the plant journal a perturbation in glutathione ...a perturbation in glutathione biosynthesis...
TRANSCRIPT

A perturbation in glutathione biosynthesis disruptsendoplasmic reticulum morphology and secretory membranetraffic in Arabidopsis thaliana
Kenneth K. C. Au1,†, Jose Perez-Gomez1,†, Helia Neto1,‡, Christopher Muller2, Andreas J. Meyer2,§, Mark D. Fricker1 and Ian
Moore1,*
1Department of Plant Sciences, University of Oxford, South Parks Rd, Oxford OX1 3RB, UK, and2Heidelberg Institute of Plant Sciences, University of Heidelberg, Im Neuenheimer Feld 360, D-69120 Heidelberg, Germany
Received 11 January 2012; revised 20 March 2012; accepted 22 March 2012; published online 12 July 2012.
*For correspondence (e-mail [email protected]).†These authors contributed equally to this work.‡Present address: The Henry Wellcome Laboratory for Cell Biology, College of Medical Veterinary and Life Sciences, University of Glasgow, Glasgow G12 8QQ, UK.§Present address: INRES – Chemical Signalling, University of Bonn, Friedrich-Ebert-Allee 144, D-53113 Bonn, Germany.
SUMMARY
To identify potentially novel and essential components of plant membrane trafficking mechanisms we
performed a GFP-based forward genetic screen for seedling-lethal biosynthetic membrane trafficking mutants
in Arabidopsis thaliana. Amongst these mutants, four recessive alleles of GSH2, which encodes glutathione
synthase (GSH2), were recovered. Each allele was characterized by loss of the typical polygonal endoplasmic
reticulum (ER) network and the accumulation of swollen ER-derived bodies which accumulated a soluble
secretory marker. Since GSH2 is responsible for converting c-glutamylcysteine (c-EC) to glutathione (GSH) in
the glutathione biosynthesis pathway, gsh2 mutants exhibited c-EC hyperaccumulation and GSH deficiency.
Redox-sensitive GFP revealed that gsh2 seedlings maintained redox poise in the cytoplasm but were more
sensitive to oxidative challenge. Genetic and pharmacological evidence indicated that c-EC accumulation
rather than GSH deficiency was responsible for the perturbation of ER morphology. Use of soluble and
membrane-bound ER markers suggested that the swollen ER bodies were derived from ER fusiform bodies.
Despite the gross perturbation of ER morphology, gsh2 seedlings did not suffer from constitutive oxidative ER
stress or lack of an unfolded protein response, and homozygotes for the weakest allele could be propagated.
The link between glutathione biosynthesis and ER morphology and function is discussed.
Keywords: secGFP, UPR, reticulon, roGFP, rml 1, GSH1, BIP, calreticulin, Arabidopsis thaliana
INTRODUCTION
Membrane trafficking is a tightly regulated and complex
process that transports macromolecules such as proteins,
polysaccharides, and lipids between endomembrane
organelles and plasma membrane. The biosynthetic and
trafficking functions of the secretory pathway organelles are
responsible for some of the most biologically important
functions of plant cells, such as the biogenesis of the plasma
membrane, vacuoles, cell walls, and cell plate. Membrane
trafficking involves the formation of a variety of specific
transport vesicles that package transported macromolecules
at one membrane compartment, followed by fusion of these
vesicles to a target membrane. This process can result in
either exchange between stable and biochemically distinct
compartments or in maturation of membrane compart-
ments over time (Huotari and Helenius, 2011). Much of our
fundamental knowledge comes from forward genetic stud-
ies in yeast, which have played a key role in uncovering
conserved regulators of the secretory pathway (Novick and
Schekman, 1979).
Although the basic molecular framework of the secretory
pathway is similar across eukaryotes, molecular and cyto-
logical studies suggest that plant cells have acquired
distinctive features in various aspects of membrane traffick-
ing, for example in relation to cell polarity and cytokinesis
(Bednarek and Falbel, 2002; Chow et al., 2008; Dettmer and
Friml, 2011; Muller et al., 2003). The existence of plant-
specific trafficking activities is also supported by phylo-
genetic studies of plant genomes which have shown extensive
diversification of important membrane trafficking protein
families such as Rab GTPases, Arf-guanine-nucleotide
ª 2012 The Authors 881The Plant Journal ª 2012 Blackwell Publishing Ltd
The Plant Journal (2012) 71, 881–894 doi: 10.1111/j.1365-313X.2012.05022.x

exchange factors (Arf-GEFs), and soluble N-ethylmaleimide-
sensitive factor attachment protein receptors (SNAREs)
(Kienle et al., 2009; Richter et al., 2007; Sanderfoot, 2007;
Woollard and Moore, 2008). In some cases, for example the
Arabidopsis syntaxin KNOLLE or the Arf-GEFs GNOM and
GNOMLIKE1 (GNL1), this reflects plant-specific neofunction-
alization (Muller et al., 2003; Richter et al., 2007; Teh and
Moore, 2007). Mutational studies have revealed significant
functional redundancy in many of these gene families,
however, which complicates reverse genetic analysis (Ebine
et al., 2011; Pinheiro et al., 2009; Sanderfoot, 2007). Conse-
quently, to identify genes with important functions in plant
membrane trafficking, several laboratories have used for-
ward genetic approaches employing fluorescent proteins as
trafficking markers in Arabidopsis thaliana.
Screens for mutants in which organelle markers showed
atypical localization have identified components that regu-
late organelle morphology (Faso et al., 2009; Matsushima
et al., 2003; Nakano et al., 2009; Rojo et al., 2001) or protein
trafficking in the endomembrane system (Agee et al., 2010;
Fuji et al., 2007; Tanaka et al., 2009; Teh and Moore, 2007).
We previously described the use of a secreted green
fluorescent protein (secGFP) as a marker to identify mem-
brane trafficking mutants in Arabidopsis (Teh and Moore,
2007; Zheng et al., 2004). Secreted GFP does not accumulate
in fluorescent form when secreted to the apoplast in wild-
type plants, but in mutants that perturb the secretory
pathway fluorescent secGFP accumulates in the cytoplasmic
organelles along the pathway and enhanced seedling fluo-
rescence is observed (Teh and Moore, 2007; Zheng et al.,
2004). Previous forward screens for membrane trafficking
mutants have identified viable plants, but it is to be expected
that perturbation of essential secretory functions will be
lethal in plants as in yeast (Novick and Schekman, 1979). It
has been argued that seedling-lethal mutants are more likely
than embryo-lethals to identify plant-specific functions that
underpin development rather than functions required sim-
ply for cell maintenance (Berna et al., 1999; Jurgens et al.,
1991). We reasoned that the same may be true of plant-
specific aspects of membrane trafficking where conserved,
core, eukaryotic trafficking functions are more likely to
mutate to embryo or gamete lethality. In support of this,
insertion mutants in conserved SNARE or Rab GTPases have
been shown to mutate to embryo- or gamete-lethal pheno-
types once redundancy is eliminated (e.g. Surpin et al., 2003;
Pinhiero et al., 2009) while the plant-specific trafficking loci
KNOLLE and GNOM were initially identified as seedling-
lethal mutants (Jurgens et al., 1991).
Here we report the use of secGFP and improved ratio-
metric fluorescent secretory markers (Samalova et al., 2006)
in a screen for seedling-lethal secretory mutants. We
describe the identification of four allelic mutations that
reveal an unexpected link between endoplasmic reticulum
(ER) organization and glutathione (GSH) biosynthesis. Glu-
tathione is a tripeptide composed of glutamate, cysteine,
and glycine and is the most abundant small thiol molecule
in cells with cytosolic concentration of 2–3 mM (Meyer et al.,
2001). It serves a diverse range of cellular functions, includ-
ing cell division (Vernoux et al., 2000), heavy metal detox-
ification (Cobbett et al., 1998), and regulation of disulfide
bond formation in the ER (Chakravarthi et al., 2006). Most
importantly, GSH plays a major role in maintenance of the
redox state and oxidative stress signaling (Meyer, 2008;
Noctor et al., 2000). Glutathione biosynthesis is a two-step
process that first combines glutamate and cysteine to form
gamma-glutamylcysteine (c-EC), followed by the addition
of glycine to form GSH. These two steps are catalyzed by
glutamate-cysteine ligase (GSH1) and glutathione synthase
(GSH2), respectively. Mutant alleles of GSH1 and GSH2 have
been described previously (Ball et al., 2004; Cairns et al.,
2006; Cobbett et al., 1998; Dubreuil-Maurizi et al., 2011; Jobe
et al., 2012; Parisy et al., 2007; Pasternak et al., 2008; Shan-
mugam et al., 2012; Vernoux et al., 2000) but their effects on
endomembrane function were not documented. Here we
show that ER morphology, but not protein folding, is
unexpectedly sensitive to a perturbation in the final step of
the GSH biosynthesis pathway.
RESULTS
To identify secretory membrane trafficking mutants we
screened for enhanced intracellular fluorescence in seed-
lings expressing the secGFP reporter (Teh and Moore, 2007;
Zheng et al., 2004) or the ratiometric secretory trafficking
markers nlsRm-2A-secGFPf and STN-Rm-2A-secGFPf
(Samalova et al., 2006). The latter two markers had the
advantage that instead of comparing absolute secGFP fluo-
rescence intensities, which can be affected by variation in
secGFP expression or imaging efficiency, putative mutants
could be identified by comparing the intensity ratio of sec-
GFP and a stoichiometrically expressed nuclear- (nlsRm) or
Golgi-targeted (STN-Rm) red fluorescent protein (RFP) mar-
ker. In previous screens for secGFP accumulators (Teh and
Moore, 2007), the brightest mutants were seedling lethal and
attempts to isolate temperature-sensitive alleles yielded few
mutants, so to recover the seedling-lethal mutations we
screened individual M2 families for secGFP accumulators
and propagated recessive mutations via heterozygous sib-
lings (see Experimental Procedures).
Four mutants accumulated secGFP in swollen bodies
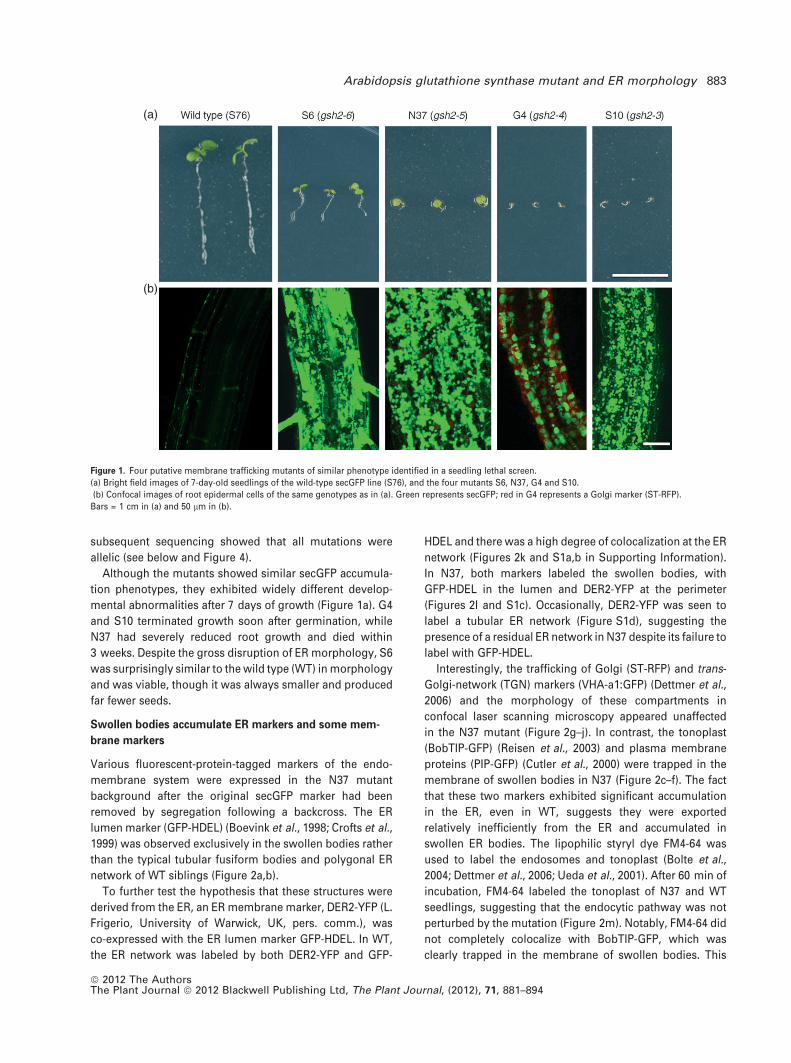
We selected four independent mutants – G4, N37, S6, and
S10 (Figure 1a)-for detailed study because they all mapped
to the same arm of chromosome V and confocal microscopy
revealed that they each accumulated secGFP in swollen
bodies 5–50 lm in diameter (Figure 1b). G4 was isolated
from a line expressing STN-Rm-2A-secGFPf, N37 from a line
expressing nlsRm-2A-secGFPf, and S6 and S10 from secGFP
line S76 (Zheng et al., 2004). Complementation tests and
882 Kenneth K. C. Au et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894
subsequent sequencing showed that all mutations were
allelic (see below and Figure 4).
Although the mutants showed similar secGFP accumula-
tion phenotypes, they exhibited widely different develop-
mental abnormalities after 7 days of growth (Figure 1a). G4
and S10 terminated growth soon after germination, while
N37 had severely reduced root growth and died within
3 weeks. Despite the gross disruption of ER morphology, S6
was surprisingly similar to the wild type (WT) in morphology
and was viable, though it was always smaller and produced
far fewer seeds.
Swollen bodies accumulate ER markers and some mem-
brane markers
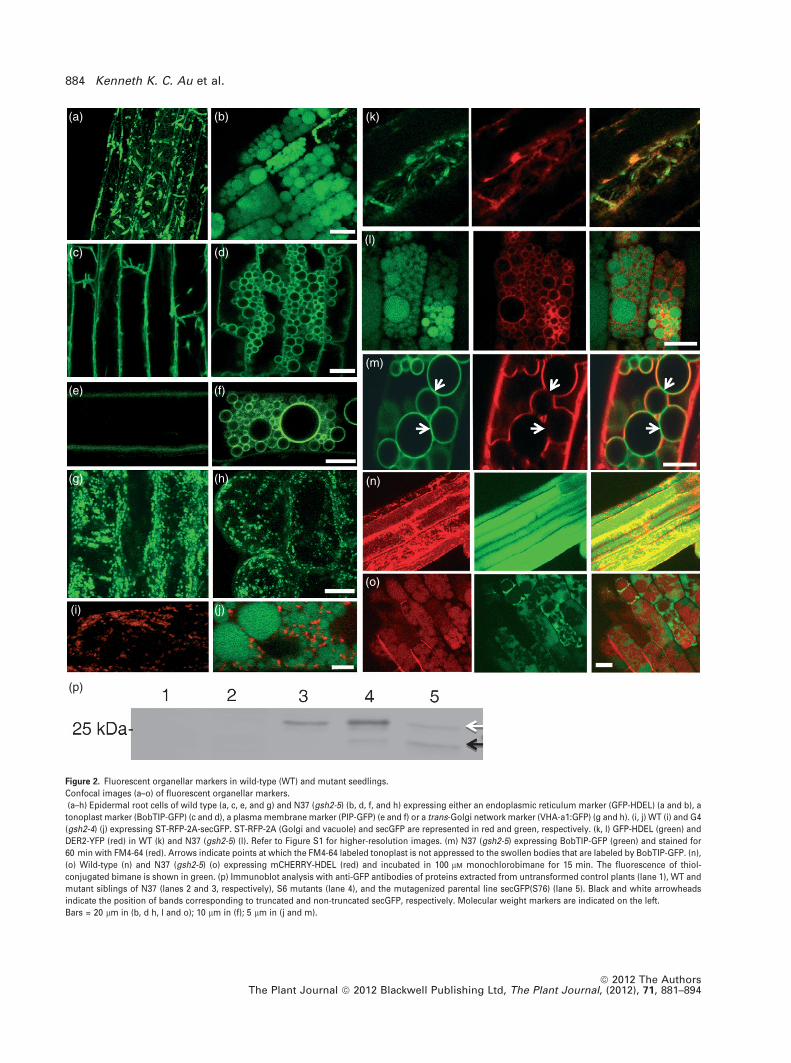
Various fluorescent-protein-tagged markers of the endo-
membrane system were expressed in the N37 mutant
background after the original secGFP marker had been
removed by segregation following a backcross. The ER
lumen marker (GFP-HDEL) (Boevink et al., 1998; Crofts et al.,
1999) was observed exclusively in the swollen bodies rather
than the typical tubular fusiform bodies and polygonal ER
network of WT siblings (Figure 2a,b).
To further test the hypothesis that these structures were
derived from the ER, an ER membrane marker, DER2-YFP (L.
Frigerio, University of Warwick, UK, pers. comm.), was
co-expressed with the ER lumen marker GFP-HDEL. In WT,
the ER network was labeled by both DER2-YFP and GFP-
HDEL and there was a high degree of colocalization at the ER
network (Figures 2k and S1a,b in Supporting Information).
In N37, both markers labeled the swollen bodies, with
GFP-HDEL in the lumen and DER2-YFP at the perimeter
(Figures 2l and S1c). Occasionally, DER2-YFP was seen to
label a tubular ER network (Figure S1d), suggesting the
presence of a residual ER network in N37 despite its failure to
label with GFP-HDEL.
Interestingly, the trafficking of Golgi (ST-RFP) and trans-
Golgi-network (TGN) markers (VHA-a1:GFP) (Dettmer et al.,
2006) and the morphology of these compartments in
confocal laser scanning microscopy appeared unaffected
in the N37 mutant (Figure 2g–j). In contrast, the tonoplast
(BobTIP-GFP) (Reisen et al., 2003) and plasma membrane
proteins (PIP-GFP) (Cutler et al., 2000) were trapped in the
membrane of swollen bodies in N37 (Figure 2c–f). The fact
that these two markers exhibited significant accumulation
in the ER, even in WT, suggests they were exported
relatively inefficiently from the ER and accumulated in
swollen ER bodies. The lipophilic styryl dye FM4-64 was
used to label the endosomes and tonoplast (Bolte et al.,
2004; Dettmer et al., 2006; Ueda et al., 2001). After 60 min of
incubation, FM4-64 labeled the tonoplast of N37 and WT
seedlings, suggesting that the endocytic pathway was not
perturbed by the mutation (Figure 2m). Notably, FM4-64 did
not completely colocalize with BobTIP-GFP, which was
clearly trapped in the membrane of swollen bodies. This
(a)
(b)
Figure 1. Four putative membrane trafficking mutants of similar phenotype identified in a seedling lethal screen.
(a) Bright field images of 7-day-old seedlings of the wild-type secGFP line (S76), and the four mutants S6, N37, G4 and S10.
(b) Confocal images of root epidermal cells of the same genotypes as in (a). Green represents secGFP; red in G4 represents a Golgi marker (ST-RFP).
Bars = 1 cm in (a) and 50 lm in (b).
Arabidopsis glutathione synthase mutant and ER morphology 883
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894
(a) (b) (k)
(l)
(m)
(n)
(o)
(c) (d)
(e) (f)
(g) (h)
(i)
(p)
(j)
Figure 2. Fluorescent organellar markers in wild-type (WT) and mutant seedlings.
Confocal images (a–o) of fluorescent organellar markers.
(a–h) Epidermal root cells of wild type (a, c, e, and g) and N37 (gsh2-5) (b, d, f, and h) expressing either an endoplasmic reticulum marker (GFP-HDEL) (a and b), a
tonoplast marker (BobTIP-GFP) (c and d), a plasma membrane marker (PIP-GFP) (e and f) or a trans-Golgi network marker (VHA-a1:GFP) (g and h). (i, j) WT (i) and G4
(gsh2-4) (j) expressing ST-RFP-2A-secGFP. ST-RFP-2A (Golgi and vacuole) and secGFP are represented in red and green, respectively. (k, l) GFP-HDEL (green) and
DER2-YFP (red) in WT (k) and N37 (gsh2-5) (l). Refer to Figure S1 for higher-resolution images. (m) N37 (gsh2-5) expressing BobTIP-GFP (green) and stained for
60 min with FM4-64 (red). Arrows indicate points at which the FM4-64 labeled tonoplast is not appressed to the swollen bodies that are labeled by BobTIP-GFP. (n),
(o) Wild-type (n) and N37 (gsh2-5) (o) expressing mCHERRY-HDEL (red) and incubated in 100 lM monochlorobimane for 15 min. The fluorescence of thiol-
conjugated bimane is shown in green. (p) Immunoblot analysis with anti-GFP antibodies of proteins extracted from untransformed control plants (lane 1), WT and
mutant siblings of N37 (lanes 2 and 3, respectively), S6 mutants (lane 4), and the mutagenized parental line secGFP(S76) (lane 5). Black and white arrowheads
indicate the position of bands corresponding to truncated and non-truncated secGFP, respectively. Molecular weight markers are indicated on the left.
Bars = 20 lm in (b, d h, l and o); 10 lm in (f); 5 lm in (j and m).
884 Kenneth K. C. Au et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894
suggests that the endocytic and biosynthetic tonoplast
markers FM4-64 and BobTIP-GFP labeled different compart-
ments in the mutant. To test this further we labeled the
vacuolar lumen using monochlorobimane (MCB), which
becomes fluorescent only after it conjugates reduced cyto-
solic thiols, principally GSH in WT cells. The conjugated
product, glutathione-S-bimane (GSB), is then actively trans-
ported to the vacuole (Fricker et al., 2000). In N37, fluores-
cent bimane-labeled vacuoles did not colocalize with the ER
luminal marker mCHERRY-HDEL (Nelson et al., 2007) in the
swollen bodies (Figure 2n,o). Hence, we concluded that the
swollen bodies were distinct from vacuoles. This conclusion
is supported by immunoblot analysis (Figure 2p), which
showed that in N37 and S6, secGFP accumulated in its full-
length form, typical of the ER, rather than the C-terminally
truncated form normally observed in the vacuole or apop-
last (Zheng et al., 2004).
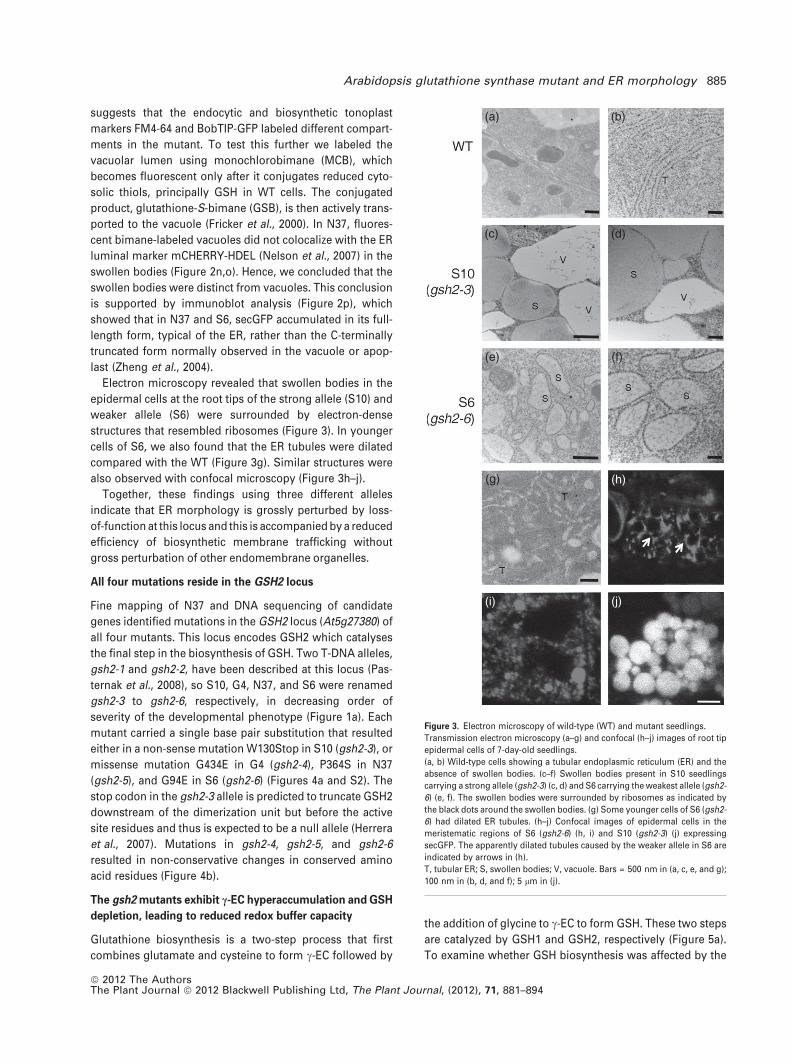
Electron microscopy revealed that swollen bodies in the
epidermal cells at the root tips of the strong allele (S10) and
weaker allele (S6) were surrounded by electron-dense
structures that resembled ribosomes (Figure 3). In younger
cells of S6, we also found that the ER tubules were dilated
compared with the WT (Figure 3g). Similar structures were
also observed with confocal microscopy (Figure 3h–j).
Together, these findings using three different alleles
indicate that ER morphology is grossly perturbed by loss-
of-function at this locus and this is accompanied by a reduced
efficiency of biosynthetic membrane trafficking without
gross perturbation of other endomembrane organelles.
All four mutations reside in the GSH2 locus
Fine mapping of N37 and DNA sequencing of candidate
genes identified mutations in the GSH2 locus (At5g27380) of
all four mutants. This locus encodes GSH2 which catalyses
the final step in the biosynthesis of GSH. Two T-DNA alleles,
gsh2-1 and gsh2-2, have been described at this locus (Pas-
ternak et al., 2008), so S10, G4, N37, and S6 were renamed
gsh2-3 to gsh2-6, respectively, in decreasing order of
severity of the developmental phenotype (Figure 1a). Each
mutant carried a single base pair substitution that resulted
either in a non-sense mutation W130Stop in S10 (gsh2-3), or
missense mutation G434E in G4 (gsh2-4), P364S in N37
(gsh2-5), and G94E in S6 (gsh2-6) (Figures 4a and S2). The
stop codon in the gsh2-3 allele is predicted to truncate GSH2
downstream of the dimerization unit but before the active
site residues and thus is expected to be a null allele (Herrera
et al., 2007). Mutations in gsh2-4, gsh2-5, and gsh2-6
resulted in non-conservative changes in conserved amino
acid residues (Figure 4b).
The gsh2 mutants exhibit c-EC hyperaccumulation and GSH
depletion, leading to reduced redox buffer capacity
Glutathione biosynthesis is a two-step process that first
combines glutamate and cysteine to form c-EC followed by
the addition of glycine to c-EC to form GSH. These two steps
are catalyzed by GSH1 and GSH2, respectively (Figure 5a).
To examine whether GSH biosynthesis was affected by the
(a) (b)
(c) (d)
(e) (f)
(g) (h)
(i) (j)
Figure 3. Electron microscopy of wild-type (WT) and mutant seedlings.
Transmission electron microscopy (a–g) and confocal (h–j) images of root tip
epidermal cells of 7-day-old seedlings.
(a, b) Wild-type cells showing a tubular endoplasmic reticulum (ER) and the
absence of swollen bodies. (c–f) Swollen bodies present in S10 seedlings
carrying a strong allele (gsh2-3) (c, d) and S6 carrying the weakest allele (gsh2-
6) (e, f). The swollen bodies were surrounded by ribosomes as indicated by
the black dots around the swollen bodies. (g) Some younger cells of S6 (gsh2-
6) had dilated ER tubules. (h–j) Confocal images of epidermal cells in the
meristematic regions of S6 (gsh2-6) (h, i) and S10 (gsh2-3) (j) expressing
secGFP. The apparently dilated tubules caused by the weaker allele in S6 are
indicated by arrows in (h).
T, tubular ER; S, swollen bodies; V, vacuole. Bars = 500 nm in (a, c, e, and g);
100 nm in (b, d, and f); 5 lm in (j).
Arabidopsis glutathione synthase mutant and ER morphology 885
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894

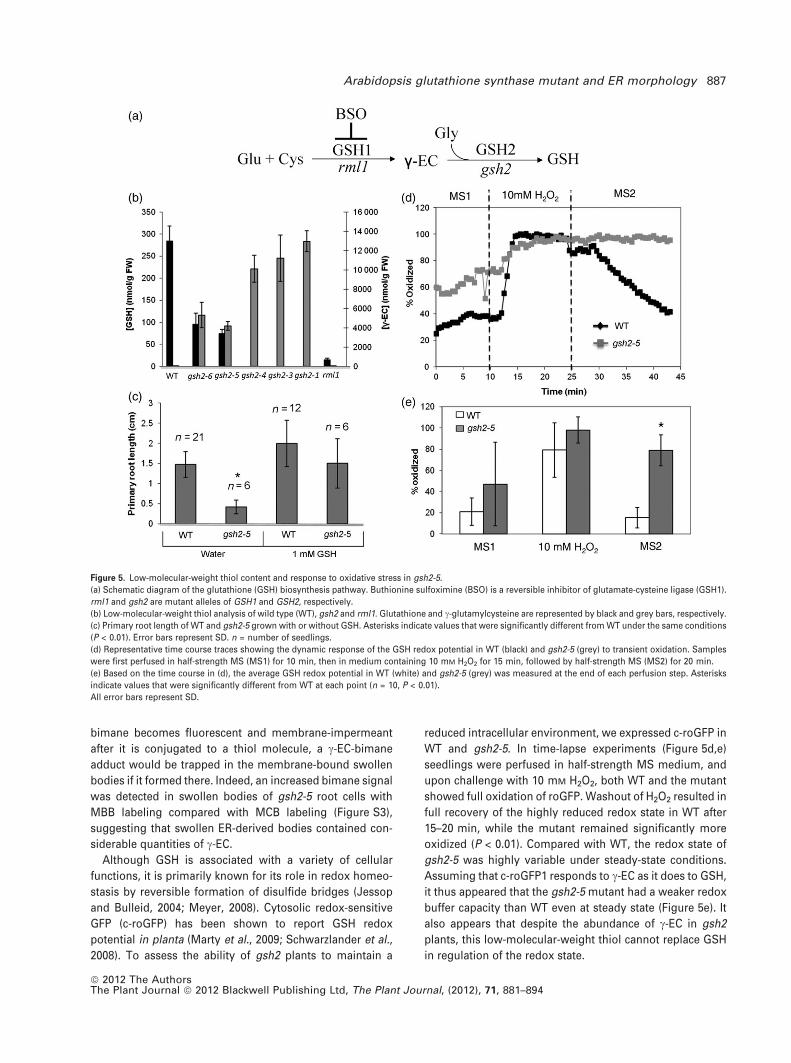
GSH2 mutations, we measured the content of cytosolic c-EC
and GSH in 10-day-old seedlings of gsh2 alleles. In this
analysis, we also included the null T-DNA allele gsh2-1
(Pasternak et al., 2008) and rml1 which is a GSH1 mutant
(Vernoux et al., 2000). While the level of GSH in WT was, as
expected, much higher than that of c-EC, c-EC was the pre-
dominant small thiol in all gsh2 mutants (Figure 5b). Among
the gsh2 mutants, gsh2-1, gsh2-3, and gsh2-4 showed the
greatest degree of c-EC hyperaccumulation and GSH defi-
ciency. The c-EC content in these mutants was 10–13 lmol
g)1 which is about 400-fold greater than in WT whilst GSH
was undetectable, suggesting that there was little or no
GSH2 activity. Hyperaccumulation of c-EC was also
observed in gsh2-5 and gsh2-6, although to a lesser extent
(170-fold greater than in WT), and their GSH content was
higher than that of rml1. Germination of gsh2-5 on medium
supplemented with 1 mM GSH rescued root growth (Fig-
ure 5c), indicating that the developmental abnormality was
attributable to the perturbation in GSH biosynthesis. Thus
each of the mutations in gsh2-3 to gsh2-6 apparently caused
a reduction in GSH2 activity, thereby increasing the abun-
dance of its substrate, c-EC, and decreasing the abundance
of its product, GSH.
To investigate the small thiol content of the swollen ER
bodies, roots were stained with monobromobimane (MBB)
and MCB. Unlike MCB that preferentially labels reduced GSH
through a GST-catalysed reaction (Meyer et al., 2001), MBB
is more reactive and labels small thiols non-specifically
(Fahey et al., 1981). As the analysis of low molecular weight
thiols of gsh2 revealed that c-EC replaced GSH as the most
abundant small thiol molecule, we expected that MBB would
mostly label c-EC in gsh2 seedlings. Furthermore, because
(a)
(b)
Figure 4. Mutations in the GSH2 locus of mutants S10, G4, N37 and S6.
(a) Location in the GSH2 locus (At5g27380) of the mutation in each allele (numbering refers to codons). S10, G4, N37, and S6 were renamed gsh2-3, gsh2-4, gsh2-5,
and gsh2-6, respectively.
(b) Alignment of glutathione synthase (GSH2) protein sequence of Arabidopsis, human (Homo sapiens) and yeast (Saccharomyces cerevisiae) using CLUSTALW.
Residues affected by mutation in G4, N37, S6 and S10 are boxed. Symbols below the sequences denote the degree of conservation. Asterisks (*) indicate identical
residues; semi-colon (:) and full-stop (.) indicate conserved and semi-conserved substitutions, respectively.
886 Kenneth K. C. Au et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894
bimane becomes fluorescent and membrane-impermeant
after it is conjugated to a thiol molecule, a c-EC-bimane
adduct would be trapped in the membrane-bound swollen
bodies if it formed there. Indeed, an increased bimane signal
was detected in swollen bodies of gsh2-5 root cells with
MBB labeling compared with MCB labeling (Figure S3),
suggesting that swollen ER-derived bodies contained con-
siderable quantities of c-EC.
Although GSH is associated with a variety of cellular
functions, it is primarily known for its role in redox homeo-
stasis by reversible formation of disulfide bridges (Jessop
and Bulleid, 2004; Meyer, 2008). Cytosolic redox-sensitive
GFP (c-roGFP) has been shown to report GSH redox
potential in planta (Marty et al., 2009; Schwarzlander et al.,
2008). To assess the ability of gsh2 plants to maintain a
reduced intracellular environment, we expressed c-roGFP in
WT and gsh2-5. In time-lapse experiments (Figure 5d,e)
seedlings were perfused in half-strength MS medium, and
upon challenge with 10 mM H2O2, both WT and the mutant
showed full oxidation of roGFP. Washout of H2O2 resulted in
full recovery of the highly reduced redox state in WT after
15–20 min, while the mutant remained significantly more
oxidized (P < 0.01). Compared with WT, the redox state of
gsh2-5 was highly variable under steady-state conditions.
Assuming that c-roGFP1 responds to c-EC as it does to GSH,
it thus appeared that the gsh2-5 mutant had a weaker redox
buffer capacity than WT even at steady state (Figure 5e). It
also appears that despite the abundance of c-EC in gsh2
plants, this low-molecular-weight thiol cannot replace GSH
in regulation of the redox state.
Figure 5. Low-molecular-weight thiol content and response to oxidative stress in gsh2-5.
(a) Schematic diagram of the glutathione (GSH) biosynthesis pathway. Buthionine sulfoximine (BSO) is a reversible inhibitor of glutamate-cysteine ligase (GSH1).
rml1 and gsh2 are mutant alleles of GSH1 and GSH2, respectively.
(b) Low-molecular-weight thiol analysis of wild type (WT), gsh2 and rml1. Glutathione and c-glutamylcysteine are represented by black and grey bars, respectively.
(c) Primary root length of WT and gsh2-5 grown with or without GSH. Asterisks indicate values that were significantly different from WT under the same conditions
(P < 0.01). Error bars represent SD. n = number of seedlings.
(d) Representative time course traces showing the dynamic response of the GSH redox potential in WT (black) and gsh2-5 (grey) to transient oxidation. Samples
were first perfused in half-strength MS (MS1) for 10 min, then in medium containing 10 mM H2O2 for 15 min, followed by half-strength MS (MS2) for 20 min.
(e) Based on the time course in (d), the average GSH redox potential in WT (white) and gsh2-5 (grey) was measured at the end of each perfusion step. Asterisks
indicate values that were significantly different from WT at each point (n = 10, P < 0.01).
All error bars represent SD.
Arabidopsis glutathione synthase mutant and ER morphology 887
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894
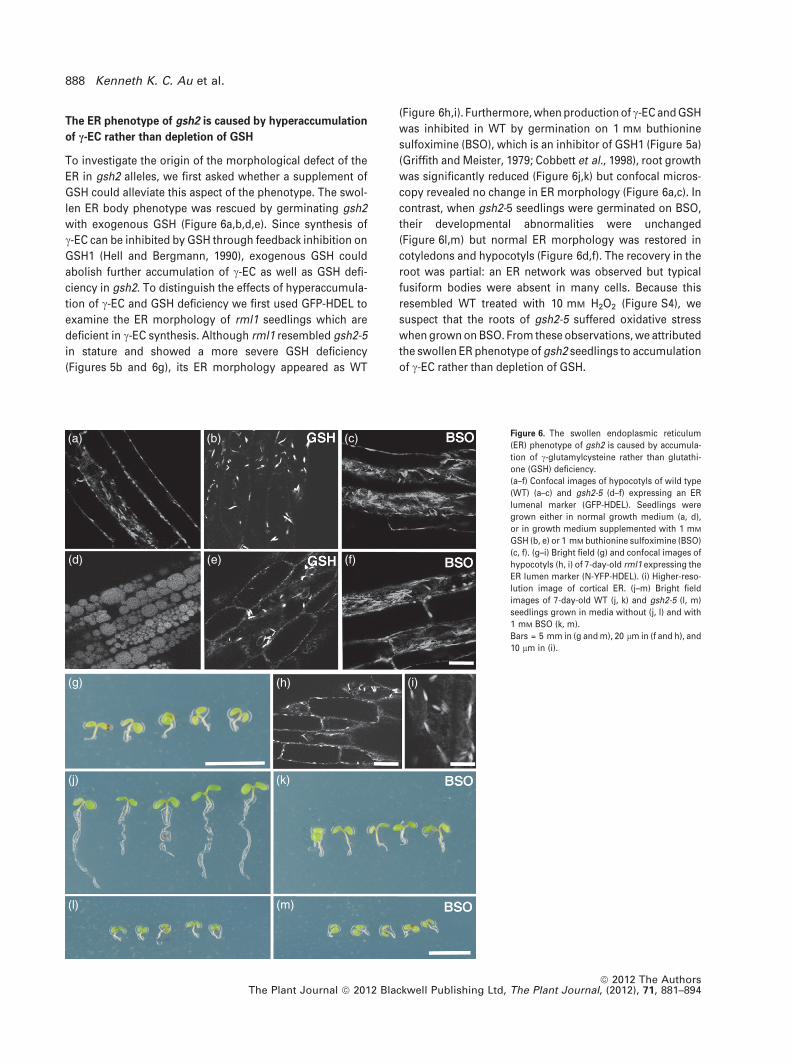
The ER phenotype of gsh2 is caused by hyperaccumulation
of c-EC rather than depletion of GSH
To investigate the origin of the morphological defect of the
ER in gsh2 alleles, we first asked whether a supplement of
GSH could alleviate this aspect of the phenotype. The swol-
len ER body phenotype was rescued by germinating gsh2
with exogenous GSH (Figure 6a,b,d,e). Since synthesis of
c-EC can be inhibited by GSH through feedback inhibition on
GSH1 (Hell and Bergmann, 1990), exogenous GSH could
abolish further accumulation of c-EC as well as GSH defi-
ciency in gsh2. To distinguish the effects of hyperaccumula-
tion of c-EC and GSH deficiency we first used GFP-HDEL to
examine the ER morphology of rml1 seedlings which are
deficient in c-EC synthesis. Although rml1 resembled gsh2-5
in stature and showed a more severe GSH deficiency
(Figures 5b and 6g), its ER morphology appeared as WT
(Figure 6h,i). Furthermore, when production of c-EC and GSH
was inhibited in WT by germination on 1 mM buthionine
sulfoximine (BSO), which is an inhibitor of GSH1 (Figure 5a)
(Griffith and Meister, 1979; Cobbett et al., 1998), root growth
was significantly reduced (Figure 6j,k) but confocal micros-
copy revealed no change in ER morphology (Figure 6a,c). In
contrast, when gsh2-5 seedlings were germinated on BSO,
their developmental abnormalities were unchanged
(Figure 6l,m) but normal ER morphology was restored in
cotyledons and hypocotyls (Figure 6d,f). The recovery in the
root was partial: an ER network was observed but typical
fusiform bodies were absent in many cells. Because this
resembled WT treated with 10 mM H2O2 (Figure S4), we
suspect that the roots of gsh2-5 suffered oxidative stress
when grown on BSO. From these observations, we attributed
the swollen ER phenotype of gsh2 seedlings to accumulation
of c-EC rather than depletion of GSH.
(a) (b) (c)
(d) (e) (f)
(g)
(j)
(l) (m)
(k)
(h) (i)
Figure 6. The swollen endoplasmic reticulum
(ER) phenotype of gsh2 is caused by accumula-
tion of c-glutamylcysteine rather than glutathi-
one (GSH) deficiency.
(a–f) Confocal images of hypocotyls of wild type
(WT) (a–c) and gsh2-5 (d–f) expressing an ER
lumenal marker (GFP-HDEL). Seedlings were
grown either in normal growth medium (a, d),
or in growth medium supplemented with 1 mM
GSH (b, e) or 1 mM buthionine sulfoximine (BSO)
(c, f). (g–i) Bright field (g) and confocal images of
hypocotyls (h, i) of 7-day-old rml1 expressing the
ER lumen marker (N-YFP-HDEL). (i) Higher-reso-
lution image of cortical ER. (j–m) Bright field
images of 7-day-old WT (j, k) and gsh2-5 (l, m)
seedlings grown in media without (j, l) and with
1 mM BSO (k, m).
Bars = 5 mm in (g and m), 20 lm in (f and h), and
10 lm in (i).
888 Kenneth K. C. Au et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894
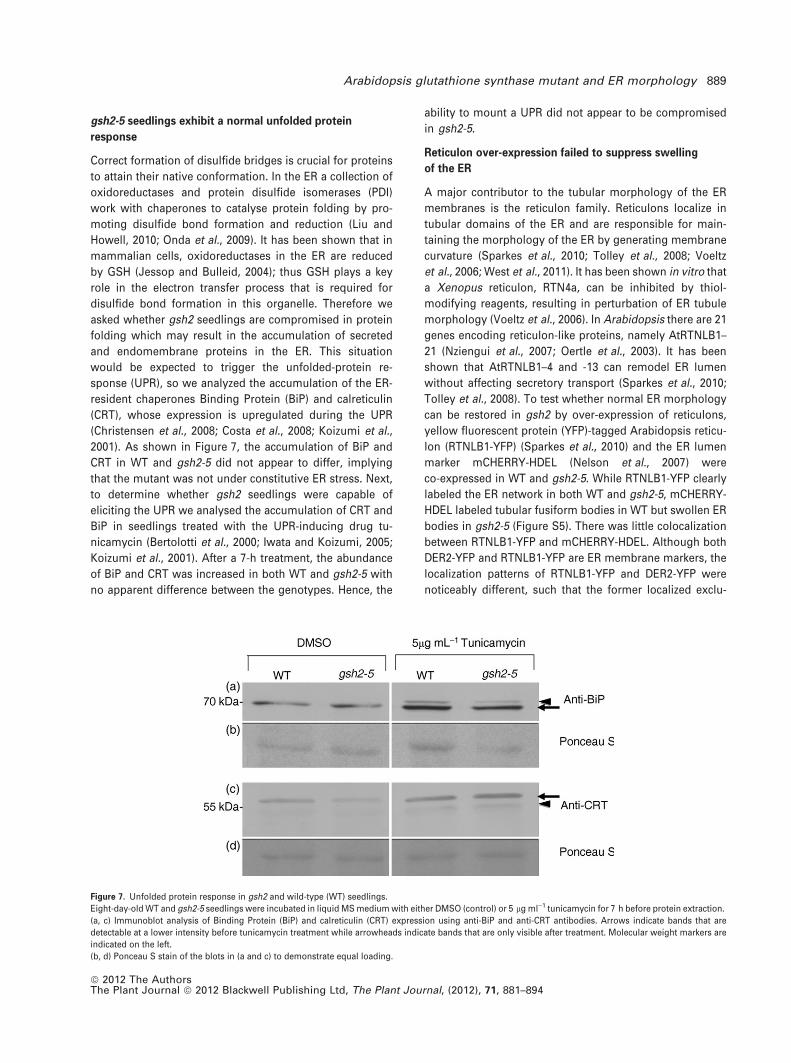
gsh2-5 seedlings exhibit a normal unfolded protein
response
Correct formation of disulfide bridges is crucial for proteins
to attain their native conformation. In the ER a collection of
oxidoreductases and protein disulfide isomerases (PDI)
work with chaperones to catalyse protein folding by pro-
moting disulfide bond formation and reduction (Liu and
Howell, 2010; Onda et al., 2009). It has been shown that in
mammalian cells, oxidoreductases in the ER are reduced
by GSH (Jessop and Bulleid, 2004); thus GSH plays a key
role in the electron transfer process that is required for
disulfide bond formation in this organelle. Therefore we
asked whether gsh2 seedlings are compromised in protein
folding which may result in the accumulation of secreted
and endomembrane proteins in the ER. This situation
would be expected to trigger the unfolded-protein re-
sponse (UPR), so we analyzed the accumulation of the ER-
resident chaperones Binding Protein (BiP) and calreticulin
(CRT), whose expression is upregulated during the UPR
(Christensen et al., 2008; Costa et al., 2008; Koizumi et al.,
2001). As shown in Figure 7, the accumulation of BiP and
CRT in WT and gsh2-5 did not appear to differ, implying
that the mutant was not under constitutive ER stress. Next,
to determine whether gsh2 seedlings were capable of
eliciting the UPR we analysed the accumulation of CRT and
BiP in seedlings treated with the UPR-inducing drug tu-
nicamycin (Bertolotti et al., 2000; Iwata and Koizumi, 2005;
Koizumi et al., 2001). After a 7-h treatment, the abundance
of BiP and CRT was increased in both WT and gsh2-5 with
no apparent difference between the genotypes. Hence, the
ability to mount a UPR did not appear to be compromised
in gsh2-5.
Reticulon over-expression failed to suppress swelling
of the ER
A major contributor to the tubular morphology of the ER
membranes is the reticulon family. Reticulons localize in
tubular domains of the ER and are responsible for main-
taining the morphology of the ER by generating membrane
curvature (Sparkes et al., 2010; Tolley et al., 2008; Voeltz
et al., 2006; West et al., 2011). It has been shown in vitro that
a Xenopus reticulon, RTN4a, can be inhibited by thiol-
modifying reagents, resulting in perturbation of ER tubule
morphology (Voeltz et al., 2006). In Arabidopsis there are 21
genes encoding reticulon-like proteins, namely AtRTNLB1–
21 (Nziengui et al., 2007; Oertle et al., 2003). It has been
shown that AtRTNLB1–4 and -13 can remodel ER lumen
without affecting secretory transport (Sparkes et al., 2010;
Tolley et al., 2008). To test whether normal ER morphology
can be restored in gsh2 by over-expression of reticulons,
yellow fluorescent protein (YFP)-tagged Arabidopsis reticu-
lon (RTNLB1-YFP) (Sparkes et al., 2010) and the ER lumen
marker mCHERRY-HDEL (Nelson et al., 2007) were
co-expressed in WT and gsh2-5. While RTNLB1-YFP clearly
labeled the ER network in both WT and gsh2-5, mCHERRY-
HDEL labeled tubular fusiform bodies in WT but swollen ER
bodies in gsh2-5 (Figure S5). There was little colocalization
between RTNLB1-YFP and mCHERRY-HDEL. Although both
DER2-YFP and RTNLB1-YFP are ER membrane markers, the
localization patterns of RTNLB1-YFP and DER2-YFP were
noticeably different, such that the former localized exclu-
Figure 7. Unfolded protein response in gsh2 and wild-type (WT) seedlings.
Eight-day-old WT and gsh2-5 seedlings were incubated in liquid MS medium with either DMSO (control) or 5 lg ml)1 tunicamycin for 7 h before protein extraction.
(a, c) Immunoblot analysis of Binding Protein (BiP) and calreticulin (CRT) expression using anti-BiP and anti-CRT antibodies. Arrows indicate bands that are
detectable at a lower intensity before tunicamycin treatment while arrowheads indicate bands that are only visible after treatment. Molecular weight markers are
indicated on the left.
(b, d) Ponceau S stain of the blots in (a and c) to demonstrate equal loading.
Arabidopsis glutathione synthase mutant and ER morphology 889
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894

sively in the tubular ER network and the latter localized in the
membrane of swollen bodies (compare Figures 2l and S5b).
Thus RTNLB1-YFP over-expression was apparently unable
to restore ER membrane morphology.
DISCUSSION
In this study we have characterized four alleles of GSH2 that
were identified in a screen for secretory membrane traffick-
ing mutants. We show that a perturbation in GSH biosyn-
thesis can affect both the operation of the secretory pathway
and the morphology of the ER. Although the four gsh2
alleles all showed a similar degree of perturbation in ER
morphology and secretory trafficking, they exhibited a wide
range of developmental abnormalities. Surprisingly, the
weakest allele, gsh2-6, was viable which indicates that gross
perturbation of ER morphology is compatible with viability
of the cell and organism. Consistent with this, the surface of
the swollen ER bodies was decorated with ribosomes and
was thus apparently competent for translocation.
The small thiol profile of the two strong alleles was
comparable to that of a previously described gsh2 null
mutant gsh2-1 (Pasternak et al., 2008). The intermediate
allele, gsh2-5, which undergoes post-germinative meriste-
matic development, had decreased redox buffer capacity
and increased sensitivity to oxidative stress. In addition,
roGFP1 expressed in this mutant exhibited a variable
cytosolic redox state that was substantially more oxidized
than WT (on average 21 and 47% oxidized in WT and gsh2-5,
respectively). We attempted to measure the GSH redox state
in the ER using modified roGFPs with a more oxidizing
midpoint potential (Lohman and Remington, 2008) but these
probes were still almost fully oxidized even in WT ER,
precluding reliable measurement of any increase in the
oxidation state in gsh2.
It has been reported previously that the complete absence
of GSH in gsh1 null mutants confers embryo lethality (Cairns
et al., 2006), whereas gsh2 null mutants are seedling lethal
owing perhaps to the accumulation of c-EC, which may
substitute for GSH in some instances (Pasternak et al., 2008).
Here, using genetic and pharmacological evidence, we show
that the deficiency in GSH determines the degree of devel-
opmental abnormality in gsh2 and rml1, whereas it is the
accumulation of c-EC, but not GSH deficiency, that affects
the morphology and secretory pathway of the ER. Thus tight
regulation of c-EC synthesis and conversion to GSH may be
required to maintain this aspect of cellular function. Indeed
GSH1 activity shows feedback inhibition by GSH (Hell and
Bergmann, 1990) and this fails when GSH2 is defective
(Pasternak et al., 2008). This also means that although
exogenous GSH can suppress the ER morphology pheno-
type of gsh2, it is likely to act indirectly by alleviating
accumulation of c-EC through feedback inhibition of GSH1
(Hell and Bergmann, 1990). This may explain why, in initial
tests, we failed to rescue the swollen ER phenotype of gsh2 if
we supplied GSH after germination when c-EC had already
accumulated to a significant level. Recently, Jobe et al.
(2012) reported a viable loss-of-function mutation at the
gsh2 locus. This allele, nrc2, was identified in a screen for
mutants with a reduced response to exogenous cadmium.
Based on growth and c-EC accumulation, ncr2 appears to be
weaker than any of the alleles described here but its
secretory activity and ER morphology have not been char-
acterized.
The regulated formation of disulfide bridges is an essen-
tial process for the proper folding and maturation of newly
synthesized proteins. Changes in the redox state of specific
thiol groups on some proteins can result in their aggregation
in the ER, inhibiting anterograde traffic (Kawagoe et al.,
2005; Orsi et al., 2001; Pompa and Vitale, 2006). Glutathione
is required directly or indirectly to reduce ER proteins via
enzymatic reactions (Jessop and Bulleid, 2004), and it has
been postulated that ER stress may lead to enlargement of
the lumen to accommodate more misfolded proteins (Banh-
egyi et al., 2007). Glutathione has been detected in the
Arabidopsis ER (Zechmann et al., 2008), but despite the
gross perturbation of small thiol accumulation, gsh2-5
apparently does not significantly accumulate unfolded pro-
tein or protein aggregates at steady state. Nevertheless, the
UPR can still be triggered in gsh2-5, showing that the
monitoring of protein misfolding is not prevented. It is
surprising that despite the expected need for GSH in
disulfide bond formation, it is ER morphology and protein
export rather than general protein folding that appear to be
most sensitive to perturbation of GSH metabolism.
Since the integral membrane markers of the Golgi (ST-
RFP) and TGN (VHA-a1:GFP) are able to reach their targeted
compartment but secGFP, the plasma membrane (PM)
marker (PIP-GFP) and the tonoplast marker (BobTIP-GFP)
are not, ER export appears to be partially blocked in gsh2.
One possible explanation is that the change in c-EC levels
causes a cargo-specific folding or selection process to be
disrupted. An alternative explanation is suggested, how-
ever, by the observation that even in WT cells PIP-GFP and
BobTIP-GFP were often detectable in the ER as well as their
resident membrane. This suggests that they may be ineffi-
ciently exported and thus more sensitive to general pertur-
bation of membrane trafficking by accumulation of c-EC.
Indeed it has been shown that early steps of secretory
membrane traffic in tobacco protoplasts are sensitive to
exogenously applied thiol-containing reagents (Pompa and
Vitale, 2006).
It is still unclear how c-EC accumulation affects ER
morphology. Animal reticulon proteins are required for the
formation of tubular ER and they are known to affect ER
morphology in plants (Sparkes et al., 2010; Tolley et al.,
2008; Voeltz et al., 2006). The function of animal reticulon
can be perturbed by treatment with thiol-modifying reagents
such as N-ethyl-maleimide (NEM), so we hypothesized that
890 Kenneth K. C. Au et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894

the accumulated c-EC in gsh2 seedlings may affect ER
morphology by interacting with one or more of the 21 plant
reticulons in Arabidopsis (Nziengui et al., 2007; Oertle et al.,
2003). However, we were unable to eliminate the swollen
bodies in gsh2 by overexpressing reticulons, nor could we
phenocopy the swollen bodies of gsh2 plants by growing
WT with various concentrations of NEM (Figure S5).
The ER is a morphologically dynamic structure that also
changes during development (Ridge et al., 1999). In many
Brassicaceae plants including Arabidopsis, parts of the ER
also exist as spindle-shaped structures known as ER bodies
(Behnke and Eschlbeck, 1978; Iversen, 1970; Matsushima
et al., 2003). These structures accumulate fluorescent pro-
teins with a C-terminal H/KDEL retrieval motif (Hawes et al.,
2001; Toyooka et al., 2000; Yamada et al., 2008). Indeed,
such markers predominantly label ER bodies in WT but
swollen ER in gsh2. Similar to fusiform ER bodies (Hawes
et al., 2001), the swollen bodies in gsh2 were also sur-
rounded by ribosomes, suggesting that they were functional
for protein translocation. Because we do not see any
spindle-shaped ER bodies in gsh2, the swollen bodies in
gsh2 may be derived from these ER bodies.
Interestingly, of all the ER markers that were tested,
RTNLB1-YFP was the only one that exclusively labeled the ER
network in gsh2-5, while all other markers (DER2-YFP, GFP-
HDEL and mCHERRY-HDEL) were localized predominantly in
swollen bodies. This may be attributed to the fact that
reticulons preferentially localize in membranes of high
curvature such as the tubular network so are excluded from
less curved surfaces like the spherical swollen bodies.
RTNLB1-YFP may have preferentially labeled the residual
ER network in gsh2. One possibility is that accumulation of
c-EC in the ER may directly cause swelling of the ER bodies.
In fact, results from MBB and MCB labeling suggested that
c-EC was abundant in the swollen bodies of gsh2-5. Even in
the weak alleles of gsh2 the total quantity of c-EC in the tissue
is approximately 20-fold greater than the total WT GSH
content. The GSH content of WT plants corresponds to a
cytosolic concentration of 2–3 mM, so c-EC may exceed
50 mM in gsh2. We speculate that the accumulation of such
quantities of c-EC in the relatively small volume of the ER
may perhaps be sufficient to induce swelling. The apparent
absence of reticulons from the surface of the spindle-shaped
ER bodies may make these regions more susceptible to
swelling if the ER volume increases, giving rise to the
swollen bodies in gsh2. It is unclear how c-EC may enter the
ER. In WT cells c-EC is undetectable, being rapidly converted
to GSH by cytosolic GSH2, so a specific transporter is
unlikely. Glutathione has been immunologically detected,
although not quantified, in the ER of Arabidopsis (Zechmann
et al., 2008) but is present in the ER lumen of animal cells at a
similar concentration to that of the cytosol (Banhegyi et al.,
1999; Hwang et al., 1992; Le Gall et al., 2004), so perhaps
c-EC could utilize the same transport pathway to enter the ER.
The phenotypes of the gsh2 alleles reported here high-
light the need for significant further investigation to fully
account for the unexpected influence of GSH metabolism on
form and function of the ER.
EXPERIMENTAL PROCEDURES
Plant material and growth conditions
Introgressions of c-roGFP1 (Schwarzlander et al., 2008), GFP-HDEL(Zheng et al., 2004), ST-RFP (Teh and Moore, 2007), BobTIP-GFP(Reisen et al., 2003), PIP-GFP (a gift of C. Hawes), VHAa1-GFP(Dettmer et al., 2006), and N-YFP-HDEL (Teh and Moore, 2007) intogsh2-5 or rml1 were performed by crossing previously describedtransgenic Arabidopsis homozygous for the fluorescent proteinmarkers with plants heterozygous for gsh2-5 or rml1. Constructs ofRTNLB1-YFP and DER2-YFP were made available by L. Frigerio,University of Warwick, UK; mCHERRY-HDEL (Nelson et al., 2007)was obtained from the Arabidopsis Biological Resource Centre(ABRC), Columbus, OH, USA. The constructs were transformed intoAgrobacterium tumefaciens and subsequently to Arabidopsis bythe floral dipping method (Clough and Bent, 1998). Seeds weresterilized with 70% ethanol, stratified at 4�C for 2 days on agar platescontaining MS medium (pH 5.7) with 1% sucrose and cultivated at20�C under a 16-h photoperiod.
Genetic screen
Seeds from transgenic Arabidopsis lines expressing secGFP (lineS76; Zheng et al., 2004), nlsRm-2A-secGf, or STN-Rm-2A-secGf
(Samalova et al., 2006) were mutagenized with ethane-methyl-sul-fonate (EMS) as described (Teh and Moore, 2007) and seed wascollected separately from over 7000 individual M1 plants. Three-day-old seedlings from 7000 M2 families, germinated and grown onMS agar, were screened for enhanced GFP fluorescence using aLeica MZFLIII Stereo Microscope (Leica Microsystems (UK) Ltd,Milton Keynes, Bucks, UK). Initially, seedlings were screened inpools comprising approximately 30 seeds from each of 10 M2
families. Individual M2 families from pools segregating fluorescentseedlings were subsequently rescreened to identify the segregatingM2 family. Intracellular accumulation of secGFP in cytoplasmicorganelles of the mutant seedlings was confirmed by confocalmicroscopy. The screen identified 97 independent putative mu-tants, and 73 of these could be propagated to the next generationvia their phenotypically WT heterozygous siblings. Initially eightphenotypically WT plants were propagated for each mutant family;if no heterozygote was found, an additional eight were tested.Heterozygous M3 or M4 plants were backcrossed to Col-0 andcrossed to Ler for approximate mapping by bulk segregant analysis(BSA) (Lukowitz et al., 2000). To do this, four phenotypically WTplants from each mutant family were selected for crossing and theirheterozygosity was subsequently assessed by scoring their selfedprogeny for mutant phenotypes; another four plants were used ifnone of the initial set were heterozygotes. Following BSA, four ofthe mutants that mapped to the top of ChrV and each exhibitedsimilar secGFP accumulation phenotypes were selected for detailedcharacterization.
Electron microscopy
Ten-day old seedlings were fixed in 2% paraformaldehyde and 1%glutaraldehyde in 20 mM K2HPO4 buffer (pH 7.0) for 3–5 h. Sampleswere washed with 50 mM K2HPO4 solution before transferring to 2%osmium tetroxide for 2 h. After washing with distilled water, thesamples were dehydrated in an ethanol series and embedded in
Arabidopsis glutathione synthase mutant and ER morphology 891
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894

Spurr’s resin. Sections were examined with a Hitachi H7650 trans-mission electron microscope (Hitachi High-Technologies EuropeGmbH, Maidenhead, Berks, UK).
Confocal laser scanning microscopy
Except for simultaneous imaging of GFP and YFP, samples wereexamined using Zeiss LSM 510 META laser scanning microscope(Carl Zeiss Ltd, Welwyn Garden City, Herts, UK). The configurationof the instrument was described previously (Samalova et al., 2006;Teh and Moore, 2007; Zheng et al., 2004) except for imagingmCHERRY, bimane and c-roGFP1. The mCHERRY was excited with a543 nm HeNe laser and a HFT 458/543 primary dichroic mirror, andwas detected with a 565–615 nm band pass filter. Bimane wasexcited with a 405 nm blue diode laser and a HFT 458/543 primarydichroic mirror, and was detected with a 475–525 nm band passfilter. For simultaneous imaging of mCHERRY and bimane, themulti-track line-sequential imaging mode with the configurationmentioned above was used. Images were processed using ZeissLSM Image Browser.
In the time course experiment with c-roGFP1, the perfusionchamber was connected to a Gilson Minipuls 2 peristaltic pump(Gilson Inc., Middleton, WI, USA) with HPLC tubing and wasmounted on the confocal microscope. Imaging and ratiometricanalysis of roGFPs was performed as described (Schwarzlanderet al., 2008). Samples coexpressing GFP and YFP were examinedusing Leica TCS SP5 II. The GFP was excited with the 458 nm argonlaser line and was detected with a PMT channel set to collect signalbetween 475 and 510 nm. The YFP was subsequently excited withthe 514 nm argon laser line and was detected with a second PMTchannel set to collect signal between 572 and 613 nm. Images wereprocessed using LAS AF (Leica Microsystems (UK) Ltd).
Fluorescent dye treatment
Seedlings were immersed in 5 lM FM4-64 (Invitrogen Ltd, Paisley,UK; from 5 mM stock in water) aqueous solution for 60 min to labelthe tonoplast. Seedlings were immersed in 100 lM MCB or MBB(Invitrogen; from 100 mM stock in ethanol) aqueous solution.
Protein domain analysis
Protein sequences of GSH2 from Arabidopsis, human (Homo sapi-ens) and yeast (Saccharomyces cerevisiae) were obtained fromGenBank (Arabidopsis, CAB51027.1; human, NP_000169.1; yeast,CAA 74136.1) and were aligned using CLUSTALW (http://www.e-bi.ac.uk/Tools/clustalw2/) to assess the conservation of eukaryoticGSH2. Putative conserved domains of Arabidopsis GSH2 wereidentified using the Basic Local Alignment Sequence Tool program(BLAST; http://blast.ncbi.nlm.nih.gov/).
Low-molecular-weight thiol molecule analysis
Low-molecular-weight thiols were extracted from 10-day-old seed-lings and analyzed by HPLC as described previously (Cairns et al.,2006).
Immunoblot analysis
For the secGFP immunoblot, proteins from 6-day-old seedlingswere extracted and analyzed by immunoblot as described(Samalova et al., 2006). For the unfolded protein response analysis,10-day-old seedlings were incubated in half-strength MS mediumwith either DMSO or 5 lg ml)1 tunicamycin (Sigma-Aldrich, Poole,UK). Proteins were extracted first by homogenization in twovolumes of extraction buffer: 4% SDS, 100 mM 2-amino-2-(hydrox-ymethyl)-1,3-propanediol (TRIS)-HCl (pH 6.8), 400 lg ml)1 brom-
ophenol blue, 200 mM DTT, 20% glycerol, 0.2% b-mercaptoethanol,and protease inhibitor cocktail (Sigma-Aldrich). Samples were thenboiled for 10 min followed by centrifugation at 10 000 g for 5 min.Then SDS-PAGE was run as described (Samalova et al., 2006).Proteins were then blotted onto a polyvinylidene fluoride mem-brane as described (Batoko et al., 2000). Before incubating withprimary antibody, the blot was stained with Ponceau S solution(0.5% Ponceau S and 1% acetic acid) for 5 min and subsequentlywashed with 1% acetic acid. An image of the blot was taken with aNikon D700 camera (Nikon UK Ltd, Kingston upon Thames, Surrey,UK) before Ponceau S was de-stained with 1% acetic acid. Anti-BiP(a gift from L. Frigerio, University of Warwick, UK) and anti-CRT (agift from J. Denecke, University of Leeds, UK) were diluted to 1:5000and 1:10 000, respectively. Alkaline phosphatase-conjugated sec-ondary antibody (anti-rabbit IgG) (Sigma-Aldrich) was used at1:10 000 dilution.
ACKNOWLEDGEMENTS
We thank Markus Schwarzlander, University of Oxford, for assis-tance on roGFP analysis and comments on the manuscript, LorenzoFrigerio, University of Warwick, for the anti-BiP antibody, RTNLB1-YFP and DER2-YFP constructs, Jurgen Denecke, University of Leeds,for the anti-CRT antibody, Barry Martin and Chris Hawes, OxfordBrookes University for assistance with electron microscopy and forPIP-GFP seeds, and Rudiger Hell, Heidelberg University, Germanyfor access to HPLC. KKCA was funded by the Clarendon Fund andLinacre Canadian Alumni Scholarship. This work was supported bygrant BBS/B/09562 from the UK Biotechnology and Biological Sci-ences Research Council.
SUPPORTING INFORMATION
Additional supporting information may be found in the online ver-sion of this article:Figure S1. GFP-HDEL and DER2-YFP labeling in N37 (gsh2-5).Figure S2. Domain structure of Arabidopsis glutathione synthase.Figure S3. Monobromobimane and M monochlorobimane labelingof small thiols in root epidermal cells.Figure S4. Buthionine sulfoximine-treated gsh2-5 root cells.Figure S5. RTNLB1-YFP and NEM treatment in N37 (gsh2-5).Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Agee, A.E., Surpin, M., Sohn, E.J. et al. (2010) MODIFIED VACUOLE PHENO-
TYPE1 is an arabidopsis myrosinase-associated protein involved in endo-
membrane protein trafficking. Plant Physiol. 152(1), 120–132.
Ball, L., Accotto, G.P., Bechtold, U. et al. (2004) Evidence for a direct link
between glutathione biosynthesis and stress defense gene expression in
Arabidopsis. Plant Cell, 16(9), 2448–2462.
Banhegyi, G., Benedetti, A., Csala, M. and Mandl, J. (2007) Stress on redox.
FEBS Lett. 581(19), 3634–3640.
Banhegyi, G., Lusini, L., Puskas, F., Rossi, R., Fulceri, R., Braun, L., Mile, V., di
Simplicio, P., Mandl, J. and Benedetti, A. (1999) Preferential transport of
glutathione versus glutathione disulfide in rat liver microsomal vesicles.
J. Biol. Chem. 274(18), 12213–12216.
Batoko, H., Zheng, H., Hawes, C. and Moore, I. (2000) A Rab1 GTPase is
required for transport between the endoplasmic reticulum and golgi
apparatus and for normal golgi movement in plants. Plant Cell, 12(11),
2201–2218.
Bednarek, S.Y. and Falbel, T.G. (2002) Membrane trafficking during plant
cytokinesis. Traffic, 3(9), 621–629.
892 Kenneth K. C. Au et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894

Behnke, H.-. and Eschlbeck, G. (1978) Dilated cisternae in capparales: an
attempt towards the characterization of a specific endoplasmic reticulum.
Protoplasma, 97(4), 351–363.
Berna, G., Robles, P. and Micol, J.L. (1999) A mutational analysis of leaf
morphogenesis in Arabidopsis thaliana. Genetics, 152(2), 729–742.
Bertolotti, A., Zhang, Y., Hendershot, L.M., Harding, H.P. and Ron, D. (2000)
Dynamic interaction of BiP and ER stress transducers in the unfolded
protein response. Nat. Cell Biol. 2(6), 326–332.
Boevink, P., Oparka, K., Cruz, S.S., Martin, B., Betteridge, A. and Hawes, C.
(1998) Stacks on tracks: the plant golgi apparatus traffics on an actin/ER
network? Plant J. 15(3), 441–447.
Bolte, S., Talbot, C., Boutte, Y., Catrice, O., Read, N.D. and Satiat-Jeune-
maitre, B. (2004) FM-dyes as experimental probes for dissecting vesicle
trafficking in living plant cells. J. Microsc. 214(2), 159–173.
Cairns, N.G., Pasternak, M., Wachter, A., Cobbett, C.S. and Meyer, A.J. (2006)
Maturation of Arabidopsis seeds is dependent on glutathione biosynthesis
within the embryo. Plant Physiol. 141(2), 446–455.
Chakravarthi, S., Jessop, C.E. and Bulleid, N.J. (2006) The role of glutathione
in disulphide bond formation and endoplasmic-reticulum-generated oxi-
dative stress. EMBO Rep. 7(3), 271–275.
Chow, C., Neto, H., Foucart, C. and Moore, I. (2008) Rab-A2 and Rab-A3
GTPases define a trans-golgi endosomal membrane domain in Arabidopsis
that contributes substantially to the cell plate. Plant Cell, 20(1), 101–123.
Christensen, A., Svensson, K., Persson, S., Jung, J., Michalak, M., Widell, S.
and Sommarin, M. . (2008) Functional characterization of Arabidopsis
Calreticulin1a: a key alleviator of endoplasmic reticulum stress. Plant Cell
Physiol. 49(6), 912–924.
Clough, S.J. and Bent, A.F. (1998) Floral dip: a simplified method for Agro-
bacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16(6),
735–743.
Cobbett, C.S., May, M.J., Howden, R. and Rolls, B. (1998) The glutathione-
deficient, cadmium-sensitive mutant, cad2-1, of Arabidopsis thaliana is
deficient in gamma-glutamylcysteine synthetase. Plant J. 16(1), 73–78.
Costa, M.D.L., Reis, P.A.B., Valente, M.A.S., Irsigler, A.S.T., Carvalho, C.M.,
Loureiro, M.E., Aragao, F.J.L., Boston, R.S., Fietto, L.G. and Fontes, E.P.B..
(2008) A new branch of endoplasmic reticulum stress signaling and the
osmotic signal converge on plant-specific asparagine-rich proteins to
promote cell death. J. Biol. Chem. 283(29), 20209–20219.
Crofts, A.J., Leborgne-Castel, N., Hillmer, S., Robinson, D.G., Phillipson, B.,
Carlsson, L.E., Ashford, D.A. and Denecke, J. (1999) Saturation of the
endoplasmic reticulum retention machinery reveals anterograde bulk flow.
Plant Cell, 11(11), 2233–2248.
Cutler, S.R., Ehrhardt, D.W., Griffitts, J.S. and Somerville, C.R. (2000) Random
GFP-cDNA fusions enable visualization of subcellular structures in cells of
Arabidopsis at a high frequency. Proc. Natl Acad. Sci. 97(7), 3718–3723.
Dettmer, J. and Friml, J. (2011) Cell polarity in plants: when two do the same,
it is not the same. Curr. Opin. Cell Biol. 23(6), 686–696.
Dettmer, J., Hong-Hermesdorf, A., Stierhof, Y. and Schumacher, K. (2006)
Vacuolar H+-ATPase activity is required for endocytic and secretory traf-
ficking in Arabidopsis. Plant Cell, 18(3), 715–730.
Dubreuil-Maurizi, C., Vitecek, J., Marty, L., Branciard, L., Frettinger, P.,
Wendehenne, D., Meyer, A.J., Mauch, F. and Poinssot, B. (2011) Glutathi-
one deficiency of the Arabidopsis mutant pad2-1 affects oxidative stress-
related events, defense gene expression, and the hypersensitive response.
Plant Physiol. 157(4), 2000–2012.
Ebine, K., Fujimoto, M., Okatani, Y. et al. (2011) A membrane trafficking
pathway regulated by the plant-specific RAB GTPase ARA6. Nat. Cell Biol.
13(7), 853–859.
Fahey, R.C., Newton, G.L., Dorian, R. and Kosower, E.M. (1981) Analysis of
biological thiols: quantitative determination of thiols at the picomole level
based upon derivatization with monobromobimanes and separation by
cation-exchange chromatography. Anal. Biochem. 111(2), 357–365.
Faso, C., Chen, Y.N., Tamura, K. et al. (2009) A missense mutation in the
Arabidopsis COPII coat protein Sec24A induces the formation of clusters of
the endoplasmic reticulum and Golgi apparatus. Plant Cell, 21(11), 3655–
3671.
Fricker, M.D., May, M., Meyer, A.J., Sheard, N. and White, N.S. (2000) Mea-
surement of glutathione levels in intact roots of Arabidopsis. J. Microsc.
198, (Pt 3) 162–173.
Fuji, K., Shimada, T., Takahashi, H., Tamura, K., Koumoto, Y., Utsumi, S.,
Nishizawa, K., Maruyama, N. and Hara-Nishimura, I. (2007) Arabidopsis
vacuolar sorting mutants (green fluorescent seed) can be identified effi-
ciently by secretion of vacuole-targeted green fluorescent protein in their
seeds. Plant Cell, 19(2), 597–609.
Griffith, O.W. and Meister, A. (1979) Potent and specific inhibition of gluta-
thione synthesis by buthionine sulfoximine (S-n-butyl homocysteine sul-
foximine). J. Biol. Chem. 254, 7558–7560.
Hawes, C., Saint-Jore, C., Martin, B. and Zheng, H.Q. (2001) ER confirmed as
the location of mystery organelles in Arabidopsis plants expressing GFP!.
Trends Plant Sci. 6(6), 245–246.
Hell, R. and Bergmann, L. (1990) k-Glutamylcysteine synthetase in higher
plants: catalytic properties and subcellular localization. Planta, 180(4),
603–612.
Herrera, K., Cahoon, R.E., Kumaran, S. and Jez, J.M. (2007) Reaction mecha-
nism of glutathione synthetase from arabidopsis thaliana. J. Biol. Chem.
282(23), 17157–17165.
Huotari, J. and Helenius, A. (2011) Endosome maturation. EMBO J. 30(17),
3481–3500.
Hwang, C., Sinskey, A.J. and Lodish, H.F. (1992) Oxidized redox state of glu-
tathione in the endoplasmic reticulum. Science, 257(5076), 1496–1502.
Iversen, T. (1970) The morphology, occurrence, and distribution of dilated
cisternae of the endoplasmic reticulum in tissues of plants of the Crucife-
rae. Protoplasma, 71(4), 467–477.
Iwata, Y. and Koizumi, N. (2005) An Arabidopsis transcription factor,
AtbZIP60, regulates the endoplasmic reticulum stress response in a man-
ner unique to plants. Proc. Natl Acad. Sci. USA, 102(14), 5280–5285.
Jessop, C.E. and Bulleid, N.J. (2004) Glutathione directly reduces an oxido-
reductase in the endoplasmic reticulum of mammalian cells. J. Biol. Chem.
279(53), 55341–55347.
Jobe, T.O., Sung, D.-Y., Akmakjian, G., Pham, A., Konives, E.A., Mendoza-
Cozatl, D.G. and Schroeder, J.I. (2012) Feedback inhibition by thiols
outranks glutathione depletion: a luciferase-based screen reveals gluta-
thione-deficient c-ECS and glutathione synthetase mutants impaired in
cadmium-induced sulfate assimilation. Plant J. 70, 783–795.
Jurgens, G., Mayer, U., Ramon, A.T.R., Berleth, T. and Misera, S. (1991) Ge-
netic analysis of pattern formation in the Arabidopsis embryo. Develop-
ment, 113, (Supplement 1) 27–38.
Kawagoe, Y., Suzuki, K., Tasaki, M., Yasuda, H., Akagi, K., Katoh, E., Nishiz-
awa, N.K., Ogawa, M. and Takaiwa, F. (2005) The critical role of disulfide
bond formation in protein sorting in the endosperm of rice. Plant Cell,
17(4), 1141–1153.
Kienle, N., Kloepper, T. and Fasshauer, D. (2009) Phylogeny of the SNARE
vesicle fusion machinery yields insights into the conservation of the
secretory pathway in fungi. BMC Evol. Biol. 9(1), 19.
Koizumi, N., Martinez, I.M., Kimata, Y., Kohno, K., Sano, H. and Chrispeels,
M.J. (2001) Molecular characterization of two Arabidopsis Ire1 homologs,
endoplasmic reticulum-located transmembrane protein kinases. Plant
Physiol. 127(3), 949–962.
Le Gall, S., Neuhof, A. and Rapoport, T. (2004) The endoplasmic reticu-
lum membrane is permeable to small molecules. Mol. Biol. Cell 15(2), 447–
455.
Liu, J. and Howell, S.H. (2010) Endoplasmic reticulum protein quality control
and its relationship to environmental stress responses in plants. Plant Cell,
22(9), 2930–2942.
Lohman, J.R. and Remington, S.J. (2008) Development of a family of redox-
sensitive green fluorescent protein indicators for use in relatively oxidizing
subcellular environments. Biochemistry, 47(33), 8678–8688.
Lukowitz, W., Gillmor, C.S. and Scheible, W.R. (2000) Positional cloning in
Arabidopsis. why it feels good to have a genome initiative working for you.
Plant Physiol. 123(3), 795–805.
Marty, L., Siala, W., Schwarzlander, M., Fricker, M.D., Wirtz, M., Sweetlove,
L.J., Meyer, Y., Meyer, A.J., Reichheld, J. and Hell, R. (2009) The NADPH-
dependent thioredoxin system constitutes a functional backup for cytosolic
glutathione reductase in Arabidopsis. Proc. Natl Acad. Sci. 106(22), 9109–
9114.
Matsushima, R., Hayashi, Y., Yamada, K., Shimada, T., Nishimura, M. and
Hara-Nishimura, I. (2003) The ER body, a novel endoplasmic reticulum-
derived structure in Arabidopsis. Plant Cell Physiol. 44(7), 661–666.
Meyer, A.J. (2008) The integration of glutathione homeostasis and redox
signaling. J. Plant Physiol. 165(13), 1390–1403.
Meyer, A.J., May, M.J. and Fricker, M. (2001) Quantitative in vivo measure-
ment of glutathione in Arabidopsis cells. Plant J. 27(1), 67–78.
Arabidopsis glutathione synthase mutant and ER morphology 893
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894

Muller, I., Wagner, W., Volker, A., Schellmann, S., Nacry, P., Kuttner, F.,
Schwarz-Sommer, Z., Mayer, U. and Jurgens, G. (2003) Syntaxin specificity
of cytokinesis in Arabidopsis. Nat. Cell Biol. 5(6), 531–534.
Nakano, R.T., Matsushima, R., Ueda, H., Tamura, K., Shimada, T., Li, L.,
Hayashi, Y., Kondo, M., Nishimura, M. and Hara-Nishimura, I. (2009)
GNOM-LIKE1/ERMO1 and SEC24a/ERMO2 are required for maintenance of
endoplasmic reticulum morphology in Arabidopsis thaliana. Plant Cell,
21(11), 3672–3685.
Nelson, B.K., Cai, X. and Nebenfuhr, A. (2007) A multicolored set of in vivo
organelle markers for co-localization studies in Arabidopsis and other
plants. Plant J. 51(6), 1126–1136.
Noctor, G., Veljovic-Jovanovic, S. and Foyer, C.H. (2000) Peroxide processing
in photosynthesis: antioxidant coupling and redox signalling. Philos.
Trans. R. Soc. Lond. B Biol. Sci. 355(1402), 1465–1475.
Novick, P. and Schekman, R. (1979) Secretion and cell-surface growth are
blocked in a temperature-sensitive mutant of saccharomyces cerevisiae.
Proc. Natl. Acad. Sci. USA, 76(4), 1858–1862.
Nziengui, H., Bouhidel, K., Pillon, D., Der, C., Marty, F. and Schoefs, B. (2007)
Reticulon-like proteins in arabidopsis thaliana: structural organization and
ER localization. FEBS Lett. 581(18), 3356–3362.
Oertle, T., Klinger, M., Stuermer, C.A. and Schwab, M.E. (2003) A reticular
rhapsody: phylogenic evolution and nomenclature of the RTN/Nogo gene
family. FASEB J. 17(10), 1238–1247.
Onda, Y., Kumamaru, T. and Kawagoe, Y. (2009) ER membrane-localized
oxidoreductase Ero1 is required for disulfide bond formation in the rice
endosperm. Proc. Nat. Acad. Sci. 106(33), 14156–14161.
Orsi, A., Sparvoli, F. and Ceriotti, A. (2001) Role of individual disulfide bonds
in the structural maturation of a low molecular weight glutenin subunit. J.
Biol. Chem. 276(34), 32322–32329.
Parisy, V., Poinssot, B., Owsianowski, L., Buchala, A., Glazebrook, J. and
Mauch, F. (2007) Identification of PAD2 as a c-glutamylcysteine synthetase
highlights the importance of glutathione in disease resistance of Arabid-
opsis. Plant J. 49(1), 159–172.
Pasternak, M., Lim, B., Wirtz, M., Hell, R., Cobbett, C.S. and Meyer, A.J. (2008)
Restricting glutathione biosynthesis to the cytosol is sufficient for normal
plant development. Plant J. 53(6), 999–1012.
Pinheiro, H., Samalova, M., Geldner, N., Chory, J., Martinez, A. and Moore, I.
(2009) Genetic evidence that the higher plant Rab-D1 and Rab-D2 GTPases
exhibit distinct but overlapping interactions in the early secretory pathway.
J. Cell Sci. 122(20), 3749–3758.
Pompa, A. and Vitale, A. (2006) Retention of a bean phaseolin/maize gamma-
zein fusion in the endoplasmic reticulum depends on disulfide bond for-
mation. Plant Cell, 18(10), 2608–2621.
Reisen, D., Leborgne-Castel, N., Ozalp, C., Chaumont, F. and Marty, F. (2003)
Expression of a cauliflower tonoplast aquaporin tagged with GFP in to-
bacco suspension cells correlates with an increase in cell size. Plant Mol.
Biol. 52(2), 387–400.
Richter, S., Geldner, N., Schrader, J., Wolters, H., Stierhof, Y., Rios, G., Koncz,
C., Robinson, D.G. and Jurgens, G. (2007) Functional diversification of
closely related ARF-GEFs in protein secretion and recycling. Nature,
448(7152), 488–492.
Ridge, R.W., Uozumi, Y., Plazinski, J., Hurley, U.A. and Williamson, R.E. (1999)
Developmental transitions and dynamics of the cortical ER of arabidopsis
cells seen with green fluorescent protein. Plant Cell Physiol. 40(12), 1253–
1261.
Rojo, E., Gillmor, C.S., Kovaleva, V., Somerville, C.R. and Raikhel, N.V. (2001)
VACUOLELESS1 is an essential gene required for vacuole formation and
morphogenesis in Arabidopsis. Dev. Cell 1(2), 303–310.
Samalova, M., Fricker, M. and Moore, I. (2006) Ratiometric fluorescence-
imaging assays of plant membrane traffic using polyproteins. Traffic, 7(12),
1701–1723.
Sanderfoot, A. (2007) Increases in the number of SNARE genes parallels the
rise of multicellularity among the green plants. Plant Physiol. 144(1), 6–17.
Schwarzlander, M., Fricker, M.D., Muller, C., Marty, L., Brach, T., Novak, J.,
Sweetlove, L.J., Hell, R. and Meyer, A.J. (2008) Confocal imaging of glu-
tathione redox potential in living plant cells. J. Microsc. 231(2), 299–316.
Shanmugam, V., Tsednee, M. and Yeh, K. (2012) ZINC TOLERANCE INDUCED
BY IRON 1 reveals the importance of glutathione in the cross-homeostasis
between zinc and iron in Arabidopsis thaliana. Plant J. 69(6), 1006–1017.
Sparkes, I., Tolley, N., Aller, I., Svozil, J., Osterrieder, A., Botchway, S., Mu-
eller, C., Frigerio, L. and Hawes, C. (2010) Five Arabidopsis reticulon iso-
forms share endoplasmic reticulum location, topology, and membrane-
shaping properties. Plant Cell, 22(4), 1333–1343.
Surpin, M., Zheng, H., Morita, M.T. et al. (2003) The VTI family of SNARE
proteins is necessary for plant viability and mediates different protein
transport pathways. Plant Cell, 15(12), 2885–2899.
Tanaka, H., Kitakura, S., De Rycke, R., De Groodt, R. and Friml, J. (2009)
Fluorescence imaging-based screen identifies ARF GEF component of early
endosomal trafficking. Curr. Biol. 19(5), 391–397.
Teh, O. and Moore, I. (2007) An ARF-GEF acting at the golgi and in selective
endocytosis in polarized plant cells. Nature, 448(7152), 493–496.
Tolley, N., Sparkes, I.A., Hunter, P.R., Craddock, C.P., Nuttall, J., Roberts,
L.M., Hawes, C., Pedrazzini, E. and Frigerio, L. (2008) Overexpression of a
plant reticulon remodels the lumen of the cortical endoplasmic reticulum
but does not perturb protein transport. Traffic, 9(1), 94–102.
Toyooka, K., Okamoto, T. and Minamikawa, T. (2000) Mass transport of
proform of a KDEL-tailed cysteine proteinase (sh-EP) to protein storage
vacuoles by endoplasmic reticulum–derived vesicle is involved in protein
mobilization in germinating seeds. The Journal of Cell Biology, 148(3), 453–
464.
Ueda, T., Yamaguchi, M., Uchimiya, H. and Nakano, A. (2001) Ara6, a plant-
unique novel type Rab GTPase, functions in the endocytic pathway of
arabidopsis thaliana. EMBO J. 20(17), 4730–4741.
Vernoux, T., Wilson, R.C., Seeley, K.A. et al. (2000) The ROOT MERISTEM-
LESS1/CADMIUM SENSITIVE2 gene defines a glutathione-dependent
pathway involved in initiation and maintenance of cell division during
postembryonic root development. Plant Cell, 12(1), 97–110.
Voeltz, G.K., Prinz, W.A., Shibata, Y., Rist, J.M. and Rapoport, T.A. (2006) A
class of membrane proteins shaping the tubular endoplasmic reticulum.
Cell, 124(3), 573–586.
West, M., Zurek, N., Hoenger, A. and Voeltz, G.K. (2011) A 3D analysis of yeast
ER structure reveals how ER domains are organized by membrane curva-
ture. The Journal of Cell Biology, 193(2), 333–346.
Woollard, A.A. and Moore, I. (2008) The functions of Rab GTPases in plant
membrane traffic. Curr. Opin. Plant Biol. 11(6), 610–619.
Yamada, K., Nagano, A.J., Nishina, M., Hara-Nishimura, I. and Nishimura,
M. (2008) NAI2 is an endoplasmic reticulum body component that en-
ables ER body formation in Arabidopsis thaliana. Plant Cell, 20(9), 2529–
2540.
Zechmann, B., Mauch, F., Sticher, L. and Muller, M. (2008) Subcellular
immunocytochemical analysis detects the highest concentrations of glu-
tathione in mitochondria and not in plastids. J. Exp. Bot. 59(14), 4017–
4027.
Zheng, H., Kunst, L., Hawes, C. and Moore, I. (2004) A GFP-based assay re-
veals a role for RHD3 in transport between the endoplasmic reticulum and
Golgi apparatus. Plant J. 37(3), 398–414.
894 Kenneth K. C. Au et al.
ª 2012 The AuthorsThe Plant Journal ª 2012 Blackwell Publishing Ltd, The Plant Journal, (2012), 71, 881–894