thèse perpignan
TRANSCRIPT

Université de Perpignan Via Domitia Année 2011
Laboratoire de Chimie des Biomolécules et de l’Environnement (LCBE) EA 4215 Laboratoire Génome et Développement des Plantes (LGDP) UMR 5096
Ecole Doctorale Energie et Environement (ED305)
N° d’ordre
THÈSE
Présentée devant l’Université de Perpignan Via Domitia pour l’obtention du grade de Docteur de l’Université
Discipline : Biologie
Par
Christophe CALVAYRAC
DÉGRADATION BIOLOGIQUE DE LA SULCOTRIONE
DANS UN SOL AGRICOLE : RECHERCHE D’UNE ÉVENTUELLE BIODÉGRADATION ACCÉLÉRÉE ET CARACTÉRISATION DE SOUCHES BACTÉRIENNES
POTENTIELLEMENT DÉGRADANTES
Soutenue publiquement le 20 septembre 2011
Composition du jury :
F. MARTIN-LAURENT Directeur de Recherches INRA, Dijon Rapporteur H. FENET Maître de Conférences, Université Montpellier 1 Rapporteur A. AMBLES Professeur, Université de Poitiers Examinateur R. ROUILLON Professeur, UPVD Examinateur C. GUYOT Expert Environnement, BayerCropScience Examinateur C. M. COSTE Professeur Émérite, UPVD Examinateur J. F. COOPER Professeur, UPVD Directeur de thèse O. PANAUD Professeur, UPVD Directeur de thèse

Université de Perpignan Via Domitia Année 2011
Laboratoire de Chimie des Biomolécules et de l’Environnement (LCBE) EA 4215 Laboratoire Génome et Développement des Plantes (LGDP) UMR 5096
Ecole Doctorale Energie et Environement (ED305)
N° d’ordre
THÈSE
Présentée devant l’Université de Perpignan Via Domitia pour l’obtention du grade de Docteur de l’Université
Discipline : Biologie
Par
Christophe CALVAYRAC
DÉGRADATION BIOLOGIQUE DE LA SULCOTRIONE DANS UN SOL AGRICOLE : RECHERCHE D’UNE
ÉVENTUELLE BIODÉGRADATION ACCÉLÉRÉE ET CARACTÉRISATION DE SOUCHES BACTÉRIENNES
POTENTIELLEMENT DÉGRADANTES
Soutenue publiquement le 20 septembre 2011
Composition du jury :
F. MARTIN-LAURENT Directeur de Recherches INRA, Dijon Rapporteur H. FENET Maître de Conférences, Université Montpellier 1 Rapporteur A. AMBLES Professeur, Université de Poitiers Examinateur R. ROUILLON Professeur, UPVD Examinateur C. GUYOT Expert Environnement, BayerCropScience Examinateur C. M. COSTE Professeur Émérite, UPVD Examinateur J. F. COOPER Professeur, UPVD Directeur de thèse O. PANAUD Professeur, UPVD Directeur de thèse


2
Ab nou cor et ab nou talen, Ab nou saber et ab nou sen, Et ab nou bel captenemen, Vuoill un bon vers commensar.
Raimbaut d’Aurenga, trobador.

3
Remerciements
Les travaux présentés dans ce manuscrit ont été effectués au sein du Laboratoire de Chimie
des Biomolécules et de l’Environnement (LCBE) et du Laboratoire Génome et
Développement des Plantes (LGDP) de l’Université de Perpignan Via Domitia (UPVD).
Au terme de ces nombreuses années passées sous la direction des mes deux directeurs de
thèse, qu’ils veuillent bien trouver dans ce travail, un modeste témoignage de mon profond
respect et de mon entière reconnaissance pour leur aide et leurs précieux conseils.
Je voudrais remercier Mr Jean-François Cooper de m’avoir accepté au sein de son équipe et
de m’avoir confié ce présent travail. Malgré les grandes difficultés et les nombreux obstacles
qui se sont présentés, il m’a toujours prodigué encouragements et conseils et a su porter un
intérêt constant à mon travail. J’ai beaucoup appris à ses cotés. Et bien au-delà de l’aspect
purement scientifique. Son érudition, son rayonnement personnel ainsi que son humanité sont
pour beaucoup dans la réussite de ce travail. Qu’il soit permis de lui exprimer mon
respectueux attachement ainsi que ma profonde admiration.
Je remercie également Mr Olivier Panaud qui a accepté de co-dirriger ce travail. Je lui suis
très reconnaissant pour la qualité de son accueil au sein de son équipe, son soutien permanent
ainsi que pour la confiance qu’il su m’accorder. Il a toujours manifesté à mon égard
gentillesse, marques de sympathie et une très grande disponibilité. La qualité de son
encadrement scientifique m’a permis de mener à bien une grande partie de ce travail. Qu’il
veuille bien trouver ici l’expression de mes très sincères remerciements.
Je tiens à exprimer toute ma gratitude à Mr Camille Michel Coste dont le précieux concours
m’a permis de réaliser une partie de mon travail. Ses conseils avisés, sa grande disponibilité,
sa bonne humeur ainsi que les nombreuses discussions scientifiques que nous avons pu
échanger ont été très constructives et m’ont été très profitables. Qu’il veuille bien trouver ici
la marque de ma sympathie et l’expression de mes très sincères remerciements.

4
Je remercie vivement les membres du jury qui ont accepté d’évaluer ce travail. Leur présence
signe l’intérêt qu’ils ont porté à la réalisation de cette thèse. Merci tout particulièrement à Mr
Fabrice Martin-Laurent et à Mme Hélène Fenet d’avoir accepté d’être les rapporteurs du
manuscrit, qu’ils soient assurés de ma profonde reconnaissance et de mes sentiments
respectueux.
J’adresse également un vif remerciement à Mr Régis Rouillon qui a accepté de présider ce
jury de thèse. Qu’il soit permis de lui exprimer toute ma profonde reconnaissance.
Je remercie BayerCropScience pour le soutien financier accordé tout au long de ce travail de
thèse. J’adresse notamment mes vifs respects à Mme Ilona Browjohn pour le rôle
d’interlocutrice qu’elle a pu jouer et pour l’intérêt qu’elle a su porter à cette étude. Je remercie
également Mr Christian Guyot pour sa collaboration.
Je ne saurai oublier dans mes remerciements tous les membres des laboratoires qui m’ont
accompagné dans la réalisation de ce travail.
Je tiens à exprimer ma profonde reconnaissance à Mr Fabrice Martin-Laurent pour son
soutien indéfectible, sa disponibilité et pour l’excellence de ses commentaires et de ses
remarques scientifiques toujours très instructifs et formateurs. Merci à Marion Devers-
Lamrani, Jérémie Beguet ainsi que tous les acteurs de l’INRA de Dijon de l’UMR
Microbiologie du Sol et de l’Environnement, pour leur accueil, leur formation et le temps
qu’ils ont su me consacrer lors de mes différents séjours.
Que tous mes collègues du LCBE, du LGDP, de l’IUT de Perpignan, Département Génie
Biologique et plus généralement de l’UPVD, soient également assurés de ma profonde
gratitude pour les nombreux conseils et les suggestions qu’ils mont prodigués. Merci pour les
sourires, les discussions, les coups de mains, les encouragements, votre bonne humeur et
votre implication dans la réalisation de cette thèse. Ce travail est aussi le vôtre.

5
Merci à Alexia pour son implication dans cette étude ainsi que pour le sérieux de son travail.
Merci à Julie pour sa bonne humeur et sa présence.
Merci également à Susan qui m’a soutenu dans les moments difficiles et qui a toujours cru en
moi.
Enfin, je souhaiterai remercier ma famille qui m’a accompagné et encouragé durant ce long
travail. Merci à mes parents, à Audrey, ma sœur. Merci à Philippe et à Gigi, qui par leur
soutien et leur présence, m’ont permis d’arriver au bout.
Je dédie cette thèse à ma fille Nina et à ma grand-mère Marie-Louise.

6
Sommaire
REMERCIEMENTS 3
SOMMAIRE 6
TABLEDESILLUSTRATIONS 9
INTRODUCTION 18
PREMIEREPARTIE:REVUEBIBLIOGRAPHIQUE 22
Chapitre1Soletdiversitémicrobienne 231‐Contexte 232‐Composantemicrobiennedusol 243‐Diversitémicrobiennedusol 26
Chapitre2Méthodologiesnonmoléculairesappliquéesàl’étudedessouchesbactériennestelluriques 271‐Méthodescultivablestraditionnelles 272‐Méthodescultivablesinnovantes 29
Chapitre3Méthodologiesmoléculairesappliquéesàl’étudedessouchesbactériennestelluriques 311‐AnalysedesprofilsderestrictiondesADNr16Samplifiés(ARDRA) 312‐AmplificationPCRdesséquencesrépétitives(REP‐PCR) 333‐SéquençagedesADNr16SetAnalysephylogénétique 35
Chapitre4Dégradationbiologiquedesproduitsphytosanitairesdanslessols 411‐Dégradationbiologiqueetrécalcitrancemoléculaire 412‐Inductionsenzymatiquesetadaptionsmicrobiennes 423‐Aspectscinétiquesdeladégradationbiologique:Métabolismedirectetco‐métabolisme. 454‐Casparticulierdedégradationbiologique:labiodégradationaccélérée(BDA)despesticides 47
DEUXIEMEPARTIE:PRESENTATIONGENERALEDEL’ETUDE 51
Chapitre1Lafamilledesherbicidestricétones 521‐Contextehistorique 522‐Lasubstanceactive 543‐Moded’actiondelasubstanceactive 564‐Approchestoxicologiqueetécotoxicologique 605‐Comportementdanslesdifférentscompartimentsdel’environnement 61
Chapitre2Matriced’étude:lesoldePerpignan 671‐Situationgéographiquedelaparcelle 672‐ParamètresédaphiquesdusoldePerpignan 683‐Historiquecultural 694‐Stratégied’échantillonnage 695‐Dispositifexpérimental:lesmicrocosmes 70
TROISIEMEPARTIE:METHODOLOGIESDETRAVAIL 73

7
Chapitre1Méthodologiedel’analysechimique 741‐Analysedeséchantillonsdesols 742‐Analysedeséchantillonsdemilieuxdeculture 77
Chapitre2Méthodologiedel’analysebiologique 871‐Cadredel’étude:organisationdestravaux 872‐Approchecultivable:essaid’isolementdesouchesbactériennesdégradantlasulcotrione 873‐Approchemoléculaire:caractérisationdesisolatsbactériensdégradantlasulcotrionevsisolatsnondégradants 99
QUATRIEMEPARTIE:RESULTATSETDISCUSSION 119
Chapitre1ConfirmationdupouvoirbiodégradantdusoldePerpignanetrecherched’uneéventuellebiodégradationaccélérée(BDA) 1201‐RéactivitébiologiquedusoldePerpignan 1202‐Recherched’uneéventuellebiodégradationaccélérée(BDA) 1243‐Conclusion 127
Chapitre2Isolementdesouchesbactériennespotentiellementdégradantesdelasulcotrione 1291‐Recherchedesouchessulcotrione‐résistantes 1292‐Isolementdesouchesbactériennesmétabolisantlasulcotrione 1303‐Etudedupotentieldedégradationdesisolatsdégradants:suivideladissipationdelasulcotrionedanslemilieudeculture(MS) 1344‐Recherchedemétabolitesdelasulcotrionedanslemilieudeculture(MS) 137
Chapitre3Caractérisationphylogénétiquedesisolats1OPet1TRANS 1421‐Vérificationdelaqualitédel’ADNmatrice 1422‐AnalysedupolymorphismedeséquencedesADNr16Samplifiés(ARDRA)desisolats 1423‐Analysedesisolatsbactériensparamplificationdesséquencesrépétitives(REP‐PCR) 1444‐Séquençagedel’ADNr16Sdesisolats1OPet1TRANS 1455‐Conclusion 154
Chapitre4Caractérisationfonctionnelledesisolats1OPet1TRANS 1591‐Préambule 1592‐DétectiondesgènesXylEcodantl’enzymecatéchol2,3‐dioxygénase 1603‐DétectiondesgènesC12Ocodantl’enzymecatéchol1,2‐dioxygénase 1644‐SéquençagedesampliconsXylE 1655‐SéquençagedesampliconsC12O 165
CONCLUSIONGENERALEETPERSPECTIVES 167
REFERENCESBIBLIOGRAPHIQUES 173
VALORISATIONDESTRAVAUXDERECHERCHE 207
Publicationn°1(SOUSPRESSE) 208
Publicationn°2 233
Communicationscientifique 241
TABLEDESANNEXES 252
ANNEXE1 253

8
ANNEXE2 260
ANNEXE3 262
TABLEDESMATIERES 263

9
Table des illustrations

10
Liste des figures
Figure I-1-1 : Principales caractéristiques du sol en tant que microhabitat (Nannipieri et al.,
2003).
Figure I-2-1 : Répartition des différents phyla (%) représentés dans la banque « Australian
Collection of Microorganisms », après entrée en collection des isolats bactériens (Hugenholtz,
2002).
Figure I-3-1 : (a) Agrandissement d’un ribosome chez une bactérie. (b) Modélisation
tridimentionnelle du ribosome 70S composé de la grande sous-unité 50S (ARNr 5S + ARNr
23S + 31 protéines) et la petite sous-unité 30S (ARNr 16S + 21 protéines). (c) Exemple de
repliement dans l’espace de l’ARNr 16S provenant de la petite sous-unité 30S du ribosome
(Stern et al., 1988b). (d) Exemple de la structure secondaire de l’ARNr 16S de Vibrio.
parahaemolyticus X56580 présentant la structure secondaire des 4 principales séquences
provenant de différents isolats de Vibrio parahaemolyticus (Harth et al., 2007).
Figure I-3-2 : Principe de la REP-PCR.
Figure I-4-1 : Principaux mécanismes moléculaires intervenant dans le transfert génétique
horizontal (HGT), d’après Zaneveld et al. (2008).
Figure I-4-2 : Modélisation conceptuelle de l’adaptation des communautés bactériennes à la
biodégradation accélérée (BDA) de l’atrazine. Dispersion (flèches rouges) des gènes atz et trz
par conjugaison plasmidique et diversification (flèches vertes) du support des gènes atz par
transposition, d’après Devers et al. (2008).
Figure II-1-1 : Formule développée de la leptospermone (C15H22O4).
Figure II-1-2 : Structure de la 2-[2,4-benzoyl disubstitué]-1,3 cyclohexanedione.

11
Figure II-1-3 : Formes tautomériques dominantes de benzoyl-cyclohexane-1,3-dione en
solution aqueuse (X = Cl pour la sulcotrione, X = NO2 pour la mésotrione.
Figure II-1-4 : Comportement d’une benzoylcyclohexane-1,3-dione à caractère acide faible
en solution aqueuse.
Figure II-1-5 : Voie métabolique de biosynthèse des tocophérols et des plastoquinones à
partir du 4-Hydroxyphénylpyruvate chez la plante : site d’inhibition de l’enzyme 4-
phénylhydroxypyruvate dioxygénase par un herbicide tricétonique (sulcotrione).
Figure II-1-6 : Formules développées des principaux produits de dégradation de la
sulcotrione issus des voies A et B décrites dans la littérature.
Figure II-1-7 : Principaux photoproduits de la sulcotrione (chaabane et al, 2007).
(A) : (2-hydroxy-4-méthylsulfonyl)-1,3-cyclohexanedione, (B) : Acide 5,7-dicéto-7-(2-
chloro-4-méthylsulfonylphényl) heptanoïque, (C) : Acide 5,7-dicéto-(2-hydroxy-4-
méthylsulfonyl) heptanoïque, (D) : 1,3-cyclohexanedione, (E) : Acide 2-chloro-4-
méthylsulfonylbenzoïque, (F) : Acide 2-hydroxy-4-méthylsulfonyl benzoïque.
Figure II-1-8 : Variation de Ln Kobs en fonction de 1/T pour la dégradation de la sulcotrione
dans le sol de Belgique dans la gamme de températures 25, 40 et 60°C.
Figure II-2-1 : Plan de la parcelle expérimentale (IUT de Perpignan, UPVD).
Figure II-2-2 : Parcelle expérimentale (avant traitement phytosanitaire) située sur le site de
l’IUT de Perpignan, Université de Perpignan Via Domitia (Pyrénées Orientales).
Figure II-2-3 : Synoptique du calendrier de traitements et de prélèvements des essais en
microcosmes.
Figure II-2-4 : Préparation et mise en place des microcosmes.
Figure III-1-1 : Appareillage CLHP/UV Shimadzu utilisé pour les analyses de la sulcotrione
à partir de la matrice sol.

12
Figure III-1-2 : Chromatogramme CLHP/UV de la sulcotrione et de ses principaux
métabolites (CHD et CMBA).
Figure III-1-3 : (a) Spectre de masse (CLHP/SM-ESI) de la 1,3-cyclohexane dione (CHD) en
mode positif. (b) Chromatogrammes du standard analytique (10 mg.L-1) par détection barrette
de diodes et (c) détection SM par extraction du fragment m/z 113 en mode positif.
Figure III-1-4: (a) Spectre de masse de l’acide 2-chloro-4 methylsulfonyl benzoïque
(CMBA) (standard analytique à 20 mg.L-1) obtenu en CPG/SM (mode positif, SM, impact
électronique). (b) Chromatogramme montrant la présence du CMBA (fragments m/z, 217, 248
et 250) dans le milieu de culture (MS) supplémenté en sulcotrione de la souche 1OP après 27
jours d’incubation.
Figure III-1-5: Droite d’étalonnage de l’ester méthylique provenant du CMBA obtenu après
dérivation et analyse en CPG/SM.
Figure III-1-6: Droite d’étalonnage de la sulcotrione surajoutée dans le milieu de culture MS,
après extraction et analyses CLHP/UV.
Figure III-1-7: Principe de la réaction de dérivation de l’acide 2-chloro-4 methylsulfonyl
benzoïque (CMBA), en présence d’un excès de triméthylsilyldiazométhane (TMSCHN2) et de
méthanol conduisant à la formation de l’ester méthylique correspondant.
Figure III-2-1 : Exemples d’appareillages et de matériels utilisés en culture Pasteurienne lors
de l’étude. IUT de Perpignan, Département Génie Biologique (UPVD).
Figure III-2-2 : Protocole d’isolement de souches bactériennes telluriques sulcotrione-
résistantes.
Figure III-2-3 : Cultures répétées des isolats sulcotrione-résistants avec pression de sélection
de l’herbicide.

13
Figure III-2-4 : Protocole de sélection de souches bactériennes sulcotrione-dégradantes après cinq cycles de cultures répétées.
Figure III-2-5 : Spectrophotomètre Nanodrop (ThermoFisherScientific) utilisé pour la
quantification des échantillons d’ADN génomique issus des isolats bactériens.
Figure III-2-6 : (a) Imageur de gel autonome (système UGenius, SynGene), (b) Système
compact d’électrophorèse, (c) Thermocycleurs PTC 200 Gradient Cycler (Bio-Rad et MJ
Research), disponibles au LGDP (UPVD).
Figure III-2-7 : Carte du vecteur de clonage utilisé lors de l’étape de ligation des produits
PCR, délimitant la positionde l’insert d’intérêt (insert cloned) situé entre les deux promoteurs
T7 et SP6 nécessaire à la transcription (plasmide pGEM®-T Easy Vector Ligation kit,
Promega, Madison, USA).
Figure III-2-8 : Croissance des colonies après étalement de 20 µL (gauche) et de 80 µL
(droite) de cellules compétentes Escherichia coli DH5α™ ayant intégré le vecteur de clonage
pGEM®-T easy sur milieu LB + ampicilline (20 µg.mL-1).
Figure III-2-9 : Séquenceur Applied Biosystems Hitachi 3130xl Genetic Analyzers.
Figure III-2-10 : Voies de clivage du noyau aromatique du catéchol (voies ortho et méta)
catalysées par les catéchol dioxygénases.
Figure IV-1-1 : Suivi de la dissipation de la sulcotrione dans les microcosmes de sol (4S) vs
(St4S) après traitements successifs.
Figure IV-1-2 : Suivi de la dissipation de la sulcotrione dans les microcosmes de sol (C) vs
(StC) après traitements successifs.
Figure IV-2-1 : Colonies isolées sur milieu minimum (MS) supplémenté en sulcotrione
comme seule source de carbone et/ou d’énergie après croissance à 24°C à l’obscurité.

14
Figure IV-2-2 : Suivi de la croissance des souches 1OP vs 1TRANS dans le milieu de culture
(MS) supplémenté en sulcotrione par mesure de l’absorbance à 600 nm.
Figure IV-2-3 : Suivi de la croissance des souches 1OP vs 1TRANS dans le milieu de culture
(MS) non supplémenté en sulcotrione par mesure de l’absorbance à 600 nm.
Figure IV-2-4 : Suivi de la dissipation de la sulcotrione en fonction du temps dans le milieu
de culture (MS) supplémenté et ensemencé par les souches 1OP ou 1TRANS. Le contrôle est
un milieu (MS) supplémenté non ensemencé.
Figure IV-2-5 : Suivi de l’accumulation de l’acide 2-chloro-4 methylsulfonyl benzoïque
(CMBA) et de la dissipation de la sulcotrione en fonction du temps dans le milieu de culture
(MS) supplémenté et ensemencé par la souche 1OP.
Figure IV-3-1 : Empreintes ARDRA des isolats 1OP et 1TRANS générées après digestion
des ADNr 16S par Alu I et migration sur gel d’agarose (1%). La taille des fragments est
indiquée en paires de bases (pb).
Figure IV-3-2 : Empreintes ARDRA des isolats 1OP et 1TRANS générées après digestion
des ADNr 16S par Hae III et migration sur gel d’agarose (1%). La taille des fragments est
indiquée en paires de bases (pb).
Figure IV-3-3 : Profils REP PCR des isolats 1OP et 1TRANS.
Puits 1, 2, 5 et 6: REP PCR réalisée avec 1 ng.µL-1 d’ADN matrice. Puits 3, 4, 7 et 8: REP
PCR réalisée avec 2 ng.µL-1 d’ADN matrice.
Figure IV-3-4 : Migration sur gel d’agarose (1%) des ADNr 16S des isolats 1OP et 1TRANS
après amplification PCR à l’aide des amorces universelles 27f et 1492r.
Figure IV-3-5 : Mise en culture des colonies recombinantes (colonies blanches T7-SP6-PCR
positives) pour la préparation de l’ADN plasmidique (MiniPreps) destiné au séquençage, à
partir des banques de clones d’ADNr 16S des isolats 1OP et 1TRANS (milieu gélosé LB +
Ampicilline).

15
Figure IV-3-6 (a) : Migration sur gel d’agarose (1% ) des amplicons après amplification PCR
à partir des colonies blanches issues de la banque d’ADNr 16S de la souche 1OP.
Figure IV-3-6 (b) : Migration sur gel d’agarose (1% ) des amplicons après amplification PCR
à partir des colonies blanches issues de la banque d’ADNr 16S de la souche 1TRANS.
Figure IV-3-7 : Alignement des séquences consensus des ADNr 16S des isolats 1OP et
1TRANS. BIODE : souche 1OP ; N-BIO : souche 1TRANS.
Figure IV-3-8 : Analyse phylogénétique des ADNr 16S des isolats Pseudomonas sp.
1TRANS (NBD) et Pseudomonas sp. 1OP (BD). Les numéros d’accession GenBank des
séquences d’ADNr16S utilisées pour l’analyse sont indiqués entre parenthèse dans la légende.
Figure IV-4-1 : Migration des produits PCR sur gel d’agarose (1,2%) après amplification de
l’ADN génomique des deux isolats 1OP et 1TRANS à l’aide des amorces XylEaf et XylEar.
Contrôle 1 : ADN génomique E. coli DH5α ; contrôle 2 : ADN plasmidique pCambia 3300.
La taille des fragments est indiquée en paires de bases (pb).
Figure IV-4-2 : Migration des produits PCR sur gel d’agarose (1,2%) après amplification de
l’ADN génomique des deux isolats 1OP et 1TRANS à l’aide des amorces C12Of et C12Or.
La taille des fragments est indiquée en paires de bases (pb).
Figure IV-4-3 : Alignement multiple des séquences provenant des fragments d’ADN (taille
approximative 500 pb) obtenus lors du ciblage du gène C12O chez les isolats 1OP et
1TRANS à l’aide du logiciel ClustalX V. 2.0.10 (Thompson et al., 1997).

16
Liste des tableaux
Tableau II-1-1 : Activité herbicide (DC50 : Dose moyenne nécessaire pour le contrôle de 50
% de 7 espèces différentes de dicotylédones) des composés 2-[2,4-benzoyl disubstitués]-1,3
cyclohexanedione (d’après Lee et al.,1998).
Tableau II-1-2 : Sensibilité des adventices à la sulcotrione pour un traitement en post-levée à
450 g.ha-1. TS = très sensible, S = sensible, MS = moyennement sensible, MR =
moyennement résistante, R = résistante (d’après Compagnon et Beraud, 1992).
Tableau II-1-3 : Toxicité et écotoxicité de la sulcotrione non formulée (The Pesticide
Manual, 2007).
Tableau II-2-1 : Propriétés pédologiques et physico-chimiques du sol de la parcelle de
l’Institut Universitaire de Technologie (IUT) de Perpignan, déterminées par les protocoles
standardisés (Laboratoire d’Analyses Agricoles de Perpignan et LCBE –UPVD).
Tableau III-1-1: Performances de la méthodologie d’analyse de la sulcotrione dans le sol.
Tableau III-1-2 : Origine, références et pureté des standards analytiques utilisés lors de
l’analyse chimique.
Tableau III-1-3 : Calibration de l’analyse sulcotrione + métabolites sur milieu de culture
avec l’appareillage CLHP Jasco).
Tableau III-2-1: Préparation du milieu de culture minimum (MS).
Tableau III-2-2: Composition du milieu de culture TS (pH 7,3 ± 0,2) pour 1 litre d’eau
purifiée.
Tableau III-2-3: Composition du tampon Knapp (pH 6,6) pour 1 litre d’eau purifiée.

17
Tableau III-2-4 : Préparation, composition et conditions de conservation du tampon Giescher
5X nécessaire à la réaction de REP-PCR.
Tableau III-2-5 : Préparation et composition du milieu Luria Bertani (LB) nécessaire à la
sélection des cellules bactériennes transformées par le plasmide pGEM®-T easy.
Tableau III-2-6 : Caractéristiques des amorces utilisées pour la détection des gènes codant la
catéchol 2,3-dioxygénase (C23O) et la catéchol 1,2-dioxygénase (C12O) par amplification
PCR.
Tableau III-2-7 : Quantité d’ADN (en ng) nécessaire par réaction de séquençage pour les
fragments PCR. Etalonnage du séquenceur Applied Biosystems Hitachi 3130xl Genetic
Analyzers.
Tableau IV-1-1 : Constantes cinétiques (k), temps de demi-vie (t1/2) et coefficients de
détermination (R2) relatifs à la cinétique de dégradation de la sulcotrione en conditions de
microcosmes pour le sol de Perpignan.
Tableau IV-1-2 : Paramètres cinétiques relatifs aux différents traitements successifs effectués
pour les sols (4S), (St4S), (C) et (StC).
Tableau IV-2-1 : Evolution des concentrations en substance active et métabolites dans le
milieu de culture (MS) supplémenté en sulcotrione et ensemencé par l’isolat bactérien 1OP
(ND : non détecté).
Tableau IV-3-1 : Quantification et évaluation de la qualité de l’ADN génomique des isolats
bactériens 1OP et 1TRANS.
Tableau IV-3-2 : Quantification et évaluation de la qualité des ADNr 16S des isolats
bactériens 1OP et 1TRANS obtenus après amplification PCR et purification.

18
Introduction

Introduction
19
La présence de résidus de composés xénobiotiques dans les principaux compartiments
de l’environnement est avérée depuis plusieurs décennies. Les diverses activités anthropiques
(développement urbain, pollutions industrielles, agriculture, épandage des boues de stations
d’épuration, …) exercent des pressions environnementales variables, que les institutions
nationales et/ou supranationales s’efforcent d’identifier et de limiter (plan national Ecophyto
2018, législation Européenne REACH,…).
L’agriculture intensive conventionnelle participe de manière notable à la dégradation de la
qualité des eaux de surface et souterraines, du sol et de l’atmosphère, consécutivement à
l’utilisation d’intrants agricoles tels que les engrais, le lisier et les produits
phytopharmaceutiques. Dans son bulletin daté de juin 2010 (L’environnement en France :
l’eau), le Service de l’Observation et des Statistiques du Commissariat Général au
Développement Durable (ex IFEN) a publié des données relatives à l’état des cours d’eau en
2007 : 82 % des stations sélectionnées présentaient une concentration en pesticides totaux <
0,5 µg.L-1 en moyenne annuelle, les teneurs supérieures (18 %) affectant les cours d’eau des
régions pratiquant une agriculture intensive (Midi-Pyrénées, bassin parisien, vallée du Rhône,
nord de la France). Les eaux souterraines apparaissent sensiblement moins contaminées.
Le rôle du sol, zone majoritaire de charge en produits phytopharmaceutiques (PPP), est
évidemment essentiel dans leur transfert vers les différents compartiments aquatiques
terrestres. Dans cette matrice, le comportement de ces composés est fortement conditionné par
plusieurs processus :
- La volatilisation, principalement en surface ;
- L’adsorption-désorption, contrôlant la distribution et la mobilité des molécules
pesticides ;
- La dégradation chimique, essentiellement hydrolyse, photolyse et oxydo-
réduction ;
- La dégradation biologique ;
- Les phénomènes de transport (diffusion moléculaire, convection, transport de
composés dissous ou en phase particulaire, …).
Les herbicides de la famille chimique des tricétones, mis sur le marché dans les années 90, ont
été homologués dans certains itinéraires techniques agricoles, notamment en maïsiculture,

Introduction
20
« bénéficiant » du retrait de substances actives comme l’atrazine. Ces molécules de nouvelle
génération (mésotrione et sulcotrione), employées à doses inférieures à celles de l’atrazine
(150 - 450 g.Ha-1 vs 1200 g.Ha-1) ont été qualifiées de composés « présentant des risques
minimes pour l’environnement (Mitchell et al., 2001). Cependant, afin de limiter les risques
de pollution future du milieu naturel et de la ressource en eau, diverses études ont été
entreprises afin d’établir le profil comportemental de cette famille chimique. Un premier
travail à été précédemment mené au Laboratoire de Chimie des Biomolécules et de
l’Environnement (LCBE) et a conduit, en collaboration avec BayerCropScience (BCS), à la
soutenance de thèse de H. Chaabane (2005). Cet auteur a étudié le comportement de la
sulcotrione dans différents sols agricoles (Belgique, Martinique, Landes et Perpignan) par une
approche phénoménologique « classique ». A propos de la rétention, quel que soit le type de
sol, la capacité d’adsorption s’est avérée modérée pour la substance active ainsi que pour un
de ses produits de dégradation principaux (acide 2-chloro-4-mésyl benzoïque, CMBA) et leur
désorption relativement aisée. La 1,3-cyclohexanedione (CHD), autre produit de dégradation,
est apparu plus fortement adsorbée sur les composantes du sol. Le transfert de ces composés a
été estimé modéré dans l’horizon 0-10 cm. Dans le cas de la dégradation abiotique, l’étude de
l’hydrolyse, en conditions naturelles puis en conditions de laboratoire, selon différentes
modalités (pH, température, présence du cation Fe++,…) a permis de qualifier la sulcotrione
de composé stable. La photolyse, étudiée dans diverses eaux (eau de laboratoire, eau de mer et
eau de rivière) a montré une vitesse de dégradation supérieure à celle observée pour
l’hydrolyse. La dégradation biologique, généralement associée à une dégradation abiotique,
est apparu prépondérante notamment dans le sol de Perpignan.
Le sol de Perpignan, présentant donc une réactivité biologique notable et ayant reçu
antérieurement des traitements répétés par la sulcotrione lors des travaux de H. Chaabane, a
constitué un outil précieux pour essayer de provoquer puis de caractériser une éventuelle
biodégradation accélérée (BDA) en conditions contrôlées, consécutivement à une possible
adaptation de la microflore tellurique par acquisition de gènes codant les enzymes
cataboliques responsables de la dégradation.
Par ailleurs, nous avons souhaité utiliser la réactivité avérée de ce sol de Perpignan afin d’en
isoler des microorganismes dégradants spécifiques, capables d’utiliser la sulcotrione comme

Introduction
21
seule source de carbone et /ou d’énergie. La culture de ces microorganismes au laboratoire, et
le souhait de les caractériser de la manière la plus formelle possible, nous ont amené à
proposer une stratégie expérimentale comprenant plusieurs étapes successives :
- sélection de milieux adaptés à la culture, au laboratoire, de souches telluriques,
généralement réputées comme difficilement cultivables sur milieux synthétiques ;
- sélection des souches d’intérêt ;
- évaluation de leur potentiel biodégradant ;
- caractérisations phylogénétique et fonctionnelle des isolats retenus.
L’ensemble de ces données sera discuté, avec pour objectif la perspective d’une valorisation
biotechnologique possible des isolats présentant le plus fort potentiel de dégradation.
En outre, ce travail devrait s’inscrire dans une thématique de recherche plus fondamentale
visant à appréhender certains mécanismes évolutifs conduisant à l’adaptation des
microorganismes en réponse aux changements de leur environnement.

22
Première partie: Revue bibliographique

Première partie – Chapitre 1 : Sol et diversité microbienne
23
Chapitre1‐Soletdiversitémicrobienne
1- Contexte
Tous les pesticides épandus sur les surfaces agricoles ou plus généralement sur tout
type de sol, sont plus ou moins rapidement dégradés. La dégradation peut ainsi s’appuyer sur
des mécanismes biochimiques consistant essentiellement en des réactions d’hydrolyse,
d’oxydation, de réduction et de déshalogénation (Van der Meer et al., 1992 ; Castillo et
Tortensson, 2007). Ce processus est qualifié de dégradation biologique, catalysée par les
microorganismes telluriques plus ou moins spécifiques, en aérobiose ou anaérobiose, avec
formation de métabolites. Ce mécanisme est dans la majorité des cas quantitativement plus
important que la dégradation chimique des composés xénobiotiques (exception faite de
composés très récalcitrants, comme les polychlorobiphényles).
Nous présenterons ici les principales composantes de ce processus avant de nous attarder sur
un cas particulier de dégradation biologique, qualifié de biodégradation accélérée (BDA).
Cependant, dans un premier temps, il nous a semblé nécessaire de considérer le sol comme
une matrice présentant ses propres spécificités, et dont le rôle sera prépondérant dans les
processus de biodégradation des produits phytosanitaires.
La phénoménologie des pesticides dans le sol fait intervenir de nombreux facteurs parmi
lesquels figurent la nature chimique de la substance, la formulation appliquée, mais également
la nature du sol, sa texture, ses propriétés physico-chimiques, sa capacité de rétention, la
diversité des communautés microbiennes présentes, …
Notre travail s’est donc limité à décrire l’étude de la biodégradation de l’herbicide
sulcotrione ; nous avons donc souhaité présenter, dans cette partie bibliographique : (i) le sol
en tant que microhabitat, source de diversité microbienne, (ii) les méthodologies retenues
pour caractériser les microorganismes et leur action et, (iii) le rôle de cette microflore
tellurique largement impliquée dans la dégradation des produits phytosanitaires.

Première partie – Chapitre 1 : Sol et diversité microbienne
24
2- Composante microbienne du sol
Le sol est un micro-habitat pourvu d’importantes propriétés distinctives (cf figure I-1-
1) qui en font une matrice originale (Kuster et Hattori, 1973). En premier lieu, la population
microbienne y est très diversifiée. En prenant la taille du génome d'Escherichia coli comme
unité de référence, Torsvik et al. (1996) ont estimé la présence d'environ 6000 génomes
bactériens différents par gramme de sol. A titre d’exemple, la biomasse microbienne dans un
sol de prairie tempérée s'élève environ à 1 à 2 t.Ha-1 de biomasse bactérienne et à 2 à 5 t.Ha-1
de biomasse fongique (Killham, 1994).
Le sol est un système structuré, hétérogène et discontinu, généralement pauvre en
éléments nutritifs. Les sources de carbone et d'énergie disponibles pour les microorganismes
vivants sont relativement plus faibles que les concentrations optimales en éléments nutritifs
obtenues au laboratoire pour assurer la croissance microbienne sur milieux synthétiques
(Stotzky, 1961). Les caractéristiques chimiques, physiques et biologiques de ces micro-
habitats diffèrent également dans le temps et l'espace. La taille des habitats dépend
principalement de la nature des organismes qui s’y trouvent : quelques µm pour les bactéries,
moins de 100 µm pour les champignons et entre 100 µm et 2 mm pour les acariens et
collemboles et, enfin, entre 2 et 20 mm pour les isopodes (Coleman et Crossley, 1996). De
plus, même si l'espace disponible est vaste dans le sol, l'espace biologique, occupé par les
microorganismes vivants, représente une faible proportion, généralement inférieure à 5% de
l'espace total disponible (Ingham et Horton, 1987). Une autre particularité du sol est la
présence de zones d'activités biologiques accrues : les agrégats de sols, présentant des
propriétés physico-chimiques différentes selon leur origine (Sexstone et al., 1985) et riches en
particules organiques (Parkin, 1987 ; Petersen et al., 1996) et la rhizosphère (Lynch, 1994 ;
Morgan et al., 2005) sont les principaux représentants de ces zones biologiquement actives.
De plus, de nombreux facteurs environnementaux contrôlent cette vie microbienne : les
sources de carbone et d'énergie, les substances minérales présentes, les facteurs de croissance,
l'eau disponible ainsi que sa composition ionique, la température et la pression, l’oxygénation,
le pH, les potentiels d'oxydo-réduction sont autant de paramètres écologiques influençant la
croissance des populations microbiennes telluriques. Enfin, la nature des surfaces et les
interactions spatiales entre les micro-organismes jouent également un rôle fondamental. Selon
Hattori (1973), près de 80 à 90% des micro-organismes telluriques se trouvent sur des

Première partie – Chapitre 1 : Sol et diversité microbienne
25
surfaces solides. Huang et al. (1998) et Chen et al. (2006) ont montré que certaines cellules
bactériennes produisent des polysaccharides extracellulaires interagissant avec les particules
d'argile et que ces complexes argilo-polysaccharides peuvent persister même après la mort
des microorganismes. Ainsi, la majorité de la microflore vit libre ou attachée à des surfaces, et
se répartit dans les films liquidiens entourant les particules solides et à l’intérieur d'agrégats
(Stotzky, 1961).
La phase solide du sol adsorbe d'importantes molécules biologiques comme les
protéines et les acides nucléiques. De cette façon, certaines enzymes extracellulaires
adsorbées par les minéraux argileux ou piégées par les molécules d’acides humiques peuvent
maintenir leur activité biologique, et sont protégés contre la protéolyse, la dénaturation
thermique et les effets du pH (Nannipieri et al., 2003). Les acides nucléiques adsorbés ou
éventuellement liés à des molécules d’acides humiques ou à des particules d'argile et de sable
sont protégés contre la dégradation par les nucléases, rendant donc possible les processus de
transfert génétique horizontal (HGT) rencontrés au sein des populations microbiennes
telluriques, facteur important de la diversité génétique des populations microbiennes
telluriques (Lorenz et al., 1991 et 1994 ; Trevors et al., 1996 ; Paget et al., 1992;
Pietramellara et al., 2002).
Figure I-1-1 : Principales caractéristiques du sol en tant que microhabitat (d’après Nannipieri et al., 2003).

Première partie – Chapitre 1 : Sol et diversité microbienne
26
3- Diversité microbienne du sol La diversité microbienne est un terme général qui comprend la diversité génétique, (c’est-à-
dire la répartition des données génétiques au sein des espèces microbiennes), la diversité des
espèces (bactéries et de champignons dans les communautés microbiennes) et la diversité
écologique correspondant à la variation de la structure des communautés, la complexité des
interactions et le nombre de niveaux trophiques. La diversité microbienne peut être mesurée
par le nombre d’espèces différentes (richness) ainsi que leur abondance relative (evenness)
dans la microflore du sol (Kennedy et Smith, 1998). En ce qui concerne la diversité
bactérienne, il est généralement admis qu’elle est comprise entre 5 x 103 et 5 x 104 espèces par
gramme de sol pour des abondances pouvant varier de 108 à 1011 cellules par gramme de sol
(Kennedy et Papendick, 1995 ; Torsvik et al., 2002 ; Curtis et al., 2002 ; Roesch et al., 2007).
Accéder à cette diversité du sol peut globalement se résumer à l’utilisation de deux
approches : (i) les techniques non moléculaires, représentées par les méthodes traditionnelles
de microbiologie pasteurienne, faisant appel aux approches cultivables par l’utilisation de
divers milieux synthétiques utilisés en fonction des objectifs définis (milieux sélectifs ou non
et/ou différentiels) ou par des comptages in situ des échantillons à l’aide de marqueurs
spécifiques ; (ii) les techniques moléculaires faisant appel à la biologie moléculaire et/ou aux
méthodes biochimiques.

Première partie – Chapitre 2 : Méthodologies non moléculaires appliquées à lʼétude des souches bactériennes telluriques
27
Chapitre2‐Méthodologiesnonmoléculairesappliquéesàl’étudedessouchesbactériennestelluriques
Dans cette partie bibliographique, nous nous limiterons à décrire brièvement le principe des
méthodologies retenues au cours de notre travail de thèse.
1- Méthodes cultivables traditionnelles
La possibilité de cultiver des souches bactériennes telluriques au laboratoire est
fondamentale car elle contribue à mieux appréhender leurs propriétés et leurs capacités
fonctionnelles. Historiquement, les techniques cultivables appliquées à la microbiologie du
sol ont connu un essor important à la fin du 19ième siècle avec les travaux de Winogradsky
(caractérisation des microorganismes responsables de la nitrification dans le sol ; Van Elsas et
al., 2007) et de Beijerinck (obtention de cultures pures de Thiobacillus denitrificans par
enrichissements successifs). Les techniques par enrichissements sélectifs, fondées sur
l’utilisation de nutriments cibles et/ou d’inhibiteurs spécifiques, ont permis de mettre en
évidence un nombre important et croissant de groupes spécifiques de microorganismes du sol.
L’unique procédure d’énumération des souches microbiennes vivantes présentes dans
leur habitat naturel a été pendant très longtemps la technique du nombre le plus probable ou
Most-Probable-Number (Bakken, 1997; Johnsen et al., 2001). La découverte de nouveaux
groupes taxonomiques ou fonctionnels n’a été possible que par la sélection de souches
microbiennes à partir de milieux gélosés spécifiques et la diversité cultivable étudiée par
isolement de ces colonies, complétées par une identification par typage moléculaire
(Vandamme et al., 1996). Certains auteurs ont également évalué la diversité des micro-
organismes cultivables en suivant la chronologie d’apparition des différentes colonies
identifiées sur le milieu gélosé choisi (Ishikuri et Hattori, 1985). Néanmoins, étudier la
diversité microbienne du sol par ce type de méthodologie présente des limites, non seulement
Première partie – Chapitre 2 : Méthodologies non moléculaires appliquées à lʼétude des souches bactériennes telluriques
28
parce que le nombre de micro-organismes cultivables sur milieux synthétiques est limité, mais
aussi parce que les procédures d’isolement sont très souvent extrêmement laborieuses. De
plus, ces techniques présentent une estimation biaisée de la diversité microbienne du sol.
Hugenholtz et al. (1998) et Hugenholtz (2002) rapportent que les microorganismes facilement
isolés représentent 0,1 à 1% de la totalité des espèces microbiennes du sol. A titre d’exemple,
pour un même échantillon de sol, le comptage in situ par microscopie à épifluorescence
permet de multiplier d’un facteur 100 à 1000 les résultats de dénombrements obtenus par les
techniques cultivables conventionnelles (Johnsen et al., 2001). Ce phénomène appelé « Great
Plate Count Anomaly » par Staley et Konopla (1985), a largement été décrit par de nombreux
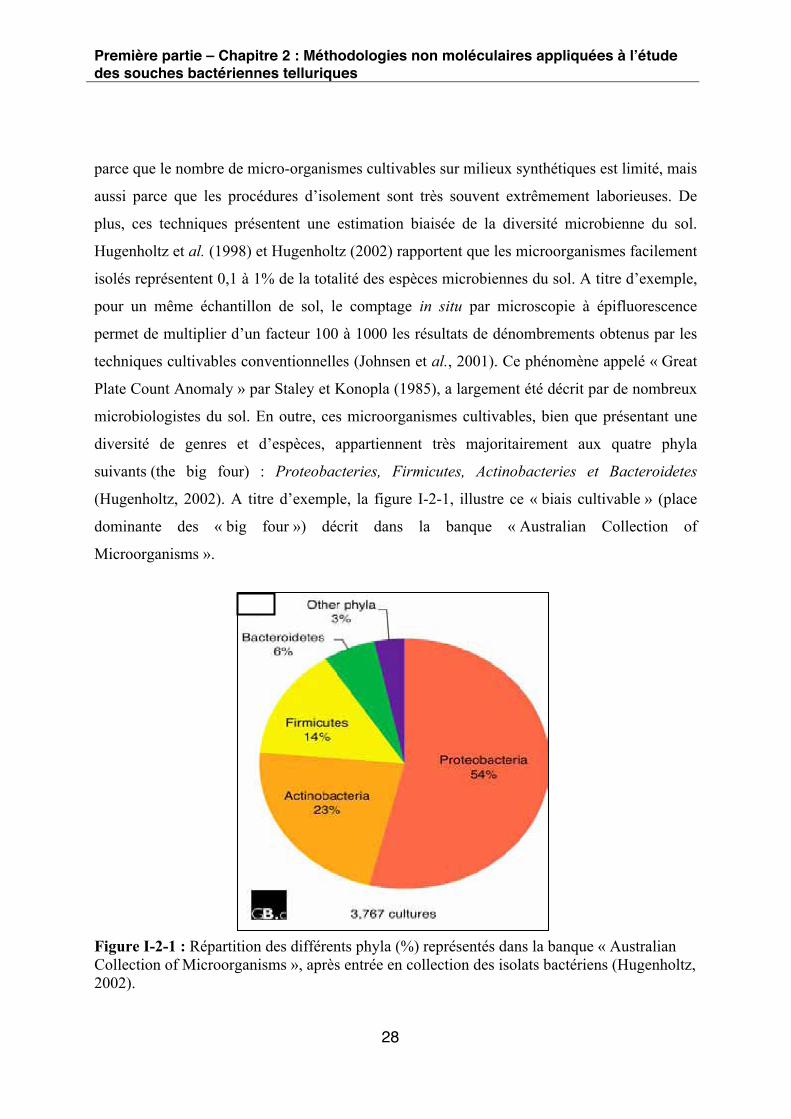
microbiologistes du sol. En outre, ces microorganismes cultivables, bien que présentant une
diversité de genres et d’espèces, appartiennent très majoritairement aux quatre phyla
suivants (the big four) : Proteobacteries, Firmicutes, Actinobacteries et Bacteroidetes
(Hugenholtz, 2002). A titre d’exemple, la figure I-2-1, illustre ce « biais cultivable » (place
dominante des « big four ») décrit dans la banque « Australian Collection of
Microorganisms ».
Figure I-2-1 : Répartition des différents phyla (%) représentés dans la banque « Australian Collection of Microorganisms », après entrée en collection des isolats bactériens (Hugenholtz, 2002).

Première partie – Chapitre 2 : Méthodologies non moléculaires appliquées à lʼétude des souches bactériennes telluriques
29
Ces données s’expliquent essentiellement par l'interdépendance trophique des
différents organismes présents dans un écosystème spécifique et à l'incapacité de créer, en
culture pure, les conditions environnementales rencontrées par les micro-organismes du sol.
Ainsi, seules certaines espèces microbiennes sont cultivables et uniquement sous certaines
conditions physiologiques (Bakken, 1997; Muyzer et Smalla, 1998; Heuer et al., 2001). De
nombreux auteurs ont en effet démontré que le choix du milieu de culture affecte
sensiblement les colonies formées (Sørheim et al., 1989; Johnsen et Nielsen, 1999). D’après
Bakken (1997), les cellules bactériennes de petites dimensions (ultramicrobactéries) ne sont
pas cultivables sur milieu gélosé car incapables de former des colonies en présence d’agar. En
considérant que les cellules de grandes dimensions occupent environ 80 % du volume total
dans lequel est confinée la totalité des bactéries telluriques, il a émis l’hypothèse que seules
ces dernières présentent une importance écologique majeure dans le sol.
Ce type de méthodologie demeure une approche fondamentale en écologie microbienne, mais
présente finalement des limites importantes d’accès à la diversité microbienne.
2- Méthodes cultivables innovantes
L’approche cultivable demeure une technique incontournable lorsque l’on souhaite
isoler et cultiver des souches microbiennes telluriques d’intérêt. De nouvelles méthodologies
alternatives sont aujourd’hui disponibles et laissent entrevoir des perspectives intéressantes
pour l’avenir. Globalement, ces technologies suivent trois stratégies distinctes mais
complémentaires (Keller et Zengler, 2004): (i) une amélioration des techniques cultivables
conventionnelles par optimisation des milieux synthétiques, (ii) une stimulation de la
croissance microbienne in situ par utilisation directe de l’échantillon environnemental comme
milieu de culture approprié, supplémenté ou non avec de faibles concentrations en nutriments
et, (iii) un développement de protocoles de cultures haut-débit accompagnées de système
haute résolution de détection de la croissance microbienne.
Ainsi, en jouant de façon synergique sur les divers paramètres contrôlant les étapes du
protocole d’isolement (sonication des particules de sol, faible concentration en nutriments,
pH, substitution de l’agar par de la gomme gellane, longue durée d’incubation et observation
minutieuse des colonies en développement sur la gélose), Janssen et al. (2002) ont pu isoler

Première partie – Chapitre 2 : Méthodologies non moléculaires appliquées à lʼétude des souches bactériennes telluriques
30
pour la première fois de nouvelles souches bactériennes réparties dans différentes divisions
(Acidobacteria, Actinobacteria, Proteobacteria et Verrucomicrobia) et jusqu’alors décrites
seulement par des méthodes non cultivables.
L’utilisation de faibles concentrations de nutriments dans les milieux d’enrichissement peut se
révéler efficace pour isoler des souches à faible vitesse de croissance, généralement perdues
dans des milieux conventionnels au profit de celles à développement rapide (Connon et
Giovannoni, 2002). Le développement de certaines colonies sur milieux synthétiques peut
être excessivement lent et demander des semaines, voire des mois avant d’être observables à
l’œil nu (Harris et Paul, 1994 ; Davis et al., 2005 et 2011).
D’après des études relativement récentes (Janssen et al., 2002 ; Joseph et al., 2003 ; Sangwan
et al., 2005 ; Stevenson et al., 2004 ; Zengler, et al., 2002 ; Kaeberlein et al., 2002 ; Ferrari et
al., 2005 ; Davis et al., 2005, 2011 ; Sait et al., 2002 et 2006 ;Vartoukian et al., 2010), la mise
au point de nouveaux milieux de culture et le développement de techniques cultivables
innovantes permettent d’envisager de nouvelles perspectives d’isolement et d’entrée en
collection de souches microbiennes telluriques non cultivables sur milieux conventionnels.
Selon certains auteurs comme Nichols (2007) ou Ellis et al. (2003), il est donc nécessaire de
nuancer quelque peu le paradigme qui consistait à considérer la majorité des espèces
microbiennes présentes dans le sol comme étant non cultivables.

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
31
Chapitre3‐Méthodologiesmoléculairesappliquéesàl’étudedessouchesbactériennestelluriques
Les techniques moléculaires impliquent généralement l'extraction des acides
nucléiques (ADN ou ARN) issus des populations microbiennes de façon directe ou indirecte à
partir du sol. Elles sont indépendantes des méthodes cultivables et ont un pouvoir de
résolution variable qui peut généralement être ajusté en fonction des objectifs, de l’analyse
des grands groupes taxonomiques jusqu’à la mise en évidence d’espèces au sein
d’échantillons de sols. Nous nous limiterons à décrire les techniques utilisées dans le cadre de
notre étude.
1- Analyse des profils de restriction des ADNr 16S amplifiés (ARDRA)
Les travaux de Carl Woese (1987) ont véritablement révolutionné l’approche
taxonomique en microbiologie. Ils ont démontré que les séquences d’ADN ribosomaux
(ADNr) présentes sous forme d’opérons chez tous les microorganismes (leur nombre variant
environ de 1 à 15 copies par génome bactérien (Rainey et al., 1996) étaient des marqueurs
évolutifs relativement robustes, semblables à des « chronomètres », très utiles pour déchiffrer
la phylogénie et l’évolution des populations microbiennes au cours du temps (Woese, 1987).
Les ribosomes (cf figure I-3-1) sont formés de deux sous-unités (une grande sous-unité 50S et
une petite sous-unité 30S) comportant chacune des protéines ribosomales (31 et 21 protéines
respectivement) assemblées sur une matrice d'acides ribonucléiques (ARNr). Les ARN
ribosomiques (petite sous unité = ARNr 16S, grande sous-unité = ARNr 23S) se sont imposés
comme référence de taxonomie moléculaire car ils réunissent l'ensemble des qualités requises.
En premier lieu, ils sont un élément clé de la synthèse protéique, fonction très conservée car
indispensable à la vie de la cellule. Ensuite, ils ont une structure particulière faite d'une
succession de domaines dont les vitesses d'évolution sont très variables, de relativement
élevée à presque nulle et chacun de ces domaines a son importance pour l'identification
moléculaire des micro-organismes. Certaines parties sont identiques chez toutes les bactéries
et donc utilisables comme sites de complémentarité pour des amorces universelles de

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
32
séquences ou d'amplification (PCR). La comparaison des domaines conservés permet de
retracer les liens de parenté qui unissent des bactéries éloignées, tandis que les domaines à
vitesse d'évolution plus rapide permettent l'étude des relations phylogénétiques d'espèces plus
proches. D'autres parties de séquences sont propres à un groupe et permettent ainsi
l'identification de séquences dites signatures caractéristiques d'ordres taxonomiques différents
(espèce, genre, famille ou royaume). A titre d’exemple, Heuer et al. (1999) ont indiqué que 14
régions différentes de l'ADNr 16S (A, B, C, D, E, F, V1–V3 et V5–V9) ont pu être utilisées
pour générer des empreintes des communautés bactériennes.
Figure I-3-1 : (a) Agrandissement d’un ribosome chez une bactérie. (b) Modélisation tridimentionnelle du ribosome 70S composé de la grande sous-unité 50S (ARNr 5S + ARNr 23S + 31 protéines) et la petite sous-unité 30S (ARNr 16S + 21 protéines). (c) Exemple de repliement dans l’espace de l’ARNr 16S provenant de la petite sous-unité 30S du ribosome (Stern et al., 1988b). (d) Structure secondaire de l’ARNr 16S de Vibrio. parahaemolyticus X56580 présentant les quatre principales séquences provenant de différents isolats de Vibrio parahaemolyticus (Harth et al., 2007).

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
33
L’ARDRA consiste, après amplification par PCR d’une partie de la séquence du gène de
l’ADNr 16S, à analyser, après digestion enzymatique, les profils de restriction obtenus entre
différents échantillons d’ADNr 16S. Cette méthodologie est, d’une façon générale et quel que
soit le gène auquel elle est appliquée, appelée analyse du polymorphisme de longueur des
fragments de restriction ou RFLP (Restriction Fragment Length Polymorphism). En revanche,
lorsqu’elle est appliquée à des ADN ribosomaux (ADNr), on parle plutôt d’analyse de
restriction d’ADN ribosomaux amplifiés ou d’ARDRA. Cette technique a été reconnue
comme étant un outil d’analyse phylogénétique fiable et a donc été largement utilisée dans de
nombreuses études taxonomiques sur divers microorganismes cultivables ou non cultivables,
issus de différents échantillons environnementaux (Vaneechoutte et al., 1992, 1993;
Martinez-Murcia et al., 1995). Les travaux de Moyer et al. (1994) ont démontré que
l’utilisation de 2 endonucléases reconnaissant des sites de restriction à 4 nucléotides
permettait de discriminer la majeure partie des ADNr 16S présents au sein d’un mélange
complexe, en révélant le polymorphisme de séquence de l’opéron ribosomique bactérien
(Massol-Deya et al., 1995). Il est communément admis que l’ARDRA permet de différencier
des échantillons jusqu’au niveau du genre bactérien (Martin-Laurent et al., 2001). Toutefois,
il est faut être très prudent dans l'interprétation de la composition des communautés
microbiennes car la méthode d'extraction des ADN peut influencer les résultats obtenus
(Martin-Laurent et al., 2001).
2- Amplification PCR des séquences répétitives (REP-PCR)
L’amplification des séquences répétitives (REP-PCR), permet l’amplification, par la
réaction de polymérisation en chaine (PCR), de fragments d’ADN de tailles différentes
(amplicons) constitutifs de séquences extragéniques (non codantes) répétitives et
palindromiques présentes dans de nombreux génomes microbiens. Les séquences REP,
comprises à l’intérieur de séquences non traduites des opérons bactériens, ont été décrites
comme étant vraisemblablement des éléments génétiques régulateurs potentiels, (Townsend et
al., 1997) intervenant notamment dans leur capacité à former des structures stables en boucles
lors du processus de transcription. Dès 1984, les premières séquences REP ont été mises en
évidence chez Escherichia coli et Salmonella typhimurium par Stern et al. (1984). Ces

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
34
éléments palindromiques, constitués de séquences consensus conservées (environ 38 paires de
bases), pouvant former une structure stable sous forme de boucle avec une région centrale
variable d’environ 5 paires de bases, sont largement présents dans les génomes de la plupart
des genres bactériens. Les séquences REP ont donc été utilisées comme amorces pour
l'amplification par PCR de régions de l'ADN génomique bactérien (de Bruijn, 1992). Ainsi,
lors de la réaction, les amorces utilisées se fixent sur plusieurs séquences d’ADN réparties
dans tout le génome des microorganismes. Plusieurs fragments amplifiés, hautement
spécifiques et de différentes longueurs sont donc ainsi facilement obtenus et séparés par
électrophorèse sur gel d’agarose. Le principe de cette technique est présenté figure I-3-2.
En écologie microbienne moléculaire, la présence des éléments REP dans la grande majorité
des génomes des Eubactéries présente une approche d’investigation moléculaire relativement
utile dans la caractérisation d’échantillons environnementaux. Cette méthodologie de travail
permet (i) de rendre compte de la variabilité de l'ensemble du génome bactérien (Versalovic et
al., 1991), (ii) d'analyser les génomes ou de différencier des souches bactériennes appartenant
à des groupes phylogénétiquement très proches pouvant aller jusqu’au niveau de l’espèce,
(iii) de rendre compte de la dispersion des séquences REP caractéristiques de chaque souche
bactérienne (Menna et al., 2009) et, enfin, (iiii) de réaliser une analyse de la diversité
génotypique plus fine que par la technique de l’ ARDRA. (Frey et al., 1996).
Figure I-3-2 : Principe de la REP-PCR

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
35
3- Séquençage des ADNr 16S et Analyse phylogénétique 3-1- Principe général
La systématique peut être définie comme une nomenclature hiérarchisée des
organismes vivants, lié à la théorie de l’évolution, et dont l’objectif principal est d’assurer une
classification des espèces s’appuyant sur les relations phylogénétiques (Harayama et Kasai,
2006). L’espèce constitue l’unité taxonomique fondamentale. Wayne et al. (1987) ont proposé
une définition de l’espèce microbienne s’appuyant sur les propriétés de dénaturation et de
renaturation de l’ADN génomique. Ainsi, les valeurs d’hybridation ADN-ADN entre deux
souches supérieures ou égales à 70 %, accompagnées de valeurs de ∆Tm inférieures ou égales
à 5°C, constituent des bornes raisonnables et acceptables de définition de l’espèce
microbienne. Néanmoins, certains auteurs ont pu mettre en évidence que cette méthodologie
présentait certains biais (Stackebrandt et Goebel, 1994 ; Vandamme et al., 1996). En outre,
Rossello-Mora et Amann (2001) ont montré que pour décrire de nouvelles espèces
microbiennes, une approche polyphasique intégrant des données à la fois génotypiques et
phénotypiques s’avérait nécessaire. Les spécialistes de la systématique bactérienne n’ont donc
pas encore totalement trouvé de consensus satisfaisant pour définir sensu stricto le terme
d’espèce (Cohan, 2002). En effet, donner une définition de l’espèce bactérienne applicable à
tous les microorganismes semble difficile, aucun critère de classification phylogénétique
n’étant suffisamment discriminant, stable et universel (Stackebrandt, 2003).
Malgré les biais inhérents à chaque méthode, l’étude de l’hybridation ADN-ADN associée à
l’analyse des séquences d’ADNr 16S constitue une méthodologie très largement admise par
les microbiologistes pour discerner les souches microbiennes étroitement apparentées.
Historiquement, le début des phylogénies moléculaires basées sur l’analyse comparée
de séquences remonte aux années 60, avec les travaux de Zucklerkandl et Pauling (1965) et
Fitch et Margoliash (1967) sur les séquences protéiques de cytochrome c et de globines.
Dans le cadre de notre étude, l’analyse phylogénétique s’est appuyée sur le séquençage des
ADNr 16S (Woese et al., 1990). Cette procédure s’est déroulée en plusieurs étapes
consécutives : (i) amplification des séquences d’ADNr 16S (environ 1500 paires de bases) à
partir d’amorces universelles de la division Bacteria, (ii) séquençage, (iii) édition et
alignement des séquences obtenues par comparaison avec des séquences de références

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
36
présentes dans les bases de données internationales, (iiii) estimation des distances évolutives
entre les séquences concernées par application de modèles évolutifs et enfin, (iiiii)
construction d’un arbre phylogénétique à partir des distances évaluées précédemment par
l’application d’un algorithme associé à une analyse statistique afin de tester la robustesse de
l’arbre retenu.
Il nous a semblé opportun, dans cette partie bibliographique, de rappeler le principe général
des méthodologies de l’après-séquençage nécessaires à l’exploitation des données
génomiques.
3-2- Méthode d’alignement
Pour construire un arbre à partir de séquences nucléotidiques, il faut tout d'abord éditer
ces séquences afin d’en éliminer le début et de la fin (baisse de qualité, donc de confiance),
d’identifier et d’éliminer les pics parasites (à géométrie irrégulière) et les fragments du
vecteur de clonage. Il s’agit également de corriger les ambiguïtés dues à des superpositions de
pics insuffisamment discriminés par le programme et assurer manuellement une correction
des erreurs.
L’alignement consiste à faire correspondre les positions dans la séquence ayant une origine
commune avec les autres séquences disponibles. Cette étape permet de mettre en regard des
nucléotides homologues présents dans les séquences d’ADNr 16S. Le travail d’alignement est
particulièrement délicat à réaliser, et, même s’il existe des logiciels adaptés, les alignements
nécessitent d’être très souvent réajustés manuellement. Deux nucléotides pourront être
homologues et informatifs pour deux espèces proches alors qu’ils ne le seront pas pour deux
espèces lointaines. En outre, les phénomènes d’homoplasie (similarité ne dérivant pas d’un
caractère ancestral) compliquent l’analyse des séquences dans les zones hypervariables. Les
homoplasies sont consécutives à des évènements de type réversion (changement dans une
séquence annulé par un second changement G→ C→G), parallélisme (changements parallèles
dans deux séquences conduisant à des nucléotides identiques) ou convergence (séquences
ancestrales différentes qui évoluent vers des séquences identiques).
En fonction des séquences à analyser, il convient donc de déterminer quelles sont les
positions informatives et de délimiter les zones réellement comparables. Pour évaluer la
confiance que l’on peut avoir dans un alignement, il est possible de s’assurer que les

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
37
séquences signatures d’un groupe phylogénétique sont bien en regard les unes des autres.
L’analyse des structures secondaires s’avère, également, très souvent nécessaire.
3-3- Reconstruction des arbres phylogénétiques
La phylogénie moléculaire a pour objectif de retrouver les liens de parenté entre des
séquences de nucléotides. Ces relations de parenté sont généralement représentées sous forme
d’arbre, composé de branches et de nœuds, les branches connectant les nœuds eux-mêmes
définis comme des points au niveau desquels deux branches ou plus divergent. Les branches
et les nœuds peuvent être internes ou externes. Dans le cas d’un arbre de gènes, un nœud
interne correspond à la séquence ancestrale hypothétique avant le ou les évènements de
duplications qui ont donné naissance aux séquences actuelles. Les nœuds terminaux
(également dénommés « Operational Taxonomic Units » ou OTU) correspondent aux
séquences à partir desquelles l’arbre a été construit. Les branches définissent les relations de
parenté entre les différents nœuds. Un nœud interne et l’ensemble des OTU qui en dérivent
forment un groupe monophylétique ou clade. Un groupe incluant un nœud interne et une
partie seulement des OTU qui en dérivent est appelé groupe paraphylétique. Enfin, un groupe
rassemblant des OTU n’ayant pas de nœud interne qui leur est spécifique est appelé groupe
polyphylétique.
L’arbre phylogénétique peut être enraciné ou pas. La racine constitue l’ancêtre commun de
toutes les OTU représentées dans l’arbre. La direction entre la racine et les différentes OTU
représente le temps évolutif. Dans un arbre non enraciné, seules les relations entre les
différentes OTU sont représentées.
Il existe différentes méthodes pour calculer des arbres phylogénétiques à partir d’alignements
multiples de séquences. Ces méthodes sont généralement regroupées en trois catégories : (i)
les méthodes de distance, (ii) les méthodes de maximum de parcimonie et (iii) les méthodes
probabilistes.
Les méthodes de distance sont les plus simples et les plus rapides à mettre en oeuvre. Elles
sont basées sur l’utilisation d’une matrice où sont consignées les distances calculées pour
toutes les combinaisons deux par deux des séquences analysées. Des algorithmes de calculs

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
38
sont ensuite utilisés pour déduire, à partir de la matrice de distance, les relations
phylogénétiques des OTU. Dans la méthode de distance, la longueur des branches sera
proportionnelle à la distance. A côté de la méthode UPGMA « Unweighted Pair Group
Method with Arithmetic mean » (Sneath et Sokal , 1973) et de la méthode de Fitch-
Margoliash (1967), nous avons utilisé la méthode de Neighbor-Joining (Saitou et Nei, 1987).
Cette méthodologie consiste à regrouper les séquences les plus similaires entre elles (c’est-à-
dire ayant une distance faible), ce regroupement se faisant par étapes, avec comme contrainte
principale de minimiser la longueur totale de l’arbre. Cette méthode donnera toujours un seul
arbre pour une matrice de distance donnée.
Le principe des méthodes de maximum de parcimonie est basé sur l’identification, parmi
tous les arbres phylogénétiques possibles, de celui qui demande le plus petit nombre de
modifications évolutives (« pas évolutifs ») pour expliquer les différences observées entre les
séquences ou OTU qui sont étudiées. La longueur des branches des arbres obtenus sera
proportionnelle au nombre de modifications évolutives requises pour passer d’un nœud à
l’autre.
Les principales différences avec la méthode de distance sont : (i) la totalité des sites de la
séquence n’est pas utilisé mais seuls les sites dits informatifs (c’est-à-dire ceux qui vont
tendre à favoriser un ou plusieurs arbres par rapport aux autres) sont pris en compte, (ii)
plusieurs arbres phylogénétiques sont possibles avec cette méthode de reconstruction.
Si la construction de l’arbre est effectuée dans le cadre d’une analyse cladistique, seront
considérés comme informatifs uniquement les sites où plusieurs OTU partagent un état
dérivé. En pratique, dès qu’il y a plusieurs OTU et un nombre de sites importants, la
construction nécessite un équipement informatique performant car le nombre d’arbres
possible augmente de manière exponentielle avec le nombre d’OTU. Il arrive donc très
souvent qu’il soit impossible d’estimer tous les arbres. La solution alternative est
généralement d’utiliser des approches « heuristiques » qui ne vont explorer qu’une partie des
arbres possibles : il s’agit généralement de partir d’un arbre choisi aléatoirement et de le
modifier tout aussi aléatoirement. Le nombre de pas évolutif est à chaque fois calculé pour
chaque arbre obtenu. L’analyse se perpétue tant que l’on obtient des arbres plus parcimonieux
que les précédents.

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
39
La méthode de maximum de parcimonie permet ainsi d’obtenir plusieurs arbres
équiparcimonieux possibles, ne diffèrant généralement les uns des autres que par quelques
nœuds. La construction d’un arbre consensus s’avère donc nécessaire.
Parmi les méthodes probabilistes, la méthode de maximum de vraisemblance ou
« Maximum of Likelihood » (Felsentein, 1973) est généralement la plus utilisée. Elle s’appuie
sur l’utilisation de modèles d’évolution, c’est-à-dire un ensemble d’hypothèses traduisant les
processus d’évolution possibles des séquences concernées. Par exemple, le principe est de
proposer des algorithmes donnant les probabilités de changement d’un nucléotide vers un
autre, différentes pour les transitions (substitution d’un nucléotide comportant une purine par
un autre nucléotide comportant également une purine ou substitution d’un nucléotide
comportant une pyrimidine par un autre nucléotide comportant également une pyrimidine) et
les transversions (susbtitution d’un nucléotide comportant une purine par un nucléotide
comportant une pyrimidine ou inversement). Cette modélisation va estimer la probabilité de
réaliser l’alignement de séquences en fonction de différentes hypothèses. Cette probabilité est
appelée la vraisemblance. La méthode de maximum de vraisemblance va donc consister à
retenir, parmi un ensemble d’arbres possibles, celui dans lequel la vraisemblance est
maximale. L’arbre retenu dans ce cas, est considéré comme la représentation phylogénétique
la plus plausible, étant donné le modèle d’évolution utilisé. La limitation principale des
méthodes probabilistes est le temps de calcul nécessaire à trouver l’arbre maximisant la
vraisemblance. En outre, cette méthodologie implique également d’utiliser le modèle
d’évolution le plus proche de la réalité.
3-4- Estimation de la robustesse des arbres
Toutes les méthodes d’analyse phylogénétique possèdent des avantages et des
inconvénients car aucune n’incorpore véritablement tous les aspects des mécanismes
biologiques de l’évolution des séquences. Ainsi, aucune méthode probabiliste disponible n’est
donc susceptible de fournir un arbre phylogénétique « vrai ». Généralement, seules les
branches des arbres retrouvées par les trois méthodes (Neighbor-Joining, maximum de
parcimonie et maximum de vraisemblance) sont susceptibles de s’approcher de la réalité
(Huelsenbeck et Hillis, 1993).

Première partie – Chapitre 3 : Méthodologies moléculaires appliquées à lʼétude des souches bactériennes telluriques
40
Néanmoins, d’autres méthodes d’évaluation sont possibles. C’est le cas de l’analyse en
boostrap (Felsenstein, 1985 ; Felsenstein et Kishino, 1993). C’est la méthode la plus souvent
utilisée pour tester la fiabilité des branches internes de l’arbre. Elle consiste à effectuer un
tirage aléatoire des sites au hasard avec remise.
Cette opération est effectuée entre 100 (pour les arbres de maximum de parcimonie ou de
maximum de vraisemblance) et 1000 fois (arbres de distances) et un arbre est proposé pour
chaque échantillonnage. Les différentes topologies d’arbres sont ensuite comparées et
compilées en un arbre consensus, présentant un pourcentage de robustesse attribué à chacun
des nœuds de l’arbre en fonction du nombre de fois où celui-ci a été retrouvé à cette position.
En pratique, seules les branches définies par une valeur supérieure à 90 % sont considérées
comme fiables.
C’est cette méthode que nous avons retenue dans notre travail de caractérisation des souches
d’intérêt.

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
41
Chapitre4‐Dégradationbiologiquedesproduitsphytosanitairesdanslessols
1- Dégradation biologique et récalcitrance moléculaire
Les microorganismes assurent une fonction essentielle du sol grâce à leur grande
diversité (Topp, 2003). Cette diversité des communautés microbiennes telluriques jouerait un
rôle important dans la dégradation des pesticides (Entry et Emmingham, 1995 ; Voos et
Groffman, 1997). Cependant, d’autres auteurs ont également observé une forte corrélation
entre la quantité de biomasse d’un sol ou l’activité microbienne et la dégradation des
pesticides (Anderson, 1984 ; Walker et Welch, 1989 ; Wardle et Parkinson, 1991 ; Flint et
Witt, 1997; Walker et al., 2001 ; de Lipthay et al., 2007). Une des conditions fondamentales à
la dégradation biologique est la présence d’accepteurs d’électrons. Dans les systèmes
aérobies, l’accepteur final d’électrons est l’oxygène moléculaire (Castillo et Tortensson,
2007) alors qu’en conditions anaérobies, les accepteurs d’électrons inorganiques sont très
majoritairement le NO3-, SO4
2-, S, CO2 ou Fe3+. Les premières étapes de la dégradation
provoquent les transformations de la structure moléculaire du pesticide avec l’apparition de
métabolites pouvant, dans certains cas, être plus toxiques que la molécule mère. La
minéralisation de la substance active, avec la transformation du carbone organique en CO2 est
le processus ultime. Il n’y a pas de relation univoque entre la stabilité chimique d’une
molécule et sa stabilité biologique.
Les réactions de transformation ou de dégradation des pesticides par les populations
microbiennes telluriques sont essentiellement des réactions d’oxydation, de réduction et
d’hydrolyse. Les réactions d’oxydation sont majoritairement représentées par les mécanismes
d’hydroxylation, déalkylation, époxydation et sulfoxydation. Ces systèmes réactionnels sont
généralement catalysés par les familles d’enzymes suivantes : les mono et di-oxygénases, les
laccases et les peroxydases. Les microorganismes ont la possibilité de dégrader les polluants
aromatiques par ortho ou méta clivage. La position des substituants a également un impact
important sur la biodégradation de la molécule (Baker et Woods, 1977, Baker et Mayfield,
1980). Outre la récalcitrance propre à la molécule, les processus de dégradation biologique

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
42
dépendent également de nombreux facteurs physiques, chimiques et physiologiques dont le
degré d’importance varie selon l’environnement (Alexander, 1994).
Cependant, les polluants présentant une structure chimique électroniquement stable ou ayant
de nombreux substituants halogénés sont généralement les plus difficiles à dégrader (Sims et
al., 1986 ; Goulding et al., 1988 ; Scheneurt et al., 1993). Dans ce cas, la stratégie
communément retenue par la microflore peut consister en l’introduction de groupes polaires
qui ont pour effet d’accroître le caractère hydrophile et de faciliter ainsi leur dégradation
ultérieure.
2- Inductions enzymatiques et adaptions microbiennes
La biodégradation d’un pesticide dans le sol suppose que les microorganismes
présents soient capables de mobiliser des fonctions catalytiques appropriées. La spécificité
enzymatique, parfois étroite, est rarement absolue (Soulas, 1990). Ainsi, la biodégradation
d’un grand nombre de composés chimiques de synthèse est très souvent accomplie par des
voies métaboliques préexistantes qui permettent aux microorganismes d’utiliser des substrats
naturels. Cette prédisposition enzymatique reposerait sur la similitude de certains composés
naturels d’origine végétale avec certaines molécules phytosanitaires, appelées molécules
analogues (Alexander, 1994). L’attaque du xénobiotique a donc un caractère fortuit
(Knackmus, 1981 ; Slater, 1984) et peut s’opérer jusqu’à un stade avancé qui peut aller
jusqu’à la minéralisation complète (Schmidt et al., 1987). En outre, certains auteurs (Parales
et al., 2002 ; Pandey et Jain, 2002) précisent que l’induction enzymatique s’accompagnerait
d’une aptitude chimiotactique des microorganismes. Pour autant, cette analogie structurale
n’est pas la seule explication possible du phénomène de biodégradation et le cas des acides
phénoxyacétiques en constitue un contre-exemple (Soulas, 1990). De plus, parler
véritablement d’adaptation microbienne vis-à-vis d’un xénobiotique donné présuppose la
mise en place d’évènements génétiques affectant des régulations ou des activités
enzymatiques (Soulas, 1990). Ainsi, certains auteurs ont mis en évidence que l’expression des
gènes codant pour la catalyse de ces molécules est étroitement liée à la concentration du
pesticide (Alexander, 1985 ; Fomsgaard et Kristenssen, 1999 ; de Lipthay et al., 2007). Dans
le cas de sols n’ayant jamais été exposés à un xénobiotique ou dans des horizons profonds

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
43
(Fomsgaard et Kristenssen, 1999), il est nécessaire d’avoir une certaine concentration afin que
le xénobiotique soit accepté en tant que substrat pour microflore.
Il existe également des systèmes enzymatiques spécialisés traduisant l’existence d’adaptations
génétiques organisées en structures simples ou polycistroniques (opérons) et codant pour des
enzymes spécifiques au sein d’une voie de biodégradation responsable d’un métabolisme
direct (Dick et Quinn, 1995 ; Mandelbaum et al.,1995 ; Sorensen et al., 2001). La sélection de
telles adaptations est guidée par le résultat du bilan entre le coût de la synthèse d’enzymes
spécialisées et le gain énergétique procuré par la dégradation du xénobiotique, sans oublier
pour autant l’avantage sélectif procuré par ce réarrangement en réponse aux pressions
environnementales subies par les communautés microbiennes (Garcia-Gonzales et al., 2003 ;
Rhine et al., 2003 ; Sims, 2006). Citons dans ce cas l’exemple de la dégradation de l’acide
2,4-dichloro-phénoxyacétique ou 2,4-D (Streber et al., 1987), de l’atrazine (de Souza et al.,
1996), des urées substituées (Turnbull et al., 2001 ; El Sebaï et al., 2004 ; Hussain et al.,
2009) comme des illustrations de ce phénomène adaptatif.
Globalement, trois mécanismes fondamentaux peuvent décrire de façon satisfaisante
ce mécanisme adaptatif des populations microbiennes : (i) soit une mutation des gènes
régulateurs contrôlant la transcription des gènes structuraux pouvant amener à une production
continue d’enzymes correspondantes à un niveau élevé, (ii) soit une amplification des gènes
structuraux aboutissant à l’existence, dans le génome microbien, de plusieurs copies d’un
même gène ou d’un ensemble de gènes responsables, par effet additifs, à des concentrations
enzymatiques suffisantes pour que la transformation d’une molécule de xénobiotique
devienne possible, (iii) soit l’apparition de mutations ponctuelles des gènes structuraux
précédée d’une amplification du ou des gènes correspondants, à l’origine de la production de
protéines enzymatiques dotées de fonctions catalytiques nouvelles ou améliorées (Soulas,
1990). L’évolution indépendante des différentes copies du gène peut ainsi permettre le
maintien de la fonction métabolique ancienne et la mise en place puis la spécialisation
progressive de fonctions métaboliques nouvelles.
Dès lors qu’une fonction nouvelle est apparue, sa dispersion au sein d’une population
microbienne emprunte classiquement les voies de reproduction sexuée (appariement

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
44
chromosomique et recombinaison) et parasexuée (transformation, transduction généralisée et
conjugaison). Ainsi, ces mécanismes adaptatifs s’appuieraient sur l’existence d’éléments
génétiques mobiles présents au sein des communautés microbiennes. Ces structures
génétiques, représentées par les plasmides, les transposons, les intégrons, les ilots génomiques
ainsi que par les bactériophages, sont également collectivement reconnues comme des
vecteurs responsables du transfert génétique horizontal (HGT), jouant un rôle fondamental
dans la dégradation des pesticides. Ces groupes de gènes, à forte mobilité, sont définis comme
étant des éléments relativement flexibles pouvant s’intégrer avec plus ou moins de facilité
dans les génomes des bactéries hôtes (Funchain et al., 2001 ; Frost et al., 2005).
Trois mécanismes moléculaires principaux contrôlent ce transfert génétique horizontal
(HGT) dans le sol : la transformation, la transduction et le mécanisme de conjugaison. La
transformation est un processus physiologique d’absorption d’un fragment d’ADN extra-
cellulaire (plasmide ou partie du chromosome bactérien) à l’intérieur de cellules dites
compétentes, qui requiert ou non la présence de séquences spécifiques (séquences
d’absorption) dans le fragment d’ADN transféré pour qu’il soit retrouvé dans le cytoplasme
de la cellule hôte (Lorenz et Wackernagel, 1994 ; Trevors, 1996 ; Thomas et Nielsen, 2005).
Le mécanisme de transduction est un processus dans lequel un fragment d’ADN pris au
hasard dans le génome d’une bactérie et provenant soit d’une cellule hôte préalablement
infectée par un bactériophage (transduction généralisée), soit d’une séquence d’ADN d’un
hôte flanquée de deux séquences d’insertion ayant préalablement intégré de l’ADN de
bactériophage excisé (transduction spécialisée), se retrouve encapsidé pour constituer de
nouvelles particules phagiques (Ochman et al., 2000 ; Ochman, 2005). De nombreuses études
ont récemment suggéré que les fréquences de transduction dans le sol sont largement sous-
estimées et que de nombreuses populations de bactériophages se trouvent dans cette matrice.
(Ashelford et al., 2000). Enfin, la conjugaison bactérienne, processus de transfert génétique
qui semble être largement majoritaire dans le sol (Burrus et Waldor, 2004), est un mécanisme
contact-dépendant où le transfert d’un fragment d’ADN se fait d’une cellule donneuse vers
une cellule hôte pour la majorité des genres bactériens (Davison, 1999). Enfin, la présence de
séquences spécifiques codant pour le contrôle de leur transfert, lors des mécanismes de
conjugaison ou de transduction, confèrent aux éléments génétiques mobiles une fréquence de
transfert plus élevée et une stabilité supérieure dans les génomes des cellules hôtes,

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
45
comparativement aux fragments d’ADN reçus par le mécanisme de transformation (Burrus et
Waldor, 2004). La figure I-4-1 présente les principaux mécanismes moléculaires impliqués
dans le transfert génétique horizontal (HGT).
Figure I-4-1 : Principaux mécanismes moléculaires intervenant dans le transfert génétique horizontal (HGT), d’après Zaneveld et al. (2008) 3- Aspects cinétiques de la dégradation biologique : Métabolisme direct et co-métabolisme.
Les microorganismes responsables de la dégradation d’un pesticide sont généralement
regroupés en deux catégories : ceux qui disposent de l’intégralité des enzymes pour
transformer le substrat (acide 2,4,5-trichlorophénoxyacétique, par exemple) par les voies du
métabolisme central (Kilbane et al., 1983), et ceux dont l’équipement enzymatique est
incomplet pour mener à terme la dégradation de la molécule (Baker et Woods, 1977 ;
Horvath, 1972 ; Dalton et al., 1982 ; Anderson, 2002).

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
46
Dans le premier cas, on parle de métabolisme direct. Le composé organique est utilisé
par les microorganismes comme source de carbone et d’énergie à des fins de croissance. Le
stade ultime correspond à une minéralisation du composé organique (acide 2,4-
dichlorophénoxyacétique, atrazine, isoproturon, organophosphorés…) se traduisant
généralement par l’apparition de CO2, d’H2O et/ou de sels inorganiques. Dans la plupart des
cas, ce processus de dégradation n’entraîne pas l’accumulation de métabolites. La courbe de
dégradation est caractérisée par une période de latence suivie d’une phase exponentielle
d’accélération (Soulas, 1990 ; Radosevitch et al., 1995 ; El Sebaï, 2004 ; …). La vitesse de
minéralisation du produit phytosanitaire augmente avec la répétition de son application. La
cinétique de dégradation est sous la dépendance de différents paramètres : (i) les facteurs
climatiques, (ii) la taille initiale de la microflore dégradante, (iii) la dose de pesticide
appliquée, (iiii) la fréquence et le nombre des apports du produit phytosanitaire. Ainsi, ce
processus de dégradation va tendre vers la sélection et le développement progressif d’une
communauté microbienne tellurique dégradante susceptible d’entraîner une biodégradation
accélérée (BDA).
Dans le second cas, la transformation partielle de la substance active ne permet pas de
récupérer l’énergie nécessaire à la croissance des souches microbiennes. On parle alors de co-
métabolisme. La dégradation ultime de la molécule nécessite la présence d’un consortium
microbien composé par des populations spécialisées chacune dans une étape métabolique bien
spécifique (Daughton et Hsieh, 1977 ; Soulas, 1990). Cette complémentarité métabolique des
voies de dégradation entre les différents microorganismes du consortium pourrait s’expliquer
par l’utilisation en cascade des métabolites produits lors des étapes de dégradation successives
(Soulas, 1990 ; de Souza et al. 1998 ; Pelz et al., 1999 ; Smith et al., 2003a, 2003b).
Cependant, ces mécanismes restent bien souvent partiels et entraînent une accumulation de
produits de dégradation qui peuvent être à leur tour transformés ou stabilisés en résidus liés
(Horvath, 1972). Dans certains cas, les métabolites produits peuvent être plus toxiques et plus
mobiles que le produit phytosanitaire dont ils sont issus (Somasundaram et Coats, 1990). Les
microorganismes participant à ce type de dégradation biologique ont nécessairement besoin
d’oxyder d’autres sources de carbone pour se développer. La matière organique, présente sous
différentes formes dans les sols, est souvent utilisée comme co-substrat (Houot et al., 1998).

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
47
De plus, les microorganismes dotés d’enzymes à large spectre d’activité (champignons par
exemple) sont souvent impliqués dans ce type de processus métabolique.
Différentes hypothèses peuvent être avancées pour expliquer la dégradation dite co-
métabolique : (i) soit les micro-organismes ne possèdent pas toutes les enzymes nécessaires à
l’ensemble des étapes métaboliques, (ii) soit les enzymes présentes sont peu efficaces vis-à-
vis du substrat, (iii) soit un métabolite présente une toxicité particulière et son accumulation
nuit au fonctionnement métabolique et éventuellement à la viabilité de la cellule microbienne
(Janke et Fritsche, 1985). Dans ce cas, la courbe de dégradation s’apparente à une
transformation chimique catalysée sans point d’inflexion. Ainsi, l’absence de croissance des
populations responsables de la dégradation, jointe à leur faible taille, fait du co-métabolisme
une voie lente de dégradation dont la vitesse n’augmente pas au cours du temps (Soulas,
1990). Il s’agit là d’une caractéristique cinétique essentielle des transformations co-
métaboliques. La plupart des produits phytosanitaires sont dégradés par voie ce type de voie
co-métabolique.
4- Cas particulier de dégradation biologique : la biodégradation accélérée (BDA) des pesticides 4-1- Contexte et définition
La biodégradation accélérée (BDA) des pesticides correspond à la diminution de la
persistance et, quelques fois, de l’efficacité agronomique d’un produit phytosanitaire,
consécutivement à des applications répétées dans une parcelle agricole (Suett, 1987 ; Suett et
Jukes, 1988 ; Suett et al., 1993 ; Arbeli et al., 2007). D’après Rouchaud et al., (1996, 1997 et
2000) ce phénomène est une des conséquences majeures de la structure et de l’évolution des
populations microbiennes impliquées dans la dégradation des pesticides. Le produit
phytosanitaire est dans ce cas utilisé comme source de nutriments et/ou d’énergie par un
nombre restreint de microorganismes adaptés conduisant à la sélection progressive de
populations microbiennes telluriques. Audus (1949) fut le premier auteur à parfaitement
décrire ce phénomène en travaillant sur la dégradation du 2,4-D. En effet, il mit en évidence
la corrélation existant entre les effets de la répétition des traitements et la vitesse de
dégradation de ce pesticide. Ces travaux ont été confirmés par de nombreuses études sur

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
48
d’autres familles phytosanitaires : carbamates (Felsot et al., 1981 ; Racke et Coats, 1988a),
organophosphorés (Racke et Coats, 1988b ; Chung et Ou, 1996), s-triazines (Barriuso et
Houot, 1996 ; Puissemier et al., 1997 ; Radosevich et al. ; 1995 ; Mandelbaum et al., 1995 ;
Yassir et al., 1999 ; Topp et al., 2000) et urées substituées notamment (Walker et Welch,
1991 ; Cox et al., 1996 ; Sorensen et al., 2001 ; El Sebaï et al., 2004 ; Sorensen et Aamand,
2003 ; Sorensen et al., 2003). Ces données ont suggéré la nécessité de prendre en compte le
risque potentiel d’adaptation des sols face aux traitements répétés de pesticides. En effet, d’un
point de vue agronomique, la BDA est un facteur limitant l’efficacité des produits
phytosanitaires puisqu’ils sont rapidement dégradés par la microflore tellurique. En revanche,
la mise en place d’une BDA au sein d’un parcelle agricole impacte positivement
l’environnement, puisque ce phénomène adaptatif peut limiter la persistance et le transfert des
produits phytosanitaires du sol vers les autres compartiments.
4-2- Mécanismes responsables de la biodégradation accélérée
L’origine du mécanisme est liée à l’activité de la biomasse microbienne du sol
(Chapman et Harris, 1990). Le schéma général est toujours identique : la première application
d’un produit phytosanitaire dans une parcelle agricole se traduit par une dégradation de la
substance active après une période plus ou moins longue selon la nature du produit, appelée
période de latence. Cette première phase correspond à l’induction enzymatique pouvant
s’expliquer par une adaptation physiologique et/ou génétique des populations microbiennes
concernées. Au cours d’une deuxième phase, la dégradation de la substance active se traduit
très souvent par une décroissance exponentielle de la concentration résiduelle. Cette phase
n’est réellement mesurable qu’à la seule condition d’avoir une densité cellulaire suffisante. Si
l’application du même produit est successivement répétée un grand nombre de fois, la
présence d’une très forte densité de souches microbiennes adaptées entraine une diminution,
voire une suppression de la phase de latence. Dans ce cas, au sein de ces sols dits « adaptés »,
80 % du produit phytosanitaire appliqué peut ainsi être rapidement biodégradé en moins de 15
jours (Devers, 2008). Ce potentiel dégradant d’un sol adapté à la substance active peut se
maintenir sur des périodes allant de quelques semaines à plusieurs années après l’arrêt total
des traitements (Roeth, 1986 ; Walker et Welch, 1991 ; Parekh et al., 1992 ; Anderson et

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
49
Lafuerza, 1992 ; Suett et al., 1993 ; Smelt et al., 1996 ; Morel-Chevillet, 1998). La disparition
de cette capacité catabolique peut résulter de la perte du caractère génétique en l’absence de
pression de sélection suffisante au sein des communautés microbiennes telluriques.
Certains auteurs (Fournier et al., 1998) précisent que le développement d’une BDA
puis sa propagation dans la parcelle agricole est généralement graduel dans le temps et dans
l’espace, et nécessite la mise en oeuvre de différents mécanismes de dispersion (travaux
agricoles, ruissellement, phénomènes météorologiques).
Le transfert génétique horizontal (HGT) présent au sein des communautés microbiennes (cf
paragraphe I-4-2) joue un rôle fondamental dans le développement du phénomène de BDA.
Cet échange de matériel génétique peut en effet s’opérer : (i) via la conjugaison bactérienne
par échange de plasmides conjugatifs (molécules d’ADN circulaires, double brins, extra-
chromosomiques, autoréplicables et transférables dans certains cas), (ii) ; via la transposition
par l’intermédiaire d’éléments transposables (petites unités d’ADN mobiles intégrées au
chromosome bactérien pouvant s’exciser et se repositionner ailleurs dans le génome) ; (iii) via
la transformation bactérienne d’ADN extra-chromosomique. Dans le cas des BDA, les
transferts génétiques sont pour la plupart pilotés par les deux premiers mécanismes.
Le modèle conceptuel de l’adaptation des communautés bactériennes à la biodégradation
accélérée (BDA) de l’atrazine proposé par Devers (2008) et Devers et al. (2004, 2005, 2007a,
2007b et 2008) est une illustration de ces mécanismes génétiques adaptatifs (cf figure I-4-2).
Ces auteurs ont pu montrer la grande plasticité de ces phénomènes, pouvant s’appuyer
partiellement sur un mécanisme déjà cité : la conjugaison plasmidique comme acteur principal
de la dispersion des gènes atz et trz. Il faut associer à ce processus l’intervention probable des
transposons via leurs séquences d’insertion IS 1071 qui jouent un rôle important dans la
diversification du support (la localisation génomique des gènes atz et tr peut être
chromosomique et/ou plasmidique) et dans leur duplication par recombinaison homologue.
Ces transferts et réarrangements génétiques concourent, in fine, (i) à augmenter le potentiel de
dispersion des gènes cataboliques de l’atrazine par augmentation du spectre d’hôte au sein des
génomes bactériens, (ii) à optimiser la voie de dégradation du pesticide par duplication en
tandem de certains gènes codant pour des enzymes clefs des voies cataboliques.

Première partie – Chapitre 4 : Dégradation biologique des produits phytosanitaires dans les sols
50
Figure I-4-2 : Modélisation conceptuelle de l’adaptation des communautés bactériennes à la biodégradation accélérée (BDA) de l’atrazine. Dispersion (flèches rouges) des gènes atz et trz par conjugaison plasmidique et diversification (flèches vertes) du support des gènes atz par transposition, d’après Devers (2008).
Dans certains cas, le processus de BDA ne se limite pas à un seul pesticide en particulier ; au
contraire, c’est une mécanisme doué d’une importante plasticité puisqu’il peut concerner des
produits phytosanitaires présentant des structures chimiques apparentées (Audus, 1951 ;
Soulas, 1985 ; Roeth et al., 1989 ; Singh et al., 2005) mais également des substances actives
appartenant à des familles totalement différentes (Morel-Chevillet et al., 1996 ; Morel-
Chevillet, 1998 ; Warton et al., 2003). De plus, la BDA est un mécanisme microbien adaptatif
qui n’est pas seulement circonscrit aux produits phytosanitaires, mais, semble être généralisé
à l’ensemble des molécules organiques polluantes massivement répandues dans
l’environnement (Erickson et Mondello, 1993 ; Field et al., 1995).

51
Deuxième partie: Présentation générale de
l’Étude

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
52
Chapitre1‐Lafamilledesherbicidestricétones
1- Contexte historique
La présence de composés β-tricétoniques d’origine naturelle a été décrite par Hellyer
(1968) dans des huiles volatiles de certaines plantes australiennes appartenant à la famille des
Myrtacées. Une activité herbicide naturelle a été ensuite envisagée par des chercheurs de la
firme agro-pharmaceutique Zeneca Ag Products, consécutivement à l’observation de
l’absence de certaines espèces d’adventices autour d’arbustes connus sous le nom de Rince-
bouteilles ou Bottle-brush (Callistemon citrinus stapf). Différentes analyses ont permis
d’attribuer l’activité herbicide à la présence d’un composé tétracétonique, la leptospermone,
substance allélopathique (figure II-1-1) efficace sur de nombreuses adventices mais non
phytotoxique sur le maïs.
Figure II-1-1 : Formule développée de la leptospermone (C15H22O4).
Ces résultats ont été à la base du développement des herbicides de la famille des tricétones.
Michaely et Kratz (1986) ont synthétisé des dérivés substitués de la 2-benzoyl-1,3-
cyclohexanedione manifestant une activité herbicide plus intéressante. Les relations structure-
activité ont été décrites par Lee et al. (1998) et ont mis en évidence le caractère obligatoire de
la présence d’un substituant en position ortho sur le noyau aromatique pour obtenir une action
herbicide. Par ailleurs, cette activité est renforcée par la présence de substituants électro-

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
53
attracteurs en position 2,4 (cf figure II-1-2 et tableau II-1-1). A partir de ces études, deux
molécules ont ainsi connu un développement commercial, notamment en maïsiculture : la
sulcotrione et la mésotrione.
Figure II-1-2 : Structure de la 2-[2,4-benzoyl disubstituée]-1,3 cyclohexanedione.
Tableau II-1-1 : Activité herbicide (DC50 : Dose moyenne nécessaire pour le contrôle de 50 % de 7 espèces différentes de dicotylédones) des composés 2-[2,4-benzoyl disubstitués]-1,3 cyclohexanedione (d’après Lee et al.,1998).
Composé Substituant A
position ortho
Substituant Q
position para
DC50 (g.ha-1) Log DC50
3 Cl OCH3 5100 3,71
4 Cl CH3 2800 3,45
5 Cl H 830 2,92
6 Cl F 1600 3,2
7 Cl Cl 180 2,26
9 Cl SO2CH3 72 1,86
10 NO2 H 810 2,91
11 NO2 Cl 42 1,62
12 NO2 CF3 14 1,16
13 NO2 SO2CH3 13 1,12

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
54
2- La substance active 2-1- Fiche d’identité Nom commun : Sulcotrione
CAS-n° : 99105-77-8
Famille chimique : Benzoyl-cyclohexane-1,3-diones ou tricétones.
Formule brute : C14H13ClO5S
Formule développée :
Noms chimiques :
IUPAC : 2-(2-chloro-4-mésylbenzoyl)cyclohexane-1,3-dione
CAS : 1,3-cyclohexanedione, 2-[2-chloro(4-méthylsulfonyl) benzoyl]
Nom de code (R&D) : ICIA0051 ; SC-0051 (The Pesticide Manual, 2006)
Activité biologique : herbicide systémique, absorbé préférentiellement par les feuilles ;
contrôle des adventices dicotylédones et certaines graminées annuelles ; homologué en France
sur maïs, lin, ray-grass, sorgho, maïs doux (Index phytosanitaire, ACTA, 2011). Utilisation
décrite également sur canne à sucre (Pesticide Manual, 2006)
2-2- Propriétés physico-chimiques Poids moléculaire : 328,7 g.mol-1

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
55
Point de fusion : 139 ° C
pKa : 2,8 ± 0,2 (acide faible)
Etat physique : solide blanc à brun, amorphe.
Tension de vapeur : 5,4 10-6 Pa = 4,18 10-8 mm Hg (molécule peu volatile)
Masse volumique : 1,15 g.mL-1 à 20°C.
Solubilité dans l’eau : 165 mg.L-1 à 25°C (solubilité « intermédiaire »).
Solubilité dans les solvants organiques :
2-3- Les formes tautomériques de la sulcotrione
La tautomérisation céto-énolique des composés de la famille des benzoyl-
cyclohexane-1,3-dione a été étudiée par divers auteurs (Rouchaud et al., 1996 ; Lin et al.,
2000 ; Huang et al., 2002 ; Wu et al., 2002). Un calcul théorique a permis de déterminer
l’existence de trente deux tautomères. Néanmoins, deux conformations sont considérées
comme dominantes en solution aqueuse (Huang et al., 2002) : il s’agit de la forme
exocyclique et de la forme endocyclique, ce dernier le tautomère apparaissant comme le plus
stable (cf figure II-1-3).
Figure II-1-3 : Formes tautomériques dominantes de la benzoyl-cyclohexane-1,3-dione en solution aqueuse (X = Cl pour la sulcotrione, X = NO2 pour la mésotrione).
Solvant Solubilité (g.L-1)
Acétone 40
Chlorobenzène 10
Ethanol < 6
Xylène < 6

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
56
En outre, le caractère acide faible de la sulcotrione (et de la mésotrione), conditionnant la
forme moléculaire ou dissociée suivant le pH du milieu, est présenté dans la figure II-1-4
(Dyson et al., 2002).
Figure II-1-4 : Comportement « acide faible » en solution aqueuse d’une benzoylcyclohexane-1,3-dione.
3- Mode d’action de la substance active La sulcotrione est un herbicide sélectif du maïs dont la sélectivité est fondée sur la
métabolisation par la plante cultivée (Pallet et al., 2001). Cet herbicide est généralement
utilisé en post-levée des adventices au stade 4-6 feuilles, mais peut être aussi utilisé en pré-
émergence et en pré-semis. La substance active peut être absorbée par la voie foliaire (Wilson
et Foy, 1992 ; Reddy et Bhowmik, 1991 ; Lee et al., 1998 ; Townson et al., 1999) mais
également par la voie racinaire puisqu’elle a montré une certaine activité résiduelle dans le sol
(Compagnon et Beraud, 1992 ; Cools et al., 1997). Cet herbicide est transporté par le phloème
dans le cas d’une application foliaire et par le xylème dans le cas d’une application sur sol nu
(Index Phytosanitaire, ACTA, 2011).
Chez les plantes sensibles, les symptômes externes se manifestent surtout par le blanchiment
des feuilles (Mayonado et al., 1989 ; Reddy et Bhowmik, 1991 ; Schultz et al., 1993 ; Lee et
al., 1998 ; Index Phytosanitaire, ACTA, 2011) qui s’accompagne rapidement d’une nécrose
puis de la mort des plantes. Néanmoins, d’autres zones autres que le système foliaire vont
également présenter signes de perturbations physiologiques importantes. Par exemple, lorsque

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
57
la sulcotrione est appliquée en pré-levée, l’herbicide agit sur les graines traitées. Ces dernières
peuvent germer mais les plantules blanchies ne tardent pas à mourir (Townson et al., 1999).
Lee et al. (1998) ont pu mettre en évidence que les tissus méristématiques les plus récents
présentent des symptômes de chlorose plus rapidement que les tissus plus âgés de l’adventice.
Mayonado et al. (1989) ont remarqué que l’application de sulcotrione entraine généralement
une diminution de la concentration en caroténoïdes et favorise l’accumulation de pigments
incolores ou phytoènes, suite à la déplétion du taux de plastoquinones. Pour Tsegaye et al.
(2002) puis Falk et al. (2003), une telle diminution accompagnée de la baisse des alpha-
tocophérols entraine nécessairement une perturbation majeure de l’appareil photosynthétique
chez la plante. Chez le maïs, l’enzyme 4-hydroxyphénylpyruvate dioxygénase a été isolée,
extraite et purifiée par Barta et Böger (1996). C’est une enzyme Fe2+-dépendante, de nature
non héminique, qui convertit le 4-hydroxyphénylpyruvate en homogentisate, précurseur clé
dans la synthèse des tocophérols et des quinones. La protéine enzymatique catalyse une
décarboxylation, une substitution et une oxydation du cycle du composé aromatique. Cette
enzyme appartient à la famille des oxygénases acides alpha-cétoniques dépendantes
(Brownlee et al., 2004 ; Moran, 2005). L’étude a notamment montré que l'enzyme avait un
pH optimum de 7,3, une masse moléculaire apparente de 43 kDa et était semblable à celle de
l'enzyme correspondante trouvée dans le foie des mammifères. Les cinétiques réalisées lors de
ces travaux ont démontré que les herbicides de la famille des tricétones étaient de puissants
inhibiteurs compétitifs de l’enzyme 4-p HPPD, notamment pour des valeurs de Ki voisines de
5 nM pour la 2-(2-nitro-4-chlorobenzoyl)-5-(2-methoxyethyl) cyclohexane-1,3-dione and et
voisine de 15 nM pour la 2-(2-chloro-4-methanesulfonylbenzoyl)-cyclohexane-1,3-dione. Les
études qui ont suivi sont venues confirmer les premiers résultats des travaux de Barta et Böger
(Secor, 1994 ; Lee et al., 1998 ; Townson et al., 1999 ; Tsegaye et al., 2002 ; Matringe et al.,
2005, Dayan et al., 2007 ; Dayan et al., 2009 ; Beaudegnies et al., 2009). Les molécules de la
famille des tricétones interagissent en tant qu’inhibiteur compétitif au niveau du site
catalytique de l’enzyme par présence du groupement 1,3-cyclohexane-dione, « analogue » de
la partie alpha-cétonique du substrat de l’enzyme 4-phénylhydroxypyruvate (cf figure II-1-5).
La synthèse de l’homogentisate étant bloquée, les plastoquinones absentes n’assurent plus
leur rôle de co-facteur de l’enzyme phytoène désaturase, la synthèse des caroténoïdes est donc
stoppée (Kim et al., 2002 ; Artetxe et al., 2006). La sulcotrione classée F2 selon la liste des
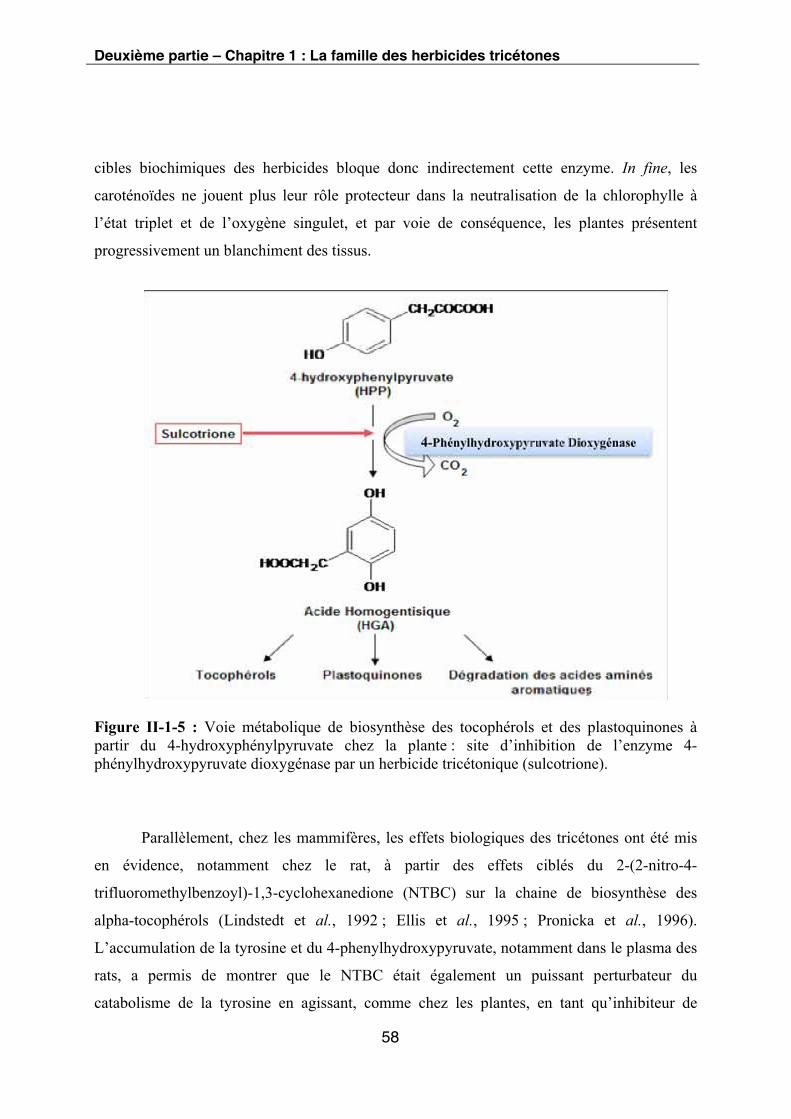
Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
58
cibles biochimiques des herbicides bloque donc indirectement cette enzyme. In fine, les
caroténoïdes ne jouent plus leur rôle protecteur dans la neutralisation de la chlorophylle à
l’état triplet et de l’oxygène singulet, et par voie de conséquence, les plantes présentent
progressivement un blanchiment des tissus.
Figure II-1-5 : Voie métabolique de biosynthèse des tocophérols et des plastoquinones à partir du 4-hydroxyphénylpyruvate chez la plante : site d’inhibition de l’enzyme 4-phénylhydroxypyruvate dioxygénase par un herbicide tricétonique (sulcotrione).
Parallèlement, chez les mammifères, les effets biologiques des tricétones ont été mis
en évidence, notamment chez le rat, à partir des effets ciblés du 2-(2-nitro-4-
trifluoromethylbenzoyl)-1,3-cyclohexanedione (NTBC) sur la chaine de biosynthèse des
alpha-tocophérols (Lindstedt et al., 1992 ; Ellis et al., 1995 ; Pronicka et al., 1996).
L’accumulation de la tyrosine et du 4-phenylhydroxypyruvate, notamment dans le plasma des
rats, a permis de montrer que le NTBC était également un puissant perturbateur du
catabolisme de la tyrosine en agissant, comme chez les plantes, en tant qu’inhibiteur de

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
59
l’enzyme 4-hydroxyphénylpyruvate dioxygénase (EC 1.13.11.27). Les travaux de Wu et al.
(2011) ont également confirmé le rôle d’inhibiteur compétitif de la sulcotrione vis-à-vis de
l’enzyme 4-pHPPD et ont mis en évidence certains troubles physiologiques associés à une
ingestion chronique de sulcotrione chez les rats.
La sulcotrione est utilisée en maïsiculture depuis l’interdiction de l’atrazine en 2003.
Elle est autorisée à la vente en France depuis février 1993 et commercialisée notamment sous
le nom de Mikado® dès 1994 par Syngenta puis, depuis 2001 par Bayer CropScience. Ce
produit est une suspension concentrée à 300 g.L-1 de sulcotrione.
Les doses d’application recommandées sont de l’ordre de 300 à 450 g.Ha-1 (Index
phytosanitaire, ACTA 2011 ; Mamy et Barriuso, 2005). La sulcotrione est utilisée pour le
désherbage en France, seule ou en association avec d’autres herbicides (bromoxynil,
isoxaflutole, acétochlore, mésotrione ou nicosulfuron) afin d’augmenter l’efficacité du
traitement tout en diminuant les doses d’application (Chaabane, 2005). Elle présente un large
spectre d’action et couvre donc une très large gamme d’adventices. L’action de la sulcotrione
sur certaines adventices monocotylédones et dicotylédones annuelles (famille des Poacées)
fréquemment rencontrées en maïsiculture (cf tableau II-1-2) a été étudiée.
Degré de sensibilité Dicotylédones Monocotylédones (famille des Poacées)
TS à S
Chenopodium album
Solanum nigrum
Polygonum persicaria
Stellaria media
Digitaria sanguinalis
Echinochloa crus galli
Panicum miliaceum
MS à MR
Amaranthus sp.
Mercurialis annua
Panicum dichotomiflorum
R
Calystegia sepium
Equisetum sp.
Setaria sp.
Cynodon dactylon
Tableau II-1-2 : Sensibilité des adventices à la sulcotrione pour un traitement en post-levée à 450 g.Ha-1. TS = très sensible, S = sensible, MS = moyennement sensible, MR = moyennement résistante, R = résistante (d’après Compagnon et Beraud, 1992).

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
60
4- Approches toxicologique et écotoxicologique 4-1- Vis-à-vis des mammifères
La sulcotrione est rapidement éliminée par voie urinaire, le métabolite majeur étant la
4-hydroxysulcotrione. La DL50 orale chez le rat est estimée supérieure à 5000 mg.kg-1. Peu
absorbé à travers la peau et peu irritant au niveau des yeux chez le lapin. Les cochons d’inde
ont cependant montré une forte sensibilité au niveau de l’épiderme. Chez le rat, la CL50 par
inhalation est > 1,6 mg.L-1.
La dose sans effet chez le rat a été estimée à 0,5 mg.kg-1 P.C par jour. La DJA a été évaluée à
0,005 mg.kg-1 PC.jour-1. Aucun effet tératogène n’a été noté chez les rats et les lapins. In vivo,
la sulcotrione n’a pas montré de propriétés génotoxiques.
4-2- Ecotoxicologie
Les données relatives à l’écotoxicologie de la sulcotrione sont reportées dans le
tableau II-1-3.
Espèce Test DL50 ou CL50
Oiseaux Toxicité aigüe par voie orale > 1350 mg.kg-1
Poissons (truite-arc-en-ciel) Toxicité aigüe par voie dermale > 4000 mg.kg-1
Abeilles Toxicité aigüe par contact
Toxicité aigüe par voie orale
> 200 µg/abeille
> 50 µg/abeille
Daphnie Toxicité aigüe après 48 heures
d’exposition
> 100 mg.L-1
Algues et plantes aquatiques Test de croissance CEBb50 = 3,5 mg.L-1
Tableau II-1-3 : Toxicité et écotoxicité de la sulcotrione non formulée (Pesticide Manual, 2006).
Des données plus exhaustives relatives à l’impact de la sulcotrione sur les organismes vivants,
extraites de la base de données Agritox, sont présentées dans l’annexe I
(http://www.dive.afssa.fr/agritox/php/sa.php?sa=1101).

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
61
5- Comportement dans les différents compartiments de l’environnement 5-1- Dans la matrice air
Très peu de données sont disponibles en ce qui concerne le comportement de la
substance active dans la matrice air. Néanmoins, la valeur de la pression de vapeur de la
sulcotrione (5,4 µPa à 25 °C) classe cet herbicide dans la famille des composés très peu
volatils.
5-2- Dans la matrice eau
L’hydrolyse de la sulcotrione a été largement décrite par Chaabane (2005). Les
données bibliographiques concernant cet herbicide faisaient état de deux voies de dégradation
hydrolytique (voies A et B) qu’elle a pu effectivement mettre en évidence. L’influence de
plusieurs paramètres a été étudiée (pH, température, durée, présence de Fe++,…). Le rôle du
pH de la solution aqueuse s’est avéré déterminant.
Un test préliminaire a été mené, selon les recommandations de la directive de l’OECD (2002),
à 50° C et sous conditions de pH fortement acide ou alcalin. Malgré la stabilité de la
substance active dans ces conditions relativement extrêmes, Chaabane a proposé un schéma
réactionnel d’hydrolyse contrôlé par le pH. A pH alcalin, la voie A (considérée comme
classique) a été prédominante, contrairement aux pH acides où la voie B a été décrite.
A pH neutre ou alcalin, la voie A, largement décrite dans la littérature (Rouchaud et al., 1996,
1998a, 1998b) est caractérisée par l’apparition de deux produits de dégradation principaux,
l’acide 2-chloro-4-méthylsulfonylbenzoïque (CMBA) ne présentant pas d’activité herbicide
(Rouchaud et al.,1998a ; Cherrier et al., 2004) et la 1,3-cyclohexanedione (CHD). Pour des
valeurs de pH < 7, la détection intermédiaire du dérivé d’acide heptanoïque indique
l’intervention d’une deuxième voie (voie B), au préalable décrite uniquement dans certains
sols (Rouchaud et al., 1996), avec cependant, pour résultat final, la présence du même
CMBA.
La figure II-1-6 décrit ces deux voies de dégradation.

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
62
Figure II-1-6 : Formules développées des principaux produits de dégradation de la sulcotrione, issus des voies A et B décrites dans la littérature. En milieu aqueux à 25°C, la sulcotrione s’est montrée chimiquement très stable. Le temps de
demi-vie a été estimé entre 200 et 400 jours selon le pH (Chaabane, 2005).
La substance active semble se dégrader plus rapidement en présence du cation Fe++, dont le
rôle de catalyseur métallique doit être confirmé.

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
63
La photolyse a également été étudiée au préalable au laboratoire (Chaabane, 2005 ;
Chaabane et al., 2007). La vitesse de cette photolyse a été nettement supérieure à celle de
l’hydrolyse, avec formation de photoproduits majoritairement comparables, laissant supposer
une hydrolyse photoassistée de la sulcotrione. Un schéma réactionnel général a été proposé (cf
figure II-1-7). La photolyse des photoproduits des herbicides tricétoniques (sulcotrione et
mésotrione) fait actuellement l’objet d’une nouvelle étude au LCBE.
Figure II-1-7 : Principaux photoproduits de la sulcotrione (Chaabane et al., 2007). (A) : (2-hydroxy-4-méthylsulfonyl)-1,3-cyclohexanedione, (B) : Acide 5,7-dicéto-7-(2-chloro-4-méthylsulfonylphényl) heptanoïque, (C) : Acide 5,7-dicéto-(2-hydroxy-4-méthylsulfonyl) heptanoïque, (D) : 1,3-cyclohexanedione, (E) : Acide 2-chloro-4-méthylsulfonylbenzoïque, (F) : Acide 2-hydroxy-4-méthylsulfonyl benzoïque.

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
64
5-3- Dans la matrice sol 5-3-1- Rétention et transfert de la sulcotrione et de ses produits de dégradation
Le processus de rétention a été largement décrit par Chaabane (2005) et Chaabane et
al. (2008), selon les directives de l’OCDE (2002) préconisant la méthode « Batch
equilibrium ». Les résultats expérimentaux ont montré une bonne concordance avec le modèle
mathématique de Freundlich, et notamment, une diminution des sites d’adsorption avec
l’augmentation de la concentration de l’herbicide dans la phase liquide.
La sulcotrione et son métabolite majeur (CMBA) montrent des capacités d’adsorption
modérées ainsi qu’une désorption relativement aisée à partir des sols étudiés lors de ces
travaux, choisis pour leur représentativité « culturale » (Perpignan, Landes, Belgique,
Martinique). La teneur en matière organique, le pH et la teneur en argiles du sol jouent un rôle
prépondérant dans ce cas. Ainsi, la persistance de la sulcotrione augmente avec la diminution
du pH mais aussi avec le contenu en matière organique (Rouchaud et al., 1998a, 1998b).
D’autres produits de dégradation (CHD, acide phénylheptanoïque) sont apparus plus
fortement adsorbés.
Le transfert de la sulcotrione (en plein champ et sur colonne de sol reconstitué) est
modéré dans le profil de sol 0-10 cm et fortement influencé par sa rétention et sa dégradation
dans le sol ainsi que par les conditions climatiques avant et après traitement.
5-3-2- Dégradation de la sulcotrione
Les travaux de Chaabane (2005) ont montré la prédominance de la voie de dégradation
biologique dans les sols étudiés, avec une cinétique de dégradation pouvant être généralement
décrite par une loi de premier ordre. Les temps de demi-vie observés en conditions de plein
champ sont comprises entre 16 et 122 jours (Rouchaud et al., 1996, 1998a et 1998b). Au
laboratoire, elles sont très variables, de 1 à 60 jours (Baer et Calvet, 1999 ; Cherrier et al.,
2004 ; Chaabane, 2005 ; Mamy et al., 2005 ; Chaabane et al., 2007 ; Doublet et al., 2009) et
sont sous l’influence de plusieurs paramètres :

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
65
La nature du sol : en accord avec les travaux de Cherrier et al. (2004), le temps de demi-vie
de la sulcotrione diminue avec l’augmentation de la teneur en matière organique du sol. Par
contre, la vitesse de dégradation semble peu influencée par la texture du sol (Rouchaud et al.,
1996).
Le taux d’humidité relative : la vitesse de dégradation augmente proportionnellement à la
teneur en eau (dans la limite 25-70 %), quel que soit le type de sol étudié. Cette activation a
été également décrite par Baer et Calvet (1999).
La température : Les expérimentations ont été menées par Chaabane (2005) à diverses
températures (25, 40 et 60° C). La figure II-1-8 présente les résultats obtenus sur le sol de
Belgique ; la non-linéarité de la courbe montre que le mécanisme de dégradation de la
sulcotrione dans le sol varie en fonction de la température. Ces travaux ont permis deux
hypothèses : i) la sulcotrione se dégrade majoritairement par voie biologique, l’activité
microbienne diminuant au-delà de 30 °C (1) ; ii) à température plus élevée (50-60°C), la
dégradation est contrôlée par la présence de bactéries themophiles (2).
Figure II-1-8 : Variation de Ln Kobs en fonction de 1/T pour la dégradation de la sulcotrione dans le sol de Belgique dans la gamme de températures 25, 40 et 60°C.

Deuxième partie – Chapitre 1 : La famille des herbicides tricétones
66
La stérilisation du sol : la dégradation est apparue très faible après stérilisation du sol à
l’autoclave (cet autoclavage pouvant cependant modifier sensiblement la modification de la
structure du sol, Cantier et al., 1986 ; Barrère et al., 1988).
La substance active sous forme de standard analytique ou formulée (Mikado®) : La
formulation joue un rôle au niveau de la vitesse de dégradation, nettement plus rapide avec le
produit formulé.
Ces résultats, obtenus au LCBE à l’issue d’un premier travail de doctorat et mettant en
exergue la prédominance de la voie de dégradation biologique, nous ont conduit à émettre
l’hypothèse d’une éventuelle biodégradation accélérée (consécutive à des traitements répétés)
et à mettre en évidence le rôle majeur d’une flore microbienne dégradante spécifique, à
l’origine de cette métabolisation. Dans ce but, le sol de la parcelle de l’IUT de Perpignan,
précédemment utilisé dans le cadre des travaux de Chaabane, a fait l’objet d’une nouvelle
campagne d’échantillonnage (cf paragraphe II-2-4 et II-2-5)
.

Deuxième partie – Chapitre 2 : Matrice dʼétude : le sol de Perpignan
67
Chapitre2‐Matriced’étude:lesoldePerpignan
1- Situation géographique de la parcelle La parcelle choisie pour cette étude a été mise à notre disposition par l’Institut
Universitaire de Technologie (IUT) de Perpignan, département Génie Biologique, Université
de Perpignan Via Domitia (UPVD). Ses coordonnées géographiques sexagésimales ont été
déterminées : 42° 40' 55" N, 2° 53' 49" E. Cette parcelle agricole (figure II-2-1), présumée
homogène, d’une surface globale voisine de 1500 m², a été préalablement subdivisée en cinq
zones convenablement matérialisées, dont deux (S1) et (S2), d’une surface équivalente (200
m² environ) ont été et exclusivement réservées à ce travail sur la sulcotrione. Des zones
« tampons » ont été délimitées, afin d’isoler chaque unité de traitement et de limiter, dans la
mesure du possible, les phénomènes de contamination (dérive, ruissellement, …). Une zone
contrôle (C) a également été réservée (environ 300 m²).
Figure II-2-1 : Plan de la parcelle expérimentale (IUT de Perpignan, UPVD).

Deuxième partie – Chapitre 2 : Matrice dʼétude : le sol de Perpignan
68
2- Paramètres édaphiques du sol de Perpignan
Les analyses pédologiques et physico-chimiques du sol ont été effectuées par le
Laboratoire d’Analyses Agricoles de Perpignan, selon des protocoles standardisés; l’humidité
relative des échantillons a été déterminée au Laboratoire de Chimie des Biomolécules et de
l’Environnement (LCBE-UPVD), suivant la méthode publiée par Mathieu et Pieltain (2003),
basée sur le calcul de la perte de poids après séchage de l’échantillon à l’étuve à 100°C
pendant 24 h. Les résultats sont présentés dans le tableau II-2-1.
Figure II-2-2 : Parcelle expérimentale (avant traitement phytosanitaire) située sur le site de l’IUT de Perpignan, Université de Perpignan Via Domitia (Pyrénées Orientales).

Deuxième partie – Chapitre 2 : Matrice dʼétude : le sol de Perpignan
69
Sable
(%)
Limon
(%)
Argile
(%)
Humidité
Relative (%)
Carbone
Orga. (%)
Azote
Orga. (g kg-1)
Matière
Orga. (%)
C/N CEC
(meq 100 g-1)
pH
25.6 60.5 13.9 24 0,9 0,98 1,55 9,64 15,5 8,1
Tableau II-2-1 : Propriétés pédologiques et physico-chimiques du sol de la parcelle de l’Institut Universitaire de Technologie (IUT) de Perpignan, déterminées par les protocoles standardisés (Laboratoire d’Analyses Agricoles de Perpignan et LCBE –UPVD).
Selon la classification FAO, le sol de l’IUT est rangé parmi les sols limono-sablo-argileux
dans le triangle des textures.
3- Historique cultural
La parcelle, mise à notre disposition à l’IUT, avait servi de système cultural en
agriculture biologique avant d’être laissée en friche agricole jusqu’en 2005. A compter de
cette date, les zones (S1) et (S2) ont reçu 4 traitements successifs à la dose agronomique
recommandée de 300 g de sulcotrione.Ha-1 par pulvérisation manuelle d’une préparation à
base de Mikado®, suspension concentrée titrant 300 g.L-1 de substance active
(BayerCropScience).
Ces traitements phytosanitaires ont été effectués selon le calendrier suivant : traitement 1 :
21 avril 2006 ; traitement 2 : 22 mai 2006 : traitement 3 : 22 juin 2006 ; traitement 4 : 21
juillet 2006.
Parallèlement, dans le contexte de notre première étude sur ces herbicides tricétones réalisée
dans le cadre d’un stage de Master (O. Moutugou Mouanda), les zones (M1) et (M2) ont été
traitées, de manière comparable, à l’aide d’une préparation à base de Callisto®, formulation
renfermant 100g.L-1 de mésotrione, substance active voisine de la sulcotrione ne différant que
par la nature du substituant en position 2 sur le noyau benzénique (substituant –NO2 au lieu de
–Cl). Par la suite, aucun autre traitement phytosanitaire n’a été appliqué sur la parcelle.
4- Stratégie d’échantillonnage
Les zones de prélèvement (S1) et (S2), présumées homogènes, ont été divisées de
manière aléatoire, en unités d’échantillonnage, chaque unité faisant l’objet d’un prélèvement

Deuxième partie – Chapitre 2 : Matrice dʼétude : le sol de Perpignan
70
élémentaire, effectué à l’aide d’une tarière dans une couche de profondeur déterminée (0-8
cm). Les prélèvements élémentaires (environ 200g de sol/prélèvement), ont été réunis et
homogénéisés pour constituer un échantillon global (4S) qui a été tamisé (≤ 2 mm) puis réduit
par quartages successifs (Mathieu et Pieltain, 2003) avant conservation au laboratoire (+ 4°C
ou - 20°C). Cet échantillon (4S) correspond donc à un sol ayant reçu 4 traitements successifs
par la sulcotrione à dose agronomique. Parallèlement, la zone « contrôle » a été
échantillonnée dans les mêmes conditions pour constituer le sol (C).
5- Dispositif expérimental : les microcosmes
La mise en place de systèmes expérimentaux permet d’appréhender le comportement
des produits phytosanitaires dans le sol, et, plus particulièrement, l’action des communautés
microbiennes du sol sur leur persistance. Ces dispositifs, appelés microcosmes ou
mésocosmes en fonction de leur taille, de leur complexité ou de leur localisation, constituent
en ce sens des modèles relativement pertinents en écologie microbienne, reconstituant des
fractions d’écosystèmes en conditions contrôlées de laboratoire.
Les microcosmes ont été préparés à partir des échantillons de sols natifs (4S) et (C),
ainsi que des sols stériles (St4S) et (StC), obtenus par autoclavage (2 cycles à 121°C pendant
20 minutes avec 1 jour de repos entre chaque autoclavage) d’une fraction des sols (4S) et (C).
Un calendrier de prélèvement a été préalablement établi (Figure II-2-3), prévoyant 1, 2, 3 ou 4
traitements successifs par la sulcotrione. Chaque date de prélèvement prévue a nécessité la
préparation initiale de 2 microcosmes (étude en duplicat) selon le protocole opératoire
suivant : 10,0 g de sol sont placés dans des boites de Pétri stériles (diamètre interne 55 mm) et
surajoutés par 0,6 mL d’une solution aqueuse de sulcotrione préalablement stérilisée par
filtration (membrane filter Minisart® 0,20µm) et de concentration 22 mg.L-1, ajout
correspondant à environ 1,2 µg de sulcotrione par g de sol. Ces microcosmes ont été ensuite
pesés individuellement avant d’être placés à l’obscurité, afin de limiter la photodégradation de
l’herbicide, et en chambre thermostatée (24,0 ± 0,2°C).

Deuxième partie – Chapitre 2 : Matrice dʼétude : le sol de Perpignan
71
Tout au long de l’expérimentation (Figure II-2-4), des pesées régulières nous ont permis de
conserver des teneurs en humidité relativement constantes, par ajout contrôlé d’eau stérile
sous hotte à flux laminaire.
Chacun des 4 sols (4S), (C), (St4S) et (StC) a nécessité la préparation de 40
microcosmes. Le premier traitement (TT1) par la sulcotrione a concerné la totalité de la
population des microcosmes. Des prélèvements ont été effectués (en duplicat) au cours du
premier mois, respectivement aux jours TT1-D0, TT1-D2, TT1-D7, TT1-D15 et TT1-D30. Un
deuxième traitement a alors été rajouté sur l’ensemble des microcosmes restants (en
conservant les mêmes conditions de température et d’humidité) et a été suivi de prélèvements
selon la même périodicité : TT2-D0, TT2-D2, TT2-D7, TT2-D15 et TT2-D30. Un troisième, puis
un quatrième traitement ont été conduits de la même manière avec les prélèvements
correspondants (TT3-D0, TT3-D2, TT3-D7, TT3-D15, TT3-D30 et TT4-D0, TT4-D2, TT4-D7, TT4-
D15, TT4-D30).
La teneur en sulcotrione extractible a été déterminée pour chacun des 160 microcosmes.
Figure II-2-3 : Synoptique du calendrier de traitements et de prélèvements des essais en microcosmes.

Deuxième partie – Chapitre 2 : Matrice dʼétude : le sol de Perpignan
72
Figure II-2-4 : Préparation et mise en place des microcosmes

73
Troisième partie: Méthodologies de travail

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
74
Chapitre1‐Méthodologiedel’analysechimique
La dégradation de la sulcotrione a été suivie successivement sur le sol de Perpignan
puis dans le milieu de culture sélectionné (cf III-2-2-2-1-2). Sur cette dernière matrice, outre
le comportement de la substance active, nous avons recherché l’apparition concomitante des
métabolites principaux, identifiés au cours d’une étude précédente (Chaabane, 2005).
Ce programme analytique a été réalisé au Laboratoire de Chimie de l’Environnement et des
Biomolécules (LCBE), en fonction de la disponibilité des moyens analytiques et notamment
de l’acquisition – tardive pour notre travail – des couplages CLHP/SM et CPG/SM.
1- Analyse des échantillons de sols 1-1-Réactifs et Matériels 1-1-1-Standard analytique Sulcotrione, référence Pestanal® 46318, pureté 98,8 %, origine Fluka analytical, France.
Solutions « mères », préparées à 200 mg.L-1 dans le méthanol de qualité CLHP ; solutions
« filles » à 0,1 ; 0,2 ; 0,5 ; 1 et 2 mg.L-1.
1-1-2- Autres réactifs
a) Méthanol et acétonitrile, qualité CLHP, origine Carlo Erba.
b) Dichlorométhane, pour analyse de pesticides, origine Riedel De Haën GmbH.
c) Acide chlorhydrique 38%, origine Prolabo, France.
d) Eau CLHP.
e) Acide trifluoroacétique (TFA), origine Sigma-Aldrich (Réf. T6508 –25mL), France.
f) Cartouches SPE SupelClean™ ENVI-Chrom, copolymère styrène/divinylbenzène, 500mg,
6 mL, origine Supelco, France.
1-1-3- Matériel classique de laboratoire

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
75
Agitateurs mécaniques à pales, évaporateurs rotatifs, chambre d’extraction sous pression
réduite, ampoules à décanter…
1-1-4- Appareillage chromatographique et conditions d’analyses Le chromatographe liquide haute performance – détection Ultra-Violet (CLHP/UV) utilisé
était une « chaine » Shimadzu équipée de deux pompes LC10ADvp, une colonne ODS
Hypersil C18 (250 mm x 4,5mm d.i., 5 µm), une boucle d’injection de 20 µL et un détecteur
SPD-10Avp UV-Visible programmé à λ = 265 nm, avec logiciel d’acquisition AZUR version
4.5 (figure III-1-1).
La phase mobile consistait en un mélange (A) : Acétonitrile et (B) : eau CLHP + 0,3% TFA,
délivré, avec un débit de 1 mL/min, selon le gradient d’élution suivant : 0 min : 30 % (A) 70
% (B), évoluant jusqu’à 50% (A) et 50 % (B) à 8 min et maintenu pendant 10 min.
L’étalonnage de cet appareillage a été effectué par injections, en triplicat, de solutions étalons
méthanoliques de sulcotrione (gamme de concentration 0,1 -2 mg.L-1). L’équation de la droite
de régression a été déterminée y = 45,0 (± 1,0) x + 2,9 (± 1,3) avec r² = 0,997 (± 0,003), ce
qui a permis d’estimer une limite de détection voisine de 0,020 mg.L-1.
Figure III-1-1 : Appareillage CLHP/UV Shimadzu utilisé pour les analyses de la sulcotrione à partir de la matrice sol.

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
76
1-2- Protocole opératoire 1-2-1- Préparation des échantillons
L’extraction de la sulcotrione à partir des échantillons de sols a été réalisée à partir
d’une méthode adaptée par Chaabane (2005) ; Trois étapes d’extraction et de purifications
successives ont été menées afin d’optimiser l’extraction de la sulcotrione tout en limitant la
co-extraction de substances interférentes.
-Extraction par solvant : les échantillons de sol (10 g) ont été mis en suspension dans 60 mL
d’un mélange binaire acétonitrile/solution aqueuse d’HCL 0,1M (90/10, v/v), puis agités
pendant 30 minutes à l’aide d’un agitateur mécanique à tige verticale muni d’une pale
d’agitation. La solution est ensuite filtrée sur fritté et papier filtre Whatmann GF/A. Le sol
partiellement extrait a été remis en suspension dans 30 mL du même mélange binaire, puis re-
agité pendant 15 minutes et filtré dans les mêmes conditions. Les filtrats ont été réunis et
évaporés sous vide (évaporateur rotatif à 30°C), afin d’éliminer l’acétonitrile. La phase
aqueuse acide (Ext 1) a été ensuite purifiée.
-Purification par partage liquide/liquide : La solution aqueuse a été extraite à deux reprises
par un volume égal de dichlorométhane (DCM) dans une ampoule à décanter. Les phases
organiques ont été réunies et évaporées juste à sec. Le résidu a été repris par 1 mL de
méthanol qualité CLHP puis « noyé » dans 20 mL de solution aqueuse d’HCl 0,1 M (Ext2).
-Purification complémentaire sur phase solide : Une nouvelle étape de purification a été
nécessaire. Elle consistait en l’extraction, à l’aide d’un dispositif adapté (chambre
d’extraction sous pression réduite) de l’extrait Ext2 sur cartouche EnviChromP®,
préalablement conditionnée successivement par 2 mL d’un mélange eau distillée/méthanol
(50/50, v/v) et 2 mL d’eau distillée. Après passage de l’extrait Ext2 et séchage de la
cartouche, la sulcotrione (ainsi que ses métabolites, bien que non recherchés dans cette étude
uniquement « sol ») ont été éluées par 5 mL de méthanol qualité CLHP (Ext3).
1-2-2- Analyse chromatographique (cf III-1-1-4)

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
77
1-3- Performances de la méthode d’analyse et validation
Après extraction des substances d’intérêt, purification de l’extrait et analyse
chromatographique, les performances de la méthodologie ont été évaluées (tableau III-1-1),
notamment en termes de répétabilité, de limites de détection (l.d.), de quantification (l.q.) et,
dans ce cas, de pourcentage de récupération des analytes. Deux niveaux de fortification (0,1 et
2 µg de sulcotrione.g-1 de sol) ont été testés en triplicat.
Ajout de 0,1 µg de sulcotrione
par g de sol (triplicat)
Ajout de 2 µg de sulcotrione
par g de sol (triplicat)
Limite de
détection
(l .d. µg.g-1 de sol)
Limite de
quantification
(l.q.µg.g-1 de
sol) %
recouvrement
Coefficient de
variation
%
recouvrement
Coefficient de
variation
0,02 0,05 84 % 21 % 92 % 10 %
Tableau III-1-1: Performances de la méthodologie d’analyse de la sulcotrione dans le sol.
Après ajouts de sulcotrione, certes supérieurs à la limite de quantification, les résultats
présentés (coefficient de variation et taux de recouvrement) paraissent acceptables, ainsi que
l’incertitude au voisinage de cette limite de quantification (0,05 µg.g-1).
Les teneurs obtenues lors de l’étude de dégradation de l’herbicide (cf IV-1-1) intègrent ces
données (limite de quantification et taux de recouvrement).
2- Analyse des échantillons de milieux de culture 2-1-Réactifs et matériels 2-1-1- Standards analytiques
Analytes Origine Référence Pureté
Sulcotrione Fluka analytical, France Pestanal® 46318 98,8 %
Acide 2-chloro-4 methylsulfonyl
benzoïque (CMBA)
Acros Organics, France 425010010 95,0 %
1,3-cyclohexane dione (CHD) Fluka analytical, France 29059 97,0 %
Tableau III-1-2 : Références et puretés des standards analytiques utilisés lors de l’analyse chimique des milieux de culture.

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
78
Pour mener à bien notre travail, trois solutions « mères » méthanoliques de sulcotrione (200
mg.L-1), de CMBA (200 mg.L-1) et de CHD (500 mg.L-1) ont été réalisées à partir des
standards analytiques présentés dans le tableau III-1-2. Les solutions « filles » ont été
prépararées à 0,1 ; 0,2 ; 0,5 ; 1 et 2 mg.L-1.
L’acide 5,7-dicéto-7-(2-chloro-4-méthylsulfonylphényl) heptanoïque a été synthétisé au
LCBE (Chaabane, 2005). Une solution méthanolique (1 mg.L-1) a été utilisée (tr = 9,95 min)
comme contrôle de présence au cours de la dégradation de la sulcotrione dans le milieu de
culture.
2-1-2- Autres réactifs a) Méthanol et acétonitrile, qualité CLHP, origine Carlo Erba.
b) Dichlorométhane, pour analyse de pesticides, origine Riedel De Haën GmbH.
c) Acide chlorhydrique 38%, origine Prolabo, France.
d) Eau CLHP.
e) Acide trifluoroacétique (TFA), origine Sigma-Aldrich (Réf. T6508 –25mL), France.
6 mL, origine Supelco, France.
f) Trimethylsilyldiazométhane 2M, solution dans diéthylether, origine Sigma-Aldrich, France
(réf.527254-100mL).
2-1-3- Matériels classiques de laboratoire Centrifugeuses de laboratoire, évaporateurs sous flux d’azote, ampoules à décanter… 2-1-4- Spectrophotomètre UV/visible
La croissance de la biomasse microbienne a été suivie par la mesure de l’absorbance à λ =
600 nm à partir d’aliquotes des milieux de cultures ensemencées. L’appareil utilisé a été un
spectrophotomètre monofaisceau équipé d’un monochromateur à réseau holographique et
d’un détecteur au silicium (Secoman, Anthelie Advanced, France).
2-1-5- Appareillages chromatographiques et conditions d’analyses

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
79
2-1-5-1- CLHP/UV Les extraits ont été analysés à l’aide d’une « chaine » JASCO équipée de deux pompes (Jasco
880PU Intelligent HPLC pump), une colonne ODS Hypersil C18 (250 mm x 4,5mm d.i., 5
µm), une boucle d’injection de 20 µL et un détecteur Jasco-875-UV programmé à λ = 265
nm, avec logiciel d’acquisition Borwin 1.50
La phase mobile consistait en un mélange (A) : Acétonitrile et (B) : eau CLHP + 0,3% TFA,
délivré, avec un débit de 1 mL/min, selon le gradient d’élution suivant : 0 min : 30 % (A) 70
% (B) jusqu’à 4 min, évoluant jusqu’à 60% (A) et 40 % (B) à 7 min et maintenu pendant 15
min.
Un chromatogramme obtenu avec un mélange de solutions « étalons » est présenté figure III-
1-2. Ce chromatogramme correspond à l’injection d’un extrait obtenu à partir d’un échantillon
de milieu de culture surajouté avant extraction, par un mélange des trois solutions étalons
(sulcotrione, CMBA et CHD). Pour ces trois composés, la gamme de concentration a été de 2
à 30 mg.L-1. Les équations correspondantes sont présentées dans le tableau III-1-3.
Figure III-1-2 : Chromatogramme CLHP/UV de la sulcotrione et de ses principaux
métabolites (CHD et CMBA).

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
80
Tableau III-1-3 : Calibration de l’analyse sulcotrione et métabolites sur milieu de culture avec l’appareillage CLHP Jasco. 2-1-5-2- CLHP/SM
Les analyses relatives à la présence de 1,3-cyclohexanedione (CHD) par CLHP/SM
ont été effectuées à l’aide d’un appareillage LCQ-Fleet (Thermo Fisher Scientific), disponible
au LCBE depuis septembre 2010, équipé d’une source d’ionisation « Electrospray » (ESI) et
d’un analyseur « ion-trap » 3D. L’analyse de la CHD a été effectuée, entre les m/z 60 et 300,
en modes « full scan » positif (4 kV) et négatif (3,5 kV). Les conditions
chromatographiques étaient: HPLC Accela Thermo Fisher, avec pompe Accela 600, passeur
automatique d’échantillons (Accela autosampler) et détecteur à barrettes de diodes (Accela
PDA detector) ; colonne phénomenex Gemini C6-phényl (150 mm x 3,0 mm ; 5µm) ; phase
mobile : mélange (A) : acide formique 0,1 % dans acétonitrile et (B) : acide formique 0,1 %
dans eau CLHP, délivré, avec un débit de 0,4 mL/min, selon le gradient d’élution suivant : 0
min : 10 % (A) 90 % (B) jusqu’à 2 min, évoluant jusqu’à 90% (A) et 10 % (B) à 20 minutes.
Le temps de rétention, pour la 1,3 cyclohexanedione (CHD), a été de 4,51 minutes (PDA).
L’ion m/z 113, en mode positif, a été retenu pour confirmer ou infirmer la présence de la 1,3
CHD dans les extraits (figure III-1-3). Dans ces conditions - non encore optimisées- la limite
de quantification a été estimée à environ 0,5 mg.L-1.
Analyte (méthode
chromatographique)
Temps de
rétention (minutes)
Gamme de
concentrations (mg.L-1)
Droite de régression
Valeurs moyennes (écart-type)
Sulcotrione
11,08
2 - 30
y = 65504 (±1420) x -19214
(± 10045)
r² = 0,997 (± 0,002)
CMBA
5,15
2 - 30
y = 14537 (± 840) x - 7247
(± 810)
r² = 0,999 (± 0,001)
CHD
4,54
2 - 30
y = 61323 (± 3101) x - 34162
(± 19205)
r² = 0,995 (± 0,003)

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
81
Figure III-1-3 : (a) Spectre de masse (CLHP/SM-ESI) de la 1,3-cyclohexanedione (CHD) en mode positif. (b) Chromatogrammes du standard analytique (10 mg.L-1) par détection barrette de diodes et (c) détection SM par extraction du fragment m/z 113 en mode positif. 2-1-5-3- CPG/SM
L’appareillage CPG/SM (Thermo Fisher Scientific) a été également disponible au
LCBE à partir de septembre 2010. Il nous a notamment permis de confirmer qualitativement
et quantitativement la présence d’acide 2-chloro-4 methylsulfonyl benzoïque (CMBA) après
dérivation (cf paragraphe III-1-3-2-3-3). Le système consiste en un CPG Focus GC couplé à
un détecteur de spectrométrie de masse DSQ2 (Thermo Fisher Scientific) fonctionnant
(impact électronique 70eV), en mode full scan (m/z 100 – 350) avec extraction, à partir du
« scan », des fragments m/z 217, 248 et 250, caractéristiques du CMBA méthylé (figure III-1-
5). Les conditions chromatographiques étaient les suivantes : injection en mode « split »,
température de la ligne de transfert 300°C, colonne G-SQC GC (5 %
phénylméthylpolysiloxane, 15m x 0,25 mm, 0,25 µm), avec un gradient de température :
température initiale 80 C° (1 min), augmentation de 10°/min jusqu’à 260°C, maintenue

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
82
pendant 15 minutes ; gaz vecteur Hélium 6.0 (1 mL.min-1). Dans ces conditions opératoires, le
temps de rétention du CMBA-méthylé a été de 14,81 minutes.
L’étalonnage de l’appareillage a été réalisé à partir de solutions méthanoliques
« étalons » méthylées, de concentrations comprises entre 0,5 et 20 mg.L-1, en mode « full
scan », et extraction de l’ion m/z 217 (figure III-1-4). La droite de régression correspondante
est présentée sur la figure III-1-5. La limite de quantification a été évaluée à 0,20 mg.L-1. Une
optimisation de cette application chromatographique est en cours (injection splitless ; mode
d’acquisition SIM : fragments m/z 250, 248, 217 et 155).
Figure III-1-4: (a) Spectre de masse du dérivé méthyle de l’acide 2-chloro-4 methylsulfonyl benzoïque (CMBA), (standard analytique à 20 mg.L-1) obtenu en CPG/SM (mode positif, SM, impact électronique). (b) Chromatogramme montrant la présence du CMBA méthylé (fragments m/z, 217, 248 et 250) dans le milieu de culture (MS) supplémenté en sulcotrione de la souche 1OP après 27 jours d’incubation.

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
83
Figure III-1-5: Droite d’étalonnage de l’ester méthylique provenant du CMBA obtenu après dérivation et analyse en CPG/SM 2-2- Protocole opératoire 2-2-1- Péparation des échantillons
L’extraction des composés d’intérêt à partir des échantillons de milieux de culture a
été mise au point au laboratoire, après une étape préalable de centrifugation. Des échantillons
de milieu de culture vierge ont été surajoutées par des quantités connues et croissantes (2-30
mg.L-1) de substances à doser (cohorte chimique sulcotrione + CMBA + CHD). Une aliquote
(1 mL) de milieu de culture a été prélevée et centrifugée pendant 15 minutes à 4000 rpm à
température ambiante. Le surnageant a été ensuite récupéré et acidifié par 0,3 mL d’HCl (0,16
M), puis amené à un volume de 2,5 mL par apport d’eau milliQ. La solution est alors extraite
deux fois avec 2,5 mL de dichlorométhane (DCM). L’extrait DCM est évaporé juste à sec
sous flux d’azote et le résidu repris par 1,0 mL de méthanol de qualité CLHP (ExtMC) avant
analyse chromatographique.

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
84
2-2-2- Analyse chromatographique
Initialement les extraits milieux de culture ont été analysés par CLHP/UV. Les
résultats obtenus ont pu être ensuite confirmés, pour la CHD, par CLHP/SM et pour le
CMBA, par CPG/SM après méthylation.
2-3- Performances de la méthode d’analyse et validation
Des échantillons non ensemencés du milieu de culture (MS) ont été supplémentés par
des quantités connues et croissantes (2-30 mg.L-1) de substances à doser (cohorte chimique
sulcotrione + CMBA + CHD), afin de déterminer les performances de la méthode d’analyse.
La méthode validée a été ensuite appliquée aux échantillons des essais de biodégradation,
obtenus après ensemencement par des souches susceptibles de dégrader la sulcotrione ajoutée
au milieu de culture (MS).
2-3-1- CLHP/UV
La droite de régression linéaire de la méthode complète, obtenue après analyse en
triplicat des échantillons témoins surajoutés, est présentée figure III-1-6.
Figure III-1-6: Droite d’étalonnage de la sulcotrione surajoutée à concentrations croissantes dans le milieu de culture (MS), après extraction et analyses CLHP/UV.

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
85
L’équation correspondante a été déterminée : y = 47154 (± 1650) x – 13276 (± 11974)
avec r² voisin de 0,9967 (± 0,0003). La limite de quantification a été estimée à 0,5 mg.L-1. Le
coefficient de détermination supérieur à 0,996 et le coefficient de variation sur la pente de la
droite inférieur à 4% nous permettent de considérer ce protocole analytique comme tout à fait
acceptable et utilisable pour l’étude de biodégradation de la sulcotrione dans ce milieu de
culture. Les échantillons provenant de l’étude de dégradation ont été ensuite quantifiés par
rapport à cet étalonnage.
Parallèlement, une étude comparable a été menée à partir d’échantillons de milieu minimum
MS, surajoutés par les métabolites principaux (CMBA et CHD). Les équations
correspondantes ont été déterminées :
Pour le CMBA
y = 10537 (± 1210) x -4224 (± 2009) avec r² = 0,9973 (± 0 ,0010)
Limite de quantification : 1mg.L-1.
Pour la 1,3-CHD
y = 34500 (± 3121) x -7240 (± 5421) avec r² = 0,9960 (± 0,0006)
Limite de quantification : 1mg.L-1.
Cette méthode s’est avérée applicable au dérivé « acide heptanoïque », initialement rajouté à
la concentration de 2 mg.L-1 (tr = 9,95 min).
2-3-2- CLHP/SM
Les échantillons de milieux de culture obtenus lors de l’étude de biodégradation de la
sulcotrione et, de manière concomitante, de l’apparition éventuelle de métabolites, ont été
conservés par congélation du fait de la non-disponibilité du couplage CLHP/SM au moment
de l’expérimentation. Dès la mise en service de cet appareillage (octobre 2010), nous avons
souhaité confirmer l’absence de résidus de CHD (du moins au-delà de la limite de
quantification) par infusion et par couplage avec la chromatographie liquide.

Troisième partie – Chapitre 1 : Méthodologie de lʼanalyse chimique
86
2-3-4- CPG/SM
En chromatographie gazeuse, l’analyse du CMBA, dérivé de l’acide benzoïque, n’est
possible qu’après une réaction de méthylation appropriée. Mashimoto et al. (1981) ont décrit
un protocole de méthylation particulièrement adapté au dosage des acides carboxyliques. Ces
auteurs indiquent que le triméthylsilyldiazométhane (TMSCHN2) réagit rapidement, à
température ambiante, en présence de méthanol et avec de bons rendements, sur des acides
carboxyliques pour donner des esters méthyliques (Figure III-1-7). Le TMSCHN2 constitue
donc une alternative intéressante au diazométhane pour l’analyse de ces acides par
chromatographie gazeuse.
25 µL de solution de TMSCHN2 2,0 M dans l’éther diéthylique et 0,5 mL de méthanol de
qualité CLHP sont ajoutés à 0,5 mL d’extrait de milieu de culture (ExtMC). L’ensemble est
ensuite agité à température ambiante pendant 2 heures.
L’estérification se produit rapidement et quantitativement en présence d’un excès
de TMSCHN2, la réaction pouvant être suivie par la disparition progressive de la couleur
jaune. L’extrait méthylé est alors analysé par CPG/SM dans les conditions décrites
précédemment. Le rendement de la méthode, après ajout d’une quantité connue de CMBA à
une aliquote du milieu de culture (MS), extraction et méthylation, a été estimé à 70%
(comparativement à une solution étalon méthanolique méthylée de concentration connue).
Cette méthode nous a permis, malgré la disponibilité tardive de l’appareillage, de confirmer la
présence et les teneurs de CMBA dans les extraits de milieux de culture après biodégradation.
Figure III-1-7: Principe de la réaction de dérivation de l’acide 2-chloro-4 methylsulfonyl benzoïque (CMBA), en présence d’un excès de triméthylsilyldiazométhane (TMSCHN2) et de méthanol conduisant à la formation de l’ester méthylique correspondant.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
87
Chapitre2‐Méthodologiedel’analysebiologique
1- Cadre de l’étude : organisation des travaux
Au cours de notre étude, les analyses microbiologiques se sont déroulées en trois
étapes successives.
En premier lieu, il a été nécessaire de développer une approche cultivable originale et
efficace, en optimisant les conditions de cultures sur milieu synthétique (intégration de l’effet
matrice de l’écosystème sol sur les conditions de cultures au laboratoire, source de carbone,
source d’azote, richesse nutritive et oxygénation), afin de favoriser, autant que possible,
l’isolement et la sélection de souches bactériennes telluriques capables de pousser sur un
milieu synthétique, en présence de sulcotrione.
Dans une deuxième étape, les isolats bactériens ont été testés, séparément, sur leurs
capacités potentielles à utiliser la sulcotrione avec plus ou moins d’efficacité, comme seule
source de carbone et d’énergie à l’aide d’essais de biodégradation en cultures liquides.
La troisième étape a consisté à identifier les isolats bactériens d’intérêt par une
méthodologie de travail s’appuyant sur une approche phylogénétique et fonctionnelle.
L’objectif était dans un premier temps de pouvoir caractériser les souches dégradantes en
comparaison de souches non dégradantes prises comme contrôles, puis dans un deuxième
temps, d’identifier formellement les isolats par un séquençage de leur ADN ribosomal 16S
(ADNr 16S), complété par une caractérisation fonctionnelle.
2- Approche cultivable : essai d’isolement de souches bactériennes dégradant la sulcotrione 2-1- Principe et stratégie de travail
Au cours de cette étude, trois types de milieux de culture ont été nécessaires pour
développer une approche cultivable efficace afin d’essayer de s’affranchir, du moins en partie,
des effets du phénomène du « Great Plate Count Anomaly » (Staley et Konopka, 1985), bien
connu des microbiologistes du sol. En effet, une grande difficulté dans l’étude de la flore

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
88
microbienne du sol réside dans l’isolement et la culture de souches microbiennes sur des
milieux de culture synthétiques conventionnels.
En premier lieu, afin de pouvoir isoler avec succès des souches bactériennes
telluriques, nous avons donc élaboré un milieu synthétique le plus voisin possible de leur
microhabitat d’origine, en incorporant l’échantillon environnemental « sol » au milieu de
culture approprié. L’objectif était double : (i) réussir à cultiver des souches au laboratoire
issues d’un échantillon de sol et (ii) sélectionner des isolats bactériens capables de se
développer en présence d’une forte concentration d’herbicide.
Un milieu complexe appelé (NS) (« nutrient source ») a donc été préparé directement à partir
d’un échantillon composite de sol provenant de la parcelle de l’IUT de Perpignan (sol C) et
supplémenté en sulcotrione (30 mg.L-1). L’ensemble sol + herbicide peut être considéré
comme la source nutritive de la microflore tellurique. En outre, le sol, présent en proportion
importante dans le milieu de culture (40 grammes), a également servi de matrice de support
aux colonies, permettant ainsi de s’affranchir en partie des effets délétères de l’agar sur la
croissance de certaines souches telluriques (Janssen et al., 2002). Cette technique cultivable a
été adaptée des travaux de Ferreira et al. (2009) qui ont étudié les effets de trois insecticides
(aldicarb, chlorpyrifos et deltaméthrine) et trois fongicides (tébuconazole, métalaxyl et
mancozebe) sur les communautés bactériennes cultivables de sols agricoles brésiliens.
Dans un deuxième temps, un milieu sélectif minimum (minéral) dans lequel
l’herbicide était la seule source de carbone et/ou d’énergie (sulcotrione 30 mg.L-1) a été
utilisé, selon une adaptation des travaux de Rousseaux et al. (2001). Ce milieu (MS) a permis
de réaliser des cultures liquides répétées, avec forte pression de sélection de l’herbicide, sur
les souches bactériennes précédemment isolées et purifiées. L’objectif était d’induire une
adaptation génétique des microorganismes présents dans le milieu au cours des cultures,
conduisant ainsi à l’apparition progressive d’une population apte à utiliser l’herbicide comme
seule source de carbone et/ou d’énergie. Par la suite, ce milieu minéral nous a également servi
à mener nos essais de dégradation en cultures liquides à partir des isolats provenant des
cultures répétées, avec pression de sélection de l’herbicide. Enfin, l’ajout d’agar dans le
milieu (MS) a également été nécessaire afin de tester la capacité de croissance des souches
isolées sur milieu gélosé en présence de l’herbicide.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
89
Un milieu riche non différentiel - trypto caséine de soja (Triptic soy broth ou TS médium)
supplémenté en sulcotrione et contenant de l’agar, a été utilisé pour réaliser les isolements sur
milieux gélosés nécessaires à l’obtention de souches pures à partir de nos isolats. Ce bouillon
de soja tryptique est un milieu liquide polyvalent préconisé dans des procédures
d’enrichissement et de culture de microorganismes aérobies non exigeants ou modérément
exigeants. Les produits issus de la digestion enzymatique de caséine et de semoule de soja,
présents dans ce milieu de culture, apportent les aminoacides et les composés azotés
complexes nécessaires à la croissance de ces microorganismes. Le glucose constitue une
source d’énergie, le chlorure de sodium assure l’équilibre osmotique et le phosphate
bipotassique (K2HPO4) permet de contrôler le pH.
Ce même milieu (TS) a également été utilisé sans ajout d’agar, afin d’obtenir par la
technique de culture en croissance discontinue ou en lot (« batch cultures»), une
concentration suffisante de biomasse microbienne en phase exponentielle de croissance. Cette
croissance a été suivie de façon régulière par la mesure de l’absorbance à λ = 600 nm à partir
d’aliquotes de cultures liquides, avant d’entreprendre les essais de biodégradation après une
étape de lavage à l’aide de tampon Knapp.
2-2- Milieux de culture et matériels utilisés 2-2-1- Préparation des milieux de culture 2-2-1-1- Milieu complexe « sol » ou nutrient soil medium (NS)
- réactifs
Sol (C) de la zone de contrôle C de la parcelle expérimentale de l’IUT de Perpignan
Agar technique en poudre AEB175006, AES Laboratoire, France
Solution de NaOH 0,16 M
Cycloheximide, minimum 94 %, référence C7698-5g, Sigma –Aldrich, France
Solution aqueuse de cycloheximide à 1g.L-1, stérilisée sur filtres stériles Acrodisc®
Sulcotrione, 98,8 %, Pestanal® 46318, Fluka analytical, France
Solution aqueuse de sulcotrione à 100 mg.L-1, stérilisée sur filtres stériles Acrodisc®
Filtres stériles Acrodisc® 32, Supor® 0,2 µm, GelmanSciences

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
90
Seringues stériles à usage unique Injekt (20 mL), référence 4606205V, Braun,
Germany.
- préparation
Le sol (C), prélevé sur la parcelle de l’IUT a été tamisé (tamis 2 mm) et séché trois
jours à l’étuve à 65°C. Le milieu (NS) est préparé à partir de 400 g de ce sol sec, 20 g d’agar
dans environ 350 mL d’eau distillée; le pH est mesuré et éventuellement amené à 8 (pH de la
solution de sol, cf tableau II-2-1) avec la solution de NaOH. Le mélange est autoclavé 1,5
heures à 120°C. Après ajout de 50 mL de solution aqueuse de cycloheximide (50 mg) et de
300 mL de solution aqueuse de sulcotrione (30mg), le volume est ajusté à 1000 mL avec de
l’eau distillée stérile.
Le mélange est homogénéisé et, avant prise en masse de l’agar, coulé dans des boites de Pétri
stériles (diamètre interne 8,6 cm).
2-2-1-2- Milieu minimum (MS)
La préparation de ce milieu de culture sélectif nécessite le mélange de plusieurs solutions
à préparer extemporanément (cf tableau III-2-1). Dans un premier temps, les solutions (A),
(B) et (C) et l’eau distillée (qsp 800 mL) sont mélangées et autoclavées 1h30 à 120°C. Les
solutions de vitamines et de sulcotrione sont ensuite rajoutées par filtration stérilisante. Le
milieu de culture (MS) liquide (en absence d’agar) a été ensuite utilisé dans des erlenmeyers
bafflés (500 mL), cotonnés et préalablement stérilisés. Après préparation, ce milieu doit être
conservé à + 4°C et utilisé rapidement.
Dans certains cas, 15 g d’agar ont pu être rajoutés, avant autoclavage, de manière à obtenir un
milieu gélosé, utilisable lors de certains tests de biodégradation sur boite de Pétri.
Le pH doit également être mesuré et éventuellement amené à 8.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
91
Solutions à
préparer
Eléments
constitutifs
Concentration de la
solution à préparer
(g.L-1)
Volume de
cette
solution qsp
1000 mL de
milieu de
culture (mL)
Concentration de
l’élément dans le
milieu de culture
(mg.L-1)
K2HPO4 100 16 1600
KH2PO4 100 4 400
MgSO4, 7H2O 40 5 200
NaCl 10 10 100
CaCl2 20 1 20
Solution (A) :
Eléments
minéraux
FeSO4, 7H2O 5 1 5
2
1,8
0,2
0,8
0,25
Solution (B) :
Mélange
d’oligo-
éléments
H3BO3
MnSO4, H2O
ZnSO4
CuSO4
Na2MoO4, 2H2O
Co(NO3)2
2
1,80
0,20
0,80
0,25
traces
1
traces
Solution (C) :
Source d’azote
(NH4)2SO4
175
20
3,5
Eau purifiée H2O - qsp 800mL -
0,1
Solution (D) :
Mélange de
vitamines
Thiamine
Biotine
0,10
0,04
1
0,04
Solution de
sulcotrione
H2O
0,150
200
30
Agar (cas d’un
milieu solide)
-
-
15 g
-
Tableau III-2-1: Préparation du milieu de culture minimum (MS), pH 8.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
92
2-2-1-3- Milieu complexe trypto - caséine de soja ou Triptic soy (TS) Le produit commercial (Bouillon Trypcase Soja, référence 51019) nous a été fourni par
BioMérieux, France. Sa composition est présentée dans le tableau III-2-2.
Elément Quantité (g)
Digestion pancréatique de caséine bovine 17,0 g
Digestion peptidique de semoule de soja 3,0 g
NaCl 5,0 g
K2HPO4 2,5 g
C6H12O6 (dextrose) 2,5 g
Tableau III-2-2: Composition du milieu de culture TS (pH 7,3 ± 0,2) pour 1 litre d’eau purifiée.
Ce milieu, a été préparé, selon les recommandations du fabricant, par l’ajout de 30 grammes
de préparation (forme pulvérulente) dans un litre d’eau purifiée. La suspension homogénéisée,
a été ensuite chauffée jusqu’à dissolution complète répartie, selon l’usage envisagé, dans des
erlenmeyers ou dans des tubes (20 x 200 mm), cotonnés et autoclavés 15 minutes à 120°C.
Dans certains cas, des milieux gélosés ont été préparés par ajout de 15 g.L-1 d’agar par litre de
milieu (TS), afin de purifier nos souches bactériennes par striages successifs sur boites de
Pétri lors de la procédure d’isolement.
Afin d’éliminer toute trace résiduelle de bouillon TS après l’étape de production de biomasse,
l’utilisation d’une solution de rinçage (Tampon Knapp) a été nécessaire avant d’entreprendre
les essais de biodégradation dans le milieu de culture (MS). La composition de ce tampon est
donnée dans le tableau III-2-3.
Elément Quantité (g)
K2HPO4 1
KH2PO4 1
MgSO4, 7 H2O 0,04
FeCl3 0,004
Tableau III-2-3: Composition du tampon Knapp (pH 6,6) pour 1 litre d’eau purifiée.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
93
2-2-2- Matériels utilisés dans le cadre de l’approche cultivable
L’ensemble des essais d’isolement et de culture des souches bactériennes telluriques
ont été menés dans les locaux de l’IUT de Perpignan, au sein du Département de Génie
Biologique. Nous avons pu disposer d’un environnement de travail bien adapté aux
techniques cultivables (figure III-2-1).
Figure III-2-1 : Exemples d’appareillages et de matériels utilisés en culture Pasteurienne lors de l’étude. IUT de Perpignan, Département Génie Biologique (UPVD).
2-3- Protocole opératoire 2-3-1- Isolement de souches bactériennes sulcotrione – résistantes
Plusieurs échantillons contenant 10 grammes de sol (4S) provenant de la surface (4S)
ayant été traitée par la sulcotrione (cf paragraphe II-2-4) ont été placés dans des erlenmeyers
stériles de 500 mL en présence de 90 mL d’eau physiologique (0,9 % NaCl) puis agités à 200
rpm pendant 30 minutes afin de constituer une suspension de sol homogène. Une aliquote de
cetet suspension (500 µL) a été ensuite étalée sur milieu gélosé (NS) contenant 30 mg.L-1 de
sulcotrione. Parallèlement, 1 mL de cette suspension de sol a été additionnée à 9 mL d’eau
physiologique stérile. Après homogénéisation au vortex, une série de dilutions décimales a été

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
94
réalisée (jusqu’à 10-4) et étalée sur le milieu (NS). Deux boites ont été réalisées pour chaque
dilution. Les boites de Pétri ont été ensuite placées à 28°C pendant 7 jours à l’obscurité et
observées régulièrement. Deux contrôles constitués d’aliquotes de 500 µL de suspension de
sol ont été étalés sur gélose (NS) sans sulcotrione et également incubés dans les mêmes
conditions. Les colonies observées sur gélose (NS) supplémentée en sulcotrione ont été
identifiées comme des souches bactériennes capables de pousser en supportant la présence de
l’herbicide concentration relativement forte dans le milieu de culture (30 mg.L-1). Après
observation et repérage de colonies bien distinctes sur la gélose, les isolats ont été purifiés.
Brièvement, chaque colonie a été prélevée à l’aide d’une anse de platine stérile, puis
ensemencée dans 100 mL de milieu liquide (TS) supplémenté en sulcotrione (30 mg.L-1)
pendant 24 heures sous agitation (200 rpm, 30°C). Plusieurs repiquages et étalements
successifs sur gélose (TS) contenant de la sulcotrione (30 mg.L-1) ont été nécessaires afin
d’obtenir des souches pures.
Cette première étape d’isolement est schématisée dans la figure III-2-2.
Figure III-2-2 : Protocole d’isolement de souches bactériennes telluriques sulcotrione-résistantes.
10 g de SOL (4S) +!90 mL H2Oavec !
0,9 % NaCl!
Agitation 30 minutes!
Aliquotes 500 μL!
Dilutions en série!
Etalement au râteau des dilutions !
Milieu (NS) + sulcotrione!
Milieu (NS) – sulcotrione!
Croissance à 28°C!à lʼobscurité!
Colonies sulcotrione-résistantes!
(TS) + sulcotrione!
Colonies ensemencées dans!bouillon (TS) + sulcotrione!
24 heures sous agitation à 30°C!Repiquages
successifs!Contrôle!
1! 2!
3!
4!
5!6!
Isolement par stries dʼépuisement!

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
95
2-3-2- Isolement de souches bactériennes dégradant la sulcotrione 2-3-2-1- Cultures liquides répétées des isolats bactériens avec pression de sélection de l’herbicide
Chaque isolat sulcotrione-résistant, caractérisé par sa capacité à pousser en présence
d’une forte concentration d’herbicide sur milieu (MS), a été mis en culture dans un milieu
liquide (MS) supplémenté en sulcotrione (30 mg.L-1). Brièvement, chaque isolat a été
ensemencé dans 100 mL de milieu (MS) supplémenté en herbicide dans des erlenmeyers
baflés (500 mL) et incubé à 28°C sous agitation (150 rpm) pendant 20 jours. Au bout de 20
jours de culture, une aliquote de 10 mL a été transférée dans un nouvel erlenmeyer stérile
(500 mL) contenant 90 mL de milieu (MS) supplémenté en herbicide et incubé à nouveau
dans les mêmes conditions. Cinq cycles de cultures répétés (C1C5) avec pression de
sélection de l’herbicide ont été ainsi conduits (cultures en triplicat) afin de provoquer une
adaptation physiologique des isolats aux conditions de culture. Au bout des cinq cycles, la
capacité de chaque isolat à pousser sur milieu gélosé (MS) supplémenté en sulcotrione (30
mg.L-1) a été testée par étalement d’aliquotes de culture (100 à 10-3) et striage sur gélose à
partir des dilutions décimales (duplicats par boite). Les milieux gélosés ont été incubés 5 jours
environ à l’obscurité à 28°C. Parallèlement, à des fins d’entrée en collection, 20 mL de
bouillon (TS) supplémenté en sulcotrione (30 mg.L-1) ont été ensemencés par une aliquote de
1 mL de chaque souche pure issue du cycle 5 et incubés 12 heures sous agitation à 28°C. La
culture a été ensuite centrifugée 10 minutes à 4000 rpm à 4°C, le surnageant jeté et le culot
bactérien resuspendu dans une solution stérile de milieu (TS) contenant 25 % de glycérol puis
congelé dans l’azote liquide et conservé à - 80°C.
La figure III-2-3 résume cette étape.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
96
Figure III-2-3 : Cultures répétées des isolats sulcotrione-résistants avec pression de sélection de l’herbicide.
2-3-2-2- Estimation de la capacité de dégradation des isolats bactériens vis-à-vis de la sulcotrione
La technique de « batch » également dénommée culture en discontinu ou en lot a été
utilisée afin d’obtenir une biomasse microbienne concentrée en phase exponentielle de
croissance, nous permettant de suivre, dans de bonnes conditions opératoires, la dissipation de
l’herbicide dans le milieu de culture (cf figure III-2-4). Une aliquote de 1 mL de chaque isolat
provenant de C5 a été ensemencée dans 49 mL de bouillon (TS) supplémenté en sulcotrione
(30 mg.L-1). Les cellules ont été récoltées par centrifugation (15 minutes, 4000 rpm, 4°C)
pendant la phase exponentielle de croissance, après 15 heures de culture à 30°C sous agitation
(150 rpm). Le surnageant a été jeté, le culot bactérien lavé deux fois avec 20 mL de tampon
Knapp afin d’éliminer toute trace de milieu (TS) résiduel. Le culot a été immédiatement
resuspendu dans un milieu minimum (MS) supplémenté en sulcotrione comme unique source
PHASE DʼADAPTATION = 5 cycles de culture (C1 à C5)" 1 cycle = 20 jours sous agitation à 28°C"
Milieu (MS) + sulcotrione"Milieu (MS) + sulcotrione (90 mL)"
C1" C5"C…"
Milieu (MS) + sulcotrione"
Transfert de 10 mL de culture au bout de 20 jours dans milieu neuf" Transfert de 10 mL"Ensemencement à"
lʼanse"
ENTREE EN COLLECTION"-80°C"
Aliquote 1 mL"
Bouillon (TS) + sulcotrione "
12 heures de culture"
Milieu (MS) + sulcotrione"
Etalement au râteau"
ESSAIS DE BIODEGRADATION " Mesure de la capacité de dégradation des isolats bactériens vis-à-vis de la
sulcotrione en cultures liquides"
1"
2"
3"4"
Milieu (MS) + sulcotrione"
Stries sur boite"
Isolat purifié"
(TS) + sulcotrione"
Dilutions en série"

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
97
de carbone et/ou d’énergie afin d’obtenir une absorbance voisine de 0,3 (λ = 600 nm) dans le
milieu de culture. Les échantillons ont été ainsi cultivés à 30°C sous agitation (200 rpm) et à
l’obscurité afin de limiter la photodégradation de la sulcotrione. Une aliquote de 1 mL de
chaque culture a été régulièrement prélevée selon un calendrier pré-établi (0, 6, 15 et 27
jours), la mesure de l’absorbance (λ = 600 nm) a été effectuée et les échantillons ont été
extraits puis analysés par chromatographie (cf paragraphe III-2-2) afin d’établir les cinétiques
de dégradation de l’herbicide et d’apparition des métabolites principaux dans le milieu de
culture. Un milieu de culture (MS) supplémenté en sulcotrione (30 mg.L-1) non ensemencé et
un milieu (MS) ensemencé non supplémenté en sulcotrione ont servi de contrôles (triplicats)
au cours de nos essais.
Figure III-2-4 : Protocole de sélection de souches bactériennes sulcotrione-dégradantes après cinq cycles de cultures répétées.
Milieu (MS) + sulcotrione!
Bouillon (TS) + sulcotrione!
15 heures sous agitation à 30°C!
Aliquote de 1 mL !de culture!
Culture C5!
Milieu (MS) + sulcotrione!
BATCH CULTURES!
Centrifugation de la culture !Lavage du culot avec
TAMPON KNAPP pH 6,6!
Absorbance voisine de 0,3 !(λ = 600 nm).!
Croissance à 28°C!à lʼobscurité avec
agitation !
Milieu (MS)! Milieu (MS) !non ensemencé!
Contrôles!
RESUSPENSION DU CULOT BACTERIEN!DANS: !
Suivi de! lʼabsorbance!
Cinétiques de dégradation de!
la sulcotrione et dʼapparition de
métabolites éventuels!
1! 2!
3!
4!
5!Aliquote 1 mL!

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
98
2-3-3- Caractérisation morphologique et biochimique des isolats dégradants
L’observation macroscopique des isolats a été réalisée à partir des cultures sur milieux
gélosés (MS) et (TS) afin d’obtenir des informations sur l’aspect des colonies, c’est-à-dire sur
leur taille, leur couleur, leur forme (contour) leur coupe (de profil), leur consistance
(crémeuse, filante ou muqueuse), l’aspect de la surface (lisse ou rugueuse) et l’opacité
(opaque, translucide ou transparente).
L’observation microscopique des isolats s’est appuyée sur la réalisation d’états frais et de
colorations de Gram. Ces observations ont été effectuées à partir des cultures liquides (MS),
supplémentées en sulcotrione lors des essais de biodégradation, mais également à partir des
colonies isolées et purifiées ayant poussé sur milieu gélosé (MS) supplémenté avec
l’herbicide. Nous avons ainsi pu acquérir des informations sur la taille et la mobilité des
isolats dégradants.
Brièvement, pour la réalisation de l’état frais, une à deux gouttes de suspension cellulaire (à
partir des colonies sur gélose) ou de milieu de culture ont été placé au centre d’une lame de
verre (Marienfeld, Germany) à l’aide d’une pipette Pasteur stérile puis recouvertes d’une
lamelle de verre (ESCO, USA). Le lutage a été effectué à l’aide de paraffine. L’observation
des isolats a été réalisée sur fond clair au microscope optique avec l’objectif X40 (Leica
CME, Leica Microsystems, France) puis avec l’objectif X100 après ajout d’une goutte d’huile
d’immersion (réactifs RAL, France) sur la lamelle.
Afin de caractériser la morphologie de la paroi de nos isolats dégradants (Gram+ ou Gram-),
des frottis ont été préparés par dépôt d’une goutte de culture sur lame (Marienfeld, Germany),
étalés sur 1cm2 environ avec l’anse de platine stérilisée puis séchés. La préparation a été fixée
par ajout d’une goutte d’éthanol puis flambée au bec bunsen. La coloration primaire a été
effectuée par trempage des lames 45 secondes dans le violet de gentiane phénique (Réactifs
RAL, France) puis rinçage à l’eau distillée. L’ajout du mordant a été réalisé par trempage des
préparations 45 secondes dans le lugol. Les lames ont été rapidement lavées avec le mélange
alcool éthylique - acétone (4/1, v/v) puis rincées à l’eau distillée lors de la décoloration. La
coloration secondaire a été réalisée par trempage pendant 45 secondes dans la safranine des
échantillons, qui sont ensuite rincés à l’eau distillée et délicatement tamponnés avec du papier
filtre. L’observation au microscope a été réalisée comme précédemment décrit pour les états
frais.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
99
L’observation d’un critère biochimique a été réalisée par l’utilisation de tests catalase, très
simples à mettre en place sur le plan pratique, afin de mettre en évidence la possible
accumulation de péroxyde d’hydrogène (H2O2) chez nos isolats dans le cas d’un métabolisme
aérobie. Après avoir déposé une goutte d’H2O2 sur une lame de verre, nous avons prélevé une
colonie à l’aide d’une pipette Pasteur boulée. L’ensemble a été délicatement mélangé. Dans le
cas d’une réaction positive, le dégagement gazeux est immédiat (bulles d’oxygène). Il traduit
la présence de l’enzyme qui catalyse la réaction suivante : 2 H2O2 → O2 + 2 H2O.
3- Approche moléculaire : caractérisation des isolats bactériens dégradant la sulcotrione vs isolats non dégradants 3-1- Principe et stratégie de travail
L’ADN total bactérien a été extrait à partir des souches cultivées en milieu liquide
(MS) supplémenté en sulcotrione (30 mg.L-1), afin de l’utiliser comme matrice pour les
différentes amplifications géniques envisagées lors de l’étude. Cette approche moléculaire a
consisté à caractériser les souches bactériennes isolées d’un point de vue phylogénétique et
fonctionnel.
3-2- Protocole opératoire
La totalité des travaux de biologie moléculaire sur les souches telluriques précédemment
isolées a été effectuée dans les locaux du Laboratoire Génome et Développement des Plantes
(LGDP) de l’Université de Perpignan Via Domitia (UPVD).
3-2-1- Extraction et purification de l’ADN génomique
L’ADN génomique des isolats bactériens d’intérêts a directement été extrait lors de nos
essais de biodégradation, à partir d’une aliquote de 100 µL de milieu de culture minimum
(MS) après 27 jours d’incubation (absorbance voisine de 0,7 à λ= 600 nm). Cette
méthodologie d’extraction avait déjà été décrite et utilisée avec succès par Chèneby et al.
(2004) au cours de leur étude sur la diversité des bactéries dénitrifiantes et leur activité
réductrice de l’azote au sein de la zone rhizosphérique du maïs.
L’échantillon, prélevé dans un tube Eppendorf stérile de 1,5 mL, a été centrifugé à 13 000 g
pendant 10 minutes à +4°C puis le culot a été lavé deux fois à l’aide d’eau ultra-pure stérilisée

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
100
et resuspendu dans 100 µL d’une solution stérile de Tris-Cl (5mM, pH 8,3). Les cellules
bactériennes ont ensuite été incubées avec 6,5 µL d’une solution de protéinase K (1 mg.mL-1)
pendant deux heures au bain-marie (+55°C). Afin de favoriser la lyse cellulaire et d’inactiver
la protéinase K, l’échantillon a été placé, successivement, à + 80°C pendant 10 minutes puis
immédiatement incubé 15 minutes dans la glace. Cette opération a été effectuée à trois
reprises. La solution a été centrifugée pendant 10 minutes (13 000 g, +4°C) et le surnageant a
été purifié par le kit GENECLEAN® Turbo Kit (MP Biomedicals, Europe) selon la procédure
préconisée par le fournisseur. Brièvement, après mesure du volume du surnageant
précédemment obtenu, les différents échantillons ont été placés dans des tubes stériles (1,5
mL) et 85 µL de GENECLEAN® Turbo Gnomic Salt Solution ont été ajoutés. L’ensemble a
légèrement été vortexé puis transféré (volume < 600 µL) sur une colonne de purification
(GENECLEAN® Turbo Cartdrige) et centrifugé 5 secondes à 14000 g à température ambiante.
Après ajout de 500 µL de GENECLEAN® Turbo Wash Solution, les colonnes ont été
centrifugées une première fois, 5 secondes à 14000 g (température ambiante), puis une
deuxième fois 4 minutes à 14000 g. Les échantillons ont été incubés 5 minutes à température
ambiante après ajout de 30 µL de GENECLEAN® Turbo Elution Solution. Les colonnes ont
été éluées par centrifugation 1 minute à 13000 g (température ambiante). Les échantillons ont
été stockés à -20°C pour analyses ultérieures.
3-2-2- Quantification et contrôle de la qualité de l’ADN génomique
L’ADN génomique des isolats bactériens précédemment extrait et purifié à été quantifié
par spectrophotométrie à λ = 260 nm à l’aide du Nanodrop® 2000 (ThermoFisherScientific)
disponible au laboratoire (cf figure III-2-5). Sa qualité a été vérifiée par le calcul des ratio
A260/A280 et A260/A230 ainsi que par une séparation électrophorétique sur gel d’agarose
1% (agarose molecular biology grade, Eurogentec France) après coloration par une solution
de GelRed™ 3X (Nucleic Acid Gel Stain, Biotum). Le gel a été révélé sous UV. Les
photographies ont été enregistrées sur l’imageur de gel autonome (système UGenius,
SynGene).

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
101
Figure III-2-5 : Spectrophotomètre Nanodrop (ThermoFisherScientific) utilisé pour la quantification des échantillons d’ADN génomique issus des isolats bactériens.
3-2-3- Caractérisation phylogénétique des isolats bactériens 3-2-3-1- Analyse des profils de restriction des ADNr 16S (ARDRA)
En vue d’une première approche globale de caractérisation taxonomique de nos
différents isolats bactériens, nous avons donc choisi d’utiliser cette méthode de travail. Nous
avons amplifié une portion de l’ADNr 16S de chacun de nos isolats en utilisant les amorces
universelles 27f (5’-AGA GTT TGA TCH TGG CTG CTC AG-3’) et 1492r (5’-TAC GGH
TAC CTT GTT ACG ACT T-3’) (Gurtler et Stanisich, 1996). La réaction d’amplification a
été réalisée dans un volume final de 25 µ L, contenant 2,5 µ L de tampon 5X GoTaq
Polymérase (Promega, USA), 200 µM de chaque dNTP (Promega, USA), 1 µM de chaque
amorce, 2,5 U de GoTaq DNA polymérase (Promega, USA) et 25 ng d’ADN matrice. La
réaction de PCR a été réalisée à l’aide d’un thermocycleur PTC 200 Gradient Cycler (MJ
Research, Waltham) avec le programme d’amplification suivant : 1 cycle de 4 minutes à
94°C ; 34 cycles de 1 minute à 94°C, 1 minute à 55°C, 2 minutes à 72°C et un cycle final de
15 minutes à 72°C. Afin de vérifier la qualité de la réaction d’amplification, 3 µL de produits

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
102
PCR ont été séparés par électrophorèse sur gel d’agarose à 1 % (Molecular Biology Grade
agarose, Eurogentec, Europe), placé à 100 V pendant 20 minutes dans une solution de TBE
1X. Le marqueur de taille moléculaire 1Kb Plus DNA Lader (Invitrogen) a été utilisé. Le gel
a été ensuite coloré par une solution de GelRed™ (Nucleic Acid Gel Stain, Biotum) et révélé
sous UV. Les photographies ont été enregistrées sur l’imageur de gel autonome (système
UGenius, SynGene). Une photographie des appareillages utilisés est présentée figure III-2-6.
Les produits PCR ont été finalement purifiés par le kit GENECLEAN® Turbo Kit (MP
Biomedicals, Europe) selon la procédure précédemment décrite. Le polymorphisme de
longueur de l’ADNr 16S a été révélé après digestion des produits PCR à l’aide des enzymes
de restriction Alu I et Hae III (New England Biolabs®). Deux mélanges réactionnels d’un
volume final de 20 µL ont été préparés, composés par 2 µL de tampon réactionnel 10X (New
England, Biolabs®) spécifique de l’enzyme de restriction utilisée, 1 U d’enzyme, 12 µL
d’H2O milliQ stérile et 5 µL de produits PCR. Un troisième mélange réactionnel d’un volume
final de 20 µL, contenant les deux enzymes de restriction précédentes Alu I et Hae III a
également été préparé avec 2 µL de tampon réactionnel 10X (New England, Biolabs®), 0,5
µL de chaque enzyme, 12 µ L d’H2O milliQ stérile et 5 µ L de produits PCR. Les trois
mélanges réactionnels ont ensuite été incubés à 37°C pendant 12 heures. Les fragments de
restriction obtenus après digestion enzymatique ont été séparés par électrophorèse sur un gel
d’agarose haute résolution à 2 % (MetaPhor®, Gentaur), placé à 80 V pendant 3,5 heures dans
du tampon TBE 1X. Le marqueur de taille moléculaire 1Kb Plus DNA Lader (Invitrogen) a
été utilisé. Après coloration par une solution de GelRed™ 3X (Nucleic Acid Gel Stain,
Biotum) le gel a été révélé sous UV. Les photographies ont été enregistrées sur l’imageur de
gel autonome (système UGenius, SynGene). Les profils de bandes caractéristiques ainsi
obtenus ont été comparés entre eux afin d’essayer de distinguer, d’un point de vue
taxonomique, les différentes souches bactériennes d’intérêt isolées lors de l’approche
cultivable.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
103
Figure III-2-6 : (a) Imageur de gel autonome (système UGenius, SynGene), (b) Système compact d’électrophorèse, (c) Thermocycleurs PTC 200 Gradient Cycler (Bio-Rad et MJ Research), disponibles au LGDP (UPVD). 3-2-3-2- Analyse des isolats bactériens par REP-PCR
Dans le cadre de notre étude phylogénétique, cette technique s’est avérée nécessaire afin
de pouvoir, dans la mesure du possible, différencier les profils génotypiques des souches
bactériennes précédemment isolées, comparant les différents profils REP-PCR obtenus pour
les isolats dégradants ou non dégradants. Cette stratégie de travail avait également pour but de
confirmer ou d’infirmer les premiers résultats comparatifs déjà obtenus par la technique de
l’ARDRA.
La réaction REP-PCR a été conduite en utilisant le couple d’amorce REP1R-I (5’-III ICG
ICG ICA TCI GGC-3’) et REP2-1 (5’-ICG ICT TAT CIG GCC TAC-3’) présentant des
inosines inclues dans la séquence oligonucléotidique. Ce couple d’amorces, utilisé par
Versalovic et al. (1991) lors de l’étude de la répartition des séquences REP chez les

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
104
Eubactéries, s’est avéré très efficace pour obtenir des empreintes génomiques très distinctes
chez Echerichia coli (souche W310), en comparaison à d’autres couples de séquences
oligonucléotidiques également testés. Leur utilisation dans l’étude du transfert génétique
horizontal des gènes cataboliques de l’atrazine chez Agrobacterium tumefaciens, par Devers
et al. (2005), a également confirmé leur capacité à donner de bons résultats.
La réaction d’amplification a été réalisée dans un volume final de 25 µL contenant 5 µL de
tampon 5X Giescher (cf composition tableau III-2-4), 0,16 mg.mL-1 de serum d’albumine
bovine (BSA, New England Biolabs®), 1,25 mM de chaque dNTP (Promega, USA), 0,1 µL.
mL-1 de dimethyl sulfoxide (Sigma Aldrich, France), 2 µM de de chaque amorce (REP1R-I et
REP2-1), 1 U de GoTaq DNA polymerase (Promega, USA) et 25 ng d’ADN matrice. La
réaction de PCR a été réalisée à l’aide d’un thermocycleur PTC 200 Gradient Cycler (MJ
Research, Waltham) avec le programme d’amplification suivant : 1 cycle de 6 minutes à
95°C ; 30 cycles de 1 minute à 94°C, 1 minute à 40°C, 4 minutes à 65°C et un cycle final de
16 minutes à 65°C. Les amplicons obtenus ont été séparés par électrophorèse sur un gel
d’agarose à 1 % placé à 100 V pendant 20 minutes dans du tampon TBE 1X. Le marqueur de
taille moléculaire 1Kb Plus DNA Lader (Invitrogen) a été utilisé. Après coloration par une
solution de GelRed™ 3X (Nucleic Acid Gel Stain, Biotum), le gel a été révélé sous UV. Les
photographies ont été enregistrées sur l’imageur de gel autonome (système UGenius,
SynGene). Des duplicats indépendants des différents échantillons d’ADN extraits à partir des
isolats bactériens ont été utilisés afin de s’assurer de la répétabilité de la méthode.
Solution de tampon Giescher 5X (mélange après auto-clavage
des solutions stock suivantes)
Concentration finale (mM)
16,6 mL d’une solution de (NH4)2SO4 1M 83
67,7 mL d’une solution de Tris-HCl 1M pH 8,8 335
1,3 mL d’une solution de MgCl2 1M 6,5
1,3 mL d’une solution d’EDTA 0,5 mM 0,0335
2,08 mL d’une solution commerciale de ß C2H6OS 14, 4 M 150
H2O milliQ qsp 200 mL
Répartir la solution de tampon Giescher 5X dans des tubes Eppendorf stériles (1,5 mL).
Stockage à -20°C pendant plusieurs mois.
Tableau III-2-4 : Préparation, composition et stockage du tampon Giescher 5X.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
105
3-2-3-3- Séquençage des ADNr 16S
Nous avons ensuite entrepris de séquencer certains des isolats bactériens issus du sol de
Perpignan à partir des ADNr 16S disponibles après étape préliminaire d’amplification par
PCR à l’aide des amorces universelles 27f (5’-AGA GTT TGA TCH TGG CTG CTC AG-3’)
et 1492r (5’-TAC GGH TAC CTT GTT ACG ACT T-3’). Afin de vérifier la qualité de la
réaction d’amplification, 3 µL de produits PCR ont été séparés par électrophorèse sur gel
d’agarose à 1 % (Molecular Biology Grade agarose, Eurogentec, Europe), et placés sous 100
V pendant 20 minutes dans une solution de TBE 1X. Le marqueur de taille moléculaire 1Kb
Plus DNA Lader (Invitrogen) a été utilisé. Le gel a été ensuite coloré par une solution de
GelRed™ (Nucleic Acid Gel Stain, Biotum) et révélé sous UV. Les photographies ont été
enregistrées sur l’imageur de gel autonome (système UGenius, SynGene). Les produits PCR
ont été purifiés par le kit GENECLEAN® Turbo Kit (MP Biomedicals, Europe) et quantifiés
au spectrophotomètre selon les procédures déjà décrites.
3-2-3-3-1- Ligation des produits PCR de l’ADNr 16S dans un vecteur de clonage
Les produit PCR de l’ADNr 16S générés à partir de chaque isolat bactérien d’intérêt
ont été raboutés au plasmide pGEM®-T easy (cf figure III-2-7). La réaction de ligation a
consisté à mélanger 1 µL de vecteur pGEM®-T Easy (50 ng. µL-1) avec 5 µL de tampon 2X
(Rapid Ligation buffer, T4DNA Ligase) pour l’enzyme, 1 µL de T4 DNA ligase (3 U. µL-1)
ainsi que 3 µL de produits PCR. Le mélange a été incubé pendant une nuit à 4°C.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
106
Figure III-2-7 : Carte du vecteur de clonage utilisé lors de l’étape de ligation des produits PCR, délimitant la position de l’insert d’intérêt (insert cloned) situé entre les deux promoteurs T7 et SP6 nécessaire à la transcription (plasmide pGEM®-T Easy Vector Ligation kit, Promega, Madison, USA). 3-2-3-3-2- Transformation des cellules compétentes avec le vecteur de clonage
Les plasmides obtenus lors de l’étape précédente sont introduits par transformation à
l’aide d’un choc thermique dans des cellules bactériennes ultra-compétentes Escherichia coli
DH5α™ (Invitrogen) selon le protocole suivant : Des aliquotes de 100 µ L de cellules
compétentes préalablement conservées à -80°C et placées dans des tubes Eppendorf (1,5 mL)
ont été incubées 10 minutes dans la glace puis 10 µL de produit de ligation ont été rajoutés et
l’ensemble a alors été placé 30 minutes dans la glace. Le mélange a été ensuite laissé 45
secondes au bain-marie à 42°C puis replacé à nouveau 2 minutes dans la glace. Enfin, 900 µL
de milieu Luria Bertani (LB) ont été rajoutés (cf tableau III-2-5) et les cellules transformées
ont été cultivées à 37°C sous agitation (150 rpm) pendant 1 heure. Des aliquotes de 80 et 20

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
107
µL de chaque culture ont alors été respectivement étalées sur du milieu LB gélosé contenant
de l’ampicilline (20 µg. mL-1), du X-Gal (40 µg. mL-) et de l’IPTG (32 µg.mL-1). Les boites
ont finalement été placées une nuit à l’étuve à 37°C.
Le plasmide pGEM®-T easy porte le gène de résistance à l’ampicilline et va donc servir de
crible de sélection pour les cellules bactériennes qui ont été transformées, c’est à dire qui ont
intégré le plasmide. De plus, ce vecteur porte également un gène codant pour l’enzyme ß
galactosidase à l’intérieur duquel se situe le site de clonage (compris entre les deux
promoteurs SP6 et T7), où peut venir se positionner l’insert d’intérêt. La présence ou
l’absence de l’insert d’intérêt dans cette zone permet ou non de sélectionner les bactéries
recombinantes en imposant un crible de sélection supplémentaire. En effet, le raboutage d’un
produit PCR au site de clonage cause dans ce cas la disruption du gène codant pour l’enzyme
ß galactosidase, entrainant nécessairement la perte de capacité de la souche recombinante à
dégrader le X-Gal et conduisant à la coloration bleue de la colonie. In fine, les colonies
bactériennes se développant sur milieu LB contenant de l’ampicilline ont donc bien intégré le
plasmide pGEM®-T easy, et parmi ces colonies, seules celles qui apparaissent blanches
possèdent un plasmide recombinant (cf figure III-2-8). Pour chaque isolat bactérien, 20
colonies blanches ont été sélectionnées à partir des boites gélosées de milieu LB +
Ampicilline, placées dans 100 µ L d’eau ultra-pure préalablement filtrée (filtres stériles
Acrodisc® 32, Supor® 0,2 µ m, GelmanSciences) et également repiquées sur boite LB +
ampicilline en vue de leur conservation (12 heures à 37°C puis placées à 4°C).
Tableau III-2-5 : Préparation et composition du milieu Luria Bertani (LB) nécessaire à la sélection des cellules bactériennes transformées par le plasmide pGEM®-T easy.
Elément Quantité (g)
Bacto-tryptone (1 %) 10
Extrait de levure (0,5 %) 5
NaCl (0,5 %) 5
Agar (1,5%) si milieu gélosé 16
H2O purifiée qsp 1 litre
Après autoclave, ajouter pour 1 litre ampicilline (20 µg.mL-1 final) 400 µL
Ajouter extemporanément (par boite, environ 20 mL) :
IPTG (32 µg.mL-1 final)
X-Gal (32 µg.mL-1 final)
3,5
40 µL

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
108
Figure III-2-8 : Croissance des colonies après étalement de 20 µL (gauche) et de 80 µL (droite) de cellules compétentes Escherichia coli DH5α™ ayant intégré le vecteur de clonage pGEM®-T easy sur milieu LB + ampicilline (20 µg.mL-1).
3-2-3-3-3- Test PCR sur colonies des clones recombinants
Ce test a consisté à confirmer par PCR, que les colonies blanches sélectionnées lors de
l’étape précédente, présentent bien le plasmide pGEM®-T easy contenant l’insert d’intérêt
(ADNr 16S des isolats bactériens) et à éviter de sélectionner des « faux positifs » avant
l’étape de séquençage des clones recombinants.
Une amplification PCR a donc été réalisée à partir des colonies blanches sélectionnées en
utilisant des amorces situées de part de d’autre du site d’insertion et composées par les
séquences des deux promoteurs SP6 et T7 du plasmide. Le mélange réactionnel a été effectué
dans un volume total de 25 µL et composé par 2,5 µL de tampon 5X GoTaq Polymérase
(Promega, USA), 200 µM de dNTP, 0,5 µM de chaque amorce (SP6 et T7), 2,5 U de GoTaq
DNA polymerase (Promega, USA) et 2,5 µL de suspension bactérienne. La réaction a été
réalisée dans un thermocycleur PTC 200 Gradient Cycler (MJ Research, Waltham) avec le
programme d’amplification suivant: 3 minutes à 94°C ; suivi de 30 cycles de 30 secondes à
94°C, 45 secondes à 55°C, 2 minutes à 72°C, et enfin un cycle d’élongation de 5 minutes à
72°C. Les amplicons obtenus ont été séparés et révélés comme décrit précédemment.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
109
Les colonies blanches sélectionnées ayant donné, après migration électrophorétique, des
fragments de taille supérieure à 3000 paires de bases (taille du plasmide pGEM®-T easy de
3015 pb) ont finalement été retenues et sélectionnées comme étant des clones recombinant
présentant le plasmide avec l’insert d’intérêt.
En vue de la préparation ultérieure des échantillons pour la réaction de séquençage, les
colonies correspondantes ont été ensemencées à partir de 6 mL de milieu liquide LB
complémenté en ampicilline (20 µg. mL-1) pendant une nuit à 37°C.
3-2-3-3-4- Préparation de l’ADN plasmidique (MiniPrep)
Une extraction de l’ADN plasmidique des clones recombinants ayant poussé pendant
une nuit à 37°C sur milieu LB complémenté d’ampicilline (colonies blanches positives après
la réaction de PCR avec les amorces T7 et SP6) a été réalisée à l’aide du kit commercial
Wizard® Plus Minipreps DNA Purification System (Promega, USA). Les cultures
ensemencées ont été placées dans des tubes Falcon stériles (10 mL) numérotés et centrifugées
en salle de culture P2 à 3000 rpm pendant 10 minutes à 4°C. Le surnageant obtenu a été
éliminé et le culot repris par 250 µL de solution de resuspension (E1) fournie par le kit. 250
µL de solution de lyse (E2) ont été rajoutés et les tubes ont été mélangés par inversion (4
fois), puis 10 µL de solution de protéase alkaline (E3) ont été rajoutés et les échantillons ont
été homogénéisés et laissés 5 minutes à température ambiante. 350 µ L de solution de
neutralisation (E4) ont été finalement été rajoutés, les échantillons homogénéisés (inversion 4
fois) et centrifugés à 13 000 rpm pendant 10 minutes à température ambiante. La totalité du
surnageant de chaque échantillon a été déposée sur spin colonne (Kit) et centrifugé à 13 000
rpm pendant 1 minute à température ambiante. Les colonnes ont été lavées deux fois (solution
E5, ajout de 750 µL puis de 250 µL) et centrifugées à nouveau à 13 000 rpm pendant 2
minutes. Les colonnes ont finalement été éluées par ajout de 50 µ L d’H2O ultra-pure
(solution E6) et centrifugation à 13000 rpm pendant 1 minute. L’ADN plasmidique ainsi
obtenu a été quantifié et sa qualité a été vérifiée comme précédemment décrit.
3-2-3-3-5- Réaction de séquençage
Les amplifiats de PCR SP6/T7, issus de la préparation plasmidique précédente, ont été utilisés
pour réaliser les réactions de séquençage. Cela consiste à réaliser l’élongation de l’insert cloné

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
110
par une PCR linéaire à l’aide d’une seule des deux amorces SP6 ou T7, en présence d’un
mélange des 4 di-désoxy nucléotides triphosphates (ddNTPs) marqués par un colorant
spécifique à chaque base. Dans le cadre de notre étude, chaque amplifiat provenant des
différents clones recombinants sélectionnés a été systématiquement amplifié en utilisant les
deux amorces séparément, en utilisant soit SP6, soit T7 dans le mélange réactionnel. Les
échantillons ont été préparés dans un volume final de 10 µL composé de 1 µL d’une des
amorces SP6 ou T7 (concentration initiale des amorces 10 pmole. µL-1), et d’un volume de
produit PCR de façon à obtenir 100 ng d’ADN dans le mélange réactionnel complété par
ajout d’eau ultra-pure (qsp 10 µL). Les échantillons ont été placés dans un thermocycleur
PTC 200 Gradient Cycler (MJ Research, Waltham) afin d’être déshydratés par chauffage à
80°C pendant 30 minutes puis repris par 20 µL de solution de ddNTPs BigDye® Terminator
v3.1 Cycle Sequencing Kits (Applied Biosystem, Fostercity, USA). Le programme
d’amplification suivant a été conduit : 5 minutes à 92°C puis 35 cycles de 30 secondes à
92°C, 20 secondes à 50°C (diminution de 1°C.seconde-1) et 4 minutes à 60°C (augmentation
de 1°C.seconde-1). La réaction d’amplification a été arrêtée par l’ajout de 75 µ L d’une
solution d’arrêt (2 mL d’acétate de sodium 3 M pH 4,5 + 48 mL d’éthanol à 95°, qsp 50 mL).
Les échantillons ont été agités au vortex et centrifugés à 3250 rpm pendant 40 minutes à 4°C.
Le surnageant a été éliminé et le culot d’ADN délicatement séché (centrifugation à 3250 rpm
pendant 30 secondes à 4°C puis égouttage). Le culot a été lavé deux fois avec 70 µL d’éthanol
à 70 % préalablement placé à -20°C puis l’ensemble est centrifugé à 3250 rpm pendant 15
minutes à 4°C. Le surnageant a été éliminé et le culot a été séché sur la paillasse puis remis en
suspension dans 20 µL d’H2O ultra-pure. Les échantillons ont été chargés dans une plaque 96
puits et analysés par un séquenceur à 16 capillaires de type Applied Biosystems Hitachi
3130xl Genetic Analyzers (cf figure III-2-9). Les données brutes générées par le séquenceur
ont été traitées à l’aide du logiciel Data Collection Software (V 3.0, Applied Biosystems,
USA) qui permet de générer un spectre de séquençage et d’aboutir à la détermination de la
séquence nucléotidique de la matrice ADN étudiée.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
111
Figure III-2-9 : Séquenceur Applied Biosystems Hitachi 3130xl Genetic Analyzers.
3-2-3-3-6- Traitement et analyse des séquences
Les séquences nucléotidiques d’ADNr 16S obtenues ont été éditées par le logiciel
4PEAKS V1.7 (http://www.mekentosj.com/science/4peaks). L’inversion des séquences
complémentaires a été réalisée à l’aide du logiciel Reverse Complement
(http://www.bioinformatics.org/sms/rev_comp.html). Des séquences consensus ont été ensuite
générées grâce au logiciel SeqManNGen® (DNASTAR, USA) à partir de nos données et
comparées aux séquences d’ADNr 16S connues et disponibles dans la banque GenBank
(http://blast.ncbi.nlm.nih.gov/Blast.cgi) par le programme BlastN (Altschul et al., 1990).
L’alignement multiple des séquences consensus d’ADNr 16S des clones et des séquences
sélectionnées dans GenBank (http://www.ncbi.nlm.nih.gov/genbank/) a été réalisé avec le
logiciel ClustalX (http://www-igbmc.u-strasbg.fr/BioInfo/; Thompson et al., 1997). La
matrice de similitude générée a été traitée par le logiciel NJPlot (ftp:://pbil.univ-

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
112
lyon1.fr/pub/mol_phylogeny/njplot; Perrière et Gouy, 1996) afin de reconstruire l’arbre
phylogénétique correspondant par la méthode du Neighbor-Joining (Saitou et Nei, 1987)
assignée de la correction de Jukes et Cantor (1969). Une analyse par boostrap (Felsentein,
1985 ; Felsentein et Kishino, 1993) a été menée pour tester la robustesse de notre arbre.
3-2-4- Caractérisation fonctionnelle des isolats bactériens : 3-2-4-1- Contexte
Nous avons voulu compléter l’identification phylogénétique de nos isolats par la
détection de deux familles de gènes impliqués dans la dégradation de xénobiotiques
aromatiques par action de métalloenzymes dioxygénases bactériennes. L’importance du
carrefour métabolique que semble constituer la voie des catéchols ou protocatéchuates laisse à
penser que les populations bactériennes ont progressivement développé des enzymes capables
de reconnaître des gammes étendues de substrat, afin de pouvoir transformer des composés
aromatiques de familles différentes en métabolites de nature catécholique (Dagley, 1986 ;
Vaillancourt et al., 2006 ; Zeyaullah et al., 2009 ; Lillis et al., 2010).
Les dioxygénases catalysent deux réactions fondamentales: en premier lieu,
l’hydroxylation du composé aromatique, puis dans une deuxième étape, l’ouverture du noyau
(Williams and Sayers, 1994 ; Hatta et al., 2003 ; Costas et al., 2004 ; Prathibba et Sumathi,
2007). Ces enzymes sont classées selon le mode d’ouverture du noyau aromatique qu’elles
catalysent : l’intradiol dioxygénase ou catéchol 1,2-dioxygénase (C12O) lorsque le clivage se
fait en position ortho (clivage entre les carbones porteurs des groupements hydroxyles) ou
l’extradiol dioxygénase ou catéchol 2,3-dioxygénase (C23O) pour clivage en position méta
(clivage entre les deux carbones adjacents au diol). Ces deux types d’enzymes constituent
deux familles distinctes et différent notamment par la valence de l’atome de fer situé au site
actif : les intradiol dioxygénases présentent un fer ferrique alors que les extradiol
dioxygénases possèdent un fer ferreux (Harayama et Rekik, 1989). La figure III-2-10
représente le mécanisme général de ces deux types d’enzymes.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
113
Figure III-2-10 : Voies de clivage du noyau aromatique du catéchol (voies ortho et méta) catalysées par les catéchol dioxygénases.
La première dioxygénase de ce type à avoir été séquencée est la catéchol 2,3-
dioxygénase, codée par le gène xylE sur le plasmide TOL (Nakai et al., 1983). Cette enzyme
catalyse l’oxydation du catéchol en acide 2-hydroxymuconique semialdéhyde. Les gènes XylE
sont constitués d’environ 923 pb correspondantes à la totalité du cadre de lecture ouvert du
gène de la catéchol-2,3-dioxygénase qui est très largement répandue dans le monde microbien
et se trouve impliquée dans des métabolismes variés (Eltis et Bolin, 1996).
Ces gènes sont potentiellement d’excellentes cibles moléculaires pour caractériser et typer la
microflore du sol impliquée dans la dégradation des composés xénobiotiques aromatiques
(Wikström et al., 1996). Ils sont généralement portés par un plasmide catabolique conjugatif,
le plasmide TOL pWW0 d’environ 117 kb (Nakai et al., 1983), organisés en opéron et se
localisent à l’intérieur d’éléments transposables (Assinder et Williams, 1990). Nous avons
recherché leur présence chez nos isolats.
Nous avons également recherché la présence des gènes codant l’enzyme catéchol-1,2-
dioxygénase (C12O) dans nos isolats. Même si la nature chimique de la sulcotrione semble
certainement incompatible avec un mécanisme de type ortho clivage, il était tout de même
important d’avoir une vision fonctionnelle élargie de nos isolats vis-à-vis de ces deux
enzymes clefs du métabolisme bactérien. Les gènes C12O sont composés d’environ 920 pb et
codent l’enzyme catéchol-1,2-dioxygénase (C12O) qui intervient également dans des
métabolismes impliquant des composés aromatiques variés (Strachan et al., 1998 ; Giedraityte
et Kalediene 2009).

114
3-2-4-2- Détection par PCR des gènes XylE codant l’enzyme catéchol 2,3-dioxygénase
Nous avons appliqué la procédure décrite par Jussila et al. (2007). Ces auteurs ont développé
une méthode de détection gène-spécifique du plasmide TOL par amplification PCR d’environ
731 pb des gènes XylE (cf tableau III-2-6) à partir des amorces consensus XylEaf (5’-TCG
AGT TGC TGG GCC TGA TCG-3’) et XylEar (5’- CCC GCA GAA CAC TTC GTT GCG-
3’). La réaction d’amplification a été réalisée dans un volume final de 25 µL, contenant 2,5
µL de tampon 5X GoTaq Polymérase (Promega, USA), 200 µM de chaque dNTP (Promega,
USA), 1 µM de chaque amorce, 2,5 U de GoTaq DNA polymérase (Promega, USA) et 25 ng
d’ADN matrice. Les amplifications ont été réalisées à partir d’ADN génomique de nos isolats.
Les deux contrôles utilisés ont été de l’ADN génomique d’Escherichia coli souche DH5α
[génotype F- φ80lacZ∆M15 ∆(lacZYA-argF)U169 deoR recA1 endA1 hsdR17(rk-, mk
+)
phoA supE44 thi-1 gyrA96 relA1 λ-] obtenu après culture de 100 µ L de suspension
bactérienne sur milieu LB (12 heures à 37°C) et extraction-purification à l’aide du kit
Wizard® Genomic DNA Purification kit (Promega, USA) ainsi que le plasmide pCambia
3300 (taille 9kb, Flag-HA /3300 : CTZ, création juin 2010, source LGDP, Equipe Nouveaux
Systèmes Transcriptionnels chez Arabidopsis thaliana). La réaction de PCR a été réalisée à
l’aide d’un thermocycleur PTC 200 Gradient Cycler (MJ Research, Waltham) avec le
programme d’amplification suivant : 1 cycle de 10 minutes à 94°C ; 30 cycles de 30 secondes
à 94°C, 1 minute à 70°C, 1 minute à 72°C et un cycle final de 3 minutes à 72°C. 8 µL de
produit PCR ont été déposés sur gel d’agarose à 1,2 % (Molecular Biology Grade agarose,
Eurogentec, Europe) et séparés par électrophorèse à 80 V pendant 35 minutes dans une
solution de TBE 1X. Le marqueur de taille moléculaire 1Kb Plus DNA Lader (Invitrogen) a
été utilisé. Le gel a été ensuite coloré par une solution de GelRed™ (Nucleic Acid Gel Stain,
Biotum) et révélé comme décrit précédemment. Les bandes obtenues ont été excisées sous
UV et purifiées par le kit GENECLEAN® Turbo Kit (MP Biomedicals, Europe) selon la
procédure précédemment décrite. 3 µL de chaque échantillon d’ADN ont été quantifiés sur
gel d’agarose 1 % (Molecular Biology Grade agarose, Eurogentec, Europe) avec 1, 2 et 5 µL
de solution de marqueur de taille moléculaire 1Kb Plus DNA Lader (Invitrogen). Les
échantillons ont également été dosés par spectrophotométrie à λ = 260 nm à l’aide du
Nanodrop® 2000 (ThermoFisherScientific) et leur qualité a été évaluée (rapports A 260/280 et
A 260/230). Les échantillons ont été conservés à -20°C pour analyses ultérieures.

115
Amorce Direction Long. (nt)
Temp. (°C)
G + C (%)
Position 5’de la séquence codante
Séquence Gène amplifié
Taille du fragment attendu
(pb)
Références
XylEaf
Foward
21
70,1
62
127
5’- TCG AGT TGC TGG GCC TGA TCG -3’
XylEar
Reverse
21
70,3
62
857
5’- CCC GCA GAA CAC TTC GTT GCG-3’
C23O 731
Jussila et al. (2007)
Nakai et al. (1983)
C12Of Foward
20 -
65
-
5’- GCC AAC GTC GAC
GTC TGG CA-3’
C12Or Reverse 25 - 52 -
5’- CGC CTT CAA AGT TGA ATC TGC GTG
GT-3’
C12O 282
Sei et al. (1999)
Guzik et al. (2011)
Tableau III-2-6 : Caractéristiques des amorces utilisées pour la détection des gènes codant la catéchol 2,3-dioxygénase (C23O) et la catéchol 1,2-dioxygénase (C12O) par amplification PCR.

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
116
3-2-4-3- Détection par PCR des gènes C12O codant l’enzyme catéchol 1,2-dioxygénase
Nous avons appliqué la procédure décrite par Sei et al. (1999) et reprise par Guzik et
al. (2011). Ces auteurs ont dessiné des amorces spécifiques (cf tableau III-2-6) à partir de 11
régions fortement homologues des gènes C12O par alignement multiple des séquences des
gènes C12O disponibles dans GenBank. Leur objectif était de pouvoir amplifier une large
gamme de gènes codant les catéchol 1,2-dioxygénases des genres bactériens suivants :
Pseudomonas, Acinetobacter, Alcaligenes et Arthrobacter. Les amorces C12Of (5’-GCC
AAC GTC GAC GTC TGG CA-3’) et C12Or (5’- CGC CTT CAA AGT TGA TCT GCG
TGG T-3’) amplifient environ 300 pb du gène C12O.
La réaction d’amplification a été réalisée dans un volume final de 25 µL, contenant 2,5 µL de
tampon 5X GoTaq Polymérase (Promega, USA), 200 µM de chaque dNTP (Promega, USA),
1 µM de chaque amorce, 2,5 U de GoTaq DNA polymérase (Promega, USA) et 25 ng d’ADN
matrice. Les amplifications ont été réalisées à partir d’ADN génomique de nos isolats. Les
deux contrôles utilisés ont été de l’ADN génomique d’Escherichia coli souche DH5α et le
plasmide pCambia 3300 décrits précédemment. La réaction PCR a été réalisée à l’aide d’un
thermocycleur PTC 200 Gradient Cycler (MJ Research, Waltham) avec le programme
d’amplification suivant : 1 cycle de 3 minutes à 94°C ; 10 cycles de 1 minute à 94°C, 30
secondes à 61°C, 30 secondes à 72°C, 15 cycles de 1 minute à 94°C, 30 secondes à 59°C, 30
secondes à 72°C, 15 cycles de 1 minute à 94°C, 30 secondes à 57°C, 30 secondes à 72°C et
un cycle final de 30 secondes à 72°C. 10 µL de produit PCR ont été déposés sur gel d’agarose
à 1% (Molecular Biology Grade agarose, Eurogentec, Europe) et séparés par électrophorèse à
80 V pendant 35 minutes dans une solution de TBE 1X. Le marqueur de taille moléculaire
1Kb Plus DNA Lader (Invitrogen) a été utilisé. Le gel a été ensuite coloré par une solution de
GelRed™ (Nucleic Acid Gel Stain, Biotum) et révélé comme décrit précédemment. Les
bandes obtenues ont été excisées sous UV et purifiées par le kit GENECLEAN® Turbo Kit
(MP Biomedicals, Europe) selon la procédure précédemment décrite. 3 µ L de chaque
échantillon d’ADN ont été quantifiés sur gel d’agarose 1 % (Molecular Biology Grade
agarose, Eurogentec, Europe) avec 1, 2 et 5 µL de solution de marqueur de taille moléculaire
1Kb Plus DNA Lader (Invitrogen). Les échantillons ont également été dosés par
spectrophotométrie à λ = 260 nm à l’aide du Nanodrop® 2000 (ThermoFisherScientific) et

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
117
leur qualité a été évaluée (rapports A 260/280 et A 260/230). Les échantillons ont été
conservés à -20°C pour analyses ultérieures.
3-2-4-4- Séquençage des produits PCR
Les échantillons ont été préparés afin de séquencer les fragments d’ADN dans les
deux sens (5’3’ et 3’5’). Ils ont été composés d’un volume final de 8 µL comprenant 1
µL d’amorces XylEaf ou XylEar (séquençage du gène XylE), soit 1 µL de d’amorces C12Of
ou C12Or (séquençage du gène C12O) de concentration de 10 pmol. µL-1 et d’un volume
d’ADN matrice variable selon la taille des fragments considérés (cf tableau III-2-7). Le
volume final a été complété par ajout d’eau ultra-pure (qsp 8 µL). La réaction de séquençage
s’est déroulée dans les conditions décrites précédemment.
Taille des fragments d’ADN
(pb)
Quantité d’ADN par réaction de séquençage
(ng)
300 7 à 14
500 à 700 9 à 18
1000 16 à 32
2000 28 à 56
Tableau III-2-7 : Quantité d’ADN (en ng) nécessaire par réaction de séquençage pour les fragments PCR. Etalonnage du séquenceur Applied Biosystems Hitachi 3130xl Genetic Analyzers.
3-2-4-5- Traitement et analyse des séquences
Les séquences nucléotidiques d’ADN obtenues ont été éditées par le logiciel FinchTV
version 1.4 (http://www.geospiza.com/Products/finchtv.shtml). L’alignement multiple entre
les séquences éditées et les séquences des gènes XylE et C12O disponibles dans GenBank
(http://www.ncbi.nlm.nih.gov/genbank/) été réalisé avec le logiciel ClustalX (http://www-
igbmc.u-strasbg.fr/BioInfo/; Thompson et al., 1997).

Troisième partie – Chapitre 2 : Méthodologie de lʼanalyse biologique
118
3-2-5- Caractérisation physiologique des isolats
En complément de l’étude moléculaire des souches bactériennes isolées, leur aptitude
à dégrader la sulcotrione a été évaluée. Cette partie de notre travail a été décrite
précédemment.

119
Quatrième partie: Résultats et discussion

Quatrième partie – Chapitre 1 : Confirmation du pouvoir biodégradant du sol de Perpignan et recherche dʼune éventuelle biodégradation accélérée (BDA)
120
Chapitre1‐ConfirmationdupouvoirbiodégradantdusoldePerpignanetrecherched’uneéventuellebiodégradation
accélérée(BDA)
1- Réactivité biologique du sol de Perpignan Les travaux précédemment réalisés au L.C.B.E par Chaabane (2005) et Chaabane et
al. (2005, 2008) ont permis de mettre en évidence la dégradation des herbicides tricétones (et
notamment la sulcotrione) dans les quatre sols étudiés (Belgique, Martinique, Landes et
Perpignan). Outre une dégradation effective par voie abiotique (hydrolyse et photolyse), cet
auteur a montré le rôle prépondérant de la dégradation biologique. Ces résultats ainsi que la
disponibilité du sol de Perpignan et les traitements qu’il avait reçus au préalable nous ont
amené à définir de nouveaux objectifs de travail : (i) confirmer le pouvoir biodégradant de ce
sol de Perpignan; (ii) rechercher une éventuelle biodégradation accélérée (BDA) de la
sulcotrione, en confrontant nos résultats aux données bibliographiques relatives à d’autres
substances actives (atrazine, isoproturon, 2,4-D,…); (iii) isoler puis caractériser des souches
bactériennes, présentes dans le sol de Perpignan et impliquées dans le processus de
dégradation biologique.
Lors des précédents travaux de H. Chaabane, la microflore du sol de Perpignan avait
montré une capacité à transformer la sulcotrione en deux métabolites majeurs, la 1,3-
cyclohexanedione (CHD) et l’acide 2-chloro-4-mésylbenzoïque (CMBA) par clivage entre la
partie CHD et le noyau benzénique. Nous avons souhaité confirmer cette potentialité
biodégradante en conditions contrôlées par l’utilisation de microcosmes.
Deux types de sols issus de la parcelle ont permis de préparer les différents microcosmes : un
sol (4S), ayant déjà subi quatre traitements au champ, deux ans plus tôt et un sol « contrôle »
(C) exempt de tout traitement préalable par la sulcotrione. Parallèlement, des essais similaires
ont été menés sur ces sols (4S) et (C) préalablement stérilisés (St4S) et (StC) (cf paragraphe
II-2-4).

Quatrième partie – Chapitre 1 : Confirmation du pouvoir biodégradant du sol de Perpignan et recherche dʼune éventuelle biodégradation accélérée (BDA)
121
Un premier « traitement » a été effectué sur l’ensemble des microcosmes (boites de Pétri
contenant 10 grammes de sol) par ajout de 0,6 mL de solution aqueuse de sulcotrione à 22
mg.L-1, se rapprochant ainsi de la teneur prévisible dans le sol (en µg.g -1 de sol dans les
premiers centimètres) après application aux doses homologuées lors des itinéraires techniques
maïsicoles (300-450 g.Ha-1).
Les cinétiques de dégradation ont été établies pour les quatre sols étudiés (4S), (C), (St4S) et
(StC) après 30 jours d’incubation dans une chambre thermostatée à 24°C (± 0,2). Les résultats
sont présentés dans le tableau IV-1-1 et les figures IV-1-1, IV-1-2). La biodégradation de la
sulcotrione est donc confirmée sur les sols (4S) et (C) conformément aux travaux présentés
par H. Chaabane (2005). Le sol de Perpignan présente une cinétique de dégradation qui peut
raisonnablement être décrite par une loi de premier-ordre avec un coefficient de détermination
r² voisin de 0,9 dans les deux cas:
Ln C = - kt + Ln C0
Avec C : concentration de pesticide restante en µg.g-1 de sol,
C0 : concentration initiale de pesticide présente dans le milieu en début d’incubation en
µg.g-1 de sol,
k : constante de cinétique de premier-ordre.
Le temps de demi-vie (t1/2), ou encore temps nécessaire pour qu’il y ait disparition de la
moitié de la quantité initiale de pesticide est calculé à partir de la relation précédente:
t1/2 = Ln 2 / k

Quatrième partie – Chapitre 1 : Confirmation du pouvoir biodégradant du sol de Perpignan et recherche dʼune éventuelle biodégradation accélérée (BDA)
122
Tableau IV-1-1 : Constantes cinétiques (k), temps de demi-vie (t1/2) et coefficients de détermination (r2) relatifs à la cinétique de dégradation de la sulcotrione en conditions de microcosmes pour le sol de Perpignan.
Figure IV-1-1 : Suivi de la dissipation de la sulcotrione dans les microcosmes de sol (4S) vs (St4S) après un, deux, trois ou quatre traitements successifs.
k (jours-1)
t1/2 (jours)
r2
Sol (C)
Traitement 1 (T1D0)
0,083
8
0.894
Sol (4S)
Traitement 1 (T1D0)
0,088
8
0.872
Sol (StC)
Traitement 1 (T1D0)
0,0004
-
-
Sol (St4S)
Traitement 1 (T1D0)
0,0004
-
-

Quatrième partie – Chapitre 1 : Confirmation du pouvoir biodégradant du sol de Perpignan et recherche dʼune éventuelle biodégradation accélérée (BDA)
123
Figure IV-1-2 : Suivi de la dissipation de la sulcotrione dans les microcosmes de sol (C) vs (StC) après un, deux, trois et quatre traitements successifs.
Après 30 jours d’incubation, environ 90 % de la sulcotrione initiale a été dégradée aussi bien
pour le sol (4S) que pour le sol (C).
Les constantes de dégradation (k ≈ 0,085) ont été voisines pour les deux sols, permettant
d’estimer des temps de demi-vie voisins de 8-10 jours, sensiblement plus courts que ceux
présentés par H. Chaabane (2005 ; Chaabane et al., 2007), mais compris dans « la fourchette »
de quelques jours à environ 60 jours, retrouvée dans d’autres données bibliographiques
(Rouchaud et al., 1996 ; 1998a, 1998b ; Cherrier et al., 2004 ; Mamy, 2005).
Aucune différence notable n’est donc apparue entre les sols (4S) et (C), malgré leur historique
de traitement volontairement différent. Ce résultat n’autorise donc pas à évoquer, à l’issue de
cette première étape, un possible « effet mémoire » sur le potentiel de dégradation du sol (4S),
lié à la répétition antérieure (36 mois auparavant) des traitements en plein champ,.
De plus, pour les deux sols, aucune phase de latence n’a été observée lors de la cinétique de
disparition, élément permettant de suggérer l’intervention d’un processus de co-métabolisme
dans la dégradation de la sulcotrione.

Quatrième partie – Chapitre 1 : Confirmation du pouvoir biodégradant du sol de Perpignan et recherche dʼune éventuelle biodégradation accélérée (BDA)
124
Les résultats relatifs à l’étude menée à partir des sols stérilisés (St4S) et (StC) sont également
présentés dans le tableau VI-1-1. La dégradation a été pratiquement négligeable, aussi bien
pour le sol (St4S) que pour le sol (StC), dans les conditions de stérilisation retenues.
Ces données confirment donc que la voie biologique est largement prépondérante dans le cas
de la sulcotrione dans le sol de Perpignan en accord avec les précédents résultats décrits par
H. Chaabane.
2- Recherche d’une éventuelle biodégradation accélérée (BDA)
Consécutivement à l’utilisation répétée de produits phytosanitaires sur de longues
périodes, une baisse progressive de l’efficacité de la substance active a été souvent décrite
dans la littérature, généralement associée à une adaptation de la microflore tellurique
impliquée dans la dissipation du pesticide dans le sol. Mise en évidence par Audus à partir des
années 50 avec le 2,4 D (Audus, 1964), la BDA est sans conséquence sur les produits à
application foliaire. Certaines familles de pesticides apparaissent plus concernées que d’autres
par ce phénomène. C’est notamment le cas des carbamates, qu’il s’agisse d’insecticides
comme le carbofuran (Felsot et al., 1981), le bendicarb et la carbaryl (Racke et Coats, 1988a)
ou d’herbicides comme le butylate et le vernolate (Gray et Joo, 1985). Une BDA a été
également observée pour certains insecticides organophosphorés comme l’isofenphos, le
fonofos et le fénamiphos par exemple (Chung et Ou, 1996) et des fongicides comme
l’iprodione et la vinchlozoline (Walker et al., 1986). Gray et Joo (1985) ont également montré
l’apparition de ce mécanisme chez les amides (napropamide), les dérivés amides (alachlore),
les benzamides (propizamide) et chez certains acides halogénés comme le dalapon. Bien sûr
dans les familles d’herbicides comme les s-triazines, ce phénomène a été largementt mis en
évidence (Barriuso et Houot , 1996 ; Pussemier et al., 1997 ; Yassir et al., 1999). Dans le cas
des urées substituées, il intéressant de voir que la BDA n’est pas systématique pour tous les
composés d’une même famille : ainsi l’isoproturon, le linuron et le chlortoluron peuvent être
sujets à la BDA alors que le diuron présente par exemple une BDA « plus modérée » malgré
une nombre de traitements importants pendant plusieurs années (Rouchaud et al., 2000).
Ainsi, l’importance de la structure chimique du composé phytosanitaire semble jouer un rôle
sur le développement d’une BDA au sein d’une parcelle agricole. En outre, ces processus

Quatrième partie – Chapitre 1 : Confirmation du pouvoir biodégradant du sol de Perpignan et recherche dʼune éventuelle biodégradation accélérée (BDA)
125
adaptatifs ont été généralement décrits pour des composés phytosanitaires, le plus souvent
herbicides, utilisés depuis plusieurs décennies à des doses d’application supérieures à celles
homologuées pour la sulcotrione, notamment en maïsiculture.
Aussi, il nous a semblé intéressant de rechercher l’existence d’un tel phénomène de BDA
dans le cas de la sulcotrione, en raison (i) de son « avenir agronomique » au sein des
itinéraires techniques en remplacement de l’atrazine, interdite au début des années 2000, (ii)
de sa structure chimique présentant un substituant halogéné (généralement décrits comme des
composés chimiques plutôt récalcitrants), (iii) des doses d’application relativement modérées.
A l’issue du premier traitement en microcosme, la probabilité d’apparition d’une BDA est
apparue relativement faible au vu des résultats relatifs aux cinétiques de dégradation,
comparables entre les sols (4S) et (C). En raison de la réactivité de ces sols, nous avons
cependant souhaité poursuivre cette étude par des traitements supplémentaires au niveau des
mêmes microcosmes. Un calendrier a été établi (cf paragraphe II-2-5). Il prévoyait un
deuxième traitement 30 jours après le premier et des prélèvements consécutifs au cours du
mois suivant. Deux traitements supplémentaires, espacés chacun de 30 jours et réalisés dans
les mêmes conditions, ont complété cette étude.
Les résultats sont présentés dans le tableau IV-1-2 ainsi que dans les figures IV-1-1 et IV-1-2.
L’examen de ces résultats, obtenus après répétition des traitements suscite plusieurs
commentaires :
(i) Les résultats des cinétiques de dissipation de l’herbicide ont été similaires sur les sols (4S)
et (C), ce qui confirme bien l’absence d’effet mémoire entre ces sols, différents au niveau de
l’historique phytosanitaire, absence déjà entrevue lors du premier traitement.
(ii) L’examen statistique (moyenne ± écart-type) des valeurs de k obtenues à l’issue de chaque
épisode (traitement et prélèvements au cours du mois d’incubation) montre une valeur
moyenne voisine de 0,085 (avec un coefficient de variation acceptable, voisin de 20 %), tout à
fait comparable à celle obtenue après le premier traitement. Le temps de demi-vie (t1/2), invariable, reste au voisinage de 8-10 jours. La répétition des traitements n’affecte donc pas la
vitesse de dégradation de la sulcotrione dans le sol de Perpignan : la vitesse n’augmente pas

Quatrième partie – Chapitre 1 : Confirmation du pouvoir biodégradant du sol de Perpignan et recherche dʼune éventuelle biodégradation accélérée (BDA)
126
au cours du temps, caractéristique cinétique essentielle du processus de dégradation par co-
métabolisme. De plus, après le premier traitement, et quels que soient les traitements
successifs (T2, T3 et T4) et le type de sol considérés, les courbes de dissipation de l’herbicide
ne font pas apparaître de temps de latence. Au contraire, la cinétique s’apparente plutôt à une
transformation chimique catalysée ne présentant pas de point d’inflexion, signature d’une
dégradation biologique par co-métabolisme.
(iii) L’examen des courbes de dissipation ainsi que les coefficients de détermination souvent
supérieurs à 0,9 corroborent le caractère de cinétique de premier-ordre, quel que soit le
nombre de traitements.
Tableau IV-1-2 : Paramètres cinétiques relatifs aux différents traitements successifs effectués
en microcosmes sur les sols (4S), (St4S), (C) et (StC).
k (jours-1) t1/2 (jours) r2
Traitement 1 (T1D0)
0,083
8
0,894 Traitement 2 (T2D30) 0,060 11,5 0,955
Sol(C) Traitement 3 (T3D60) 0,100 7 0,989 Traitement 4 (T4D90) 0,100 7 0,962
Moyenne
0,085 ± 0,020
8 ± 2,5
-
Traitement 1 (T1D0)
0,088
8
0,872
Traitement 2 (T2D30) 0,093 7 0,936 Sol (4S) Traitement 3 (T3D60) 0,068 10 0,955
Traitement 4 (T4D90) 0,089 8 0,828
Moyenne
0,084 ± 0.012
8 ± 2,0
-
Sol (StC)
Traitement 1 (T1D0)
0,0004
-
-
Sol (St4S)
Traitement 1 (T1D0)
0,0004
-
-

Quatrième partie – Chapitre 1 : Confirmation du pouvoir biodégradant du sol de Perpignan et recherche dʼune éventuelle biodégradation accélérée (BDA)
127
3- Conclusion
Le sol de Perpignan présente bien une réactivité biologique importante vis-à-vis de la
sulcotrione, et ces résultats confirment le rôle majeur des communautés microbiennes dans la
dégradation de la substance active. Cette réactivité se caractérise, dans ce cas, par un potentiel
de dégradation notable puisque 50 % de la substance active est dégradée après seulement 7
jours d’incubation en microcosmes. Les profils cinétiques observés traduisent une
prépondérance du processus de co-métabolisme dans la dégradation de cet herbicide dans le
sol. L’enchaînement de quatre cycles de traitement successifs, de 30 jours chacun, n’a pas été
à l’origine de réponses différentes des communautés microbiennes, et cela, autant pour les
microcosmes de sol (4S) et que ceux de sol (C), opposés pourtant, par des historiques de
traitement volontairement différents. La diminution de la demi-vie de la molécule active,
évènement signature de la mise en place du phénomène de biodégradation accélérée (Topp et
al. 2003) n’a donc pas été observée. L’adaptation de la microflore tellurique n’a, à priori, pas
été suffisamment importante pour générer une biodégradation accélérée (BDA) en conditions
contrôlées (microcosmes).
Ces résultats peuvent être expliqués par un nombre de traitement peut-être insuffisant. Cette
conclusion va dans le sens de F. Martin-Laurent (communication personnelle), la
biodégradation accélérée d’un produit phytosanitaire ne pouvant être valablement mise en
évidence qu’à partir de traitements s’étalant sur des durées beaucoup plus longues (plusieurs
décennies ?).
D’autre part, comparativement aux doses généralement utilisées pour les pesticides ayant
généré des processus de biodégradation accélérée, la dose d’application de la sulcotrione
(300-450 g.Ha-1 en maïsiculture) apparaît sensiblement inférieure, ce qui pourrait limiter ou
retarder l’apparition des phénomènes d’adaptation biologique. Cette problématique avait déjà
été abordée par El Sebaï (2004), qui rapportait que la majorité des BDA avaient été observées
pour des produits phytopharmaceutiques dont les doses utilisées étaient généralement égales
ou supérieures à 1000 g.Ha-1. Il s’était notamment interrogé sur les effets imputés à
l’utilisation de ces composés sur la flore microbienne tellurique: (i) Des doses inférieures à
1000 g.Ha-1 sont-elles susceptibles de générer une pression environnementale suffisante sur
les communautés microbiennes pour provoquer des phénomènes adaptatifs ? (ii) La quantité
de carbone disponible issue du pesticide est-elle, dans ce cas, suffisante pour générer des

Quatrième partie – Chapitre 1 : Confirmation du pouvoir biodégradant du sol de Perpignan et recherche dʼune éventuelle biodégradation accélérée (BDA)
128
effectifs microbiens en nombre important afin d’observer une BDA à l’échelle d’une parcelle
agricole ?
Au début de cette étude, nous avions fait l’hypothèse que le sol de Perpignan pourrait
éventuellement présenter une aptitude « naturelle » à la BDA de la sulcotrione, facilement
observable dans des conditions de microcosmes, en réponse à des cycles de traitements
répétés traduisant une stimulation de la biomasse dégradante en taille et/ou en activité. Cela
n’a pas été le cas. Néanmoins, le potentiel de dégradation de ce sol a été confirmé et laisse
entrevoir tout de même, la possibilité d’y trouver des microorganismes dotés de capacités
génétiques à dégrader ce pesticide. Au regard de ces résultats et conscient des difficultés
expérimentales inhérentes à la culture des microorganismes telluriques sur milieux
synthétiques, nous avons toutefois tenté d’isoler à partir du sol (4S) des souches bactériennes
susceptibles de biodégrader l’herbicide sulcotrione.

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
129
Chapitre2‐Isolementdesouchesbactériennespotentiellementdégradantesdelasulcotrione
1- Recherche de souches sulcotrione-résistantes
La première étape de notre travail a consisté à développer une méthode de culture
utilisant les échantillons environnementaux de sol (4S) et (C) sous forme de suspensions dans
du sérum physiologique, avant étalement, à diverses dilutions (100 à 10-4) sur le milieu (NS),
constitué de sol (C) stérilisé, d’agar et supplémenté en sulcotrione (30 mg.L-1) et
cycloheximide (cf figure III-2-2). Sur ce milieu, synthétique mais se rapprochant du micro-
habitat initial des souches telluriques, nous avons pu faire certaines observations :
- seule la dilution 100 (direct) a permis une croissance exploitable de micro-organismes après
sept jours d’incubation à l’obscurité;
- dans ces conditions, de nombreuses colonies bactériennes ont poussé, sans contamination
fongique ; divers morphotypes ont été observés ;
- un morphotype dominant a été retenu, caractérisé par des colonies blanches de petite taille et
de consistance visqueuse ; ces isolats ont montré une morphologie comparable, formant des
colonies « lisses », plus ou moins opaques, convexes, de forme circulaires et de diamètre
voisin de 2,5 mm.
- présence d’un nombre de colonies plus élevé pour le contrôle [milieu (MS) non supplémenté
en sulcotrione] que pour les géloses supplémentées avec l’herbicide ;
- huit colonies bactériennes ont été isolées après purification par étalements successifs sur
milieu gélosé à partir de ce morphotype ;
- la croissance de ces colonies est possible malgré par la présence de sulcotrione à des teneurs
importantes dans le milieu de culture (30 mg.L-1) ;
Cette méthode « hybride », a permis de cultiver, à partir d’un échantillon environnemental de
sol (milieu complexe naturel), des souches bactériennes difficilement cultivables sur milieux
synthétiques conventionnels, avec un temps d’incubation relativement court et en l’absence de
toute contamination mycélienne. Elle est directement adaptée des travaux de Ferreira et al.
(2009) ; son originalité réside dans l’utilisation simultanée de sol initial stérilisé et de gélose,

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
130
à des concentrations respectives optimisées, limitant ainsi les effets potentiellement délétères
de l’agar sur certaines colonies bactériennes et se rapprochant des conditions naturelles
propres aux communautés microbiennes du sol (4S). Ainsi, en jouant de façon synergique sur
divers paramètres contrôlant les étapes du protocole d’isolement, comme l’avaient déjà décrit
Zengler et al. (2002), il semblerait que les conditions physiologiques de culture développées
dans notre approche soient favorables à la croissance d’espèces bactériennes du sol. Ces
résultats vont dans le sens des travaux déjà développés par Janssen et al. (2002), qui ont pu
isoler du sol de nouvelles souches bactériennes décrites jusqu’alors comme non cultivables et
caractérisables uniquement que par des techniques moléculaires.
La comparaison du nombre de colonies obtenues à partir des géloses additionnées d’herbicide
et celles obtenues à partir du contrôle (sans sulcotrione) révèle que l’ajout d’herbicide dans le
milieu de culture a un impact sur la croissance des colonies cultivables. Malgré ce stress
« environnemental », une croissance des colonies, issue du morphotype dominant est tout de
même observée, ce qui nous autorise à qualifier ces isolats de souches résistantes à la
sulcotrione.
L’obtention de ces huit souches nous a fourni le support favorable à la poursuite de notre
travail.
2- Isolement de souches bactériennes métabolisant la sulcotrione
Chacune de ces huit colonies résistantes, préalablement isolée et purifiée, a été
cultivée sur milieu liquide (MS) supplémenté en sulcotrione comme seule source de carbone
et/ou d’énergie. Cinq cycles de culture successifs de 20 jours ont été menés de façon à exercer
une importante pression de sélection sur la population microbienne issue de chaque isolat et à
induire des mécanismes génétiques adaptatifs (cf figure III-2-3). A l’issue de ces cinq cycles,
nous avons souhaité évaluer la capacité des isolats à utiliser la sulcotrione comme source de
carbone : la croissance de la biomasse microbienne a donc été suivie par méthode
spectrophotométrique (à 600 nm) et la dissipation de l’herbicide par méthodes
chromatographiques.

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
131
A ce stade, quelle qu’ait été la souche étudiée, l’augmentation de la biomasse a été très limitée
(faible vitesse de croissance), la dégradation de la substance active n’a été que peu
significative, le pourcentage de dégradation (calculé par rapport à la teneur initiale)
n’excédant pas 10 ± 4 % pour la souche la plus active. A la lumière de ces premiers résultats,
il est possible de considérer que le nombre de cycles de cultures avec pression de sélection de
l’herbicide, volontairement limité à cinq pour des raisons pratiques, ait été insuffisant.
Afin de mieux caractériser visuellement la croissance des colonies, nous avons ensuite
ensemencé des aliquotes des milieux de culture précédents sur milieu gélosé (MS)
supplémenté en sulcotrione, au niveau duquel la croissance est généralement plus nette.
Chacune des huit souches a plus ou moins poussé sur ce milieu solide (dilutions décimales
100 et 10-1 après étalement au râteau et striage direct sur gélose), semblant montrer, à ce stade,
une aptitude des diverses souches bactériennes à métaboliser la sulcotrione avec plus ou
moins d’efficacité. Cependant, l’agar peut, dans certains cas, représenter une source parasite
de carbone du fait de ses impuretés éventuelles et générer des biais expérimentaux. La figure
IV-2-1 représente des exemples de colonies cultivées dans ces conditions. La suite de notre
travail a donc consisté à mesurer la capacité de dégradation des isolats après ensemencement
dans des cultures liquides (MS) supplémentées en sulcotrione.
Nous avons donc préféré, à ce stade, utiliser, sur les huit souches isolées, la technique de
« batch » (cf figure III-2-4), méthodologie qui nous a permis d’augmenter, par concentration
préalable, la biomasse microbienne en phase exponentielle de croissance, au sein du milieu
liquide (MS) et, de façon concomitante, l’impact sur la dégradation de la substance active et
l’apparition de métabolites. Afin d’obtenir une biomasse importante sans pour autant perdre la
capacité dégradante vis-à-vis de l’herbicide, chaque souche a été cultivée pendant 15 heures
en milieu riche (bouillon TS supplémenté en sulcotrione) puis re-ensemencée par dilution en
milieu liquide (MS + sulcotrione) de façon à obtenir une absorbance voisine de 0,300 à 600
nanomètres.

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
132
Figure IV-2-1 : Colonies isolées sur milieu minimum (MS) supplémenté en sulcotrione comme seule source de carbone et/ou d’énergie après croissance à 24°C à l’obscurité.
Parmi les huit souches testées, seule la souche d’apparence opaque, dénommée 1OP, a
présenté une croissance notable visualisée par le suivi de l’absorbance. Sa cinétique a présenté
deux phases : une phase de latence d’environ six jours et une phase de croissance continue
atteignant une absorbance de 0,700, trente jours après l’ensemencement. Aucune
augmentation de l’absorbance, donc de la biomasse microbienne, n’a été observée au bout des
trente jours pour les 7 autres souches, incubées dans les mêmes conditions.
La figure IV-2-2 présente le suivi de l’absorbance pour la souche 1OP (dans le milieu MS
supplémenté en sulcotrione) vs la souche 1TRANS, choisie comme contrôle négatif de

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
133
croissance parmi les 7 souches n’ayant pas présenté de croissance suffisamment significative
lors de nos essais pour être retenues comme souches à potentiel de dégradation intéressant.
Figure IV-2-2 : Suivi de la croissance des souches 1OP vs 1TRANS dans le milieu de culture (MS) supplémenté en sulcotrione par mesure de l’absorbance à 600 nm.
Parallèlement, une expérimentation comparable (contrôles) mais menée en milieu liquide
(MS) non supplémenté en sulcotrione, à partir de ces deux mêmes souches 1OP et 1TRANS,
n’a montré aucune augmentation de l’absorbance, donc de la biomasse, à l’issue des 30 jours
de culture (cf figure IV-2-3).
!"
!#$"
!#%"
!#&"
!#'"
!#("
!#)"
!#*"
!#+"
!" (" $!" $(" %!" %(" &!"
Abs
orba
nce
(600
nm
)
Temps d'incubation (jours)
Souche 1OP Souche 1TRANS

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
134
Figure IV-2-3 : Suivi de la croissance des souches 1OP vs 1TRANS dans le milieu de culture (MS) non supplémenté en sulcotrione par mesure de l’absorbance à 600 nm.
Ces résultats nous permettent de faire les remarques suivantes :
- Le milieu (MS) ne contient donc pas, a priori, de source nutritive carbonée assimilable par
ces deux souches.
- Les souches 1OP et 1TRANS ne présentent pas un caractère de croissance de type
autotrophe chimiolithotrophe.
- La sulcotrione semble constituer la seule source de carbone et/ou d’énergie du milieu (MS)
supplémenté pour la souche 1OP.
3- Etude du potentiel de dégradation des isolats dégradants : suivi de la dissipation de la sulcotrione dans le milieu de culture (MS)
L’augmentation significative de la biomasse pour la souche 1OP constitue un élément
important permettant de préjuger de son rôle biodégradant vis-à-vis de la sulcotrione.
!"
!#$"
!#%"
!#&"
!#'"
!#("
!#)"
!#*"
!#+"
!" (" $!" $(" %!" %(" &!"
Abs
orba
nce
(600
nm
)
Temps d'incubation (jours)
Souche 1OP Souche 1TRANS

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
135
Néanmoins, il était indispensable de confirmer la probable métabolisation de l’herbicide par
(i) le suivi de la concentration de la sulcotrione dans le milieu de culture (MS) supplémenté au
cours de ces 30 jours d’incubation ; (ii) la mise en évidence de l’apparition concomitante de
métabolites de l’herbicide.
Dans ce but, les méthodes chromatographiques (CLHP/UV), décrites au chapitre III-1, ont été
largement utilisées, après extraction d’aliquotes de milieu de culture par solvant organique.
La dissipation de l’herbicide a été très nettement mise en évidence dans le milieu (MS
supplémenté) consécutivement à son ensemencement avec la souche 1OP :
- Une phase de latence d’une durée d’environ 6 jours a été observée avant le début de la
dissipation effective de la sulcotrione. Ce résultat est en accord avec la phase de latence
observée, dans les mêmes conditions, lors du suivi de la croissance de la biomasse par mesure
de l’absorbance (cf figureIV-2-2). Ce délai peut s’expliquer par le temps nécessaire pour
induire l’expression des gènes dans le génome de la souche 1OP et la synthèse des enzymes
cataboliques impliquées dans la transformation de l’herbicide.
- A partir du 6ième jour d’incubation, la cinétique de dissipation de la sulcotrione peut
raisonnablement être décrite par une loi de premier-ordre :
Ln C = - 0,081 t + Ln 121
avec r² = 0,993 (n=3) et C représentant la concentration de l’herbicide en µmoles.L-1.
Le temps de demi-vie (t1/2) a pu être estimé à environ 15 jours après le début de l’incubation.
Dans ces conditions, le pourcentage de dégradation, lié à l’activité métabolique de la souche
1OP, a été estimé à 88 % au bout de 27 jours.
Lors de cette étude, deux « contrôles » ont été utilisés : un contrôle avec la souche 1TRANS
ensemencée et incubée dans les mêmes conditions, et un deuxième contrôle mené sur le
milieu (MS) supplémenté mais non ensemencé. L’ensemble de ces résultats est présenté dans
la figure IV-2-4.

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
136
Figure IV-2-4 : Suivi de la dissipation de la sulcotrione en fonction du temps dans le milieu de culture (MS) supplémenté : ensemencé par 1OP ; ensemencé par 1TRANS ; non ensemencé.
Au cours de l’incubation des 2 lots « contrôles », aucune dissipation significative de la
sulcotrione n’a été observée. Les équations relatives à la dégradation n’ont pas été établies.
L’absence d’évolution du « contrôle » non ensemencé comparativement à la souche 1OP
permet d’affirmer que la part de dégradation par voie chimique (hydrolyse principalement, les
incubations ayant été réalisées à l’obscurité) apparaît négligeable par rapport au processus
biologique.
Par ailleurs, ces résultats confirment une des données obtenues par spectrophotométrie visible
lors du suivi de la croissance bactérienne à 600 nm: le profil cinétique de la souche 1TRANS
étant similaire à celui du contrôle non ensemencé, cette souche ne présente donc pas
d’aptitude à métaboliser la substance active.

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
137
La capacité de la souche 1OP à utiliser la sulcotrione (88 % de dégradation), lorsque ce
composé constitue la seule source de carbone et/ou d’énergie du milieu, est donc confirmée,
résultat corroboré par l’augmentation importante de la biomasse déjà décrite (absorbance
variant de 0,300 à 0,700 au bout de 30 jours).
4- Recherche de métabolites de la sulcotrione dans le milieu de culture (MS)
La recherche de l’apparition concomitante des métabolites principaux : l’acide 2-
chloro-4 méthyl sulfonyl benzoïque (CMBA) et la 1,3-cyclohexane dione (CHD) a été
entreprise pour confirmer la métabolisation et acquérir, si possible, des informations relatives
aux voies métaboliques empruntées.
Ces deux métabolites ont été recherchés tout au long du processus d’incubation par méthode
de chromatographie liquide avec détection UV (λ = 265 nm). Les chromatogrammes
correspondants sont joints en annexe 2. La présence de CMBA a été formellement confirmée
pour l’isolat 1OP vs 1TRANS par CPG/SM (annexe 3) après méthylation de la fonction
carboxylique. L’acquisition récente d’un couplage CLHP/SM nous a permis de confirmer les
résultats concernant la CHD.
Les résultats sont présentés dans le tableau IV-2-1 et repris dans la figure IV-2-5 qui montre
clairement que le CMBA commence à se former dans le milieu de culture (MS) supplémenté
et ensemencé par la souche 1OP dès la fin de la phase de latence préalable à la dissipation
effective de la sulcotrione. Cette augmentation est constante entre 6 et 27 jours, le rapport
molarité en CMBA / molarité initiale en sulcotrione atteignant 44 % à l’issue de l’étude.
Le bilan molaire (teneur en sulcotrione non métabolisée + teneur CMBA formé x 100 / teneur
initiale en sulcotrione) fait état d’une diminution sensible entre le 15ième et le 27ième jour
d’incubation. A ce stade, nous ne pouvons qu’avancer l’hypothèse d’une possible dégradation
du CMBA lui-même par la souche 1OP, consécutivement à l’appauvrissement en sulcotrione
du milieu.

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
138
Figure IV-2-5 : Suivi de l’accumulation de l’acide 2-chloro-4 methylsulfonyl benzoïque (CMBA) et de la dissipation de la sulcotrione en fonction du temps dans le milieu de culture (MS) supplémenté et ensemencé par la souche 1OP.
La 1,3-cyclohexanedione (CHD), identifiée au cours du travail de H.Chaabane dans le sol de
Perpignan à des concentrations très faibles, n’a jamais été détectée par CLHP/UV (< l.d. : 0, 5
mg.L-1) dans les milieux de culture (MS) ensemencés par 1OP ou 1TRANS. Ce résultat
négatif concernant la 1,3-cyclohexanedione a été confirmé par SM et SM-SM (CLHP et
méthode « infusion »).
L’attaque de la partie cyclohexanedione de la molécule de sulcotrione et son utilisation
comme source de carbone et/ou d’énergie par la souche dégradante 1OP semblent donc
probables. A ce stade de nos travaux, et en se référant à des données de la littérature sur des
études ciblées relatives au métabolisme de diverses di- et tri-cétones, plusieurs hypothèses ont
pu être envisagées, concernant la voie métabolique empruntée ; nous en avons retenu trois,
sans exclure cependant d’autres possibilités :
!"
#!"
$!"
%!"
&!"
'!!"
'#!"
'$!"
!"
#!"
$!"
%!"
&!"
'!!"
'#!"
'$!"
!" (" '!" '(" #!" #(" )!"
Con
cent
ratio
n en
CM
BA
(µM
)
Con
cent
ratio
n en
sulc
otri
one
(µM
)
Temps d'incubation (jours)
Concentration sulcotrione 1OP Concentration CMBA 1OP

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
139
- (a) une coupure hydrolytique au niveau du carbonyle en β entrainant la formation exclusive
de la CHD et du CMBA, suivie d’une attaque plus ou moins rapide de la CHD par des
systèmes enzymatiques adaptés (hydrolases ?). Dans notre cas, cette hypothèse semble
cependant peu probable du fait de la non-détection de la 1,3-cyclohexanedione dans les
prélèvements effectués à partir de ce milieu de culture, quel que soit le stade d’incubation.
- (b) le clivage de la molécule de sulcotrione par ouverture du noyau 1,3-cyclohexanedione,
aboutissant à un premier métabolite dérivé de l’acide heptanoïque : l’acide 5,7-dicéto-7-(2-
chloro-4-méthylsulfonylphényl) heptanoïque, déjà décrit dans certains sols par Rouchaud et
al. (1998a). Ce composé subit ensuite un clivage hydrolytique enzymatique aboutissant à un
composé aliphatique en C5 (acide glutarique) métabolisable via le cycle des acides
tricarboxyliques (TCA) et un dérivé de l’acétophénone (1-acétyl-2-chloro-4-méthylsulfonyl-
benzène) se transformant rapidement en CMBA. Le dérivé heptanoïque n’a jamais été
retrouvé au cours de notre étude.
- (c) le clivage de la sulcotrione par un mécanisme oxydatif, déjà décrit pour certaines
dicétones par Dangel et al. (1989) et par Grogan (2002, 2005), catalysé par des systèmes
enzymatiques de type catéchol-dioxygénases et aboutissant à une coupure de la liaison C-C au
niveau de la β-dicétone et à la formation d’un composé aliphatique empruntant également la
voie métabolique des TCA. Plusieurs auteurs (Rouchaud et al., 1996 ; Lin et al., 2000 ;
Huang et al., 2002 ; Wu et al., 2002 ; Szczecinski et al., 2008) ont décrit l’existence d’un
grand nombre de formes tautomères céto-énoliques dans la famille des benzoylcyclohexane-
1,3-diones. L’existence de formes tautomères de la partie 1,3 cyclohexanedione de la
sulcotrione, présentant une relative analogie structurale avec certains dérivés catécholiques,
soit directement, soit après intervention préalable sur le carbone 3 d’une monooxygénase
flavinique de type hydroxylase à large spécificité, pourrait expliquer, via l’intervention de la
catéchol 2,3-dioxygénase, une attaque enzymatique ciblée de la substance active par la souche
1OP. Une des caractéristiques essentielles de ce type d’enzyme est d’avoir une spécificité de
substrat généralement plus large que les catéchol 1,2-dioxygénases (Hirose et al., 1994). Le
fer à l’état réduit (Fe2+) permet l’interaction directe de l’oxygène diatomique avec le métal
(Kita et al., 1998). Au cours de la catalyse enzymatique lors de l’interaction avec le site actif

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
140
de l’enzyme, le substrat n’a pas forcément besoin de subir un changement d’orientation aussi
important et aussi précis que dans le cas d’une catéchol 1,2-dioxygénase (Arciero et
Lipscomb, 1986). Cette moins grande spécificité de substrat pourrait éventuellement
expliquer l’attaque de la partie 1,3-cyclohexanedione sous une des ses formes céto-énoliques.
A ce stade, il nous a semblé intéressant de rappeler que l’action herbicide de la sulcotrione
(Pallett et al., 2001 ; Dayan et al., 2007) s’exerce par inhibition compétitive de la 4-
hydroxyphénylpyruvate dioxygénase (HPPD, EC 1.13.11.27, EC 1.14.2.2), ce qui permet
d’envisager d’autres interactions possibles dioxygénase - sulcotrione.
Jours
Concentration en
sulcotrione
mg.L-1
(µM)
Concentration
en CMBA
mg.L-1
(µM)
Concentration
en CHD
mg.L-1
(µM)
Rapport molarité
CMBA/sulcotrione
initiale
(%)
Bilan
molaire
(%)
0 36 ± 3
(110 ± 10)
ND (< 0,5) ND (< 0,5) - -
6 37 ± 3
(114 ± 13)
ND (< 0,5) ND (< 0,5) - -
15 21 ± 2
(65 ± 7)
6 ± 0,5
(25 ± 2)
ND (< 0,5) 22 82
27 5 ± 1
(16 ± 3)
12 ± 1
(48 ± 3)
ND (< 0,5) 44 59
Tableau IV-2-1 : Evolution des concentrations en substance active et métabolites dans le milieu de culture (MS) supplémenté en sulcotrione et ensemencé par l’isolat bactérien 1OP (ND : non détecté).
A titre comparatif, dans le cas de la mésotrione, Durand et al. (2006b et 2010) et Batisson et
al. (2009) ont proposé un mécanisme de dégradation apparenté par clivage oxydatif de la
partie 1,3-cyclohexanedione catalysé par le genre Bacillus, via deux voies différenciées : une
voie métabolique majeure, dans laquelle l’herbicide est d’abord attaqué par une nitroréductase
au niveau du radical nitro en position 4 du noyau aromatique, donnant un dérivé

Quatrième partie – Chapitre 2 : Isolement de souches bactériennes potentiellement dégradantes de la sulcotrione
141
hydroxylamine intermédiaire, puis subit un clivage oxydatif de la partie 1,3-cyclohexanedione
pour donner l’acide glutarique et l’acide 2-amino-4-méthylsulfonylbenzoïque (AMBA). Dans
le cas de la voie métabolique mineure, le clivage oxydatif de la 1,3-cyclohexanedione semble
être la seule attaque enzymatique permettant de cliver la mésotrione en acide glutarique et en
acide 4-méthylsulfonyl-2-nitrobenzoïque (MNBA) donnant presque directement l’AMBA
après réduction du MNBA.
La connaissance précise du mécanisme de métabolisation de la sulcotrione par la souche 1OP
nécessiterait, à notre avis, une étude spécifique beaucoup plus approfondie.
A ce stade de notre étude, parmi les huit souches résistantes à la sulcotrione isolées sur milieu
(NS supplémenté) et purifiées, seule l’une d’entre elles (isolat 1OP) a montré une forte
aptitude à métaboliser la sulcotrione en l’utilisant comme source de carbone et /ou d’énergie
sur milieu (MS), avec une vitesse de dégradation élevée (temps de ½ vie de 15 jours et
pourcentage de dégradation voisin de 88 %). Néanmoins, ce potentiel biodégradant n’a pu
être caractérisé qu’à la suite d’une étape de concentration de la biomasse microbienne (batch
culture), ce qui permet d’évoquer la notion de seuil de population microbienne, nécessaire à
l’observation du processus métabolique.
La suite de ce travail a donc consisté : i) à identifier, d’un point de vue phylogénétique, cette
souche bactérienne efficace en l’opposant à la souche 1TRANS, choisie parmi les sept
souches à faible activité biodégradante et ii) à caractériser d’un point de vue fonctionnel ces
deux isolats par l’utilisation de cibles moléculaires adaptées.

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
142
Chapitre3‐Caractérisationphylogénétiquedesisolats1OPet1TRANS
1- Vérification de la qualité de l’ADN matrice
Après extraction et purification de l’ADN génomique des isolats, nous avons effectué
une quantification et une évaluation de la qualité des échantillons par dosage au
spectrophotomètre à 260 et 280 nm. La qualité des ADN obtenus pour les souches 1OP et
1TRANS s’est avérée satisfaisante au regard des rapports d’absorbance (rapports A260/A280
et A260/A230 proches de 1,80 pour les deux isolats). Les résultats sont présentés dans le
tableau IV-3-1.
SOUCHE 1OP ADN génomique
SOUCHE 1TRANS ADN génomique
Concentration (ng.µL-1)
486 ± 51
371 ± 56
A260/A280
2,09 ± 0,01
2,06 ± 0,01
A260/A230
1,88 ± 0,19
1,70 ± 0,23
Tableau IV-3-1 : Quantification et évaluation de la qualité de l’ADN génomique des isolats bactériens 1OP et 1TRANS. 2- Analyse du polymorphisme de séquence des ADNr 16S amplifiés (ARDRA) des isolats Les empreintes moléculaires ont été obtenues après utilisation des deux enzymes de
restriction Alu I et Hae III, largement utilisées pour la caractérisation de communautés
bactériennes et/ou d’isolats bactériens issus de nombreux groupes (Pukall et al., 1998). Les
souches bactériennes 1OP et 1 TRANS ont montré des empreintes identiques après digestion
par Alu I, composées de quatre bandes majoritaires d’environ 70, 210, 380 et 550 paires de
bases. Dans le cas de la digestion avec Hae III, les profils ARDRA ont été également

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
143
identiques pour les deux isolats, mettant en évidence cinq bandes majoritaires d’environ 30,
120, 180, 210 et 300 paires de bases. Les résultats obtenus sont présentés dans les figures IV-
3-1 et IV-3-2.
Figure IV-3-1 : Empreintes ARDRA des isolats 1OP et 1TRANS générées après digestion des ADNr 16S par Alu I et migration sur gel d’agarose (1%). La taille des fragments est indiquée en paires de bases (pb).
Figure IV-3-2 : Empreintes ARDRA des isolats 1OP et 1TRANS générées après digestion des ADNr 16S par Hae III et migration sur gel d’agarose (1%). La taille des fragments est indiquée en paires de bases (pb).

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
144
Les profils de restriction obtenus après digestion des ADNr 16S des deux isolats 1OP et
1TRANS sont donc apparus identiques, et cela, quelle que soit l’endonucléase utilisée. En
outre, l’utilisation simultanée des deux enzymes de restriction Hae III et Alu I a également
généré des profils ARDRA identiques pour les deux isolats.
A ce stade, il nous a donc été impossible de différencier taxonomiquement les deux souches
bactériennes par l’utilisation de cette méthode moléculaire, l’analyse du polymorphisme de
longueur des séquences de l’ADNr 16S de l’opéron ribosomique bactérien n’ayant pas révélé
de différence entre 1OP et 1TRANS. Pourtant, cette technique de caractérisation moléculaire
a largement été décrite dans la littérature comme suffisamment fiable, robuste et
particulièrement adaptée comme outil d’approche taxonomique globale des microorganismes
cultivables et non-cultivables (Vaneechoutte et al., 1992 ; Martinez-Murcia et al., 1995). De
plus l’ARDRA permet généralement la discrimination de populations microbiennes jusqu’au
niveau du genre bactérien (Martin-Laurent et al., 2001).
A ce stade de notre travail, les résultats de l’ARDRA semblent donc indiquer que les deux
isolats 1OP et 1TRANS appartiennent donc au même genre bactérien. Afin d’améliorer la
discrimination taxonomique des deux isolats, nous avons souhaité utiliser une méthodologie
différente et plus résolutive que l’ARDRA, la REP-PCR.
3- Analyse des isolats bactériens par amplification des séquences répétitives (REP-PCR)
L’utilisation de la REP-PCR comme méthode d’étude taxonomique complémentaire a
été retenue pour deux raisons majeures : (i) cette méthode permet généralement de
différencier des souches bactériennes appartenant à des groupes phylogénétiquement très
proches (Versalovic et al., 1991 ; Schneider et de Bruijn, 1996 ), (ii) elle permet de réaliser
une analyse de la diversité génétique plus fine que la technique de l’ARDRA, pouvant dans
certain cas discriminer des souches microbiennes jusqu’au niveau de l’espèce (Frey et al.,
1996). Les profils REP-PCR des isolats 1OP et 1TRANS sont présentés figure IV-3-3.

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
145
Figure IV-3-3 : Profils REP-PCR des isolats 1OP et 1TRANS. Puits 1, 2, 5 et 6: REP PCR réalisée avec 1 ng.µL-1 d’ADN matrice. Puits 3, 4, 7 et 8: REP PCR réalisée avec 2 ng.µL-1 d’ADN matrice.
A l’issue de cette étape, les profils REP-PCR respectifs des isolats 1OP et 1TRANS se sont
avérés similaires, confirmant ainsi les premiers résultats obtenus par l’ARDRA. De plus, cinq
bandes majoritaires d’environ 550, 400, 270, 150 et 100 paires de bases ont été retrouvées
pour les deux isolats 1OP et 1TRANS.
Ces résultats suggèrent donc que les deux souches 1OP et 1TRANS appartiennent très
probablement à la même espèce bactérienne.
4- Séquençage de l’ADNr 16S des isolats 1OP et 1TRANS L’amplification, à partir de l’ADN génomique, d’une portion importante de l’ADNr 16S des
isolats 1OP et 1TRANS à l’aide des amorces universelles 27f et 1492r (Gurtler et Stanisich,
1996), a généré des amplicons de bonne qualité, de taille comprise entre 1000 et 1650 paires
de bases pour les deux souches bactériennes. Les résultats sont illustrés dans la figure IV-3-4
et le tableau IV-3-2.

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
146
Figure IV-3-4 : Migration sur gel d’agarose (1%) des ADNr 16S des isolats 1OP et 1TRANS après amplification PCR à l’aide des amorces universelles 27f et 1492r.
SOUCHE 1OP ADNr 16S
SOUCHE 1TRANS ADNr 16S
Concentration (ng.µL-1)
32,2 ± 1,2
40,0 ± 2,9
A260/A280
1,84 ± 0,03
1,92 ± 0,03
A260/A230
1,76 ± 0,06
1,79 ± 0,13
Tableau IV-3-2 : Quantification et évaluation de la qualité des ADNr 16S des isolats bactériens 1OP et 1TRANS, obtenus après amplification PCR et purification.
Deux réactions d’amplification PCR sur colonies (l’une pour les clones provenant de 1OP,
l’autre pour les clones provenant de 1TRANS), à l’aide des amorces T7 et SP6 et à partir de
20 colonies blanches bien distinctes, obtenues après transformation des souches Escherichia
coli DH5α™ (Invitrogen) ultra-compétentes, (1OP ou 1TRANS), ont été effectuées afin de ne
pas sélectionner de faux positifs avant le séquençage. Cela a permis de confirmer, de facto,
que les clones retenus pour le séquençage (couleur blanche des colonies) présentent bien le

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
147
vecteur de clonage contenant comme insert d’intérêt les ADNr 16S des isolats bactériens
respectifs. La figure IV-3-5 présente une photographie des deux banques de clones d’ADNr
16S-PCR-colonies positives utilisées pour le séquençage.
Figure IV-3-5 : Mise en culture des colonies recombinantes (colonies blanches T7-SP6-PCR positives) pour la préparation de l’ADN plasmidique (MiniPreps) destiné au séquençage, à partir des banques de clones d’ADNr 16S des isolats 1OP et 1TRANS (milieu gélosé LB + Ampicilline).
Nous avons obtenu 15 colonies blanches positives présentant l’insert d’intérêt à partir de la
banque d’ADNr 16S 1TRANS (les clones 5, 6, 11, 13 et 20 n’ayant pas été retenus) et 12
colonies positives en ce qui concerne la banque 1OP (les clones 27, 31, 32, 33, 36, 37, 38 et
39 n’ayant pas été retenus). Les figures IV-3-6 (a) et (b) présentent le résultat des deux
réactions d’amplification PCR sur colonies après migration sur gel d’agarose 1%.

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
148
Figure IV-3-6 (a) : Migration sur gel d’agarose (1% ) des amplicons après amplification PCR à partir des colonies blanches issues de la banque d’ADNr 16S de la souche 1OP.
Figure IV-3-6 (b) : Migration sur gel d’agarose (1% ) des amplicons après amplification PCR à partir des colonies blanches issues de la banque d’ADNr 16S de la souche 1TRANS.
Nous avons réalisé un séquençage systématique de 12 clones positifs extraits des banques
pour chaque isolat. Pour chaque séquence d’ADNr 16S, un séquençage a été effectué dans les

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
149
sens 5’→3’ et 3’→5’ à l’aide des amorces T7 ou SP6, afin de disposer d’un nombre suffisant
de données pour mener à bien notre analyse phylogénétique.
A l’issue du séquençage d’environ 1500 pb, soit la quasi-totalité de l’ADNr 16S de chaque
isolat, nous avons obtenu 24 séquences de très bonne qualité pour chaque isolat, douze dans
chacun des sens 5’→3’ et 3’→5’.
Le traitement des séquences provenant de 1OP et de 1TRANS par le logiciel SeqManNGen®
(DNASTAR, USA) a permis d’obtenir une séquence consensus d’ADNr 16S représentative
de chaque souche. L’alignement des deux séquences consensus a révélé une identité de
séquence voisine de 100 %. Le résultat de l’alignement est présenté figure IV-3-7.
Les séquences ont été déposées dans la base de données GenBank
(http://www.ncbi.nlm.nih.gov/genbank/)sous les numéros d’accession JF303892 et JF303891
pour 1OP et 1TRANS respectivement.
A la lumière de ces résultats, nous pouvons donc conclure que les isolats 1OP et 1TRANS
sont deux souches appartenant à la même espèce bactérienne. Ce résultat est donc totalement
en accord avec les données obtenues lors de la caractérisation taxonomique, initiée par
l’analyse du polymorphisme de l’ADNr 16S (ARDRA), puis par REP PCR, ces deux
techniques n’ayant pas révélé de différence significative entre les deux isolats.
BIODE 1 AGAGTTTGATCMTGGCTCAGATTGAACGCTGGCGGCAGGCCTAACACATGCAAGTCGAGC 60 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 1 AGAGTTTGATCMTGGCTCAGATTGAACGCTGGCGGCAGGCCTAACACATGCAAGTCGAGC 60 BIODE 61 GGATGAYRGGAGCTTGCTCCTGGATTCAGCGGCGGACGGGTGAGTAATGCCTAGGAATCT 120 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 61 GGATGAYRGGAGCTTGCTCCTGGATTCAGCGGCGGACGGGTGAGTAATGCCTAGGAATCT 120 BIODE 121 GCCTGGTAGTGGGGGACAACGTTTCGAAAGGAACGCTAATACCGCATACGTCCTACGGGA 180 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 121 GCCTGGTAGTGGGGGACAACGTTTCGAAAGGAACGCTAATACCGCATACGTCCTACGGGA 180 BIODE 181 GAAAGCAGGGGACCTTCGGGCCTTGCGCTATCAGATGAGCCTAGGTCGGATTAGCTAGTT 240 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 181 GAAAGCAGGGGACCTTCGGGCCTTGCGCTATCAGATGAGCCTAGGTCGGATTAGCTAGTT 240 BIODE 241 GGTGGGGTAATGGCTCACCAAGGCGACGATCCGTAACTGGTCTGAGAGGATGATCAGTCA 300 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 241 GGTGGGGTAATGGCTCACCAAGGCGACGATCCGTAACTGGTCTGAGAGGATGATCAGTCA 300 BIODE 301 CACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGGACA 360 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 301 CACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGGACA 360 BIODE 361 ATGGGCGAAAGCCTGATCCAGCCATGCCGCGTGTGTGAAGAAGGTCTTCGGATTGTAAAG 420 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 361 ATGGGCGAAAGCCTGATCCAGCCATGCCGCGTGTGTGAAGAAGGTCTTCGGATTGTAAAG 420

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
150
BIODE 421 CACTTTAAGTTGGGAGGAAGGGCAGTAAGCGAATACCTTGCTGTTTTGACGTTACCGACA 480 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 421 CACTTTAAGTTGGGAGGAAGGGCAGTAAGCGAATACCTTGCTGTTTTGACGTTACCGACA 480 BIODE 481 GAATAAGCACCGGCTAACTCTGTGCCAGCAGCCGCGGTAATACAGAGGGTGCAAGCGTTA 540 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 481 GAATAAGCACCGGCTAACTCTGTGCCAGCAGCCGCGGTAATACAGAGGGTGCAAGCGTTA 540 BIODE 541 ATCGGAATTACTGGGCGTAAAGCGCGCGTAGGTGGTTCGTTAAGTTGGATGTGAAAGCCC 600 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 541 ATCGGAATTACTGGGCGTAAAGCGCGCGTAGGTGGTTCGTTAAGTTGGATGTGAAAGCCC 600 BIODE 601 CGGGCTCAACCTGGGAACTGCATCCAAAACTGGCGAGCTAGAGTATGGTAGAGGGTGGTG 660 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 601 CGGGCTCAACCTGGGAACTGCATCCAAAACTGGCGAGCTAGAGTATGGTAGAGGGTGGTG 660 BIODE 661 GAATTTCCTGTGTAGCGGTGAAATGCGTAGATATAGGAAGGAACACCAGTGGCGAAGGCG 720 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 661 GAATTTCCTGTGTAGCGGTGAAATGCGTAGATATAGGAAGGAACACCAGTGGCGAAGGCG 720 BIODE 721 ACCACCTGGACTGATACTGACACTGAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGAT 780 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 721 ACCACCTGGACTGATACTGACACTGAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGAT 780 BIODE 781 ACCCTGGTAGTCCACGCCGTAAACGATGTCAACTAGCCGTTGGAATCCTTGAGGTTTTAG 840 ||||||||||||||||||||||||||||||||||||||||||||||||||||| |||||| N-BIO 781 ACCCTGGTAGTCCACGCCGTAAACGATGTCAACTAGCCGTTGGAATCCTTGAGRTTTTAG 840 BIODE 841 TGGCGCAGCTAACGCATTAAGTTGACCGCCTGGGGAGTACGGCCGCAAGGTTAAAACTCA 900 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 841 TGGCGCAGCTAACGCATTAAGTTGACCGCCTGGGGAGTACGGCCGCAAGGTTAAAACTCA 900 BIODE 901 AATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGAAGCAACGCG 960 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 901 AATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGAAGCAACGCG 960 BIODE 961 AAGAACCTTACCAGGCCTTGACATGCAGAGAACTTTCCAGAGATGGATTGGTGCCTTCGG 1020 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 961 AAGAACCTTACCAGGCCTTGACATGCAGAGAACTTTCCAGAGATGGATTGGTGCCTTCGG 1020 BIODE 1021 GAACTCTGACACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAA 1080 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 1021 GAACTCTGACACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAA 1080 BIODE 1081 GTCCCGTAACGAGCGCAACCCTTGTCCTTAGTTACCAGCACGTTATGGTGGGCACTCTAA 1140 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 1081 GTCCCGTAACGAGCGCAACCCTTGTCCTTAGTTACCAGCACGTTATGGTGGGCACTCTAA 1140 BIODE 1141 GGAGACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTT 1200 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 1141 GGAGACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTT 1200 BIODE 1201 ACGGCCTGGGCTACACACGTGCTACAATGGTCGGTACAGAGGGTTGCCAAGCCGCGAGGT 1260 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 1201 ACGGCCTGGGCTACACACGTGCTACAATGGTCGGTACAGAGGGTTGCCAAGCCGCGAGGT 1260 BIODE 1261 GGAGCTAATCTCACAAAACCGATCGTAGTCCGGATCGCAGTCTGCAACTCGACTGCGTGA 1320 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 1261 GGAGCTAATCTCACAAAACCGATCGTAGTCCGGATCGCAGTCTGCAACTCGACTGCGTGA 1320 BIODE 1321 AGTCGGAATCGCTAGTAATCGCGAATCAGAATGTCGCGGTGAATACGTTCCCGGGCCTTG 1380 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 1321 AGTCGGAATCGCTAGTAATCGCGAATCAGAATGTCGCGGTGAATACGTTCCCGGGCCTTG 1380 BIODE 1381 TACACACCGCCCGTCACACCATGGGAGTGGGTTGCACCAGAAGTAGCTAGTCTAACCTTC 1440 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| N-BIO 1381 TACACACCGCCCGTCACACCATGGGAGTGGGTTGCACCAGAAGTAGCTAGTCTAACCTTC 1440 BIODE 1441 GGGGGGACGGTTACCACGGTGTGATTCATGACTGGGGTGAAGTCGTAACAAGGTAWCCGT 1500 ||||||||||||||||||||||||||||||||||||||||||||||||||||||| |||| N-BIO 1441 GGGGGGACGGTTACCACGGTGTGATTCATGACTGGGGTGAAGTCGTAACAAGGTADCCGT 1500

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
151
BIODE 1501 A 1501 | N-BIO 1501 A 1501 Figure IV-3-7 : Alignement des séquences consensus des ADNr 16S des isolats 1OP et 1TRANS. BIODE : souche 1OP ; N-BIO : souche 1TRANS.
La comparaison des deux séquences consensus d’ADNr 16S avec les séquences disponibles
dans la base de données GenBank (http://www.ncbi.nlm.nih.gov/genbank/) nous a permis de
conclure que les isolats 1OP et 1TRANS présentaient un haut niveau d’homologie (99 %)
avec le genre Pseudomonas de la classe des γ Protéobactéries.
Nous avons donc construit l’arbre phylogénétique à partir d’alignements multiples des
séquences de l’ADNr 16S de nos deux isolats et des séquences d’Eubactéries présentes dans
GenBank, correspondant aux souches bactériennes suivantes: Arthrobacter creatinolyticus
F75208 (HQ908759), Arthrobacter nicotianae HMF5 (GU220490), Streptomyces sp. SNI5
(GU320191), Rhizobium leguminosarum V-6 (GU306144), Agrobacterium tumefaciens
YQH34 (HQ143653), Methylopila sp. JZL-4 (FJ502233), Burkholderia cepacia RFW-8
(HQ650586), Comamonas testosteroni H18 (EU887829), Acidovorax sp. p4(HQ652596),
Pseudomonas putida A (HQ697262), Pseudomonas putida T2g (AB539810), Pseudomonas
aeruginosa P28 (HQ697285), Pseudomonas fluorescens C7R12 (AM229082), Pseudomonas
fluorescens (HQ880245), Pseudomonas sp. ADP (AM088478).
L’analyse phylogénétique nous a révélé que nos deux isolats 1OP et 1TRANS sont deux
souches bactériennes appartenant à l’espèce Pseudomonas putida.
Nous avons donc proposé de nommer nos deux isolats Pseudomonas sp. 1OP et Pseudomonas
sp. 1TRANS. Ces deux souches ont donc été classées comme Bacteries, phylum des
Proteobacteries, classe des γ protéobactéries, ordre des Pseudomonadales, famille
Pseudomonaceae, genre Pseudomonas.
L’observation macroscopique des isolats sur milieu gélosé nous a permis de mettre en
évidence que les deux souches présentaient des colonies de type S (smooth), c’est-à-dire à
surface lisse et à bords réguliers, bombées, de couleur blanche (légèrement plus opaque pour
l’isolat 1OP), de consistance plutôt crémeuse, de taille réduite et donnant des suspensions
homogènes.

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
152
L’observation microscopique de Pseudomonas sp. 1OP et Pseudomonas sp. 1TRANS a mis
en évidence des bactéries de type Gram- et non sporulantes. En milieu liquide, la lecture de
l’état frais nous a permis d’observer des cellules à mobilité importante de type polaire. Quel
que soit l’isolat observé, il s’agit de bacilles simples, de petite taille et de forme plutôt
allongée.
L’observation du critère biochimique par le test catalase a révélé un phénotype catalase+ pour
les deux isolats, cela étant en accord avec les données de la littérature concernant le
métabolisme de type aérobie de Pseudomonas sp.
L’arbre phylogénétique obtenu en utilisant la méthode du Neighbor-Joining est présenté
figure IV-3-8. La robustesse de l’arbre a été vérifiée par analyse par bootstraps. Les valeurs
supérieures à 90 % sont représentées par les cercles noirs sur la figure. La distance
phylogénétique est indiquée sur la barre d'échelle.
Page suivante, figure IV-3-8 : Analyse phylogénétique des ADNr 16S des isolats Pseudomonas sp. 1TRANS (NBD) et Pseudomonas sp. 1OP (BD). Les numéros d’accession GenBank des séquences d’ADNr16S utilisées pour l’analyse sont indiqués entre parenthèse dans la légende.

153

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
154
5- Conclusion Les résultats du séquençage nous permettent donc de dégager trois grandes réflexions:
(i) Nous avons pu identifier de façon formelle les deux souches bactériennes retenues à l’issue
de l’étude de biodégradation de la sulcotrione comme étant affiliées au genre Pseudomonas ;
(ii) les deux isolats obtenus lors de l’approche cultivable appartiennent au phylum des
Proteobacteria, un des quatre phyla dominants les plus fréquemment retrouvés dans les
banques d’ADNr 16S isolés à partir d’échantillons environnementaux (Dunbar et al., 1999 et
2002 ; Mc Garvey et al., 2004 ; Schloss et al., 2004 ; Acinas et al., 2005 ; Janssen, 2006 ;
Herrera et al., 2007),
(iii) Ce phylum appartient également au groupe des « big four » (Proteobacteria, Firmicutes,
Actinobacteria et Bacteroidete), décrit par Hugenholtz (2002) comme majoritaire dans les
collections de souches cultivables provenant d’échantillons environementaux (cf paragraphe
I-2-1);
(iiii) l’isolement des deux souches bactériennes Pseudomonas sp. 1OP et Pseudomonas sp.
1TRANS, appartenant à la même espèce mais présentant des phénotypes dégradants
différents, met en exergue le caractère versatile de cette capacité de biodégradation vis-à-vis
de la sulcotrione.
En ce qui concerne le phénotype dégradant de l’isolat Pseudomonas sp. 1OP, nos
résultats sont en accord avec les données de la littérature, à propos de la capacité du Genre
Pseudomonas à dégrader un nombre important de xénobiotiques présents dans
l’environnement, et notamment dans les sols. Diverses souches appartenant à ce genre
bactérien ont en effet été décrites comme capables de dégrader un certain nombre de produits
phytosanitaires issus de différentes familles. C’est notamment le cas de la souche
Pseudomonas sp. ADP, isolée à partir d'un site de stockage d'herbicides par Mandelbaum et

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
155
al. (1995), capable de métaboliser l'atrazine à des concentrations très élevées (80 % environ
de minéralisation à des concentrations > 1000 ppm).
Daugherty et Karel (1994) ont eux aussi isolé une souche de Pseudomonas cepacia DBO1
utilisant l’acide 2,4-dichlorophénoxyacétique comme source de carbone et/ou d’énergie.
Kamanavalli et Ninnekar (2000) ont également isolé du sol, par enrichissements successifs,
un Pseudomonas sp. dégradant le propoxur (2-isopropoxyphényl-N-méthylcarbamate). Cette
souche a été capable de se développer en utilisant ce composé comme seule source de carbone
et d'azote, à partir de concentrations relativement élevées du produit phytosanitaire (2 g.L-1) et
accumulant au cours du temps, le 2-isopropoxyphenol comme métabolite principal dans le
milieu de culture. Bajaj et al. (2010) ont également caractérisé un Pseudomonas sp. IITR01
capable de métaboliser l’α-endosulfan et l’endosulfan-sulfate. Fan et al. (2010) ont isolé puis
identifié par séquençage de l’ADNr 16S, un Pseudomonas sp. capable de dégrader un
fongicide, le carbendazime, avec un taux de dégradation voisin de 90 % au bout de 3 jours.
Ces auteurs ont proposé une voie de biodégradation dans laquelle le carbendazime est d'abord
converti en 2-aminobenzimidazole, puis transformé en 2-hydroxybenzimidazole, en 1,2-
diamino, en catéchol, et finalement en dioxyde de carbone.
A notre connaissance, il n’existait pas dans la littérature de données relatives à la dégradation
de la sulcotrione catalysée par des microorganismes isolés.
Dans le cas d’autres herbicides appartenant à la famille des tricétones, Durand et al. (2006a,
2006b et 2010) ont pu isoler, à partir d’eau de nuage, une souche bactérienne affiliée au genre
Bacillus, capable de dégrader la mésotrione en moins de 10 heures de culture et cela, pour une
large gamme de concentration en substance active (0,1 à 5 mmol.L-1). Ces auteurs ont
également pu mettre en évidence l’apparition de plusieurs métabolites différents au cours de
la période d’incubation et proposer deux voies métaboliques possibles. De même, Batisson et
al. (2009) ont isolé du sol une autre souche, également affiliée au genre Bacillus (isolat
Mes11), utilisant la mésotrione comme source de carbone et/ou d’énergie et dégradant
totalement la substance active en moins de 24 heures pour une concentration de 1 mmol.L-1.
Trois métabolites déjà identifiés par Durand et al. (2006a, 2006b et 2010) ont été retrouvés
dans le milieu de culture. Dans ce cas, il est intéressant de noter que les auteurs ont montré
que la souche tellurique Bacillus sp. Mes11 était incapable de dégrader la sulcotrione.

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
156
En conclusion, puisque les deux isolats 1OP et 1TRANS présentent deux phénotypes distincts
vis-à-vis du caractère dégradant mais sont deux souches appartenant à la même espèce, ce
résultat nous amène à proposer deux hypothèses pouvant probablement expliquer cette
différence phénotypique:
(i) la présence d’événements mutationnels pléiotropes chez les isolats sulcotrione-résistants,
selon un modèle aléatoire (Lederberg J. et E., 1952) ou non aléatoire, aurait sélectionné,
uniquement chez l’isolat 1OP, des cellules bactériennes aptes à se développer sur milieu
sélectif (MS) après cinq cycles de cultures liquides répétées avec pression de sélection de
l’herbicide (C1→C5);
(ii) l’intervention d’éléments génétiques mobiles (MGE), présents dans le génome de l’isolat
1OP et ayant été activés consécutivement aux conditions de cultures répétées, serait
responsable de l’apparition du phénotype dégradant.
La première hypothèse nous semble la moins probable, pour plusieurs raisons : en premier
lieu, la probabilité d’apparition de mutation est un phénomène rare (taux de mutation moyen
est de l'ordre de 10-6 à 10-7, le temps de génération constituant généralement l’unité de temps
retenue), n'affectant généralement qu'un faible pourcentage de cellules au sein d'une
population abondante. Dans le cas de nos cultures répétées (C1→C5), il semblerait que le
nombre de cycles soit insuffisant et le temps de génération trop long pour générer l’apparition
de phénotypes dégradants.
Nous préférons privilégier la deuxième hypothèse : en effet, l'implication des éléments
génétiques mobiles (MGE) tels que les bactériophages, plasmides, et transposons dans ces
processus cataboliques adaptatifs a été largement décrite dans la littérature comme des acteurs
fondamentaux du transfert génétique horizontal dans la matrice sol (Sikorski et al., 2002 ;
Thomas et Nielsen, 2005). Ces structures génétiques sont des éléments clés pour le transfert
de gènes entre bactéries (Wilkins, 1995 ; Davison, 1999 ; Nielsen et al., 2000 ; Jain et al.,
2002), contribuent directement à leur évolution, et, dans certains cas, jouent un rôle dans la
spéciation bactérienne (Majewski et Cohen, 1999 ; Bergstrom et al., 2000 ; Ochman et al.,

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
157
2000 ; Nielsen et van Elsas, 2001 ; Majewski et al., 2000). Dans le cas des biodégradations,
ces MGE sont de puissants outils adaptatifs pour le développement de métabolismes
alternatifs au sein des populations microbiennes (Ashelford et al., 1999, 2000 et 2003 ;
Ashelford et al., 2000 ; Top et Springael, 2003 ; Thomas et al., 2005 ; Frost et al., 2005).
L’intervention des MGE dans les biodégradations a été particulièrement décrit pour le genre
bactérien Pseudomonas, sur une gamme étendue de composés organiques (Kostal et al.,
1998 ; Chang et al., 2001; Top et al., 2002 ; Bandounas et al., 2011 ; Zhou et al., 2011 ; Zhao
et al., 2011 ; Chanika et al., 2011). De plus, de nombreuses études ont démontré que ce
potentiel métabolique modelable était souvent dû, au sein des MGE, à l’activité d’un ou
plusieurs plasmides cataboliques de type conjugatif, de taille comprise entre 50 à 220 kb
environ et /ou à la présence de gènes cataboliques à l’intérieur d’éléments transposables
(Lilley et Bailey 1997 ; Ogawa et al., 2004 ; Williams et al., 2004 ; Yano et al., 2007).
A ce stade de notre travail, nous proposons plusieurs possibilités à propos de la nature des
MGE responsables du phénotype dégradant, parmi lesquelles :
i) la présence d’un plasmide de type conjugatif, portant le ou les gènes cataboliques
impliqués ;
ii) la présence d’éléments transposables localisés sur le chromosome bactérien et
contenant le ou les gènes cataboliques ;
iii) la présence d’éléments transposables, eux-mêmes localisés sur un plasmide de type
conjugatif et contenant le ou les gènes cataboliques.
À partir de nos résultats, et en les confrontant aux schémas de dégradation classiquement
proposés pour Pseudomonas sp. il est possible d’envisager la présence simultanée de gènes
codant pour des dioxygénases à affinité et/ou activité accrues pour la sulcotrione,
responsables de l’attaque de la substance active au niveau de la partie 1,3-cyclohexanedione.
Nous avons souhaité vérifier cette hypothèse en essayant d’identifier et de localiser les gènes
cataboliques impliqués dans la dégradation de la sulcotrione chez l’isolat Pseudomonas sp.
1OP. Nous avons donc recherché la présence de gènes codant deux familles de dioxygénases
(catéchol 2,3-dioxygénase et catéchol 1,2-dioxygénase) chez Pseudomonas à l’aide d’amorces

Quatrième partie – Chapitre 3 : Caractérisation phylogénétique des isolats 1OP et 1TRANS
158
hautement spécifiques, ayant déjà donné des résultats probants lors d’études similaires (Sei et
al., 1999 ; Jussila et al., 2006 ; Guzik et al., 2011).

Quatrième partie – Chapitre 4 : Caractérisation fonctionnelle des isolats 1OP et 1TRANS
159
Chapitre4‐Caractérisationfonctionnelledesisolats1OPet1TRANS
1- Préambule
Le rôle métabolique des intermédiaires catécholiques, impliqués dans les
biodégradations microbiennes en conditions aérobies et notamment chez Pseudomonas sp., a
largement été décrit depuis plusieurs décennies (Chatterjee et al., 1981 ; Chatterjee et
Chakrabarty, 1982 ; Hickey et Focht, 1990 ; Aemprapa et Williams, 1998 ; Nakai et al.,
1995 ; Lee et al., 1995 ; Wikström et al., 1996 ; Joshi et Walia, 1996a et 1996b ; Kitayama et
al., 1996a et 1996b ; Moiseeva et al., 2002 ; Otenio et al., 2005 ; Guzik et al., 2009). Le
catéchol (1,2-dihydroxybenzène) et par extension les catéchols substitués, constituent des
intermédiaires essentiels dans ces carrefours métaboliques microbiens, et, notamment, pour
des composés tels que les benzoates, naphtalènes, toluènes, salicylates, phénols et dérivés
chlorés et/ou méthylés (Dagley et Gibson, 1965; Dagley, 1986 ; Reineke et Knackmuss, 1988
; Jeenes et al., 1982). Un large éventail d'enzymes est élaboré par les microorganismes, afin
de convertir progressivement les composés aromatiques au cours d’étapes successives et de
les amener, jusqu’au stade ultime des intermédiaires catécholiques. Deux voies enzymatiques
parfaitement définies et spécifiques, la voie méta et/ou la voie ortho prennent en charge les
catéchols et leurs dérivés (cf paragraphe III-3-2-4). En ce qui concerne la voie méta, elle fut
tout d’abord mis en évidence chez des souches de Pseudomonas qui avaient la capacité de
métaboliser le phénol et également le crésol ou le méthylphénol (Dagley et Gibson, 1965).
Herrmann et al. (1995 et 1998) ont également montré que la souche Pseudomonas putida H
métabolisait le crésol grâce à la présence du gène phlH situé sur le plasmide PHE (220 kb) à
l’intérieur d’éléments transposables. Cette voie fonctionne également dans la dégradation de
phénols et de méthyl-phénols substitués pour un nombre important d'espèces de Pseudomonas
différentes (Kukor et Olsen, 1991; Shingler et al., 1989 ; Bartilson, 1990 ; Takenaka et al.,
1998). Au cours des dernières décennies, les données récoltées à propos de ces deux voies
métaboliques se sont depuis largement enrichies (Assinder et Williams, 1990 ; Loh et Chua,
2002 ; Cao et al., 2002 et 2009 ; Jimenez et al., 2002 ; Toyama et al., 2010).
Au regard de ces données, et en les confrontant à nos précédents résultats (étude du potentiel
de biodégradation et caractérisation phylogénétique des isolats), nous avons souhaité savoir si

Quatrième partie – Chapitre 4 : Caractérisation fonctionnelle des isolats 1OP et 1TRANS
160
nos deux souches Pseudomonas sp. 1OP et Pseudomonas sp. 1TRANS pouvaient
éventuellement exprimer ce potentiel génétique catécholique.
2- Détection des gènes XylE codant l’enzyme catéchol 2,3-dioxygénase
La catéchol 2,3-dioxygénase, codée par le gène XylE se trouvant sur le plasmide TOL pWW0
(117 kb environ), constitue une des enzymes clés de la voie catabolique méta, puisqu’elle
catalyse l’ouverture du cycle aromatique du catéchol (Kita et al., 1999). Les gènes Xyl se
trouvant sur le plasmide TOL sont organisés en opérons et sont situés à l’intérieur d’éléments
transposables (Assinder et Williams, 1994). Nous avons cherché à savoir si les gènes XylE
étaient présents dans le génome de nos deux isolats. Les résultats de l’amplification des gènes
XylE sont présentés dans la figure IV-4-1.
Figure IV-4-1 : Migration des produits PCR sur gel d’agarose (1,2%) après amplification de l’ADN génomique des deux isolats 1OP et 1TRANS, à l’aide des amorces XylEaf et XylEar. Contrôle 1 : ADN génomique E. coli DH5α ; contrôle 2 : ADN plasmidique pCambia 3300. La taille des fragments est indiquée en paires de bases (pb).
Les résultats obtenus entraînent plusieurs remarques :
(i) Le couple d’amorces XylEaf et XylEar (Jussila et al., 2007) nous a permis d’amplifier de
l’ADN génomique pour les deux isolats ;
Contrôle 1 Contrôle 2
1 Kb+
300
500
800
Isolat 1TRANS
Isolat 1OP

Quatrième partie – Chapitre 4 : Caractérisation fonctionnelle des isolats 1OP et 1TRANS
161
(ii) Nous avons obtenu plusieurs bandes, et cela quelle que soit la souche bactérienne
considérée ;
(iii) Les profils de migration sont différents entre les deux isolats : trois bandes d’ADN
majoritaires de 300, 500 et 800 pb environ sont observées pour l’isolat Pseudomonas sp.
1TRANS contre deux seulement (300 et de 800 pb) pour l’isolat Pseudomonas sp. 1OP ;
(iiii) La bande de taille approximative de 500 pb est retrouvée uniquement chez l’isolat
Pseudomonas sp. 1TRANS, contrairement aux bandes 300 et 800 pb environ communes aux
deux isolats,
Les deux contrôles négatifs constitués d’ADN génomique de la souche E. coli DH5α de
génotype F- (contrôle 1) et de l’ADN plasmidique (contrôle 2) n’ont donné aucun signal
d’amplification.
Jussila et al. (2007) ont montré que les amorces XylEaf et XylEar qu’ils ont dessinées,
amplifient de façon spécifique la région de 731 pb du gène XylE porté par le plasmide
archétypal TOL pWW0. Au cours de leurs travaux, ces auteurs ont également montré qu’une
température d’hybridation de 70°C lors de la PCR permettait de s’affranchir de l’obtention de
faux-positifs, consécutivement à une amplification possible des gènes nahH présentant des
séquences très comparables avec celles des gènes XylE déjà décrites par Ghosal et al. (1987).
Dans ces conditions, la bande de taille légèrement inférieure à 800 pb, retrouvée chez les deux
isolats, pourrait constituer le signal d’amplification attendu.
En revanche, l’obtention de deux bandes supplémentaires de 300 et 500 pb environ chez
l’isolat Pseudomonas sp. 1TRANS, et d’une seule bande supplémentaire de 300 pb chez
l’isolat Pseudomonas sp. 1OP, nous amène à nous interroger sur ce résultat.
A ce stade, nous avons envisagé deux hypothèses:
(i) des amplifications non spécifiques seraient responsables de la présence des fragments
additionnels de 300 et/ou de 500 pb,
(ii) l’amplification à l’aide du couple d’amorces XylEaf et XylEar est véritablement spécifique
des séquences du gène XylE, et ce résultat met donc en évidence une organisation génétique
différente de l’opéron Xyl pour les deux souches bactériennes étudiées. Dans ce cas, la

Quatrième partie – Chapitre 4 : Caractérisation fonctionnelle des isolats 1OP et 1TRANS
162
présence de réarrangements génétiques dans le génome des deux isolats est fortement
envisageable.
Dans la première hypothèse, il est possible que des hybridations non spécifiques soient
intervenues. Dans le cas de travaux similaires, à partir d’alignements multiples de 20
séquences de C23O extraites de GenBank, Sei et al. (1999) ont montré que l’analyse par PCR
de 12 souches bactériennes de référence exprimant la catéchol 2,3-dioxygénase, avait
également donné des fragments additionnels de différentes tailles pour quatre isolats, dont
trois souches de Pseudomonas putida (souche BH-pheB, Takeo et al., 1995 ; souche MT15-
cdo, Keil et al., 1985 ; souche mt2-pWW0 XylE, Nakai et al., 1983). Ces auteurs ont
également été confrontés au même phénomène en détectant des fragments non spécifiques
lors de « southerns » effectués sur les souches de références PCR-positives. En outre,
l’analyse par PCR de 106 isolats inconnus, présentant un phénotype dégradant vis-à-vis du
phénol et du benzoate, a également généré 35 % de fragments de taille non attendue (> 380
pb).
Jussila et al. (2007) ont dessiné les amorces XylEaf et XylEar à partir de la séquence du gène
XylE (GenBank V01161 ; Nakai et al., 1983) et par comparaison avec des séquences
homologues retrouvées dans GenBank. La spécificité de ces amorces a été ensuite testée avec
différentes températures d’hybridation (61 à 72°C), à partir d’ADN génomique purifié de 12
souches reconnues comme exprimant ou non la catéchol 2,3-dioxygénase. Ces auteurs ont
retenu une température d’hybridation de 70°C lors des cycles de PCR et ont décrit que, pour
des identités de séquences du gène XylE inférieures ou égales à 81%, des amplicons non
spécifiques avaient été obtenus avec un signal cependant très faible (souche Pseudomonas
putida HS1 avec le plasmide TOL/pDK1 ; Kunz et Chapman, 1981). Dans notre cas et quels
que soient les fragments considérés, nous avons obtenu un signal intense des bandes d’ADN,
et cela pour les deux isolats 1OP et 1TRANS.
En ce qui concerne la seconde hypothèse, plusieurs remarques sont à faire : certaines souches
de Pseudomonas sp. peuvent présenter plusieurs copies de l’opéron de la voie méta et/ou
plusieurs copies des gènes codant l’enzyme catéchol 2,3-dioxygénase. Pseudomonas putida
MT53 (DSMZ 4303) porte à la fois deux copies de l’opéron de la voie méta sur le plasmide
pWW53 et deux copies du gène codant la catéchol 2,3-dioxygénase, l’un se trouvant sur le

Quatrième partie – Chapitre 4 : Caractérisation fonctionnelle des isolats 1OP et 1TRANS
163
plasmide archétypal pWW0, l’autre sur le plasmide de type pDK1 (Keil et al., 1985a et
1985b ; Kok et al., 1999 ; Nojiri et al., 2004). Cette souche arbore donc trois plasmides
distincts codant pour des catéchol 2,3-dioxygénases présentant des spécificités de substrat
nettement différentes (Jussila et al., 2007). Chez Pseudomonas aeruginosa JI104, trois copies
homologues du gène XylE ont été caractérisées. Jussila et al. (2007) ont pu vérifier ces
données lors de leurs travaux avec l’utilisation du couple d’amorces XylEaf et XylEar.
La présence des gènes XylE, situés à l’intérieur d’éléments transposables sur le plasmide
archétypal TOL pWW0 (Assinder et Williams, 1990 ; Williams et al., 2004), pourrait
autoriser la mise en place d’événements génétiques de duplication/délétion, pouvant expliquer
la présence des deux fragments supplémentaires (300-500 pb). Dans ce cas, la différence de
signal obtenu entre l’isolat 1OP et l’isolat 1TRANS pourrait mettre en évidence un fort lien
de causalité entre phénotypes dégradant ou non dégradant et nombre de fragments obtenus.
Lors de la phase d’adaptation des isolats sulcotrione-resistants (développée au cours de notre
stratégie cultivable), les cultures répétées avec pression de sélection de l’herbicide auraient pu
provoquer des réarrangements génétiques par transposition et/ou recombinaison chez les
différents isolats. Jeenes et al. (1982) ont déjà décrit ce type d’évènements chez Pseudomonas
sp. Ces auteurs ont décrit des mutants présentant un nouveau phénotype dégradant vis-à-vis
de composés benzoïques halogénés, conféré par l’expression d’une catéchol 2,3-dioxygénase
non spécifique mais fonctionnelle, après une réorganisation structurale du plasmide pWW0
(délétions) et une mutation du gène XylE. A titre purement hypothétique, nous pourrions donc
envisager que les réarrangements génétiques opérés chez l’isolat Pseudomonas sp.1OP
auraient pu favoriser son potentiel dégradant vis-à-vis de la sulcotrione.
A l’issue de nos résultats, il semble intéressant d’approfondir ce ciblage des gènes codant la
catéchol 2,3 dioxygénase par l’optimisation des conditions analytiques, en testant nos deux
isolats avec d’autres couples d’amorces. Le couple 230f et C23Or, dessiné par Sei et al.
(1999) à partir d’alignements multiples de 20 séquences des gènes C23O extraites de
GenBank (http://www.ncbi.nlm.nih.gov/genbank/), ciblant une région plus courte du gène
(380 pb), pourrait être testé à partir de l’ADN génomique des isolats 1OP et 1TRANS. Enfin,
l’utilisation de contrôles positifs avec des souches de référence affiliées au genre
Pseudomonas exprimant la catéchol 2,3-dioxygénase pourrait être envisagé.

Quatrième partie – Chapitre 4 : Caractérisation fonctionnelle des isolats 1OP et 1TRANS
164
Toutefois, afin d’identifier de façon formelle les séquences des fragments amplifiés, nous
avons souhaité séquencer l’ensemble des amplicons (300, 500 et 800 pb) pour les deux isolats
1OP et 1TRANS. Le résultat du séquençage nous permettra d’évaluer la qualité de notre
méthodologie de ciblage des gènes.
3- Détection des gènes C12O codant l’enzyme catéchol 1,2-dioxygénase
Le gène codant la catéchol 1,2-dioxygénase (C12O) a également été recherché chez nos deux
isolats Pseudomonas sp. 1OP et Pseudomonas sp. 1TRANS, selon la méthode décrite par Sei
et al. (1999), également utilisée par Guzik et al. (2011). Les résultats de l’amplification sont
présentés dans la figure IV-4-2.
Figure IV-4-2 : Migration des produits PCR sur gel d’agarose (1,2%) après amplification de l’ADN génomique des deux isolats 1OP et 1TRANS à l’aide des amorces C12Of et C12Or. La taille des fragments est indiquée en paires de bases (pb). Nous avons obtenu des produits PCR pour les deux souches bactériennes. Après migration
des amplicons, les deux isolats présentent une bande majoritaire de bonne qualité, de taille
approximative de 500 pb avec, toutefois, un signal d’intensité légèrement plus élevée pour
l’isolat 1OP. Un fragment de taille voisine de 100 pb, présentant un signal plus faible est
également retrouvé pour 1OP et 1TRANS. Toutefois, l’utilisation des amorces C12Of et
C12Or n’a pas généré, chez nos deux isolats, les amplicons de 282 pb que Sei et al. (1999)
avaient obtenus à partir de 9 souches bactériennes de référence (sur 11 au total) exprimant la

Quatrième partie – Chapitre 4 : Caractérisation fonctionnelle des isolats 1OP et 1TRANS
165
catéchol 1,2-dioxygénase, dont 7 souches affiliées au genre Pseudomonas. Les deux contrôles
négatifs (non présentés sur la figure IV-4-2), identiques à ceux utilisés lors de l’amplification
des gènes XylE, n’ont pas donné de réponse.
Plus récemment, avec les mêmes amorces, Guzik et al. (2011) ont également obtenu un
fragment d’amplification unique à partir de la souche Pseudomonas putida N6 (EF208895.1),
mais de taille légèrement supérieure à celui attendu (380 pb au lieu de 282 pb).
Nous avons donc effectué le séquençage de l’amplicon de 500 pb provenant de Pseudomonas
sp. 1OP et de Pseudomonas sp. 1TRANS, afin de savoir s’ils correspondaient bien à la
séquence codante du gène C12O.
4- Séquençage des amplicons XylE
Deux tentatives de séquençage des différents fragments d’ADN obtenus (800, 500 et 300 pb)
ont donné des chromatogrammes de trop mauvaise qualité pour qu’ils puissent être
correctement exploités. Un nouveau séquençage est actuellement en cours.
5- Séquençage des amplicons C12O
Nous avons séquencé la totalité des 500 paires de bases des fragments d’ADN obtenus après
amplification de l’ADN génomique des isolats 1OP et 1TRANS. Les alignements multiples,
effectués à l’aide du logiciel CLUSTALX, ont montré que ces séquences présentaient entre
elles un très fort pourcentage d’identité (supérieur à 90 %). Le résultat des alignements
multiples est présenté dans la figure IV-4-3.

Quatrième partie – Chapitre 4 : Caractérisation fonctionnelle des isolats 1OP et 1TRANS
166
Figure IV-4-3 : Alignements multiples des séquences provenant des fragments d’ADN (taille approximative 500 pb) obtenus lors du ciblage du gène C12O chez les isolats 1OP et 1TRANS, à l’aide du logiciel ClustalX V. 2.0.10 (Thompson et al., 1997).
Le résultat du BLAST effectué à l’aide du logiciel blastn, dans GenBank a montré que les
séquences obtenues ne correspondaient pas au gène C12O codant l’enzyme catéchol 1,2-
dioxygénase.
Toutefois, la séquence a présenté 89 % d’identité avec le gène codant l’enzyme 4-
diphosphocytidyl-2-C-méthyl-D-érythritol kinase (CMK) de Pseudomonas sp. Une analyse à
l’aide de blastx a bien confirmé que ces séquences nucléotidiques (500 pb) étaient
relativement proches de la séquence de la kinase (95 % de similarité).
Ces résultats constituent une première étape dans la caractérisation fonctionnelle des deux
isolats à propos des gènes C12O. Toutefois, il est nécessaire d’améliorer nos conditions
opératoires (PCR), puisque la détection des gènes codant l’enzyme catéchol 1,2-dioxygénase
chez 1OP et 1TRANS, à partir des conditions décrites par Sei et al. (1999), n’a pas été
possible.

167
Conclusion générale et perspectives

Conclusion générale et perspectives
168
Le travail présenté a permis d’approfondir les connaissances relatives au
comportement d’un herbicide de la famille chimique des tricétones, la sulcotrione. Cette
substance active est notamment proposée dans les itinéraires techniques maïsicoles, avec des
doses d’application nettement inférieures à celles antérieurement préconisées pour des
herbicides plus anciens.
La littérature spécialisée et, plus particulièrement une étude précédemment réalisée au
Laboratoire de Chimie des Biomolécules et de l’Environnement, nous a fourni des données
importantes sur la phénoménologie comportementale de cet herbicide, que nous avons
cependant souhaité compléter par une approche « plus biologique » des processus de
biodégradation.
Grâce à des méthodes d’analyses chimiques validées et adaptées aux exigences de ce travail,
nous avons pu confirmer la réactivité biologique du sol de Perpignan vis-à-vis de la
sulcotrione. Ce sol se caractérise par un potentiel de dégradation important, puisque 90%
environ de la substance active initiale est dégradée après 30 jours, en conditions de laboratoire
(temps de ½ vie de 8 à 10 jours). Aucune différence n’a été observée entre des prélèvements
d’origine identique mais présentant des historiques de traitement différents. Parallèlement,
une étude comparable menée sur ces mêmes sols, préalablement stérilisés, a montré une
dégradation négligeable, confirmant le rôle prépondérant de la biodégradation.
Le précédent travail (H. Chaabane) faisait état, à titre de perspective, de l’éventualité de
l’installation d’un phénomène de biodégradation accélérée, lié à un processus adaptatif de la
microflore indigène consécutivement à des traitements répétés. Notre étude a donc consisté à
exposer les sols de Perpignan à quatre traitements successifs en microcosmes, afin de
caractériser d’éventuelles différences au niveau des profils cinétiques de dissipation de la
substance active. Quels qu’aient été le nombre de traitements et l’historique des sols non
stérilisés, ces profils cinétiques ont tous été très comparables. Dans nos conditions
opératoires, l’apparition d’une biodégradation accélérée de la sulcotrione n’a donc pas été
observée. Ce résultat va dans le sens de données bibliographiques qui font état de la nécessité
de traitements répétés sur des périodes beaucoup plus longues et/ou avec des doses
d’application nettement supérieures avant d’observer la mise en place d’une biodégradation
accélérée à l’échelle de la parcelle agricole. Par ailleurs, l’observation des profils de

Conclusion générale et perspectives
169
dégradation, et notamment l’absence de délai de latence en début d’incubation, orientent vers
une approche co-métabolique du phénomène de biodégradation dans ce sol.
La réactivité biologique intrinsèque des sols de Perpignan (4S) et (C) a suggéré la possibilité
d’y trouver des souches microbiennes dotées de capacités génétiques à dégrader plus ou
moins spécifiquement la sulcotrione. Afin de contourner les difficultés inhérentes à la
problématique d’isolement de souches telluriques sur milieu synthétique, nous avons donc
entrepris une approche cultivable originale, s’appuyant sur l’utilisation combinée de divers
milieux de culture, dans le but de sélectionner, à partir du sol (4S), des isolats potentiellement
biodégradants. Cette stratégie expérimentale nous a permis d’isoler huit souches sulcotrione-
résistantes, parmi lesquelles une seule s’est avérée capable de métaboliser la sulcotrione. A
notre connaissance, il s’agit de la première description d’une telle souche, cultivable en
laboratoire et utilisant cet herbicide comme source de carbone et/ou d’énergie. Cet isolat
bactérien, dénommé 1OP lors de nos essais, a été formellement identifié, après séquencage de
l’ADNr 16S, comme étant affilié au genre Pseudomonas, espèce putida. Cette souche, apte à
utiliser la sulcotrione en culture liquide, a présenté un potentiel de dégradation élevé : 88 %
de la sulcotrione initiale a été dégradé au bout de 30 jours de culture, le temps de ½ vie étant
estimé à environ 15 jours. Contrairement aux données relevées lors de l’étude en
microcosmes de sol, la dissipation de la substance active correspond dans ce cas à un
processus de métabolisme direct, une phase de latence (5-6 jours) ayant été clairement mise
en évidence lors de la dégradation de la sulcotrione, qui, par ailleurs, constitue la seule source
de carbone et d’énergie du milieu. Dès la fin de cette période de latence, la cinétique observée
montre l’apparition concomitante de l’acide 2-chloro-4-méthylsulfonyl benzoïque (CMBA),
un des métabolites majeurs connus. Son augmentation est constante entre 6 et 27 jours, le
rapport molarité en CMBA / molarité initiale en sulcotrione atteignant 44 % à l’issue de
l’étude.
En revanche, la 1,3-cyclohexanedione (CHD) n’a jamais été détectée au cours de la période
de culture. Le processus de dégradation peut, dès lors, être interprété comme une attaque
ciblée de la souche Pseudomonas sp. 1OP au niveau de la partie CHD de la molécule de
sulcotrione, avec utilisation de cette partie comme source de carbone et/ou d’énergie.

Conclusion générale et perspectives
170
Les données recueillies au cours de notre travail vont dans le sens de la littérature,
relativement fournie, soulignant le large potentiel catabolique du genre Pseudomonas ainsi
que son rôle largement décrit dans le domaine des biodégradations. Cependant, des études
récentes ont mis en évidence l’intervention d’un autre genre bactérien (Bacillus) dans la
biodégradation de la mésotrione, herbicide également de la famille des tricétones ; cette
souche de Bacillus sp. s’est avérée incapable de métaboliser la sulcotrione. Ce travail entraine
également une réflexion sur la place de la souche Pseudomonas sp.1OP vis-à-vis de la
diversité des communautés microbiennes du sol étudié.
Notre étude confirme également la représentativité du Phylum des Proteobacteria au niveau
de l’ensemble des souches bactériennes telluriques cultivables présentes au sein de
l’écosystème sol.
Parmi les sept autres souches bactériennes isolées du sol et sulcotrione-résistantes, mais
incapables de dégrader la sulcotrione, l’isolat 1TRANS présentait un morphotype très
semblable à celui de 1OP sur milieu gélosé. Il a été retenu comme « contrôle » comparatif lors
de l’identification phylogénétique et a été identifié comme appartenant également au genre
Pseudomonas espèce putida.
La mise en évidence d’un phénotype dégradant la sulcotrione a mis en lumière
l’extraordinaire plasticité génétique d’une fraction de la microflore du sol génétiquement apte
à dégrader certains produits phytopharmaceutiques. De plus, la caractérisation de deux
souches appartenant à la même espèce, mais présentant des phénotypes différents en ce qui
concerne le caractère dégradant vis-à-vis de l’herbicide, souligne le caractère versatile de ce
potentiel de biodégradation. Ce résultat nous amène à proposer l’intervention d’éléments
génétiques mobiles régulant ces mécanismes moléculaires adaptatifs, via la présence de
plasmides conjugatifs et/ou d’éléments transposables (séquences d’insertion ou transposons)
codant les enzymes cataboliques chez Pseudomonas sp. 1OP, intervention déjà décrite pour
diverses souches dans le cas de l’atrazine. Cette perspective évolutive, en attente de
confirmation par séquençage, va cependant dans le sens des premières données recueillies lors
notre étude fonctionnelle, effectuée sur les isolats 1OP et 1TRANS, par ciblage des gènes
codant les enzymes impliquées dans le métabolisme de la voie catécholique. La mise en
évidence, au niveau génomique, d’une réponse différente entre l’isolat 1OP et l’isolat

Conclusion générale et perspectives
171
1TRANS lors du ciblage du gène codant la catéchol 2,3 dioxygénase, pourrait étayer cette
hypothèse de travail. De plus, les gènes codant la voie de catabolisme catécholique méta sont
présents chez Pseudomonas sp. et sont, dans la majorité des cas, portés par des plasmides
conjugatifs de taille supérieure à 50 kb, eux-mêmes situés à l’intérieur d’éléments
transposables.
Perspectives de travail Ces résultats expérimentaux nous ont donc permis d’approfondir l’état des connaissances sur
la dégradation biologique de la sulcotrione dans un sol agricole, mais également d’ouvrir
certaines perspectives de travail, tant dans le domaine fondamental que dans un domaine plus
appliqué pouvant conduire à une valorisation technologique.
L’isolement des éléments génétiques mobiles intervenant dans le processus de biodégradation
constituerait une première étape incontournable avant de les identifier, de la façon la plus
formelle possible, et de caractériser les mécanismes moléculaires impliqués, par
l’intermédiaire d’une approche de génomique évolutive. L’évaluation de la mobilité de ces
gènes cataboliques et leur rôle potentiel au niveau du transfert génétique horizontal au sein de
la microflore tellurique constituerait une avancée capitale dans la valorisation de ce travail.
Notre étude n’a pu que proposer des hypothèses concernant les voies métaboliques impliquées
dans la biodégradation de la sulcotrione par Pseudomonas sp.1OP en culture liquide. Ce
travail pourrait être complété, dans ce sens, par la combinaison de l’exploitation des données
du séquençage des gènes codant la catéchol 2,3-dioxygénase et l’utilisation de techniques de
mutagénèse dirigée par transposons à partir de notre souche. L’étude parallèle des cinétiques
enzymatiques à partir des phénotypes mutants obtenus aiderait également à mieux
appréhender les voies de régulation métaboliques.

Conclusion générale et perspectives
172
Sur un plan purement cultivable, le pouvoir dégradant de l’isolat 1OP pourrait être testé, dans
un premier temps, sur les métabolites identifiés de l’herbicide, mais également sur divers
composés plus ou moins proches chimiquement de la sulcotrione, et, à terme, sur certains
cocktails de xénobiotiques.
Une étape supplémentaire de valorisation serait l’exploitation « conditionnelle » de la souche
à des fins de dépollution environnementale.

173
Références Bibliographiques

Références Bibliographiques
174
Acinas, Silvia G., Sarma-Rupavtarm, R., Klepac-Ceraj, V., et Polz, M. F. (2005). PCR-
induced sequence artifacts and bias: insights from comparison of two 16S rRNA clone
libraries constructed from the same sample. Applied Environmental Microbiology, 71, 8966-
8969.
Aemprapa, S. et Williams, P A (1998). Implications of the xylQ gene of TOL plasmid pWW102
for the evolution of aromatic catabolic pathways. Microbiology (Reading, Engl.), 144 (Pt 5),
1387-1396.
Aislabie, J. M., Richards, N. K., et Boul, H. L. (1997). Microbial degradation of DDT - and its
residues- A review. New Zealand Journal of Agricultural Research, 40, 269.
Alexander, M. (1994). Biodegradation and bioremediation. Academic Press, A division of
Harcourt Brace & Company. New York.
Alexander, M. (1985). Biodegradation of organic chemicals. Environmental Science &
Technology, 19, 106-111.
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., et Lipman, D. J. (1990). Basic local
alignment search tool. Journal of Molecular Biology, 215, 403-410.
Anderson, J. P. E. (1984). Herbicide degradation in soil: Influence of microbial biomass. Soil
Biology and Biochemistry, 16, 483-489.
Anderson, J. P. E., et Lafuerza, A. (1992). Microbiological aspects of accelerated pesticide
degradation. In: Anderson, J.P.E., Arnold, D.J., Lewis, F., Torstensson, L. (Eds.),
Proceedings of the International Symposium on Environmental Aspects of Pesticide
Microbiology. Department of Microbiology, Swedish University of Agricultural Sciences,
Uppsala, pp. 184-192.
Anderson, K. L., Wheeler, K. A., Robinson, J. B., Tuovinen, O. H., (2002). Atrazine
mineralization in two wetlands. Water Research, 36, 4785-4794.
Anzai, Y., Kim, H., Park, J. Y., Wakabayashi, H., et Oyaizu, H. (2000). Phylogenetic
affiliation of the Pseudomonads based on 16S rRNA sequence. International Journal of
Systematic and Evolutionary Microbiology, 50 Pt 4, 1563-1589.
Arbeli, Z., et Fuentes, C. L. (2007). Accelerated biodegradation of pesticides: An overview of the
phenomenon, its basis and possible solutions; and a discussion on the tropical dimension.
Crop Protection, 26 1733-1746.

Références Bibliographiques
175
Arciero, D. M. et Lipscomb, J. D. (1986a). Binding of 17O-labeled substrate and inhibitors to
protocatechuate 4,5-dioxygenase-nitrosyl complex. Evidence for direct substrate binding to
the active site Fe2+ of extradiol dioxygenases. Journal of Biological Chemistry, 261, 2170 -
2178.
Arciero, D. M. et Lipscomb, J. D. (1986). Binding of 17O-labeled substrate and inhibitors to
protocatechuate 4,5-dioxygenase-nitrosyl complex. Journal of Biological Chemistry, 261,
2170–2178.
Artetxe, U., Hernández, A., García-Plazaola, J. I., et Becerril, J. M. (2006). Do light
acclimation mechanisms reduce the effects of light-dependent herbicides in Duckweed
(Lemna minor)? Weed Science, 54, 230-236.
Ashelford, K. E., Day, M. J., Bailey, Mark J., Lilley, Andrew K., et Fry, J. C. (1999). In Situ
Population Dynamics of Bacterial Viruses in a Terrestrial Environment. Applied
Environmental Microbiology, 65, 169-174.
Ashelford, K. E., Norris, S. J., Fry, J. C., Bailey, Mark J., et Day, M. J. (2000). Seasonal
population dynamics and interactions of competing bacteriophages and their host in the
rhizosphere. Applied Environmental Microbiology, 66, 4193-4199.
Ashelford, K. E., Day, M. J., et Fry, J. C. (2003). Elevated abundance of aacteriophage infecting
bacteria in Soil. Applied Environmental Microbiology, 69, 285-289.
Assinder, S. J., et Williams, P. A. (1990). The TOL plasmids: determinants of the catabolism of
toluene and the xylenes. Academic Press, p. 1-69.
Audus, L. J. (1949). The biological detoxication of 2,4-dichlorophenoxyacetic acid in soil. Plant
Soil, 2, 31-36.
Audus, L. J., (1951). The biological detoxification of hormone herbicides in soil. Plant Soil, 3,
170-192.
Baer, U. et Calvet, R. (1999). Fate of soil applied herbicides: Experimental data and prediction of
dissipation kinetics. Journal of Environmental Quality, 28, 1765-1777.
Bajaj, A., Pathak, A., Mudiam, M. R., Mayilraj, S., et Manickam, N. (2010). Isolation and
characterization of a Pseudomonas sp. strain IITR01 capable of degrading α-endosulfan and
endosulfan sulfate. Journal of Applied Microbiology, 109, 2135-2143.
Bakken, L.R. (1997). Culturable and nonculturable bacteria in soil. Modern Soil Microbiology.
eds van Elsas, J. D., Trevors, J. T. et Wellington, E. M. H., pp. 47–61. Marcel Dekker, New
York.

Références Bibliographiques
176
Baker, P. B. et Woods, D. R. (1977). Co-metabolism of the Ixodicide Amitraz. Journal of
Applied Microbiology, 42, 187-196.
Baker, M. D., et Mayfield, C. I. (1980). Microbial and non-biological décomposition of
chlorophenols and phénol in soil. Water Air Soil Pollution, 13, 411-424.
Bandounas, L., Ballerstedt, H., de Winde, J. H., et Ruijssenaars, H. J. (2011). Redundancy in
putrescine catabolism in solvent tolerant Pseudomonas putida S12. Journal of Biotechnology,
154, 1-10.
Barrere, C., Bastide, J., et Coste, C. M. (1988). Relations entre la vitesse de dégradation du
propyzamide et les propriétés physicochimiques des sols. Weed Research, 28, 93-99.
Barriuso, E. et Houot, S. (1996). Rapid meneralization of the s-triazine ring of atrazine in soils in
relation to soil management. Soil Biology and Biochemistry, 28, 1341-1348.
Barta, I. C. et Böger, P. (1996). Purification and characterization of 4-Hydroxyphenylpyruvate
dioxygenase from maize. Pesticide Science, 48, 109-116.
Bartilson, M., Nordlund, I., et Shingler, V. (1990). Location and organization of the
dimethylphenol catabolic genes of Pseudomonas CF600. Molecular and General Genetics,
220.
Batisson, I., Crouzet, O., Besse-Hoggan, P., Sancelme, M., Mangot, J.-F., Mallet, C., et
Bohatier, J. (2009). Isolation and characterization of mesotrione-degrading Bacillus sp. from
soil. Environmental Pollution, 157, 1195-1201.
Beaudegnies, R., Edmunds, A. J. F., Fraser, T. E. M., Hall, R. G., Hawkes, T. R., Mitchell,
G., Schaetzer, J., Wendeborn, S., et Wibley, J. (2009). Herbicidal 4-
hydroxyphenylpyruvate dioxygenase inhibitors. A review of the triketone chemistry story
from a Syngenta perspective. Bioorganic & Medicinal Chemistry, 17, 4134-4152.
Bergstrom, C. T., Lipsitch, M., et Levin, B. R. (2000). Natural selection, infectious transfer and
the existence conditions for bacterial plasmids. Genetics, 155, 1505-1519.
Brownlee, J. M., Johnson-Winters, K., Harrison, D. H. T., et Moran, G. R. (2004). Structure
of the ferrous form of (4-Hydroxyphenyl)pyruvate dioxygenase from Streptomyces avermitilis
in complex with the therapeutic herbicide, NTBC. Biochemistry, 43, 6370-6377.
de Bruijn, F. J. (1992). Use of repetitive (repetitive extragenic palindromic and enterobacterial
repetitive intergeneric consensus) sequences and the polymerase chain reaction to fingerprint
the genomes of Rhizobium meliloti isolates and other soil bacteria. Applied Environmental
Microbiology, 58, 2180-2187.

Références Bibliographiques
177
Burrus, V. et Waldor, M. K. (2004). Shaping bacterial genomes with integrative and conjugative
elements. Research in Microbiology, 155, 376-386.
Cantier, J. M., Bastide, J., et Coste, C. M. (1986). Structure degradability relationships for
propyzamide analogues in soil. Pesticide Science, 17, 235-241.
Cao, B., Geng, A., et Loh, K.-C. (2008). Induction of ortho- and meta-cleavage pathways in
Pseudomonas in biodegradation of high benzoate concentration: MS identification of
catabolic enzymes. Applied Microbiology Biotechnology, 81, 99-107.
Cao, B., Nagarajan, K., et Loh, K.-C. (2009). Biodegradation of aromatic compounds: current
status and opportunities for biomolecular approaches. Applied Microbiology Biotechnology
85, 207-228.
Castillo, M. del P. et Torstensson, L. (2007). Effect of Biobed Composition, Moisture, and
Temperature on the Degradation of Pesticides. Journal of Agricultural and Food Chemistry,
55, 5725-5733.
Chaabane, H., Vulliet, E., Joux, F., Lantoine, F., Conan, P., Cooper, J.-F., et Coste, C.-M.
(2007). Photodegradation of sulcotrione in various aquatic environments and toxicity of its
photoproducts for some marine micro-organisms. Water Research, 41, 1781-1789.
Chaabane, H. (2005). Etude de la stabilité et du comportement de la sulcotrione, herbicide du
maïs, dans l’environnement: eaux et sols agricoles. Thèse de doctorat. Université de
Perpignan Via Domitia.
Chaabane, H., Cooper, J.-F., Azouzi, L., et Coste, C.-M. (2005). Influence of Soil Properties on
the Adsorption−Desorption of Sulcotrione and Its Hydrolysis Metabolites on Various Soils.
Journal of Agricultural and Food Chemistry, 53, 4091-4095.
Chaabane, H., Vulliet, E., Calvayrac, C., Coste, C., et Cooper, J.-F. (2008). Behaviour of
sulcotrione and mesotrione in two soils. Pest Management Science, 64, 86-93.
Chang, J.-S., Chou, C., Lin, Y.-C., Lin, P.-J., Ho, J.-Y., et Lee Hu, T. (2001). Kinetic
characteristics of bacterial azo-dye decolorization by Pseudomonas luteola. Water Research,
35, 2841-2850.
Chanika, E., Georgiadou, D., Soueref, E., Karas, P., Karanasios, E., Tsiropoulos, N. G.,
Tzortzakakis, E. A., et Karpouzas, D. G. (2011). Isolation of soil bacteria able to hydrolyze
both organophosphate and carbamate pesticides. Bioresource Technology, 102, 3184-3192.

Références Bibliographiques
178
Chapman, R. A. et Harris, C. R. (1990). Enhanced degradation of insecticides in soil: Factors
influencing the development and effects of enhanced microbial activity. Racke, K.D., Coats,
J.R. Enhanced Biodegradation of Pesticides in the Environment. American Chemical Society,
Washington, DC.
Chatterjee, D K, Kellogg, S. T., Hamada, S., et Chakrabarty, A. M. (1981). Plasmid specifying
total degradation of 3-chlorobenzoate by a modified ortho pathway. J. Bacteriol, 146, 639-
646.
Chatterjee, Deb K. et Chakrabarty, A. M. (1982). Genetic rearrangements in plasmids
specifying total degradation of chlorinated benzoic acids. Molecular General Genetics, 188,
279-285.
Chèneby, D., Perrez, S., Devroe, C., Hallet, S., Couton, Y., Bizouard, F., Iuretig, G., Germon,
J. C., et Philippot, L. (2004). Denitrifying bacteria in bulk and maize-rhizospheric soil:
diversity and N2O-reducing abilities. Canadian Journal of Microbiology, 50, 469-474.
Cherrier, R., Perrin-Ganier, C., et Schiavon, M. (2004). Degradation of sulcotrione in a brown
soil amended with various organic matters. Agronomie, 24, 5.
Chung, K. Y. et Ou, L. T. (1996). Degradation of fenamiphos sulfoxide and fenamiphos sulfone
in soil with a history of continuous applications of fenamiphos. Archives of Environmental
Contamination and Toxicology, 30, 452-458.
Cohan, F. M. (2002). What are bacterial species ? Annu. Rev. Microbiol., 56, 457-487.
Coleman, D. C., et Crossley, D. A. (1996). Fundamentals of Soil Ecology. Academic Press,
London.
Compagnon, J. M., et Beraud, J. M. (1992). Journées Internationales sur la lutte contre les
mauvaises herbes, Versailles, France. 351-356.
Commissariat général au développement durable. (2010). Service de l'observation et des
statistiques. L’environnement en France. L’eau. In Références.
Connon, S. A. et Giovannoni, S. J. (2002). High-throughput methods for culturing
microorganisms in very-low-nutrient media yield diverse new marine isolates. Applied
Environmental Microbiology, 68, 3878-3885.
Cools, K., Bulcke, R., et Callens, D. (1997). Response of replacement crop to soil-applied
sulcotrione. Med. Fac. Landbouw. Univ. Gent., 64,721-726.

Références Bibliographiques
179
Costas, M., Mehn, M. P., Jensen, M. P., et Que, L., Jr (2004). Dioxygen activation at
mononuclear nonheme iron active sites: enzymes, models, and intermediates. Chemistry
Review, 104, 939-986.
Cox, L., Walker, Allan, et Welch, Sarah J (1996). Evidence for the accelerated degradation of
isoproturon in soils. Pesticide Science, 48, 253-260.
Crouzet, O., Batisson, I., Besse-Hoggan, P., Bonnemoy, F., Bardot, C., Poly, F., Bohatier, J.,
et Mallet, C. (2010). Response of soil microbial communities to the herbicide mesotrione: A
dose-effect microcosm approach. Soil Biology and Biochemistry, 42, 193-202.
Curtis, T. P., Sloan, W. T., et Scannell, J. W. (2002). Estimating prokaryotic diversity and its
limits. Proceedings of the National Academy of Sciences, 99, 10494 -10499.
Dagley S. et Gibson D. T. (1965). The bacterial degradation of catechol.
Dagley, S. (1986). Biochemistry of aromatic hydrocarbon degradation in Pseudomonads. In, The
bacteria. C. Gunsalus, J. R. Sokatch, and L. N. Omston, New-York, p. 527-555.
Dalton, H., Stirling, D. I., et Quayle, J. R. (1982). Co-Metabolism [and Discussion].
Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences,
297, 481-496.
Dangel, W., Tschech, A., et Fuchs, G. (1989). Enzyme reactions involved in anaerobic
cyclohexanol metabolism by a denitrifying Pseudomonas species. Archives of Microbiology,
152, 271-279.
Daugherty, D. D. et Karel, S. F. (1994). Degradation of 2,4-dichlorophenoxyacetic acid by
Pseudomonas cepacia DBO1(pRO101) in a dual-substrate chemostat. Appl. Environ.
Microbiol, 60, 3261-3267.
Daughton, C. G. et Hsieh, D. P. H. (1977). Parathion utilization by bacterial symbionts in a
chemostat. Applied Environmental Microbiology, 34, 175-184.
Davis, K. E. R., Joseph, S. J., et Janssen, P. H. (2005). Effects of growth medium, inoculum
size, and incubation time on culturability and isolation of soil bacteria. Applied Environmental
Microbiology, 71, 826-834.
Davis, K. E. R., Sangwan, P., et Janssen, P. H. (2011). Acidobacteria, Rubrobacteridae and
Chloroflexi are abundant among very slow-growing and mini-colony-forming soil bacteria.
Environmental Microbiology, 13, 798-805.
Davison, J. (1999). Genetic Exchange between Bacteria in the Environment. Plasmid, 42, 73-91.

Références Bibliographiques
180
Dayan, F. E., Duke, S. O., Sauldubois, A., Singh, N., McCurdy, C., et Cantrell, C. (2007). p-
Hydroxyphenylpyruvate dioxygenase is a herbicidal target site for [beta]-triketones from
Leptospermum scoparium. Phytochemistry, 68, 2004-2014.
Dayan, F. E., Singh, N., McCurdy, C. R., Godfrey, C. A., Larsen, L., Weavers, R. T., Van
Klink, J. W., et Perry, N. B. (2009). β -Triketone inhibitors of plant p-
Hydroxyphenylpyruvate Dioxygenase: modeling and comparative molecular field analysis of
their interactions. Journal of Agricultural and Food Chemistry, 57, 5194-5200.
Devers, M., Soulas, Guy, et Martin-Laurent, Fabrice (2004). Real-time reverse transcription
PCR analysis of expression of atrazine catabolism genes in two bacterial strains isolated from
soil. Journal of Microbiological Methods, 56, 3-15.
Devers, M., Henry, S., Hartmann, A., et Martin-Laurent, F. (2005). Horizontal gene transfer
of atrazine-degrading genes (atz) from Agrobacterium tumefaciens St96-4 pADP1::Tn5 to
bacteria of maize-cultivated soil. Pest Management Science, 61, 870-880.
Devers, M., Azhari, N. E., Kolic, N., et Martin-Laurent, F. (2007). Detection and organization
of atrazine-degrading genetic potential of seventeen bacterial isolates belonging to divergent
taxa indicate a recent common origin of their catabolic functions. FEMS Microbiology
Letters, 273, 78-86.
Devers, M., Rouard, N., et Martin-Laurent, Fabrice (2007). Genetic rearrangement of the
atzAB atrazine-degrading gene cassette from pADP1::Tn5 to the chromosome of Variovorax
sp. MD1 and MD2. Gene, 392, 1-6.
Devers, M., Rouard, N., et Martin-Laurent, F. (2008). Fitness drift of an atrazine-degrading
population under atrazine selection pressure. Environmental Microbiology, 10, 676-684.
Devers-Lamrani, M. (2008). Etude des mécanismes à l’origine de la dispersion des gènes codant
les enzymes responsables de la minéralisation de l’atrazine au sein de la microflore des sols.
Présentation des activités de recherche, Prix Phytopharma 2008.
Dick, R. E. et Quinn, J. P. (1995). Glyphosate-degrading isolates from environmental samples:
occurrence and pathways of degradation. Applied Microbiology and Biotechnology, 43, 545-
550.
Doublet, J., Mamy, L., et Barriuso, Enrique (2009). Delayed degradation in soil of foliar
herbicides glyphosate and sulcotrione previously absorbed by plants: Consequences on
herbicide fate and risk assessment. Chemosphere, 77, 582-589.

Références Bibliographiques
181
Dunbar, J., Barns, S. M., Ticknor, L. O., et Kuske, C. R. (2002). Empirical and theoretical
bacterial diversity in four Arizona soils. Applied Environmental Microbiology, 68, 3035-3045.
Dunbar, J., Takala, S., Barns, S. M., Davis, J. A., et Kuske, C. R. (1999). Levels of bacterial
community diversity in four arid soils Compared by cultivation and 16S rRNA gene cloning.
Applied Environmental Microbiology., 65, 1662-1669.
Durand, S, Amato, P., Sancelme, M, Delort, A-M, Combourieu, B, et Besse-Hoggan, P
(2006a). First isolation and characterization of a bacterial strain that biotransforms the
herbicide mesotrione. Letters in Applied. Microbiology, 43, 222-228.
Durand, St., Légeret, B., Martin, A.-S., Sancelme, M., Delort, A.-M., Besse-Hoggan, P. et
Combourieu, Bruno (2006b). Biotransformation of the triketone herbicide mesotrione by a
Bacillus strain. Metabolite profiling using liquid chromatography/electrospray ionization
quadrupole time-of-flight mass spectrometry. Rapid Communications in Mass Spectrometry,
20, 2603-2613.
Durand, S., Sancelme, M., Besse-Hoggan, P., et Combourieu, B. (2010). Biodegradation
pathway of mesotrione: Complementarities of NMR, LC-NMR and LC-MS for qualitative
and quantitative metabolic profiling. Chemosphere, 81, 372-380.
Dyson, J. S., Beulke, S., Brown, C. D., et Lane, M. C. G. (2002). Adsorption and degradation of
the weak acid mesotrione in soil and environmental fate implications. In Organic componds in
the environment. Journal of Environmental Quality, 31, 613-618.
El Sebai, T. (2004). Biodégradation accélérée de l’isoproturon: un phénomène présentant une
variabilité spatiale et catalysé par des microorganismes dégradants spécifiques. Thèse de
doctorat. Université de Perpignan Via Domitia.
El Sebai, T., Lagacherie, B., Soulas, Guy, et Martin-Laurent, F. (2004). Isolation and
characterisation of an isoproturon-mineralising Methylopila sp. TES from French agricultural
soil. FEMS Microbiology Letters, 239, 103-110.
Ellis, M. K., Whitfield, A. C., Gowans, L. A., Auton, T., Provan, W. M., Lock, E. A., et Smith,
L. L. (1995). Inhibition of 4-hydroxyphenylpyruvate dioxygenase by 2-(2-nitro-4-
trifluoromethylbenzoyl)-cyclohexane-1,3- dione and 2-(2-chloro-4-methanesulfonylbenzoyl)-
cyclohexane-1,3- dione. Toxicology and Applied Pharmacology, 133, 12-19.
Ellis, R. J., Morgan, P., Weightman, A. J., et Fry, J. C. (2003). Cultivation-dependent and -
independent approaches for determining bacterial diversity in heavy-metal-contaminated soil.
Applied Environmental Microbiology, 69, 3223-3230.

Références Bibliographiques
182
Eltis, L. et Bolin, J. (1996). Evolutionary relationships among extradiol dioxygenases. Journal of
Bacteriology, 178, 5930-5937.
Entry, J. A. et Emmingham, W. H. (1995). The Influence of dairy manure on atrazine and 2,4-
dichlorophenoxyacetic acid mineralization in pasture soils. Canadian Journal of Soil Science,
75, 379-383.
Erickson, B. D., et Mondello, F. J. (1993). Enhanced biodégradation of polychlorinated biphenyls
after site-directed mutagenesis of a biphenyl dioxygenase gene. Applied and Environmental
Microbiology, 59, 3858-3862.
Falk, J., Andersen, G., Kernebeck, B., et Krupinska, K. (2003). Constitutive overexpression of
barley 4-hydroxyphenylpyruvate dioxygenase in tobacco results in elevation of the vitamin E
content in seeds but not in leaves. FEBS Letters, 540, 35-40.
Felsenstein, J. (1973). Maximum-likelihood estimation of evolutionary trees from continuous
characters. American Journal Human Genetics, 25, 471-492.
Felsenstein, J. (1981). Evolutionary trees from gene frequencies and quantitative characters:
finding maximum likelihood estimates. Evolution, 35, 1229-1242.
Felsenstein, J. (1985). Confidence limits on phylogenies: An approach using the bootstrap.
Evolution, 39, 783-791.
Felsenstein, J. et Kishino, H. (1993). Is There Something Wrong with the Bootstrap on
Phylogenies? A Reply to Hillis and Bull. Systematic Biology, 42, 193-200.
Felsot, A., Maddox, J. V., et Bruce, W. (1981). Enhanced microbial degradation of carbofuran in
soils with histories of Furadan® use. Bulletin of Environmental Contamination and
Toxicology, 26, 781-788.
Ferrari, B. C., Binnerup, S. J., et Gillings, M. (2005). Microcolony cultivation on a soil substrate
membrane system selects for previously uncultured soil bacteria. Applied Environmental
Microbiology, 71, 8714-8720.
Ferreira, E. P. de B., Dusi, A. N., Costa, J. R., Xavier, G. R., et Rumjanek, N. G. (2009).
Assessing insecticide and fungicide effects on the culturable soil bacterial community by
analyses of variance of their DGGE fingerprinting data. European Journal of Soil Biology, 45,
466-472.
Field, J. A., Stams, A. J. M., Kato, M, et Schraa, G. (1995). Enhanced biodégradation of
aromatic pollutants in coculture of anaerobic and aérobic bacterial consortia. Antonie Van
Leeuwenhoek, 67, 47-77.

Références Bibliographiques
183
Fitch, W. M. et Margoliash, E. (1967). Construction of phylogenetic trees. Science, 155, 279-
284.
Flint, J. L. et Witt, W. W. (1997). Microbial degradation of imazaquin and imazethapyr. Weed
Science, 45, 586-591.
Fomsgaard, I. S. et Kristensen, K. (1999). Influence of microbial activity, organic carbon
content, soil texture and soil depth on mineralisation rates of low concentrations of 14C-
mecoprop-development of a predictive model. Ecological Modelling, 122, 45-68.
Fournier, J.-C., Morel-Chevillet, C., Catroux, C., et Arrault, S. (1998). La dégradation
accélérée des pesticides : contraintes et solutions. Pages 289-296. In 17ième conférence du
Columa, journées internationales de lutte contre les mauvaises herbes, Dijon, France.
Frey, P., Smith, J. J., Albar, L., Prior, P., Saddler, G. S., Trigalet-Demery, D., et Trigalet., A.
(1996). Bacteriocin typing of Burkholderia (Pseudomonas) solanacearum race 1 of the
French West Indies and correlation with genomic variation of the pathogen. Applied
Environmental Microbiology, 62, 473-479.
Frost, L. S., Leplae, R., Summers, A. O., et Toussaint, A. (2005). Mobile genetic elements : the
agents of open source evolution. Nature Reviews Microbiology, 3, 722-732.
Funchain, P., Yeung, A., Stewart, J., Clendenin, W. M., et Miller, J. H. (2001). Amplification
of mutator cells in a population as a result of horizontal transfer. Journal of Bacteriology, 183,
3737-3741.
García-González, V., Govantes, F., Porrúa, O., et Santero, E. (2005). Regulation of the
Pseudomonas sp. strain ADP cyanuric acid degradation operon. Journal of Bacteriology, 187,
155-167.
Garcia-Gonzalez, V., Govantes, F., Shaw, L. J., Burns, R. G., et Santero, E. (2003). Nitrogen
control of atrazine utilization in Pseudomonas sp. strain ADP. Applied Environmental
Microbiology, 69, 6987-6993.
Ghosal, D., In-Soon, Y., et Gunsalus, I. C. (1987). Nucleotide sequence and expression of gene
nahH of plasmid NAH7 and homology with gene xylE of TOL pWWO. Gene, 55, 19-28.
Giedraityte, G. et Kalėdienė, L. (2008). Catechol 1,2-dioxygenase from α -naphthol degrading
thermophilic Geobacillus sp. strain: purification and properties. Central European Journal of
Biology., 4, 68-73.
Goulding, C., Gillen, C. J., et Bolton, E. (1988). Biodegradation of substituted benzenes. Journal
of Applied Microbiology, 65, 1-5.

Références Bibliographiques
184
Gray, R. A. et Joo, G. K. (1985). Reduction in weed control after repeat applications of
thiocarbamate and other herbicides. Weed Science, 33, 698-702.
Grogan, G. (2002). [beta]-Diketone hydrolases. Journal of Molecular Catalysis B: Enzymatic, 19-
20, 73-82.
Grogan, G. (2005). Emergent mechanistic diversity of enzyme-catalysed β -diketone cleavage.
Biochem J, 388, 721-730.
Gurtler, V. et Stanisich, V. A. (1996). New approaches to typing and identification of bacteria
using the 16S-23S rDNA spacer region. Microbiology, 142, 3-16.
Guzik, U., Gren, I., Hupert-Kocurek, K., et Wojcieszynska, D. (2011). Catechol 1,2-
dioxygenase from the new aromatic compounds-degrading Pseudomonas putida strain N6.
International Biodeterioration & Biodegradation, 65, 504-512.
Hamzah, R. Y. et Al-Baharna, B. S. (1994). Catechol ring-cleavage in Pseudomonas cepacia: the
simultaneous induction of ortho and meta pathways. Applied Microbiology Biotechnology, 41,
250-256.
Harayama, S. et Rekik, M. (1989). Bacterial aromatic ring-cleavage enzymes are classified into
two different gene families. Journal of Biological Chemistry, 264, 15328 -15333.
Harayama, S., et Kasai, H. (2006). Bacterial phylogeny recon- struction from molecular
sequences. In Stackebrandt E. (ed). Molecular identification, systematics, and population
structure of Prokaryotes. Springer : New York, pp 105-140.
Harris, D., et Paul, E. A. (1994). Measurements of bacterial growth rates in soil. Applied Soil
Ecology. 1:277–290.
Harth, E., Romero, J., Torres, R., et Espejo, R. T. (2007). Intragenomic heterogeneity and
intergenomic recombination among Vibrio parahaemolyticus 16S rRNA genes. Microbiology,
153, 2640-2647.
Hatta, T., Mukerjee-Dhar, G., Damborsky, J., Kiyohara, H., et Kimbara, K. (2003).
Characterization of a novel thermostable Mn(II)-dependent 2,3-dihydroxybiphenyl 1,2-
dioxygenase from a polychlorinated biphenyl- and naphthalene-degrading Bacillus sp. JF8.
Journal of Biology Chemistry, 278, 21483-21492.
Hattori, T. (1973). Microbial Life in Soil. Marcel Dekker, New York.
Hellyer, R. O. (1968). The occurrence of ß-triketones in the steam-volatile oils of some
myrtaceous Australian plants. Australian Journal of Chemistry, 21, 2826-2828.

Références Bibliographiques
185
Herrera, A., Hery, M., Stach, J. E. M., Jaffre, T., Normand, P., et Navarro, E. (2007). Species
richness and phylogenetic diversity comparisons of soil microbial communities affected by
nickel-mining and revegetation efforts in New Caledonia. European Journal of Soil Biology,
43, 130-139.
Herrmann, Heidrun, Muller, C., Schmidt, I., Mahnke, J., Petruschka, L., et Hahnke, K.
(1995). Localization and organization of phenol degradation genes of Pseudomonas putida
strain H. Molecular and General Genetics, 247, 240-246.
Herrmann, H., Muller, C., et Lauf, U. (1998). The transposable elements resident on the
plasmids of Pseudomonas putida strain H, Tn ::5501 and Tn ::5502 , are cryptic transposons
of the Tn 3 family. Molecular and General Genetics MGG, 259, 674-678.
Heuer, H., Hartung, K., Wieland, G., Kramer, I., et Smalla, Kornelia (1999). Polynucleotide
Probes That Target a Hypervariable Region of 16S rRNA Genes To Identify Bacterial Isolates
Corresponding to Bands of Community Fingerprints. Applied Environmental Microbiology,
65, 1045-1049.
Heuer, H., Wieland, G., Schonfeld, J., Schnwalder, A., Gomes, N. C. M. et Smalla, K. (2001).
Bacterial community profiling using DGGE or TGGE analysis. Environmental Molecular
Microbiology: Protocols and Applications, ed. P.A. Rochelle. Horizon Scientific Press,
Wymondham, UK. pp. 177–190.
Hickey, W. J. et Focht, D. D. (1990). Degradation of mono-, di-, and trihalogenated benzoic acids
by Pseudomonas aeruginosa JB2. Applied Environmental Microbiology, 56, 3842-3850.
Hirose, J., Kimura, N., Suyama, A., Kobayashi, A., Hayashida, S., et Furukawa, K. (1994).
Functional and structural relationship of various extradiol aromatic ring-cleavage
dioxygenases of Pseudomonas origin. FEMS Microbiology Letters, 118, 273-277.
Horvath, R. S. (1972). Cometabolism of the herbicide, 2,3,6-trichlorobenzoate by natural
microbial populations. Bulletin of Environmental Contamination Toxicology, 7, 273-276.
Houot, Sabine, Barriuso, Enrique, et Bergheaud, V. (1998). Modifications to atrazine
degradation pathways in a loamy soil after addition of organic amendments. Soil Biology and
Biochemistry, 30, 2147-2157.
Huang, W., Yu, H., et Weber, W. J. (1998). Hysteresis in the sorption and desorption of
hydrophobic organic contaminants by soils and sediments: 1. A comparative analysis of
experimental protocols. Journal of Contaminant Hydrology, 31, 129-148.

Références Bibliographiques
186
Huang, M.-lan, Zou, J.-wei, Yang, D.-yah, Ning, B.-zhu, Shang, Z.-cai, et Yu, Q.-sen (2002).
Theoretical studies on tautomerism of benzoylcyclohexane-1,3-dione and its derivatives.
Journal of Molecular Structure: THEOCHEM, 589-590, 321-328.
Huelsenbeck, J. P. et Hillis, D. M. (1993). Success of Phylogenetic Methods in the Four-Taxon
Case. Systematic Biology, 42, 247 -264.
Hugenholtz, P, Goebel, B. M., et Pace, N. R. (1998). Impact of culture-independent studies on
the emerging phylogenetic view of bacterial diversity. Journal of Bacteriology, 180, 4765-
4774.
Hugenholtz, Philip (2002). Exploring prokaryotic diversity in the genomic era. Genome Biology,
3, reviews0003.
Hughes, J. B., Hellmann, J. J., Ricketts, T. H., et Bohannan, B. J. M. (2001). Counting the
uncountable: statistical approaches to estimating microbial diversity. Applied Environmental
Microbiology, 67, 4399-4406.
Hussain, S., Sørensen, Sebastian R., Devers-Lamrani, M., El-Sebai, T., et Martin-Laurent,
Fabrice (2009). Characterization of an isoproturon mineralizing bacterial culture enriched
from a French agricultural soil. Chemosphere, 77, 1052-1059.
Index Phytosanitaire. (2011). Herbicides, sulcotrione. 47ième édition de l’Association de
Coordination Technique Agricole, Paris., 440-441.
Ingham, E. R. et Horton, K. A. (1987). Bacterial, fungal and protozoan responses to chloroform
fumigation in stored soil. Soil Biology and Biochemistry, 19, 545-550.
Ishikuri, S. et Hattori, T. (1985). Formation of bacterial colonies in successive time intervals.
Appl. Environ. Microbiol., 49, 870-873.
Jain, R., Rivera, M. C., Moore, J. E., et Lake, J. A. (2002). Horizontal Gene Transfer in
Microbial Genome Evolution. Theoretical Population Biology, 61, 489-495.
Janke, D. et Fritsche, W. (1985). Nature and significance of microbial cometabolism of
xenobiotics. Journal of Basic Microbiology, 25, 603-619.
Janssen, P. H. (2006). Identifying the Dominant Soil Bacterial Taxa in Libraries of 16S rRNA and
16S rRNA Genes. Applied Environmental Microbiology, 72, 1719-1728.
Janssen, P. H., Yates, P. S., Grinton, B. E., Taylor, P. M., et Sait, M. (2002). Improved
culturability of soil bacteria and isolation in pure culture of novel members of the divisions
Acidobacteria, Actinobacteria, Proteobacteria, and Verrucomicrobia. Applied Environmental
Microbiology, 68, 2391-2396.

Références Bibliographiques
187
Jeenes, D. J., Reineke, W., Knackmuss, H. J., et Williams, P A (1982). TOL plasmid pWW0 in
constructed halobenzoate-degrading Pseudomonas strains: enzyme regulation and DNA
structure. Journal of Bacteriology, 150, 180-187.
Jimenez, J. I., Minambres, B., Garcia, J. L., et Diaz, E. (2002). Genomic analysis of the
aromatic catabolic pathways from Pseudomonas putida KT2440. Environmental
Microbiology, 4, 824-841.
Johnsen, K., et Nielsen, P. (1999). Diversity of Pseudomonas strains isolated with King’s B and
Gould’s S1 agar determined by repetitive extragenic palindromic-polymerase chain reaction,
16S rDNA sequencing and Fourier transform infrared spectroscopy characterisation. FEMS
Microbiological Letters, 173, 155–162.
Johnsen, K., Jacobsen, C., Torsvik, Vigdis, et Sorensen, J. (2001). Pesticide effects on bacterial
diversity in agricultural soils - a review. Biology and Fertility of Soils, 33, 443-453.
Joseph, S. J., Hugenholtz, Philip, Sangwan, P., Osborne, C. A., et Janssen, P. H. (2003).
Laboratory Cultivation of Widespread and Previously Uncultured Soil Bacteria. Applied
Environmental Microbiology, 69, 7210-7215.
Joshi, B. et Walia, S. (1996a). PCR amplification of catechol 2,3-dioxygenase gene sequences
from naturally occurring hydrocarbon degrading bacteria isolated from petroleum
hydrocarbon contaminated groundwater. FEMS Microbiology Ecology, 19, 5-15.
Joshi, B. et Walia, S. K. (1996b). Detection of metapyrocatechase homologous genes in
petroleum hydrocarbon contaminated groundwater by polymerase chain reaction. Journal of
Microbiological Methods, 27, 121-128.
Jukes, T. et Cantor, C. (1969). Evolution of protein molecules. Mammalian Protein Metabolism.
Jussila, M. M., Zhao, J., Suominen, L., et Lindström, K. (2007). TOL plasmid transfer during
bacterial conjugation in vitro and rhizoremediation of oil compounds in vivo. Environmental
Pollution, 146, 510-524.
Kaeberlein, T., Lewis, K., et Epstein, S. S. (2002). Isolating « uncultivable » microorganisms in
pure culture in a simulated natural environment. Science, 296, 1127-1129.
Kamanavalli, C. M., et Ninnekar, H. Z. (2000). Biodegradation of propoxur by Pseudomonas
species. World Journal of Microbiology and Biotechnology,16, 329-331.
Keil, H., Keil, S., Pickup, R. W., et Williams, P A (1985a). Evolutionary conservation of genes
coding for meta pathway enzymes within TOL plasmids pWW0 and pWW53. Journal of
Bacteriology, 164, 887-895.

Références Bibliographiques
188
Keil, H., Lebens, M. R., et Williams, P. A. (1985b). TOL plasmid pWW15 contains two
nonhomologous, independently regulated catechol 2,3-oxygenase genes. Journal of
Bacteriology,, 163, 248-255.
Keller, M. et Zengler, K. (2004). Tapping into microbial diversity. Nature Reviews Microbiology,
2, 141-150.
Kennedy, A. C., et Papendick, R. I. (1995). Microbial characteristics of soil quality. Journal of
Soil and Water Conservation, 50, 243-248.
Kennedy, A. C., et Smith, K. L. (1998). Soil microbial diversity and the sustainability of
agricultural soils. Plant and Soil, 170, 75-86.
Kilbane, J. J., Chatterjee, D K, et Chakrabarty, A. M. (1983). Detoxification of 2,4,5-
trichlorophenoxyacetic acid from contaminated soil by Pseudomonas cepacia. Applied
Environmental Microbiology, 45, 1697-1700.
Killham, K. (1994). Soil ecology. Cambridge University Press.
Kim, J.-S., Kim, T.-J., Kwon, O. K., et Cho, K. Y. (2002). Mechanism of action of sulcotrione, a
4-hydroxyphenylpyruvate dioxygenase inhibitor, in developed plant tissues. Photosynthetica,
40, 541-545.
Kita, A., Kita, S., Fujisawa, I., Inaka, K., Ishida, T., Horiike, K., Nozaki, Mitsuhiro, et Miki,
K. (1999). An archetypical extradiol-cleaving catecholic dioxygenase: the crystal structure of
catechol 2,3-dioxygenase (metapyrocatechase) from Pseudomonas putida mt-2. Structure, 7,
25-34.
Kitayama, A., Achioku, T., Yanagawa, T., Kanou, K., Kikuchi, M., Ueda, H., Suzuki, E.,
Nishimura, H., Nagamune, T., et Kawakami, Y. (1996a). Cloning and characterization of
extradiol aromatic ring-cleavage dioxygenases of Pseudomonas aeruginosa JI104. Journal of
Fermentation and Bioengineering, 82, 217-223.
Kitayama, A., Suzuki, E., Kawakami, Y., et Nagamune, T. (1996b). Gene organization and low
regiospecificity in aromatic-ring hydroxylation of a benzene monooxygenase of Pseudomonas
aeruginosa JI104. Journal of Fermentation and Bioengineering, 82, 421-425.
Knackmuss, H. J. (1981). Degradation of halogenated and sulfonated hydrocarbons. Academic
Press. Leisenger, T.; Cook, A.M.; Huller, R.; Nuesch, J.

Références Bibliographiques
189
Kok, R. G., Young, D. M., et Ornston, L. N. (1999). Phenotypic Expression of PCR-Generated
Random Mutations in a Pseudomonas putida Gene after Its Introduction into an Acinetobacter
Chromosome by Natural Transformation. Applied Environmental Microbiology, 65, 1675-
1680.
Kostal, J., Suchanek, M., Klierova, H., Demnerova, K., Kralova, B., et McBeth, D. L. (1998).
Pseudomonas C12B, an SDS degrading strain, harbours a plasmid coding for degradation of
medium chain length n-alkanes. International Biodeterioration & Biodegredation, 42, 221-
228.
Kukor, J. J. et Olsen, R. H. (1991). Genetic organization and regulation of a meta cleavage
pathway for catechols produced from catabolism of toluene, benzene, phenol, and cresols by
Pseudomonas pickettii PKO1. Journal of Bacteriology, 173, 4587-4594.
Kunz, D. A. et Chapman, P. J. (1981). Catabolism of pseudocumene and 3-ethyltoluene by
Pseudomonas putida (arvilla) mt-2: evidence for new functions of the TOL (pWWO)
plasmid. Journal of Bacteriology, 146, 179-191.
Küster, E., et Hattori, T. (1973). Microbial Life in the Soil; an Introduction. Marcel Dekker, Inc.
New York 1973, 427 S., 67 Tabellen, 272 Figuren im Text. Zeitschrift für Pflanzenernährung
und Bodenkunde, 136, 248-249.
Lederberg, E. M. (1952). Allelic Relationships and Reverse Mutation in Escherichia Coli.
Genetics, 37, 469-483.
Lee, D. L., Knudsen, C. G., Michaely, W. J., Chin, H., Nguyen, N. H., Carter, C. G.,
Cromartie, T. H., Lake, B. H., Shribbs, J. M., et Fraser, T. (1998). The structure–activity
relationships of the triketone class of HPPD herbicides. Pesticide Science, 54, 377-384.
Lee, J., Min, K. R., Kim, Y.-C., Kim, C.-K., Lim, J.-Y., Yoon, H., Min, K.-H., Lee, K.-S., et
Kim, Y. (1995). Cloning of salicylate hydroxylase gene and catechol 2,3-dioxygenase gene
and sequencing of an intergenic sequence between the two genes of Pseudomonas putida
KF715. Biochemical and Biophysical Research Communications, 211, 382-388.
Lilley, A. K. et Bailey, M. J. (1997). Impact of Plasmid pQBR103 Acquisition and Carriage on
the Phytosphere Fitness of Pseudomonas fluorescens SBW25: Burden and Benefit. Applied
Environmental Microbiology, 63, 1584-1587.
Lillis, L., Clipson, N., et Doyle, E. (2010). Quantification of catechol dioxygenase gene
expression in soil during degradation of 2,4-dichlorophenol. FEMS Microbiology Ecology,
73, 363-369.

Références Bibliographiques
190
Lin, Y. L., Wu, C. S., Lin, S. W., et Yang, D. Y. (2000). SAR studies of 2-o-substituted-benzoyl-
and 2-alkanoyl-cyclohexane-1,3-diones as inhibitors of 4-hydroxyphenylpyruvate
dioxygenase. Bioorganics and Medicinal Chemistry Letters, 10, 843-845.
Lindstedt, S., Holme, E., Lock, E. A., Hjalmarson, O., et Strandvik, B. (1992). Treatment of
hereditary tyrosinaemia type I by inhibition of 4-hydroxyphenylpyruvate dioxygenase. The
Lancet, 340, 813-817.
de Lipthay, J. R., Sørensen, Sebastian R., et Aamand, Jens (2007). Effect of herbicide
concentration and organic and inorganic nutrient amendment on the mineralization of
mecoprop, 2,4-D and 2,4,5-T in soil and aquifer samples. Environmental Pollution, 148, 83-
93.
Loh K.-C. et Chua S.-S. (2002). Ortho pathway of benzoate degradation in Pseudomonas putida:
induction of meta pathway at high substrate concentrations. Enzyme and Microbial
Technology, 30, 620-626.
Loh, K.-C. et Chua, S.-S. (2002). Ortho pathway of benzoate degradation in Pseudomonas putida:
induction of meta pathway at high substrate concentrations. Enzyme and Microbial
Technology, 30, 620-626.
Lorenz, M G et Wackernagel, W (1994). Bacterial gene transfer by natural genetic
transformation in the environment. Microbiology Reviews, 58, 563-602.
Lorenz, Michael G., Gerjets, D., et Wackernagel, Wilfried (1991). Release of transforming
plasmid and chromosomal DNA from two cultured soil bacteria. Archives of Microbiology,
156, 319-326.
Lynch, J. M. (1994). The rhizosphere : form and function. Applied Soil Ecology, 1, 193-198.
Majewski, J. et Cohan, F. M. (1999). DNA Sequence Similarity Requirements for Interspecific
Recombination in Bacillus. Genetics, 153, 1525 -1533.
Majewski, J., Zawadzki, P., Pickerill, P., Cohan, F. M., et Dowson, C. G. (2000). Barriers to
Genetic Exchange between Bacterial Species: Streptococcus pneumoniae Transformation.
Journal of Bacteriology, 182, 1016-1023.
Mamy, L. (2004). Comparaison des impacts environnementaux des herbicides à large spectre et
des herbicides sélectifs: Caractérisation de leur devenir dans le sol et modélisation. Thèse de
doctorat. Institut Agronomique National Paris-Grignon.
Mamy, L. et Barriuso, E. (2005). Glyphosate adsorption in soils compared to herbicides replaced
with the introduction of glyphosate resistant crops. Chemosphere, 61, 844-855.

Références Bibliographiques
191
Mamy, L., Barriuso, E., et Gabrielle, B. (2005). Environmental fate of herbicides trifluralin,
metazachlor, metamitron and sulcotrione compared with that of glyphosate, a substitute broad
spectrum herbicide for different glyphosate-resistant crops. Pest Management Science, 61,
905-916.
Mandelbaum, R., Allan, D., et Wackett, L. (1995). Isolation and Characterization of a
Pseudomonas sp. That Mineralizes the s-Triazine Herbicide Atrazine. Applied Environmental
Microbiology, 61, 1451-1457.
Martin-Laurent, F., Philippot, L., Hallet, S., Chaussod, R., Germon, J. C., Soulas, G., et
Catroux, G. (2001). DNA Extraction from Soils: Old Bias for New Microbial Diversity
Analysis Methods. Applied Environmental Microbiology, 67, 2354-2359.
Martínez-Murcia, A. J., Acinas, S. G, et Rodriguez-Valera, F. (1995). Evaluation of
prokaryotic diversity by restrictase digestion of 16S rDNA directly amplified from
hypersaline environments. FEMS Microbiology Ecology, 17, 247-255.
Mashimoyo, N., Aoyama, N., et Shiori, T. (1981). New methods and reagents in organic
synthesis. A simple efficient preparation of methyl esters with trimethylsilyldiazomethane
(TMSCHN2) and its application to gas chromatographic analysis of fatty acids. Chemical and
Pharmaceutical Bulletin, 29, 1475-1478.
Massol-Deya, A. A., Odelson, D. A., Hickey, R. F., et Tiedje, J. M. (1995). Bacterial community
fingerprinting of amplified 16S and 16S-23S ribosomal DNA gene sequences and restriction
endonuclease analysis (ARDRA). Molecular Microbial Ecology Manual. 3.3.2 : 1-8.
Mathieu, C. et Pieltain, F. (2003). Analyse chimique des sols; méthodes choisies. Londres, Paris,
New-York. Tec et Doc.
Matringe, M., Sailland, A., Pelissier, B., Rolland, A., et Zink, O. (2005). p-
Hydroxyphenylpyruvate dioxygenase inhibitor-resistant plants. Pest Management Science, 61,
269-276.
Mayonado, D. J., Hatzios, K. K., Orcutt, D. M., et Wilson, H. P. (1989). Evaluation of the
mechanism of action of the bleaching herbicide SC-0051 by HPLC analysis. Pesticide
Biochemistry and Physiology, 35, 138-145.
McGarvey, J. A., Miller, W. G., Sanchez, S., et Stanker, L. (2004). Identification of bacterial
populations in dairy wastewaters by use of 16S rRNA gene sequences and other genetic
markers. Applied Environmental Microbiology, 70, 4267-4275.

Références Bibliographiques
192
Meazza, G., Scheffler, B. E., Tellez, M. R., Rimando, A. M., Romagni, J. G., Duke, S. O.,
Nanayakkara, D., Khan, I. A., Abourashed, E. A., et Dayan, F. E. (2002). The inhibitory
activity of natural products on plant p-hydroxyphenylpyruvate dioxygenase. Phytochemistry,
60, 281-288.
Menna, P., Pereira, A. A., Bangel, E. V., et Hungria, M. (2009). Rep-PCR of tropical rhizobia
for strain fingerprinting, biodiversity appraisal and as a taxonomic and phylogenetic tool.
Symbiosis, 48, 120-130.
Michaely, W. J., et Kratz, G. W. (1986). Certains 2-(2-substituted benzoyl)-1,3-
cyclohexanediones. In European patent application 0135191, Stauffer Chemical Compagny,
assignée.
Mitchell, G., Bartlett, D. W., Fraser, T. E. M., Hawkes, T. R., Holt, D. C., Townson, J. K., et
Wichert, R. A. (2001). Mesotrione : a new selective herbicide for use in maize. Pest
Management Science, 57, 120-128.
Moiseeva, O. V., Solyanikova, I. P., Kaschabek, S. R., Gröning, J., Thiel, M., Golovleva, L.
A., et Schlömann, M. (2002). A New modified ortho cleavage pathway of 3-chlorocatechol
degradation by Rhodococcus opacus 1CP: genetic and biochemical evidence. Journal of
Bacteriology, 184, 5282-5292.
Moran, G. R. (2005). 4-Hydroxyphenylpyruvate dioxygenase. Archives of Biochemistry and
Biophysics, 433, 117-128.
Morel-Chevillet, C. (1998). La biodégradation accélérée croisée des méthylcarbamates
insecticides et nématicides. Possibilités d’interactions entre ces produits et avec certains
organophosphorés. Thèse de doctorat. Université de Lyon.
Morel-Chevillet, C., Parekh, N. S., Pautrel, D., et Fournier, J. (1996). Cross enhancement of
carbofuran biodegradation in soil samples previously treated with carbamate pesticides. Soil
Biology and Biochemistry, 28, 1767–1776.
Morgan, J. A. W., Bending, G. D., et White, P. J. (2005). Biological costs and benefits to plant–
microbe interactions in the rhizosphere. Journal of Experimental Botany, 56, 1729 -1739.
Moyer, C. L., Dobbs, F. C., et Karl, D. M. (1994). Estimation of diversity and community
structure through restriction fragment length polymorphism distribution analysis of bacterial
16S rRNA genes from a microbial mat at an active, hydrothermal vent system, Loihi
Seamount, Hawaii. Applied Environmental Microbiology, 60, 871-879.

Références Bibliographiques
193
Muyzer, G., et Smalla, K. (1998). Application of denaturing gradient gel electrophoresis (DGGE)
and temperature gradient gel electrophoresis (TGGE). in microbial ecology. Antonie van
Leeuwenhoeck, International Journal of General and Molecular Microbiology, 73, 127-141.
Nakai, C., Kagamiyama, H., Nozaki, M, Nakazawa, T., Inouye, S., Ebina, Y., et Nakazawa,
A. (1983). Complete nucleotide sequence of the metapyrocatechase gene on the TOI plasmid
of Pseudomonas putida mt-2. Journal of Biological Chemistry, 258, 2923 -2928.
Nakai, C., Uyeyama, H., Kagamiyama, H., Nakazawa, T., Inouye, S., Kishi, F., Nakazawa, A.,
et Nozaki, M. (1995). Cloning, DNA sequencing, and amino acid sequencing of catechol 1,2-
dioxygenases (pyrocatechase) from Pseudomonas putida mt-2 and Pseudomonas arvilla C-1.
Archives of Biochemistry and Biophysics, 321, 353-362.
Nannipieri, P., Ascher, J., Ceccherini, M. T., Landi, L., Pietramellara, G., et Renella, G.
(2003). Microbial diversity and soil functions. European Journal of Soil Science, 54, 655-670.
Nichols, D. (2007). Cultivation gives context to the microbial ecologist. FEMS Microbiology
Ecology, 60, 351-357.
Nielsen, K. M. et van Elsas, J. D. (2001). Stimulatory effects of compounds present in the
rhizosphere on natural transformation of Acinetobacter sp. BD413 in soil. Soil Biology and
Biochemistry, 33, 345-357.
Nielsen, Kaare M., van Elsas, Jan D., et Smalla, K. (2000). Transformation of Acinetobacter sp.
strain BD413(pFG4ΔnptII) with transgenic plant DNA in soil microcosms and effects of
kanamycin on selection of transformants. Applied Environmental Microbiology, 66, 1237-
1242.
Nojiri, H., Shintani, M., et Omori, T. (2004). Divergence of mobile genetic elements involved in
the distribution of xenobiotic-catabolic capacity. Applied Microbiology and Biotechnology,
64, 154-174.
Ochman, H., Lawrence, J. G., et Groisman, E. A. (2000). Lateral gene transfer and the nature of
bacterial innovation. Nature, 405, 299-304.
Ochman, H. (2005). Examining bacterial species under the specter of gene transfer and exchange.
Proceedings of the National Academy of Sciences, 102, 6595-6599.
OECD guideline. (2002). OECD guideline for testing of chemicals. Hydrolysis as a function of
pH (proposal for udapting guideline 111). 15 p.
Ogawa, N., Chackrabarty, A. M., et Zaborina, O. (2004). Degradative plasmid. In Plasmid
Biology. ASM Press. Funnell, B. E., Philipps, G. J., Washington, DC.

Références Bibliographiques
194
Otenio, M .H., da Silva, M. T. L., Marques, M. L. O., Roseiro, J. C., et Bidoia, E. D. (2005).
Benzene, toluène and xylene biodegradation by Pseudomonas putida CCMI 852, Brazilian
Journal of Microbiology, 36, 258-261.
Paget, E., Monrozier, L. J., et Simonet, Pascal (1992). Adsorption of DNA on clay minerals:
protection against DNaseI and influence on gene transfer. FEMS Microbiology Letters, 97,
31-39.
Pallett, K. E., Cramp, S. M., Little, J. P., Veerasekaran, P., Crudace, A. J., et Slater, A. E.
(2001). Isoxaflutole: the background to its discovery and the basis of its herbicidal properties.
Pest Management Science, 57, 133-142.
Pandey, G. et Jain, R. K. (2002). Bacterial Chemotaxis toward Environmental Pollutants: Role in
Bioremediation. Applied Enviromental Microbiology, 68, 5789-5795.
Parales, R. E., Bruce, N. C., Schmid, A., et Wackett, L. P. (2002). Biodegradation,
biotransformation, and biocatalysis (B3). Applied Enviromental Microbiology, 68, 4699-4709.
Parekh, N. R., McKeown, T., Roberts, S. J., Walker, A., et Suett, D. L., (1992). Bacterial
systems involved in the enhanced degradation of soil-applied pesticides. In Anderson, J.P.E.,
Arnold, D.J., Lewis, F., Torstensson, L. (Eds.), Proceedings of the International Symposium
on Environmental Aspects of Pesticide Microbiology. Department of Microbiology, Swedish
University of Agricultural Sciences, Uppsala, pp. 153-158.
Parkin, T. B. (1987). Soil microsites as a source of denitrification variability. Soil Science Society
of America Journal, 51, 1194–1199.
Pelz, O., Tesar, M., Wittich, R. - M., Moore, E. R. B., Timmis, K. N., et Abraham, R. (1999).
Towards elucidation of microbial community metabolic pathways: unravelling the network of
carbon sharing in a pollutant-degrading bacterial consortium by immunocapture and isotopic
ratio mass spectrometry. Environ Microbiol, 1, 167-174.
Perrière, G., et Gouy, M. (1996) WWW-Query: An on-line retrieval system for biological
sequence banks. Biochimie, 78, 364-369.
Pesce, S., Batisson, I., Bardot, C., Fajon, C., Portelli, C., Montuelle, B., et Bohatier, J. (2009).
Response of spring and summer riverine microbial communities following glyphosate
exposure. Ecotoxicology and Environmental Safety, 72, 1905-1912.
Pesticide Manual. (2006). The Pesticide Manual. British Crop Protection Council. Farnham.
XXième édition, Editeur Tomlin, C.D.S., 1124-1125.

Références Bibliographiques
195
Petersen, S. O., Nielsen, T. H., Frostegard, A. et Olesen, T. (1996). O2 uptake, C metabolism
and denitrification associated with manure hot-spots. Soil Biology and Biochemistry, 28, 341-
349.
Peterson, G. A., Halvorson, A. D., Havlin, J. L., Jones, O. R., Lyon, D. J., et Tanaka, D. L.
(1998). Reduced tillage and increasing cropping intensity in the Great Plains conserves soil C.
Soil and Tillage Research, 47, 207-218.
Pietramellara J., Ascher M. T., Ceccherini G., et Renella G. (2002). Soil as a biological system.
Annals of Microbiology, 52, 119-131.
Prathibha, K. et Sumathi, S. (2007). Biodegradation of mixture containing
monohydroxybenzoate isomers by Acinetobacter calcoaceticus. World Journal of
Microbiology and Biotechnology, 24, 813-823.
Pronicka, E., Rowinska, E., Bentkowski, Z., Zawadzki, J., Holme, E., et Lindstedt, S. (1996).
Treatment of two children with hereditary tyrosinaemia type I and long-standing renal disease
with a 4-hydroxyphenylpyruvate dioxygenase inhibitor (NTBC). J Inherit Metab Dis, 19,
234-238.
Pukall, R., Brambilla, E., et Stackebrandt, E. (1998). Automated fragment length analysis of
fluorescently-labeled 16S rDNA after digestion with 4-base cutting restriction enzymes.
Journal of Microbiological Methods, 32, 55-63.
Puissemier, L., Goux, S., Vanderhayden, V., Debongnie, P., Tresinie, I., et Foucart, G. (1997).
Rapid dissipation of atrazine in soils taken from various maize fields. Weed Research, 37,
171-179.
Racke, K. D. et Coats, J. R. (1988a). Comparative degradation of organophosphorus insecticides
in soil: specificity of enhanced microbial degradation. Journal of Agricultural and Food
Chemistry, 36, 193-199.
Racke, K. D. et Coats, J. R. (1988b). Enhanced degradation and the comparative fate of
carbamate insecticides in soil. Journal of Agricultural and Food Chemistry, 36, 1067-1072.
Radosevich, M., Traina, S. J., Hao, Y. L., et Tuovinen, O. H. (1995). Degradation and
mineralization of atrazine by a soil bacterial isolate. Appl. Environ. Microbiol, 61, 297-302.
Rainey, F. A., N. L. Ward-Rainey, Janssen, P. H., et Hippe, H. (1996). Clostridium paradoxum
DSM 7308(T) contains multiple 16S rRNA genes with heterogeneous intervening sequences.
Microbiology, 142, 2087-2095.
Ramos, J.-L. (2004). Pseudomonas: Genomics, life style and molecular architecture Birkhäuser.

Références Bibliographiques
196
Reddy, K. N. et Bhowmik, P. C. (1991). ICIA-0051 for Postemergence Weed Control in
Conventional Corn (Zea mays). Weed Technology, 5, 509-512.
Reineke, W. et Knackmuss, H. J. (1988). Microbial Degradation of Haloaromatics. Annual
Review of Microbiology, 42, 263-287.
Rentz, J. A., Alvarez, P. J. J., et Schnoor, J. L. (2004). Repression of Pseudomonas putida
phenanthrene-degrading activity by plant root extracts and exudates. Environmental
Microbiology, 6, 574-583.
Rhine, E. D., Fuhrmann, J. J., et Radosevich, M. (2003). Microbial community responses to
atrazine exposure and nutrient availability: Linking degradation capacity to community
structure. Microbial Ecology, 46, 145-160.
Roesch, L. F. W., Fulthorpe, R. R., Riva, A., Casella, G., Hadwin, A. K. M., Kent, A. D.,
Daroub, S. H., Camargo, F. A. O., Farmerie, W. G., et Triplett, E. W. (2007).
Pyrosequencing enumerates and contrasts soil microbial diversity. ISME J, 1, 283-290.
Roeth, F.W. (1986). Enhanced herbicide degradation in soil with repeat application. Review of
Weed Science, 2, 45-65.
Roeth, F. W., Wilson, R. G., Martin, A. R., et Shea, P. J. (1989). Enhanced soil degradation of
butylate and EPTC in nebraska fields. Weed Technology, 3, 24-29.
Rosselló-Mora, R. et Amann, R. (2001). The species concept for prokaryotes. FEMS
Microbiology Reviews, 25, 39-67.
Rouchaud, J., Thirion, A., Callens, D., et Bulcke, R. (1996). Soil Dissipation of the Post-
Emergence Herbicide Sulcotrione in Maize Crops Treated with Organic Fertilizers. Bulletin of
Environmental Contamination and Toxicology, 57, 398-405.
Rouchaud, J., Neus, O., Callens, D., et Bulcke, R. (1997). Enhanced Soil Biodegradation of
Prosulfocarb Herbicide in Barley Crop. Bulletin of Environmental Contamination and
Toxicology, 58, 752-757.
Rouchaud, J., Neus, O., Bulcke, R., Cools, K., et Eelen, H. (1998a). Sulcotrione soil metabolism
in summer corn crops. Bulletin of Environmental Contamination and Toxicology, 61, 669-
676.
Rouchaud, J., Neus, O., Callens, D., et Bulcke, R. (1998b). Sulcotrione soil persistence and
mobility in summer maize and winter wheat crops. Weed Research, 38, 361-371.

Références Bibliographiques
197
Rouchaud, J., Neus, O., Bulcke, R., Cools, K., Eelen, H., et Dekkers, T. (2000). Soil dissipation
of diuron, chlorotoluron, simazine, propyzamide, and diflufenican herbicides after repeated
applications in fruit tree orchards. Archives of Environmental Contamination and. Toxicology,
39, 60-65.
Rousseaux, S., Hartmann, A., et Soulas, Guy (2001). Isolation and characterisation of new
Gram-negative and Gram-positive atrazine degrading bacteria from different French soils.
FEMS Microbiology Ecology, 36, 211-222.
Sait, M., Davis, K. E. R., et Janssen, P. H. (2006). Effect of pH on Isolation and Distribution of
Members of Subdivision 1 of the Phylum Acidobacteria Occurring in Soil. Applied
Environmental Microbiology., 72, 1852-1857.
Sait, M., Hugenholtz, Philip, et Janssen, P. H. (2002). Cultivation of globally distributed soil
bacteria from phylogenetic lineages previously only detected in cultivation-independent
surveys. Environmental Microbiology, 4, 654-666.
Saitou, N. et Nei, M. (1987). The neighbor-joining method: a new method for reconstructing
phylogenetic trees. Molecular Biology and Evolution, 4, 406-425.
Sangwan, P., Kovac, S., Davis, K. E. R., Sait, M., et Janssen, P. H. (2005). Detection and
Cultivation of Soil Verrucomicrobia. Appl. Environ. Microbiol., 71, 8402-8410.
Scheunert, I., Mansour, M., Dörfler, U., et Schroll, R. (1993). Fate of pendimethalin, carbofuran
and diazinon under abiotic and biotic conditions. The Science of The Total Environment, 132,
361-369.
Schloss, P. D., Larget B. R., et Handelsman, J. (2004) Integration of microbial ecology and
statistics: a test to compare gene libraries. Applied Environmental Microbiology, 70, 5485-
5492.
Schmidt, S.K., Scow, K.M., et Alexander, M. (1987). Kinetics of p-nitrophenol mineralization
by a Pseudomonas sp.: effects of second substrates. Applied. Environmental Microbiology.,
53, 2617-2623.
Schneider, M. et Bruijn, F. J. (1996). Rep-PCR mediated genomic fingerprinting of rhizobia and
computer-assisted phylogenetic pattern analysis. World Journal of Microbiology and
Biotechnology, 12, 163-174.
Schulz, A., Ort, O., Beyer, P., et Kleinig, H. (1993). SC-0051, a 2-benzoyl-cyclohexane-1,3-
dione bleaching herbicide, is a potent inhibitor of the enzyme p-hydroxyphenylpyruvate
dioxygenase. FEBS Letters, 318, 162-166.

Références Bibliographiques
198
Sei, K., Asano, K.-I., Tateishi, N., Mori, K., Ike, M., et Fujita, M. (1999). Design of PCR
primers and gene probes for the general detection of bacterial populations capable of
degrading aromatic compounds via catechol cleavage pathways. Journal of Bioscience and
Bioengineering, 88, 542-550.
Secor, J. (1994). Inhibition of barnyadgrass 4-hydroxyphenylpyruvate dioxygenase by sulcotrione.
Plant Physiology, 106, 1429-1433.
Sexstone, A.J., Revsbech, N.P., Parkin, T.B., et Tiedje, J.M. (1985). Direct measurement of
oxygen profiles and denitrification rates in soil aggregates. Soil Science Society of America
Journal, 49, 645–651.
Shingler, V., Franklin, F. C. H., Tsuda, M., Holroyd, D., et Bagdasarian, M. (1989). Molecular
Analysis of a Plasmid-encoded Phenol Hydroxylase from Pseudomonas CF600. Journal of
General Microbiology, 135, 1083-1092.
Sikorski, J., Teschner, N., et Wackernagel, W. (2002). Highly different levels of natural
transformation are associated with genomic subgroups within a local population of
Pseudomonas stutzeri from soil. Applied and Environmental Microbiology, 68, 865-873.
Sims, G.K. (2006). Nitrogen starvation promotes biodegradation of N-heterocyclic compounds in
soil. Soil Biology and Biochemistry, 38, 2478-2480.
Sims, G. K., Sommers, L. E., et Konopka, Allan (1986). Degradation of pyridine by
Micrococcus luteus isolated from soil. Applied Environmental Microbiology, 51, 963-968.
Singh, B. K., Walker, A., et Wright, D. J. (2005). Cross-enhancement of accelerated
biodegradation of organophosphorus compounds in soils: Dependence on structural similarity
of compounds. Soil Biology and Biochemistry 37, 1675-1682.
Slater, J. H. (1984). Genetic interactions in microbial communities. Current perspectives in
microbial ecology ASM. Klug, M.J., Reddy, C.A.
Smelt, J. H., Van de Peppel-Groen, A. E., Van der Plas, L. J. T., et Dijksterhuis, A. (1996).
Development and duration of accelerateddegradation of nematicides in different soils. Soil
Biology and Biochemistry, 28, 1757-1765 .
Smith, K. L. et Kennedy, A. C. (1995). Soil microbial diversity and the sustainability of
agricultural soils. Plant and Soil, 170, 75-86.
Smith, C. A., O’Reilly, K. T., et Hyman, M. R. (2003a). Characterization of the initial reactions
during the cometabolic degradation of methyl tert-butyl ether (MTBE) by propane-grown
Mycobacterium vaccae JOB5. Applied Environmental Microbiology, 69, 796-804.

Références Bibliographiques
199
Smith C. A., O’Reilly K. T., et Hyman, M. R. (2003b) Cometabolism of methyl tertiary butyl
ether and gaseous n-alkanes by Pseudomonas mendocina KR-1 grown on C5 to C8 n-
alkanes. Applied Environmental Microbiology, 69, 7385-7394.
Sneath, P. H. A., et Sokal, R. R. (1973). Numerical taxonomy. W. H. Freeman and Company, San
Francisco.
Somasundaram L., et J. R. Coats, (1990). Pesticide transformation products in the environment.
ACS symposium, series 459, 2-9.
Sørensen, S. R., Ronen, Z., et Aamand, J. (2001). Isolation from agricultural soil and
characterization of a Sphingomonas sp. able to mineralize the phenylurea herbicide
isoproturon. Applied Environmental Microbiology, 67, 5403-5409.
Sørensen, S. R., et Aamand, J. (2003). Rapid mineralisation of the herbicide isoproturon in soil
from a previously treated Danish agricultural field. Pest Management Science, 59, 1118-1124.
Sørensen, S. R., Bending, G. D., Jacobsen, C. S., Walker, A., et Aamand, J. (2003). Microbial
degradation of isoproturon and related phenylurea herbicides in and below agricultural fields.
FEMS Microbiology Ecology, 45, 1-11.
Sørheim, R., Torsvik, V. L., et Goksøyr, J. (1989). Phenotypical Divergences between
Populations of Soil Bacteria Isolated on Different Media. Microbial Ecology, 17, 181-192.
Soulas, G. (1985). La dégradation des pesticides dans le sol. Aspects microbiens et cinétiques.
Science du sol, 43-57.
Soulas, G. (1990). Dégradation biologique d’un herbicide, l’acide 2,4-dichlorophénoxyacétique
(2,4-D) dans le sol. Aspects cinétiques. Thèse de doctorat. Institut National Polytechnique de
Lorraine. ENSAIA.
de Souza, M., Sadowsky, M., et Wackett, L. (1996). Atrazine chlorohydrolase from
Pseudomonas sp. strain ADP: gene sequence, enzyme purification, and protein
characterization. Journal of Bacteriology, 178, 4894-4900.
de Souza, M. L., Seffernick, J., Martinez, B., Sadowsky, M. J., et Wackett, Lawrence P.
(1998). The atrazine catabolism genes atzABC are widespread and highly conserved.
Joournal of Bacteriology, 180, 1951-1954.
Stackebrandt, E., et Goebel, B. M. (1994). Taxonomic note: a place for DNA-DNA reassociation
and 16S rRNA sequence analysis in the present species définition. International Journal of
Systematics and Evolutionary Bacteriology, 44, 846–849.

Références Bibliographiques
200
Stackebrandt, E. (2003). The richness of prokaryotic diversity : there must be a species
somewhere. Food Technology Biotechnology, 41, 17-22.
Staley, J. T. et Konopka, A. (1985). Measurement of in situ activities of nonphotosynthetic
microorganisms in aquatic and terrestrial habitats. Annual Review of Microbiology, 39, 321-
346.
Stern, M. J., Ames, G. F.-L., Smith, N. H., Clare Robinson, E., et Higgins, C. F. (1984).
Repetitive extragenic palindromic sequences: A major component of the bacterial genome.
Cell, 37, 1015-1026.
Stern, S., Weiser, B., et Noller, H. F. (1988b). Model for the three-dimensional folding of 16S
ribosomal RNA. Journal of Molecular Biology, 204, 447-481.
Stevenson, B. S., Eichorst, S. A., Wertz, J. T., Schmidt, T. M., et Breznak, J. A. (2004). New
strategies for cultivation and detection of previously uncultured microbes. Applied
Environmental Microbiology, 70, 4748-4755.
Stockinger, J., Hinteregger, C., Loidl, M., Ferschl, A., et Streichsbier, F. (1992).
Mineralization of 3-chloro-4-methylaniline via an ortho-cleavage pathway by Pseudomonas
cepacia strain CMA1. Applied Microbiology and Biotechnology, 38.
Stotzky, G., et Norman, A.G. (1961). Factors limiting microbial activities in soil. I. The level of
substrate, nitrogen and phosphorus. Archives of Microbiology, 40, 341–369.
Strachan, P. D., Freer, A. A., et Fewson, C. A. (1998). Purification and characterization of
catechol 1,2-dioxygenase from Rhodococcus rhodochrous NCIMB 13259 and cloning and
sequencing of its catA gene. Biochemistry Journal, 333 (Pt 3), 741-747.
Streber, W. R., Timmis, K. N., et Zenk, M. H. (1987). Analysis, cloning, and high-level
expression of 2,4-dichlorophenoxyacetate monooxygenase gene tfdA of Alcaligenes
eutrophus JMP134. Journal of Bacteriology, 169, 2950-2955.
Suett, D. L. (1987). Influence of treatment of soil with carbofuran on the subsequent performance
of insecticides against cabbage root fly (Delia radicum) and carrot fly (Psila rosae). Crop
Protection, 6, 371-378.
Suett, D. L. et Jukes, A. A. (1988). Accelerated degradation of aldicarb and its oxidation products
in previously treated soils. Crop Protection, 7, 147-152.

Références Bibliographiques
201
Suett, D. L., Jukes, A. A., et Phelps, K., (1993). Stability of accelerated degradation of soil
applied insecticides: laboratory behaviour of aldicarb and carbofuran in relation to their
efficacy against cabbage root fly (Delia radicum) in previously-treated field soils. Crop
Protection, 12, 431-442.
Szczecinski, P., Lamparska, D., Gryff-Keller, A., et Gradowska, W. (2008). Identification of 2-
[2-nitro-4-(trifluoromethyl)-benzoyl]-cyclohexane-1,3-dione métabolites in urine of patients
suffering from tyrosinemia type I with the use of 1H and 19F NMR spectroscopy. Acta
Biochimica Polonica, 55, 749-752.
Takenaka, S., Murakami, S., Shinke, R., et Aoki, K. (1998). Metabolism of 2-aminophenol by
Pseudomonas sp. AP-3: modified meta -cleavage pathway. Archives of Microbiology, 170,
132-137.
Takeo, M., Maeda, Y., Okada, H., Miyama, K., Mori, K., Ike, M., et Fujita, M. (1995).
Molecular cloning and sequencing of the phenol hydroxylase gene from Pseudomonas putida
BH. Journal of Fermentation and Bioengineering, 79, 485-488.
Takeo, Masahiro, Maeda, Yoshimichi, Okada, Hiroshi, Miyama, Kanako, Mori, Kazuhiro,
Ike, Michihiko, et Fujita, Masanori (1995). Molecular cloning and sequencing of the phenol
hydroxylase gene from Pseudomonas putida BH. Journal of Fermentation and
Bioengineering, 79, 485-488.
Thomas, C. M. et Nielsen, K. M. (2005). Mechanisms of, and Barriers to, Horizontal Gene
Transfer between Bacteria. Nature Reviews Microbiology, 3, 711-721.
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., et Higgins, D. G. (1997). The
CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by
quality analysis tools. Nucleic Acids Research, 25, 4876-4882.
Top, E. M., Springael, D., et Boon, N. (2002). Catabolic mobile genetic elements and their
potential use in bioaugmentation of polluted soils and waters. FEMS Microbiology Ecology,
42, 199-208.
Top, E. M. et Springael, D. (2003). The role of mobile genetic elements in bacterial adaptation to
xenobiotic organic compounds. Current Opinion in Biotechnology, 14, 262-269.
Topp, E., Zhu, H., Nour, S. M., Houot, S., Lewis, M., et Cuppels, D. (2000). Characterization of
an Atrazine-Degrading Pseudaminobacter sp. Isolated from Canadian and French
Agricultural Soils. Applied Environmental Microbiology, 66, 2773-2782.

Références Bibliographiques
202
Topp, E. (2003). Bacteria in agricultural soils: Diversity, role and future perspectives. Canadian
Journal of Soil Science, 83, 303-309.
Torsvik, V., Goksøyr, J, et Daae, F. L. (1990). High diversity in DNA of soil bacteria. Applied
Environmental Microbiology, 56, 782-787.
Torsvik, V., Sørheim, R, et Goksøyr, J (1996). Total bacterial diversity in soil and sediment
communities-A review. Journal of Industrial Microbiology and Biotechnology, 17, 170-178.
Torsvik, V., Øvreås, L., et Thingstad, T. F. (2002). Prokaryotic diversity-magnitude, dynamics,
and controlling factors. Science, 296, 1064 -1066.
Townsend, K. M., Dawkins, H. J. S., et Papadimitriou, J. M. (1997). REP-PCR analysis of
Pasteurella multocida isolates that cause haemorrhagic septicaemia. Research in Veterinary
Science, 63, 151-155.
Townson, J., Kemp, J., Cory, P., Bartlett, D., Sutton, P., Collins, D., et Wichert, R. (1999).
Mode of action of HPPD-inhibiting herbicides. The XIVth International Plant Protection
Congress (IPPC). Jerusalem, Israel, Phytoparasitica, 28,2.
Toyama, T., Maeda, N., Murashita, M., Chang, Y.-C., et Kikuchi, S. (2010). Isolation and
characterization of a novel 2-sec-butylphenol-degrading bacterium Pseudomonas sp. strain
MS-1. Biodegradation, 21, 157-165.
Trevors, J. T. (1996). DNA in soil: adsorption, genetic transformation, molecular evolution and
genetic microchip. Antonie van Leeuwenhoek, 70, 1-10.
Tsai, Y. L. et Olson, B. H. (1992). Rapid method for separation of bacterial DNA from humic
substances in sediments for polymerase chain reaction. Applied Environmental Microbiology,
58, 2292-2295.
Tsegaye, Y., Shintani, D. K., et DellaPenna, D. (2002). Overexpression of the enzyme p-
hydroxyphenolpyruvate dioxygenase in Arabidopsis and its relation to tocopherol
biosynthesis. Plant Physiology and Biochemistry, 40, 913-920.
Turnbull, G., Cullington, J., Walker, A., et Morgan, J. (2001). Identification and
characterisation of a diuron-degrading bacterium. Biology and Fertility of Soils, 33, 472-476.
Vaillancourt, F. H., Bolin, J. T., et Eltis, L. D. (2006). The ins and outs of ring-cleaving
dioxygenases. Critical Reviews in Biochemistry and Molecular Biology, 41, 241-267.
Van der Meer, J. R., de Vos, W. M., Harayama, S., et Zehnder, A. J. (1992). Molecular
mechanisms of genetic adaptation to xenobiotic compounds. Microbiology and Molecular
Biology Reviews, 56, 677-694.

Références Bibliographiques
203
Van Elsas, J. D., Jansson, J. K., et Trevors, J. T. (2007). Modern soil microbiology. Second
Edition. CRC Press. Taylor and Francis Group. Boca, Raton, London, New-York.
Vandamme, P., Pot, B., Gillis, M., de Vos, P., Kersters, K., et Swings, J. (1996). Polyphasic
taxonomy, a consensus approach to bacterial systematics. Microbiology and Molecular
Biology Reviews, 60, 407-438.
Vaneechoutte, M., Rossau, R., De Vos, P., Gillis, M., Janssens, D., Paepe, N., De Rouck, A.,
Fiers, T., Claeys, G., et Kersters, K. (1992). Rapid identification of bacteria of the
Comamonadaceae with amplified ribosomal DNA-restriction analysis (ARDRA). FEMS
Microbiology Letters, 72, 227-233.
Vaneechoutte, M., De Beenhouwer, H., Claeys, G., Verschraegen, G., De Rouck, A., Paepe,
N., Elaichouni, A., et Portaels, F. (1993). Identification of Mycobacterium species by using
amplified ribosomal DNA restriction analysis. Journal of Clinical Microbiology, 31, 2061-
2065.
Vartoukian, S. R., Palmer, R. M., et Wade, W. G. (2010). Strategies for culture of
‘unculturable’ bacteria. FEMS Microbiology Letters, 309, 1-7.
Versalovic, J., Koeuth, T., et Lupski, J. R. (1991). Distribution of repetitive DNA sequences in
eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Research, 19,
6823-6831.
Voos, G. et Groffman, P. M. (1997). Relationships between microbial biomass and dissipation of
2,4-D and dicamba in soil. Biology and Fertility of Soils, 24, 106-110.
Walker, A., Jurado-Exposito, M., Bending, G. D., et Smith, V. J. R. (2001). Spatial variability
in the degradation rate of isoproturon in soil. Environmental Pollution, 111, 407-415.
Walker, A., et Welch, S. J. (1989). The relative movement and persistence in soil of
chlorsulfuron, metsulfuron-methyl and triasulfuron. Weed Research, 29, 375-383.
Walker, A., et Welch, S. J. (1991). Enhanced degradation of some soil-applied herbicides. Weed
Research, 31, 49-57.
Walker, A., Brown, P. A., et Entwistle, A. R. (1986). Enhanced degradation of iprodione and
vinclozolin in soil. Pesticide Science, 17, 183-193.
Wardle, D. A. et Parkinson, D. (1991). Relative importance of the effect of 2,4-D, glyphosate,
and environmental variables on the soil microbial biomass. Plant Soil, 134, 209-219.

Références Bibliographiques
204
Warton, B., Matthiessen, J. N., et Shackleton, M. A. (2003). Cross-enhancement: enhanced
biodegradation of isothiocyanates in soils previously treated with metham sodium. Soil
Biology and Biochemistry, 35, 1123-1127.
Wayne, L. G., Brenner, D. J., Colwell, R. R., Grimont, P. A. D., Kandler, O., Krichevsky, L.,
Moore, L. H., Moore, W. C., Murray, R. G. E., Stackebrandt, E., Starr, M. P., et
Trüper, H. G. (1987). Report of the ad hoc committee on reconciliation of approaches to
bacterial systematics. International Journal of Systematic and Evolutionary Bacteriology 37,
463-464.
Wikström, P., Wiklund, A., Andersson, A.-C., et Forsman, M. (1996). DNA recovery and PCR
quantification of catechol 2,3-dioxygenase genes from different soil types. Journal of
Biotechnology, 52, 107-120.
Wilkins, B. M. (1995). Gene transfer by bacterial conjugation: diversity of systems and functional
specializations. In Population Genetics of Bacteria Cambridge University Press. Baumberg,
S., Young, J. P. W., Saunders, S. R. & Wellington, E. M. H., Cambridge, UK.
Williams, P. A., Jones, R. M., et Zylstra, G.J. (2004). Genomics of catabolic plasmids. Kluwer
Academic/Plenum Publishers. Ramos, J.L., New-York.
Williams, P. A. et Sayers, J. R. (1994). The evolution of pathways for aromatic hydrocarbon
oxidation in Pseudomonas. Biodegradation, 5, 195-217.
Wilson, J. S., et Foy, C. L. (1992). Weed control in no-tillage and conventional corn (Zea mays)
with ICIA-0051 and SC-0074. Weed technology, 4, 731-738.
Wiszniowski, J., Halle, A. T., Richard, C., Hitmi, A., et Ledoigt, G. (2009). Photodegradation
product of sulcotrione and the physiological response of maize (Zea mays) and white mustard
(Sinapis alba). Chemosphere, 74, 1224-1230.
Woese, C. R. (1987). Bacterial evolution. Microbiol. Rev, 51, 221-271.
Woese, C. R., Winker, S., et Gutell, R. R. (1990). Architecture of ribosomal RNA: constraints on
the sequence of « tetra-loops ». Proceedings of the National Academy of Sciences U.S.A, 87,
8467-8471.
Wu, C.-S., Huang, J.-L., Sun, Y.-S., et Yang, D.-Y. (2002). Mode of action of 4-
hydroxyphenylpyruvate dioxygenase inhibition by triketone-type inhibitors. Journal of
Medicinal Chemistry, 45, 2222-2228.

Références Bibliographiques
205
Wu, N., Jin, Y., Jin, F., Tan, Y., Tao, H., Zheng, M., Chen, R., Liu, K., et Gao, M. (2011).
Effects of sulcotrione [2-(2-chloro-4-mesylbenzoyl)-cyclohexane-1,3-dione] on enzymes
involved in tyrosine catabolism and the extent of the resulting tyrosinemia and its relationship
with corneal lesions in rats. Pesticide Biochemistry and Physiology, 99, 162-169.
Yano, H., Garruto, C. E., Sota, M., Ohtsubo, Y., Nagata, Y., Zylstra, Gerben J, Williams,
Peter A, et Tsuda, M. (2007). Complete sequence determination combined with analysis of
transposition/site-specific recombination events to explain genetic organization of IncP-7
TOL plasmid pWW53 and related mobile genetic elements. Journal of Molecular Biology,
369, 11-26.
Yano, H., Miyakoshi, M., Ohshima, K., Tabata, M., Nagata, Y., Hattori, M., et Tsuda, M.
(2010). Complete nucleotide sequence of TOL plasmid pDK1 provides evidence for
evolutionary history of IncP-7 catabolic plasmids. Journal of Bacteriology, 192, 4337-4347.
Yassir, A., Rieu, C., et Soulas, G. (1998). Microbial N-dealkylation of atrazine : effect of
exogenous organic substrates and behaviour of the soil microflora. Pesticide Science, 54, 75-
82.
Yassir, A., Lagacherie, B., Houot, S., et Soulas, G. (1999). Microbial aspects of atrazine
biodegradation in relation to history of soil treatment. Pesticide Science, 55, 799-809.
Yee, T. W. et Smith, D. W. (1990). Pseudomonas chromosomal replication origins: a bacterial
class distinct from Escherichia coli-type origins. Proc Natl Acad Sci U S A, 87, 1278-1282.
Zaneveld, J. R., Nemergut, D. R., et Knight, R. (2008). Are all horizontal gene transfers created
equal? Prospects for mechanism-based studies of HGT patterns. Microbiology (Reading,
Engl.), 154, 1-15.
Zengler, K., Toledo, G., Rappé, M., Elkins, J., Mathur, E. J., Short, J. M., et Keller, M.
(2002). Cultivating the uncultured. Proceedings of the National Academy of Sciences, 99,
15681 -15686.
Zeyaullah, M., Abdelkafe, A. S., Zabya, W. B., et Ali, A. (2009). Biodegradation of catechols by
microorganisms. A short review. African journal of Biotechnology, 8, 2916-2922.
Zhao, C., Zhang, Y., Li, X., Wen, D., et Tang, X. (2011). Biodegradation of carbazole by the
seven Pseudomonas sp. strains and their denitrification potential. Journal of Hazardous
Materials, 190, 253-259.

Références Bibliographiques
206
Zhou, N.-Y., Fuenmayor, S. L., et Williams, Peter A. (2001). Nag genes of Ralstonia (formerly
Pseudomonas) sp. strain U2 encoding enzymes for gentisate catabolism. Journal of
Bacteriology, 183, 700-708.
Zhou, Y.-Y., Chen, D.-Z., Zhu, R.-Y., et Chen, J.-M. (2011). Substrate interactions during the
biodegradation of BTEX and THF mixtures by Pseudomonas oleovorans DT4. Bioresource
Technology, 102, 6644-6649.
Zuckerkandl, E. et Pauling, L. (1965). Molecules as documents of evolutionary history. Journal
of Theoretical Biology, 8, 357-366.

207
Valorisation des travaux de recherche

Valorisation des travaux de recherche
208
Publicationn°1(SOUSPRESSE)
Article soumis le 30 mars 2011 à la revue Pest Management Science, corrigé le 24 mai 2011, accepté le 24 juin 2011.
Isolation and Characterization of a Bacterial Strain Degrading the Herbicide Sulcotrione from an Agricultural Soil.
Christophe CALVAYRACa,b, Fabrice MARTIN-LAURENTc, Alexia FAVEAUXa, Nathalie
PICAULTb, Olivier PANAUDb, Camille-Michel COSTE a, Hanene CHAABANEd and Jean-
François COOPERa∗
a Laboratoire de Chimie des Biomolécules et de l’Environnement (LCBE, EA 4215),
Université de Perpignan Via Domitia (UPVD), 52 avenue Paul Alduy 66860 Perpignan,
France.
b Laboratoire Génome et Développement des Plantes (UMR, 5096 CNRS-UPVD), Université
de Perpignan Via Domitia (UPVD), 52 avenue Paul Alduy 66860 Perpignan, France.
c INRA, Université de Bourgogne, UMR1229 Microbiologie du Sol et de l'Environnement,
CMSE, 17 rue Sully, BP 86510, F-21065 Dijon, France.
d Unité de recherche Protection Intégrée des Cultures, Institut National Agronomique de
Tunisie
43, av Charles Nicolle, 1082 Tunis Mahrajène, Tunisie.
∗ Corresponding author. Tel: +33 (4) 68 66 20 81 Fax: +33 (4) 68 66 81 38; E-mail : [email protected]

Valorisation des travaux de recherche
209
Keywords: Sulcotrione, Biodegradation, Soil, Pseudomonas putida, Metabolites.
Running title : Biodegradation of sulcotrione in soil. Intervention of Pseudomonas sp.
1- Abstract
BACKGROUND: Dissipation kinetics of sulcotrione herbicide, previously four times sprayed
on a French soil, was studied using a laboratory microcosm approach. Then, advanced
cultivation-based approach was performed to isolate bacteria responsible for
biotransformation of sulcotrione. Chromatographic methods were used as complementary
tools to define its metabolic pathway.
RESULTS: Soil microflora was able to quickly biotransform the herbicide (DT50 ca 8 days).
2-chloro-4-mesylbenzoic acid, one of its main metabolites, was clearly detected. However, no
accelerated biodegradation process was observed. Eight pure sulcotrione-resistant strains were
isolated but only one (1OP) was capable to degrade this herbicide with a relative high
efficiency and to use it as sole carbon source and energy. In parallel, another sulcotrione-
resistant strain (1TRANS) was shown to be unable to degrade the herbicide. Amplified
ribosomal restriction analysis (ARDRA) and repetitive extragenic palendromic PCR genomic
(REP-PCR) fingerprintings of strains 1OP and 1TRANS gave indistinguishable profiles.
CONCLUSION: Sequencing and aligning analysis of 16S rDNA genes of each pure strain
revealed identical sequences and a close phylogenetic relationship (99 % sequence identity) to
Pseudomonas putida. Such physiological and genetic properties of 1OP to metabolize
sulcotrione were probably governed by mobile genetic elements in the genome of the bacteria.
1 INTRODUCTION
Sulcotrione [2-(2-chloro-4-methylsulfonylbenzoyl)-1,3-cyclohexanedione] is a triketone
herbicide applied at 300-450 g ha-1 on corn crop to control the development of a wide range
of annual and perennial broadleaf weeds.
A large proportion of this herbicide reaches the soil where both abiotic (i.e sorption, chemical
degradation) and biotic (biodegradation) processes control the fate and the activity of this

Valorisation des travaux de recherche
210
herbicide. 2-chloro-4-mesylbenzoic acid (CMBA) and 1,3-cyclohexanedione (CHD) have
been described as two biodegradation and hydrolysis transformation products.1-8 A derivative
of phenylheptanoic acid was also described under special conditions.6-7,9 Up to now, no report
showed the existence of accelerated biodegradation of sulcotrione as a result of repeated
applications. Soil microorganisms able to transform sulcotrione are not yet described,
contrarily to mesotrione, another herbicide belonging to the triketone family.10-14 In this
context, the aim of our work was to evaluate the response of an agricultural soil, regularly
treated with sulcotrione, possibly adapted to its biodegradation, in order to isolate novel and
efficient sulcotrione degraders. Sulcotrione transformation was monitored in the soil using
chromatographic methods. Sulcotrione-degrading bacterial strain was isolated by conducting
an adaptation culture to the sulcotrione. Isolate was characterized by ARDRA, REP-PCR and
by cloning/sequencing of partial 16S rDNA sequence. Its capacity to degrade aerobically
sulcotrione was estimated.
2 MATERIALS AND METHODS
2.1. Experimental design
2.1.1. Experimental field site
The experimental field site was set up at the University of Perpignan, France (42° 40' 55" N,
2° 53' 49" E). The field was divided in two plots of 184 m² each, separated by a buffer strip.
One plot (4S) was treated with 300 g ha-1 of sulcotrione (Mikado®) four times with one month
interval. After this, plot 4S was neither pesticide treated nor cultivated for 24 months. The
second plot (C), used as control, was never treated with sulcotrione. According to FAO
classification, the soil is a clayey sandy loam soil. Soil physicochemical characteristics were:
clay 13.9 %, silt 60.5%, sand 25.6 %, soil moisture 25 %, organic carbon 0.9 %, organic
nitrogen 0.98 g kg-1, C/N ratio 9.64, cation exchange capacity (CEC) 15.5 meq 100 g-1,
Ca2+/CEC 214 % and pH in water 8.1. Soil samples were collected from the surface layer (0-7
cm) of the experimental field. Composite samples were sieved to 2 mm and stored in the dark
at 4°C until used.

Valorisation des travaux de recherche
211
2.1.2 Sulcotrione biodegradation
The ability of the microflora of the soil samples collected on the experimental field (4S and
C) to degrade sulcotrione was tested under laboratory conditions. Each microcosm, prepared
with 10 g of moist soil placed in Petri dishes (5.5 cm i.d.), were treated with 550 µL sterile
aqueous sulcotrione solution at 22 mg L-1 (i.e. 1.2 µ g of sulcotrione per g of soil) and
homogeneized. Treatments were applied four times at day 0, 30, 60 and 90 (T1D0, T2D30,
T3D60 and T4D90), necessiting the preparation of forty microcosms for each soil type. All
analysis were made in duplicate. In addition, autoclaved controls were set up to estimate the
occurrence of abiotic biodegradation (St4S and StC). All samples were kept in the dark to
avoid photodegradation of the sulcotrione. Thorough the incubation period, soil moisture
content was kept at 25 % (moist weight), making it comparable to field moisture. After each
treatment, each entire soil aliquot (10 g) was regularly sampled (i.e. after 0, 2, 7, 15 and 30
days) to monitor sulcotrione dissipation.
2.2. Chemical Analysis
2.2.1. Reagents
Standard of sulcotrione (98.8 % purity) – a weak acidic herbicide, pKa value 2.87, MW
328.77 g mol-1 and water solubility 165 mg L-1 at 25°C - was purchased from Sigma-Aldrich,
France. Standards of 1,3-cyclohexanedione (CHD) (97.0% purity) and 2-chloro-4-
mesylbenzoic acid (CMBA) (95.0% purity) were purchased from Fluka and Acros Organics,
respectively.
Methanol and acetonitrile (HPLC quality) were purchased from Carlo Erba, dichloromethane
for pesticide analysis from Riedel-deHaën GmbH, trifluoroacetic acid (TFA) (99.0% purity)
from Aldrich, hydrochloric acid (38.0%) from Prolabo and water was Milli-Q quality.
(Trimethylsilyl) diazomethane 2.0 M in diethyl ether was supplied by Sigma-Aldrich.
Supelclean TM ENVI-Chrom P SPE cartridges were supplied by Supelco, France.
2.2.2. Analytical procedure
2.2.2.1. Degradation of sulcotrione in soil samples
Extraction step
Soil samples (10 g) were extracted twice with 60 and 30 mL of a mixture acetonitrile /0.1 M
HCl (90/10; v/v) over 30 and 15 min periods, then filtered on a Whatman filter GF/A. The

Valorisation des travaux de recherche
212
organic filtrate was evaporated at 30 °C and the acidic aqueous solution was then extracted
twice with 5.0 mL of dichloromethane. The organic phase was evaporated to dryness, and the
extract was solubilized in 0.10 M HCl. It was then purified on an EnviChrom P cartridge,
eluted with methanol (5 mL) and concentrated to 1 mL. The extract was finally analysed by
HPLC. One blank (sulcotrione-free soil) was systematically included and analysed under the
same conditions.
Chromatographic analysis
Soil extracts were analysed using Shimadzu HPLC apparatus equipped with a Supelco ODS
Hypersil C18 column (5 µm, 250 mm x 4.6) and a SPD-10 Avp UV/Vis detector set at 265 nm.
The mobile phase consisted of a mixture of water acidified by 0.3 % trifluoroacetic acid (AW)
and acetonitrile (ACN), delivered at a flow rate of 1 mL min-1 and using a gradient system
[70/30 (AW/ACN) at t = 0 min to 50/50 (AW/ACN) at 8 min, then maintained for 10 min].
Analytical performance
This analytical method was an adaptation of the procedure previously published by Chaabane
et al.9 Quantification limit was estimated to 0.05 µg g-1 for sulcotrione herbicide, with a mean
recovery rate of 87 % ± 10.
2.2.2.2. Degradation of sulcotrione by the bacterial culture
Extraction step
One mL aliquots of the bacterial culture (selected among sulcotrione-resistant ones) were
centrifuged (15 min, 1100 g) at room temperature. The supernatant was recovered, acidified
with 0.3 mL of 0.16 M HCl and brought to 2.5 mL with distilled water. Then, the acidic
aqueous solution was extracted twice with 2.5 mL dichloromethane. The organic phase was
evaporated to dryness, solubilized in 1 mL methanol before analysis. No adsorption of
sulcotrione was observed on centrifuge tubes.
High performance liquid chromatography (HPLC/UV) of sulcotrione and its main métabolites
Culture media extracts were analyzed using Jasco HPLC apparatus equipped with a Supelco
ODS Hypersil C18 column (5 µm, 250 mm x 4.6) and an UV/Visible Detector Jasco 875-UV
set at 265 nm. The mobile phase consisted of a mixture of water acidified by 0.3 %

Valorisation des travaux de recherche
213
trifluoroacetic acid (AW) and acetonitrile (ACN), delivered at a flow rate of 1 mL.min-1. To
improve the separation of the active ingredient and its metabolites during degradation
kinetics, a gradient system [70/30 (AW/ACN) from 0 to 4 min, evolving to 40/60 (AW/ACN)
at 7 min, then maintained for 15 min] was used. Detection of sulcotrione and its main
metabolites (1,3-cyclohexanedione and 2-chloro-4-mesylbenzoic acid) was done at 265 nm.
Retention times were estimated as follows: sulcotrione (11.1 min), CMBA (5.1 min) and
CHD (4.5 min).
High performance liquid chromatography/Mass spectrometry (HPLC/ESI/MS) analysis of
metabolite 1,3- cyclohexanedione (CHD)
LC/MS analysis was carried out on a LCQ-Fleet (Thermo Fisher Scientific) equipped with an
Electrospray Ionisation (ESI) source and a 3D ion-trap analyser. LC analysis was achieved
using an Accela Thermo Fisher chromatograph equipped with an Accela 600 pump, an Accela
Autosampler and a diode array detector (Accela PDA detector). Full scan spectra from m/z 60
to 300 in both positive (source voltage 4 kV) and negative (source voltage 3.5 kV) ion modes
were recorded. Separation was carried out at room temperature using a Phenomenex Gemini
C6-Phenyl column (150 x 3.0 mm; 5 µm). The mobile phase consisted of a mixture of 0.1 %
formic acid water (A-W) and 0.1 % formic acid acetonitrile (A-ACN) delivered at a flow rate
of 0.4 mL min-1, with a gradient system: 90/10 (A-W/A-ACN) from 0 to 2 min, evoluting to
10/90 (A-W/A-ACN) at 20 min. Under these conditions, retention time of 1,3 CHD was 4.51
min. Quantification limit was estimated in positive mode to 0.5 mg L-1.
Gas chromatography/mass spectrometry (GC/MS) analysis of metabolite 2-chloro-4-
mesylbenzoic acid (CMBA) after derivatization
In presence of methanol, trimethylsilyldiazomethane (TMS) gives methyl esters in good
yields at room temperature with various carboxylic acid including aromatic, heteroaromatic,
alicyclic and aliphatic ones. The esterification proceeds instantaneously and quantitatively
when an excess of TMS is used and the reaction can be easily monitored by the disappearance
of the yellow colour.15
Twenty five µL of TMS prepared in diethyl ether solution was added to 0.5 mL of culture
media extract mixed with 0.5 mL of methanol. After 2 hours reaction time, the extract was
analysed by GC/MS.

Valorisation des travaux de recherche
214
GC/MS analysis was carried out on a Focus GC connected to a mass spectrometric detector
DSQII (Thermo Fisher Scientific) equipped with a column G-SQC GC, 15 m x 0.25 mm x
0.25 µm (5% Phenyl Methylpolysiloxane), initial oven temperature 80 °C, for 1 min raised at
10°C min-1 to 260 °C for 15 min; carrier gas helium 6.0; electron impact mode 70 eV; full
scan mode; split injection mode; transfer line temperature 300 °C. Under such conditions,
retention time of methylated CMBA was 14.81 min and characteristic m/z ions were 155, 217
and 248 (Fig. 1). A calibration curve was established on m/z 217 for the range of
concentration 0.4 to 20 mg L-1 (y = 364 x – 62, r² = 0,996).
Analytical performances
Culture media samples were spiked with known amounts of analytical standards of
sulcotrione, CHD and CMBA (2 to 30 mg L-1). Calculations were made by HPLC/UV and/or
GC/MS, after analysis in duplicate, using calibration curves. The mean recoveries were
estimated at 75 ± 6 %, 55 ± 8 %, 75 ± 10 % for sulcotrione, CHD and CMBA, respectively.
The limits of quantification, defined as the sample concentration required giving a signal-to-
noise ratio of 5:1, were estimated to 0.5 to 1.0 mg L-1.
2.3. Isolation of sulcotrione degrader
2.3.1. Culture media and cells preparation
A culture medium (NS), where the nutrient source was the control soil C, was prepared and
used to promote the growth of bacteria previously described as hardly cultivable on the
conventional culture media.16 Briefly, soil was sieved through a 2 mm mesh, dried at 65 °C
for 3 days. The culture media was prepared by adding 40 g of soil and 2 g of agar in a final
volume of 100 mL of deionised water. After sterilization the medium was supplemented with
sterilized solutions of sulcotrione (30 mg L-1) and of cycloheximide (50 mg L-1) and poured
into Petri dishes to solidify.
A sterile mineral salt medium (MS), modified from Rousseaux et al.,17 was used for both the
adaptation culture and the biodegradation assays of sulcotrione-degrading strains. This
medium contained per litre: 1.6 g K2HPO4, 0.4 g KH2PO4, 0.2 g MgSO4.7H2O, 0.1 g NaCl,
0.002 g CaCl2, 3.5g (NH4)2SO4, 30 mg sulcotrione, 1 mL of salt stock solution, 1mL of
vitamin stock solution, 1mL of FeSO4.6H2O stock solution (5 g L-1). The salt stock solution
contained 2 g boric acid; 1.8 g MnSO4.H2O, 0.2 g ZnSO4, 0.1 g CuSO4, 0.25 g Na2MoO4 (per

Valorisation des travaux de recherche
215
litre). The vitamin stock solution contained 100 mg L-1 of thiamine-HCl and 40 mg L-1 of
biotin. 0.2 µm filter sterilized sulcotrione, FeSO4.6H2O, and vitamin stock solutions were
added to the medium after sterilization. pH was adjusted at 7.7 with aqueous 0.1 M NaOH
solution. A solid sulcotrione-MS medium obtained by adding 15 g L-1 agar (Biokar
Diagnostics, France) was used to test the capacity of the strains to growth on such solid
medium.
Triptic soy broth medium (TS) obtained by adding 15 g L-1 agar (Biokar Diagnostics, France)
supplemented with sulcotrione (30 mg L-1) was used to purify bacterial strains by successive
stricking. A sulcotrione-(TS) medium (BioMérieux, France) was also used to improve
microbial biomass culture during batch cultures for degradation assays. Microbial biomass
was harvested by centrifugation (15 min, 1600 g at 4°C) and the pellet was washed twice with
Knapp buffer (1 g KH2PO4 ; 1 g K2HPO4 ; 4 mg FeCl3 ; 40 mg MgSO4.7H2 , per litre of
deionized water, pH 6.6) as described earlier18 before being resuspended in sulcotrione-MS
medium to obtain an absorbance of ca 0.3 at 600 nm.
2.3.2. Adaptation cultures to the sulcotrione
Ten grams of soil 4S were suspended in 90 mL 0.9% NaCl and placed on orbitary shaker at
200 rpm for 30 min. Duplicates were carried out. Five hundred µL soil suspension aliquots
were plated on NS-sulcotrione or NS medium. Plates were incubated in the dark at 28°C for 7
days. Growing colonies identified as sulcotrione-resistant strains and/or possible degraders
were selected, grown in 100 mL of Triptic soy broth (TS) medium supplemented with
sulcotrione for 24 hours at 30°C under agitation. They were then plated on TS agar with
sulcotrione.
For each isolated bacterial strain, a selective adaptation step was then carried out by
performing five successive cultures (A1 to A5) in 100 mL sulcotrione MS medium at 28°C
under agitation at 150 rpm for 20 days. Subsequent studies were performed using A5
adaptation solutions.
2.3.3. Estimation of sulcotrione-degrading capabilitiy of the bacterial isolates
One mL of the A5 bacterial culture was grown in TS supplemented with sulcotrione. After 15
hours of culture, bacterial cells were harvested in the late-exponential-growth phase, by
centrifugation (15 min, 1100 g, 4 °C). The bacterial pellet was washed twice with Knapp

Valorisation des travaux de recherche
216
buffer and was resuspended in sulcotrione-MS medium to obtain an absorbance of ca 0.3 at
600 nm. They were then incubated at 28°C in the dark on an orbital shaker (200 rpm). One
mL aliquot of each bacterial suspension was then regularly sampled (0, 6, 15 and 27 days) and
analyzed to establish the kinetics of degradation of sulcotrione and of appearance of its main
metabolites. These degradation tests were realized in triplicate. One control without bacterial
inoculums was also included.
2.3.4. ARDRA, REP-PCR, partial 16S rRNA cloning and sequencing
The extraction of bacterial genomic DNA was done following the protocol previously
described by Cheneby et al.19 and purified using the GENECLEAN® Turbo Kit according to
the manufacturer’s instructions (MP Biomedicals, Europe). DNA quality was checked by 1%
(w/v) agarose gel 1x TBE buffer and quantified by spectrophotometry (NanoDrop® 2000,
Thermo Fisher Scientific).
Partial 16S rRNA sequences were amplified by PCR in 50 µL volume reaction containing 1
µM concentration of the universal primers 27f (5’-AGA GTT TGA TCM TGG CTC AG-3’)
and 1492r (5’-TAC GGH TAC CTT GTT ACG ACT T-3’), 200 µM of each dNTP (Promega,
USA), 2.5 U of GoTaq DNA polymerase (Promega, USA) and 25 ng of DNA.20 PCR
reactions were carried out in a PTC-200 gradient cycler (MJ Research) under the following
conditions: 5 min at 94°C, 35 cycles of 1 min at 94°C, 1 min at 55°C, and 2 min at 72°C, plus
an additional 15-min cycle at 72°C. PCR products were then separated by electrophoresis on a
1% agarose gel, purified using the GENECLEAN® Turbo Kit and quantified by
spectrophotometry.
ARDRA profiles were obtained by digesting 16S rRNA PCR fragments (7 µL) with the
restriction endonucleases Alu I and Hae III (New England Biolabs®) overnight at 37°C. The
obtained restriction fragments for each isolate were separated on a 2% (v/w) Metaphor
agarose gel in 1x TBE buffer (80 V for 3.5 h).
Repetitive extragenic palendromic PCR (REP-PCR) was carried out with REP1R-I and
REP2-1 primers which targets the conserved stem of each REP-like sequence in the
genome.21 DNA amplification was carried out in a 25 µL mixture using 50 ng of extracted
DNA as template in 5 µL of 5x Giescher buffer [83 mM (NH4)2SO4; 335 mM Tris HCl, pH
8.8; 150 mM β -mercaptoethanol; 33.5 µM EDTA, pH 8.8], 6.5 mM MgCl2, 0.16 mg mL-1
Bovine Serum Albumin (BSA, New England Biolabs®), 1.25 mM of each dNTP, 0.1 µL mL-1

Valorisation des travaux de recherche
217
dimethyl sulfoxide, 2 µM of each of the two primers (REP1R-I and REP2-1) and 1 U of
GoTaq DNA polymerase. Amplifications were carried out in a PTC-200 gradient cycler (MJ
Research): 1 cycle at 95 °C for 6 min, 30 cycles at 94°C for 1 min, 40°C for 1 min and 65°C
for 4 min, plus an extension cycle at 65°C for 16 min.17 Amplified DNA fragments were
separated by electrophoresis on 1% agarose gel. PCR reactions were performed independently
in duplicate to ensure the consistency of the REP-profiles obtained.
16S rRNA PCR products were cloned into the pGEM-T Easy vector according to provider’s
recommendation (Promega, USA). Recombinant plasmids of interest were then purified by
using Wizard® Plus Minipreps DNA Purification Systems kit (Promega). Both DNA strands
of the insert were sequenced with an Applied Biosystems Hitachi 3130xl Genetic Analyzers
by using SP6 and T7 primers. Sequences of recombinant clones from the degrading and the
non-degrading strains were treated by SeqManNGen® software (DNASTAR, USA) to provide
consensus sequences and then were compared to known sequences found in the GenBank
database using Blast.22 Sequences were deposited in GenBank under the accession numbers
JF303891 (1TRANS strain) and JF303892 (1OP strain). Multiple alignments were realised
using the ClustalX programme.23 Phylogenetic tree was elaborated using the NJ plot
programme.24
3 RESULTS AND DISCUSSION
3.1. Sulcotrione degradation kinetics in Perpignan soils
The microflora of the Perpignan soil was previously shown to transform sulcotrione to CHD
and CMBA by cleaving the bound between 1,3-cyclohexanedione (CHD) and the benzoic
ring.3 Here we report the transformation of sulcotrione observed only in 4S and C soils and
not in both sterilized soil (StC and St4S), further confirming that the predominace of the biotic
pathway for sulcotrione dissipation in the soil of Perpignan. Similar kinetics of dissipation
were observed in C and 4S soils which were differing by the herbicide treatment, the first
being never treated with it and the last having been treated four times at the agronomic dose
24 months ago. Indeed, after 30 days of incubation (treatment 1), almost 90 % of the applied
sulcotrione was dissipated with half-lives averaging 8 days, without significant differences
between C and 4S plots. For both soil no lag phase period was observed, which suggests that
co-metabolic process is responsible for sulcotrione dissipation. By fitting sulcotrione
dissipation kinetics with R² values close to 0.9, one could observe similar half life (DT50 ≈ 8

Valorisation des travaux de recherche
218
days) and dissipation rates (k ≈ 0.08 day-1) in C and 4S soils (Table 1). As compared to the
literature, sulcotrione dissipation recorded in the soil of Perpignan is rather rapid when
considering DT50 values.3,4,6-8,25 In addition, since the plots C and 4S showed similar capacity
to dissipate sulcotrione, beside different herbicide treatment, the possible accelerated
biodegradation (ABD) of sulcotrione seems unlikely. To further explore this possibility, C
and 4S soils were submitted to 3 additional sulcotrione treatments under laboratory conditions
(T2D30, T3D60 and T4D90). Similar kinetics of dissipation of sulcotrione and kinetics
parameters (k and DT50) were observed in these two plots differing by their scenario of
exposure to the sulcotrione (Table 1). In addition, no correlation could be found between the
rate of degradation (k) and the number of sulcotrione treatments. These results further confirm
that sulcotrione degradation is most likely resulting from co-metabolic process and that
accelerated biodegradation was not occurring in the soil of Perpignan. This feature was also
observed for several other herbicides belonging to s-triazine or substituted ureas families, for
which degradation was for a long period resulting from co-metabolic activity of the soil
microflora prior to the adaptation of the soil microflora to metabolic degradation relying on
specific catabolic pathways. The switch between co-metabolic and metabolic degradation of
these two classes of herbicides occurred after almost 30 years of repeated exposure to these
compounds of the agricultural fields. In order to get a better insight in the processes involved
in the dissipation of sulcotrione from the soil of Perpignan, adaptation cultures already shown
to be successful to isolate different pesticide degrading strains including atrazine, isoproturon,
carbofuran, mesotrione was established.10-11,26-29
3.2. Isolation of sulcotrione-degrading bacterial cultures
The first step of the isolation process relied on the use of a soil agar medium NS
supplemented with sulcotrione, adapted from Ferreira et al.,16 and which was shown to
promote the growth of pesticide-degrading bacterial strains known to be difficult to cultivate
on conventional media, like most of the soil microbes. Starting from soil suspensions plated
on NS-sulcotrione medium, numerous bacterial colonies showing different morphotypes
grew. Eight colonies of the dominant morphotype, forming little white colonies, showing
viscid consistency without mycelial growth were further purified by successive plating.
Excepted their opaque or translucent appearance, these isolates showed comparable
morphology, forming white colonies with a diameter of ca 2.5 mm and following

Valorisation des travaux de recherche
219
characteristics: smooth, circular form, convex elevation. Each of the eight colonies was then
cultivated repeatedly on MS-sulcotrione (A1 → A5) where they were able to grow.
Nevertheless, as agar can also represent a source of carbon, the aptitude of this bacterial
isolates to degrade sulcotrione was checked in MS-sulcotrione liquid medium.
3.3. Estimation of sulcotrione-degrading capacity of bacterial isolates
To get enough microbial biomass to set up a batch experiment to follow sulcotrione
transformation, each of the eight cultures were grown for 15 h in TS-herbicide medium. They
were then diluted to 0.3 OD at 600 nm in MS herbicide-supplemented medium. Dissipation of
herbicide and concomitant accumulation of CMBA and CHD in the culture medium was
monitored by high performance liquid chromatography. Among the eight isolates tested here,
only 1OP showed an increased OD from 0.3 to ca 0.7 after 30 days of incubation (Fig. 2),
reflecting a bacterial growth using sulcotrione as carbon source. 1TRANS, one of the seven
non growing strains was chosen as a non-degrading control. No significant growth was
measured neither for 1OP nor 1TRANS in MS sulcotrione-free medium under the same
incubation conditions. One could conclude that no interfering carbon sources are contained in
MS medium and, that sulcotrione most likely constitutes the sole carbon and energy source
for the growth of 1OP strain. HPLC/UV monitoring clearly showed the dissipation of
sulcotrione in 1OP culture and not in the two controls 1TRANS, the non-degrading isolate,
and the uninoculated MS-sulcotrione medium (Fig. 3). Sulcotrione dissipation was clearly
effective after six days lag phase which was in accordance with the lag phase observed for the
biomass development (Fig. 2). One could hypothesize that this lag phase is the time necessary
to induce the expression of the genes and the synthesis of the catabolic enzymes responsible
for sulcotrione transformation. From the seventh day of incubation, sulcotrione dissipation
kinetics could be reasonably described by a first-order kinetics (C = 121 e-0.081t, R² = 0,993, n
= 3). DT50 could be estimated to ca 15 days after the beginning of the incubation.
During the time course of sulcotrione transformation, one could observe the appearance of 2-
chloro-4-mesylbenzoic acid (CMBA), known as the main metabolite of sulcotrione herbicide
(Fig. 3). It has to be noticed that this metabolite was not observed in none of the two controls
1TRANS, the non-degrading isolate, and in the uninoculated MS-sulcotrione medium. CMBA
was detected from the sixth day of incubation which is in accordance with the lag phase
observed for the initiation of sulcotrione disappearance. After 1 month of incubation, the

Valorisation des travaux de recherche
220
concentration of CMBA reached 12 ± 1 mg L-1 (48 ± 3 µM) in the culture medium. This
result was confirmed by GC/MS analysis after derivatization of CMBA to give the
corresponding methyl ester. In addition, one could observe that molar recovery taking into
account sulcotrione and CMBA was slightly decreased between day 15 (0.065 ± 7 µmole L-1
+ 0.025 ± 2 µmole L-1, respectively) and day 27 (0.017 ± 2 µmole L-1 + 0.048 ± 3 µmole L-1,
respectively), suggesting a possible degradation of benzoic acid derivative CMBA by 1OP.
Finally, based on the results presented here, one could conclude that 1OP strain was capable
of using sulcotrione as a carbon source for its growth by degrading sulcotrione to CMBA.
1,3-cyclohexanedione (CHD), the other degradation product of sulcotrione, previously
identified at very low concentrations in the Perpignan soil3 was not detected (< 0.2 mg L-1) in
the culture media of strain 1OP neither by HPLC/UV nor by HPLC/MS/MS. One could
explain this discrepancy by chemical transformation by hydrolysis of CHD to 5-oxohexanoic
acid at pH>7, or by an oxidation mechanism catalysed by enzymatic cleavage of C-C bonds
in the β -diketone before entering the tricarboxylic acid cycle (TCA), as previously
reported.30,31-32 It is noteworthy that for mesotrione, another herbicide belonging to triketone
family but with a nitro radical, a minor metabolic route catalysed by a Bacillus sp. with a
comparable mechanism was reported.11-12
3.4. Characterization of sulcotrione degrading (1OP) and non-degrading (1TRANS) strains
by ARDRA, REP-PCR and 16S rDNA sequencing
The amplified ribosomal rDNA restriction analysis technique (ARDRA) has been reported as
a reliable and valuable tool for phylogenetic and taxonomic studies of large sets of cultured or
uncultured microorganisms from different habitats.33-36 In this study, 1OP and 1TRANS
isolates strains were characterized by ARDRA with AluI and HaeIII restriction
endonucleases. 1OP and 1TRANS bacterial strains showed similar ARDRA profiles made of
four major bands of approximately 550, 380, 210 and 70 bp in length. In order to get more
taxonomical information than the genus level offered by ARDRA fingerprinting 1OP and
1TRANS isolates were analysed by repetitive extragenic palindromic-PCR (REP-PCR) which
provides a rapid identification of bacterial isolates and are useful for the analysis of
prokaryotic genomes.18,21 Again 1OP and 1TRANS bacterial strains showed similar REP-
PCR profiles made of five major bands of approximately 550, 400, 270, 150 and 100 bp in
length (Fig. 4). This observation suggests that 1OP and 1TRANS belong to the same bacterial

Valorisation des travaux de recherche
221
species. 16S rRNA sequencing further revealed that these two isolates showed identical 16S
rRNA sequences. We conclude that these isolates were different strains from the same
species. Comparison of 16S rRNA sequence with those found in GenBank showed highest
level of similarity (99% of similarity) to Pseudomonas sp. Furthermore, multiple alignments
revealed that 1OP and 1TRANS 16S rRNA sequences were closely related to Pseudomonas
putida strains both for 1OP and 1TRANS (Fig. 5). Based on 16S rDNA phylogenetic data, we
suggest naming our sulcotrione-degrading isolate Pseudomonas sp. 1OP. It was classified as
Bacteria, phylum Proteobacteria, class Gammaproteobacteria, order Pseudomonadales,
family Pseudomonaceae and genus Pseudomonas, with the following characteristics: rod
shaped, Gram-negative, one or more flagella providing motility, positive catalase test and
non-spore forming bacteria. Several strains belonging to Pseudomonas genus able to degrade
various pesticides have been described earlier among which Pseudomonas sp. ADP degrading
atrazine,37Pseudomonas cepacia DBO1 degrading 2,4-dichlorophenoxyacetic,38 Pseudomonas
sp. degrading propoxur (2-isopropoxyphenyl-N-methylcarbamate).39 Interestingly, the
isolation of two strains identical from the taxonomical point of view but differing by their
sulcotrione-degrading capacity, highlights the versatility of the degrading character. One
could hypothesize that such versatility might be due to mobile genetic elements (MGE)
surrounding catabolic genes in the genome of 1OP. MGE as plasmids, transposons, integrons,
genomics islands and bacteriophages were acting as a powerful evolutionary engine and
regulator on microbial populations and were collectively referred as drivers or vectors of “the
horizontal gene pool”40,41. Such properties are known to be associated with various strains of
genus Pseudomonas and its relatives, and were often specified by large (> 50kb) catabolic
plasmids, though the catabolic genes could also be loaded on transposable elements.42-44
Further work will aim to differentially characterize the genetic determinants of the
sulcotrione-degrading ability of Pseudomonas sp. 1OP.
4 CONCLUSION
This work confirms the aptitude of the soil microflora of Perpignan to transform the
sulcotrione, although no enhanced biodegradation was observed after repeated exposure to
this herbicide. We report for the first time the isolation of a sulcotrione-degrading strain,
identified as Pseudomonas putida strain 1OP. This bacterial strain was shown to transform
sulcotrione to 2-chloro-4-mesylbenzoic acid (CMBA). Nevertheless, 1,3-cyclohexanedione

Valorisation des travaux de recherche
222
(CHD), another known sulcotrione metabolite was not detected. Based on the evolution of the
molar recovery of sulcotrione and CMBA during the time course of dissipation kinetics, we
hypothesized that Pseudomonas putida strain 1OP may start degrading the benzoic acid
derivative (CMBA) several days after its formation in the liquid medium.
ACKNOWLEDGEMENTS
The PhD work of Christophe Calvayrac was funded by Bayer Cropscience. The authors also
acknowledge the members of Laboratories of UPVD (LCBE, LGDP, IMAGE and IUT),
INSA Toulouse and INRA Dijon, France.
REFERENCES
1 Mamy L, Gabrielle B and Barriuso E, Comparative environmental impacts of
glyphosate and conventional herbicides when used with glyphosate-tolerant and non-
tolerant crops. Environ Pollut 158:3172-3178 (2010).
2 Chaabane H, Vulliet E, Joux F, Lantoine F, Conan P, Cooper JF and Coste CM,
Photodegradation of sulcotrione in various aquatic environments and toxicity of its
photoproducts for some marine micro-organisms. Water Res 41:1781-1789 (2007).
3 Chaabane H, Vulliet E, Calvayrac C, Coste CM and Cooper JF, Behaviour of
sulcotrione and mesotrione in two soils. Pest Manag Sci 64:86-93 (2008).
4 Cherrier R, Perrin-Ganier C and Schiavon M, Degradation of sulcotrione in a brown
soil with various organic matters. Agronomie 24:29-33 (2004).
5 Maeghe L, Desmet EM and Bulcke R, Soil activity and persistence of sulcotrione and
mesotrione. Commun Agric Appl Biol Sc 69:41-48 (2004).
6 Rouchaud J, Thirion A, Callens D and Bulcke R, Soil Dissipation of the Post-
Emergence Herbicide Sulcotrione in Maize Crops Treated with Organic Fertilizers.
Bull Environ Contam Toxicol 57:398-405 (1996).
7 Rouchaud J, Neus O, Bulcke R, Cools, K and Eelen H, Sulcotrione Soil Metabolism in
Summer Corn Crops. Bull Environ Contam Toxicol 61:669-676 (1998a).
8 Rouchaud J, Neus O, Callens D and Bulcke R, Sulcotrione soil persistance and
mobility in summer maize and winter wheat crops. Weed Res 38:361-371 (1998b).

Valorisation des travaux de recherche
223
9 Chaabane H, Cooper JF, Azouzi L and Coste CM, Influence of soil properties on the
adsorption-desorption of sulcotrione and its hydrolysis metabolites on various soils. J
Agric Food Chem 53:4091-4095 (2005).
10 Durand S, Amato P, Delort AM, Combourieu B and Besse-Hogan P, First isolation
and characterization of a bacterial strain that biotransforms the herbicide mesotrione.
Lett Appl Microbiol 43:222-228 (2006a).
11 Durand S, Legeret B, Martin A, Sancelme M, Delort A, Besse-Hoggan P and
Combourieu B, Biotransformation of the triketone herbicide mesotrione by a Bacillus
strain. Metabolite profiling using liquid chromatography/electrospray ionization
quadrupole time-of-flight mass spectrometry. Rapid Commun Mass Spectrom
20:2603-2613 (2006b).
12 Durand S, Sancelme M, Besse-Hoggan P and Combourieu B, Biodegradation pathway
of mesotrione: Complementarities of NMR, LC-NMR and LC-MS for qualitative and
quantitative metabolic profiling. Chemosphere 81:372-380 (2010).
13 Batisson I, Crouzet O, Besse-Hoggan P, Sancelme M, Mangot JF, Mallet C and
Bohatier J, Isolation and characterization of mesotrione-degrading Bacillus sp. from
soil. Environ Pollut 157:1195-1201 (2009).
14 Crouzet O, Batisson I, Besse-Hoggan P, Bonnemoy F, Bardot C, Poly F, Bohatier J
and Mallet C, Response of soil microbial communities to the herbicide mesotrione: A
dose-effect microcosm approach. Soil Biol Biochem 42:193-202 (2010).
15 Mashimoto N, Aoyama T and Shiori T, New methods and reagents in organic
synthesis. A simple efficient preparation of methyl esters with
trimethylsilyldiazomethane (TMSCHN2) and its application to gas chromatographic
analysis of fatty acids. Chem Pharm Bull 29:1475-1478 (1981).
16 Ferreira EPDB, Dusi AN, Costa JR, Xavier GR and Rumjanek NG, Assessing
insecticide and fungicide effects on the culturable soil bacterial community by
analyses of variance of their DGGE fingerprinting data. European Journal of Soil
Biology 45:466-472 (2009).

Valorisation des travaux de recherche
224
17 Rousseaux S, Hartmann A and Soulas G, Isolation and characterisation of a new
Gram-negative and Gram-positive atrazine degrading bacteria from different French
soils. FEMS Microbiol Ecol 36:211-222 (2001).
18 Devers M, Henry S, Hartmann A and Martin-Laurent F, Horizontal gene transfert of
atrazine-degrading genes (atz) from Agrobacterium tumefaciens St96-4 Padp::Tn5 to
bacteria of maize-cultivated soil. Pest Manag Sc 61:870-880 (2005).
19 Cheneby D, Perrez S, Devroe C, Hallet S, Couton Y, Bizouard F, Luterig G, Germon
JC and Philippot L, Denitrifying bacteria in bulk and maize-rhizospheric soil: diversity
and N2O-reducing abilities. Can J Microbiol 50:469-474 (2004).
20 Gurtler V and Stanisich VA, New approaches to typing and identification of bacteria
using the 16S-23S rDNA spacer region. Microbiology 142:3-16 (1996).
21 Versalovic J, Koeuth TR and Lupski J, Distribution of repetitive DNA sequences in
eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Res
19:6823-6831 (1991).
22 GenBank®. Available : http://blast.ncbi.nlm.nih.gov/Blast.cgi
23 Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F and Higgins DG, The ClustalX
windows interface: flexible strategies for multiple sequence alignment aided by
quality analysis tools. Nucleic Acids Res 24:4876-4882 (1997). Available :
http://www-igbmc.u-strasbg.fr/BioInfo/
24 Perriere G and Gouy M, WWW-Query: An on-line retrieval system for biological
sequence banks. Biochimie 78:364-369 (1996). Available: ftp:://pbil.univ-
lyon1.fr/pub/mol_phylogeny/njplot
25 Cherrier R, Boivin A, Perrin-Ganier C and Schiavon M, Sulcotrione versus atrazine
transport and degradation in soil columns. Pest Manag Sci 61:899-904 (2005).
26 Govantes F, García-Gonzalez V, Porrua O, Platero AI, Jimenez-Fernandez A and
Santero E, Regulation of the atrazine-degradative genes in Pseudomonas sp. strain
ADP. FEMS Microbiol Lett 310:1-8 (2010).
27 El-Sebai T, Lagacherie B, Soulas G and Martin-Laurent F, Isolation and
characterisation of an isoproturon-mineralising Methylopila sp. TES from french
agricultural soil. FEMS Microbiol Lett 239:103-110 (2004).

Valorisation des travaux de recherche
225
28 Hussain S, Devers-Lamrani M, El Azhari N and Martin-Laurent F, Isolation and
characterization of an isoproturon mineralizing Sphingomonas sp. strain SH from a
French agricultural soil. Biodegradation DOI: 10.1007/s10532-010-9437-x. (2010).
29 Yano H, Garruto CE, Sota M, Ohtsubo Y, Nagata Y, Zylstra GJ, Williams PA and
Tsuda M, Complete sequence determination combined with analysis of
transposition/site-specific recombination events to explain genetic organization of
IncP-7 TOL plasmid pWW53 and related mobile genetic elements. J Mol Biol 369:11-
26 (2007).
30 Dangel W, Tschech A and Fuchs G, Enzyme reactions involved in anaerobic
cyclohexanol metabolism by a denitrifying Pseudomonas species. Arch Microbiol
62:3245-3250 (1989).
31 Grogan G, [beta]-Diketone hydrolases. International Symposium on Biocatalysis and
Biotransformations No5, Darmstadt Germany. Journal of molecular catalysis. B,
Enzymatic vol 19-20:73-82 (2002).
32 Grogan G, Emergent mechanistic diversity of enzyme-catalysed [beta]-Diketone
cleavage. Biochem J 388: 721-730 (2005).
33 Vaneechoutte M, Rossau R, de Vos P, Gillis M, Janssens D, Paepe N, De Rouck A,
Fiers T, Claeys G and Kersters K, Rapid identification of bacteria of the
Comamonadaceae with amplified ribosomal DNA-restriction analysis (ARDRA).
FEMS Microbiol Lett 72:227-233 (1992).
34 Vaneechoutte M, De Beenhouwer H, Claeys G, Verschraegen G, De Rouck A, Paepe
N, Elaichouni A and Portaels F, Identification of Mycobacterium species by using
amplified ribosomal DNA-restriction analysis. J Clin Microbiol 31:2061-2065 (1993).
35 Martinez-Murcia AJ, Acinas SG and Rodriguez-Valera F, Evaluation of prokaryotic
diversity by restrictase digestion of 16S rDNA directly amplified from hypersaline
environments. FEMS Microbiol Ecol 36:247-256 (1995).
36 Martin-Laurent F, Philippot L, Hallet S, Chaussod R, Germon JC, Soulas G, and
Catroux G, DNA extraction from soils: old bias for new microbial diversity analysis
method. Appl Environ Microbiol 67: 2354-2359 (2001).
37 Mandelbaum R, Allan D and Wackett LP, Isolation and characterization of a
Pseudomonas sp. that mineralizes the s-triazine herbicide atrazine. Appl Environ
Microbiol 61:1451-1457 (1995).

Valorisation des travaux de recherche
226
38 Daugherty DD and Karel SF, Degradation of 2,4-dichlorophenoxyacetic acid by
Pseudomonas cepacia DBO1(pRO101) in a dual-substrate chemostat. Appl Environ
Microbiol 60:3261-7 (1994).
39 Kamanavalli CM and Ninnekar HZ, Biodegradation of propoxur by Pseudomonas
species. World Journal of Microbiology & Biotechnology 16:329-331 (2000).
40 Thomas CM and Nielsen KM, Mechanisms and barriers to horizontal gene transfer
between bacteria. Nat Rev Microbiol 3:711-721 (2005).
41 Frost LS, Leplae R, Summers AO and Toussaint A, Mobile genetic elements: the
agents of open source evolution. Nat Rev Microbiol 3:722-732 (2005).
42 Ogawa N, Chackrabarty AM and Zaborina O, Degradative plasmid, in Plasmid
Biology, ed. by Funnell B, E, Philipps G J, ASM Press Washington, DC, pp. 341-392
(2004).
43 Williams PA, Jones RM and Zylstra GJ, Genomics of catabolic plasmids, in
Pseudomonas, ed. by Ramos JL, Kluwer Academic/Plenum Publishers, New-York,
vol 1, pp. 165-195 (2004).
44 Yano H, Garruto CE, Sota M, Ohtsubo Y, Nagata Y, Zylstra GJ, Williams PA and
Tsuda M, Complete sequence determination combined with analysis of
transposition/site-specific recombination events to explain genetic organization of
IncP-7 TOL plasmid pWW53 and related mobile genetic elements. J Mol Biol 369:11-
26 (2007).

Valorisation des travaux de recherche
227
Fig. 1: Electronic impact/mass spectrometry (EI/MS) spectrum of methylated CMBA
analytical standard
(20 mg L-1).
!
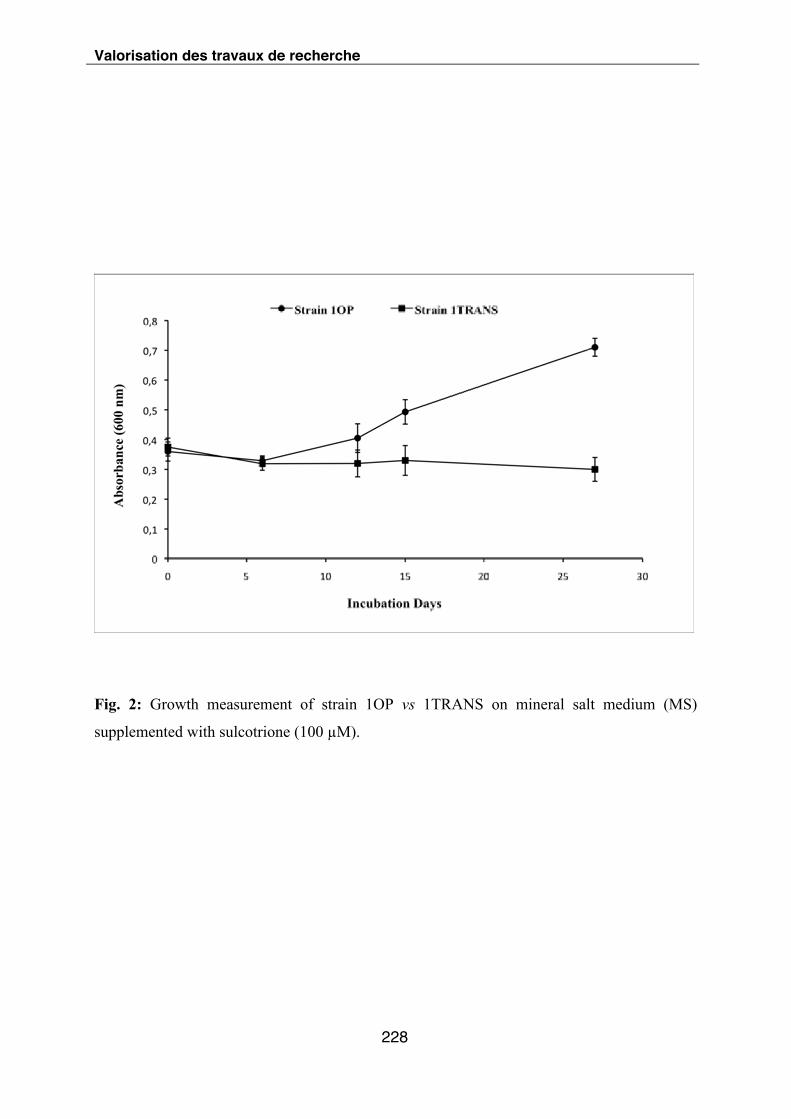
Valorisation des travaux de recherche
228
Fig. 2: Growth measurement of strain 1OP vs 1TRANS on mineral salt medium (MS)
supplemented with sulcotrione (100 µM).
!

Valorisation des travaux de recherche
229
Fig. 3: Dissipation of sulcotrione and accumulation of CMBA by strain 1OP vs control and
strain 1TRANS in liquid mineral salt medium MS supplemented with sulcotrione (100 µM).
Errors bars indicated standard deviation.
!

Valorisation des travaux de recherche
230
Fig. 4: REP PCR analysis of the isolates 1OP and 1TRANS. REP-PCR were performed in
duplicate for each isolate with 1 ng µL -1 or 2 ng µL -1 of DNA. Lane M represents the
molecular-weight marker 1 Kb Plus DNA Ladder (sizes indicated in base pairs). Lanes 1 and
2: 1TRANS (1 ng µL-1), lanes 3 and 4: 1TRANS (2 ng µL-1), lanes 5 and 6: 1OP (1 ng µL-1),
lanes 7 and 8: 1OP (2 ng µL-1).
!

Valorisation des travaux de recherche
231
Fig. 5 Neighbor-joining phylogenetic analysis resulting from the multiple alignment of 16S
rRNA gene sequences of Pseudomonas putida strains 1TRANS (NBD) and 1OP (BD) with
those retrieved from GenBank database Arthrobacter creatinolyticus strain F75208
(HQ908759), Arthrobacter nicotianae strain HMF5 (GU220490), Streptomyces sp. SNI5
(GU320191), Rhizobium leguminosarum strain V-6 (GU306144), Agrobacterium tumefaciens
strain YQH34 (HQ143653), Methylopila sp. JZL-4 (FJ502233), Burkholderia cepacia strain
RFW-8 (HQ650586), Comamonas testosteroni strain H18 (EU887829), Acidovorax sp.
p4(HQ652596), Pseudomonas putida strain A (HQ697262), Pseudomonas putida strain T2g
(AB539810), Pseudomonas aeruginosa strain P28 (HQ697285), Pseudomonas fluorescens
strain C7R12 (AM229082), Pseudomonas fluorescens strain (HQ880245), Pseudomonas sp.
strain ADP (AM088478). Bootstrap values greater than 90 % are marked as black circles. The
phylogenetic distance is shown on a scale bar.
!

Valorisation des travaux de recherche
232
Table 1: Degradation coefficients (k) and half-live (DT50) for all soils.
k (days-1) DT50 (days) R2
Treatment 1 (T1D0)
0.083
8
0.894 Treatment 2 (T2D30) 0.060 11.5 0.955
C Plot Treatment 3 (T3D60) 0.100 7 0.989 Treatment 4 (T4D90) 0.100 7 0.962
Mean value
0.085 ± 0.020
8 ± 2.5
-
Treatment 1 (T1D0)
0.088
8
0.872
Treatment 2 (T2D30) 0.093 7 0.936 4S Plot Treatment 3 (T3D60) 0.068 10 0.955
Treatment 4 (T4D90) 0.089 8 0.828
Mean value
0.084 ± 0.012
8 ± 2.0
-
StC Plot
Treatment 1 (T1D0)
0.0004
-
-
St4S Plot
Treatment 1 (T1D0)
0.0004
-
-

Valorisation des travaux de recherche
Publication n° 2
Pest Management Science Pest Manag Sci 64:86-93 (2008)
Behaviour of sulcotrione and mesotrione scí where sciencemeets business
in two soils www.sod.org
Hanene Chaabane, 1 Emmanuelle Vulliet,2 Christophe Calvayrac, 1
Camille-Michel Coste 1 and Jean-Franyois Cooper 1 * 1 Laboratoíre de Chímíe des Bíomo/écu/es et de /'Envíronnement, Centre de Phytopharmacíe, Uníversíté de Perpignan, 52 avenue Pau/
A/duy, 66860 Perpignan, France
2Service Central d'Analyses, CNRS, Chemin du Canal - BP 22- 69390 Vemaison, France
Abstract: The behaviour of sulcotrione, a recendy introduced triketone herbicide, in various soil types was studied under laboratory conditions. In particular, degradation and sorption processes were examined on Ghent and Perpignan soils. Kinetics showed that the degradation ofsulcotrione \vas influenced by biotic andlor abiotic factors. Half-lives ranged between 45 and 65 days. Among the degradation compounds identified were 1,3-cyc1ohexanedione (CHD) and 2-chloro-4-mesyl benzoic acid (CMBA) , previously described as hydrolysis products, and, under special conditions, a derivative ofphenylheptanoic acid (PHD). This new degradation product suggested that sulcotrione could follow two possible pathways in the soil, as in water. During the sorption study, a moderate retention of sulcotrione and CMBA relative to CHD and PHD, which were high1y adsorbed whatever the soil type, was reported. Experlments carrled out under the same conditions for sulcotrione and mesotrione, another triketone herbicide recommended in maize culture, made it possible to compare the two·triketones and to conclude that they exhibited relatively similar behaviour in the soil, Le. that their leaching potential needs to be proper1y addressed and risks evaluated. © 2007 Society of Chemical Industry
Keywords: sulcotrione; mesotrione; sol1; degradation; degradation products; sorption
1 INTROOUCTION Among the long list of pesticides used for weed contro 1 in maize, two triketone herbicides, sulcotrione [2-(2-chloro-4-mesylbenzoyl) cyclohexane-l ,3-dione] and mesotrione [2-( 4-mesyl-;2-nitrobenzoyl) cydohexane-1,3-dione] (Fig. 1), have proved their efficiency against a broad spectrum of weeds. 1,2 Their target enzyme is 4-hydroxyphenylpyruvate dioxygenase,3-5 the inhibition of which leads to disturbance of plastoquinone and a-tocophérol biosynthesis. Intermil symptoms are bleaching followed by necrosis and death oftreated plants. Sulcotrione and mesotrione are mainly recommended for post-emergence treatments, at respective application doses of 300 and 150 g ha -[ , lower than those of atrazine or pendimethaline.
After direct application andlor foliage wash-off, a large proportion of these triketone herbicides reaches the soil, which always plays a key role in their environmental fate. Soil sorption and degradation, the major processes controlling pesticide actlVlty, persistence and availability for transfer, are affected by soil properties such as pH, day and organic matter corrtents, tJ.~e nature of the chemical arra environmental conditions.
Literature concerning sulcotrione and mesotrione provides only a few reports about their behaviour in soil. Recent work has shown that sulcotrione exhibits
a moderate retention capacity, whatever the selected soil. 6 Comparable results have beeri t;eported for mesotrione.7
For the degradation process, half-lives calculated for sulcotrione and mesotrione have varied respectively between 2 and 144 days and between 4 and 32days.7-10
The aim of this work was to assess the degradation and sorption of sulcotrione and to compare these with the degradation and sorption of mesotrione under the same conditions. Their leaching potential towards water resources could then be evaluated according to their adsorption coefficients and half-lives.
2 MATERIALS ANO METHOOS 2.1 Chemicals Standards of sulcotrione (purity 99.2%, molecular mass 328 and water soIubility 165 mg L -1 at 25 oC) and mesotrione (99.6%, molecular mass 339 and water solubility 160 mg L -1 at 20 OC) were respectively purchased from RiedeI-deHaen Gmbh (Germany) and supplied by Syngenta (France). Both sulcotrione ana mesotrione are weak acidic herbicides, with pKa values of 2.87 and 3.12.
Standards of 1,3-cydohexanedione (CHD) (97.0%) and 2-chloro-4-mesylbenzoic acid (CMBA) (95.0%)
233

Valorisation des travaux de recherche
o o el
6:~ 1iSO'CH'
Sulcotrione Mesotrione
Figure 1. Molecular structures of sulcotrione and mesotrione.
were respective1y supplied by Fluka and Acros Organics. The phenylheptanoic acid derivative (PHD) was synthesised as previously described. 11
Methanol and acetonítrile (HPLC quality) were purchased from Carlo Erba, dichloromethane (for pesticide analysis) from Riede1-deHaen GmbH, trifluoroacetic acid (TFA) (99.0%) fromAldrich, hydroch10-ríe acid (38.0%) from Prolabo and water was Milli-Q quality. Stock solutions of each compound were prepared at 1000 mg L -1 in methanol. For retention studíes, working solutions (0.25, 0.5, 1, 2.5, 10, 20 and 30 mg L -1) were prepared by di1ution of each stock solution in distilled water, with the methano1 percentage never exceeding 3%.
Supe1clean™ EnviChrom P eartridges (3 mL, 0.25 g) were purchased from Supelco and used for solid-phase extraction of soil samples.
2.2 Soils Two soils, Perpignan (France) and Ghent (Be1gium), were se1eeted aecording to their physicoehemical properties (Tab1e 1). The Freneh soi1 (a clayey sandy loam soil, aeeording to FAO classificatíon) was free of any sulcotrione and/or mesotrione treatments, whereas the Be1gian one (a sandy 10am soíl) had been treated yearly with sulcotrione from 1992 until 2003. The two soi1 types were presieved to <2 mm and air dried at ambient temperature. Pedo10gica1 analyses were earried out by Laboratoire d' Ana1yses Agrieoles, Perpignan, Franee, and Laboratory of Herbology, Ghent Universíty, Belgium. Soíl moisture contents were determined in the authors' 1aboratory aecording to the method published by Mathieu and Pieltain 12 as 3.4 and 1.60% for the soil of Perpignan and Ghent respective1y.
2.3 Degradation process In order to assess the relative parts of the abiotic and biotic ways in the degradation proeess, the kinetics of degradation were determined using three protocols:
Table 1. Physicochemical properties of selected soils
CECb Clay (%)
Silt
(%) Sand OCa (meq Ca2+¡ (%) pH (%) 1009-1) CEC (%)
Perpignan 13.9 60.5 25.6 8.1 0.9 Ghent 23.0 41.4 35.6 6.6 2
a Organic Carbono b Cation Exchange Capacity.
Pest Manag Sci 64:86-93 (2008)
15.5 9.4
214.0
Behaviour of sulcotrione and mesotrione in soils
• Perpignan and Ghent unsterilised soi1s maintained at 25 oC (protoco1 1);
• Ghent unsterilised soil maintained at higher temperatures, 40 and 60 oC (protocol 2);
• Perpignan and Ghent soíls twice autoclaved at 120 oC over a period of 30 min, maintained at 25 oC (protocol 3).
Sulcotrione additions were earried out under a laminar flow hood.
For ea eh soíl and for the three protoco1s, seven soíl samp1es (10.0 g) were treated in glass petri dishes with a methanolic solution of sulcotrione (10.0 mg L -1) or mesotrione (5.0 mg L -1, on1y for protocol 1) to give 0.5 or 0.25!J,gg-l, which corresponds to the recommended application dose for each triketone. AH samples were kept in darkness to avoid photodegradation. Soil moisture content was fixed at 30% of dry weíght and maintained constant during the experimento At fixed dates (O, 3, 7, 14, 30, 60 and 120 days), one petri dish of eaeh soil (10.0 g) was taken, extraeted and analysed by high-performance liquid chromatography (HPLC) (see Section 2.5).
2.4 Batch sorption study Aecording to OECD guideline,13 a sorption kinetic study is needed to evaluate the agitation period required to reaeh the equilibrium state, corresponding to adsorption on the most accessible sites. For eaeh compound, 1.0 g of soil was introduced into a centrifugation glass tube contaíning 5.0 mL of working solution (initial concentration 30 mg L -1) and shaken mechanicaHy for 0.5, 1, 2, 4, 8 and 24 h. The soil/solution suspension was then centrifuged at 3500 x g for 10 min and the supematant was analysed by HPLC (Section 2.5). Calcu1ations were performed using an indirect method13 based on the difference between initia1 and equilibrium concentrations in the supernatant, and led to the establishment of distribution coefficient I~ from the following equation:
Kd = (A eq x Vo)/(100 - Aeq) x msoil (1)
where Aeq is the adsorption percentage at the equilibrium state, Va is the ¡ni tia! vo1ume of liquid phase and msoil is the mass of soiL
The agitation time (8 h for sulcotrione and 24 h for its degradation products and mesotrione) deduced from adsorption kinetic plots was used for determining the adsorption isot..1-}erms. Glass tubes containing 5.0 mL oí working solution and 1.0 g of soil were shaken mechanically during this agitation periodo Samples were then centrifuged at 3500 x g for 10 min and the supernatants were analysed by HPLC. Two blanks (soíl with aqueous solution free of chemicals and aqueous solution without soil) were performed under the same eonditions. Adsorption isotherms were obtained by plotting solid-phase concentration, Os (mg kg- 1
), versus liquid phase concentration at equilibrium
87
234

Valorisation des travaux de recherche
H Chaabane er al.
state, Ce,ads (mg L -1). The Freundlich model was used to describe experimental results according to the equation
(2)
where Kfa and linfa are empirical constants representing respectívely the Freundlích coefficient for adsorption and the slope of the isotherm which expresses its degree of non-linearity.
Desorptíon experiments were carried out on soil samples treated at the highest concentration in the adsorption process. After HPLC analysis, the supernatant was removed and replaced with the same volume of distilled water. The remainíng slurry was shaken for the same period (8 and 24 h) and centrifuged at 3500 x g for 10 min, and the supernatant was then removed and analysed. This desorption procedure was repeated 3 times consecutively. The amount of each chemical remaining adsorbed in the soíl after each desorption step, cs, was evaluated. Desorption isotherms were obtained by plotting Cs
against Ce (the concentration of the compound in the liquid phase after each desorption step). The fo11owing Freundlich relationship was used to characterise the desorption process:
where G so is the solid-phase concentration (mg kg- 1)
of the compound adsorbed on the soil at equilibrium status, Ceo is the compound concentration in the liquid phase at equilibrium after adsorption and Kfd
and 11 nfd are respectively the empirical coefficient for desorption and the slope of the curve which expresses the desorption intensity of the compound from the soil.
For both kínetic and isotherm studies, experiments were carried out in duplicate.
2.5 Analytical methodology 2.5. 1 Soil samples Extractíon step. In order to fo11ow triketone degradation, soil samples were extracted with 60 mL and then with 30 mL of acetonitrile + 0.1 M hydrochloric acid (90 + 10 by volume) over 30 and 15 min periods, then filtered on a Whatman filter GF/A. The organic filtrate was evaporated at 30 oC and the acidic aqueous solution was then extracted with dichloromethane (2 x 5.0 mL). The organic phase was evaporated to dryness, solubilised in 0.1 M hydrochloric acid, purified on preconditioned EnvíChrom P cartridge and fina11y eluted with methanol (5 mL). This final extract was analysed by HPLC. One blank (soíl with the
respective volume of methanol free from triketone) was analysed under the same conditions.
Ghromatographíc analysis. Soil extracts were analysed usíng a Shimadzu HPLC apparatus equipped with a Supelco ODS Hypersil C I8 column (5 11m, 250 mm x 4.6mm) and a SPD-I0 Avp UVNis detector set at 254 nm. The mobile phase consisted of a mixture of water acidified' with 0.3 % trifl uoroacetic acid (A W) and acetonitrile (ACN) delivered at a flowrate of 1 mL mín-l. A gradíent system [70/30 (AW/ACN) at t = O min to 50/50 (AW/ACN) at 8 min, maintained for 10 mín] was used to ímprove the separation of the active ingredient and its metabolites during degradatíon and adsorption kinetics.
Analytical performance. The analytical method (soíl extractíon and HPLC analysis) was based on the procedures previously published3,14 and performed for a11 soil/compound couples (Table 2).
2.5.2 Water samples Water samples (for batch sorption study) were directly analysed without any extraction concentratíon step. Chromatographíc analysis was performed under the same condítions as for soíl samples, using isocratic. mode when analysis concerned only one compound Cfor isotherm study, since no degradation was observed during adsorption kinetics). The proportions of mobile phase were respectively 80/20 (AW/ACN) for CHD, 70/30 for CMBA and 50/50 for PHD, sulcotrione and mesotrione. The detection limit was 0.05 mg L -1 for all compounds under our chromatographic conditions.
3 RESUL TS ANO OISCUSSION 3.1 Triketone degradation and soil properties The decline of sulcotrione concentration in soils was plotted against incubation period as presented in Figs 2a and b. Degradation rates, fo11owed over a period of 120 days in Perpignan and Ghent ~oils according to protocol 1, were represented reasonably by first-order kinetics
C = Co x e-kt (4)
(with r2 values higher than 0.9), from which halflife values were determined (these are reported in Table 3). In equation (4), Co is the initial concentration of the triketone, C is the concentration after each incubation period t (days) and k is the apparent kinetic constant.
The disappearance ofsulcotrione over an incubatíon period of 60 days was estimated as 55 and 45% of the initial application dose and half-lives as 45 and 65 days, respectively, on Ghent and Perpignan soils. .
Table 2. Analytical performance of the method used tor soil extraction and HPLC analysis
Oe!ection limi! (¡.¡.g g-1) Recovery rate (%)
88
Sulcotrione
0.05 90(± 10)
CHD
0.001 50 (± 10)
235
CMBA
0.05 80 (± 10)
PHO
0.05
70 (± 10)
Mesotrione
0.01
70 (± 10)
Pes! Manag Sci 64:86-93 (2008)

Valorisation des travaux de recherche
'1 + mesotrione sulcotrione
í \. 01 0:6: 01 .6 (,)
+ ;:1!
• o 50 100 150
(a) Incubation period (days)
'1 + mesotrione -'~ sulcotrione
~---- : tO.5'~> .. , ~
O+------J~--.-~--------._--~~----~
o 50 100
(b) Incubation period (days)
Figure 2. Rate of degradation of sulcotrione and mesotrione, according to protocol 1, in (a) Ghent and (b) Perpignan soil.
Table 3. Degradation coefficients, K, and half-lives, DTsQ , for all
soil/triketone couples
Sulcotrione Mesotrione
OT50 OT50
150
K(day-1) (days) (2 K(day-1) (days) (2
Ghent
Perpignan
0.015 0.006
45 0.99 65 0.91
0.02 0.13
34 0.95 5 0.92
During this monitoring, degradation produets oE sulcotrione were identified as CHD and CMBA and sometimes PHD, and quantified. Their levels are plotted against incubation period in Figs 3a and b. These degradation produets had previously been Eound during hydrolysis in aqueous media, where their presence was particularly pH dependent. 11 Whatever the soil type, analyses oE soil extracts showed the presence oE CHD and CMBA, but PHD was deteeted only in the Ghent soil which had a pH (6.6) slightly
·lower than that of the Perpignan soi1. Therefore, the two degradarion pathways already described in
,aqueous media can be found in the soi1, depending on ,pH value.
The firsr degradation pathway (A) consists oE a hydrolytic cleavage berween 1,3-cyclohexanedione and rhe benzoic ring, leading ro CHD and CMBA. In Perpignan soil, only rhis degradarion pathway occurred, since pH condirions were re1atively alkaline.
In Belgian soil, in parallel ro this firsr pathway, sulcotrione could Eollow another mechanism (B) leading to PHD which is larer rransformed into CMBA. In
Pest Mana!! Sci 64:86-93 (2008)
Behaviour of sulcotrione and mesotrione in soils
0.003 s CMBA X
i: 0.002 Si:
"O E ¿ (.) 0.001
o~: :
PHD :¡:: sulcotrione + CHD
:¡::
~----, O 50 100 150
(a) Incubation period (days)
0.003
• CHD R CMBA )K Sulcotrione
í 0.002 C1
"O E ¿
:tSK (.) 0.001 )K
ilIi ¡¡¡
i!I )K !I
O • • O 50 100
(b) Incubation period (days)
Figure 3. Formation of degradation products of sulcotrione in (a) Ghent and (b) Perpignan soil.
150
water and using a high initial sulcotrione concentration (30 mg L -1), during this last step, 1-(2-chloro-4-mesyl)ethanone was detected as an intermediary compound (lC) (Fig. 4), while it was nor Eound in the soil under the present experimental conditions.
However, although the pH role appears evident, it is suggested that biotic Eactors have a possible infiuence on the degradation proeess. In fact, experiments carried out in Perpignan soil after sterilisation (protocol 3) showed that such treatment seemed hardly to affeet sulcotrione degradation (Fig. 5b). However, the predominance of the biotic pathway over ehemical hydrolysis was les s evident in Ghent soil (Fig. 5a). These results suggest that sulcotrione can be degraded Eollowing both biotie and abiotic ways, depe~ding on soil origino This observation was confirmed at high temperatures (protocol 2, 40 and 60 oC), especial1y in Ghent soil, where sulcotrione appeared more stable at 40 oC (half-life not evaluated) than at 25 oC (45 days) and at 60 oC (32 days). An Arrhenius diagram, plottirtg In Kobs
against 1/ T (K) (Fig. 6), confirms that sulcotrione follows three types oE pathway: a biotic pathway at 25 oC, a predominant abiotic pathway at 40 oC (where microorganisms can be affected) and an abiotic pathway with a possible intervention oE thermophile bacteria at 60 oc.
Furthermore, an available comparison between the degradation oE sulcotrione and mesotrione was possible; using the same experimental conditions as described Eor prorocol l. The disappearance oE mesotrione 14 days aEter treatment was estimared as 65 and 90% oE the initial applied dose, against 16
89
236

Valorisation des travaux de recherche
H Chaabane et al.
Sulcotrione CHD
(a)
o O CI
~:~SO-2C-H-3----~ Sulcotrione
(b) CMBA
Figure 4. Degradation pathways (A and B) of sulco trio ne .
... Autoclaved soil • Unsterilised soil * Autoclaved soil • Unsterilised soil
0.8 0.8
0.6 0.6
• ::j~ • 0.2
O+----~-----,-------.
O 50 100 150 O 50 100 150
(a) Incubation period (days) (b) Incubation period (days)
Figure 5. Degradation rate of sulcotrione under sterilised conditions in (a) Ghent and (b) Perpignan soil (protoGol 3).
o 0.0 29 0.003 0.0031 0.0032 0.0033 0.0034
-1
-2
'" .c
'::t:.0
-3 E
-4 •• A
-5 A"
-6
Figure 6. Arrhenius diagram for degradation of sulcotrione at 25, 40 and 60 oc in Ghent soil.
90
and 33% for sulcotrione, respectively, on Ghent and Perpignan soils (Figs 2a and b). Whatever the soil type, mesotrione (with the highest apparent constant, k = 0.13 day-l) seemed to be more rapidly degraded than sulcotrione. Half-lives for the two soils varied between 5 and 34 days for mesotrione and between 45 and 65 days for sulcotrione (Table 3).
The present results suggested that physicochemical properties of each soil (pH and OC) (see Table 1) infiuenced the degradation process. Owing to their weak acidic nature, the two triketones dissociate from molecular to anionic form as the pH rises. Moreover, the authors reponed in earlier work11 that their anionic form was more resistant to hydrolysis and photolysis processes. However, in soil, the proportion ofionic and
Pest Manag Sci 64:86-93 (2008)
237

Valorisation des travaux de recherche
molecular forms is often difficult ro evaluate exactly owing to the hard-to-foresee difference between the pH value on the soil surface and in the outer bulk soil solution. 7
Under these experimental conditions (25 oC, unsterilised soils and a soil moisture content of 30%), close to ~atural conditions, the relatively high stability of sulcotrione (under its anionic form) in Perpignan soil could be attributed ro the alkaline pH of this soil. For mesotrione, the present experimental results showed that it disappeared more rapidly than sulcotrione in Perpignan soil, whereas on the Ghent soil there was no important dífference between the degradation behaviour of the two compounds (45 and 34 days respectively for sulcotrione and mesotrione). This observation suggested the presence of other factors influencing the degradatíon process. The rapid disappearance of mesotrione in Perpígnan soil could
. be attributed to its possible reduction to the corresponding amino compound owing to the hypothetical presence of specific microflora.
With the exception of the half-life of mesotrione i in Perpignan soil (5 days), all other half-lives were , relatively high (34-65 days) and could be due ro the
presence of nitro and halogen moieties that did not , allow rapid mineralisation of the compounds. 15
3.2 Sorption behaviour of sulcotrione and mesotrione The sorption behaviour of sulcotrione, CHD and CMBA has been reported previously. 6 It was shown that sulcotrione and CMBA are moderately adsorbed whatever the soil type, whereas CHD presents the highest adsorption capacity. Values of adsorption coefficients are reported in Table 4.
As a consequence of identification of degradatíon pathway B, identification of PHD as a new metabolite led the authors to study ¡ts sorption characteristics under the same conditions. Using kinetic and isotherm
, studies, the distribution and sorption coefficients Kct , and K fa were determined for PHD. Kinetic results showed that PHD exhibits an adsorprion behaviour relatively close to that of CHD (Kd varied between
: 0.18 and 5.40 L kg- 1 for CHD and between 2.00 and • 5.00 L kg- I for PHD). This preliminary observation
was confirmed by an isotherm study, with K fa values of 1.6 and 25.0 respectively for Perpignan and Ghent soil. As with CHD, the metabolire PHD seemed ro be more adsorbed than CMBA and sulcorrione (Fig. 7). Perpignan soil had the iowest Kfa values. Even if it is difficult to classify soils using Kfa values when linfa is significantly different from 1, it seemed possible to consider Perpignan soil as having the lowest adsorption capacity. This soil is characterised by a low organic matrer content and a slightly alkaline pH (Table 1) compared with Ghent soil. The acidic character of sulcotrione and its degradation products allow possible repulsion between their anionic forms and negatively charged soil surfaces. This observation has al so be en
Pest Manag Sci 64:86-93 (2008)
Behaviour of sulcotrione and mesotrione in soils
Table 4. Adsorption coefficients of sulcotrione, its degradation
products and mesotrione
Sorptlon coefficlent Ghent soll Perplgnan soll
Sulcotrlone Kfa 1.54 1.30a
1¡nfa 1.02 0.87a ·
Koc 74.0 144.0a
Kfd 0.026 n.d. Total desorptlon (TO%) 91.0 n.d.
CHO Kta 10.0 1.4
1/nfa 0.52 0.64
Koc 492.0 155.0
Ktd n.d. n.d. TO% 2.5 11.0
CMBA Kfa 0.39 0.35 1¡nfa 1.02 0.87 Koc 20.0 39.0
Kfd n.d. n.d. TD% n.d. 4.0
PHO Kfa 25.0 1.6 1¡nfa 0.91 0.85 Koc 1231.0 178.0
Kfd n.d. n.d. TO% 12.0 14.0
Mesotrlone Kfa 1.57 0.14 1¡nfa 0.55 1.43 K~c 78.0 16.0 Kfd 10.08 n.d.
TO% 30.00
a Adsorption coefficient only far low concentrations.
150 -*-sulcotrione --+-CHD ___ CMBA -lf- PHD
30
1501-*-sulcotrione --+-CHD ___ CMBA -lf- PHD
í 100 el el
~ ~ O· 50 I 0~.14~:)(
o 5 10 15 20 25 30
(b) Ce (mg L-1)
Figure 7. Adsorption isotherms of sulcatrione and its degradation products in (a) Ghent and (b) Perpignan soil.
reported for other weak acidic herbicides such as 2,4-D 16 and 1FT. 17
In parallel to sulcotrione and its degradation products, a sorption srudy of mesotrione was conducted under the same conditions. Kinetic results
91
238
-.0;,.., • .-.

Valorisation des travaux de recherche
H Chaabane et al.
150
í 100 el
~ O'" 50
__ Ghent ..... ~_. Perpignan
o -~..--' -,-----,--:_
(a)
I el
~
150
100
O"' 50
o 5 10 20 25 30
__ Ghent ~- Perpignan
O~~~~;!l'i- -.... _ ..... _ .. _"', ... =======-.. ~-+
O 5 10 15 20 25 30
(b)
Figure 8. Adsorption isotherms of (a) sulcotrione and (b) mesotrione on Ghent and Perpignan soils.
showed that Kd for mesotrione ranged from 0.22 to 0.44 L kg- 1• For sulcotrione, no adsorption was observed on Perpignan soil, but on Belgian soil Kd was 0.66 L kg- 1
• This moderate adsorption of the two triketones was verified by isotherm results. As shown in Fig. 8, according to the classification ofGiles et al., 18 adsorption isotherms are L-type, expressing a moderate affinity between soil and herbicide and a decrease in the availability of adsorption sites with increasing liquid concentration. Experimental data were well fitted by the Freundlich model [equation (2), r2 values ranging between 0.89 and 0.91], and a log-log transformation allowed us to estimate Kfa and linfa reported in Table 4. The adsorption coefficient normalised to the organic carbon content of the soil, K oc, is a useful indicator of the binding capacity of a chemical on organic matter of soil and allows a comparison between different compounds. According 10 K fa and Koc values, adsorption capacity was slightly more important on Ghent than on Perpignan soil, where K fa was established only for low concentrations as no adsorption was determined using high concentrations. Adsorption of the two triketones was slightly different, and this was probably due to their molecular structure, particularly the moiety in position 2 of the cyc1ic ring (Fig. 1). The N02 moiety could exercise more steric hindrance than CI to reaching adsorption sites.
As previously reported,6 sulcotrione adsorption in soils increased with c1ay content, while organic matter content and pH played a minor role, and this result seemed to be verified for mesotrione.
92
The Freundlich relationship was tried for al! compounds to evaluate desorption experimental results. For sulcotrione, the Kfd values and the total desorption percentage (70%) showed its relatively high desorption capacity. For the degradation products, the present results did not fit the Freuncilich equation well. However, the total desorption percentage ( < 15 %) (Table 4) showed their lower aptitude for desorption under the present experimental conditions. For mesotrione, the desorption percentage (30%) was lower than for sulcotrione, suggesting that the adsorption process was not totally reversible and led to the presence of a hysteresis phenomenon.
4 CONCLUSION This study, carried out on two soil types with different physicochemical properties, provides supplementary information about sulcotrione behaviour in soils. A comparison with mesotrione could be established as experiments were carried out under the same conditions. The two triketones exhibited moderate adsorption in both soil types tested. However, degradatíon processes depended on soil type and campound properties.
It was proposed to evaluate the potential for reaching water resources accordiilg to the sorption and degradation coefficients. Among the list of mathematical models, the groundwater ubiquity score (GUS)19 is the most simple model giving preliminary but available information about the leaching potential of the two triketones. According to the equatíon
GUS = 10g(tl/2soi') x [4 -log(Koc)] (5)
the GUS values are 3.5, 3.2, 3.3 and l.9 for the respecti~e couples sulcotrione or mesotrione/Ghent and sulcotrione or mesotrione/Perpignan. In both soil types, sulcotrione and mesotrione could be c1assified as substances for which the leaching potential must be addressed (with GUS values higher than 2.8), even if this potential seemed to be limited for mesotrione in Perpignan soil. In order ro refine the leaching patential, these results might be complemented with experimental values obtained using leaching soil column or lysimeter devices. However, a proper assessment of risk for leaching should be performed with data obtained from GLP studies (adsorption, laboratory degradation rate) and with calculations of predicted environmental concentrations under the scenarios developed by the European FOCUS group.20
REFERENCES Wilson JS and Foy CL, Weed control in no-tillage and
conventional corn (Zea mays) with lClA-0051 and SC-0074. Weed Technol4:731-738 (1992).
2 Compagnon JM and Béraud JM, Adaptation de la sulcotrione seule ou assocíée á la résolutíon des problémes régíonaux de désherbage du mais. Journées Internacionales sur la Lurte Contre
Pes, Manag Sci 64:86-93 (2008)
239

Valorisation des travaux de recherche
les Mauvaises Herbes, Reims, France, 6-8 December 1995, pp. 805-812 (1995).
3 Surton P, Richards C, Buren L and Glasgow L, Activity of mesotrione on resistant weeds in maize. Pest Manag Sci 58:981-984 (2002).
4 Rouchaud J, Thirion A, Callens D and Bu1cke R, Soil dissipation of the post-emergence su1cotrione in maize crops treated with organic ferólisers. Bull Environ Contam Toxicol 57:398-405 (1996).
Rouchaud J, Neus O, Cools K and Bu1cke R, Dissipation of the triketone mesotrione herbicide in the soil of corn crops grown on different soil types. Toxicol Environ Chem 77:31-40 (2000).
6 Chaabane H, Cooper ]F, Azouzi L and Coste CM, Influence of soil properties on adsorption-desorption of su1cotrione and its hydrolysis metabolites on various soils. J Ag Food Chem 53:4091-4095 (2005).
7 Dyson JS, Beulke S, Brown CD and Lane MCG, Adsorption and degradation of the weak acid mesotrione in soil and environmental fate implieations. Organic eompounds in the environment. J Environ Qual31:613-618 (2002).
8 Hatzios KK, Herbicide Handbook, Supplement te 7th edition. Weed Science Society of America, pp. 34-36 (1998).
9 Rouehaud J, Neus O, Bu1cke R, Cools K and Eelen H, Suleotrione soil metabolism in summer corn crops. Bull Environ Cantam Toxicol61:669-676 (1998).
10 Baer U and Calvet R, Fate of soil applied herbicides: experimental data and prediction of clissipation kinetics. J Enviran QuaI28:1765-1777 (1999).
11 Chaabane H, Etude de la stabilité et du componement de la suIcotrione dans l'environnement: eaux et sois agricoles. PhD Thesis, Perpignan University, Franee, 154 pp. (2005).
Pest Manag Sci 64:86-93 (2008) DO!: IO.1002/ps
Behaviour of sulcotrione and mesotrione in soils
12 Mathieu C and Pieltain F, Prélevement et préparation des éehantillons de terre, in Analyse Chimique des Sois. Tec and Doe, France; pp. 20-21 (2003).
13 Adsorption-desorption using a bateh equilibrium method. OECD Test Guideline 106 (2000).
14 Cherrier R, Impaet sur I'environnement de deux herbicides du mai"s: la su1cotrione et l'atrazine. Influence du ehangemept d'apports organiques. PhD Thesis, National Polytechnic Institute of Lorraine, Nancy, Franee, 175 pp. (2003).
15 Calvet R, Barriuso E, Bedos C, Benoit P, Charnay MP and Coquet Y, Rétention des pesticides dans le sol. Ch 4, in Les Pesticides dans le· Sol : Canséquences Agranomiques et Environnementales. FranceAgricoles, pp. 161-233 (2005).
16 Rütters H, Hollrigl-Rosta A, Kreuzig R and Bahadir M, Sorption behaviour of prochloraz in different soils. J Ag Food Chem 47:1242-1246 (1999).
17 Bresnahan GA, Koskinen WC, Dexter AG and Luesehen WE, Influence of soil pH-sorption interactions on imazethapyr carryover. J Ag Faod Chem 48: 1929-1934 (2000).
18 Giles CH, McEwan TH, Nakhwa SN and Smith D, Studies in adsorption. Pan XI: asystem of c1assifieation of solution adsorption isotherms and its use in diagnosis of adsorption mechanisms and in measurement of specific surface areas of solido J Chem Soc 3:3973-3993 (1960).
19 Gustafson DI, Groundwater Ubiquity Score: a simple method for assessing pesticide leachability. Enviran Toxical Chem 8:339-357 (1989).
20 SANCO DocumenrJ32112000 rey. 2; FOCUS groundwater scenarios in the EU, Review of active substances, 202 pp. (2000).
93
240

Valorisation des travaux de recherche
241
Communicationscientifique
Actes du 37ème congrès du Groupe Français des Pesticides, Bordeaux (mai 2007)
Transfert de la Sulcotrione, Herbicide du Maïs, sur Mini-colonnes de Sol Reconstitué
Hanène Chaabane, Christophe Calvayrac et Jean-François Cooper
Laboratoire de Chimie des Biomolécules et de l’Environnement, Centre de Phytopharmacie,
Université de Perpignan, 52 avenue Paul Alduy, 66860 Perpignan, France
1- Introduction
Le devenir d’un produit phytopharmaceutique et de ses produits de dégradation, après
application au champ, est fortement conditionné par les processus de dégradation et de
rétention. Ces processus ont été étudiés dans le cas de la sulcotrione [2-(2-chloro-4-
methylsulfonylbenzoyl)-1,3-cyclohexanedione], substance active herbicide de la famille des
tricétones, aussi bien dans l’eau (avec ou sans irradiation) que dans le sol (Chaabane, 2005,
Chaabane et al., 2005, Chaabane et al., 2007a et b). Les mêmes produits de dégradation (1,3-
cyclohexanedione ou CHD, acide 2-chloro-4-méthylsulfonylbenzoïque ou CMBA, dérivés de
l’acétophénone et de l’acide phényl heptanoïque) ont été identifiés dans les deux cas
(Chaabane et al., 2007a et b).
Les processus de dégradation et d’adsorption-désorption participent au transfert horizontal
et/ou vertical de la substance active et des produits de dégradation. Le modèle empirique de
Gustafson (1989), qui met en relation le temps de demi-vie ainsi que les constantes
d’adsorption, nous a permis d’estimer le potentiel de lessivage de la sulcotrione (Chaabane,
2005): cette molécule pourrait être classée parmi les substances présentant un risque de
transfert relativement facile vers les eaux souterraines. Dans la littérature, certains résultats

Valorisation des travaux de recherche
242
corroborent notre estimation (Cherrier, 2003), d’autres indiquent l’absence de transfert de la
sulcotrione au delà de 20 cm de profondeur (Rouchaud et al., 1996 et 1998a et b, Baer et
Calvet, 1999). Cela nous a conduit à étudier le mécanisme de transfert de la sulcotrione à
l’aide d’un dispositif expérimental approprié.
En parallèle, un autre objectif de cette étude était de comparer le potentiel de lessivage de la
sulcotrione à celui de deux autres substances actives souvent détectées dans les ressources en
eau: l’atrazine et la bentazone, dont l’utilisation a été ou reste homologuée en maïsiculture.
2- Matériels et méthodes
2-1 Réactifs chimiques
- Standards analytiques: La sulcotrione (99,2%), l’atrazine [(2-chloro-4-ethylamino-6-
isopropylamino-S-triazine] et la bentazone [3-isopropyl-(1H)-Benzo-2,1,3-thiadiazin-4-(3H)-
one 2,2-dioxide] proviennent de Riedel-de Haën. Leurs propriétés physico-chimiques ainsi
que les doses agronomiques recommandées sont réunies dans le tableau 1. Une solution mère
de chaque substance active a été préparée dans le méthanol à 1000 mg.L-1 et conservée à
l’obscurité à 4°C. Les solutions de traitement ont été préparées à 100 mg.L-1 par dilution des
solutions mères dans l’eau distillée. La CHD (97,0%), le CMBA (95,0%) et la 1-(2-chloro-4-
methylsulfonyl) éthanone (96,6%) proviennent respectivement de Fluka Chemica, Acros
Organics et Bayer CropScience. L’acide 5,7-dicéto-7-(2-chloro-4-méthylsulfonylphényl)
heptanoïque (96,6%) a été synthétisé suivant la méthode publiée par Chaabane et al. (2007b).
- Autres réactifs : Les solvants organiques (Méthanol, acétonitrile sont de qualité
CLHP, le dichlorométhane est de qualité pour analyses de résidus de pesticides) sont fournis
par Carlo Erba, l’acide trifluoroacétique (TFA, 99,0%) de la part d’Aldrich, l’acide
chlorhydrique (HCl, 38%) de Prolabo et l’eau est de qualité milliQ.

Valorisation des travaux de recherche
243
Tableau 1 : Propriétés physico-chimiques de substances actives utilisées.
2-2 Sols
Deux types de sols, nommés Perpignan (sol limono-sablo argileux, pH= 8,1 et M.O= 1,7%)
et Belgique (Sol sablo-limoneux, pH= 6,6 et M.O= 3,5%) ont été sélectionnés, tamisés (<
2mm), séchés à l’air libre et conservés à +4°C jusqu’à leur utilisation.
2-3 Dispositif experimental
Des mini-colonnes de sol reconstitué (diamètre interne 8,0 cm et hauteur 15,0 cm, munies
d’un tamis et d’un entonnoir) ont été placées en plein champ ou sous serre. Dans chaque mini-
colonne a été placée une couche fine de bille en verre de 0,6 mm de diamètre puis 10,0 cm de
chaque type de sol correspondant respectivement à 550 g et 625 g (Perpignan et Belgique) en
tenant compte de la densité de chaque type de sol, respectivement 1,1 et 1,3.
Afin d’étudier l’effet de l’état initial du sol avant traitement, deux états hydriques des sols ont
été choisis correspondant aux sols "naturels" de Belgique et de Perpignan sans humidification
préalable : BN et PN et aux sols saturés BS et PS, préalablement saturés en eau, par un
passage régulier d’eau distillée durant un jour avant traitement.
Les sols (PN, PS, BN et BS) ont été traités au double de la dose recommandée pour la
sulcotrione (600 g.ha-1). Les mini-colonnes ont été placées sous serre et ont reçu un volume
Standard
analytique
Poids moléculaire
g.mol-1
pKa Hydrosolubilité
mg.L-1
Dose agronomique
recommandée g.ha-1
Sulcotrione 328 2,87 165 300-450
Atrazine 215 1,7 33 1100
Bentazone 240 3,3 570 1392

Valorisation des travaux de recherche
244
d’eau de 50 mL, tous les quatre jours environ, afin de se rapprocher des conditions naturelles.
Dans le cas des traitements avec l’atrazine, la bentazone et la sulcotrione, nous avons opté
pour des traitements simulant les doses agronomiques recommandées, soit 550, 696 et 150 µg
de substance active par mini-colonne de sol (tableau 1). L’expérimentation a été menée sous
serre seulement avec le sol de Perpignan initialement saturé en eau avant le traitement.
Parallèlement aux mini-colonnes traitées, deux blancs de chaque type de sol ont été conduits
dans les mêmes conditions pour vérifier l’absence d’interférence au niveau des
chromatogrammes des extraits de sol ou des percolats.
2-4 Méthodologie analytique
Afin de suivre l’évolution de la sulcotrione, de ses métabolites, de l’atrazine et de la
bentazone, les percolats ont été récupérés après chaque apport d’eau puis analysés sans
concentration préalable par Chromatographie Liquide Haute Performance (CLHP).
L’évolution de l’ensemble des molécules à travers le profil du sol a été suivie, aux dates
préfixées (0, 4, 7, 15, 30 et 60 jours après le traitement), en procédant à un fractionnement du
profil en trois horizons (h1=0-3, h2=3-6 et h3=6-10 cm). Chaque horizon a été par la suite
homogénéisé et un échantillon de 10 g a été extrait suivant la méthode précédemment publiée
(Chaabane et al., 2007a) puis analysé par CLHP. Le système chromatographique utilisé est
une chaîne CLHP Shimadzu équipée de deux pompes LC-10ADvp (débit 1mL.min-1, les
modes en gradient d’élution utilisés sont présentés dans le tableau 2), d’une colonne Supelco
ODS Hypersil C18 (5µm, 250 mm x 4,6 mm) et d’un détecteur UV/Visible SPD-10Avp fixé à
254 nm.

Valorisation des travaux de recherche
245
Molécule Phase mobile Gradient d’élution
Sulcotrione et ses
métabolites
Bentazone
A = acétonitrile
B = eau acidifiée par
le TFA à 0,3 %
30% à 50% (A) en 8 min, puis
maintien à 50% (A) pendant 10 min
Atrazine et ses
métabolites
A = acétonitrile
B = eau milliQ
30% (A) / 70% (B) : 1min
30% à 60% (A) en 6 min puis
maintien à 60% (A) pendant 8.50 min.
Tableau 2 : Conditions d’analyse chromatographique.
3- Résultats et discussion
3-1Transfert de la sulcotrione sur mini-colonnes de sol reconstitué, sous serre
3-1-1 Dynamique de lessivage de la sulcotrione : Analyse des percolats
L’évolution des volumes d’eau percolés ainsi que la concentration de la sulcotrione dans les
deux types de sols de Perpignan et de Belgique, quel que soit leur état avant traitement, sont
présentés dans les figures 1a et 1b. Au cumul des volumes d’eau percolés à la fin du suivi,
nous pouvons conclure que quel que soit l’état initial du sol, le sol de Belgique semble
montrer une rétention d’eau légèrement supérieure à celle du sol de Perpignan
(respectivement 339, 98, 415 et 141 mL sur BS, BN, PS et PN, figure 1). Ce comportement
hydrique, avec une différence significative suivant l’état du sol avant traitement, aura une
influence sur la dynamique de lessivage de la sulcotrione et éventuellement de ses
métabolites. En effet, les premiers résidus de sulcotrione ont été détectés dès le 3ème apport
d’eau (150 mL), ce qui correspondrait au volume de la macroporosité occupée par l’eau
mobile du sol saturé.

Valorisation des travaux de recherche
246
a) b)
Fig. 1 : Dynamique de lessivage de la sulcotrione dans les percolats a)- du sol de Perpignan et
b)- du sol de Belgique.
Les concentrations sont nettement supérieures dans les percolats du sol de Perpignan par
rapport à celles enregistrées sur le sol de Belgique (figure 1). Ce résultat laisse penser à un
mouvement de la sulcotrione (moyennement soluble et faiblement retenue sur ce type de sol
(Chaabane et al., 2005) en profondeur avec le flux d’eau à travers les macropores. Ces
concentrations tendent à diminuer malgré un apport d’eau régulier, expliquant soit une
dispersion de la substance active et formation de résidus liés, soit l’intervention d’une
dégradation biotique.
En résumé, les quantités exportées semblent fortement corrélées aux volumes d’eau drainés,
et montrent que malgré des concentrations sensiblement identiques dans le cas du sol de
Belgique, les quantités exportées sont trois fois plus importantes sur le sol saturé (2%) que sur
le sol naturel (0,85%). Ces quantités restent nettement inférieures à celles détectées sur le sol
de Perpignan avec un maximum de 18% sur le sol initialement saturé en eau (PS).

Valorisation des travaux de recherche
247
Concernant les métabolites détectés dans les percolats, nous avons identifié uniquement le
CMBA (précédemment décrit en conditions de laboratoire par Chaabane et al. (2005)) qui a
manifesté le même comportement de lessivage que la molécule mère. Dans les percolats du
sol de Belgique, aucun métabolite n’a été détecté à des teneurs supérieures à la limite de
détection de notre méthode d’analyse.
Au vue des quantités de sulcotrione et de ses métabolites exportées tout au long du suivi, nous
pouvons déduire que la sulcotrione est assez faiblement transférée au delà de 10 cm de
profondeur.
3-1-2 Dynamique de lessivage de la sulcotrione : Analyse des horizons de sol
Les teneurs en résidus de sulcotrione distribués dans les horizons des sols de Belgique et de
Perpignan sont présentées dans les figures 2a et 2b, aussi bien pour les sols naturels
qu’initialement saturés en eau. L’analyse des horizons de sols permet de confirmer les
résultats de l’analyse des percolats (2.1.1). Les concentrations de la sulcotrione détectées dans
le sol de Perpignan sont nettement plus faibles que dans les horizons du sol de Belgique.
a)- Sol de Perpignan b)- Sol de Belgique
Fig. 2 : Dynamique de lessivage de la sulcotrione à travers le profil du sol.
4J8J
32J61J
PNPS
4J8J
32J
BNBS
61J

Valorisation des travaux de recherche
248
Fig. 3: Comparaison des résidus de la sulcotrione par rapport à ceux de l’atrazine et de la
bentazone, dans les mini-colonnes de sol reconstitué.
Les propriétés physico-chimiques de ce type de sol, entraînant une rétention plus importante
de la sulcotrione comparativement au sol de Perpignan, pourraient expliquer ces teneurs
élevées et la persistance surtout sur sol "naturel". Ces résultats montrent : (i) une stabilisation
des résidus dans l’horizon superficiel (h1=0-3cm) jusqu’à 15 jours après le traitement sur le
sol de Belgique (ii) une distribution plus homogène sur le sol de Perpignan entre les horizons
h1 et h2 et (iii) une absence de résidus dans l’horizon h3 expliquant l’absence de sulcotrione
dans les percolats.
L’analyse des horizons de sols a révélé la présence des produits de dégradation identifiés,
notamment le CMBA (précédemment détecté dans les percolats) et la CHD sur le sol de
Perpignan (naturel et saturé) et en plus le dérivé de l’acide phényl heptanoïque sur le sol de
Belgique. Ce métabolite a été particulièrement identifié dans l’horizon superficiel (0-3cm) ce
qui pourrait être expliqué soit par sa forte rétention et donc sa stabilisation en surface ou
encore par son unique formation par voie biotique qui serait absente en profondeur.
3-2 Transfert de la sulcotrione sur mini-colonnes de sol reconstitué, en plein champ

Valorisation des travaux de recherche
249
En parallèle, le dispositif expérimental a été placé en plein champ en conditions naturelles de
température et de précipitations. Trois épisodes pluvieux ont été enregistrés tout au long du
suivi, dont seulement un survenu le 12/02/2005, qui était efficace. Les percolats analysés ont
révélés des quantités de sulcotrione exportées nettement supérieures à celles enregistrées en
conditions sous serre (particulièrement sur sol naturel BN et PN respectivement 5 et 3%).
L’analyse des horizons de sols montre également des teneurs plus importantes sur le sol de
Belgique, quel que soit l’état initial du sol dans les divers horizons jusqu’à 53 jours après
traitement. Ce résultat peut expliquer une certaine activité résiduelle de la sulcotrione dans le
sol, précédemment soulignée par Cools et al. (1997).
Par contre, trois mois après traitement, la sulcotrione n’a été détectée ni dans les percolats ni
dans les horizons de sol, à une profondeur de 10cm.
3-3 Comparaison du potentiel de lessivage de la sulcotrione, de la bentazone et de l’atrazine
Les résultats de l’analyse des percolats révèlent que les pourcentages d’exportation de la
sulcotrione et de l’atrazine (sensiblement identiques, respectivement 2 et 3%) restent
nettement inférieurs à celui de la bentazone (13% de la dose initiale appliquée).
L’évolution des résidus de chaque substance active dans la mini-colonne durant le suivi est
représentée par la figure 3.
Sur le sol de Perpignan (PS), les teneurs en sulcotrione n’excèdent pas 1% jusqu’à la fin du
suivi. Ces faibles teneurs par rapport aux résultats précédents seraient expliquées par des
conditions climatiques, notamment de température, différentes par rapport au premier suivi.
Dans le cas de l’atrazine, les teneurs, importantes au début du suivi, tendent à diminuer quel
que soit l’horizon de sol pour atteindre au total 10% de la dose initiale sur l’ensemble du
suivi. Malgré une dose d’application 5 fois plus importante que celle de la sulcotrione, les
teneurs restent faibles témoignant une dégradation rapide de cette substance active dans ce
type de sol. Un résultat similaire, sur des colonnes de sol non perturbé, a été précédemment
souligné par Cherrier (2003).

Valorisation des travaux de recherche
250
Dans le cas de la bentazone, nous avons enregistré une répartition plus homogène à travers le
profil du sol témoignant un déplacement rapide avec le mouvement de l’eau, lié à sa forte
hydrosolubilité. Au cours du temps, les teneurs en bentazone tendent à diminuer montrant le
mouvement de transfert avec le flux préférentiel et rappelant les fortes concentrations
enregistrées dans les percolats.
4- Conclusion
Les résultats de ce présent travail nous ont permis de modérer les prévisions établies d’après
les calculs moyennant le modèle GUS. L’état du sol sec avant traitement contribue à limiter le
transfert de la sulcotrione et favoriser sa stabilisation dans les horizons superficiels. Par
contre, elle semble facilement lessivée sur sol initialement saturé en eau malgré une
dégradation plus rapide. Les résultats en plein champ restent tributaires de la fréquence, du
volume et des débits des apports d’eau après traitement.
La comparaison du comportement de lessivage de la sulcotrione, de l’atrazine et de la
bentazone nous a permis de conclure que cet herbicide tricétone présenterait un risque de
transfert et de stabilisation limité par rapport à celui des deux autres substances actives.
5- Références
Baer, U. et Calvet, R. Fate of soil applied herbicides: experimental data and prediction of
dissipation kinetics. J. Environ. Qual. 1999, 28, 1765-1777.
Chaabane, H.; Cooper, J.F.; Azouzi, L. et Coste, C.M. Influence of soil properties on
adsorption-desorption of sulcotrione and its hydrolysis metabolites on various soils. J. Agric.
Food Chem. 2005, 53, 4091-4095.
Chaabane, H. Etude de la stabilité et du comportement de la sulcotrione dans l'environnement
: eaux et sols agricoles. Thèse de l'Université de Perpignan, 2005, 154 p.
Chaabane, H.; Vulliet, E.; Cooper, J.F.; Calvayrac, C. et Coste, C.M. Behaviour of sulcotrione
on various soils : comparison with another triketone herbicide. Pest Management Science
2007a, Sous press.

Valorisation des travaux de recherche
251
Chaabane, H.; Vulliet, E.; Joux, F., Lantoine, F.; Conan, P.; Cooper, J.F. et Coste C.M.
Photodegradation of sulcotrione in various aquatic environments and toxicity of its
photoproducts for some micro-organisms. Water Research 2007b, 41, 1781-1789.
Cherrier, R. Impact sur l'environnement de deux herbicides du maïs: la sulcotrione et
l'atrazine Influence du changement d'apports organiques. Thèse de l'INPL-Nancy, 2003, 175
p.
Cools, K.; Bulcke, R. et Callens, D. Response of replacement crops to soil-applied
sulcotrione. Med. Fac. Landbouww. Univ. Gent. 1997, 64, 721-726.
Gustafson, D. I. Groundwater Ubiquity Score : a simple method for assessing pesticide
leachability. Environmental Toxicology and Chemistry 1989, 8, 339-357.
Rouchaud, J.; Thirion, A.; Callens, D. et Bulcke, R. Soil dissipation of the Post-Emergence
Sulcotrione in maize crops treated with organic fertilisers. Bull. Environ. Contam. Toxicol
1996, 57, 398-405.
Rouchaud, J.; Neus, O.; Bulcke, R.; Cools, K. et Eelen,H. Sulcotrione soil metabolism in
summer corn crops. Bull. Environ. Contam. Toxicol 1998a, 61, 669-676.
Rouchaud, J.; Neus, O.; Callens, D. et Bulcke, R. Sulcotrione soil persistence and mobility in
summer maize and winter wheat crops. Weed Research 1998b, 38, 361-371.

Annexes
252
Table des annexes
Annexe 1 : Fiche AGRITOX sulcotrione. Annexe 2 : Chromatogrammes CLHP/UV du milieu de culture (MS) pour l’isolat bactérien 1OP. Annexe 3 : Chromatogrammes CPG/SM du milieu de culture (MS) pour les isolats bactériens 1OP et 1TRANS au bout de 27 jours d’incubation.

Annexes
253
ANNEXE1
AGRITOX - Base de données sur les substances actives
phytopharmaceutiques
FICHE SULCOTRIONE Dernière mise à jour de la fiche d'information le 05/11/2002
1 - IDENTITÉ Substance active : sulcotrione Source de l'information : union européenne Activité biologique principale : herbicide Famille chimique : tricétone Dénominations ISO : sulcotrione NOU : sulcotrione PRO : sulcotrione ANC : chlormesulone CAS : 99105-77-8 Noms chimiques CAS : 1,3-CYCLOHEXANEDIONE, 2-[2-CHLORO-4-(METHYLSULFONYL)BENZOYL]- CAS : 2-[2-chloro-4-(methylsulfonyl)benzoyl]-1,3-cyclohexanedione Formule brute : C14 H13 Cl O5 S IUPAC : 2-(2-chloro-4-mesylbenzoyl)cyclohexane-1,3-dione Poids moléculaire : 328.77 Formule plane :
2 - PROPRIÉTÉS PHYSIQUES ET CHIMIQUES 2.1 ETAT PHYSIQUE
• solide amorphe 2.2 PRESSION DE VAPEUR (TENSION DE VAPEUR)
• 5.39 µPa à 25 °C 2.3 CONSTANTE DE HENRY

Annexes
254
2.4 SOLUBILITÉS DANS L'EAU • 0.165 g/l
2.5 SOLUBILITÉS DANS LES SOLVANTS ORGANIQUES • acetone : 40 g/l • chlorobenzene : 10 g/l • ethanol : <6 g/l • xylene : <6 g/l
2.6 COEFFICIENT DE PARTAGE OCTANOL/EAU •
2.7 VITESSE D'HYDROLYSE (STABILITE) • temps de demi-vie : 100 jour(s) au pH de 7 - Méthode : EPA
2.8 VITESSE DE PHOTOLYSE DANS L'EAU 2.9 RENDEMENT QUANTIQUE de la phototransformation dans l'eau à lambda > 290 nanomètres 2.10 DISSOCIATION DANS l'EAU 3 - TOXICITÉ 3.1 ABSORPTION, DISTRIBUTION, ÉLIMINATION ET MÉTABOLISME 3.2 ABSORPTION DERMALE 3.3 TOXICITÉ AIGUË PAR VOIE ORALE
• DL50 rat : >5000 mg/kg - Sexe : MF - Souche : Sprague Dawley - Véhicule : en solution dans l'huile de mais - Méthode : EPA / OCDE
3.4 TOXICITÉ AIGUË PAR VOIE DERMALE • DL50 lapin : >4000 mg/kg - Sexe : MF - Souche : Stauffland - Méthode :
EPA / OCDE 3.5 TOXICITÉ AIGUË PAR INHALATION
• Toxicité aigue par inhalation, 4 heures, rat - Sexe : MF - Souche : Wistar derive CL50 : >=1.63 mg/l d'air
3.6 IRRITATION CUTANÉE • Test d'irritation cutanée, lapin : non irritant - Méthode : Draize
3.7 IRRITATION OCULAIRE • Test d'irritation oculaire, lapin : moderement irritant - Méthode : Draize
3.8 SENSIBILISATION CUTANÉE • Sensibilisation retardée par voie cutanée, cobaye : sensibilisant
Magnusson et Kligmann 3.9 GÉNOTOXICITÉ 3.10 TOXICITÉ À TERME ET CANCÉROGÉNÈSE
• Toxicité par voie orale et étude de la cancérogenèse, 18 mois , souris - Sexe : M - Souche : CD-1 DSE/CSE = 3000 ppm
• Toxicité par voie orale et étude de la cancérogenèse, 18 mois , souris - Sexe : F - Souche : CD-1 DSE/CSE = 350 ppm
• Toxicité par voie orale, 2 ans , rat - Sexe : M - Souche : CD DSE/CSE = 1 ppm
• Toxicité par voie orale, 2 ans , rat - Sexe : F - Souche : CD DSE/CSE = 20 ppm
3.11 TOXICITÉ SUR LA REPRODUCTION ET LE DÉVELOPPEMENT

Annexes
255
• Reproduction sur deux générations par voie orale, rat DSE parents = 0.05 mg/kg/j DSE descendance = 0.05 mg/kg/j DSE reproduction = 0.5 mg/kg/j toxicite generale
• Tératogenèse par voie orale, rat DSE foetus = 100 mg/kg/j
• Tératogenèse par voie orale, lapin DSE foetus = 300 mg/kg/j
3.12 NEUROTOXICITÉ 3.13 AUTRES ÉTUDES 3.14 DOSE JOURNALIÈRE ACCEPTABLE (DJA)
• DJA = 0.0004mg/kg/j DSE = 0.04 mg/kg p.c./j (2 ans, voie orale, rat ) (Décision de l'Union Européenne )
Observations : la DJA est basée sur une LOAEL mais le facteur de sécurité 100 est suffisant compte tenu de la pente de la courbe dose-réponse 3.15 DOSE DE RÉFÉRENCE AIGUË (DRfA ou ARfD) Observations : non pertinent 3.16 NIVEAU D'EXPOSITION ACCEPTABLE POUR L'OPÉRATEUR (NEAO ou AOEL)
• AOEL = 0.0006 DSE = 0.06 mg/kg p.c./j (Reproduction sur deux générations, rat ) (Décision de l'Union Européenne )
4 - COMPORTEMENT DANS L'ENVIRONNEMENT 4.1 COMPORTEMENT DANS LE SOL 4.1.1 VOIES DE DEGRADATION 4.1.2 VITESSE DE DEGRADATION 4.1.2.1 VITESSE DE DEGRADATION : EN LABORATOIRE, CONDITIONS AEROBIES
• Substance active DT50 (exprimé en jours) : 24 DT90 Temp : 25 °C Sol : limon fin - USA - Atterberry / Iowa - MO : 4.2 % - pH : 5.6
• Substance active DT50 (exprimé en jours) : 15 DT90 Temp : 25 °C Sol : sable - FRA - Toulouse - MO : 1.4 % - pH : 5.2
• Substance active DT50 (exprimé en jours) : 74 DT90 Temp : 25 °C Sol : limon sableux - USA - San Jose - MO : 0.3 % - pH : 7.3
4.1.2.2 VITESSE DE DEGRADATION : EN LABORATOIRE, CONDITIONS ANAEROBIES

Annexes
256
4.1.2.3 VITESSE DE DEGRADATION : AU CHAMP • Substance active
DT50 (exprimé en jours) - Min : 2 - Max : 6 DT90 Sol :
4.1.2.3 VITESSE DE DEGRADATION 4.1.2.4 PHOTODEGRADATION : EN LABORATOIRE, CONDITIONS AEROBIES 4.1.2.5 PHOTODEGRADATION : EN LABORATOIRE, CONDITIONS ANAEROBIES 4.1.2.5 PHOTODEGRADATION 4.1.3 MOBILITE DANS LE SOL : ADSORPTION
• Substance active Koc : 6.6 - Sol : sable - FRA - Toulouse - %MO : 1.4 % - pH : 5.2
• Substance active Koc : 5.3 - Sol : limon sableux - FRA - Velisy - %MO : 6.6 % - pH : 4.9
• Substance active Koc : 8.98 - Sol : limon sableux - USA - San Jose - %MO : 0.3 % - pH : 7.3
• Substance active Koc : 1.82 - Sol : limon fin - USA - Atterberry / Iowa - %MO : 4.2 % - pH : 5.6
• Substance active Koc : 1.08 - Sol : limon argileux - USA - Luling - %MO : 1.8 % - pH : 7.5
• Substance active Koc : 123 - Sol : limon sableux - USA - %MO : 2 %
• Substance active Koc : 160 - Sol : argile - X - %MO : 1.7 %
• Substance active Koc : 17 - Sol : limon - X - %MO : 2.1 %
• Substance active Koc Min : 1.08 - Max : 8.98 Sol :
4.1.4 MOBILITE DANS LE SOL 4.1.4.1 MOBILITE DANS LE SOL : AUTRES ETUDES AU LABORATOIRE 4.1.4.2 MOBILITE DANS LE SOL : AUTRES ETUDES AU CHAMP 4.2 COMPORTEMENT DANS L'EAU 4.2.1 PRODUITS D'HYDROLYSE 4.2.2 VITESSE D'HYDROLYSE
• temps de demi-vie : 100 jour(s) au pH de 7 - Méthode : EPA 4.2.3 PRODUITS DE PHOTOLYSE DANS L'EAU 4.2.4 VITESSE DE PHOTOLYSE DANS L'EAU 4.2.5 BIODEGRADATION FACILE

Annexes
257
4.2.6 VOIES DE DEGRADATION DANS LE SYSTEME EAU-SEDIMENT 4.2.7 VITESSE DE DISSIPATION DANS LE SYSTEME EAU-SEDIMENT 4.2.7.1 VITESSE DE DISSIPATION - CONDITIONS AEROBIES, OBSCURITE 4.2.7.2 VITESSE DE DISSIPATION - CONDITIONS AEROBIES, LUMIERE 4.2.7.3 VITESSE DE DISSIPATION - CONDITIONS ANAEROBIES, OBSCURITE 4.2.7.4 VITESSE DE DISSIPATION - CONDITIONS ANAEROBIES, LUMIERE 4.2.7.5 VITESSE DE DISSIPATION 4.3 COMPORTEMENT DANS L'AIR 4.3.1 PRESSION DE VAPEUR
• 5.39 µPa à 25 °C 4.3.2 CONSTANTE DE HENRY 4.3.3 VITESSE DE DEGRADATION DANS L'AIR
5 - ÉCOTOXICITÉ 5.1 EFFETS SUR LES OISEAUX ET SUR LES AUTRES VERTÉBRÉS TERRESTRES 5.1.1 TOXICITÉ ORALE AIGUË CHEZ LES OISEAUX
• Anas platyrhynchos - DL50 : >1350 mg/kg p.c. - Véhicule : en solution dans l'huile de mais - Méthode : EPA
• Colinus virginianus - DL50 : >2250 mg/kg p.c. - Véhicule : en solution dans l'huile de mais - Méthode : EPA
5.1.2 TOXICITÉ ALIMENTAIRE CHEZ LES OISEAUX • Anas platyrhynchos - CL50 : >5620 mg/kg aliment - Durée d'exposition : 5
jours - Véhicule : en solution dans l'huile de mais - Méthode : EPA • Colinus virginianus - CL50 : >5620 mg/kg aliment - Durée d'exposition : 5
jours - Véhicule : en solution dans l'huile de mais - Méthode : EPA 5.1.3 TOXICITÉ SUR LA REPRODUCTION CHEZ LES OISEAUX 5.1.4 TOXICITÉ ORALE AIGUË CHEZ LES AUTRES VERTÉBRÉS TERRESTRES 5.1.5 TOXICITÉ CHRONIQUE CHEZ LES AUTRES VERTÉBRÉS TERRESTRES 5.2 EFFETS SUR LES ORGANISMES AQUATIQUES 5.2.1 TOXICITÉ AIGUË CHEZ LES POISSONS
• Cyprinus carpio - CL50 : 240 mg/l - Durée d'exposition : 96 heures - Exposition statique
• Oncorhynchus mykiss (ex Salmo gairdneri) - CL50 : 227 mg/l - Durée d'exposition : 96 heures - Exposition statique
5.2.2 TOXICITÉ CHRONIQUE CHEZ LES POISSONS 5.2.3 BIOCONCENTRATION CHEZ LES POISSONS 5.2.4 TOXICITÉ AIGUË CHEZ LES INVERTÉBRÉS AQUATIQUES VIVANT DANS L'EAU OU LE SÉDIMENT
• Daphnia magna - CE50 : >100 mg/l - Durée d'exposition : 24 heures • Daphnia magna - CE50 : >100 mg/l - Durée d'exposition : 48 heures
5.2.5 TOXICITÉ CHRONIQUE CHEZ LES INVERTÉBRÉS AQUATIQUES VIVANT DANS L'EAU OU LE SÉDIMENT 5.2.6 EFFETS SUR LA CROISSANCE DES ALGUES ET SUR LES PLANTES AQUATIQUES
• algue - CEb50 : 3.5 mg/l

Annexes
258
5.2.7 AUTRES ÉTUDES AQUATIQUES - EFFETS SUR MICRO ET MÉSOCOSMES 5.2.8 CONCENTRATION SANS EFFET PREVISIBLE POUR LES ORGANISMES AQUATIQUES (PNEC)
• PNEC = 5.1 µg/L basée sur : étude : toxicité chez la plante aquatique (Lemna gibba) CE50 = 0.051 mg/L FS = 10 décision de AF le 30/03/10
5.3 EFFETS SUR LES ABEILLES 5.3.1 TOXICITÉ AIGUË
• Abeille - DL50 Contact : >200 µg/abeille - DL50 Orale : >50 µg/abeille 5.4 EFFETS SUR LES ARTHROPODES TERRESTRES AUTRES QUE LES ABEILLES 5.5 EFFETS SUR LES VERS DE TERRE ET AUTRES MACRO-ORGANISMES NON CIBLES DU SOL supposés être exposés à un risque 5.5.1 TOXICITÉ AIGUË 5.5.2 EFFETS SUR LA REPRODUCTION 5.6 EFFETS SUR MICRO-ORGANISMES NON CIBLES DU SOL 5.7 EFFETS SUR D'AUTRES ORGANISMES NON CIBLES (FLORE ET FAUNE) supposés être exposés à un risque 5.8 EFFETS SUR LES MÉTHODES BIOLOGIQUES DE TRAITEMENT DES EAUX USÉES 6 - VALEURS REGLEMENTAIRES CLASSEMENT TOXICOLOGIQUE selon les règles de la directive 67/548/CEE Décision de la Commission des Toxiques le 13/01/93 Xn R36 R40 R43 Classe(s) CMR : Substance cancérogène, troisième catégorie Nocif (Xn) R36 irritant pour les yeux R40 effet cancérogène suspecté - preuves insuffisantes R43 peut entraîner une sensibilisation par contact avec la peau
CLASSEMENT TOXICOLOGIQUE / ETIQUETAGE selon les règles du réglement (CE) N°1272/2008 Classement adapté par ANSES selon les règles de traduction du règlement CLP

Annexes
259
Pictogramme(s) de danger :
SGH07 SGH08 Mention avertissement : attention Cancérogénicité, catégorie 2 H351 Susceptible de provoquer le cancer Irritation oculaire, catégorie 2 H319 Provoque une sévère irritation des yeux Sensibilisation cutanée, catégorie 1 H317 Peut provoquer une allergie cutanée

Annexes
260
ANNEXE2Chromatogrammes CLHP/UV du milieu de culture (MS) pour l’isolat
bactérien 1OP
Figure 1 : 0 jour de culture
Figure 2 : 6 jours de culture

Annexes
261
Figure 3 : 15 jours de culture
Figure 4 : 30 jours de culture

Annexes
262
ANNEXE3
Chromatogrammes montrant (a) la présence de CMBA (extraction des fragments m/z, 110, 217, 248 et 250) dans le milieu (MS) supplémenté en sulcotrione, ensemencé par la souche 1OP au bout de 27 jours et (b) sa non détection avec 1TRANS.

263
Table des matières
REMERCIEMENTS 3
SOMMAIRE 6
TABLEDESILLUSTRATIONS 9
INTRODUCTION 18
PREMIEREPARTIE:REVUEBIBLIOGRAPHIQUE 22
Chapitre1Soletdiversitémicrobienne 231‐Contexte 232‐Composantemicrobiennedusol 243‐Diversitémicrobiennedusol 26
Chapitre2Méthodologiesnonmoléculairesappliquéesàl’étudedessouchesbactériennestelluriques 271‐Méthodescultivablestraditionnelles 272‐Méthodescultivablesinnovantes 29
Chapitre3Méthodologiesmoléculairesappliquéesàl’étudedessouchesbactériennestelluriques 311‐AnalysedesprofilsderestrictiondesADNr16Samplifiés(ARDRA) 312‐AmplificationPCRdesséquencesrépétitives(REP‐PCR) 333‐SéquençagedesADNr16SetAnalysephylogénétique 353‐1‐Principegénéral 353‐2‐Méthoded’alignement 363‐3‐Reconstructiondesarbresphylogénétiques 373‐4‐Estimationdelarobustessedesarbres 39
Chapitre4Dégradationbiologiquedesproduitsphytosanitairesdanslessols 411‐Dégradationbiologiqueetrécalcitrancemoléculaire 412‐Inductionsenzymatiquesetadaptionsmicrobiennes 423‐Aspectscinétiquesdeladégradationbiologique:Métabolismedirectetco‐métabolisme. 454‐Casparticulierdedégradationbiologique:labiodégradationaccélérée(BDA)despesticides 474‐1‐Contexteetdéfinition 474‐2‐Mécanismesresponsablesdelabiodégradationaccélérée 48
DEUXIEMEPARTIE:PRESENTATIONGENERALEDEL’ETUDE 51
Chapitre1Lafamilledesherbicidestricétones 521‐Contextehistorique 522‐Lasubstanceactive 542‐1‐Fiched’identité 542‐2‐Propriétésphysico‐chimiques 542‐3‐Lesformestautomériquesdelasulcotrione 55
3‐Moded’actiondelasubstanceactive 564‐Approchestoxicologiqueetécotoxicologique 604‐1‐Vis‐à‐visdesmammifères 60

264
4‐2‐Ecotoxicologie 605‐Comportementdanslesdifférentscompartimentsdel’environnement 615‐1‐Danslamatriceair 615‐2‐Danslamatriceeau 615‐3‐Danslamatricesol 645‐3‐1‐Rétentionettransfertdelasulcotrioneetdesesproduitsdedégradation 645‐3‐2‐Dégradationdelasulcotrione 64
Chapitre2Matriced’étude:lesoldePerpignan 671‐Situationgéographiquedelaparcelle 672‐ParamètresédaphiquesdusoldePerpignan 683‐Historiquecultural 694‐Stratégied’échantillonnage 695‐Dispositifexpérimental:lesmicrocosmes 70
TROISIEMEPARTIE:METHODOLOGIESDETRAVAIL 73
Chapitre1Méthodologiedel’analysechimique 741‐Analysedeséchantillonsdesols 741‐1‐RéactifsetMatériels 741‐1‐1‐Standardanalytique 741‐1‐2‐Autresréactifs 741‐1‐3‐Matérielclassiquedelaboratoire 741‐1‐4‐Appareillagechromatographiqueetconditionsd’analyses 75
1‐2‐Protocoleopératoire 761‐2‐1‐Préparationdeséchantillons 761‐2‐2‐Analysechromatographique 76
1‐3‐Performancesdelaméthoded’analyseetvalidation 772‐Analysedeséchantillonsdemilieuxdeculture 772‐1‐Réactifsetmatériels 772‐1‐1‐Standardsanalytiques 772‐1‐2‐Autresréactifs 782‐1‐3‐Matérielsclassiquesdelaboratoire 782‐1‐4‐SpectrophotomètreUV/visible 782‐1‐5‐Appareillageschromatographiquesetconditionsd’analyses 782‐1‐5‐1‐CLHP/UV 792‐1‐5‐2‐CLHP/SM 802‐1‐5‐3‐CPG/SM 81
2‐2‐Protocoleopératoire 832‐2‐1‐Péparationdeséchantillons 832‐2‐2‐Analysechromatographique 84
2‐3‐Performancesdelaméthoded’analyseetvalidation 842‐3‐1‐CLHP/UV 842‐3‐2‐CLHP/SM 852‐3‐4‐CPG/SM 86
Chapitre2Méthodologiedel’analysebiologique 871‐Cadredel’étude:organisationdestravaux 872‐Approchecultivable:essaid’isolementdesouchesbactériennesdégradantlasulcotrione 872‐1‐Principeetstratégiedetravail 872‐2‐Milieuxdecultureetmatérielsutilisés 892‐2‐1‐Préparationdesmilieuxdeculture 892‐2‐1‐1‐Milieucomplexe«sol»ounutrientsoilmedium(NS) 892‐2‐1‐2‐Milieuminimum(MS) 902‐2‐1‐3‐Milieucomplexetrypto‐caséinedesojaouTripticsoy(TS) 92
2‐2‐2‐Matérielsutilisésdanslecadredel’approchecultivable 932‐3‐Protocoleopératoire 932‐3‐1‐Isolementdesouchesbactériennessulcotrione–résistantes 93

265
2‐3‐2‐Isolementdesouchesbactériennesdégradantlasulcotrione 952‐3‐2‐1‐Culturesliquidesrépétéesdesisolatsbactériensavecpressiondesélectiondel’herbicide 952‐3‐2‐2‐Estimationdelacapacitédedégradationdesisolatsbactériensvis‐à‐visdelasulcotrione 96
2‐3‐3‐Caractérisationmorphologiqueetbiochimiquedesisolatsdégradants 983‐Approchemoléculaire:caractérisationdesisolatsbactériensdégradantlasulcotrionevsisolatsnondégradants 993‐1‐Principeetstratégiedetravail 993‐2‐Protocoleopératoire 993‐2‐1‐Extractionetpurificationdel’ADNgénomique 993‐2‐2‐Quantificationetcontrôledelaqualitédel’ADNgénomique 1003‐2‐3‐Caractérisationphylogénétiquedesisolatsbactériens 1013‐2‐3‐1‐AnalysedesprofilsderestrictiondesADNr16S(ARDRA) 1013‐2‐3‐2‐AnalysedesisolatsbactériensparREP‐PCR 1033‐2‐3‐3‐SéquençagedesADNr16S 1053‐2‐3‐3‐1‐LigationdesproduitsPCRdel’ADNr16Sdansunvecteurdeclonage 1053‐2‐3‐3‐2‐Transformationdescellulescompétentesaveclevecteurdeclonage 1063‐2‐3‐3‐3‐TestPCRsurcoloniesdesclonesrecombinants 1083‐2‐3‐3‐4‐Préparationdel’ADNplasmidique(MiniPrep) 1093‐2‐3‐3‐5‐Réactiondeséquençage 1093‐2‐3‐3‐6‐Traitementetanalysedesséquences 111
3‐2‐4‐Caractérisationfonctionnelledesisolatsbactériens: 1123‐2‐4‐1‐Contexte 1123‐2‐4‐2‐DétectionparPCRdesgènesXylEcodantl’enzymecatéchol2,3‐dioxygénase 1143‐2‐4‐3‐DétectionparPCRdesgènesC12Ocodantl’enzymecatéchol1,2‐dioxygénase 1163‐2‐4‐4‐SéquençagedesproduitsPCR 1173‐2‐4‐5‐Traitementetanalysedesséquences 117
3‐2‐5‐Caractérisationphysiologiquedesisolats 118
QUATRIEMEPARTIE:RESULTATSETDISCUSSION 119
Chapitre1ConfirmationdupouvoirbiodégradantdusoldePerpignanetrecherched’uneéventuellebiodégradationaccélérée(BDA) 1201‐RéactivitébiologiquedusoldePerpignan 1202‐Recherched’uneéventuellebiodégradationaccélérée(BDA) 1243‐Conclusion 127
Chapitre2Isolementdesouchesbactériennespotentiellementdégradantesdelasulcotrione 1291‐Recherchedesouchessulcotrione‐résistantes 1292‐Isolementdesouchesbactériennesmétabolisantlasulcotrione 1303‐Etudedupotentieldedégradationdesisolatsdégradants:suivideladissipationdelasulcotrionedanslemilieudeculture(MS) 1344‐Recherchedemétabolitesdelasulcotrionedanslemilieudeculture(MS) 137
Chapitre3Caractérisationphylogénétiquedesisolats1OPet1TRANS 1421‐Vérificationdelaqualitédel’ADNmatrice 1422‐AnalysedupolymorphismedeséquencedesADNr16Samplifiés(ARDRA)desisolats 1423‐Analysedesisolatsbactériensparamplificationdesséquencesrépétitives(REP‐PCR) 1444‐Séquençagedel’ADNr16Sdesisolats1OPet1TRANS 1455‐Conclusion 154
Chapitre4Caractérisationfonctionnelledesisolats1OPet1TRANS 1591‐Préambule 1592‐DétectiondesgènesXylEcodantl’enzymecatéchol2,3‐dioxygénase 1603‐DétectiondesgènesC12Ocodantl’enzymecatéchol1,2‐dioxygénase 1644‐SéquençagedesampliconsXylE 1655‐SéquençagedesampliconsC12O 165

266
CONCLUSIONGENERALEETPERSPECTIVES 167
REFERENCESBIBLIOGRAPHIQUES 173
VALORISATIONDESTRAVAUXDERECHERCHE 207
Publicationn°1(SOUSPRESSE) 208
Publicationn°2 233
Communicationscientifique 241
TABLEDESANNEXES 252
ANNEXE1 253
ANNEXE2 260
ANNEXE3 262
TABLEDESMATIERES 263

DEGRADATION BIOLOGIQUE DE LA SULCOTRIONE DANS UN SOL AGRICOLE : RECHERCHE D’UNE EVENTUELLE BIODEGRADATION
ACCELEREE ET CARACTERISATION DE SOUCHES BACTERIENNES POTENTIELLEMENT DEGRADANTES.
RESUME La dégradation des produits phytopharmaceutiques dans le sol conditionne fortement leur devenir et participe à la réduction des risques de pollution diffuse dans les divers milieux aquatiques, et notamment dans les eaux souterraines. Notre travail avait donc pour objectif d’enrichir les connaissances sur la dégradation biologique dans un sol agricole de la sulcotrione, herbicide largement utilisé dans les itinéraires techniques maïsicoles. Le potentiel dégradant du sol de Perpignan a été mis en évidence en conditions de microcosmes. En revanche, nos résultats ont montré l’absence de phénomène de biodégradation accélérée (BDA), malgré la répétition des traitements. La deuxième partie de l’étude a consisté à isoler et caractériser des microorganismes telluriques intervenant au niveau de la réactivité dégradante de ce sol. Après isolement sur milieux de culture synthétiques de huit souches bactériennes résistantes à la sulcotrione, une seule s’est avérée présenter un potentiel dégradant important, en utilisant l’herbicide comme source de carbone et/ou d’énergie. Après séquençage de l’ADNr 16S, cette souche a été identifiée comme appartenant au genre Pseudomonas espèce putida. Notre étude a permis de localiser le site d’attaque enzymatique au niveau de la partie 1,3-cyclohexanedione de la molécule de sulcotrione. Mots clés : sulcotrione, sol, biodégradation, biodégradation accélérée, Pseudomonas putida, séquençage ADNr 16S.
BIOLOGICAL DEGRADATION OF SULCOTRIONE IN AN AGRICULTURAL SOIL : RESEARCH OF A POSSIBLE ENHANCED
BIODEGRADATION PROCESS AND CHARACTERISATION OF DEGRADING BACTERIAL ISOLATES.
SUMMARY Degradation of pesticides in soil plays a large part in the limitation of risks towards aquifers. This study investigates the biological response of an agricultural soil of south of France after several treatments with sulcotrione, a maize herbicide belonging the triketones group. Although reactivity of the Perpignan soil was clearly confirmed, no enhanced biodegradation was observed after repeated herbicide treatments, under microcosms conditions. The second part of our work consisted on isolating and characterising bacterial strains showing a notable capability to intervene in this biodegradation process. Eight sulcotrione-resisting strains were isolated and tested in liquid culture with sulcotrione as sole carbone and/or energy source. Only one was able to efficiently degrade the herbicide. Partial 16S rDNA sequencing allows identification of this strain as Pseudomonas putida. The enzymatic catalysis occurred at the level of the 1,3-cyclohexanedione moiety of the sulcotrione molecule. Key words: sulcotrione, soil, biodegradation, enhanced biodegradation, Pseudomonas putida, 16S rDNA sequencing