time-sensitive effects of hypoxia on differentiation of neural stem cells derived from mouse...
TRANSCRIPT

Original Research Paper
Time-sensitive effects of hypoxia ondifferentiation of neural stem cells derivedfrom mouse embryonic stem cells in vitro
Nguyen Huy Binh1, Hitomi Aoki2, Manabu Takamatsu1, Yuichiro Hatano1,Akihiro Hirata1,3, Hiroyuki Tomita1, Akira Hara1
1Department of Tumor Pathology, Gifu University Graduate School of Medicine, Japan, 2Department of Tissueand Organ Development, Gifu University Graduate School of Medicine, Japan, 3Division of Animal Experiment,Life Science Research Center, Gifu University, Japan
Objectives: Oxygen tension is an important component of microenvironment for the differentiation ofembryonic stem cells including neural lineage. However, the comprehensive influence of hypoxia on neuraldifferentiation during embryonic neural development has not yet been examined.Methods: In this study, we investigated the effect of low oxygen levels (5% O2), or hypoxia, in two stages ofneural differentiation in vitro: (1) inducing mouse embryonic stem cells into neural stem cells (NSCs); andthen (2) inducing NSCs into neural progenitor cells in neurospheres.Results: In the first stage, NSCs generation was reduced under hypoxia. Less mature morphologicalchanges (including neural marker) of NSCs were observed, suggesting the prevention of earlydifferentiation under hypoxic conditions. Thus undifferentiated stem cells were maintained in this stage.However, in the second stage, hypoxia induced neural differentiation in neurospheres. Nevertheless, non-neural progenitor cell formation, such as mesoderm progenitor cell lines or epithelial cell lines, wasrestricted by low oxygen tension.Discussions: Our results demonstrate that hypoxia is essential for regulating neural differentiation and showthe different effects on NSC differentiation dependent on the time-course of NSC development. In the earlystage of NSCs induction, hypoxia inhibits neural differentiation and maintains the undifferentiated state; inthe later stage of NSCs induction, hypoxia induces neural differentiation. Our study may contribute to thedevelopment of new insights for expansion and control of neural differentiation.
Keywords: Embryonic stem cells, Neural stem cells, Neural progenitor cells, Hypoxia, Neuronal development
IntroductionIn aerobic organisms, oxygen regulates various intra-
cellular pathways involved in cellular metabolism,
proliferation, survival, and fate, so that it is a critical
factor in tissue and organ morphogenesis during
mammalian embryonic development and throughout
post-natal life.1 Oxygen plays an important role in
regulating the growth and differentiation state of
neural stem cells (NSCs) in the developing embryo
stage of the mammalian central nervous system.1,2
Neural stem cells, which are multipotent precursor cells
that reside in specialized regions of the fetus, have long
life, self-renewable, and generate neurons, astrocytes,
and oligodendrocytes.3 This makes them attractive
entities for study to gain a better understanding of the
effects of oxygen levels on embryonic neural develop-
ment, as well as for possible therapeutic applications.
It is known that, in the early stage of embryonic
development, mammalian blastocysts are exposed to a
low concentration of O2 ranging from 1.5 to 5.3% in
the reproductive tract.4–6 Several studies have revealed
that hypoxia might profoundly influence stem cells
microenvironment and can promote the differentia-
tion of certain types of stem or progenitor cells, while
inhibiting the differentiation of others.7 Thus hypoxia
may be regarded as driving the regulation of stem/
progenitor cell differentiation, especially the regula-
tion of neural differentiation. In vivo, neural differ-
entiation during embryonic development includes two
stages: early stage and later stage of NSCs induction.
In the early stage of NSCs induction, the first NSCs to
arise are primitive NSCs, which can be isolated from
the anterior embryo from embryonic day E5.5 epiblast
until E8.5 neuroectoderm. In the later stage of NSCs
Correspondence to: Akira Hara, Department of Tumor Pathology, GifuUniversity Graduate School of Medicine, Yanagido 1-1, Gifu City, Gifu501-1194, Japan. Email: [email protected]
804� W. S. Maney & Son Ltd 2014DOI 10.1179/1743132814Y.0000000338 Neurological Research 2014 VOL. 36 NO. 9

induction, the transition of primitive NSCs to defini-
tive NSCs in the embryo at approximately E8.5 occurs
and definitive NSCs are present in the brain through-
out the life of the organism.8 In the early stage of neural
induction from embryonic stem cells, the transcription
of stem cell pluripotency genes such as Pou5f1 (Oct4),
Nanog, and Sox2 is not completely suppressed and
researchers have reported that NSCs are less likely to
be generated in low oxygen conditions.9,10 Fur-
thermore, during the later stages of neural induction,
low oxygen tension may activate molecular pathways
that regulate Wnt/beta-catenin, Oct4, and Notch
signaling and exert a positive effect on neural differen-
tiation of ES cells, resulting in a faster commitment
toward neural progenitors.1 Consequently, under-
standing the effects of hypoxia during neural commit-
ment is important for scientific and therapeutic
purposes and may provide new insights into ES cell
proliferation and differentiation. However, studies on
the influence of hypoxia in the early and later stages of
differentiation of NSC derived from ES cells have not
been conducted.
In this study, we demonstrate that hypoxia is
essential for regulating neural differentiation and that
hypoxia has various effects on the NSC differentiation
dependent on the time-course of NSC development.
We expect essential findings for the improvement of
current therapeutic strategies for the differentiation of
NSCs.
Materials and MethodsLow-oxygen cultureIn the normoxic condition, mouse embryonic stem
(mES) cells and neurospheres were placed in a 37uCincubator supplemented with 20% O2 and 5% CO2. In
the hypoxic condition, 5% O2, 5% CO2, and N2 gas
were mixed using compressed air and supplied into a
sealed container with a small outtake valve placed
inside a 37uC incubator (Wakenyaku CO2 incubator
9000E series, Kyoto, Japan).
ES cell cultureGreen fluorescent protein (GFP)-expressing ES cells11
were maintained on gelatin-coated dishes in Dulbecco’s
modified Eagle’s medium (DMEM, Invitrogen, CA,
USA) supplemented with 10% Hyclone Fetal Bovine
Serum (Thermo Scientific, Massachusetts, USA),
1024 M 2-mercaptoethanol, non-essential amino acid
solution (Invitrogen, California, USA), and human
leukemia inhibitory factor (LIF, 20 ng/ml ESGROH,
Millipore, Massachusetts, USA).
For differentiation, 2000 trypsinized mES cells
were placed in six-well plates seeded with mitomycin-
C treated mouse embryonic fibroblasts PA6 cells in
advance and differentiated in a minimal essential
medium (alphaMEM, Invitrogen, California, USA)
supplemented with 10% fetal calf serum (FBS,
Thermo Scientific, USA). Unless otherwise men-
tioned, 20 pM fibroblast growth factor (FGF),
10 pM cholera toxin, and 100 nM dexamethasone
were added from days 0 to 6, days 0 to 3, and days 3
to 6 of the culture, respectively. The medium was
changed twice per week.
Induction of neurospheresTo induce neurospheres, after washing the dish with
phosphate-buffered saline (PBS, Invitrogen, CA,
USA) to remove floating cells in the medium, only
differentiated mES colonies on PA6 were treated with
0.2% trypsin and dissociated into single cells and then
transferred to a growth medium in culture flasks
(Nunclon, Thermo Scientific) at a concentration of
150 000 cells/ml. The detached mES colonies were
mechanically dissociated in serum-free medium con-
sisting of DMEM and F-12 nutrient (1:1, Invitrogen,
La Jolla, CA, USA). The cells were grown in growth
medium (DMEM) and F-12 nutrient, epidermal
growth factor (EGF), FGF (20 ng/ml each; R&D
Systems, Minneapolis, MN, USA), and 2% B27H
supplement (Invitrogen, CA, USA). Half of the
medium was replaced every 2 days with fresh medium
containing the same concentrations of growth factors.
Analysis of neural differentiation in early stage(experiment-1) and later stage (experiment-2) ofNSCs induction under hypoxiaExperiment-1: Undifferentiated mES cells were co-
cultured with PA6 stromal cells for 6 days to induce
NSCs. In the normoxia group, mES was incubated in
20% oxygen from days 0 to 6. In the hypoxia group,
mES cells were incubated in 20% oxygen from days 0
to 1, then incubated in 5% oxygen from days 1 to 6.1,12
An outline of the experiment-1 is shown in Fig. 1A.
Experiment-2: undifferentiated mES cells were co-
cultured with PA6 stromal cells for 6 days to induce
NSCs. Then these cells were enzymatically digested
into single cells and transferred to a growth medium
with EGF, FGF, and B27 supplement in culture
flasks for 6 days to induce neurospheres containing
NSCs and neural progenitor cells. In the normoxia
group, neurospheres were incubated in 20% oxygen
from days 0 to 12. In the hypoxia group, neuro-
spheres were incubated in 20% oxygen from days 0 to
7, then incubated in 5% oxygen from days 7 to 12.1,12
An outline of the experiment-2 is shown in Fig. 1B.
Immunocytochemistry for NSCs in six-well plates(for experiment-1)Cells grown on six-well plates were fixed with 4%
paraformaldehyde (4% PFA) for 20 minutes, then
washed in 0.01 M PBS and incubated in blocking
solution (2% bovine serum albumin – 2% BSA) for
30 minutes. The cells were incubated with anti-Tuj-1
(mouse IgG, 1:1000, Covance, Princeton, NJ, USA)
in 2% BSA/PBS overnight at 4uC. The secondary
Binh et al. Time-sensitive effects of hypoxia on stem cells
Neurological Research 2014 VOL. 36 NO. 9 805

antibody Alexa Fluor 568 (red) goat anti-mouse IgG
(1:1,000, Invitrogen, Eugene, OR, USA) was used to
visualize the signal for 60 minutes at room temperature.
After washing with PBS, the cell nuclei were stained
with 49,6-diamino-2-phenylindole (DAPI, 1:1000,
Wako Pure Chemical, Osaka, Japan) for 5 minutes at
room temperature.
Immunohistochemistry for NSCs in neurospheres(for experiment-2)Neurospheres were collected and fixed overnight in 10%
phosphate-buffered formalin (pH 7.0), embedded in
paraffin, and then sectioned. The deparaffinized sec-
tions were blocked to endogenous peroxidase activity
by incubation in distilled water containing 3% hydrogen
peroxide for 5 minutes. Antigen retrieval was per-
formed using 0.01 M citrate buffer (pH 6.0) by the
Pascal heat-induced target retrieval system (DAKO,
Glostrup, Denmark). Non-specific binding sites were
blocked in 0.01 M PBS containing 2% BSA (Wako
Pure Chemical, Osaka, Japan) for 60 minutes.
Anti-Desmin (mouse IgG, 1:100; DAKO, Glostrup,
Denmark), anti-Cytokeratin AE1/3 (mouse IgG, 1:100;
DAKO, Glostrup, Denmark), anti-Ki-67 (rat IgG,
clone TEC-3, 1:100, DAKO, Glostrup, Denmark),
and anti-hypoxia-inducible factor-1alpha (anti-HIF-
1alpha) (mouse IgG, 1:500; Millipore, Massachusetts,
USA) in 2% BSA/PBS were added to the slides and
incubated overnight at 4uC. Primary antibodies were
detected with biotinylated anti-mouse IgG (Elite
PK 6102 Mouse IgG Vestastain ABC kit Vector
Laboratories, Burlingame, CA, USA) and biotinylated
anti-rat IgG (1:200, DAKO, Glostrup, Denmark) for
30 minutes, respectively, followed by incubation with
avidin-coupled peroxidase (Vectastain ABC kit, Vector
Laboratories, Burlingame, CA, USA) for 30 minutes.
The peroxidase binding sites were detected by staining
with 3,39-diaminobenzidine (DAB) in 50 mM Tris–
EDTA buffer (pH 7.6). Finally, counterstaining was
performed using Mayer’s hematoxylin. For negative
controls, the primary antibody was substituted with
Figure 1 (A) Experiment-1: undifferentiated mouse embryonic stem (mES) cells were co-cultured with PA6 stromal cells for
6 days to induce neural stem cells (NSCs). Normoxia group: from days 0 to 6, incubated in 20% O2 Hypoxia group: from days 0
to 1, incubated in 20% O2; from days 1 to 6, incubated in 5% O2. (B) Experiment-2: undifferentiated mES cells were co-cultured
with PA6 stromal cells for 6 days to induce NSCs. Then these cells were enzymatically digested into single cells and transferred
to a growth medium with fibroblast growth factor (FGF), epidermal growth factor (EGF), and B27 supplement in culture flasks
for 6 days to induce neurospheres containing NSCs and neural progenitor cells. Normoxia group: from days 0 to 12, incubated
in 20% O2. Hypoxia group: from days 0 to 7, incubated in 20% O2; from days 7 to 12, incubated in 5% O2.
Binh et al. Time-sensitive effects of hypoxia on stem cells
806 Neurological Research 2014 VOL. 36 NO. 9

the buffer (PBS) or with non-immune immunoglo-
bulin G.
Immunofluorescence chemistry for NSCs inneurospheres (for experiment-2)The first antibodies, anti-Nestin (rabbit IgG, 1:200;
Immuno-Biological Laboratory, Gunma, Japan), anti-
microtubule associated protein-2 (anti-MAP2) (mouse
IgG, 1:500; Sigma-Aldrich, St. Louis, MO, USA), and
anti-glial fibrillary acidic protein (anti-GFAP) (rabbit
IgG, 1:500; DAKO, Glostrup, Denmark) in 2% BSA/
PBS were added to the slides and incubated overnight
at 4uC. The second antibodies, Alexa Fluor 568 (red)
goat anti-mouse IgG (1:1,000, Invitrogen, Eugene,
OR, USA) and Alexa Fluor 488 (green) goat anti-
rabbit IgG (1:1,000, Invitrogen, Eugene, OR, USA)
were used to visualize the signal for 60 minutes at
room temperature. After washing with PBS, the cell
nuclei were stained with 4,6-diamino-2-phenylindole
(DAPI, 1:1,000, Wako Pure Chemical, Osaka, Japan)
for 5 minutes at room temperature.
Cell counts and statistical analysisPhotographs for immunohistochemistry staining were
taken under a microscope with a high-resolution digital
camera (Olympus, Tokyo, Japan). Fluorescence was
also photographed under a fluorescence microscope
with a high-resolution digital camera (Olympus,
Tokyo, Japan). The number of immunoreactive cells
in five visual fields (50–100 cells per field) in each
sample was counted in a randomized fashion. All
results are expressed as mean¡standard error (SE).
Reverse Transcription Polymerase ChainReaction (RT-PCR)Total RNA was extracted from cells using the RNeasy
Mini kit (Qiagen, Valencia, CA, USA). Total RNA
(0.5 mg each) was reverse transcribed using Superscript
III Reverse Transcriptase (Invitrogen, Carlsbad, CA,
USA). Quantitative real-time PCR was carried out
with the Thermal Cycler Dice Real Time System Single
(Takara, Kyoto, Japan) using the SYBR Green
(Takara, Kyoto, Japan) method. The primers were
as follows:
MAP2 (ATGACAGGCAAGTCGGTGAAG and
TTGAGTCCACTGGTCGAGGTT)
GFAP (CGGAGACGCATCACCTCTG and TG-
GAGGAGTCATTCGAGACAA)
Nestin (GTGCCTCTGGATGATG and TTGAC-
CTTCCTCCCCCTC)
Oct4 (ACCAGTTGCCATTGGTGGAAA and C-
ATGAGGAGAGTCCGGTACTT)
HIF-1alpha (GTCCCAGCTACGAAGTTACAG-
C and CAGTGCAGGATACACAAGGTTT)
Beta-actin (ATGGAGCCACCGATCCACA and
CATCCGTAAAGACCTCTATGCCAAC)
For evaluation of gene expression, beta-actin was
used as internal control.
ResultsThe hypoxic condition inhibited generation ofNSC from mES cells co-cultured with PA6To investigate the effect of hypoxia on generation of
NSCs from mES cells, we incubated mES cells co-
cultured with PA6 for 5 days under the hypoxic or
normoxic condition (Fig. 1A). This culture technique
is known as an efficient method for induction of
NSCs from mES cells under the normoxic condition.6
Morphological differences of the outgrowth cells
were easily recognizable between the two conditions
as prominent neurite outgrowth emerging from the
colonies was observed in the normoxic culture
whereas less frequent neurite outgrowth was seen in
the hypoxic culture (Fig. 2D and E).
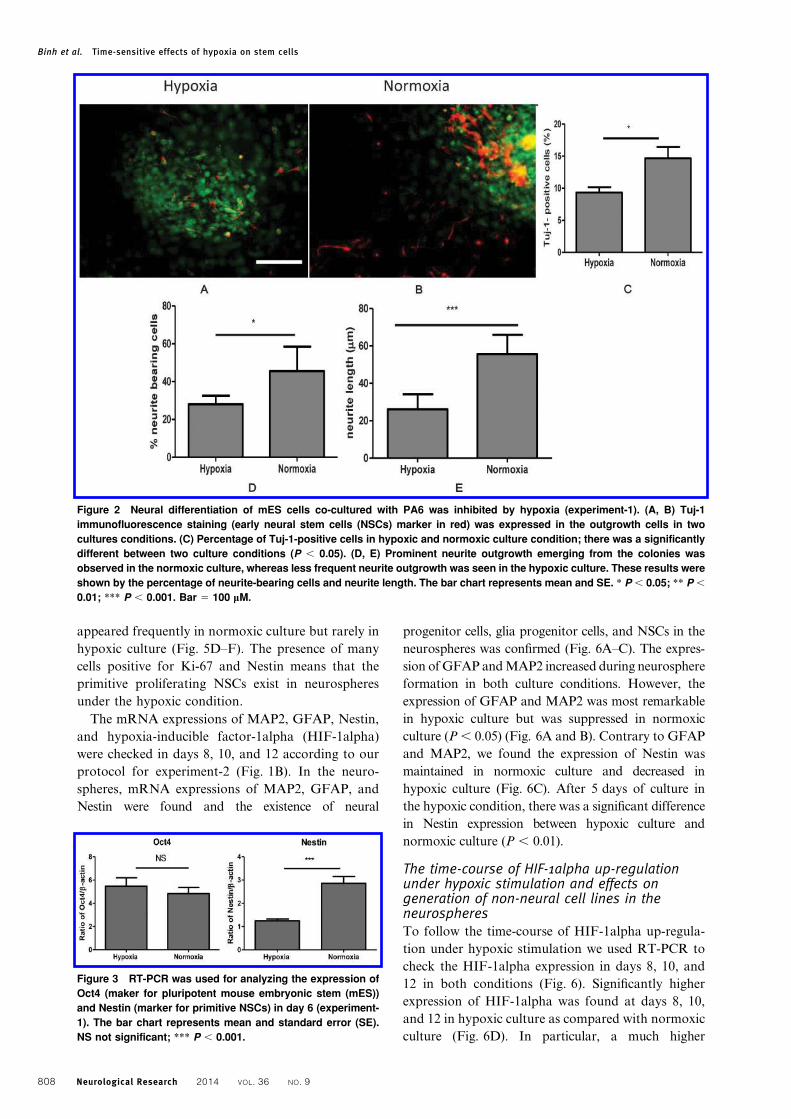
Neurite outgrowths from NSC colonies on PA6
feeder were assessed by the immunocytochemical
method. Tuj-1 (early NSC marker) positive cells
appeared prominently in normoxic cell culture, while
a limited number of Tuj-1-positive cells were
observed in hypoxic cell culture (Fig. 2C). The results
from RT-PCR (Fig. 3) showed the expressions of
Nestin (NSC marker) were most remarkable in
normoxic culture and suppressed in a hypoxic culture
(P , 0.001). These results suggested that hypoxia
inhibited the generation of NSCs from mES cells.
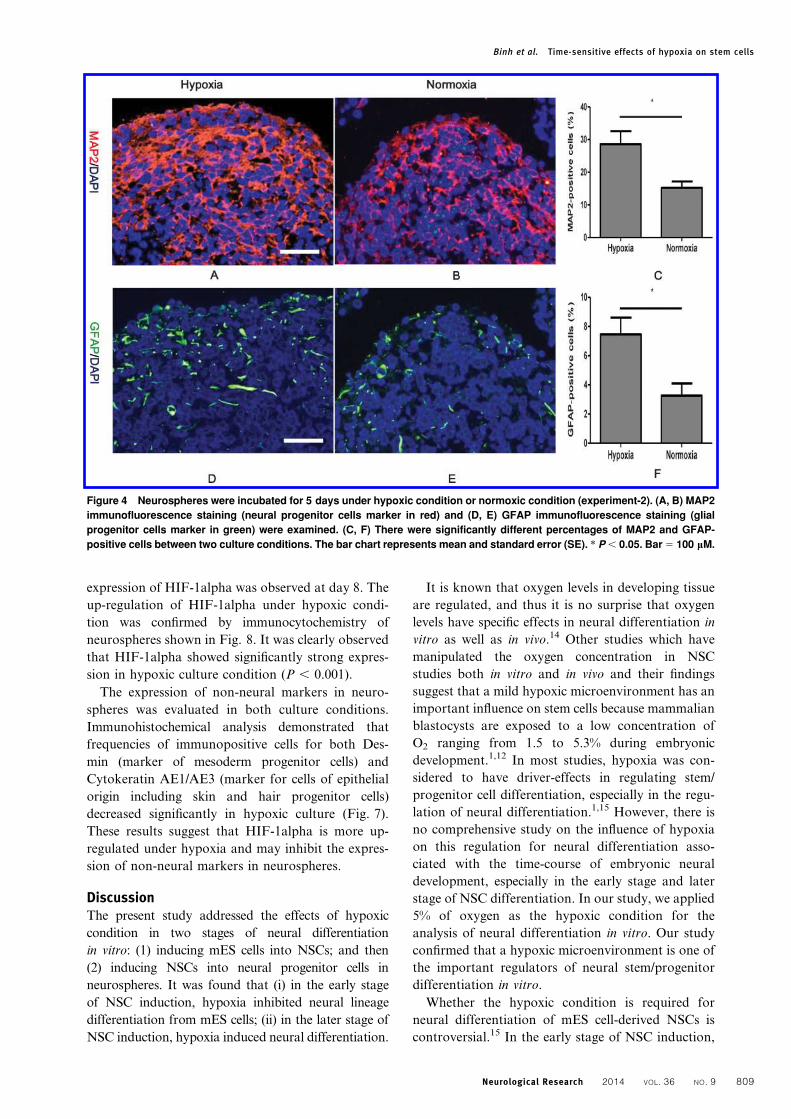
Hypoxic condition promoted the generation ofneural progenitor cells from NSCs contained inneurospheresNext, to determine the effect of hypoxia on genera-
tion of neural progenitor cells from NSCs in neuro-
spheres, we generated neurospheres composed of
free-floating clusters of NSCs supplied with EGF,
FGF, and B27 in hypoxic or normoxic conditions
(Fig. 1B). This culture technique is known as an
efficient method for induction of neural progenitor
cells from NSCs.13 Immunohistochemical staining
examinations showed that neurospheres in hypoxic
culture were composed of more MAP2 (neural
progenitor cells marker) positive cells and GFAP
(glial progenitor cells marker) positive cells as com-
pared with those in normoxic culture (Fig. 4A, B, D,
and E). The frequencies of MAP2-positive cells in
hypoxic culture and normoxic culture were approxi-
mately 28.6 and 15.2%, respectively (Fig. 4C).
Similarly, the frequencies of GFAP-positive cells in
hypoxic culture and normoxic culture were approxi-
mately 7.4 and 3.2%, respectively (Fig. 4F). There
was a significant difference in the percentages of
GFAP and MAP2 expressing cells between the two
culture conditions (P , 0.05). In addition, over 50%
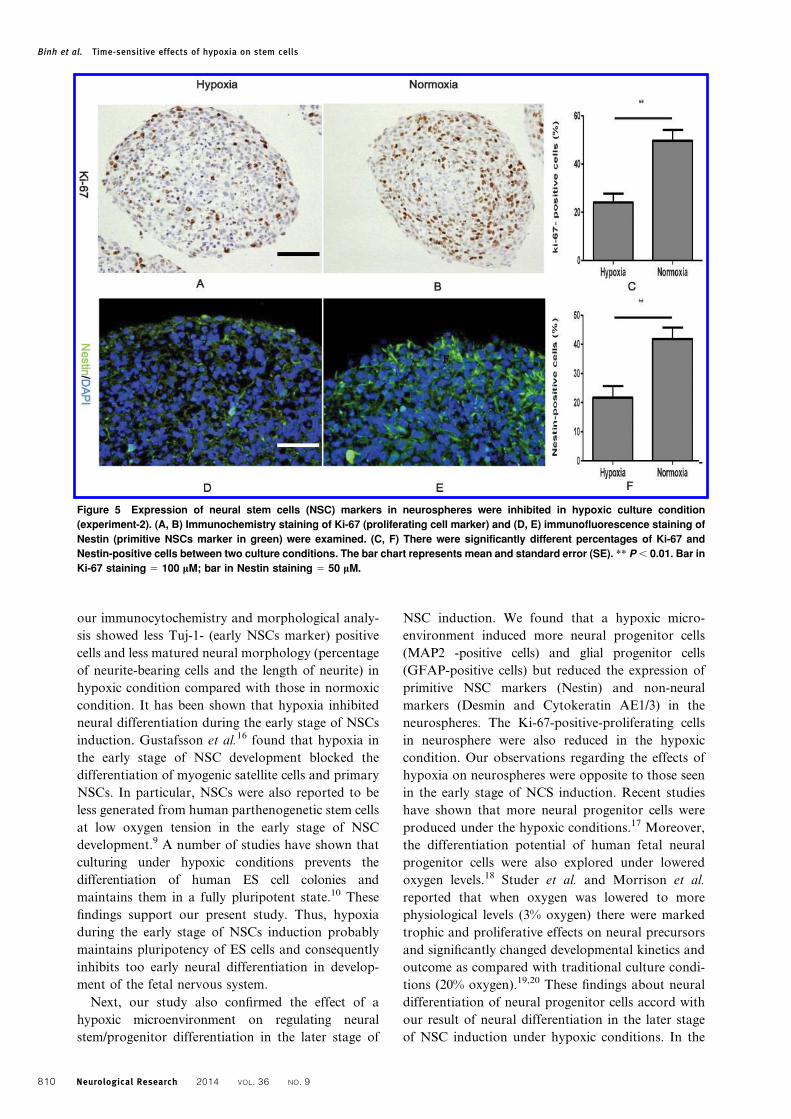
of the cells in the neurospheres in normoxic culture
were ki-67 (proliferating cell marker) positive,
whereas only about 20% positive cells were found in
hypoxic culture (Fig. 5A–C). Furthermore, Nestin
(primitive NSC marker) immunopositive cells
Binh et al. Time-sensitive effects of hypoxia on stem cells
Neurological Research 2014 VOL. 36 NO. 9 807
appeared frequently in normoxic culture but rarely in
hypoxic culture (Fig. 5D–F). The presence of many
cells positive for Ki-67 and Nestin means that the
primitive proliferating NSCs exist in neurospheres
under the hypoxic condition.
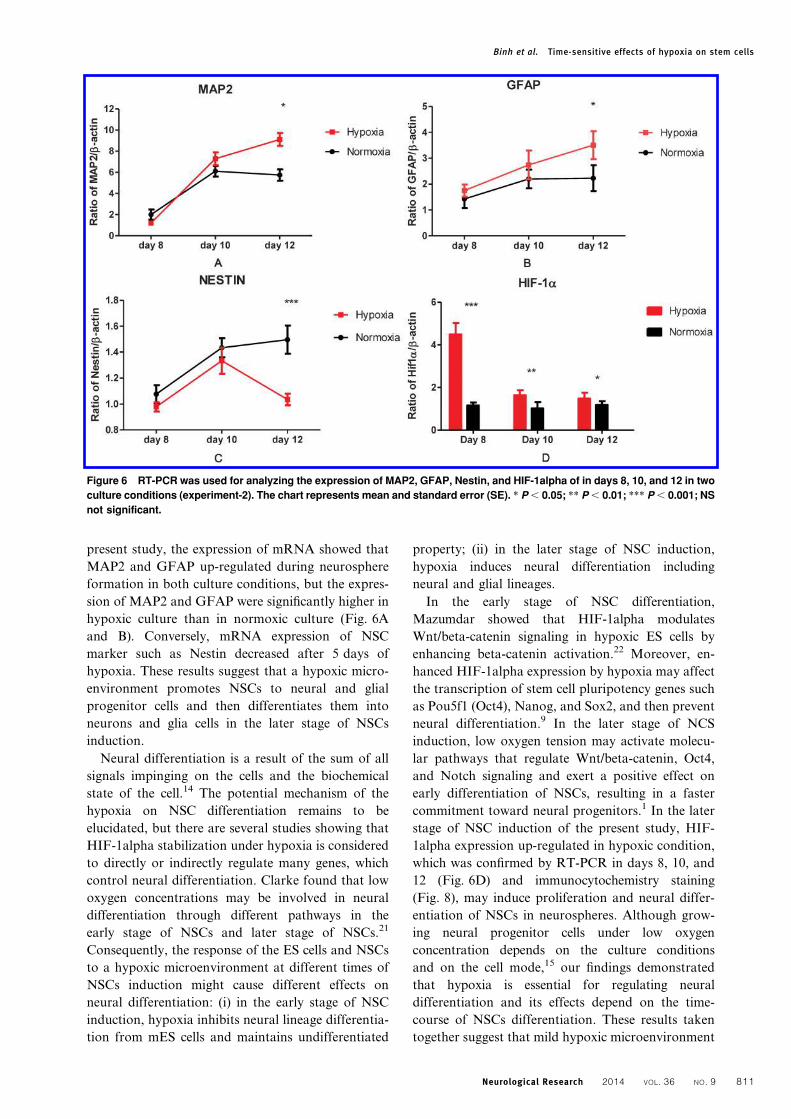
The mRNA expressions of MAP2, GFAP, Nestin,
and hypoxia-inducible factor-1alpha (HIF-1alpha)
were checked in days 8, 10, and 12 according to our
protocol for experiment-2 (Fig. 1B). In the neuro-
spheres, mRNA expressions of MAP2, GFAP, and
Nestin were found and the existence of neural
progenitor cells, glia progenitor cells, and NSCs in the
neurospheres was confirmed (Fig. 6A–C). The expres-
sion of GFAP and MAP2 increased during neurosphere
formation in both culture conditions. However, the
expression of GFAP and MAP2 was most remarkable
in hypoxic culture but was suppressed in normoxic
culture (P , 0.05) (Fig. 6A and B). Contrary to GFAP
and MAP2, we found the expression of Nestin was
maintained in normoxic culture and decreased in
hypoxic culture (Fig. 6C). After 5 days of culture in
the hypoxic condition, there was a significant difference
in Nestin expression between hypoxic culture and
normoxic culture (P , 0.01).
The time-course of HIF-1alpha up-regulationunder hypoxic stimulation and effects ongeneration of non-neural cell lines in theneurospheresTo follow the time-course of HIF-1alpha up-regula-
tion under hypoxic stimulation we used RT-PCR to
check the HIF-1alpha expression in days 8, 10, and
12 in both conditions (Fig. 6). Significantly higher
expression of HIF-1alpha was found at days 8, 10,
and 12 in hypoxic culture as compared with normoxic
culture (Fig. 6D). In particular, a much higher
Figure 3 RT-PCR was used for analyzing the expression of
Oct4 (maker for pluripotent mouse embryonic stem (mES))
and Nestin (marker for primitive NSCs) in day 6 (experiment-
1). The bar chart represents mean and standard error (SE).
NS not significant; *** P , 0.001.
Figure 2 Neural differentiation of mES cells co-cultured with PA6 was inhibited by hypoxia (experiment-1). (A, B) Tuj-1
immunofluorescence staining (early neural stem cells (NSCs) marker in red) was expressed in the outgrowth cells in two
cultures conditions. (C) Percentage of Tuj-1-positive cells in hypoxic and normoxic culture condition; there was a significantly
different between two culture conditions (P , 0.05). (D, E) Prominent neurite outgrowth emerging from the colonies was
observed in the normoxic culture, whereas less frequent neurite outgrowth was seen in the hypoxic culture. These results were
shown by the percentage of neurite-bearing cells and neurite length. The bar chart represents mean and SE. * P , 0.05; ** P ,
0.01; *** P , 0.001. Bar 5 100 mM.
Binh et al. Time-sensitive effects of hypoxia on stem cells
808 Neurological Research 2014 VOL. 36 NO. 9
expression of HIF-1alpha was observed at day 8. The
up-regulation of HIF-1alpha under hypoxic condi-
tion was confirmed by immunocytochemistry of
neurospheres shown in Fig. 8. It was clearly observed
that HIF-1alpha showed significantly strong expres-
sion in hypoxic culture condition (P , 0.001).
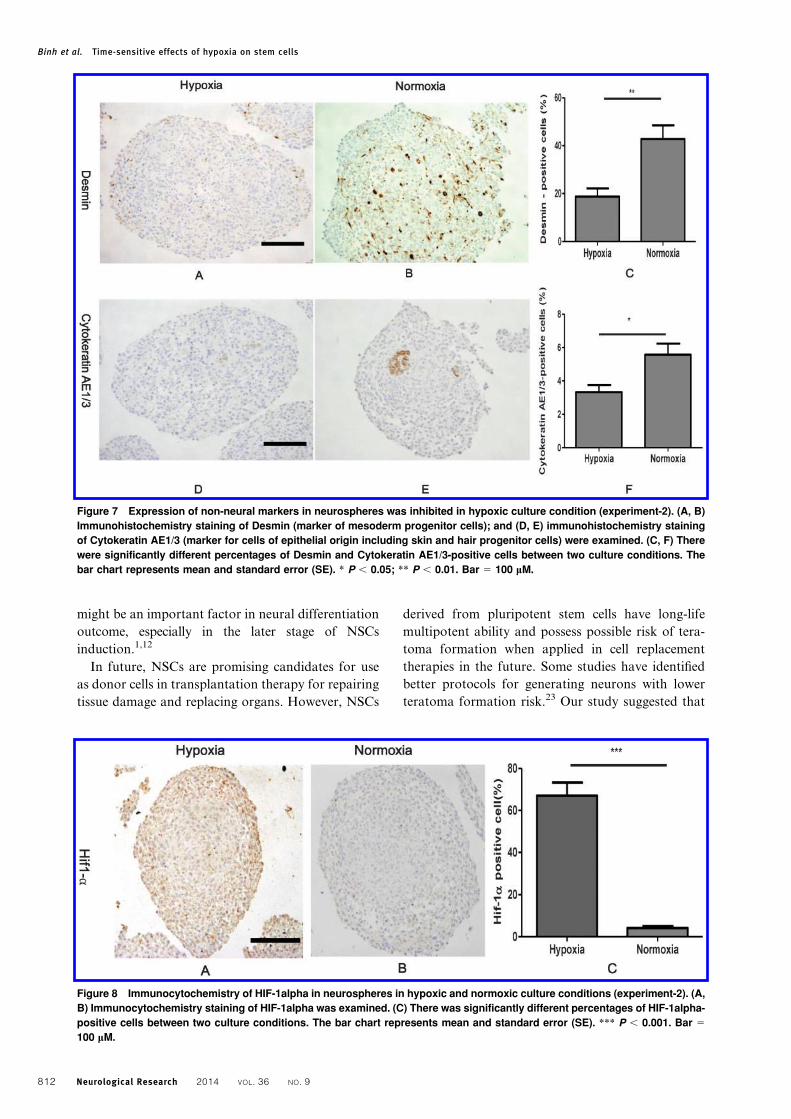
The expression of non-neural markers in neuro-
spheres was evaluated in both culture conditions.
Immunohistochemical analysis demonstrated that
frequencies of immunopositive cells for both Des-
min (marker of mesoderm progenitor cells) and
Cytokeratin AE1/AE3 (marker for cells of epithelial
origin including skin and hair progenitor cells)
decreased significantly in hypoxic culture (Fig. 7).
These results suggest that HIF-1alpha is more up-
regulated under hypoxia and may inhibit the expres-
sion of non-neural markers in neurospheres.
DiscussionThe present study addressed the effects of hypoxic
condition in two stages of neural differentiation
in vitro: (1) inducing mES cells into NSCs; and then
(2) inducing NSCs into neural progenitor cells in
neurospheres. It was found that (i) in the early stage
of NSC induction, hypoxia inhibited neural lineage
differentiation from mES cells; (ii) in the later stage of
NSC induction, hypoxia induced neural differentiation.
It is known that oxygen levels in developing tissue
are regulated, and thus it is no surprise that oxygen
levels have specific effects in neural differentiation in
vitro as well as in vivo.14 Other studies which have
manipulated the oxygen concentration in NSC
studies both in vitro and in vivo and their findings
suggest that a mild hypoxic microenvironment has an
important influence on stem cells because mammalian
blastocysts are exposed to a low concentration of
O2 ranging from 1.5 to 5.3% during embryonic
development.1,12 In most studies, hypoxia was con-
sidered to have driver-effects in regulating stem/
progenitor cell differentiation, especially in the regu-
lation of neural differentiation.1,15 However, there is
no comprehensive study on the influence of hypoxia
on this regulation for neural differentiation asso-
ciated with the time-course of embryonic neural
development, especially in the early stage and later
stage of NSC differentiation. In our study, we applied
5% of oxygen as the hypoxic condition for the
analysis of neural differentiation in vitro. Our study
confirmed that a hypoxic microenvironment is one of
the important regulators of neural stem/progenitor
differentiation in vitro.
Whether the hypoxic condition is required for
neural differentiation of mES cell-derived NSCs is
controversial.15 In the early stage of NSC induction,
Figure 4 Neurospheres were incubated for 5 days under hypoxic condition or normoxic condition (experiment-2). (A, B) MAP2
immunofluorescence staining (neural progenitor cells marker in red) and (D, E) GFAP immunofluorescence staining (glial
progenitor cells marker in green) were examined. (C, F) There were significantly different percentages of MAP2 and GFAP-
positive cells between two culture conditions. The bar chart represents mean and standard error (SE). * P , 0.05. Bar 5 100 mM.
Binh et al. Time-sensitive effects of hypoxia on stem cells
Neurological Research 2014 VOL. 36 NO. 9 809
our immunocytochemistry and morphological analy-
sis showed less Tuj-1- (early NSCs marker) positive
cells and less matured neural morphology (percentage
of neurite-bearing cells and the length of neurite) in
hypoxic condition compared with those in normoxic
condition. It has been shown that hypoxia inhibited
neural differentiation during the early stage of NSCs
induction. Gustafsson et al.16 found that hypoxia in
the early stage of NSC development blocked the
differentiation of myogenic satellite cells and primary
NSCs. In particular, NSCs were also reported to be
less generated from human parthenogenetic stem cells
at low oxygen tension in the early stage of NSC
development.9 A number of studies have shown that
culturing under hypoxic conditions prevents the
differentiation of human ES cell colonies and
maintains them in a fully pluripotent state.10 These
findings support our present study. Thus, hypoxia
during the early stage of NSCs induction probably
maintains pluripotency of ES cells and consequently
inhibits too early neural differentiation in develop-
ment of the fetal nervous system.
Next, our study also confirmed the effect of a
hypoxic microenvironment on regulating neural
stem/progenitor differentiation in the later stage of
NSC induction. We found that a hypoxic micro-
environment induced more neural progenitor cells
(MAP2 -positive cells) and glial progenitor cells
(GFAP-positive cells) but reduced the expression of
primitive NSC markers (Nestin) and non-neural
markers (Desmin and Cytokeratin AE1/3) in the
neurospheres. The Ki-67-positive-proliferating cells
in neurosphere were also reduced in the hypoxic
condition. Our observations regarding the effects of
hypoxia on neurospheres were opposite to those seen
in the early stage of NCS induction. Recent studies
have shown that more neural progenitor cells were
produced under the hypoxic conditions.17 Moreover,
the differentiation potential of human fetal neural
progenitor cells were also explored under lowered
oxygen levels.18 Studer et al. and Morrison et al.
reported that when oxygen was lowered to more
physiological levels (3% oxygen) there were marked
trophic and proliferative effects on neural precursors
and significantly changed developmental kinetics and
outcome as compared with traditional culture condi-
tions (20% oxygen).19,20 These findings about neural
differentiation of neural progenitor cells accord with
our result of neural differentiation in the later stage
of NSC induction under hypoxic conditions. In the
Figure 5 Expression of neural stem cells (NSC) markers in neurospheres were inhibited in hypoxic culture condition
(experiment-2). (A, B) Immunochemistry staining of Ki-67 (proliferating cell marker) and (D, E) immunofluorescence staining of
Nestin (primitive NSCs marker in green) were examined. (C, F) There were significantly different percentages of Ki-67 and
Nestin-positive cells between two culture conditions. The bar chart represents mean and standard error (SE). ** P , 0.01. Bar in
Ki-67 staining 5 100 mM; bar in Nestin staining 5 50 mM.
Binh et al. Time-sensitive effects of hypoxia on stem cells
810 Neurological Research 2014 VOL. 36 NO. 9
present study, the expression of mRNA showed that
MAP2 and GFAP up-regulated during neurosphere
formation in both culture conditions, but the expres-
sion of MAP2 and GFAP were significantly higher in
hypoxic culture than in normoxic culture (Fig. 6A
and B). Conversely, mRNA expression of NSC
marker such as Nestin decreased after 5 days of
hypoxia. These results suggest that a hypoxic micro-
environment promotes NSCs to neural and glial
progenitor cells and then differentiates them into
neurons and glia cells in the later stage of NSCs
induction.
Neural differentiation is a result of the sum of all
signals impinging on the cells and the biochemical
state of the cell.14 The potential mechanism of the
hypoxia on NSC differentiation remains to be
elucidated, but there are several studies showing that
HIF-1alpha stabilization under hypoxia is considered
to directly or indirectly regulate many genes, which
control neural differentiation. Clarke found that low
oxygen concentrations may be involved in neural
differentiation through different pathways in the
early stage of NSCs and later stage of NSCs.21
Consequently, the response of the ES cells and NSCs
to a hypoxic microenvironment at different times of
NSCs induction might cause different effects on
neural differentiation: (i) in the early stage of NSC
induction, hypoxia inhibits neural lineage differentia-
tion from mES cells and maintains undifferentiated
property; (ii) in the later stage of NSC induction,
hypoxia induces neural differentiation including
neural and glial lineages.
In the early stage of NSC differentiation,
Mazumdar showed that HIF-1alpha modulates
Wnt/beta-catenin signaling in hypoxic ES cells by
enhancing beta-catenin activation.22 Moreover, en-
hanced HIF-1alpha expression by hypoxia may affect
the transcription of stem cell pluripotency genes such
as Pou5f1 (Oct4), Nanog, and Sox2, and then prevent
neural differentiation.9 In the later stage of NCS
induction, low oxygen tension may activate molecu-
lar pathways that regulate Wnt/beta-catenin, Oct4,
and Notch signaling and exert a positive effect on
early differentiation of NSCs, resulting in a faster
commitment toward neural progenitors.1 In the later
stage of NSC induction of the present study, HIF-
1alpha expression up-regulated in hypoxic condition,
which was confirmed by RT-PCR in days 8, 10, and
12 (Fig. 6D) and immunocytochemistry staining
(Fig. 8), may induce proliferation and neural differ-
entiation of NSCs in neurospheres. Although grow-
ing neural progenitor cells under low oxygen
concentration depends on the culture conditions
and on the cell mode,15 our findings demonstrated
that hypoxia is essential for regulating neural
differentiation and its effects depend on the time-
course of NSCs differentiation. These results taken
together suggest that mild hypoxic microenvironment
Figure 6 RT-PCR was used for analyzing the expression of MAP2, GFAP, Nestin, and HIF-1alpha of in days 8, 10, and 12 in two
culture conditions (experiment-2). The chart represents mean and standard error (SE). * P , 0.05; ** P , 0.01; *** P , 0.001; NS
not significant.
Binh et al. Time-sensitive effects of hypoxia on stem cells
Neurological Research 2014 VOL. 36 NO. 9 811
might be an important factor in neural differentiation
outcome, especially in the later stage of NSCs
induction.1,12
In future, NSCs are promising candidates for use
as donor cells in transplantation therapy for repairing
tissue damage and replacing organs. However, NSCs
derived from pluripotent stem cells have long-life
multipotent ability and possess possible risk of tera-
toma formation when applied in cell replacement
therapies in the future. Some studies have identified
better protocols for generating neurons with lower
teratoma formation risk.23 Our study suggested that
Figure 8 Immunocytochemistry of HIF-1alpha in neurospheres in hypoxic and normoxic culture conditions (experiment-2). (A,
B) Immunocytochemistry staining of HIF-1alpha was examined. (C) There was significantly different percentages of HIF-1alpha-
positive cells between two culture conditions. The bar chart represents mean and standard error (SE). *** P , 0.001. Bar 5
100 mM.
Figure 7 Expression of non-neural markers in neurospheres was inhibited in hypoxic culture condition (experiment-2). (A, B)
Immunohistochemistry staining of Desmin (marker of mesoderm progenitor cells); and (D, E) immunohistochemistry staining
of Cytokeratin AE1/3 (marker for cells of epithelial origin including skin and hair progenitor cells) were examined. (C, F) There
were significantly different percentages of Desmin and Cytokeratin AE1/3-positive cells between two culture conditions. The
bar chart represents mean and standard error (SE). * P , 0.05; ** P , 0.01. Bar 5 100 mM.
Binh et al. Time-sensitive effects of hypoxia on stem cells
812 Neurological Research 2014 VOL. 36 NO. 9

hypoxia could be considered as a potential element
for controlling proliferation and differentiation of
NSCs possessing multipotent ability.
In conclusion, we showed that hypoxia is essential
for regulating neural differentiation and we demon-
strated the different effects on the NSC differentia-
tion occurring during development. These results
highlight the importance of oxygen homeostasis in
regulating neural differentiation in cell fate commit-
ment and maturation. It is suggested that oxygen
tension control may be crucial for neural embryonic
development and neural generation in vitro. The
hypoxic culture of the present study expanded ES
cells into neuron progenitor cells with low risk of
teratoma formation and also provided a useful and
effective tool for future cell therapy.
Disclaimer statementsContributors None.
Funding None.
Conflicts of interest No competing financial inter-
ests exist.
Ethics approval We fully complied with the
‘Guidelines Concerning Experimental Animals’ issued
by the Japanese Association for Laboratory Animal
Science and exercised due consideration so as not to
cause any ethical problems.
AcknowledgementsThe authors are indebted to Ms. Kyoko Takahashi
for her excellent technical assistance. This work was
supported by grants from the Ministry of Education,
Culture, Sports, Science and Technology of Japan.
References1 De Filippis L, Delia D. Hypoxia in the regulation of neural
stem cells. Cell Mol Life Sci. 2011;68:2831–44.2 Panchision DM. The role of oxygen in regulating neural stem
cells in development and disease. J Cell Physiol. 2009;220:562–8.3 Breunig JJ, Haydar TF, Rakic P. Neural stem cells: historical
perspective and future prospects. Neuron. 2011;70:614–25.4 Burton G.J. Oxygen, the Janus gas; its effects on human
placental development and function. J Anat. 2009;215:27–35.5 Fischer B, Bavister BD. Oxygen tension in the oviduct and
uterus of rhesus monkeys, hamsters and rabbits. J ReprodFertil. 1993;99:673–9.
6 Jauniaux E, Watson A, Ozturk O, et al. In-vivo measurement ofintrauterine gases and acid-base values in early humanpregnancy. Hum Reprod. 1999;14:2901–4.
7 Simon MC, Keith B. The role of oxygen availability inembryonic development and stem cell function. Nat Rev MolCell Biol. 2008;9:285–96.
8 Hitoshi S, Seaberg R, Koscik C et al. Primitive neural stem cellsfrom the mammalian epiblast differentiate to definitive neuralstem cells under the control of Notch signaling. Genes Dev.2004;18:1806–11.
9 Abramihina TV, Isaev DA, Semechkin RA. Effect of hypoxiaon neural induction in colonies of human parthenogenetic stemcells. Bull Exp Biol Med. 2012;154:130–2.
10 Ezashi T, Das P, Roberts RM. Low O2 tensions and theprevention of differentiation of hES cells. Proc Natl Acad SciUSA. 2005;102:4783–8.
11 Hirano M, Yamamoto A, Yoshimura N, et al. Generation ofstructures formed by lens and retinal cells differentiating fromembryonic stem cells. Dev Dyn. 2003;228:664–71.
12 Santilli G, Lamorte G, Carlessi L. Mild hypoxia enhancesproliferation and multipotency of human neural stem cells.PLoS One.. 2010;5(1):e8575.
13 Kitajima H, Yoshimura S, Kokuzawa J, et al. (2005). Culturemethod for the induction of neurospheres from mouseembryonic stem cells by coculture with PA6 stromal cells. JNeurosci Res. 2005;80:467–74.
14 Prithi R, Evan S. Neuronal stem cells and their manipulation.In: Robert L and Irina K. Essential Stem Cell Methods:Elsevier, 2009: pp. 23–51.
15 Vieira HL, Alves PM, Vercelli A. Modulation of neuronal stemcell differentiation by hypoxia and reactive oxygen species. ProgNeurobiol. 2011;93(3):444–55.
16 Zheng X, Pereira T, et al. Hypoxia requires notch signaling tomaintain the undifferentiated cell state. Dev Cell. 2005;9:617–28.
17 Zhang CP, Zhu LL, Zhao T, et al. Characteristics of NeuralStem Cells Expanded in Lowered Oxygen and the PotentialRole of Hypoxia-Inducible Factor-1Alpha. Neurosignals.2007;15:259–65.
18 Giese AK, Frahm J, Hubner R, et al. Erythropoietin and theeffect of oxygen during proliferation and differentiation ofhuman neural progenitor cells. BMC Cell Biology. 2010;11:94.
19 Morrison SJ, Csete M, Groves AK, et al. Culture in reducedlevels of oxygen promotes clonogenic sympathoadrenal differ-entiation by isolated neural crest stem cells. J Neurosci.2000;20:7370–6.
20 Studer L, Csete M, Lee SH, et al. Enhanced proliferation,survival, and dopaminergic differentiation of CNS precursorsin lowered oxygen. J Neurosci. 2000;20:7377–83.
21 Clarke L, van der Kooy D. Low Oxygen Enhances Primitiveand Definitive Neural Stem Cell Colony Formation byInhibiting Distinct Cell Death Pathways. Stem Cells.2009;27:1879–86.
22 Mazumdar J, O’Brien WT, Johnson RS, et al. (2010). O2regulates stem cells through Wnt/b-catenin signalling. Nat CellBiol. 2010;12:1007–13.
23 Hara A, Taguchi A, Aoki H, et al. Folate antagonist,methotrexate induces neuronal differentiation of humanembryonic stem cells transplanted into nude mouse retina.Neurosci Lett. 2010;477(3):138–43.
Binh et al. Time-sensitive effects of hypoxia on stem cells
Neurological Research 2014 VOL. 36 NO. 9 813