the survival of propagules of macrophomina phaseoli and diplodia maydis in nigerian soils
TRANSCRIPT

J. Basic Microbiol. 27 (1987) 2, 67 - 73
(Department of Agricultural Biology, University of Ibadan, Ibadan, Nigeria' and International Institute of Tropical Agriculture, Ibadan, Nigeria*)
The survival of propagules of Mucrophorninu phuseoli and Diplodia rnuydis in Nigerian soils
J. A. DUROJAIYE~,~), T. IKOTUN~) and J. M. FAJEMISIN~)
(Received August 22, 1986)
Weekly isolations from sterile and unsterile soils artificially inoculated with Macrophomina phaseoli and Diplodia maydis showed a decline in numbers of the propagules of these pathogens. Sclerotia of M. phaseoli end spores of D. maydis could not be recovered either on dilution plate or on soil plates after four weeks of inoculation of unsterile soil and after fifteen weeks for D. maydis and twenty-five weeks for M . phaseoli in sterile soil. I n unsterile soil, sclerotia-like struc- tures were observed with the aid of a light microscope up to the tenth week after soil inoculation by the flotation technique but their viability could not be determined due to active parasitization by Pemicillium sp. and Trichoderma sp. The high number of these hyperparasiteslg of soil may be responsible for the non-detection of these pathogens in soils of some maize-growing states in Nigeria, as well as a reduction in their longevity in soil. It thus seems, from this study, that the host tissue is the site of survival of propagules of these pathogens from one season to another.
Maize (Zea mays) is perhaps one of the most valuable cereal crop in Nigeria today. Not only is it eaten in various forms, it is also used as an important component of animal feeds (OYENUGA 1968). The effect of its scarcity on poultry production in recent years in Nigeria, underscores the importance of the crop. Diseases, however, constitute a major constraint t o increased production and yield of this crop the world over. Some of these diseases include the stripe and streak caused by viruses (WELLMAN 1934), wilt and blight caused by bacteria (ELLIOTT 1941), blight, rust, smut, kernel mould, root rot, ear rot and stalk rot caused by fungi (OYEKAN 1977). Among the pathogens causing some of these diseases are Macrophomina phaseoli (MAUBL) ASHBY and Diplodia maydis (BERK) SACO. M.phaseoli is known to cause charcoal rot of many valuable crops and is soil-borne (OYEKAN and OGUNREMI 1978). It causes stalk rot of maize and had been associated with other pathogens in causing ear rot (FRANCIS and BURGESS 1975).
Diplodia maydis caused stalk rot, dry or white ear rot, root rot and seedling blight or maize (ANDREW 1954). These diseases have been reported in Nigeria (NWICWE 1974). Virtually all Nigerian varieties of maize are susceptible t o these two pathogens (FAJEMISIN 1981). The overall effect of stalk rot is that maize plants lodge and the stem breaks. Infected ears become discoloured and unmarketable.
Elsewhere in the world, losses due to stalk and ear rot caused by these two patho- gens range from 4-10%. In Nigeria, losses are still very low, but could increase with time and with increased maize production. The fact that M . phaseoli has a host range of over 300 plants indicated that it may be a difficult pathogen to control once it is established. However, in Nigeria, the survival of these pathogens in soil has not been studied. It is very important t o study this aspect of the pathogens since they are soil-borne, in order t o be able to formulate an effective method of control. This paper reports the investigation on the survival of these pathogens in soil and in host tis- sues. 6'

68 J. A. DUROJAIYE, T. IKOTUN and J. M. FAJEMISIN
Materials and mothods
Soils were collected from several maize farms mainly in the University of Ibadan and I.I.T.A. campuses. The soil samples were collected from the top 15 cm of soil and were screened for the number of propagules of M . phaseoli and D. maydis they contained. The soils were well mixed and placed in 20 cm-diameter plastic pots. Each pot was then inoculated with M . phaseoli and D. maydis inoculum prepared as follows:
Isolates of M . phaseoli were obtained from rotted stalks of maize plants collected from the field and those of D. maydis were obtained from diseased maize grains. The stalks and grains were surface-sterilized in a 1 : 9 solution of clorox for 2 min, rinsed in five changes of sterile distilled water in watch glasses and the pieces were then plated on potato dextrose agar (PDA) in petri dishes. Plates were incubated a t 25 "C in alternating light and darkness regime for 2 weeks and subcultures were made after pure cultures were established. Pure cultures were allowed to grow for three weeks for the maturation of spores and sclerotia.
The mycelial mats and agar from the pure cultures were slowly blended in a blender to separate the single sclerotia of M . phaseoli and spores of D. mnydis from the pycnidia. The number of spores and sclerotia present in the suspension was estimated using a SPENCER haemacytometer slide with an improved NEUBAUER linear scale under a light microscope. For D. maydis, each pot containing unsterile soil was inoculated with 500 ml of inoculum containing about 2.9 x lo4 spores per ml of suspension. For M . phaseoli, each pot containing unsterile soil was inoculated with 500 ml of inoculum containing 3.9 x lo2 sclerotia/ml of suspension. The pots were placed in the greenhouse and records of the relative humidity and temperature were taken using a thermo- hygrograph. For each pathogen, the experiment was replicated thrice with one sterile inoculated (with both pathogen) and uninoculated pots serving as the controls. The soils were sterilized a t 1.5 kg/cm2 for 1 hour everyday for three consecutive days. Samples of 1 g of soil were taken in triplicates from each of the pots at weekly intervals t o monitor the number of spores and sclerotia still surviving in the soil using the dilution plate method (PAHARIA and KOMMEDAHL 1956), soil plate method (WARCUP 1955), and for M . phaseoli, the flotation technique (WEINER and LUTTREL 1945), was also used in addition to the earlier two methods.
As a parallel experiment, debris of plants infected by both pathogens separately was buried in sterilized and unsterilized soils in 20 cm-diameter pots and a t weekly intervals the soils were sampled for the presence of the pathogens using the methods cited above, except that 1 g wt of plant debris had t o be slowly blended t o homogenize the suspension before serial dilution and plating took place.
I n surveying some Nigerian soils for the presence and survival of sclerotia of M . phaseoli and spores of D. maydis, soil samples were collected from some maize growing states such as Bendel, Kwara, Kaduna, Niger, Ogun, Ondo and Oyo. Soil samples were collected in triplicate from the top 15 cm of soil of each area where maize was cultivated. From these samples, sclerotia and spores of both pathogens were screened for their presence following the methods enumerated above.
Results
In unsterile soils inoculated with the propagules of M . phaseoli and D. maydis, there was a general decline of numbers after inoculation. No propagules of these pathogens were isolated from sterile and unsterile soils not inoculated with propagules of the pathogens.
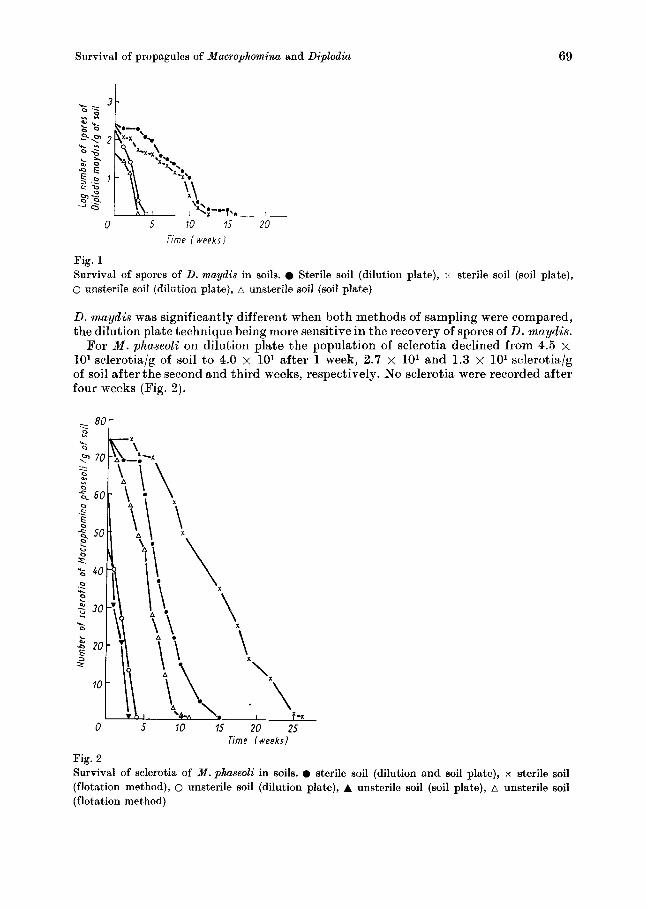
For D. maydis, using the dilution plate method, there was a decline in the number of spores present in soil from 2.2 x 102 spores/g of soil initially to 8.0 x lo1 spores/g of soil after one week of inoculation. By the second and third weeks, the number of spores had declined further to 4.0 x 101 spores/g of soil and to 0.4 x lo1 spores/g of soil, respectively. No spore was recorded on dilution plate four weeks after inocula- tion (Fig. 1). A similar result was obtained with the soil plate method. Initially only 6.7 x lo1 spores/g of soil was recovered from soil and this declined t o 4.2 x lo1 and 1.9 x lo1 spores/g of soil after one and two weeks, respectively. No spore was recovered three weeks after soil inoculation. Using a split plot with complete randomized block design for the analysis of data, the rate of decline in the number of spores/g of soil of
Survival of propagules of Macrophomina and Diplodiu 69
0 5 I0 15 Time ( w e e k s }
..I 20
Fig. 1 Survival of spores of D. maydis in soils. Sterile soil (dilution plate), x sterile soil (soil plate), 0 unsterile soil (dilution plate), A unsterile soil (Boil plate)
D. rnaydis was significantly different when both methods of sampling were compared, the dilution plate technique being more sensitive in the recovery of spores of D. maydis.
For M . phaseoli on dilution plate the population of sclerotia declined from 4.5 x 10’ sclerotia/g of soil to 4.0 x lo1 after 1 week, 2.7 x lo1 and 1.3 x lo1 sclerotia/g of soil after the second and third weeks, respectively. No sclerotia were recorded after four weeks (Fig. 2 ) .
Time /weekr]-
Fig. 2 Survival of sclerotia of M.phaseoZi in soils. 0 sterile soil (dilution and soil plate), x sterile soil (flotation method), 0 unsterile soil (dilution plate), A unsterile soil (soil plate), A unsterile soil (flotation method)

70 J. A. DUROJAIYE, T. IKOTUN and J. M. FAJEMISIN
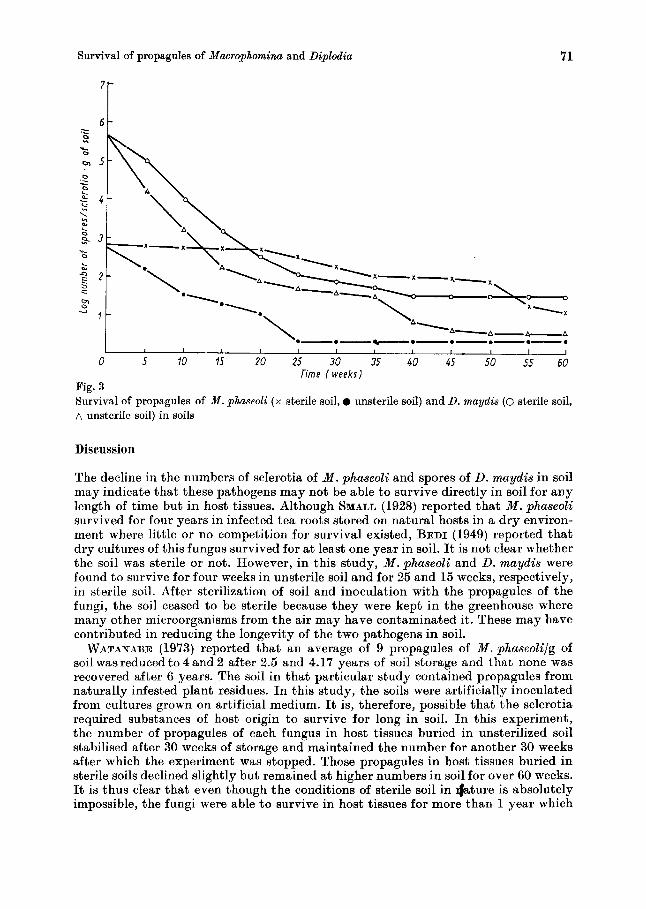
The result of the soil plate method showed a similar trend. Initially 5.9 x lo1 sclero- tia/g of soil was estimated and these declined to 3.0 x lo1 and 2.1 x 10' sclerotia/g of soil after the first and second weeks of inoculation, respectively. No sclerotia were recorded after the third week (Fig. 2). Using the flotation technique, higher numbers of sclerotia (7.4 x 101 sclerotia/g of soil) were recovered initially and while the decline in numbers followed the same trend as in other methods, the numbers of sclerotia recoveredlg of soil were higher than those recovered from same samples by the other two methods. The sclerotia survived up t o seven weeks (2.2 x 10' sclerotia/g of soil) b u t none was isolated after ten weeks.
Also, using the split plot with complete randomized block design, there was no statistically significant difference between the methods of sampling of M . phaseoli and there was no interaction between the times of sampling. The decline of sclerotia was not significant at 1% level but was significant a t 5 % level.
From the sterilized soils, however, the rate of decline of both pathogens was slower and they lasted longer in such treated soil. Fig. 1 shows that from the 2.2 x lo2 spores/g of soil initially isolated for D. maydis there was a slight decrease to 2.0 x lo2 spores/g of soil after one weekin sterile soil. Thereafter, the rate of decline was slow but the spores were not isolated by both isolating techniques after fourteen weeks.
Fig. 2 also shows that from sterile soil a higher number of sclerotia of M . phaseoli were isolated and for a longer time. Thus, from the initial 7.4 x lo1 sclerotia/g of soil using the flotation technique, there was no change in sclerotial population for the first three weeks. In the fourth week the was a slight reduction to 7.0 x 10' sclero- tia/g of soil. Population reduction continued gradually until after twenty-five weeks when sclerotia could not be isolated anymore.
With the soil plate technique, the story was a little different while the rate of decline of population of sclerotia/g of soil was slower, they did not last as long as those isolated by the flotation technique. They were last detected 15 weeks after inoculation into the sterile soil.Using the dilution plate technique the trend of sclerotial population was similar to that of soil plate technique.
Fig. 3 shows that the population of spores of D. maydis and sclerotia of M . phaseoli survived longer in buried host tissues than when naked spores were buried in soil, and for longer in host tissues buried in sterile soil than in unsterilized soil. Although there was some gradual decline in the population of spores and sclerotia. There were more propagules of D. maydislg of host tissues than of M . phaseoli. Out of about 6.0 x 106 spores of D. maydis in host tissues buried in sterile soil about 1.5 x 101 spores/g of host survived for 60 weeks while out of about 8.0 x lo2 sclerotia/g of host tissue only about 1.0 x lo1 survived until the 60th week. In unsterilized soil, the population of spores of D. maydis declined immediately reaching a trough in 35 weeks and thereafter the population declined further t o 0.5 x lo1 spores/g of host tissue. With the sclerotia of M . phaseoli in unsterilized soil, there was an im- mediate gradual decline in population/g of host tissue until it reached a trough twenty weeks after burial. Thereafter the population stablized a t about 0.3 x 101 sclerotia/g of host tissue.
While spores of D. maydis were not recovered from soils brought in from Oyo, Ondo, Ogun, Bendel, Kwara, Kaduna and Nigeria States using the dilution and soil plate met hods, sclerotia-like structures of varying numbers were obtained from all the soil samples by the flotation technique, but a test of their viability was made impossible by the presence of a Penicillium species and TTichoderma sp. which were found to be actively parasitizing the sclerotia. These same fungi were also isolated in unsterilized soils used in the experiments throughout this study.
Survival of propagules of Macrophomina and Diplodia
6 - - .-
71
Discussion
The decline in the numbers of sclerotia of M . phaseoli and spores of D. rnaydis in soil may indicate that these pathogens may not be able to survive directly in soil for any length of time but in host tissues. Although SMALL (1928) reported that M. phaseoli survived for four years in infected tea roots stored on natural hosts in a dry environ- ment where little or no competition for survival existed, BEDI (1949) reported that dry cultures of this fungus survived for at least one year in soil. It is not clear whether the soil was sterile or not. However, in this study, M . phaseoli and D. maydis were found to survive for four weeks in unsterile soil and for 25 and 15 weeks, respectively, in sterile soil. After sterilization of soil and inoculation with the propagules of the fungi, the soil ceased to be sterile because they were kept in the greenhouse where many other microorganisms from the air may have contaminated it. These may have contributed in reducing the longevity of the two pathogens in soil.
WATANABE (1973) reported that an average of 9 propagules of M.phaseoli/g of soil was reduced to 4 and 2 after 2.5 and 4.17 years of soil storage and that none was recovered after 6 years. The soil in that particular study contained propagules from naturally infested plant residues. I n this study, the soils were artificially inoculated from cultures grown on artificial medium. It is, therefore, possible that the sclerotia required substances of host origin to survive for long in soil. I n this experiment, the number of propagules of each fungus in host tissues buried in unsterilized soil stabilised after 30 weeks of storage and maintained the number for another 30 weeks after which the experiment was stopped. Those propegules in host tissues buried in sterile soils declined slightly but remained a t higher numbers in soil for over 60 weeks. It is thus clear that even though the conditions of sterile soil in *ture is absolutely impossible, the fungi were able to survive in host tissues for more than 1 year which

72 J. A. DUROJAIYE, T. IKOTUN and J. M. FAJEXISIN
spans two growing seasons in the Southern part of Nigeria and one growing season in the Northern parts. Thus the infected plant debris and other alternate hosts may be the source of carryover of inoculum from season to season. M . phaseoli is known to have over 300 host plant species.
NORTON (1953) observed that sclerotia or mycelia were not formed to any extent in unsterilized soil away from a food base and added that sclerotia were formed on the organic substrate colonized by the fungus. Thus sclerotia formed on the organic residue or on the host plants during the parasitic phase of the fungus were released into the soil upon the decay of such substrate, and this may increase its longevity in soil. NORTON, therefore, concluded that plant debris was the principal means of survival of the fungus subterraneously. Thus if materials of the host origin are needed for the germination of sclerotia of M . phaseolina, this may lead to a delay in germination on agar plates. This is likely to affect the isolation of the fungus from soil when a non- selective medium is used. If COOK et al. (1973) report is to be followed, then our results indicate about 5 % recovery of sclerotia after inoculation into soil using a non-selective medium such as PDA.
CHRISTENSEN and WILCOXSON (1966) reported that D. maydis survived on plant residues left over from earlier crops from which i t forms dark brown to black pycnidia which clusters near nodes in the subepidermal region of infected maize plants and that this may increase their longevity in soil.
If the pathogens were present in soil, they would have invaded plant tissues during the sampling period which was the normal maize-growing season in the sampling areas. This could have been responsible for the absence of these fungal propagules in sampled Nigerian soils. AGNIHOTHRUDU (1953) was unable to obtain propagules of iW. phaseoli on soil dilution plates but detected the fungus on roots of most stages of pigeon pea (Cajanus cajan L.). Thus the pathogen may probably be found host plants rather than in soil.
Moreover, in the same soil samples, microorganisms such as Trichoderma longi- brachiatum, Penicillium cyclopium and Aspergillus niger were isolated. I n separate study, these microorganisms were found to be antagonistic on plate cultures to both pathogens. This result is similar to that of PAPAVISAS and KLAG (1975) who reported that M . phaseoli was a poor competitor on plates. Even a few colonies of saprophytic fungi growing next to M . phaseoli inhibited or reduced its growth and production of sclerotia needed for identifying the fungus in soil. VASUDERA and SIKKA (1941) reported that Trichoderma lignorum and A. niger directly parasitized hyphae of M . phaseoli, coiling around it and resulting in the coagulation of the protoplasmic content of the hyphal cells followed by cell wall degradation and dissolution. The survival of both pathogens in soil could have been affected by the antagonists isolated and may have affected our results. Other antagonists of M . phaseoli isolated by other workers from soil include Arachiotus sp. and Aspergillus aculeatus (DHINGRA and KHARE 1973). These may also have affected D. maydis.
It is recommended from our results that after harvest, corn stubble be uprooted and burnt. Also it is recommended that the flotation technique is best for isolating sclerotia of M . phaseolifrom soil while the dilution plate method is better for (isolating D . maydis.
Acknowledgemets
The authors are grateful to the International Institute of Tropical Agriculture (IITA) for providing travelling facilities f4 collecting soil samples and for granting working space for this study and to the Director-General for giving permission for this work to be published.

Survival of propagules of Macrophomina and Diplodia 73
Re forenees
AGNIROTRHUDU, V., 1953. Soil conditions and root diseases. VII I . Rhizosphere microflora and some of the important plants of South India. Proc. Indian Acad. Sci. Section B, 37, 1-13.
ANDREW, R. H., 1954. Breeding for stalk rot resistance in maize: A review. Euphytica, 3, 43-48. BEDI, K. S., 1949. Infectivity and longevity of dried cultures of Ascochyta rabiei (PASS) LAB.,
CHRISTENSEN, J. J. and WILCOXSON, R. D., 1966. Stalk rot of corn. Monograph 3. Pub]. Anier.
COOK, G. E., BOOSALIS, M. G., DUNKLE, L. D. and ODVODY, G. N., 1973. Survival of Macrophomina
DHINGRA, 0. D. and KHARE. M. N.. 1973. Biological control of Rhizoctonia bataticola on urid beans.
the causal fungus of gram blight. Indian Phytopathol., 2, 6-10.
Phytopath. Soc.
phaseoli in corn and sorghum stalk residue. PI. Dis. Reptr., 67, 873-875.
Phytopathol. Z., 76, 23-'29. '
- ELLIOT, C.. 1941. Bacterial wilt of corn. U. S. D. A. Farmers Bulletin 1878. FAJEMISIN, J. M., 1981. I. I. T. A. Annual Report - Grain Legume Improvement Programme. FRANCIS, R. G. and BURGESS, L. W., 1975. Survey of Fusarium and other fungi apsociated nith
stalk of maize in Eastern Australia. Australian J. Agric. Res., 26, 801-807. NORTON, D. C., 1953. Linear growth of Sclerotium bataticola through soil. Phytopathology, 43,
633-636. NWIGWE, C., 1874. Effects of Diplodia zeae and Botryodiplodia phomopsis on germination of seeds
of Maize (Zea mays L.). P1. Dis. Reptro., 68, 414-415. OYEKAN, P. O., 1977. Effect of planting data on the incidence and severity of common fungal
diseases of maize in Western State, Nigeria. Nig. J. Plant Protection, 3, 11-15. OYEKAN, P. 0. and OGUNREMI, E. A., 1978. Damping-off of melon (Cucumeropsis edul is) in Niger,
caused by Macrophomina phaseolina. P1. Dis. Reptr., 62, 174-175. OYENUGA, V. A., 1968. Nigerian Foods and Feeding Stuffs. Ibadan University Press, 99 p. PAHARIA, K. D. and KOMMEDAHL, T., 1956. The effect of time and adding suspensions in soil
mycoflora assays. P1. Dis. Reptr., 40, 1029-1031. PAPAVISAS, G. C. and KLAG, N. G., 1975. Isolation and quantitative determination of Macro-
phomina phaseoli from soil. Phytopathol., 66, 624-625. SMALL, W., 1928. Note on the parasitism of Macrophomino phaseoli (MAUBL) ASHBY (Rhizoctonia
bataticola (TAuB.) BUTLER)). Trop. Agric., 6, 315-319. VASUDERA, R. S. and SIKKA, M. R., 1941. Studies on the root disease of cotton in the Punjab. X.
Effect of certain fungi on the growth of root rot fungi. Indian J. Agric. Sci., 11,422-431. WARCUP, J. H., 1955. Isolation of fungi from hyphae present in soil. Nature, 175, 953-954. WATANABE, T., 1973. Survivability of soil Macrophomina phaseoli (MAUBL.) ASHBY in naturally
infected soil and longevity of the sclerotia formed in vitro. Ann. Phytopathol. SOC. Japan, 39, 337.
WEINER, J. L. and LUTTREL, E. S., 1945. A canker of cowpea and Macrophomina leaf spot of cowpea and snap bean. P1. Dis. Reptr., 29, 27-29.
WELLMAN, F. L., 1934. Infection of Zea mays and various Gramineae by the celery virus in Florida. Phytopathol., 24, 1035-1037.
Mailing address: Dr. T. IKOTUN, Department of Agricultural Biology, University of Ibadan, Ibadan, Nigeria