt size due to changes in
both deposition and winnowing with current speed. The mag-
netic properties of sediments (concentration of magnetic
particles) have been used to infer flow paths and strength of
the overflows in the North Atlantic (Ballini et al., 2006; Kissel
et al., 1999).
Benthic foraminiferal assemblages have been related to dis-
tinct deep water masses and have been used for tracking these
444 Tracers of Past Ocean Circulation
water masses during the past (Schnitker, 1974; Streeter, 1973).
In more recent years, it has been determined that the deep-sea
benthic assemblage responds strongly to the flux of organic
material from overlying surface productivity (e.g., Loubere,
1991; Miller and Lohmann, 1982), limiting its use as a water
mass tracer to low-productivity regions (Schnitker, 1994).
0
(a)
(b)
Dep
th (m
)
(c)
1000
2000
3000
4000
5000
6000
0
0
1000
2000
3000
4000
5000
1000
2000
3000
4000
5000
40°S EQ20°S
40°S EQ20°S
40°S EQ20°S
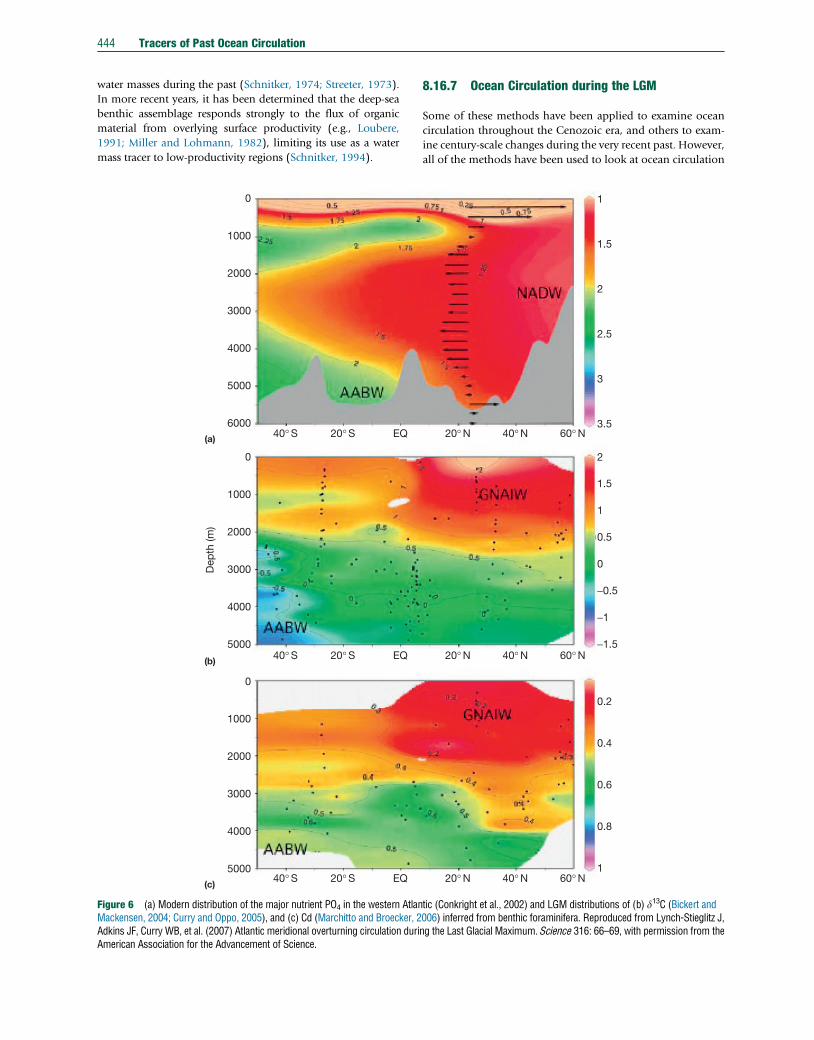
Figure 6 (a) Modern distribution of the major nutrient PO4 in the western AtlanMackensen, 2004; Curry and Oppo, 2005), and (c) Cd (Marchitto and Broecker, 2Adkins JF, Curry WB, et al. (2007) Atlantic meridional overturning circulation duriAmerican Association for the Advancement of Science.
8.16.7 Ocean Circulation during the LGM
Some of these methods have been applied to examine ocean
circulation throughout the Cenozoic era, and others to exam-
ine century-scale changes during the very recent past. However,
all of the methods have been used to look at ocean circulation
1
1.5
2
2
1.5
1
0.5
0
−0.5
−1
−1.5
0.2
0.4
0.6
0.8
1
2.5
3
3.540°N20°N 60°N
40°N20°N 60°N
40°N20°N 60°N
tic (Conkright et al., 2002) and LGM distributions of (b) d13C (Bickert and006) inferred from benthic foraminifera. Reproduced from Lynch-Stieglitz J,ng the Last Glacial Maximum. Science 316: 66–69, with permission from the
Tracers of Past Ocean Circulation 445
during the LGM (about 19000–23000 years ago) and thus
have their maximum geographic coverage during this time.
Here, the picture of ocean circulation that these techniques
have produced for the LGM is briefly examined. The various
paleoceanographic methods outlined above each have their
advantages and pitfalls, times and places where they should
work well, and environments where variables other than ocean
circulation complicate the interpretations. There is no single
ocean circulation scenario for the LGM that is consistent with
all of the available data from all of the tracers. In the following
description, regionally coherent signals are weighted more
heavily than data from isolated sediment cores. Data in regions
where the complications may overwhelm the signal (e.g., high-
productivity regions for d13C data, the low-carbonate very deep
ocean for Cd data) are weighted less heavily. A quantitative
description of past ocean circulation will require many more
data and better understanding of the tracers, combined with
better techniques for dealing with the kinds of uncertainties
and multiple processes that affect the paleoceanographic
proxies.
Past circulation in the Atlantic Ocean basin has been
reviewed by Lynch-Stieglitz et al. (2007). The most prominent
feature of the glacial Atlantic is a water mass divide at about
2-km water depth. Both Cd/Ca and d13C distributions suggest
a strong stratification in the North Atlantic Ocean, with a low
nutrient, high-d13C water mass (often called Glacial North
Atlantic Intermediate Water, GNAIW) occupying depths
down to about 2000 m and a high nutrient, low-d13C water
mass of southern origin underneath (Figure 6; e.g., Boyle and
Keigwin, 1987; Curry and Oppo, 2005; Duplessy et al., 1988;
Marchitto and Broecker, 2006; Oppo and Lehman, 1993). An
expansion of southern-source waters into the deep Atlantic is
supported by higher Nd isotope ratios (Roberts et al., 2010)
40°W2000
1500
1000
500
0−2 0−1
26
27
25.5
26.5
27.5
sθ (kg m3)d18O
0°20°W
Holocene
20°E
(a) (b)
Dep
th (m
)
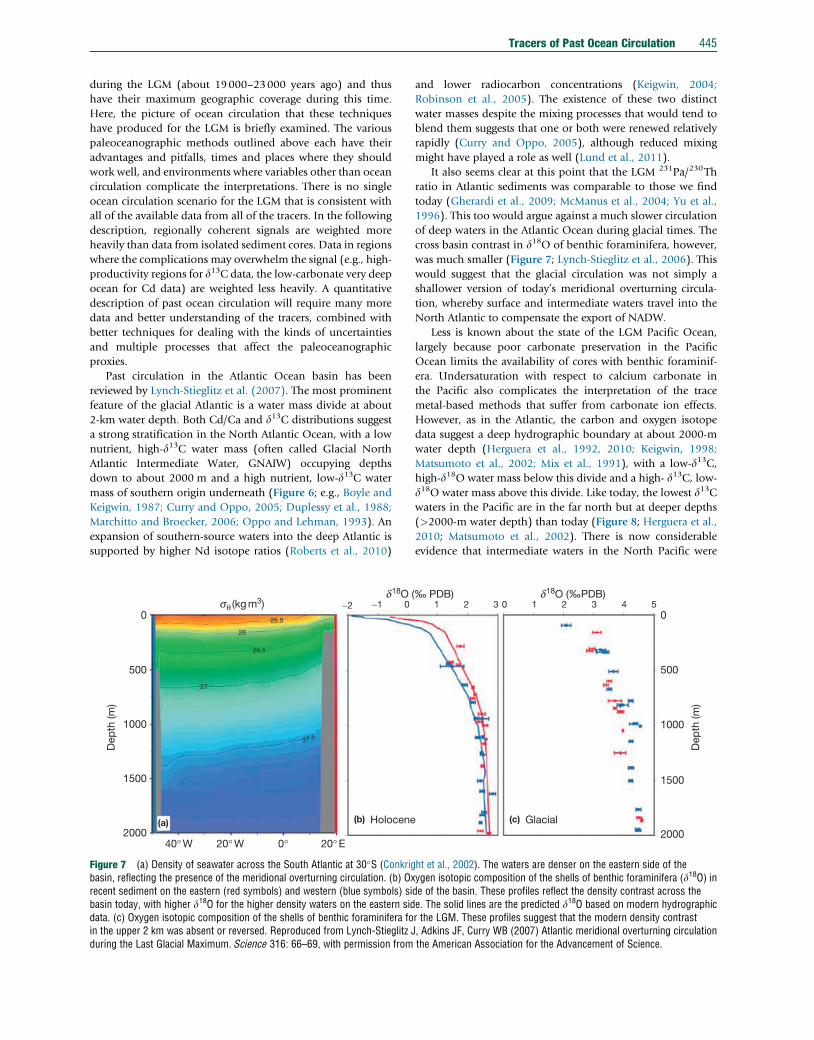
Figure 7 (a) Density of seawater across the South Atlantic at 30�S (Conkrigbasin, reflecting the presence of the meridional overturning circulation. (b) Oxrecent sediment on the eastern (red symbols) and western (blue symbols) sibasin today, with higher d18O for the higher density waters on the eastern siddata. (c) Oxygen isotopic composition of the shells of benthic foraminifera foin the upper 2 km was absent or reversed. Reproduced from Lynch-Stieglitz Jduring the Last Glacial Maximum. Science 316: 66–69, with permission from
and lower radiocarbon concentrations (Keigwin, 2004;
Robinson et al., 2005). The existence of these two distinct
water masses despite the mixing processes that would tend to
blend them suggests that one or both were renewed relatively
rapidly (Curry and Oppo, 2005), although reduced mixing
might have played a role as well (Lund et al., 2011).
It also seems clear at this point that the LGM 231Pa/230Th
ratio in Atlantic sediments was comparable to those we find
today (Gherardi et al., 2009; McManus et al., 2004; Yu et al.,
1996). This too would argue against a much slower circulation
of deep waters in the Atlantic Ocean during glacial times. The
cross basin contrast in d18O of benthic foraminifera, however,
was much smaller (Figure 7; Lynch-Stieglitz et al., 2006). This
would suggest that the glacial circulation was not simply a
shallower version of today’s meridional overturning circula-
tion, whereby surface and intermediate waters travel into the
North Atlantic to compensate the export of NADW.
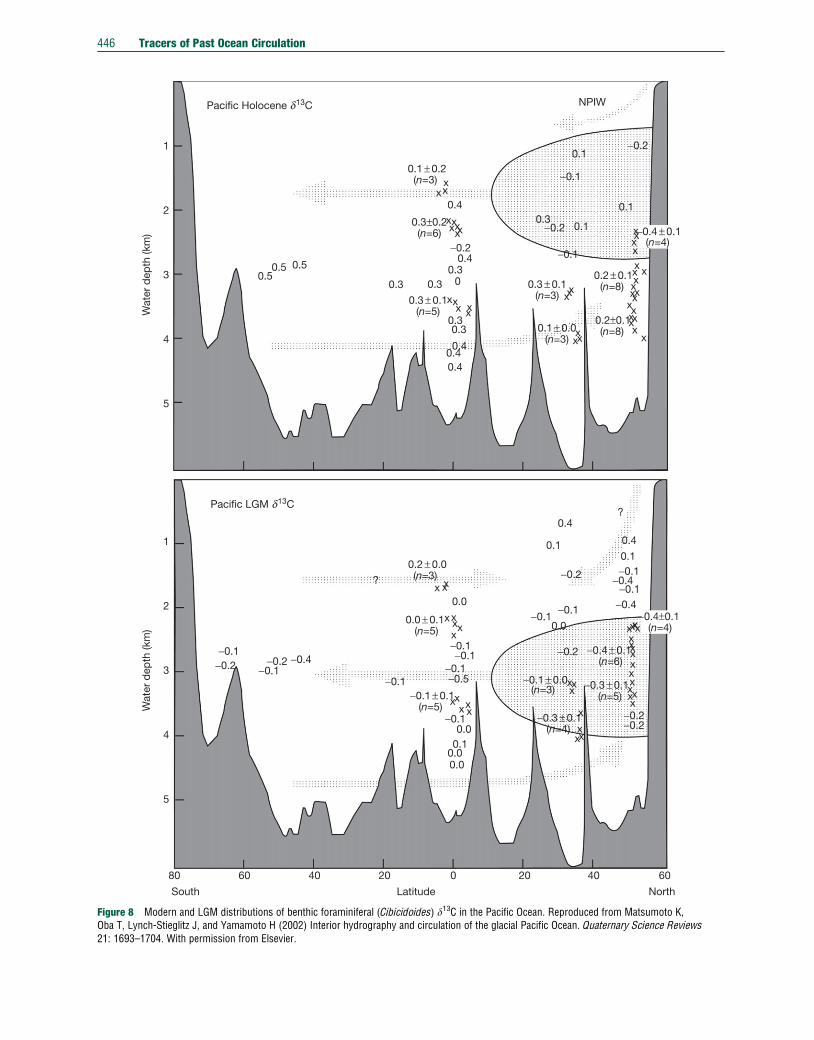
Less is known about the state of the LGM Pacific Ocean,
largely because poor carbonate preservation in the Pacific
Ocean limits the availability of cores with benthic foraminif-
era. Undersaturation with respect to calcium carbonate in
the Pacific also complicates the interpretation of the trace
metal-based methods that suffer from carbonate ion effects.
However, as in the Atlantic, the carbon and oxygen isotope
data suggest a deep hydrographic boundary at about 2000-m
water depth (Herguera et al., 1992, 2010; Keigwin, 1998;
Matsumoto et al., 2002; Mix et al., 1991), with a low-d13C,high-d18O water mass below this divide and a high- d13C, low-d18O water mass above this divide. Like today, the lowest d13Cwaters in the Pacific are in the far north but at deeper depths
(>2000-m water depth) than today (Figure 8; Herguera et al.,
2010; Matsumoto et al., 2002). There is now considerable
evidence that intermediate waters in the North Pacific were
43210321 5
2000
1500
1000
500
0
(‰ PDB) d18O (‰PDB)
Glacial(c)
Dep
th (m
)
ht et al., 2002). The waters are denser on the eastern side of theygen isotopic composition of the shells of benthic foraminifera (d18O) inde of the basin. These profiles reflect the density contrast across thee. The solid lines are the predicted d18O based on modern hydrographicr the LGM. These profiles suggest that the modern density contrast, Adkins JF, Curry WB (2007) Atlantic meridional overturning circulationthe American Association for the Advancement of Science.
1
2
3
4
5
0.50.5 0.5
0.3 0.30.3
0
0.4−0.2
0.3 ± 0.1(n=5)
0.3
xxx xx
xxxxxx xxxxxx
xxxx
x
xx
xxxx
xxx
x?
xxxxx
xx xx x
xx
x
x
xxx
x
xxxxxx
x
xxxxx
xxxx
x
xxx
xxxxx
0.3
0.40.40.4
0.4
0.4
0.10.1
0.0
0.0
0.00.00.10.0
−0.4
−0.4
−0.4
−0.1
−0.1−0.1
−0.1−0.1
−0.1 −0.1−0.1
−0.1
−0.1
−0.5
−0.1−0.2
−0.2−0.2−0.2
−0.2−0.2
?
0.3±0.2
0.4
(n=6)
0.3 ± 0.1(n=3)
0.1 ± 0.0(n=3)
−0.4±0.1(n=4)
0.2 ± 0.0(n=3)
0.0 ± 0.1(n=5)
−0.1 ± 0.1(n=5)
−0.1 ± 0.0(n=3)
−0.3 ± 0.1(n=4)
−0.4 ± 0.1(n=6)
−0.3 ± 0.1(n=5)
0.2±0.1(n=8)
0.2 ± 0.1(n=8)
−0.4 ± 0.1(n=4)
0.1 ± 0.2
Pacific Holocene d13C
Pacific LGM d13C
Wat
er d
epth
(km
)
1
2
3
4
5
80
South Latitude North
40200204060 60
Wat
er d
epth
(km
)−0.2
−0.2
−0.1
−0.1
0.1
0.1
0.10.3
NPIW
(n=3)
Figure 8 Modern and LGM distributions of benthic foraminiferal (Cibicidoides) d13C in the Pacific Ocean. Reproduced from Matsumoto K,Oba T, Lynch-Stieglitz J, and Yamamoto H (2002) Interior hydrography and circulation of the glacial Pacific Ocean. Quaternary Science Reviews21: 1693–1704. With permission from Elsevier.
446 Tracers of Past Ocean Circulation
Tracers of Past Ocean Circulation 447
more oxygen-rich during the LGM (van Geen et al., 2003),
consistent with the d13C-based scenario of waters more de-
pleted in nutrients at intermediate depths. The LGM Indian
Ocean is also characterized by a benthic divide at about
2000 m, with higher d13C and lower Cd reconstructed for
the upper water mass (Boyle et al., 1995; Kallel et al., 1988;
McCorkle et al., 1998). This deep hydrographic boundary is
also seen off southern Australia (Lynch-Stieglitz et al., 1996).
For both the upper and lower water masses, there is a
general progression from highest d13C in the North Atlantic
to intermediate values in the South Atlantic and South
Australia to progressively lower d13C values in the North Pacific
(excluding the deepest Southern Ocean). Goldstein et al.
(2001) also argue, based on limited glacial-age radiocarbon
data, that the modern progression of youngest deep waters in
the North Atlantic, intermediate values in the Southern Ocean,
and oldest values in the deep Pacific was preserved during
glacial times. The d18O values in the upper water mass are
similar from North Atlantic to North Pacific, within the con-
siderable scatter in the data. Whether or not the GNAIW
reached the Pacific or there was another source of nutrient-
poor upper deep water forming in the North Pacific remains a
source of debate (Keigwin, 1998; Lynch-Stieglitz et al., 1996;
Matsumoto et al., 2002; Oppo and Horowitz, 2000), although
box model results do argue for GNAIW export out of the
Atlantic (Sigman et al., 2003).
Finally, there appears to be an even denser water mass that
occupies the deepest portion of the Southern Ocean. This deep
water has high d18O values and very low d13C, with lowest d13Cvalues (about �1%) observed in the Atlantic sector of the
Southern Ocean (Figure 6). While there was some thought
that the low d13C values may reflect a productivity-related
overprint (Mackensen et al., 1993), a convincing case has
been made that the d13C values are regionally coherent and
34.5
3
2.5
2
1.5
1
0.5
0
−0.5
−1
−1.5
−2
3.5
35.535
Modern North Atlantic
Modern South Pacific
Thet
a (°
C)
Modern Southern Ocean
Site 9812184 m N. Atla
Site 10634584 m N. Atlantic
Figure 9 Estimates of LGM potential temperature (theta) and salinity fromsalinities (assuming 125 m of eustatic seawater change) are indicated with thfor the LGM are close to the freezing point of seawater (�1.98 �C). The data tReproduced from Adkins JF, McIntyre K, and Schrag DP (2002) The salinity, twith permission from the American Association for the Advancement of Scie
unrelated to other measures of overlying productivity
(Mackensen et al., 2001; Ninnemann and Charles, 2002).
Nd isotope values within this water mass have less of a
North Atlantic signature than today (Rutberg et al., 2000).
Unfortunately, there is not enough spatial coverage at present
to document convincingly the extent of this deep water mass in
the Indian and Pacific sectors of the Southern Ocean. The Cd/
Ca data are even more limited than the d13C data in the
Southern Ocean, due largely to low foraminiferal abundance
and potential overprints from saturation state; these data none-
theless suggest that the low d13C values are not accompanied
by a significant increase in nutrients (Rosenthal et al., 1997a).
This suggests that reduced air–sea exchange may have been
responsible, at least in part, for the lower d13C in the deep
Southern Ocean relative to the rest of the deep LGM ocean
(Marchitto and Broecker, 2006; Toggweiler, 1999).
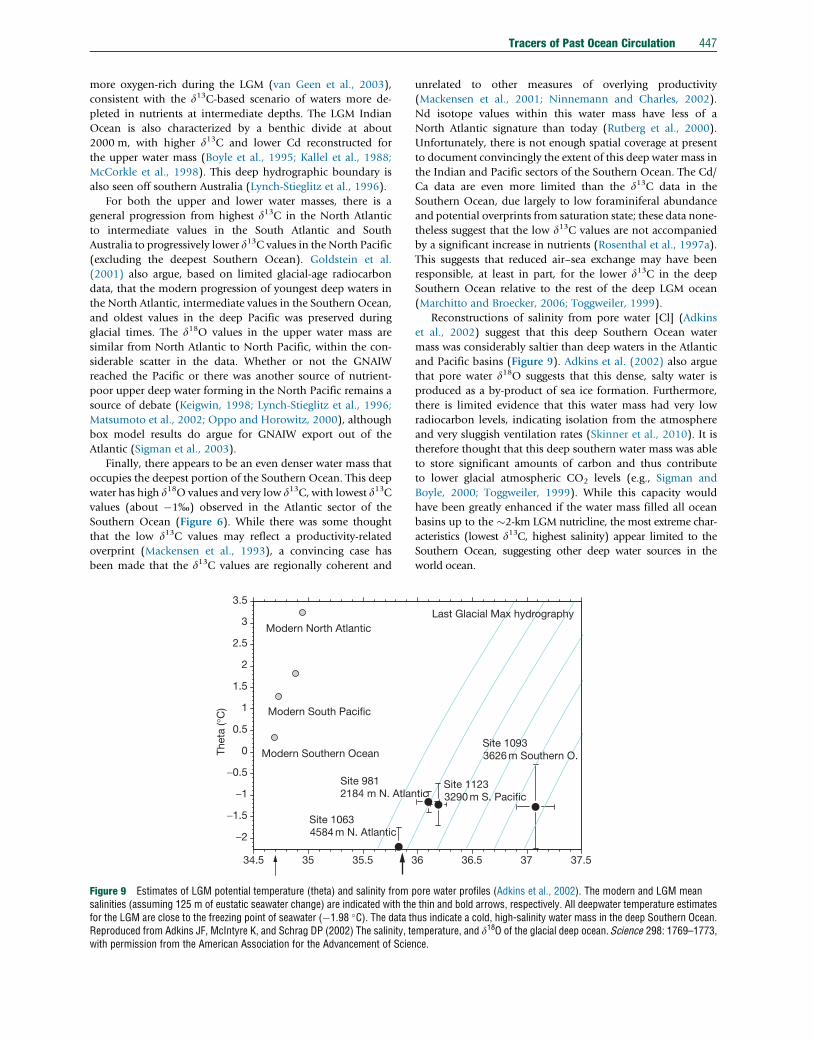
Reconstructions of salinity from pore water [Cl] (Adkins
et al., 2002) suggest that this deep Southern Ocean water
mass was considerably saltier than deep waters in the Atlantic
and Pacific basins (Figure 9). Adkins et al. (2002) also argue
that pore water d18O suggests that this dense, salty water is
produced as a by-product of sea ice formation. Furthermore,
there is limited evidence that this water mass had very low
radiocarbon levels, indicating isolation from the atmosphere
and very sluggish ventilation rates (Skinner et al., 2010). It is
therefore thought that this deep southern water mass was able
to store significant amounts of carbon and thus contribute
to lower glacial atmospheric CO2 levels (e.g., Sigman and
Boyle, 2000; Toggweiler, 1999). While this capacity would
have been greatly enhanced if the water mass filled all ocean
basins up to the �2-km LGM nutricline, the most extreme char-
acteristics (lowest d13C, highest salinity) appear limited to the
Southern Ocean, suggesting other deep water sources in the
world ocean.
3736.536 37.5
Last Glacial Max hydrography
nticSite 11233290 m S. Pacific
Site 10933626 m Southern O.
pore water profiles (Adkins et al., 2002). The modern and LGM meane thin and bold arrows, respectively. All deepwater temperature estimateshus indicate a cold, high-salinity water mass in the deep Southern Ocean.emperature, and d18O of the glacial deep ocean. Science 298: 1769–1773,nce.

448 Tracers of Past Ocean Circulation
8.16.8 Conclusions
Considerable strides have been made in the last twenty-five
years in developing methods to reconstruct past ocean circula-
tion. There are now a number of different geochemical
methods that can yield information on paleo-ocean flow.
However, as the use of each method develops, it becomes
clear that there are multiple processes affecting both the tracers
themselves and their imprint into the sediments. We are still
just beginning to understand most of these methods and there
are limited amounts of data available. A full understanding of
these factors and how to separate out their effects, combined
with multiple measurements using methods affected by differ-
ent processes, can ultimately lead to well-constrained recon-
structions of past ocean circulation. However, even now, these
methods are capable of giving us a tantalizing glimpse of the
ocean during the LGM, when the water masses were arranged
in a distinctly different pattern than they are today.
References
Adkins JF and Boyle EA (1997) Changing atmospheric D14C and the record of deepwater paleoventilation ages. Paleoceanography 12: 337–344.
Adkins JF, Cheng H, Boyle EA, Druffel ERM, and Edwards RL (1998) Deep-sea coralevidence for rapid change in ventilation of the deep North Atlantic 15,400 years ago.Science 280: 725–728.
Adkins JF, McIntyre K, and Schrag DP (2002) The salinity, temperature, and d18O of theglacial deep ocean. Science 298: 1769–1773.
Adkins JF and Schrag DP (2001) Pore fluid constraints on deep ocean temperature andsalinity during the Last Glacial Maximum. Geophysical Research Letters28: 771–775.
Albarede F and Goldstein SL (1992) World map of Nd isotopes in sea-floorferromanganese deposits. Geology 20: 761–763.
Albarede F, Goldstein SL, and Dautel D (1997) The neodymium isotopic composition ofmanganese nodules from the Southern and Indian oceans, the global oceanicneodymium budget, and their bearing on deep ocean circulation. Geochimica etCosmochimica Acta 61: 1277–1291.
Anderson RF, Bacon MP, and Brewer PG (1983) Removal of 231Pa and 230Th from theopen ocean. Earth and Planetary Science Letters 62: 7–23.
Ballini M, Kissel C, Colin C, and Richter T (2006) Deep-water mass source and dynamicassociated with rapid climatic variations during the last glacial stage in the NorthAtlantic: A multi-proxy investigation of the detrital fraction of deep-sea sediments.Geochemistry, Geophysics, Geosystems 7: Q02N01.
Belanger PE, Curry WB, and Matthews RK (1981) Core-top evaluation of benthicforaminiferal isotopic ratios for paleo-oceanographic interpretations.Palaeogeography, Palaeoclimatology, Palaeoecology 33: 205–220.
Bickert T and Mackensen A (2004) Last Glacial to Holocene changes in South AtlanticDeep Water Circulation. In: Wefer G, Mulitza S, and Ratmeyer V (eds.) The SouthAtlantic in the Late Quaternary: Reconstruction of Material Budget and CurrentSystems, pp. 671–693. Berlin: Springer-Verlag.
Bostock HC, Opdyke BN, Gagan MK, Kiss AE, and Fifield LK (2006) Glacial/interglacialchanges in the East Australian current. Climate Dynamics 26: 645–659.
Boyle EA (1981) Cadmium, zinc, copper, and barium in foraminifera tests. Earth andPlanetary Science Letters 53: 11–35.
Boyle EA (1988) Cadmium: Chemical tracer of deepwater paleoceanography.Paleoceanography 3: 471–489.
Boyle EA (1992) Cadmium and d13C paleochemical ocean distributions during thestage 2 glacial maximum. Annual Reviews of Earth and Planetary Sciences20: 245–287.
Boyle EA and Keigwin LD (1982) Deep circulation of the North Atlantic over the last200,000 years: Geochemical evidence. Science 218: 784–787.
Boyle EA and Keigwin LD (1987) North Atlantic thermohaline circulation during the last20,000 years linked to high latitude surface temperature. Nature 330: 35–40.
Boyle EA, Labeyrie L, and Duplessy J-C (1995) Calcitic foraminiferal data confirmed bycadmium in aragonitic Hoeglundina : Application to the Last Glacial Maximum in thenorthern Indian Ocean. Paleoceanography 10: 881–900.
Boyle EA and Rosenthal Y (1996) Chemical hydrography of the South Atlantic duringthe Last Glacial Maximum: Cd vs. d13C. In: Wefer G, Berger WH, Siedler G, andWebb DJ (eds.) The South Atlantic: Present and Past Circulation, pp. 423–443.Berlin: Springer-Verlag.
Boyle EA, Sclater FR, and Edmond JM (1976) On the marine geochemistry of cadmium.Nature 263: 42–44.
Broecker WS (1982) Glacial to interglacial changes in ocean chemistry. Progress inOceanography 11: 151–197.
Broecker WS (2002) Constraints on the glacial operation of the Atlantic Ocean’sconveyor circulation. Israel Journal of Chemistry 42: 1–14.
Broecker WS, Andree M, Bonani G, et al. (1988) Preliminary estimates for theradiocarbon age of deep water in the glacial ocean. Paleoceanography3: 659–669.
Broecker WS and Maier-Reimer E (1992) The influence of air and sea exchange on thecarbon isotope distribution in the sea. Global Biogeochemical Cycles 6: 315–320.
Broecker W, Matsumoto K, Clark E, Hajdas I, and Bonani G (1999) Radiocarbon agedifferences between coexisting foraminiferal species. Paleoceanography14: 431–436.
Broecker WS and Peng T-H (1974) Gas exchange rates between air and sea. Tellus26: 21–35.
Broecker WS and Peng T-H (1982) Tracers in the Sea. Palisades, NY: Eldigio Press.Broecker WS, Peng T-H, Trumbore S, Bonani G, and Wolfli W (1990) The distribution of
radiocarbon in the glacial ocean. Global Biogeochemical Cycles 4: 103–107.Bruland KW, Knauer GA, and Martin JH (1978) Zinc in north-east Pacific water. Nature
271: 741–743.Bryan SP and Marchitto TM (2008) Mg/Ca-temperature proxy in benthic foraminifera:
New calibrations from the Florida Straits and a hypothesis regarding Mg/Li.Paleoceanography 23: 1–17.
Bryan SP and Marchitto TM (2010) Testing the utility of paleonutrient proxies Cd/Caand Zn/Ca in benthic foraminifera from thermocline waters. Geochemistry,Geophysics, Geosystems 11: Q01005.
Burke A, Marchal O, Bradtmiller LI, McManus JF, and Francois R (2011) Application ofan inverse method to interpret 231Pa/230Th observations from marine sediments.Paleoceanography 26: PA1212.
Conkright ME, Locarnini RA, Garcia TD, et al. (2002)World Ocean Atlas 2001: ObjectiveAnalyses, Data Statistics, and Figures, CD-ROM Documentation. Silver Spring, MD:National Oceanographic Data Center.
Corliss BH, Sun X, Brown CW, and Showers WJ (2006) Influence of seasonalprimary productivity on d13C of North Atlantic deep-sea benthic foraminifera. DeepSea Research Part I: Oceanographic Research Papers 53: 740–746.
Craig H (1970) Abyssal carbon 13 in the South Pacific. Journal of GeophysicalResearch 75: 691–695.
Craig H and Gordon LT (1965) Deuterium and oxygen 18 variations in the ocean and themarine atmosphere: Stable isotopes in oceanographic studies andpaleotemperatures. In: Tongiori E (ed.) Proceedings of the Third SpoletoConference, Spoleto, Italy, pp. 9–130. Pisa: Sischi and Figli.
Curry WB, Duplessy J-C, Labeyrie LD, and Shackleton NJ (1988) Changes in thedistribution of d13C of deep water SCO2 between the last glaciation and theHolocene. Paleoceanography 3: 317–341.
Curry WB and Lohmann GP (1982) Carbon isotopic changes in benthic foraminiferafrom the western South Atlantic: Reconstructions of glacial abyssal circulationpatterns. Quaternary Research 18: 218–235.
Curry WB and Oppo DW (2005) Glacial water mass geometry and the distribution ofd13C of SCO2 in the western Atlantic Ocean. Paleoceanography 20: PA1017.
Deuser WG and Hunt JM (1969) Stable isotope ratios of dissolved inorganiccarbon in the Atlantic. Deep Sea Research and Oceanographic Abstracts16: 221–225.
Duplessy JC, Labeyrie L, and Waelbroeck C (2002) Constraints on the ocean oxygenisotopic enrichment between the Last Glacial Maximum and the Holocene:Paleoceanographic implications. Quaternary Science Reviews 21: 315–330.
Duplessy J-C, Shackleton NJ, Fairbanks RG, Labeyrie L, Oppo D, and Kallel N (1988)Deepwater source variations during the last climatic cycle and their impact on theglobal deepwater circulation. Paleoceanography 3: 343–360.
Duplessy J-C, Shackleton NJ, Matthews RK, et al. (1984) 13C record of benthicforaminifera in the last interglacial ocean: Implications for the carbon cycle and theglobal deep water circulation. Quaternary Research 21: 225–243.
Eberwein A and Mackensen A (2006) Regional primary productivity differences offMorocco (NW-Africa) recorded by modern benthic foraminifera and their stablecarbon isotopic composition. Deep Sea Research Part I: Oceanographic ResearchPapers 53: 1379–1405.
Elderfield H, Bertram CJ, and Erez J (1996) A biomineralization model for theincorporation of trace elements into foraminiferal calcium carbonate. Earth andPlanetary Science Letters 142: 409–423.

Tracers of Past Ocean Circulation 449
Elderfield H, Greaves M, Barker S, et al. (2010) A record of bottom water temperatureand seawater d18O for the Southern Ocean over the past 440 kyr based on Mg/Ca ofbenthic foraminiferal Uvigerina spp. Quaternary Science Reviews 29: 160–169.
Elderfield H and Rickaby REM (2000) Oceanic Cd/P ratio and nutrient utilization in theglacial Southern Ocean. Nature 405: 305–310.
Elderfield H, Yu J, Anand P, Kiefer T, and Nyland B (2006) Calibrations for benthicforaminiferal Mg/Ca paleothermometry and the carbonate ion hypothesis. Earth andPlanetary Science Letters 250: 633–649.
Frank M (2002) Radiogenic isotopes: Tracers of past ocean circulation and erosionalinput. Reviews of Geophysics 40(1): 1-1–1-38.
Gherardi JM, Labeyrie L, Nave S, Francois R, McManus JF, and Cortijo E (2009)Glacial-interglacial circulation changes inferred from 231Pa/230Th sedimentaryrecord in the North Atlantic region. Paleoceanography 24: PA2204.
Ghosh P, Adkins J, Affek H, et al. (2006) 13C-18O bonds in carbonate minerals: A newkind of paleothermometer. Geochimica et Cosmochimica Acta 70: 1439–1456.
Gil IM, Keigwin LD, and Abrantes FG (2009) Deglacial diatom productivity and surfaceocean properties over the Bermuda Rise, northeast Sargasso Sea.Paleoceanography 24: PA4101.
Goldstein SJ and Hemming SR (2003) Long-lived isotopic tracers in oceanography,paleoceanography, and ice-sheet dynamics. In: Elderfield (ed.) Treatise onGeochemistry, Vol. 6: The Oceans and Marine Geochemistry, pp. 453–489. Oxford:Elsevier.
Goldstein SJ, Lea DW, Chakraborty S, Kashgarian M, and Murrell MT (2001)Uranium-series and radiocarbon geochronology of deep-sea corals: Implicationsfor Southern Ocean ventilation rates and the oceanic carbon cycle. Earth andPlanetary Science Letters 193: 167–182.
Graham DW, Corliss BH, Bender ML, and Keigwin LD (1981) Carbon and oxygenisotopic disequilibria of recent deep-sea benthic foraminifera. MarineMicropaleontology 6: 483–497.
Grossman EL (1984a) Stable isotope fractionation in live benthic foraminifera from thesouthern California Borderland. Palaeogeography, Palaeoclimatology,Palaeoecology 47: 301–327.
Grossman EL (1984b) Carbon isotopic fractionation in live benthicforaminifera-comparison with inorganic precipitate studies. Geochimica etCosmochimica Acta 48: 1505–1512.
Gruber N, Keeling CD, Bacastow RB, et al. (1999) Spatiotemporal patterns of carbon-13in the global surface oceans and the oceanic Suess effect. Global BiogeochemicalCycles 13: 307–335.
Gutjahr M, Frank M, Stirling CH, Klemm V, van de Flierdt T, and Halliday AN (2007)Reliable extraction of a deepwater trace metal isotope signal from Fe-Mnoxyhydroxide coatings of marine sediments. Chemical Geology 242: 351–370.
Havach SM, Chandler GT, Wilson-Finelli A, and Shaw TJ (2001) Experimentaldetermination of trace element partition coefficients in cultured benthic foraminifera.Geochimica et Cosmochimica Acta 65: 1277–1283.
Hendry KR, Georg RB, Rickaby REM, Robinson LF, and Halliday AN (2010) Deep oceannutrients during the Last Glacial Maximum deduced from sponge silicon isotopiccompositions. Earth and Planetary Science Letters 292: 290–300.
Herguera JC, Herbert T, Kashgarian M, and Charles C (2010) Intermediate and deepwater mass distribution in the Pacific during the Last Glacial Maximum inferred fromoxygen and carbon stable isotopes. Quaternary Science Reviews 29: 1228–1245.
Herguera JC, Jansen E, and Berger WH (1992) Evidence for a bathayal front at 2000 mdepth in the glacial Pacific, based on a depth transect on Ontong Java Plateau.Paleoceanography 7: 273–288.
Hughen KA, Overpeck JT, Lehman SJ, et al. (1998) Deglacial changes in oceancirculation from an extended radiocarbon calibration. Nature 391: 65–68.
Inoue H and Sugimura Y (1985) Carbon isotopic fractionation during CO2 exchangeprocess between air and sea water under equilibrium and kinetic conditions.Geochimica et Cosmochimica Acta 49: 2453–2460.
Kallel N, Labeyrie LD, Juillet-Leclerc A, and Duplessy J-C (1988) A deep hydrologicalfront between intermediate and deep water masses in the glacial Indian Ocean.Nature 333: 651–655.
Keigwin LD (1998) Glacial-age hydrography of the far northwest Pacific Ocean.Paleoceanography 13: 323–339.
Keigwin LD (2004) Radiocarbon and stable isotope constraints on Last GlacialMaximum and Younger Dryas ventilation in the western North Atlantic.Paleoceanography 19: PA4012.
Keigwin LD and Schlegel MA (2002) Ocean ventilation and sedimentation since theglacial maximum at 3 km in the western North Atlantic. Geochemistry, Geophysics,Geosystems 3: 1034.
Kim S-T and O’Neil J (1997) Equilibrium and nonequilibrium oxygen isotope effects insynthetic carbonates. Geochimica et Cosmochimica Acta 61: 3461–3475.
Kissel C, Laj C, Labeyrie L, Dokken T, Voelker A, and Blamart D (1999) Rapidclimatic variations during marine isotopic stage 3: Magnetic analysis of sediments
from Nordic Sea and North Atlantic. Earth and Planetary Science Letters171: 489–502.
Klevenz V, Vance D, Schmidt DN, and Mezger K (2008) Neodymium isotopes inbenthic foraminifera: Core-top systematics and a down-core record from theNeogene south Atlantic. Earth and Planetary Science Letters 265: 571–587.
Kroopnick PM (1985) The distribution of 13C of SCO2 in the world oceans. Deep SeaResearch Part A. Oceanographic Research Papers 32: 57–84.
Labeyrie LD (1992) Changes in the vertical structure of the North Atlantic Oceanbetween glacial and modern times. Quaternary Science Reviews 11: 401–413.
Lacan F and Jeandel C (2005) Neodymium isotopes as a new tool for quantifyingexchange fluxes at the continent-ocean interface. Earth and Planetary Science Letters232: 245–257.
Lal D and Charles C (2007) Deconvolution of the atmospheric radiocarbon record in thelast 50,000 years. Earth and Planetary Science Letters 258: 550–560.
Lea DW (1993) Constraints on the alkalinity and circulation of glacial circumpolar deepwater from benthic foraminiferal barium. Global Biogeochemical Cycles7: 695–710.
Lea DW and Boyle EA (1989) Barium content of benthic foraminifera controlled bybottom-water composition. Nature 338: 751–753.
Lea DW and Boyle EA (1990) Foraminiferal reconstructions of barium distributions inwater masses of the glacial oceans. Paleoceanography 5: 719–742.
Lear CH, Mawbey EM, and Rosenthal Y (2010) Cenozoic benthic foraminiferal Mg/Caand Li/Ca records: Toward unlocking temperatures and saturation states.Paleoceanography 25: PA4215.
Ledbetter MT and Johnson DA (1976) Increased transport of Antarctic Bottom Water inthe Vema Channel during the Last Ice Age. Science 194: 837–839.
LeGrand P and Wunsch C (1995) Constraints from paleotracer data on the NorthAtlantic circulation during the Last Glacial Maximum. Paleoceanography10: 1011–1045.
LeGrande AN and Lynch-Stieglitz J (2007) Surface currents in the western NorthAtlantic during the Last Glacial Maximum. Geochemisty, Geophysics, Geosystems8: Q01N09.
Levitus S and Boyer T (1994) World Ocean Atlas, vol. 4, Temperature. NOAA NESDIS 4.Washington, DC: US Department of Commerce.
Lippold J, Grutzner J, Winter D, Lahaye Y, Mangini A, and Christl M (2009) Doessedimentary 231Pa/230Th from the Bermuda Rise monitor past Atlantic MeridionalOverturning Circulation? Geophysical Research Letters 36: L12601.
Loubere P (1991) Deep-sea benthic foraminiferal assemblage response to a surfaceocean productivity gradient: A test. Paleoceanography 6: 193–204.
Lund DC, Adkins JF, and Ferrari R (2011) Abyssal Atlantic circulation during the LastGlacial Maximum: Constraining the ratio between transport and vertical mixing.Paleoceanography 26: PA1213.
Lund DC, Lynch-Stieglitz J, and Curry WB (2006) Gulf Stream density structure andtransport during the past millennium. Nature 444: 601–604.
Luo S and Ku T-L (1999) Oceanic 231Pa/230Th ratio influenced by particle compositionand remineralization. Earth and Planetary Science Letters 167: 183–195.
Lutze GF and Thiel H (1989) Epibenthic foraminifera from elevated microhabitats:Cibicidoides wuellerstorfi and Planulina ariminensis. Journal of ForaminiferalResearch 19: 153–158.
Lynch-Stieglitz J (2001) Using ocean margin density to constrain ocean circulationand surface wind strength in the past. Geochemistry, Geophysics, Geosystems2(12): 1069.
Lynch-Stieglitz J, Adkins JF, Curry WB, et al. (2007) Atlantic meridional overturningcirculation during the Last Glacial Maximum. Science 316: 66–69.
Lynch-Stieglitz J, Curry WB, and Lund DC (2009) Florida Straits density structure andtransport over the last 8000 years. Paleoceanography 24: 1–9.
Lynch-Stieglitz J, Curry WB, Oppo DW, Ninnemann U, Charles CD, and Munsen J(2006) Meridional overturning circulation in the South Atlantic at the Last GlacialMaximum. Geochemistry, Geophysics, Geosystems 7: Q10N03.
Lynch-Stieglitz J, Curry WB, and Slowey N (1999a) A geostrophic transport estimate forthe Florida Current from the oxygen isotope composition of benthic foraminifera.Paleoceanography 14: 360–373.
Lynch-Stieglitz J, Curry WB, and Slowey N (1999b) Weaker gulf stream in the FloridaStraits during the Last Glacial Maximum. Nature 402: 644–648.
Lynch-Stieglitz J and Fairbanks RG (1994) A conservative tracer for glacial oceancirculation from carbon isotope and paleonutrient measurements in benthicforaminifera. Nature 369: 308–310.
Lynch-Stieglitz J, Schmidt MW, and Curry WB (2011) Evidence from the FloridaStraits for Younger Dryas ocean circulation changes. Paleoceanography26: PA1205.
Lynch-Stieglitz J, Stocker TF, Broecker WS, and Fairbanks RG (1995) The influence ofair-sea exchange on the isotopic composition of oceanic carbon: Observations andmodeling. Global Biogeochemical Cycles 9: 653–665.

450 Tracers of Past Ocean Circulation
Lynch-Stieglitz J, van Geen A, and Fairbanks RG (1996) Interocean exchange of GlacialNorth Atlantic Intermediate Water: Evidence from subantarctic Cd/Ca and carbonisotope measurements. Paleoceanography 11: 191–201.
Mackensen A and Douglas RG (1989) Down-core distribution of live and deaddeep-water benthic foraminifera in box cores from the Weddell Sea and theCalifornia continental borderland. Deep Sea Research Part A. OceanographicResearch Papers 36: 879–900.
Mackensen A, Hubberten H-W, Bickert T, Fischer G, and Futterer DK (1993) The d13C inbenthic foraminiferal tests of Fontbotia wuellerstorfi (Schwager) relative to the d13Cof dissolved inorganic carbon in Southern Ocean Deep Water: Implications forglacial ocean circulation models. Paleoceanography 8: 587–610.
Mackensen A, Rudolph M, and Kuhn G (2001) Late Pleistocene deep-water circulationin the subantarctic eastern Atlantic. Global and Planetary Change 30: 197–229.
Mangini A, Lomitschka M, Eichstadter R, et al. (1998) Coral provides way to age deepwater. Nature 392: 347–348.
Marchal O, Stocker TF, and Muscheler R (2001) Atmospheric radiocarbon during theYounger Dryas: Production, ventilation, or both? Earth and Planetary Science Letters185: 383–395.
Marchitto TM (2004) Lack of a significant temperature influence on the incorporation ofCd into benthic foraminiferal tests. Geochemistry, Geophysics, Geosystems5: Q10D11.
Marchitto TM and Broecker WS (2006) Deep water mass geometry in the glacial AtlanticOcean: A review of constraints from trace metal paleonutrient proxies. Geochemistry,Geophysics, Geosystems 7(12): Q12003.
Marchitto TM, Curry WB, and Oppo DW (2000) Zinc concentrations in benthicforaminifera reflect seawater chemistry. Paleoceanography 15: 299–306.
Marchitto TM, Oppo DW, and Curry WB (2002) Paired benthic foraminiferal Cd/Ca andZn/Ca evidence for a greatly increased presence of Southern Ocean Water in theglacial North Atlantic. Paleoceanography 17(3): 1038.
Martin EE and Haley BA (2000) Fossil fish teeth as proxies for seawater Sr and Ndisotopes. Geochimica et Cosmochimica Acta 64: 835–847.
Martin PA, Lea DW, Rosenthal Y, Shackleton NJ, Sarnthein M, and Papenfuss T (2002)Quaternary deep sea temperature histories derived from benthic foraminiferalMg/Ca. Earth and Planetary Science Letters 198: 193–209.
Matsumoto K and Lynch-Stieglitz J (1999) Similar glacial and Holocene deep watercirculation inferred from southeast Pacific benthic foraminiferal carbon isotopecomposition. Paleoceanography 14: 149–163.
Matsumoto K and Lynch-Stieglitz J (2003) Persistence of Gulf Stream separation duringthe Last Glacial Maximum: Implications for current separation theories. Journal ofGeophysical Research 108(C6): 3174.
Matsumoto K, Lynch-Stieglitz J, and Anderson RF (2001) Similar glacial and HoloceneSouthern Ocean hydrography. Paleoceanography 16: 445–454.
Matsumoto K, Oba T, Lynch-Stieglitz J, and Yamamoto H (2002) Interior hydrographyand circulation of the glacial Pacific Ocean. Quaternary Science Reviews21: 1693–1704.
McCave IN, Manighetti B, and Robinson SG (1995) Sortable silt and fine sedimentsize/composition slicing: Parameters for palaeocurrent speed andpalaeoceanography. Paleoceanography 10: 593–610.
McCorkle DC, Corliss BH, and Farnham CA (1997) Vertical distributions and stableisotopic compositions of live (stained) benthic foraminifera from the North Carolinaand California continental margins. Deep Sea Research Part I: OceanographicResearch Papers 44: 983–1024.
McCorkle DC, Heggie DT, and Veeh HH (1998) Glacial and Holocene stable isotopedistributions in the southeastern Indian Ocean. Paleoceanography 13: 20–34.
McCorkle DC, Keigwin LD, Corliss BH, and Emerson SR (1990) The influence ofmicrohabitats on the carbon isotopic composition of deep-sea benthic foraminifera.Paleoceanography 5: 161–185.
McCorkle DC and Klinkhammer GP (1991) Porewater cadmium geochemistry and theporewater cadmium–d13C relationship. Geochimica et Cosmochimica Acta55: 161–168.
McCorkle DC, Martin PA, Lea DW, and Klinkhammer GP (1995) Evidence of adissolution effect on benthic foraminiferal shell chemistry: d13C, Cd/Ca, Ba/Ca, andSr/Ca results from the Ontong Java Plateau. Paleoceanography 10: 699–714.
McManus JF, Francois R, Gherardi JM, Keigwin LD, and Brown-Leger S (2004)Collapse and rapid resumption of Atlantic meridional circulation linked to deglacialclimate changes. Nature 428: 834–837.
Miller KG and Lohmann GP (1982) Environmental distribution of Recent benthicforaminifera on the northeast United States continental Slope. Geological Society ofAmerica Bulletin 93: 200–206.
Mix AC, Pisias NG, Zahn R, Rugh W, Lopez C, and Nelson K (1991) Carbon 13 inPacific deep and intermediate waters, 0–370 ka: Implications for ocean circulationand Pleistocene CO2. Paleoceanography 6: 205–226.
Mook WG, Bommerson JC, and Staverman WH (1974) Carbon isotope fractionationbetween dissolved bicarbonate and gaseous carbon dioxide. Earth and PlanetaryScience Letters 22: 169–176.
Ninnemann US and Charles CD (2002) Changes in the mode of Southern Oceancirculation over the last glacial cycle revealed by foraminiferal stable isotopicvariability. Earth and Planetary Science Letters 201: 383–396.
O’Neil JR, Clayton RN, and Mayeda TK (1969) Oxygen isotope fractionation in divalentmetal carbonates. Journal of Chemical Physics 5: 5547–5558.
Oppo DW and Horowitz M (2000) Glacial deep water geometry: South Atlantic benthicforaminiferal Cd/Ca and d13C evidence. Paleoceanography 15: 147–160.
Oppo DW and Lehman SJ (1993) Mid-depth circulation of the subpolar North Atlanticduring the Last Glacial Maximum. Science 259: 1148–1152.
Ortiz J, Mix A, Hostetler S, and Kashgarian M (1997) The California current of the LastGlacial Maximum: Reconstruction at 42� N based on multiple proxies.Paleoceanography 12(2): 191–205.
Ostermann DR and Curry WB (2000) Calibration of stable isotopic data: An enrichedd18O standard used for source gas mixing detection and correction.Paleoceanography 15: 353–360.
Peng T-H, Broecker WS, Kipphut G, and Shackleton N (1977) Benthic mixing in deepsea cores as determined by 14C dating and its implications regarding climatestratigraphy and the fate of fossil fuel CO2. In: Anderson NR and Malahoff A (eds.)Fate of Fossil Fuel CO2 in the Oceans, pp. 355–373. New York: Plenum Press.
Piepgras DJ and Wasserburg GJ (1980) Neodymium isotopic variations in seawater.Earth and Planetary Science Letters 50: 128–138.
Rathburn AE, Corliss BH, Tappa KD, and Lohmann KC (1996) Comparisons of theecology and stable isotopic compositions of living (stained) benthic foraminiferafrom the Sulu and South China Seas. Deep Sea Research Part I: OceanographicResearch Papers 43: 1617–1646.
Rathburn AE and DeDeckker P (1997) Magnesium and strontium compositions ofRecent benthic foraminifera from the Coral Sea, Australia and Prydz Bay, Antarctica.Marine Micropaleontology 32: 231–248.
Rickaby REM and Elderfield H (2005) Evidence from the high-latitude North Atlantic forvariations in Antarctic Intermediate water flow during the last deglaciation.Geochemistry, Geophysics, Geosystems 6: Q05001.
Rickaby REM, Greaves MJ, and Elderfield H (2000) Cd in planktonic and benthicforaminiferal shells determined by thermal ionisation mass spectrometry.Geochimica et Cosmochimica Acta 64: 1229–1236.
Roberts NL, Piotrowski AM, McManus JF, and Keigwin LD (2010) Synchronousdeglacial overturning and water mass source changes. Science 327(5961):75–78.
Robinson LF, Adkins JF, Keigwin LD, et al. (2005) Radiocarbon variability in thewestern North Atlantic during the last deglaciation. Science 310: 1469–1473.
Romanek CS, Grossman EL, and Morse JW (1992) Carbon isotope fractionation insynthetic aragonite and calcite: Effect of temperature and precipitation rate.Geochimica et Cosmochimica Acta 56: 419–430.
Rosenthal Y, Boyle EA, and Labeyrie L (1997a) Last Glacial Maximum paleochemistryand deepwater circulation in the Southern Ocean: Evidence from foraminiferalcadmium. Paleoceanography 12: 787–796.
Rosenthal Y, Boyle EA, Labeyrie L, and Oppo D (1995a) Glacial enrichments ofauthigenic Cd and U in subantarctic sediments – A climatic control on the elements’oceanic budget? Paleoceanography 10: 395–413.
Rosenthal Y, Boyle EA, and Slowey N (1997b) Temperature control on the incorporationof magnesium, strontium, fluorine, and cadmium into benthic foraminiferal shellsfrom Little Bahama Bank: Prospects for thermocline paleoceanography. Geochimicaet Cosmochimica Acta 61: 3633–3643.
Rosenthal Y, Lam P, Boyle EA, and Thomson J (1995b) Authigenic cadmiumenrichments in suboxic sediments – Precipitation and postdepositional mobility.Earth and Planetary Science Letters 132: 99–111.
Rutberg RL, Hemming SR, and Goldstein SL (2000) Reduced North Atlantic DeepWater flux to the glacial Southern Ocean inferred from neodymium isotope ratios.Nature 405: 935–938.
Schnitker D (1974) West Atlantic abyssal circulation during the past 120,000 years.Nature 248: 385–387.
Schnitker D (1994) Deep-sea benthic foraminifers: Food and bottom water masses.In: Zahn R, Pedersen TF, Kaminiski MA, and Labeyrie L (eds.) Carbon Cycling in theGlacial Ocean: Constraints on the Ocean’s Role in Global Change, NATO ASI Series,vol. 17, pp. 539–554.
Schrag DP, Adkins JF, McIntyre K, et al. (2002) The oxygen isotopic composition ofseawater during the Last Glacial Maximum. Quaternary Science Reviews21: 331–342.
Schrag DP and DePaolo DJ (1993) Determination of d18O of seawater in the deep oceanduring the Last Glacial Maximum. Paleoceanography 8: 1–6.

Tracers of Past Ocean Circulation 451
Shackleton NJ (1977) Tropical rainforest history and the equatorial Pacific carbonatedissolution cycles. In: Anderson NR and Malahoff A (eds.) Fate of Fossil Fuel CO2 inthe Oceans, pp. 401–427. New York: Plenum Press.
Shackleton NJ, Duplessy JC, Arnold M, Maurice P, Hall MA, and Cartlidge J (1988)Radiocarbon age of last glacial Pacific deep water. Nature 335: 708–711.
Siddall M, Stocker TF, Henderson GM, et al. (2007) Modeling the relationship between231Pa/230Th distribution in North Atlantic sediment and Atlantic meridionaloverturning circulation. Paleoceanography 22: PA2214.
Sigman DM and Boyle EA (2000) Glacial/interglacial variations in atmospheric carbondioxide. Nature 407: 859–869.
Sigman DM, Lehman SJ, and Oppo DW (2003) Evaluating mechanisms of nutrientdepletion and 13C enrichment in the intermediate-depth Atlantic during the last iceage. Paleoceanography 18(3): 1072.
Skinner LC, Fallon S, Waelbroeck C, Michel E, and Barker S (2010) Ventilation of the deepSouthern Ocean and deglacial CO2 rise. Science 328(5982): 1147–1151.
Skinner LC and Shackleton NJ (2004) Rapid transient changes in northeast Atlantic deepwater ventilation age across Termination I. Paleoceanography 19: 1–11.
Spero HJ, Bijma J, Lea DW, and Bemis BE (1997) Effect of seawater carbonate chemistryon planktonic foraminiferal carbon and oxygen isotope values. Nature390: 497–500.
Spero HJ and Lea DW (1996) Experimental determination of stable isotope variability inGlobigerina bulloides : Implications for paleoceanographic reconstructions. MarineMicropaleontology 28: 231–246.
Streeter SS (1973) Bottom water and benthonic foraminifera in the North Atlanticglacial-interglacial contrasts. Quaternary Research 3: 131–141.
Stuiver M, Quay PD, and Ostlund HG (1983) Abyssal water carbon-14 distribution andthe age of the world oceans. Science 219: 849–851.
Tachikawa K and Elderfield H (2002) Microhabitat effects on Cd/Ca and d13C of benthicforaminifera. Earth and Planetary Science Letters 202: 607–624.
Tans PP (1980) On calculating the transfer of carbon-13 in reservoir models of thecarbon cycle. Tellus 32: 464–469.
Thiagarajan N, Adkins J, and Eiler J (2011) Carbonate clumped isotope thermometryof deep-sea corals and implications for vital effects. Geochimica et CosmochimicaActa 75: 4416–4425.
Toggweiler JR (1999) Variation of atmospheric CO2 by ventilation of the ocean’s deepestwater. Paleoceanography 14: 571–588.
Tripati AK, Eagle RA, Thiagarajan N, et al. (2010) 13C-18O isotope signatures and‘clumped isotope’ thermometry in foraminifera and coccoliths. Geochimica etCosmochimica Acta 74: 5697–5717.
van Geen A, McCorkle DC, and Klinkhammer GP (1995) Sensitivity of thephosphate-cadmium-carbon isotope relation in the ocean to cadmium removal bysuboxic sediments. Paleoceanography 10: 159–169.
van Geen A, Zheng Y, Bernhard JM, et al. (2003) On the preservation of laminatedsediments along the western margin of North America. Paleoceanography18: 1098–1114.
Vance D, Scrivner AE, Beney P, Staubwasser M, Henderson GM, and Slowey NC (2004)The use of foraminifera as a record of the past neodymium isotope compositionof seawater. Paleoceanography 19: PA2009.
Wefer G and Berger WH (1991) Isotope paleontology: Growth and composition of extantcalcareous species. Marine Geology 100: 207–248.
Woodruff F, Savin SM, and Douglas RG (1980) Biological fractionation of oxygenand carbon isotopes by recent benthic foraminifera. Marine Micropaleontology5: 3–11.
Yu JM and Elderfield H (2007) Benthic foraminiferal B/Ca ratios reflect deep watercarbonate saturation state. Earth and Planetary Science Letters 258: 73–86.
Yu E-F, Francois R, and Bacon MP (1996) Similar rates of modern and last-glacialocean thermohaline circulation inferred from radiochemical data. Nature379: 689–694.
Yu E-F, Francois R, Bacon MP, et al. (2001) Trapping efficiency of bottom-tetheredsediment traps estimated from the intercepted fluxes of 230Th and 231Pa. Deep SeaResearch Part I: Oceanographic Research Papers 48: 865–889.
Zahn R and Mix AC (1991) Benthic foraminiferal d18O in the ocean’stemperature-salinity-density field: Constraints on Ice Age thermohaline circulation.Paleoceanography 6: 1–20.
Zahn R and Stuber A (2002) Suborbital intermediate water variability inferred frompaired benthic foraminiferal Cd/Ca and d13C in the tropical West Atlantic andlinking with North Atlantic climates. Earth and Planetary Science Letters200: 191–205.
Zarriess M and Mackensen A (2011) Testing the impact of seasonal phytodetritusdeposition on d13C of epibenthic foraminifer Cibicidoides wuellerstorfi: A 31,000year high-resolution record from the northwest African continentalslope. Paleoceanography 26: PA2202.