tim-3 signaling in peripheral nk cells promotes maternal ... · tim-3 signaling in peripheral nk...
TRANSCRIPT

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
IMMUNOLOGY
1Laboratory for Reproductive Immunology, Hospital ofObstetrics andGynecology, FudanUniversity Shanghai Medical College, Shanghai 200011, China. 2Key Laboratory of Repro-duction Regulation of National Population and Family Planning Commission, ShanghaiInstitute of Planned Parenthood Research, Institute of Reproduction and Development,Fudan University, Shanghai 200032, China. 3Shanghai Key Laboratory of Female Repro-ductiveEndocrineRelatedDiseases, Shanghai 200011,China. 4KeyLaboratoryof StemCellBiology, Institute of Health Sciences, Shanghai Jiao Tong University School of Medicineand Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Universityof Chinese Academy of Sciences, Shanghai 200031, China. 5Department of Immunol-ogy, School of Basic Medical Sciences, Fudan University, Shanghai 200032, China.6In Vitro Fertilization–Embryo Transfer Center, Hospital of Obstetrics and Gynecology,Fudan University Shanghai Medical College, Shanghai 200011, China. 7Department ofObstetrics, Hospital of Obstetrics and Gynecology, Fudan University Shanghai Med-ical College, Shanghai 200011, China.*These authors contributed equally to this work.†Corresponding author. Email: [email protected] (X.C.); [email protected] (M.D.)
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
Copyright © 2017
The Authors, some
rights reserved;
exclusive licensee
American Association
for the Advancement
of Science. No claim
to original U.S.
Government Works
http://stke.D
ownloaded from
Tim-3 signaling in peripheral NK cells promotesmaternal-fetal immune tolerance and alleviatespregnancy lossYanhong Li,1,2,3* Jiayuan Zhang,4* Di Zhang,1,2,3 Xiaowu Hong,5 Yu Tao,1 Songcun Wang,1,2,3
Yuanyuan Xu,1 Hailan Piao,1 Weijie Yin,4 Min Yu,1,6 Yin Zhang,1,7 Qiang Fu,1 Dajin Li,1,2,3
Xing Chang,4† Meirong Du1,2,3†
Pregnancy loss occurs in about 15% of clinically recognized pregnancies, and defective maternal-fetal immunetolerance contributes to more than 50% of these events. We found that signaling by the type I membraneprotein T cell immunoglobulin and mucin-containing protein 3 (Tim-3) in natural killer (NK) cells had an essen-tial protective role during early pregnancy. Tim-3 on peripheral NK (pNK) cells was transiently increased inabundance during the first trimester of pregnancy, which depended on interleukin-4 (IL-4)–signal transducerand activator of transcription 6 (STAT6) and progesterone signaling. Tim-3+ pNK cells displayed immunosuppressiveactivities, including the production of anti-inflammatory cytokines and the induction of regulatory T cells (Tregs) in atransforming growth factor–b1 (TGF-b1)–dependent manner. Tim-3 on pNK cells was stimulated by its ligandgalectin-9 (Gal-9), leading to signaling by the kinases c-Jun N-terminal kinase (JNK) and AKT. In recurrent mis-carriage (RM) patients, Tim-3 abundance on pNK cells was reduced and the immunosuppressive activity of Tim-3+
pNK cells was impaired. Compared to Tim-3+ pNK cells from donors with normal pregnancies, RM patient Tim-3+
pNK cells exhibited changes in DNA accessibility in certain genetic loci, which were reversed by inhibiting accessiblechromatin reader proteins. Furthermore, Tim-3+ pNK cells, but not Tim-3− pNK cells, reduced fetal loss in abortion-prone and NK cell–deficient mice. Together, our findings reveal a critical role for Tim-3–Gal-9 signaling–mediatedimmunoregulation by pNK cells in maternal-fetal immune tolerance and suggest that Tim-3 abundance on pNK cellsis a potential biomarker for RM diagnosis.
scie
on January 27, 2020ncemag.org/
INTRODUCTIONHow the semiallogeneic fetus can develop without being attacked or re-jected by the maternal immune system has been a fascinating questionin reproductive biology and immunology. Complex immunoregulationis required for maternal immune tolerance toward the semiallogeneicfetus and the maintenance of pregnancy (1, 2). Once maternal-fetal im-mune tolerance is disturbed, various complications of pregnancy occur(3, 4). Spontaneous abortion is the most common complication of ges-tation, occurring in about 15%of humanpregnancies (5).When a spon-taneous abortion occurs, the risk of subsequent miscarriage is markedlyincreased. Recurrent miscarriage (RM) is defined as two or moreconsecutive miscarriages. More than 50% of these events remain un-explained after standard investigations (6). There is also no reliable pre-dictor or effective prevention strategy for RM. A systematic review andmeta-analysis demonstrated that the most commonly used biomarkers[serum human chorionic gonadotropin (hCG) and progesterone] are
not useful in predicting pregnancy outcome (7). Therefore, an accuratediagnostic biomarker to predict miscarriage is urgently needed.
Multiple mechanisms for evading the rejection of embryos by thematernal immune system have been proposed (8–10). As a critical reg-ulator of active immune tolerance, inducible regulatory T cells (iTregs)express the transcription factor Foxp3 and increase in number in theperipheral blood and decidua during pregnancy, playing indispensableroles in the generation of maternal-fetal tolerance in mice and humans(11, 12). Natural killer (NK) cells with a high functional diversity havemany regulatory roles in pregnancy. Althoughmuch attention has beenpaid to studying decidual NK (dNK) cells in pregnancy because of theirmarked abundance (13–15), NK cells in the peripheral blood (pNK)may also play indispensable roles in pregnancy. These cells participatein increasing the number of T helper 2 (TH2) cells while decreasing thenumber of TH1 cells during gestation (16, 17). In addition, an increasedproportion of CD56+CD16+NK cells is found in womenwith recurrentpregnancy loss, which is accompanied by an increase in the cytotoxicactivity of pNK cells (18–20). Nevertheless, the precise role of pNK cellsduring normal pregnancy (NP) and their regulatory mechanisms re-main poorly understood.
T cell immunoglobulin and mucin-containing protein 3 (Tim-3), atype I membrane protein, was initially identified on terminally differen-tiated TH1 cells. The identification of galectin-9 (Gal-9) as a ligand forTim-3 has now firmly established the Gal-9–Tim-3 interaction as animportant regulator of TH1 immunity by inducing TH1 cell apoptosis(21, 22). Tim-3 is also found onTH17 andT cytotoxic 1 cells. Tim-3 actsas a negative regulatorymolecule to suppress cell-mediated immune re-sponses and promote immune tolerance (23–25). The engagement ofTim-3 by Gal-9 suppresses allograft rejection and improves the sur-vival of allogeneic skin grafts (26). Because embryos express paternal
1 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
ohttp://stke.sciencem
ag.org/D
ownloaded from
antigens, NP is often regarded as a successful allogeneic transplan-tation. In this regard, it is tempting to explore whether Gal-9–Tim-3signaling plays a role in protecting the fetus from being attacked bythe maternal immune system.
In addition to being found on T cells, Tim-3 is also found on innateimmune cells, including mast cells, macrophages, dendritic cells (DCs),and NK cells (23, 25, 27). Tim-3 not only serves as a marker of cellularactivation or maturation but also suppresses NK cell cytotoxicity (28).An increase in the cell surface abundance of Tim-3 on NK cells leads toNK cell dysfunction during chronic viral infections (29, 30). Therefore,Tim-3 exerts diverse regulatory effects on NK cells under differentimmune contexts, involving numerous pathological and physiologicalprocesses. Whether Tim-3–mediated NK cell functions influencematernal-fetal immune tolerance and pregnancy outcome is currentlyunclear.
Here, we investigated the role of Tim-3 on pNK cells during earlypregnancy inmice.We found that Tim-3+NKcells proliferate in the firsttrimester through signaling by interleukin-4 (IL-4) and signal transducerand activator of transcription 6 (STAT6) or by progesterone signaling.Tim-3+ NK cells displayed tolerant phenotypes and induced the differ-entiation of naïve CD4+ T cells into iTregs in a transforming growthfactor–b1 (TGF-b1)–dependent manner. Furthermore, in patients withRM, NK cells had reduced cell surface expression of Tim-3, and Tim-3+
NK cells produced less anti-inflammatory cytokines and lost their abilityto induce iTreg generation. A loss of the suppressive activity of Tim-3+
NK cells in RMwas associated with a global change in chromatin acces-sibility, which was partially reversed by the inhibition of accessible chro-matin reader proteins. In abortion-prone (AP) or NK cell–deficientmice, the adoptive transfer ofTim-3+NKcells reduced fetal loss, whereasthe transfer of Tim-3− NK cells had no effect. Our data suggest thattheGal-9–Tim-3 axis plays a role in themaintenance of NP through theregulation of pNK cell functions, suggesting that Tim-3+ pNK cellsmight be a promising biomarker for the prediction of RM.
n January 27, 2020
RESULTSTim-3+ NK cells are transiently increased in number inperipheral blood during early human pregnancyTo characterize Tim-3 function in maternal-fetal immune tolerance,we first analyzed the expression of Tim-3 in peripheral blood mono-nuclear cells (PBMCs) during pregnancy. Leukocytes, NK cells, CD11c+
DCs, and CD14+ monocytes all showed a substantial cell surfaceabundance of Tim-3 (fig. S1, A and B). Although DCs and mono-cytes expressed Tim-3, the percentages of these cell populations thatwere Tim-3 did not vary during pregnancy (fig. S1C). In contrast, thepercentage of Tim-3+ NK cells in the first trimester of pregnancy wasconsiderably greater than that in either nonpregnant donors or those inthe second trimester (Fig. 1, A and B). Among pNK cells (including twomajor subsets, CD56dim andCD56highNKcells), nearly 90%ofCD56dim
NK cells expressed Tim-3, whereas only about 40% of CD56high NKcells were Tim-3+ (Fig. 1C). Together, these data suggest that thenumber of Tim-3+ NK cells is transiently increased in the peripheralblood during the first trimester of pregnancy, a critical period forestablishing maternal immune tolerance to the fetus.
Tim-3+ NK cells display immunosuppressive activityduring pregnancyTo further investigate the role of Tim-3+ NK cells in human pregnancy,we analyzed the cytokines that they produce. Compared to Tim-3−NK
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
cells, Tim-3+ NK cells produced more anti-inflammatory cytokines,including TGF-b1, IL-10, and IL-4, but less of the pro-inflammatorytumor necrosis factor–a (TNF-a) (Fig. 1, D and E). The abundance ofperforin (Fig. 1E) and the extent of cytotoxicity against HTR-8/SVneocells (a first trimester human trophoblast cell line) (Fig. 1F) were alsosubstantially reduced in Tim-3+NK cells compared to Tim-3−NK cells.Similar findings were obtained from a comparison of the abundances ofthe mRNAs encoding these cytokines between these two subpopula-tions of NK cells (Fig. 1G). These results suggest that Tim-3+ NK cellshave a robust immunosuppressive activity, which was further con-firmed by microarray analysis (Fig. 1, H and I). A gene ontology(GO) analysis revealed that cytokines and cytokine activity representeda major transcriptional difference between Tim-3+ and Tim-3− pNKcells in early pregnancy (fig. S2). These data suggest that Tim-3+ pNKcells have a high capacity to produce anti-inflammatory cytokines and adiminished cytotoxicity toward trophoblasts. These traits are conduciveto an immune tolerant microenvironment.
The cell surface abundance of Tim-3 on NK cells is enhancedthrough IL-4–STAT6 or progesterone signalingWe next investigated the factors that contributed to the increasedabundance of Tim-3 on NK cells in early pregnancy. Because of thepredominance of TH2 over TH1 cytokines during normal gestation(31, 32), we treated pNK cells with the TH2 cytokine IL-4 and foundthat Tim-3 abundance was substantially increased (Fig. 2A). Treat-ment with the TH1 cytokine IFN-g reduced the abundance of Tim-3under both basal and IL-4–stimulated conditions (Fig. 2A).Moreover,IL-4 activity was dependent on its downstream transcription factorSTAT6, because a STAT6 inhibitor (A77-1726) abrogated the in-creased Tim-3 abundance induced by IL-4 (Fig. 2, B and C). We alsoobserved the potential regulatory effects of pregnancy-associated hor-mones on Tim-3 abundance on NK cells. Progesterone exerted a bi-phasic regulatory effect on Tim-3 abundance, which depended on itsconcentration (Fig. 2D). Progesterone at concentrations of 0.01 to1.0 nM increased the abundance of Tim-3 on pNK cells in a dose-dependent manner, whereas greater concentrations of progesterone(>100 nM) had an inhibitory effect, suggesting that physiologicalconcentrations of progesterone promoted the cell surface expressionof Tim-3. However, estradiol and b-hCG did not affect Tim-3 abun-dance on NK cells (fig. S3). Together, these data suggest that theabundance of Tim-3 on pNK cells increased during early pregnancythrough IL-4–STAT6 or progesterone signaling.
Gal-9–Tim-3 interactions induce the immunosuppressiveactivity of NK cells in maternal bloodPrevious studies indicated that the immunoregulatory effect of Tim-3depends on the interaction with its ligand, Gal-9 (21, 33, 34). Thus, wetreated pNK cells with recombinant human Gal-9 (rhGal-9) and de-tected substantial increases in the amounts of TGF-b1 and IL-10produced by pNK cells but a decreased amount of TNF-a production(Fig. 2E). The activity of rhGal-9 was dependent on Tim-3, because ananti–Tim-3 neutralizing antibody blocked rhGal-9 activity, whereas aneutralizing antibody against CD44, another target receptor for Gal-9,failed to abrogate the effect of rhGal-9 onNK cells (Fig. 2E). Similarly,rhGal-9 suppressed the production of perforin (Fig. 2E) and inhibitedpNK cytotoxicity (Fig. 2F), and these effects were abrogated by theanti–Tim-3 antibody but not the anti-CD44 antibody. Therefore,these data suggest that Gal-9 might modulate pNK cell functiontoward an immune tolerant state by binding Tim-3.
2 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
To identify the signaling pathways downstream of Gal-9–Tim-3, westimulated pNK cells with rhGal-9 and examined the phosphorylationof key kinases involved inNK cell function.We found that rhGal-9 sub-stantially increased the abundances of total AKT and phosphorylatedAKT (pAKT), as well as of phosphorylated c-Jun N-terminal kinase(pJNK) and phosphorylated extracellular signal–regulated kinase 1/2(pERK1/2) (Fig. 2G). Furthermore, the specific inhibition of the JNKor AKT signaling pathways, but not ERK1/2 signaling, eliminated theeffect of rhGal-9 on pNK cells (Fig. 2H). These results suggest that theGal-9–Tim-3 interactionmay induce a tolerant phenotype in pNK cellsthrough the activation of JNK and AKT signaling. In addition to thesedownstream signaling molecules, we also analyzed the expression ofgenes encoding transcription factors that are associated with NK cell
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
development. After rhGal-9 stimulation, the abundances of mRNAsof ID2, BLIMP1, andGATA3were substantially increased, whereas thatof EOMESwasmarkedly reduced (Fig. 2I). Similar results were found atthe protein level (Fig. 2J). The altered expression of ID2, BLIMP1,GATA3, and EOMES induced by rhGal-9 was abrogated by the specificinhibition of JNK or AKT (Fig. 2K). All ex vivo analyses suggested aGal-9–dependentmechanism, whereby Tim-3 inhibits pNK cell activity.
The immunosuppressive activity of Tim-3+ NK cells isimpaired in patients with RMTo evaluate the potential relevance of Tim-3+ NK cells inmaternal-fetaltolerance, we first quantified these cells in patients with RM by flow cy-tometry. Although the frequency of Tim-3+ NK cells was comparable
A
D
E F
C
Tim-3 Non 1st 2nd
β-Tubulin
B
G
H I
–3.0 –2.0 –1.0 0.0 1.0 2.0 3.0 4.0 5.0 6.0 7.0
0 2
4
6 8
10
13
16
19
22
25
Tim-3+ NK Tim-3– NK (n = 865 )
Log2 Fold change (Tim3+/Tim3–)
–Log
10 (P
)
(n = 100)
R2
R1
CD3
CD
56
Tim-3
89.3%
38.6% CD
56
CD56
TGF-
β1
Tim-3+ NK
Tim-3– NK
IL-1
0
IL-4
TNF-
α
IFN
-γ
Per
forin
43.1% 2.4% 15.9%
3.37%
28.5% 8.63%
37.4% 8.51%
59.6%
72.7% 0.52% 30.1%
Fig. 1. Human Tim-3+ NK cells increase in num-ber and have an immune tolerant phenotypeduring early pregnancy. (A to C) Tim-3 abun-dance on NK cells is increased during the first tri-mester of pregnancy. (A and B) The percentage ofperipheral CD56+ cells from nonpregnant do-nors (Non, 10 donors), first-trimester donors(1st, 30 donors), and second-trimester donors(2nd, 9 donors) that were Tim-3+ was deter-mined by flow cytometric analysis (A), whereasthe abundance of Tim-3 in the same cells wasdetermined by Western blotting analysis (B). (C)The percentages of CD3−CD56+, CD3−CD56dim,and CD3−CD56bright pNK cells in early pregnancythat were Tim-3+ were determined by flow cytom-etry. Data are from 30 donors. (D and E) HumanTim-3+ pNK cells show an immunosuppressivephenotype. Purified Tim-3+ or Tim-3− CD56+ pNKcells were stimulated with phorbol 12-myristate13-acetate, ionomycin, and brefeldin A for 4 hours.The cells were analyzed by flow cytometry to de-tect the indicated cytokines and perforin. (D)Numbers in the plots show the percentage of cellsin the boxed regions from a representative exper-iment. IFN-g, interferon-g. (E) Data are means ±SEM of the percentage of cells positive for the in-dicatedmarker from six independent experiments.(F) Tim-3+ NK cells have a reduced cytotoxicitytoward trophoblast cells. Cytotoxicity was deter-mined with CytoTox 96 Non-Radioactive Cyto-toxicity Assay. The trophoblast cell line HTR-8/SVneowas used as the target (T) cell for NK cells. Data aremeans ± SEM of nine samples per group. (G) Tim-3+
and Tim-3− pNK cells were analyzed by real-timepolymerase chain reaction (PCR) to determine the re-lative abundance of the indicated mRNAs. Data aremeans ± SEM of three experiments. *P < 0.05, **P <0.01, and ***P < 0.001 by Student’s t test. (H andI) Tim-3+ and Tim-3− NK cells were sorted fromthe PBMCs of a pregnant donor at first trimester.RNA was extracted, and global gene expressionwas analyzed by microarray. Data are represen-tative of six donors. The unique gene signaturesbased on a >2-fold difference in expression forTim-3− NK cells (100 probes, gray) and Tim-3+
NK cells (865 probes, yellow) are highlighted. (I)The heat map depicts the expression of genes as-sociated with immune tolerance and activation inTim-3− and Tim-3+ NK cells.
3 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
between patients with RM and those control donors with NPs, the cellsurface abundance of Tim-3 [as assessed by measurement of the meanfluorescence intensity (MFI)] on pNK cells was substantially decreasedin patients with RM (Fig. 3A). Furthermore, the plasma Gal-9 concen-tration (Fig. 3B) and the amount of intracellular Gal-9 produced in
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
peripheral immune cells (Fig. 3C) were reduced in patients with RMcompared to controls.
Compared toTim-3+NK cells from control donorswithNPs, Tim-3+
pNK cells from patients with RMproduced less of the anti-inflammatorycytokinesTGF-b1, IL-10, and IL-4butmorepro-inflammatory cytokines,
ID2
GATA3 EOMES GAPDH
BLIMP1
Ctrl Gal-9
37 KDa
17 KDa 90 KDa 48 KDa 17 KDa
pSTAT6 STAT6
β-Tubulin
110 kDa 100 kDa 54 kDa
GAPDH
pJNK JNK
pAKT AKT
NF-κB pNF-κB
ERK
pERK
Gal-9
110 kDa 100 kDa
37 kDa
60 kDa 60 kDa 48 kDa 48 kDa 44 kDa
44 kDa
0 10 30 60 (min)
IL-4+vehicle IL-4+A77-1726
Tim
-3
CD56
32.1% 57.4%
Tim
-3
Ctrl IL-4 IFN-γ IL-4+IFN-γ
CD56
63.4% 28.1% 17.1% 36.9%
JI
F
H
E
G
DC
A B
K
analyzed by real-time PCR to determine the relative abundances of the indicated mRNAs (I) and by Western blotting
Fig. 2. The IL-4–STAT6pathwayandprogester-one promote the increased number of Tim-3+ NKcells in early pregnancy, whereas Gal-9–Tim-3interactions suppress NK cell activation. (A andB) NK cells isolated from nonpregnant donorswere left untreated (Ctrl) or were treated with IL-4(100 ng/ml), IFN-g (35 ng/ml), or both for 48 hours.The cells were then analyzed by flow cytometry todetermine thepercentageof Tim-3+ cells (A) andbyWestern blotting to detect the phosphorylationof STAT6 (B). pSTAT6, phosphorylated STAT6;tSTAT6, total STAT6. (C) NK cells were stimulatedwith IL-4 alone or in the presence of the STAT6 in-hibitor A77-1726 for 48 hours. The cells were thenanalyzed by flow cytometry to determine the per-centage of Tim-3+ cells. *P < 0.05, **P < 0.01, and***P < 0.001 by analysis of variance (ANOVA).DMSO, dimethyl sulfoxide. (D) NK cells stimulatedwith the indicated concentrations of progesteronewere analyzed by flow cytometry to determinethe percentage of Tim-3+ cells. ***P < 0.001 byStudent’s t test. Data in (A) to (D) are means ±SEM of three independent experiments. (E) pNKcells from early pregnant donors were stimulatedby rhGal-9 alone or in the presence of blocking anti-bodies against Tim-3 or CD44, as indicated, for48 hours. The cells were then analyzed by flow cy-tometry to determine the percentage of cells pos-itive for the indicated cytokines or perforin. Dataare means ± SEM of 12 donors. IgG, immuno-globulin G. (F) NK cells were treated with rhGal-9in the presence or absence of the indicatedblocking antibodies before being incubated withtarget cells at the indicated ratios to determinecytotoxicity as described inMaterials andMethods.Data are means ± SEM of four experiments. *P <0.05, **P < 0.01, and ***P < 0.001 by ANOVA.(G) NK cells were stimulated with rhGal-9 for theindicated times before being analyzed by Westernblotting with antibodies against the indicated pro-teins. Bar graphs show pooled densitometric datafrom three experiments. *P < 0.05, **P < 0.01, and***P < 0.001 by Student’s t test. pNF-kB, phosphory-lated nuclear factor kB; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NS, not significant.(H) Purified NK cells were stimulated with rhGal-9in 15 mMLY294002, 10 mMSP600125, 10 mMU0126,or DMSO, as indicated. The cells were analyzed byflow cytometry to determine the percentage of cellspositive for the indicated cytokines or perforin. Dataaremeans ± SEMof 10 experiments. *P< 0.05, **P<0.01, ***P < 0.001, and ****P < 0.0001 by ANOVA.(I and J) NK cells were left unstimulated or werestimulated with rhGal-9 for 48 hours and then
with antibodies against the indicated proteins (J).*P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001 by Student’s t test. (K) NK cells were left untreated or treated with rhGal-9 in the presence or absence of the indicatedinhibitors. The relative abundances of the indicated mRNAs were then determined by real-time PCR analysis. Data in (I) to (K) are means ± SEM of three experiments. *P <0.05, **P < 0.01, and ***P < 0.001 by ANOVA.
4 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
including TNF-a and IFN-g, as well as more perforin (Fig. 3D). Thecytotoxicity of Tim-3+ pNK cells from patients with RM was alsoenhanced (Fig. 3E), suggesting that there were functional defects inthe Tim-3+ NK cells of patients with RM. However, cytokine produc-tion by the Tim-3−NK cells and their cytotoxicity were comparable be-tween RMpatients and donors withNPs (Fig. 3, D and E). Accordingly,the immunosuppressive activity of Tim-3+ NK cells was markedly re-duced in patients with RM, which may account for the uncontrolledallogeneic response to the fetus.
Chromatin accessibility of genes encoding anti- andpro-inflammatory cytokines is altered in Tim-3+ NK cellsfrom patients with RMTo systemically characterize the difference in Tim-3+ pNK cellsbetween RM patients and donors with NPs, we assessed the land-
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
scape of chromatin accessibility in theirTim-3+ and Tim-3−NK cells. An assayfor transposase-accessible chromatinsequencing (ATAC-seq) was used toanalyze global chromatin accessibility(35). To determine whether globalchromatin accessibility reflected thedifferences among samples, we per-formed a principal components analy-sis (PCA). We found that the samplesformed three well-separated groups,including Tim-3+ NK cells from RMpatients, Tim-3−NK cells from RM pa-tients, and NK cells (either Tim-3+ orTim-3−) from normal donors (Fig. 4A).PC1, which accounted for 50% of thetotal variance, separated the Tim-3+ cellsfrom the Tim-3− NK cells of RM pa-tients. PC2, which accounted for 15%of the variance, distinguished the NKcells of normal donors from those ofRM patients (Fig. 4A). Note that theNK cell subsets from the same normaldonor tended to cluster together, sug-gesting that individual differences over-shadowed the differences betweenTim-3+ and Tim-3− cells in normal do-nors. Similarly, hierarchical clusteringconfirmed the separation between pa-tient NK cells and normal donor NKcells (Fig. 4B). These results suggest thatthe chromatin accessibility of pNK cellscan capture different pregnancy out-comes. Thus, these findings suggest thatTim-3+ pNKcells represent an indicatorof pregnancy status, which promptedour desire to investigate the underlyingmechanism by which Tim-3+ NK cellsare involved in gestation.
We next compared the Tim-3+ NKcells from RM patients to those fromnormal donors and identified 260 re-gions with statistically significantly re-duced accessibility but 360 regions
with significantly increased accessibility in the cells from the RM pa-tients (P < 0.05) (Fig. 4C). On the basis of published chromatin immu-noprecipitation sequencing (ChIP-seq) results for human CD56+ NKcells, the regions with reduced accessibility in the RM patients were as-sociated with H3K4me1hi H3K4me3low (the two forms of methylationof Lys4 on histone H3) histone modifications and were distal totranscription start sites (TSSs), suggesting that these regions weremainly enhancers. In contrast, regions with increased accessibility inthe RM patient cells were enriched for H3K4me1low H3K4m3hi andwere proximal to the TSS, suggesting that they weremainly promoters(Fig. 4, D and E). These data suggest that promoters and enhancerswere regulated by different mechanisms in Tim-3+ NK cells.
Furthermore, one of the regions with the most diminished accessi-bility in the Tim-3+ NK cells from RM patients was the 3′ end of theTGFB1 gene, which was associated with a reduced promoter accessibility
Fig. 3. Tim-3+ NK cells from RM patients have reduced immunosuppressive activity. (A) The relative abundances ofTim-3 onpNK cells fromdonorswithNP andpatientswith RMwere determined by flow cytometry. Data aremeans ± SEMof20 donors per group. (B) The concentrations of soluble Gal-9 in the plasma of NP donors and RM patients were determinedby enzyme-linked immunosorbent assay. Data aremeans ± SEMof six NP donors and eight RM patients. (C) PBMCs fromNPdonors and RMpatients were analyzed by flow cytometry to determine the percentages of Gal-9+ cells among the indicatedimmune cell subsets. Data aremeans ± SEM of six NP donors and eight RM patients. (D) Tim-3+ NK cells (left) and Tim-3−NKcells (right) from NP donors and RM patients were analyzed by flow cytometry to determine the percentages of cells thatwere positive for the indicated cytokines or perforin. Data are means ± SEM of 8 NP donors and 10 RM patients. (E) Tim-3+
andTim-3−NKcells fromNPdonors andRMpatientswere analyzed todetermine their cytotoxicity, as describedearlier.Dataare means ± SEM of nine donors per group. *P < 0.05, **P < 0.01, and ***P < 0.001 by Student’s t test.
5 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
of TGFB1, suggesting that this region functions as an enhancer tocoordinate with the TGFB1 promoter (Fig. 4, C and F). This enhancerin the Tim-3−NK cells from RM patients showed a similar accessibilityto that of the enhancer in Tim-3− NK cells from normal donors, sug-gesting that a cell-specific mechanism regulates TGFB1 expression inTim-3+ NK cells (Fig. 4F). Similarly, the accessibility of the IL10 locuswas also reduced in the Tim-3+ NK cells from RM patients (Fig. 4G).These data suggest that Tim-3 activation in NK cells during pregnancy
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
promotes the accessibility of loci encoding anti-inflammatory cytokines,which enhances their expression and induces immune tolerance to thefetus. A decreased expression of Tim-3, together with other defects,may inhibit the accessibility of these genes encoding anti-inflammatorycytokines in RM patients and contribute to immune-based rejection ofthe fetus.
Chromatin-accessible regions are usually associated with histoneacetylation, which is recognized by bromodomain-containing proteins
A RM Tim-3+ NP Tim-3+
NP Tim-3–
NP1 NP2
RM1
RM2
NP3
RM Tim-3–
–200 –100 0 100 200
200
100
–100
–2
00
0
PC1 (50%)
PC
2 (1
5%)
NP3+
NP1+
NP3–
NP2+
NP2–
RM1+
NP1–
RM2+
RM1–
RM2–
Normal Patients
B
NP
1+
NP
3–
NP
2+
NP
2–
NP
3+
NP
1–
RM
1–
RM
2–
RM
1+
RM
2+
D
-4 -2 0 2 40
2
4
6Tim-3+
Log2 (patient/normal)
P < 0.05
TGF-β1NFIX
POTEAMIR5583-1
F
TGFB1
55
55
55
55
55
55
55
55
3'Enhancer Promoter NP 1
NP 2
RM 1
RM 2 NP 1
NP 2
RM1
RM2
Tim-3
+
Enhancer
IL10
25
25
25
25
25
25
25
25
G
H3K4me1
H3K
4me3
Distance to TSS (bp)
Accessibility unchanged
Accessibility decreased in RM
Accessibility increased in RM C
E
Tim-3
–
H
-Log
10 P
(adj
)
Fig. 4. Changes in chromatin accessibilityin Tim-3+ NK cells are involved in the ab-normal cytokine profile associated withRM. (A and B) Tim-3+ and Tim-3− NK cellswere sorted from NP donors (n = 3) or RMpatients in the first trimester (n = 2). Globalchromatin accessibility was analyzed byATAC-seq and was compared using PCA(A). On the basis of a hierarchical clusteringanalysis, RM patient Tim-3+ NK cells dis-played distinct global chromatin accessibilitycompared with NK cells from NP donors (B).(C) ATAC-seq analysis was used to comparechromatin accessibility between Tim-3+ NKcells from NP donors and RM patients. Reg-ulatory regions showing a statistically sig-nificant difference (P < 0.05) are labeled inred. (D andE) Tim-3+NK cells fromNPdonorsand RM patients were analyzed to assesschromatin accessibility at enhancer and pro-moter regions. Regulatory regions that showedreduced accessibility (left), unaltered accessi-bility (middle), or increased accessibility (right)were analyzed for the presence of H3K4me3andH3K4me1 (D)or for distance to the closestTSS (E). bp, base pair. (F andG) Tim-3+NK cellsfrom NP donors and RM patients were ana-lyzed to assess accessibility at the TGFB1 (F)and IL10 (G) loci. Top to bottom: The genomebrowser tracks ofATAC-seq for Tim-3+NKcellsfrom twohealthydonors and twoRMpatientsand then those for the Tim-3− NK cells fromthe same individuals. The numbers on the leftindicate the ATAC-seq reads after normaliza-tion to 10 × 106 mapped reads. (H) SortedTim-3+ and Tim-3− NK cells from NP donorsand RM patients were treated with the indi-cated concentrations of JQ1, and the percen-tages of the NK cells that were positive forTNF-a (left) and IFN-g (right) were determinedby flow cytometry. Data are means ± SEM ofthree independent experiments. *P < 0.05,**P< 0.01, and ***P< 0.001 by Student’s t test.6 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
(36). To investigate the relationship between altered chromatin accessi-bility and cytokine-encoding gene expression, we treated the Tim-3+ orTim-3−NK cells with JQ1, an inhibitor of the BET family of bromodo-main proteins (37). We observed that the percentages of Tim-3+ NKcells from RM patients that were positive for the inflammatory cyto-kines TNF-a and IFN-g were substantially reduced by JQ1 (Fig. 4H).Treatment with JQ1 also reduced the percentages of Tim-3− NK cellsfromRMpatientswith normal donors thatwere positive for TNF-a andIFN-g, although to a lesser degree. Furthermore, JQ1 had no effect onthe percentages of Tim-3+ NK cells from normal donors that were pos-itive for inflammatory cytokines (Fig. 4H). The production of TGF-b1 and perforin was unaffected by JQ1 (fig. S4). These data suggest thatthe increased pro-inflammatory cytokine production by Tim-3+ NKcells frompatientswithRMwas at least partially caused by the increasedaccessibility of the loci of the corresponding genes.
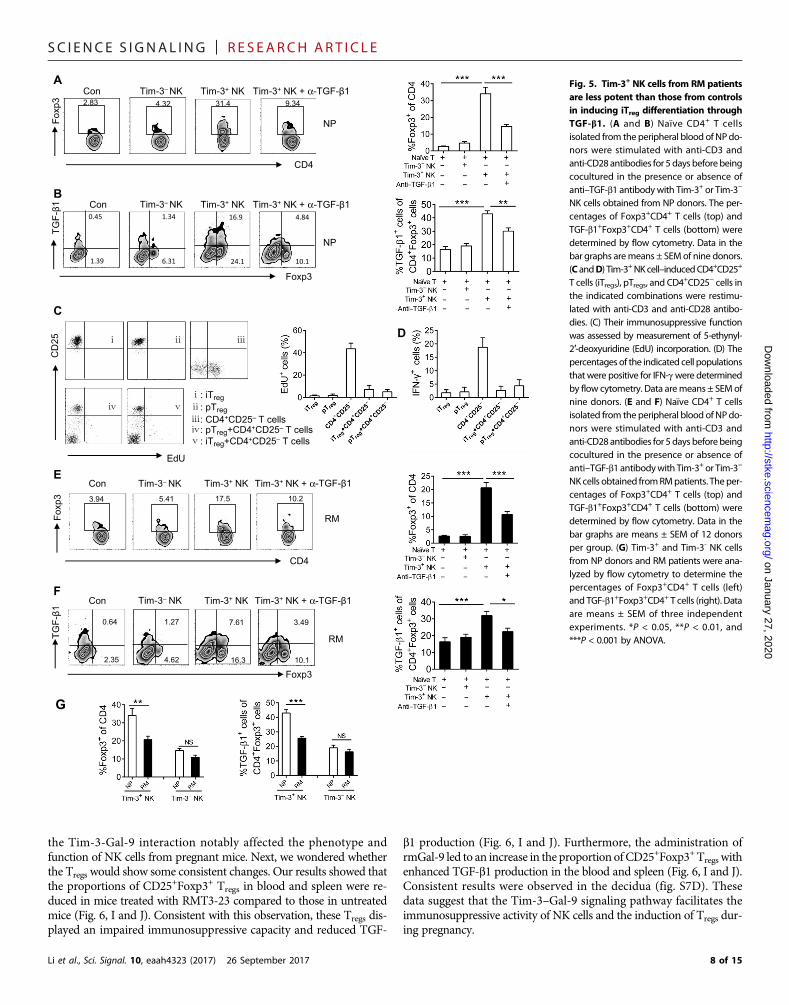
Tim-3+ NK cells from RM patients are less potent than thosefrom normal donors in promoting iTreg differentiationTGF-b1 is a key factor in inducing the differentiation of naïve CD4+ Tcells to iTregs. Our earlier results (Figs. 3D and 4, C and F) showed thatthe Tim-3+ NK cells from normal donors produced large amounts ofTGF-b1 compared to those from RM patients, which also showed themost diminished promoter accessibility of TGFB1. Therefore, we hy-pothesized that Tim-3+ NK cells could promote the generation of iTregs
and that this activity of Tim-3+ NK cells might be impaired in patientswith RM, which could contribute to the pathogenic role of this NK cellpopulation in RM.
To investigate this hypothesis, we first cultured Tim-3+ or Tim-3−
NK cells with CD4+CD45RA+ naïve T cells from the same donor at ear-ly pregnancy. Coculture with Tim-3+NK cells, but not with Tim-3−NKcells, induced the production of Foxp3 in the CD4+ T cells, and this wasinhibited by an anti–TGF-b1 neutralizing antibody (Fig. 5A). The pro-duction of TGF-b1 in the iTregs was also promoted by Tim-3+ NK cells(Fig. 5B). In addition, we investigated the in vitro capacity of the iTregsthat were induced byTim-3+NK cells to inhibit proliferation and IFN-gproduction from CD4+CD25− T cells. We found that the proliferationof CD4+CD25− T cells in response to anti-CD3 and anti-CD28 anti-bodies was markedly inhibited by peripheral naïve CD4+CD25+ T cells(pTregs) and CD4+CD25+Foxp3+ T cells induced by Tim-3+ NK cells(iTregs) (Fig. 5C). IFN-g production by the CD4
+CD25− T cells was alsoreduced by pTregs and iTregs (Fig. 5D). These results suggest that Tim-3+
NK cells not only induce the differentiation of iTregs but also promotetheir immunosuppressive function in a TGF-b1–dependent manner.
We next compared Tim-3+ NK cells from RM patients andmatched controls for their capacity to induce the differentiationof CD4+CD45RA+ naïve T cells into Tregs. In coculture experimentswith naïve CD4+ T cells, Tim-3+ NK cells from RM patients wereless capable than control cells to induce Foxp3+ T cell generation(Fig. 5, E and F). Using a crossover approach to compare the effectof Tim-3+ NK cells derived from RM patients and from controls onTreg differentiation and function, we found that the percentage ofFoxp3+ and TGF-b1+ cells induced by RM patient Tim-3+ NK cellswas substantially less than that induced by control Tim-3+ NK cells(Fig. 5G). This is consistent with our observation that Tim-3+ NKcells from RM patients produce less TGF-b1 compared with thosefrom controls. From these results, we conclude that Tim-3+ NKcells in maternal peripheral blood can promote iTreg differentiationand enhance iTreg function through TGF-b1 but that this capacityis attenuated in RM patients.
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
Tim-3 blockade disrupts immune tolerance and inducespregnancy failureTo further explore the function of Tim-3 in pregnancy, we used a well-established AP mouse model (38, 39). Consistent with observations inhuman pregnancies, the percentage of Tim-3+ NK cells in the first tri-mester of pregnancywas substantially greater than in the second trimes-ter in mice (fig. S5A). In addition, in the AP mice, the abundances ofTim-3 and Gal-9 on NK cells were decreased (fig. S5B), which was ac-companied by abnormal cytokine production and cytotoxicity (fig. S5,C andD). The conserved function of Tim-3+ NK cells between humansandmice further highlights the importance of theTim-3 signaling in theimmunoregulation of NK cells during pregnancy.
We next treated the AP and NP mice with a Tim-3–blockingantibody, RMT3-23. This antibody substantially decreased littersize and increased the resorption rate in NP and AP mice (Fig.6, A to C). The fetus size for mice treated with RMT3-23 was alsomarkedly decreased (Fig. 6, D and E), suggesting that Tim-3 block-ade had an adverse effect on embryo development. In addition, wefound that the NK cell number in peripheral blood declined mark-edly (fig. S6A) in mice treated with RMT3-23. Flow cytometricanalysis showed that the greatest abundance of Tim-3 was onmature CD11b single-positive (SP) cells but that the lowest abun-dance of Tim-3 was on immature CD27 SP cells (fig. S6, B and C).Moreover, Tim-3 abundance was gradually increased during theprocess of NK cell differentiation from CD34+ cells to matureNK cells (fig. S6D). Together, these data suggest that Tim-3 is closelyassociated with NK cell development and maturation. Furthermore,Tim-3 blockade inhibited the production of anti-inflammatory cyto-kines but promoted pro-inflammatory cytokine production by thesplenic NK cells in both NP and AP mice (Fig. 6F). Similar resultswere obtained in dNK cells (fig. S7A). However, we observed that ac-tivating Tim-3 by administering recombinant mouse Gal-9 (rmGal-9)reduced embryo resorption in AP mice but not in normal mice (Fig. 6,A and D). The RMT3-23–induced fetal loss in both NP and AP micewas not reversed by rmGal-9 administration (Fig. 6, A and C).Consistent results were obtained for cytokine production and the cy-totoxicity of NK cells in the spleen (Fig. 6F) and decidua (fig. S7A).These observations suggest a possible role for Gal-9 and Tim-3signaling in the regulation of the immune tolerant propensity ofNK cells in pregnancy.
To assess the signaling pathways downstream of Gal-9 and Tim-3,the relative abundances of pJNK and pAKT were measured byperforming intracellular staining. The inhibitory effect of RMT3-23on JNK and AKT phosphorylation in NK cells was observed, andthe administration of rmGal-9 failed to reverse that inhibitory effect.However, we detected substantial JNK and AKT phosphorylation inNK cells from mice treated with rmGal-9 (Fig. 6G). Furthermore, NKcells from RMT3-23–challenged mice had decreased amounts of thetranscriptional regulators Id2, Blimp-1, and Gata-3 but an increasedamount of Eomes compared to NK cells from control mice. Conversely,NK cells from mice treated with rmGal-9 exhibited increased amountsof Id2, Blimp-1, and Gata-3 but a decreased amount of Eomes (Fig.6H). Consistent results were observed in the decidua (fig. S7, B andC). Thus, the regulatory effect of Tim-3 and Gal-9 on NK cells duringpregnancy might depend on the JNK and AKT signaling pathwaysand might be mediated through the modulation of the transcriptionfactors Id2, Blimp-1, Gata-3, and Eomes.
The in vitro experiments revealed that Tim-3+ NK cells can induceTreg generation. Here, our in vivo studies showed that interfering with
7 of 15
SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
the Tim-3-Gal-9 interaction notably affected the phenotype andfunction of NK cells from pregnant mice. Next, we wondered whetherthe Tregs would show some consistent changes. Our results showed thatthe proportions of CD25+Foxp3+ Tregs in blood and spleen were re-duced in mice treated with RMT3-23 compared to those in untreatedmice (Fig. 6, I and J). Consistent with this observation, these Tregs dis-played an impaired immunosuppressive capacity and reduced TGF-
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
b1 production (Fig. 6, I and J). Furthermore, the administration ofrmGal-9 led to an increase in the proportion ofCD25+Foxp3+Tregs withenhanced TGF-b1 production in the blood and spleen (Fig. 6, I and J).Consistent results were observed in the decidua (fig. S7D). Thesedata suggest that the Tim-3–Gal-9 signaling pathway facilitates theimmunosuppressive activity of NK cells and the induction of Tregs dur-ing pregnancy.
CD4
Foxp
3
Con Tim-3+ NK Tim-3+ NK + α-TGF-β1 Tim-3– NK
RM
: iTreg : pTreg : CD4+CD25– T cells : pTreg+CD4+CD25– T cells : iTreg+CD4+CD25– T cells
CD
25
EdU
Con Tim-3+ NK Tim-3+ NK + α-TGF-β1 Tim-3– NK
CD4
Foxp
3
NP
Foxp3
TGF-
β1
Con Tim-3+ NK Tim-3+ NK + α-TGF-β1 Tim-3– NK
NP
Foxp3
TGF-
β1 Con Tim-3– NK Tim-3+ NK Tim-3+ NK + α-TGF-β1
RM
2.83 4.32 31.4 9.34
3.94 5.41 17.5 10.2
7.61
16.3 4.62
0.64
2.35
3.49
10.1
1.27
A
B
C
F
G
E
D
Fig. 5. Tim-3+ NK cells from RM patientsare less potent than those from controlsin inducing iTreg differentiation throughTGF-b1. (A and B) Naïve CD4+ T cellsisolated from the peripheral blood of NPdo-nors were stimulated with anti-CD3 andanti-CD28 antibodies for 5 days beforebeingcocultured in the presence or absence ofanti–TGF-b1 antibodywith Tim-3+ or Tim-3−
NK cells obtained from NP donors. The per-centages of Foxp3+CD4+ T cells (top) andTGF-b1+Foxp3+CD4+ T cells (bottom) weredetermined by flow cytometry. Data in thebar graphs aremeans ± SEMof nine donors.(C andD) Tim-3+NKcell–inducedCD4+CD25+
T cells (iTregs), pTregs, and CD4+CD25− cells in
the indicated combinations were restimu-lated with anti-CD3 and anti-CD28 antibo-dies. (C) Their immunosuppressive functionwas assessed by measurement of 5-ethynyl-2′-deoxyuridine (EdU) incorporation. (D) Thepercentages of the indicated cell populationsthat were positive for IFN-gwere determinedby flow cytometry. Data aremeans ± SEMofnine donors. (E and F) Naïve CD4+ T cellsisolated from the peripheral blood of NPdo-nors were stimulated with anti-CD3 andanti-CD28 antibodies for 5 days beforebeingcocultured in the presence or absence ofanti–TGF-b1 antibodywith Tim-3+ or Tim-3−
NKcells obtained fromRMpatients. The per-centages of Foxp3+CD4+ T cells (top) andTGF-b1+Foxp3+CD4+ T cells (bottom) weredetermined by flow cytometry. Data in thebar graphs are means ± SEM of 12 donorsper group. (G) Tim-3+ and Tim-3- NK cellsfrom NP donors and RM patients were ana-lyzed by flow cytometry to determine thepercentages of Foxp3+CD4+ T cells (left)and TGF-b1+Foxp3+CD4+ T cells (right). Dataare means ± SEM of three independentexperiments. *P < 0.05, **P < 0.01, and***P < 0.001 by ANOVA.
8 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
Tim-3+ NK cells facilitate pregnancy maintenanceTo confirm the function of Tim-3+ NK cells in NP, we purified Tim-3+,Tim-3−, or total NK cells frommice withNP and adoptively transferredthese cells into APmice (Fig. 7A).We observed that the resorption ratesof mice that received Tim-3+ NK cells and total NK cells from the micewith NP were substantially reduced. The adoptive transfer of Tim-3−
NK cells from mice with NP had no obvious effects on the resorptionrate. In contrast, Tim-3+ NK cells and total NK cells isolated from the
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
AP mice failed to reverse the pregnancy outcome (Fig. 7, B and C). Inaddition, the effect of Tim-3+ NK cells from APmice was rescued aftertreatment with IL-4 or progesterone, which was concomitant with theincreasedTim-3 abundance on the treated cells (Fig. 7, B toD). Further-more, we found a marked increase in the proportion of CD25+Foxp3+
Tregs in the blood of AP mice that received Tim-3+ NK cells and totalNK cells from mice with NP (Fig. 7E). The transfer of Tim-3+ NK cellsand total NK cells from the APmice had no effect on the proportion of
Isotype PBS RMT3-23 Gal-9 RMT3-23+Gal-9
pJNK
Cel
ls
pAKT
Fluorescence intensity
CD25
Foxp
3 PBS RMT3-23 Gal-9 RMT3-23+Gal-9
Sple
en
Bloo
d
A
E
F
B
C
G
H
I J
( ) ( ) ( ) ( )
( ) ( ) ( ) ( )
( ) ( ) ( ) ( )
( ) ( ) ( ) ( )( ): NP+RMT3-23+Gal-9 ( ): NP+Gal-9 ( ): NP+RMT3-23
( ): AP+PBS ( ): AP+RMT3-23 ( ): AP+Gal-9 ( ): AP+RMT3-23+Gal-9
D ( ): NP+PBS
13.9 2.2 6.8 1.84
22.1 6.1 11.6 5.21
Fig. 6. Tim-3 blockade breaks maternal-fetal tolerance and impairs pregnancymaintenance. (A) Representative picturesshowing the number of embryos per uterusfromNPand APmousemodels treatedwithRMT3-23 and Gal-9 alone or in combination,as indicated by conditions (i) to (viii). (B) Anal-ysis of the percentage of fetal resorption inthe NP and AP mouse models treated as in-dicated in (A). (C) Summary of litter sizes atgestational day (GD) 14.5 in the NP and APmousemodels thatwere treated as indicatedin (A). (D) Representative pictures showingembryos from theNPandAPmousemodelsthat were treated as described in (A). PBS,phosphate-buffered saline. (E) Analysis offetal size (volume) in the NP andAPmousemodels at GD 14.5 after treatment as indi-cated in (A). (F) Spleens from the NP and APmouse models treated with the indicatedcombinations of RMT2-23 and Gal-9 wereanalyzed by flow cytometry to determinethe percentages of NK cells that were posi-tive for the indicated cytokines. Data aremeans ± SEMof five to eightmice per group.(G) Left: Splenic NK cells from the AP micetreated as described in (A) were analyzedby flow cytometry to determine the relativeabundances of pJNK andpAKT. Right: Calcu-lation of the MFIs for pJNK and pAKT. Dataare means ± SEM of five to eight mice pergroup. (H) Splenic NK cells from the APmicetreated as described in (A) were analyzed todetermine the percentage of cells positivefor the indicated transcriptional regulators.Data are means ± SEM of five to eight micepergroup. (Iand J) Left: Peripheralblood (top)and splenic (bottom) CD4+ T cells from APmice thatwere treatedasdescribed in (A)wereanalyzed by flow cytometry to determinethe percentage of Foxp3+ cells. Representa-tive dot plots are shown. Right: Calculationof the percentages of Foxp3+CD4+ T cells inthe blood and spleen under the indicatedconditions. Data are means ± SEM of fiveto eight mice per group. *P < 0.05, **P <0.01, and ***P < 0.001 by ANOVA.
9 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
CD25+Foxp3+ Tregs in the blood of the recipient APmice. Nevertheless,Tim-3+ NK cells and total NK cells isolated from the AP mice treatedwith IL-4 or progesterone had a similar function as those cells isolatedfrommicewithNP that increased the proportion of CD25+Foxp3+ Tregs
in the blood of the recipient mice (Fig. 7E). Furthermore, the transfer ofTim-3+ NK cells and total NK cells isolated from mice with NP led tothe increased production of TGF-b1 by CD25+Foxp3+ Tregs in the
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
blood. A similar effect was observed after AP mice received Tim-3+
NK cells and total NK cells isolated from the AP mice treated withIL-4 or progesterone (fig. S8A). Consistent results were obtained inthe spleen and decidua (fig. S8, B to E).
The potent capacity of Tim-3+ NK cells to induce the production ofiTregs and improve the pregnancy outcome was further confirmed byusing the Nfil3−/− pregnant mouse model. Because they are deficient
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
FinfosepTceinTtrAoflTtoN(NremininAtrbthfethoacodethNnthoaTtrdpm0
CD25Foxp
3
Ctrl Tim-3+ NK Tim-3– NK
Sple
en
Bloo
d
CD25
TGF-
β1
Sple
en
Bloo
d
Ctrl
Tim-3+ NK of NP NK of NP
Tim-3+ NK of AP treated with IL-4
Tim-3+ NK of AP treated with P Tim-3+ NK of AP
Nfil3–/– + Ctrl
Nfil3–/– + Tim-3+ NK
Nfil3–/– + Tim-3– NK
A
Normal pregnancy mouse Abortion-prone mouse Spleen
NK + IL-4
NK + Progesterone
FACS FACS
Abortion-prone mouse Spleen
FACS
Spleen Nfil3+/+ mouse Nfil3
–/–
Tim-3+ NK
NK Tim-3– NK
Tim-3+ NK
NK Tim-3– NK
FACS
FACS Tim-3+ NK
NK Tim-3– NK
Tim-3+ NKTim-3– NK
Tim-3+ NK
NK Tim-3– NK
Abortion-prone mouse
C
F
B
D
H
E
G 11.9 22 9.02
13.6 26.2 13.8
4.48
12.3 24 9.03
20.2 9.8
mouse
ig. 7. Tim-3+ NK cells alleviate fetal lossAP and NK-deficient mice. (A) Schematicr the adoptive transfer of different NK cell sub-ts from the indicated pregnant mice to APregnant mice or Nfil3−/− pregnant mice. Top:im-3+ NK cells, Tim-3− NK cells, and total NKlls were sorted from NP mice and transferredto the tail vein of AP pregnant mice. Middle:otal NK cells sorted from the AP models wereansferred directly into the tail vein of pregnantP mice or were treated with IL-4 or progester-ne for 48 hours in vitro before being sorted byuorescence-activated cell sorting (FACS) intoim-3+ and Tim-3−NK cells and then transferredAP pregnantmice. Bottom: Tim-3+ and Tim-3−
K cells were sorted from normal C57BL/6fil3+/+) pregnant mice and transferred di-ctly into the tail vein of Nfil3−/− pregnantice. (B) Representative pictures of embryosthe uterus of AP mice that had received thedicated NK cells. (C) NK cells isolated fromP mice were left untreated (Ctrl) or wereeated with IL-4 or progesterone (P) beforeeing analyzed by flow cytometry to determinee percentage of Tim-3+ cells. (D) Analysis oftal resorption in AP mice that had receivede indicated NK cells. Data are means ± SEMf 9 to 12 mice per group. (E) Flow cytometricnalysis of the percentage of CD25+Foxp3+
ells within the peripheral blood CD4+ T cellsf AP pregnant mice that had received the in-icated NK cells. (F) Representative pictures ofmbryos in the uterus of controlNfil3−/−miceorose that received either Tim-3+ or Tim-3-
K cells. (G) Analysis of fetal resorption in preg-ant controlNfil3−/−mice or those that receivede indicated NK cells. Data are means ± SEMf five mice per group. (H) Left: Flow cytometricnalysis of the percentages of CD25+Foxp3+
cells and TGF-b1+CD25+Foxp3+ T cells in con-ol Nfil3−/− mice or those that received the in-icated NK cells. Right: Measurement of theercentages of the indicated cells. Data areeans ± SEM of five mice per group. *P <.05, **P < 0.01, and ***P < 0.001 by ANOVA.
10 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
in NK cells (40), Nfil3−/− pregnant mice display an increased rate ofembryo resorption than do the control mice. We sorted Tim-3+ andTim-3− NK cells from the spleens of Nfil3+/+ mice (which have NP)and injected them into Nfil3−/− pregnant mice. The adoptive transferof Tim-3+ NK cells, but not Tim-3− NK cells, substantially reducedthe embryo resorption rate of the recipient Nfil3−/− mice (Fig. 7, Fand G). Furthermore, we found that the proportion of CD25+Foxp3+
Tregs was increased in the blood and spleen ofNfil3−/−mice that received
Tim-3+ NK cells. The production of TGF-b1 by the CD25+Foxp3+ Tregs
was also substantially greater than that in themice that receivedTim-3−NKcells. The transfer of Tim-3− NK cells had no effect on the proportion ofCD25+Foxp3+ Tregs or their production of TGF-b1 (Fig. 7H). Consistentresults were observed in the decidua (fig. S8, F and G). These data sug-gest that Tim-3+ NK cells with the capacity to induce Treg differenti-ation contribute to immune tolerance and pregnancy maintenance.
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
DISCUSSIONNK cells are crucial in the establishment and maintenance of preg-nancy because of their functional diversity, including their roles inplacental vascular remodeling and immunoregulation. Emergingevidence indicates an association between the abnormality of NKcells and the occurrence of RM. Ameta-analysis showed that womenwith RM have statistically significantly more pNK cells than thosewith a healthy pregnancy (7, 20). For patients with a history of RM,the increased activity of pNK cells might be predictive of subsequentmiscarriage. In addition, cytokine production by pNK cells is dysre-gulated in women with a history of reproductive failure (41). Thesestudies suggest that women with RM have altered pNK parameters,which can therefore be recommended as biological markers to iden-tify RM and assess the effect of immunotherapy. However, the spe-cific role of pNK cells in pregnancy is poorly defined, and little isknown about the underlying molecular mechanism. Here, we de-scribed Tim-3+ NK cells with tolerance phenotypes that are charac-terized by an increased production of anti-inflammatory cytokines(including TGF-b1, IL-10, and IL-4), a reduced production of pro-inflammatory cytokines (including TNF-a), and reduced cyto-toxicity toward trophoblast cells. Furthermore, this subtype of NK cellsefficiently induced Treg differentiation. These results suggest that Tim-3+
pNK cells can directly contribute to the anti-inflammatory micro-environment and, more selectively, facilitate iTreg proliferation, whichis conducive to maternal-fetal immune tolerance.
The systemic generation of Tregs during early gestation is pivotalfor successful pregnancy in both human andmouse models. The im-portance of Tregs for NP has been demonstrated in studies showing thatthe absence of Tregs can impair murine pregnancy, whereas the adop-tive transfer of Tregs can prevent fetal rejection (42, 43). However, themechanism underlying Treg generation during early pregnancy is un-clear. Here, we found that the accessibility of TGFB1 in Tim-3+ pNKcells frompatients withRMwas reduced, resulting in amarked decreasein the production of TGF-b1 and a subsequent deficiency in Treg differ-entiation. Thus, our results suggest that previous reports of diminishedTreg responses in patients with RM may be attributable to abnormalTim-3+ NK cells. These findings provide new insights into the mecha-nism of RM and the versatility of NK cells (44).
We found that Tim-3 abundance on pNK cells was decreased inpatients with RM, indicating that Tim-3+ NK might be a predictor ofaberrant pregnancy. However, the regulation of Tim-3 abundance onNK cells is unclear. Here, we found that the amount of cell surface
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
Tim-3 on pNK cells was increased during the first trimester in NPbecause of the typical TH2 polarization signaling by the IL-4–STAT6pathway and physiological concentrations of progesterone. However,increased concentrations of progesterone showed an inhibitory effecton Tim-3 abundance, which may explain the decreased abundance ofTim-3 on NK cells in the second trimester of pregnancy, when theprogesterone concentration is several times greater than that in earlypregnancy. Estrogen and b-hCG showed no effect on Tim-3 abun-dance on pNK cells, which is consistent with the wide use of proges-terone, but not estrogen or b-hCG, in the clinical treatment of earlythreatened abortion. Moreover, progesterone promotes TH2 responsesby inhibiting TH1 cytokine production (45, 46). Thus, we suggest thatprogesterone induces Tim-3 abundance on pNK cells through a directeffect as well as an indirect effect by regulating the TH2-TH1 balance.This result provides a potential theoretical basis for the clinicalapplication of progesterone in the treatment of RM.
Accompanying the deficient Tim-3 expression on pNK cells in RM,we found a substantial decrease in the concentration of the naturalligand Gal-9 in the blood circulation. Furthermore, rmGal-9 admin-istration reduced embryo resorption in AP mice, and the disorderedcytokine production of NK cells was efficiently reversed, consistentwith the results of our in vitro experiment. We therefore propose thatthe Tim-3–mediated immunomodulation by pNK cells during preg-nancy is enhanced by Gal-9. In addition, Gal-9–Tim-3 activated JNKand AKT, leading to increased ID2, BLIMP1, and GATA3 mRNAabundance and decreased EOMES mRNA abundance. These tran-scription factors are associated with NK cell development and func-tions (47–49). Previous reports demonstrated that Tim-3 ligationenhances the activation of theMAPK (mitogen-activated protein kinase)and NF-kB pathways (50). Tim-3 enhances phosphoinositide 3-kinase(PI3K)–AKT activation and A20 activity, inhibiting Toll-like receptor 4(TLR4)–stimulated NF-kB activation (51). Nevertheless, we showedthat JNK and AKT were downstream mediators of Gal-9–Tim-3signaling. Differences among these studies might be explained bythe specific intracellular tail of Tim-3 found in different cell types(50, 52). Thereby, the influence of Gal-9 on pregnancy outcome andthe underlying mechanism of its interaction with Tim-3 indicate itspotential application in the diagnosis and treatment of RM.
We also showed that Tim-3 blockade substantially increasedembryo resorption and led to the appearance of dysfunctional pNKcells, further confirming the crucial modulatory effects of Gal-9–Tim-3 onpNKcells in pregnancy. Note that blocking Tim-3 decreasedpNK cell numbers (fig. S6A), which is consistent with Tim-3 activa-tion by Gal-9 inducing the expression of genes encoding transcriptionfactors associated with NK development. To find the mechanismunderlying the decrease in the number of pNK cells after RMT3-23treatment, we analyzed the expression of Tim-3 on CD27−CD11b−
(double-negative), CD27+CD11b− (CD27 SP), CD27+CD11b+ (double-positive), and CD27−CD11b+ (CD11b SP) populations among NK cells(CD3−Nkp46+) in mice. The results showed that the greatest abundanceof Tim-3was onmature CD11b SP cells but that the lowest abundance ofTim-3 was on immature CD27 SP cells (fig. S6, B and C). Consistently,flow cytometric analysis of human samples showed that about 80% ofmature CD56dim NK cells had cell surface Tim-3, whereas only about40% immature CD56bright NK cells were Tim-3+ (Fig. 1C). These dataimply that Tim-3 might be associated with NK cell development andmaturation. Moreover, we analyzed the published microarray data (28)and found that Tim-3 expression was gradually increased during theprocess of NK cell differentiation from CD34+ cells to mature NK cells
11 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on http://stke.sciencem
ag.org/D
ownloaded from
(fig. S6D). In addition, Ndhlovu et al. (28) demonstrated that Tim-3marks human NK cell maturation. We speculate that Tim-3 mightbe a maturation marker of NK cells in both humans and mice.Treatingmice with RMT3-23, the antibody against Tim-3, had a simi-lar effect to that of the administration of antibodies to either NK1.1 orasialo-GM1. These two antibodies are usually used in NK cell deple-tion assays. That is, RMT3-23 administration may inhibit the mat-uration of NK cells. Because mature CD11b SP NK cells are the majorsubset of peripheral blood NK cells (accounting for 70 to 80% of thetotal) in mice and nearly 80% of CD11b SP NK cells are Tim-3+ (fig.S6, B and C), RMT-23 led to amarked reduction in the number of pNKcells. On the basis of these results, we suggest that Gal-9–Tim-3signaling not only regulates pNK cell function but also influencespNK cell development.
In conclusion, we identified a distinct NK cell subpopulation, Tim-3+
NK cells, with immunosuppressive activity in early pregnancy (fig. S9).The reduced abundance of Tim-3 on NK cells was accompanied bydisordered anti- and pro-inflammatory cytokine profiles in patientswith RM. Moreover, the Treg differentiation–inducing activity ofTim-3+ NK cells was attenuated in patients with RM. Together, theseresults suggest a protective role for this NK cell subset in pregnancy.Currently, the most commonly used biomarkers (serum hCG andprogesterone) are regarded as ineffective in predicting pregnancyoutcome (7). Thus, an accurate and reliable diagnostic biomarkeris urgently needed to reflect the maternal-fetal tolerance status andto enable preventative intervention. The Tim-3+ NK cells defined inour studymay serve as biological markers during early pregnancy forpredicting the occurrence of RM. Our findings also underscore theimportant functional diversity that exists within humanNK cell sub-populations and provide insights into the molecular mechanisms bywhichNKcellsmediate immune tolerance. Because the embryo expressespaternal antigens foreign to the mother, it has been regarded as an allo-graft or a pseudotumor. Paradigms from our studymay also be useful forinvestigating transplantation immunology and tumor immunology.
January 27, 2020
MATERIALS AND METHODSHuman sample collectionThis study was approved by the Research Ethics Committee of Obstet-rics and Gynecology Hospital, Fudan University (Shanghai, China). Allsubjects signed informed written consent for the collection and study ofsamples. Peripheral blood samples were obtained from normal non-pregnant women, women with clinically normal pregnancy, and pa-tients with RM that occurred during the first trimester. NPs wereterminated for nonmedical reasons, and RMs were classified as un-explained after the exclusion of maternal anatomic or hormonal ab-normalities or paternal and maternal chromosomal defects.
Flow cytometry and cell sortingCells were washed and incubated with appropriate fluorochrome-conjugated antibodies for staining. Flow cytometric analysis was per-formed on a CyAn ADP analyzer (Beckman Coulter), and data wereanalyzed with FlowJo version 6.1 software (Tree Star). For flow cyto-metric sorting, cells were stained with specific antibodies and isolatedon a BD FACSAria cell sorter (BD Biosciences).
Western blotting analysisWhole-cell protein extracts were prepared by lysing cells in radioim-munoprecipitation assay buffer supplemented with proteinase inhibi-
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
tors and phosphatase inhibitors (Beyotime Institute of Biotechnology).Protein yield was quantified using the bicinchoninic acid protein assay.After denaturation, equal amounts of protein were separated by SDS–polyacrylamide gel electrophoresis before wet-transfer onto polyvinylidenedifluoride membranes (Amersham Biosciences). Nonspecific bindingsites were blocked by incubating themembranes with 5% bovine serumalbumin (BSA) in tris-buffered saline (TBS) with 0.1% Tween 20(TBS-T) for 1 hour, whichwere rinsed and incubatedwith primary anti-body solutions (1:500 for Tim-3; 1:2000 for ID2, BLIMP1, GATA3, andEOMES; 1:1000 for STAT6, pSTAT6, pERK1/2-Thr202/Tyr204, pNF-kBp65, pJNK-Thr183/Tyr185, pp38, pAKT, ERK1/2, NF-kB p65, JNK, p38,and AKT; and 1:3500 for GAPDH) diluted in blocking buffer (5% BSAand 1× TBS-T) overnight at 4°C. All of the antibodies used forWesternblotting analysis were purchased from Cell Signaling Technology.Primary antibodies were removed by washing the membranes fourtimes in TBS-T, and the membranes were then incubated for 1 hourwith horseradish peroxidase–conjugated secondary antibody (1:5000).After three washes with TBS-T, immunopositive bands on the blotswere visualizedwith the enhanced chemiluminescence detection system(Merck Millipore).
Quantitative real-time PCRTotal RNAwas extracted from pNK cells with TRIzol Reagent (Invitro-gen) and then reverse-transcribed into first-strand complementaryDNA (cDNA) (RR036A, TaKaRa Biotechnology) according to themanufacturer’s instructions. The synthesized cDNAwas amplified withspecific primers (Sagon) and SYBR Green (TaKaRa Biotechnology)withABI PRISM7900 SequenceDetection System (Applied Biosystems).Triplicate samples were examined in each condition. A comparativethreshold cycle (CT) value was normalized for each sample by the 2−DDCT
method.
NK cell cytotoxicity assayThe cytotoxicity activity of NK cells was determined with the CytoTox96 Non-Radioactive Cytotoxicity Assay Kit (G1780; Promega) accordingto the manufacturer’s instructions. pNK cells (effector cells; 100 ml) atconcentrations of 1.0 × 106/ml, 5.0 × 105/ml, 2.5 × 105/ml, and 1.3 ×105/ml were mixed with 100 ml of HTR-8/SVneo cells (target cells)at a concentration of 2.0 × 104/ml, resulting in E:T ratios of 50:1, 25:1,12.5, and 6.5:1, respectively. The percentage of cytotoxicity wascalculated after correcting for lactate dehydrogenase (LDH) release fromHTR-8/SVneo cells using the following formula: Percentage cytotoxicity =100 × (Experimental values − Culture medium background)/(MaximumLDH release −Culturemedium background). Experimental values re-present LDH release activity from cocultures of effector and targetcells. LDH culturemedium background represents LDH release fromtrophoblasts. Maximum LDH release represents LDH release fromtrophoblasts that were lysed by sonication.
Murine models of pregnancyMale DBA/2 and female CBA/J mice (8 to 10 weeks old) were pur-chased from Beijing HFK Bioscience Co. Male Balb/c mice (8 to10 weeks old) were purchased from the Department of LaboratoryAnimal Science, Fudan University. All animals were kept under spe-cific pathogen–free conditions. All of the experimental proceduresinvolving animals were conducted in accordance with the Guidefor the Care and Use of Laboratory Animals (China), and permissionwas approved by the Human Research Ethics Committee of Obstet-rics and Gynecology Hospital of Fudan University. CBA/J × Balb/c
12 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
mating combinations (NPmodel) were established as follows: femaleCBA/J mice were mated in natural cycling with male Balb/c mice.CBA/J × DBA/2 mating combinations (AP model) were establishedas follows: female CBA/J mice were mated in natural cycling with maleDBA/2 mice. Detection of a vaginal plug was chosen to indicate day 0.5of gestation. Pregnant females received three injections of RMT3-23(anti–Tim-3 antibody) intraperitoneally at doses of 500, 250, and 250mg at days 3.5, 5.5, and 7.5, respectively. In other instances, pregnantmice were injected with rmGal-9 intraperitoneally at a dose of 100 mgat days 3.5, 5.5, and 7.5, respectively. The percentage of resorbed embryoswas calculated as follows: resorbed embryos/total embryos × 100.
Adoptive transfer of Tim-3+ NK cellsTim-3+NK cells, Tim-3−NKcells, and total NK cells were isolated fromthe spleens of CBA/J mice with an NP (GD 7.5) by FACS. Tim-3+ NKcells, Tim-3− NK cells, or total NK cells (3 × 105) were resuspended in200 ml of PBS and injected into the tail vein of pregnant AP CBA/J mice(GD 4.5). Tim-3+ NK cells, Tim-3−NK cells, and total NK cells isolatedfrom the spleens of AP CBA/J mice (GD 7.5) by FACS were transferredin the same way. NK cells isolated from the spleens of AP CBA/J mice(GD 7.5) were stimulated with IL-4 or progesterone for 48 hours beforebeing sorted by FACS to isolate the Tim-3+ and Tim-3− NK cells. Thesorted cells were resuspended in 200 ml of PBS and injected into the tailvein of pregnant AP CBA/J mice (GD 4.5). Tim-3+ NK cells, Tim-3−
NK cells, and total NK cells were isolated from the spleens of Nfil3+/+
mice with NP (GD 7.5) by FACS. Tim-3+ NK cells, Tim-3−NK cells,and total NK cells (3 × 105) were resuspended in 200 ml of PBS andinjected into the tail vein of pregnant (GD 4.5) Nfil3−/− mice (a giftfrom H. Wei, University of Science and Technology of China). Thesame volume of PBS was injected into the control mice. All of therecipient mice were sacrificed and examined to calculate the embryoresorption rate.
Expression profiling by microarrayTim-3+ pNK cells and Tim-3− pNK cells from six women with NPwere sorted by FACS. Total RNA was directly isolated with TRIzolaccording to the manufacturer’s instructions. RNA was analyzed ona NanoDrop spectrophotometer (Thermo Fisher Scientific) and aBioanalyzer (Agilent) to determine RNA yield and integrity. RNAwas subjected to one round of amplification and biotinylation withthe Amino Allyl MessageAmp II aRNA Amplification Kit (ThermoFisher Scientific). BiotinylatedRNAwas hybridized to human genomeHOA 6.2 chips. Gene expression analysis was performed using theWhole Genome OneArray Microarray (Phalanx Biotech Group).The log2 ratio of each probe was calculated by pairwise combinationand error-weighted average. Probes showing a log2 ratio of ≥1 or≤−1 and a P value of <0.05 were defined as differentially expressedgenes.
Assay for transposase-accessible chromatin sequencingThe ATAC-seq protocol (35, 53) was performed to profile openchromatin. The ATAC-seq libraries were sequenced on a HiSeq2500 next-generation sequencer (Illumina) with paired-end 50-bpreads. The ATAC-seq reads were mapped to Hg19 with Bowtie2, andmitochondrial or duplicate PCRs were removed. ATAC-seq peakswere identified with MACS2, and peaks from all of the samples weremerged with Homer. The number of reads falling into each peak wasthen calculated with Homer, and differentially accessible regions weredetermined with DESeq2 (P < 0.05).
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
Statistical analysisThe statistical software Prism 6 (GraphPad) was used for data analyses.Statistical significance was determined by Student’s t test. Multiplemeans were compared by ANOVA. Error bars in figures indicate theSEM. Statistical significance was set at P < 0.05. Statistically significantresults are expressed using asterisks, where *P < 0.05, **P < 0.01, and***P < 0.001.
SUPPLEMENTARY MATERIALSwww.sciencesignaling.org/cgi/content/full/10/498/eaah4323/DC1Fig. S1. Detection of Tim-3 on different cell types in the peripheral blood of patients in earlypregnancy.Fig. S2. GO analysis of differentially expressed genes.Fig. S3. Estrogen and b-hCG have no effect on Tim-3 abundance on pNK cells.Fig. S4. The amounts of TGF-b1 and perforin in Tim-3+ and Tim-3− NK cells are unchanged bythe BET inhibitor JQ1.Fig. S5. AP mice show decreased Tim-3 and Gal-9 abundance and have dysfunctional NK cells.Fig. S6. The percentage of NK cells in the blood is reduced by RMT3-23.Fig. S7. The function of dNK cells is impaired after Tim-3 blockade.Fig. S8. Tregs are generated in mice that received Tim-3+ NK cells.Fig. S9. Model of how Tim-3 on pNK cells may participate in establishing an immune tolerantphenotype during early pregnancy.
REFERENCES AND NOTES1. A. Erlebacher, Immunology of the maternal-fetal interface. Annu. Rev. Immunol. 31,
387–411 (2013).2. M. PrabhuDas, E. Bonney, K. Caron, S. Dey, A. Erlebacher, A. Fazleabas, S. Fisher,
T. Golos, M. Matzuk, J. M. McCune, G. Mor, L. Schulz, M. Soares, T. Spencer, J. Strominger,S. S. Way, K. Yoshinaga, Immune mechanisms at the maternal-fetal interface: Perspectivesand challenges. Nat. Immunol. 16, 328–334 (2015).
3. P. C. Arck, K. Hecher, Fetomaternal immune cross-talk and its consequences for maternaland offspring’s health. Nat. Med. 19, 548–556 (2013).
4. M. E. Solano, M. K. Kowal, G. E. O’Rourke, A. K. Horst, K. Modest, T. Plosch, R. Barikbin,C. C. Remus, R. G. Berger, C. Jago, H. Ho, G. Sass, V. J. Parker, J. P. Lydon, F. J. DeMayo,K. Hecher, K. Karimi, P. C. Arck, Progesterone and HMOX-1 promote fetal growth by CD8+
T cell modulation. J. Clin. Invest. 125, 1726–1738 (2015).5. O. B. Christiansen, R. Steffensen, H. S. Nielsen, K. Varming, Multifactorial etiology of
recurrent miscarriage and its scientific and clinical implications. Gynecol. Obstet. Invest.66, 257–267 (2008).
6. E. C. Larsen, O. B. Christiansen, A. M. Kolte, N. Macklon, New insights into mechanismsbehind miscarriage. BMC Med. 11, 154 (2013).
7. S. Seshadri, S. K. Sunkara, Natural killer cells in female infertility and recurrentmiscarriage: A systematic review and meta-analysis. Hum. Reprod. Update 20, 429–438(2014).
8. M. A. Ivarsson, L. Loh, N. Marquardt, E. Kekäläinen, L. Berglin, N. K. Björkström,M. Westgren, D. F. Nixon, J. Michaëlsson, Differentiation and functional regulation ofhuman fetal NK cells. J. Clin. Invest. 123, 3889–3901 (2013).
9. Z. Madeja, H. Yadi, R. Apps, S. Boulenouar, S. J. Roper, L. Gardner, A. Moffett,F. Colucci, M. Hemberger, Paternal MHC expression on mouse trophoblast affectsuterine vascularization and fetal growth. Proc. Natl. Acad. Sci. U.S.A. 108, 4012–4017(2011).
10. S. Xiong, A. M. Sharkey, P. R. Kennedy, L. Gardner, L. E. Farrell, O. Chazara, J. Bauer,S. E. Hiby, F. Colucci, A. Moffett, Maternal uterine NK cell-activating receptor KIR2DS1enhances placentation. J. Clin. Invest. 123, 4264–4272 (2013).
11. D. A. Somerset, Y. Zheng, M. D. Kilby, D. M. Sansom, M. T. Drayson, Normal humanpregnancy is associated with an elevation in the immune suppressive CD25+ CD4+
regulatory T-cell subset. Immunology 112, 38–43 (2004).12. R. M. Samstein, S. Z. Josefowicz, A. Arvey, P. M. Treuting, A. Y. Rudensky, Extrathymic
generation of regulatory T cells in placental mammals mitigates maternal-fetal conflict.Cell 150, 29–38 (2012).
13. C. Carlino, H. Stabile, S. Morrone, R. Bulla, A. Soriani, C. Agostinis, F. Bossi, C. Mocci,F. Sarazani, F. Tedesco, A. Santoni, A. Gismondi, Recruitment of circulating NK cells throughdecidual tissues: A possible mechanism controlling NK cell accumulation in the uterusduring early pregnancy. Blood 111, 3108–3115 (2008).
14. J. Kieckbusch, L. M. Gaynor, A. Moffett, F. Colucci, MHC-dependent inhibition of uterineNK cells impedes fetal growth and decidual vascular remodelling. Nat. Commun. 5,3359 (2014).
13 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from
15. P. Vacca, M. C. Mingari, L. Moretta, Natural killer cells in human pregnancy. J. Reprod.Immunol. 97, 14–19 (2013).
16. A. M. Borzychowski, B. A. Croy, W. L. Chan, C. W. Redman, I. L. Sargent, Changes insystemic type 1 and type 2 immunity in normal pregnancy and pre-eclampsia may bemediated by natural killer cells. Eur. J. Immunol. 35, 3054–3063 (2005).
17. W. L. Chan, N. Pejnovic, C. A. Lee, N. A. Al-Ali, Human IL-18 receptor and ST2L are stableand selective markers for the respective type 1 and type 2 circulating lymphocytes.J. Immunol. 167, 1238–1244 (2001).
18. K. Aoki, S. Kajiura, Y. Matsumoto, M. Ogasawara, S. Okada, Y. Yagami, N. Gleicher,Preconceptional natural-killer-cell activity as a predictor of miscarriage. Lancet 345,1340–1342 (1995).
19. N. Karami, M. G. Boroujerdnia, R. Nikbakht, A. Khodadadi, Enhancement of peripheralblood CD56dim cell and NK cell cytotoxicity in women with recurrent spontaneousabortion or in vitro fertilization failure. J. Reprod. Immunol. 95, 87–92 (2012).
20. K. King, S. Smith, M. Chapman, G. Sacks, Detailed analysis of peripheral bloodnatural killer (NK) cells in women with recurrent miscarriage. Hum. Reprod. 25, 52–58(2010).
21. C. Zhu, A. C. Anderson, A. Schubart, H. Xiong, J. Imitola, S. J. Khoury, X. X. Zheng,T. B. Strom, V. K. Kuchroo, The Tim-3 ligand galectin-9 negatively regulates T helper type1 immunity. Nat. Immunol. 6, 1245–1252 (2005).
22. A. Sánchez-Fueyo, J. Tian, D. Picarella, C. Domenig, X. X. Zheng, C. A. Sabatos,N. Manlongat, O. Bender, T. Kamradt, V. K. Kuchroo, J.-C. Gutiérrez-Ramos, A. J. Coyle,T. B. Strom, Tim-3 inhibits T helper type 1–mediated auto- and alloimmune responsesand promotes immunological tolerance. Nat. Immunol. 4, 1093–1101 (2003).
23. T. Flecken, P. Sarobe, Tim-3 expression in tumour-associated macrophages: A new playerin HCC progression. Gut 64, 1502–1503 (2015).
24. S. Koyama, E. A. Akbay, Y. Y. Li, G. S. Herter-Sprie, K. A. Buczkowski, W. G. Richards,L. Gandhi, A. J. Redig, S. J. Rodig, H. Asahina, R. E. Jones, M. M. Kulkarni, M. Kuraguchi,S. Palakurthi, P. E. Fecci, B. E. Johnson, P. A. Janne, J. A. Engelman, S. P. Gangadharan,D. B. Costa, G. J. Freeman, R. Bueno, F. S. Hodi, G. Dranoff, K.-K. Wong, P. S. Hammerman,Adaptive resistance to therapeutic PD-1 blockade is associated with upregulation ofalternative immune checkpoints. Nat. Commun. 7, 10501 (2016).
25. B. L. Phong, L. Avery, T. L. Sumpter, J. V. Gorman, S. C. Watkins, J. D. Colgan, L. P. Kane,Tim-3 enhances FceRI-proximal signaling to modulate mast cell activation. J. Exp. Med.212, 2289–2304 (2015).
26. F. Wang, W. He, J. Yuan, K. Wu, H. Zhou, W. Zhang, Z. K. Chen, Activation of Tim-3–Galectin-9 pathway improves survival of fully allogeneic skin grafts. Transpl. Immunol. 19,12–19 (2008).
27. G. J. Freeman, J. M. Casasnovas, D. T. Umetsu, R. H. DeKruyff, TIM genes: A family of cellsurface phosphatidylserine receptors that regulate innate and adaptive immunity.Immunol. Rev. 235, 172–189 (2010).
28. L. C. Ndhlovu, S. Lopez-Vergès, J. D. Barbour, R. B. Jones, A. R. Jha, B. R. Long,E. C. Schoeffler, T. Fujita, D. F. Nixon, L. L. Lanier, Tim-3 marks human natural killercell maturation and suppresses cell-mediated cytotoxicity. Blood 119, 3734–3743(2012).
29. S. Jost, U. Y. Moreno-Nieves, W. F. Garcia-Beltran, K. Rands, J. Reardon, I. Toth,A. Piechocka-Trocha, M. Altfeld, M. M. Addo, Dysregulated Tim-3 expression on naturalkiller cells is associated with increased Galectin-9 levels in HIV-1 infection. Retrovirology10, 74 (2013).
30. Y. Ju, N. Hou, J. Meng, X. Wang, X. Zhang, D. Zhao, Y. Liu, F. Zhu, L. Zhang, W. Sun,X. Liang, L. Gao, C. Ma, T cell immunoglobulin- and mucin-domain-containing molecule-3(Tim-3) mediates natural killer cell suppression in chronic hepatitis B. J. Hepatol. 52,322–329 (2010).
31. P.-F. Guo, M.-R. Du, H.-X. Wu, Y. Lin, L.-P. Jin, D.-J. Li, Thymic stromal lymphopoietinfrom trophoblasts induces dendritic cell–mediated regulatory TH2 bias in the deciduaduring early gestation in humans. Blood 116, 2061–2069 (2010).
32. L.-P. Jin, Y.-H. Zhou, X.-Y. Zhu, M.-Y. Wang, D.-J. Li, Adoptive transfer of paternalantigen-hyporesponsive T cells facilitates a Th2 bias in peripheral lymphocytes and atmaterno-fetal interface in murine abortion-prone matings. Am. J. Reprod. Immunol.56, 258–266 (2006).
33. Y. Liu, H. Ji, Y. Zhang, X. Shen, F. Gao, X. He, G. A. Li, R. W. Busuttil, V. K. Kuchroo,J. W. Kupiec-Weglinski, Recipient T cell TIM-3 and hepatocyte galectin-9 signallingprotects mouse liver transplants against ischemia-reperfusion injury. J. Hepatol.62, 563–572 (2015).
34. K. Sakuishi, P. Jayaraman, S. M. Behar, A. C. Anderson, V. K. Kuchroo, EmergingTim-3 functions in antimicrobial and tumor immunity. Trends Immunol. 32, 345–349 (2011).
35. J. D. Buenrostro, P. G. Giresi, L. C. Zaba, H. Y. Chang, W. J. Greenleaf, Transpositionof native chromatin for fast and sensitive epigenomic profiling of openchromatin, DNA-binding proteins and nucleosome position. Nat. Methods 10,1213–1218 (2013).
36. R. E. Thurman, E. Rynes, R. Humbert, J. Vierstra, M. T. Maurano, E. Haugen, N. C. Sheffield,A. B. Stergachis, H. Wang, B. Vernot, K. Garg, S. John, R. Sandstrom, D. Bates, L. Boatman,
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
T. K. Canfield, M. Diegel, D. Dunn, A. K. Ebersol, T. Frum, E. Giste, A. K. Johnson,E. M. Johnson, T. Kutyavin, B. Lajoie, B.-K. Lee, K. Lee, D. London, D. Lotakis, S. Neph,F. Neri, E. D. Nguyen, H. Qu, A. P. Reynolds, V. Roach, A. Safi, M. E. Sanchez,A. Sanyal, A. Shafer, J. M. Simon, L. Song, S. Vong, M. Weaver, Y. Yan, Z. Zhang,Z. Zhang, B. Lenhard, M. Tewari, M. O. Dorschner, R. S. Hansen, P. A. Navas,G. Stamatoyannopoulos, V. R. Iyer, J. D. Lieb, S. R. Sunyaev, J. M. Akey, P. J. Sabo,R. Kaul, T. S. Furey, J. Dekker, G. E. Crawford, J. A. Stamatoyannopoulos, The accessiblechromatin landscape of the human genome. Nature 489, 75–82 (2012).
37. P. Filippakopoulos, J. Qi, S. Picaud, Y. Shen, W. B. Smith, O. Fedorov, E. M. Morse, T. Keates,T. T. Hickman, I. Felletar, M. Philpott, S. Munro, M. R. McKeown, Y. Wang, A. L. Christie,N. West, M. J. Cameron, B. Schwartz, T. D. Heightman, N. La Thangue, C. A. French,O. Wiest, A. L. Kung, S. Knapp, J. E. Bradner, Selective inhibition of BET bromodomains.Nature 468, 1067–1073 (2010).
38. E. K. Haddad, A. J. Duclos, M. G. Baines, Early embryo loss is associated with local productionof nitric oxide by decidual mononuclear cells. J. Exp. Med. 182, 1143–1151 (1995).
39. A. Zhao, M. Xiong, Y. Zhang, S. Bao, J. Zhang, L. Qiu, Q. Lin, Adoptive transfer ofmFas ligand into dendritic cells influences the spontaneous resorption rate in theCBA/J × DBA/2 mouse model. Fertil. Steril. 93, 1700–1705 (2010).
40. B. Fu, X. Li, R. Sun, X. Tong, B. Ling, Z. Tian, H. Wei, Natural killer cells promote immunetolerance by regulating inflammatory TH17 cells at the human maternal–fetal interface.Proc. Natl. Acad. Sci. U.S.A. 110, E231–E240 (2013).
41. A. Fukui, J. Kwak-Kim, E. Ntrivalas, A. Gilman-Sachs, S.-K. Lee, K. Beaman, Intracellularcytokine expression of peripheral blood natural killer cell subsets in women withrecurrent spontaneous abortions and implantation failures. Fertil. Steril. 89, 157–165(2008).
42. V. R. Aluvihare, M. Kallikourdis, A. G. Betz, Regulatory T cells mediate maternal toleranceto the fetus. Nat. Immunol. 5, 266–271 (2004).
43. A. Leber, A. Teles, A. C. Zenclussen, Regulatory T cells and their role in pregnancy.Am. J. Reprod. Immunol. 63, 445–459 (2010).
44. E. Vivier, E. Tomasello, M. Baratin, T. Walzer, S. Ugolini, Functions of natural killer cells.Nat. Immunol. 9, 503–510 (2008).
45. H. Miyaura, M. Iwata, Direct and indirect inhibition of Th1 development by progesteroneand glucocorticoids. J. Immunol. 168, 1087–1094 (2002).
46. T. Karasu, T. H. Marczylo, M. Maccarrone, J. C. Konje, The role of sex steroid hormones,cytokines and the endocannabinoid system in female fertility. Hum. Reprod. Update17, 347–361 (2011).
47. R. Schotte, W. Dontje, M. Nagasawa, Y. Yasuda, A. Q. Bakker, H. Spits, B. Blom, Synergybetween IL-15 and Id2 promotes the expansion of human NK progenitor cells, whichcan be counteracted by the E protein HEB required to drive T cell development.J. Immunol. 184, 6670–6679 (2010).
48. J. J. Knox, G. L. Cosma, M. R. Betts, L. M. McLane, Characterization of T-bet and eomes inperipheral human immune cells. Front. Immunol. 5, 217 (2014).
49. R. B. Delconte, W. Shi, P. Sathe, T. Ushiki, C. Seillet, M. Minnich, T. B. Kolesnik, L. C. Rankin,L. A. Mielke, J.-G. Zhang, M. Busslinger, M. J. Smyth, D. S. Hutchinson, S. L. Nutt,S. E. Nicholson, W. S. Alexander, L. M. Corcoran, E. Vivier, G. T. Belz, S. Carotta,N. D. Huntington, The helix-loop-helix protein ID2 governs NK cell fate by tuning theirsensitivity to Interleukin-15. Immunity 44, 103–115 (2016).
50. A. C. Anderson, D. E. Anderson, L. Bregoli, W. D. Hastings, N. Kassam, C. Lei,R. Chandwaskar, J. Karman, E. W. Su, M. Hirashima, J. N. Bruce, L. P. Kane, V. K. Kuchroo,D. A. Hafler, Promotion of tissue inflammation by the immune receptor Tim-3 expressedon innate immune cells. Science 318, 1141–1143 (2007).
51. X. Yang, X. Jiang, G. Chen, Y. Xiao, S. Geng, C. Kang, T. Zhou, Y. Li, X. Guo, H. Xiao, C. Hou,R. Wang, Z. Lin, X. Li, J. Feng, Y. Ma, B. Shen, Y. Li, G. Han, T cell Ig mucin-3 promoteshomeostasis of sepsis by negatively regulating the TLR response. J. Immunol. 190,2068–2079 (2013).
52. M. K. Gleason, T. R. Lenvik, V. McCullar, M. Felices, M. S. O’Brien, S. A. Cooley,M. R. Verneris, F. Cichocki, C. J. Holman, A. Panoskaltsis-Mortari, T. Niki, M. Hirashima,B. R. Blazar, J. S. Miller, Tim-3 is an inducible human natural killer cell receptor thatenhances interferon gamma production in response to galectin-9. Blood 119,3064–3072 (2012).
53. D. Lara-Astiaso, A. Weiner, E. Lorenzo-Vivas, I. Zaretsky, D. A. Jaitin, E. David,H. Keren-Shaul, A. Mildner, D. Winter, S. Jung, N. Friedman, I. Amit,Immunogenetics. Chromatin state dynamics during blood formation. Science 345,943–949 (2014).
Acknowledgments: We thank D. Li (The Shanghai Information Center for Life Sciences ofthe Chinese Academy of Sciences) for his critical revision of the manuscript and H. Weifor his gift of the Nfil3−/− mice, which were originally from TakWahMak (University ofToronto, Toronto, Canada). Funding: This work was supported by the National Basic ResearchProgram of China (2015CB943300, 2014CB943600, and 2017YFC1001403), the National NaturalScience Foundation of China (81630036, 91542116, 31570920, 81490744, 31171437,
14 of 15

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
31270969, 31370858, 81671552, and 81571512), the Program of Shanghai Academic/Technology Research Leader (17XD1400900), the Key Project of Shanghai Municipal EducationCommission (14ZZ013), the Key Project of Shanghai Basic Research from Shanghai Municipal Scienceand Technology Commission (12JC1401600), and Shanghai Sailing Program (no. 17YF1411600).Author contributions: Y.L. designed and performed most experiments; J.Z. designed andperformed ATAC-seq procedures and analyses; M.D. and X.C. conceived the study and designedexperiments; D.Z., Y.X., S.W., Y.T., X.H., andH.P. coordinated the sample collection, literature search,data analysis, and figure preparation; and Y.L. drafted the manuscript. All authors analyzedand interpreted data and critically revised the manuscript. Competing interests: The authorsdeclare that they have no competing interests.
Li et al., Sci. Signal. 10, eaah4323 (2017) 26 September 2017
Submitted 27 June 2016Resubmitted 22 March 2017Accepted 12 September 2017Published 26 September 201710.1126/scisignal.aah4323
Citation: Y. Li, J. Zhang, D. Zhang, X. Hong, Y. Tao, S. Wang, Y. Xu, H. Piao, W. Yin, M. Yu,Y. Zhang, Q. Fu, D. Li, X. Chang, M. Du, Tim-3 signaling in peripheral NK cells promotesmaternal-fetal immune tolerance and alleviates pregnancy loss. Sci. Signal. 10, eaah4323(2017).
15 of 15
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from

alleviates pregnancy lossTim-3 signaling in peripheral NK cells promotes maternal-fetal immune tolerance and
Yin Zhang, Qiang Fu, Dajin Li, Xing Chang and Meirong DuYanhong Li, Jiayuan Zhang, Di Zhang, Xiaowu Hong, Yu Tao, Songcun Wang, Yuanyuan Xu, Hailan Piao, Weijie Yin, Min Yu,
DOI: 10.1126/scisignal.aah4323 (498), eaah4323.10Sci. Signal.
pNK cells promote maternal-fetal immune tolerance.+protected against fetal loss, suggesting that Tim-3 pNK cells,− pNK cells, but not Tim-3+in immunosuppression. Experiments with abortion-prone mice showed that Tim-3
with normal pregnancies, those from recurrent miscarriage patients had decreased Tim-3 abundance and were defective anti-inflammatory cytokines and induced the generation of immunosuppressive cells. In contrast to pNK cells from donorsthe surface of peripheral natural killer (pNK) cells in the first trimester of pregnancy. These cells produced
. found that the abundance of the type I membrane receptor Tim-3 was increased onet alsuggest possible therapies. Li fetus. Determining the underlying mechanism may aid in the identification of biomarkers for recurrent miscarriage and
Many cases of recurrent miscarriage are due to a breakdown in immune tolerance between the mother and theMediating maternal-fetal tolerance
ARTICLE TOOLS http://stke.sciencemag.org/content/10/498/eaah4323
MATERIALSSUPPLEMENTARY http://stke.sciencemag.org/content/suppl/2017/09/22/10.498.eaah4323.DC1
CONTENTRELATED
http://immunology.sciencemag.org/content/immunology/2/15/eaao3543.fullhttp://immunology.sciencemag.org/content/immunology/2/9/eaah4569.fullfile:/contenthttp://stm.sciencemag.org/content/scitransmed/8/350/350ra102.fullhttp://stke.sciencemag.org/content/sigtrans/10/504/eaar3893.fullhttp://stke.sciencemag.org/content/sigtrans/10/500/eaaq1408.fullhttp://stke.sciencemag.org/content/sigtrans/10/500/eaal4055.fullhttp://stke.sciencemag.org/content/sigtrans/10/499/eaaq0229.fullhttp://stke.sciencemag.org/content/sigtrans/10/499/eaaq0904.fullhttp://science.sciencemag.org/content/sci/350/6258/334.fullhttp://science.sciencemag.org/content/sci/348/6230/56.fullhttp://stke.sciencemag.org/content/sigtrans/8/399/ra104.full
REFERENCES
http://stke.sciencemag.org/content/10/498/eaah4323#BIBLThis article cites 53 articles, 15 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.Science SignalingYork Avenue NW, Washington, DC 20005. The title (ISSN 1937-9145) is published by the American Association for the Advancement of Science, 1200 NewScience Signaling
Science. No claim to original U.S. Government WorksCopyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of
on January 27, 2020http://stke.sciencem
ag.org/D
ownloaded from