the nutritional status modulates preservation-reperfusion injury in rat fatty liver
TRANSCRIPT

Journal of Surgical Research 127, 190–196 (2005)
The Nutritional Status Modulates Preservation-Reperfusion Injury in RatFatty Liver
Paolo Caraceni,*,†,1 Marco Domenicali,*,† Anna Maria Pertosa,*,† Elisabetta Maiolini,* Ignazio Grattagliano,‡Alessandro Principe,*,† Giuseppe Palasciano,‡ Franco Trevisani,* and Mauro Bernardi*,†
*Department of Internal Medicine, Cardioangiology, and Hepatology, University of Bologna, Bologna, Italy; †Center for BiomedicalApplied Research (C.R.B.A.), University of Bologna, Bologna, Italy; and ‡Department of Internal Medicine and Public Health, University
of Bari, Bari, Italy
Submitted for publication October 28, 2004
doi:10.1016/j.jss.2005.02.018
Background. Microcirculation disturbances are es-sential factors of preservation injury in fatty liver.However, hepatocyte injury is also markedly excessivein fatty liver resulting, at least in part, from energymetabolism impairment and oxidative stress. Thus,this study aimed to determine whether nutritional sta-tus influences preservation injury in fatty liver andwhether energetic substrate supplementation, aloneor with a vasodilator, is protective.
Materials and methods. Normal or fatty livers in-duced by a choline-deficient diet were isolated fromfed and fasted rats, preserved in University of Wiscon-sin solution at 4°C for 18 h, and then reperfused withKrebs–Henseleit solution at 37°C for 120 min. Fastedrats with fatty liver were also treated as follows: (1)Glucose supplementation: rats had access to a glucosesolution for 18 h prior procurement; (2) Prostaglandin(PG): alprostadil was continuously infused duringreperfusion; (3) Combined treatment: Glucose supple-mentation � PG.
Results. Fasting-induced liver injury was signifi-cantly greater in fatty than normal liver. In fatty liversfrom fasted rats, all treatments reduced the alanineaminotransaminase release. Hepatic oxygen consump-tion improved in the glucose and glucose � PG groups,while PG infusion had no effect. Glucose supplemen-tation did not affect portal pressure, which, in con-trast, was reduced in livers receiving PG. Finally, alltreatments lowered oxidative injury.
Conclusions. Preservation injury in fatty liver isgreatly related to nutritional status. Energetic sub-strate supplementation may represent a clinically fea-
1 To whom correspondence and reprint requests should be ad-dressed at Dipartimento di Medicina Interna, Cardioangiologia,Epatologia, University of Bologna, Policlinico Sant’Orsola, Via Al-
bertoni 15, 40138 Bologna, Italy. E-mail: [email protected].1900022-4804/05 $30.00© 2005 Elsevier Inc. All rights reserved.
sible protective strategy and a multistep approachadding vasodilators could offer further benefit by act-ing on different pathogenetic mechanisms. © 2005
Elsevier Inc. All rights reserved.
Key Words: fatty liver; experimental liver transplan-tation; preservation injury; reperfusion injury; nutri-tional status; prostaglandins.
INTRODUCTION
Fatty liver is the most frequent alteration of the liverfound in the general population and its prevalencereaches about 25% of the potential donors for ortho-topic liver transplantation and living donor liver trans-plantation. The presence of fatty degeneration reducesthe tolerance of the liver to the ischemia-reperfusioninjury invariably associated with these surgical proce-dures, leading to higher mortality and postoperativecomplication rates. Thus, albeit the persistent short-age of donor organs, most centers do not transplantlivers with moderate-massive fatty infiltration [1].
Impaired microcirculation has been proposed as thekey event in the pathogenesis of preservation-reperfusion injury in steatotic livers [1]. The sinusoidalblood flow is greatly reduced after transplantation offatty livers as the result of the narrowed sinusoidallumen, which is compressed by the swollen fat-loadedhepatocytes and partially or completely obstructed bycell debris and platelets and leukocytes adherent to theendothelium [1–3]. Thus, it is not surprising that sub-stances with vasodilator and antiinflammatory proper-ties, such as prostaglandins, have been found to beprotective in animal models of fatty liver transplanta-tion [4, 5].
However, other evidences indicate that hepatocyte

191CARACENI ET AL.: PRESERVATION INJURY IN FATTY LIVER
injury is also markedly excessive in fatty liver exposedto cold preservation compared to that observed in nor-mal organs [5]. Several potential events have beensuggested as mediators of this parenchymal cell injuryincluding oxidative stress [6], mitochondrial dysfunc-tion [7], and energy metabolism impairment [8].
The preexisting nutritional status is a major deter-minant of hepatocyte injury associated with ischemia-reperfusion [9]. In normal livers, it is universally ac-cepted that fasting exacerbates warm ischemic injury[9, 10]. Less homogenous results have been obtained ifnormal livers are exposed to cold ischemia; when fast-ing is prolonged up to 4 days, the starved livers appearcapable of tolerating the transplant procedure betterthan do livers obtained from fed donors [11, 12]. Nev-ertheless, preharvesting nutritional repletion by glu-cose supplementation improves survival and liverfunction in both fasted and fed animals [13, 14].
Conversely, the data on the effect of nutritional sta-tus in fatty livers subjected to ischemia-reperfusion arevery limited. We previously reported that the preexist-ing nutritional status of the animal adversely affectswarm ischemia-reperfusion injury in fatty livers muchmore than it does in normal livers [15]. A preliminaryreport suggested that fasting rats with fatty liver for 4days reduces early reperfusion injury after cold isch-emia likely by suppressing Kupffer cell function [16].However, long-term starvation is not applicable in thesetting of clinical transplantation and the effect of ashorter period of fasting has never been tested thus far.Thus, no clear data exist on the relation between fattyliver, preservation injury, and nutritional status.
Therefore, the aim of this study was to compare in amodel of isolated perfused rat liver the influence ofanimal starvation on cold ischemia-reperfusion injuryexperienced by normal and fatty liver induced by acholine-deficient diet. Furthermore, since the alter-ation of the nutritional status is a common condition ofpotential donors staying in intensive care units forseveral days and the beneficial effect of a nutritionalsupport has been already demonstrated in normal liv-ers exposed to cold ischemia [13, 14], experiments werealso designed to determine whether administration ofenergetic substrates, alone or with a vasodilator, isprotective against preservation injury in fatty liverfrom fasted rats.
MATERIALS AND METHODS
Animals and Induction of Fatty Liver
Male Sprague Dawley rats (Charles River, Calco, LC, Italy),weighing 150–175 g, were allowed to acclimate to the animal quar-ters and were given free access to a standard chow diet and water for1 week. Then, rats were fed a choline-deficient diet (Dyets Inc.,Bethlehem, PA) for an additional 5 days. With this procedure, theliver of rats fed the choline-deficient diet presented a moderate
steatosis, predominantly macrovesicular and involving about 50-60%of the hepatocytes, without evidence of inflammation and/or fibrosis[15]. Triglycerides are the main component of accumulated lipidswith an increased molar percentage of palmitic and oleic acids as itoccurs in fatty liver developing in humans for an excessive dietaryintake of carbohydrates [17]. Prior to sacrifice, the animal weightwas similar in the two groups ranging between 230 and 280 g.
All procedures involving rats were conducted according to theguidelines for the care and use of laboratory animals approved by ourInstitution.
Procurement and Preservation of the Liver
The rats were anesthetized with enflurane (Abbott Laboratories,Chicago, IL); the abdomen was opened with a midline incision andthe portal vein was cannulated with a 16-G angiocath. The liver wasflushed out with 20 ml of the University of Wisconsin (UW) solutionat 4°C and then immediately removed. After 18 h of preservation inthe UW solution at 4°C, all livers were exposed to room temperatureon a Petri dish for 15 min prior to reperfusion to mimic the period ofrewarming during surgical implantation in vivo. The organs werethen rinsed with 10 ml of a Krebs–Henseleit bicarbonate (KHB)solution and connected to the perfusion circuit.
Isolated Perfused Rat Liver Model
The livers were perfused in a non-recirculating fashion by using aperistaltic pump for 120 min at 37°C with a KHB buffer containing5 mM glucose and 0.5% serum bovine albumin within a thermostat-ically controlled plexiglas cabinet designed to perform two perfusionssimultaneously (Ditta Disa, Milan, Italy) [8]. The KHB solution wasexposed to a gas mixture of 95% O2 and 5% CO2 in a pressuredmembrane oxygenator, which generated a partial pressure of O2
greater than 500 mm Hg. The organs were initially perfused at a flowrate of 0.5 ml/min/g liver and then increased progressively up to 3.0ml/min/g liver in a 5-min period to avoid barotrauma. The portalpressure was continuously monitored through a three-way stopcockplaced before the portal vein cannula and expressed as cm H2O.Samples were taken from a portal inflow port and from the effluentperfusate for measurement of the alanine aminotransaminase (ALT)level and oxygen consumption. In most of the experiments, livertissue was also collected for the biochemical determinations beforeexplantation (basal conditions) and at the end of reperfusion.
Study Design
The day before the experiment, both the rats with fatty liver andthe rats with normal liver were further divided as follows: (a) ratswith free access to food and water until the time of surgery (fedgroups) (n � 6–8); and (b) rats with access only to water for the 18 hprior surgery (fasted groups) (n � 8).
Besides the group receiving no treatment, fasted rats with fattyliver were also subjected to three different treatment modalities andaccordingly divided: (1) Glucose supplemented group: fasted rats hadfree access to a 40% glucose oral solution for the 18 h prior procure-ment (n � 8); (2) Prostaglandin (PG) group: the livers explanted fromfasted rats were infused with alprostadil (Prostin VR, Upjohn-Pharmacia, Puurs, Belgium), a PGE1 analog, during the 120 min ofreperfusion at the rate of 1 �g/kg rat/min (n � 8); (3) Combinedtreatment group: fasted rats had access to a 40% glucose oral solutionfor the 18 h prior procurement and the explanted livers were reper-fused with alprostadil during the 120 min of reperfusion at the rateof 1 �g/kg rat/min (n � 6). The experimental design is summarized inFig. 1.
Liver Oxygen Consumption
The perfusate was sampled from a portal inflow port and from anoutflow port during reperfusion to determine the oxygen concentra-
tion with a blood gas analyzer (ABL 5, ABL Instruments, Copenha-
192 JOURNAL OF SURGICAL RESEARCH: VOL. 127, NO. 2, AUGUST 2005
gen, Denmark). Rates of oxygen consumption were calculated frominfluent minus effluent concentration differences, liver wet weight,and flow rate.
Biochemical Methods
ALT Release
The effluent perfusate from the liver was collected during reper-fusion for measurement of the ALT levels using a commerciallyavailable kit test (Sigma-Aldrich, Milan, Italy).
Glycogen
The preischemic hepatic glycogen content was quantified usingthe method of Keppler and Decker [18] and expressed as mg/�g DNA.The liver DNA concentration was measured according the method ofSchneider and Greco [19].
Thiobarbituric Acid-Reactive Substances (TBARs)
Approximately 100 mg of liver tissue was homogenized in 5 vol of180 mM KCl, 50 mM Tris HCl, and 10 mM EDTA (pH 7.4) containing0.02% (w/v) butylated hydroxytoluene. Then, the homogenate wasprecipitated with 10% trichloroacetic acid and the supernatant wasincubated at 100°C for 45 min with an equal volume of 0.67% thio-barbituric acid. After cooling, the supernatant was extracted with 1ml of n-butanol and the absorption peak was measured at 532 nm[20].
Protein Carbonyl (PC)
Aliquots of liver tissue homogenates containing approximately 2mg of proteins were incubated for 60 min at room temperature with1 ml of 2% dinitrophenylhydrazine (DNPH) in 2 N HCl or 1 ml of 2N HCl as control blank. Next, proteins were precipitated with 200 �lof 50% TCA and washed three times with 1:1 (w:v) ethanol:ethylac-etate and three times with 10% TCA. The final precipitate was solvedin 6 M guanidine and the spectrum of the DNPH versus HCl controlswas followed at 350–375 nm [21]. The concentration of carbonylgroups was then calculated using 21.5 nM�1 cm�1 as the extinctioncoefficient for aliphatic hydrazones.
Total Protein
Total protein concentration in liver homogenate was determinedby the method of Lowry [22]. The protein concentration expressed asmg protein/g of liver homogenate did not differ significantly between
FIG. 1. Flow diagram of the experimental design. PG � alpros-tadil during reperfusion.
the experimental groups.
Chemicals
All chemicals were purchased from Sigma-Aldrich.
Statistical Analysis
The values of the ALT release, oxygen consumption, and portalpressure resulted normally distributed as assessed by theKolmogorov–Smirnov test and were reported as mean � SD. Thedifference between groups was calculated by using a two-wayANOVA for repeated samples. The data obtained for the liver glyco-gen content and the oxidative parameters did not result normallydistributed and were reported as median � range. Statistically sig-nificant differences between the two groups were determined by theWilcoxon rank sum test. Statistical analysis was performed by run-ning the SPSS Version 10.0 for Windows package on a personalcomputer. A P value less than 0.05 was considered to be significant.
RESULTS
Preischemic Hepatic Glycogen Content
Under fed conditions, the preischemic hepatic con-tent of glycogen was significantly lower in fatty thannormal rats (median: 1.62; range: 1.00–2.10 mg/�gDNA versus median: 1.07; range: 0.50–1.59 mg/�gDNA; P � 0.05). As expected, in all of the experimentalgroups exposed to 18 h of fasting, the glycogen storeswere almost completely depleted. Nutritional supple-mentation of fasted rats with fatty liver restored ap-proximately 60% of the glycogen stores measured un-der fed conditions in the group supplemented with oralglucose alone (median: 0.62; range: 0.58–0.69 mg/�gDNA) and in the group supplemented with glucose andtreated with PG (median: 0.63; range 0.52–0.72 mg/�gDNA) (Fig. 2).
Effect of Fasting on Preservation-Reperfusion Injury inNormal and Fatty Liver
Under fed conditions, as expected, the ALT releasewas greater in fatty than in normal livers throughoutall of the reperfusion time (at 30 min: 236 � 71 versus
FIG. 2. Preischemic hepatic glycogen content in the differentexperimental groups (n � 4–6). G: glucose supplementation priorharvesting; PG: alprostadil during reperfusion. Fatty fed versus nor-mal fed � P � 0.05. Fasted versus fed groups � P � 0.001. Glucosesupplementation and combined treatment versus untreated fatty
fasted � P � 0.01.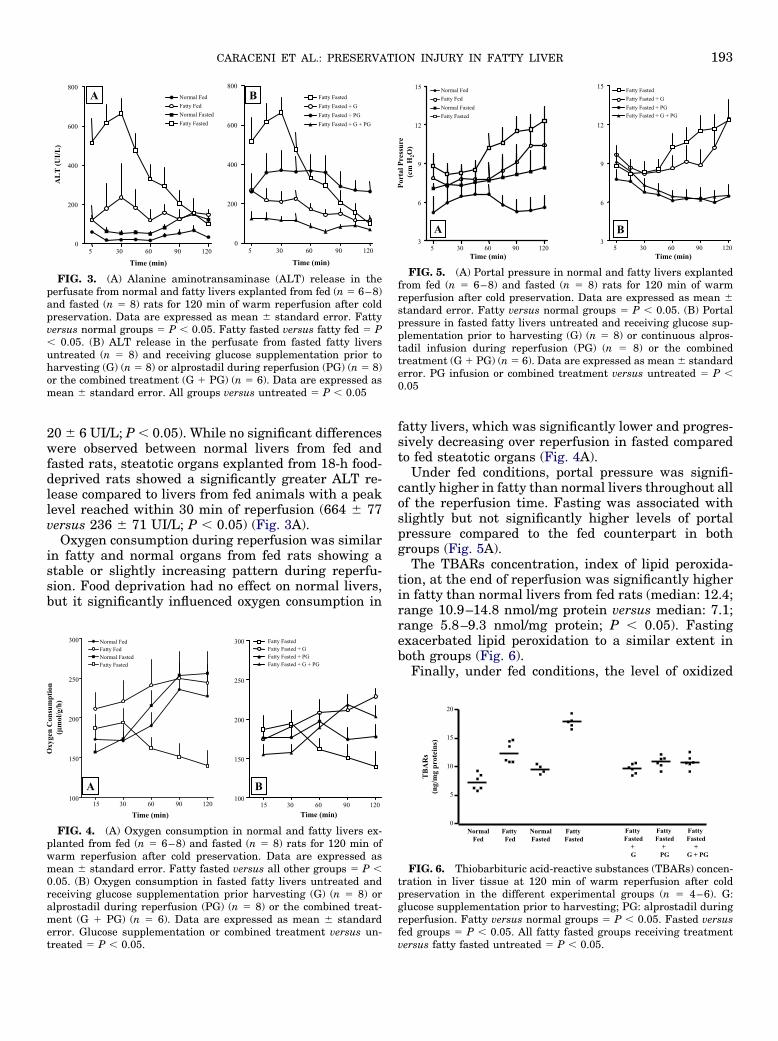
193CARACENI ET AL.: PRESERVATION INJURY IN FATTY LIVER
20 � 6 UI/L; P � 0.05). While no significant differenceswere observed between normal livers from fed andfasted rats, steatotic organs explanted from 18-h food-deprived rats showed a significantly greater ALT re-lease compared to livers from fed animals with a peaklevel reached within 30 min of reperfusion (664 � 77versus 236 � 71 UI/L; P � 0.05) (Fig. 3A).
Oxygen consumption during reperfusion was similarin fatty and normal organs from fed rats showing astable or slightly increasing pattern during reperfu-sion. Food deprivation had no effect on normal livers,but it significantly influenced oxygen consumption in
FIG. 4. (A) Oxygen consumption in normal and fatty livers ex-planted from fed (n � 6–8) and fasted (n � 8) rats for 120 min ofwarm reperfusion after cold preservation. Data are expressed asmean � standard error. Fatty fasted versus all other groups � P �0.05. (B) Oxygen consumption in fasted fatty livers untreated andreceiving glucose supplementation prior harvesting (G) (n � 8) oralprostadil during reperfusion (PG) (n � 8) or the combined treat-ment (G � PG) (n � 6). Data are expressed as mean � standarderror. Glucose supplementation or combined treatment versus un-
FIG. 3. (A) Alanine aminotransaminase (ALT) release in theperfusate from normal and fatty livers explanted from fed (n � 6–8)and fasted (n � 8) rats for 120 min of warm reperfusion after coldpreservation. Data are expressed as mean � standard error. Fattyversus normal groups � P � 0.05. Fatty fasted versus fatty fed � P� 0.05. (B) ALT release in the perfusate from fasted fatty liversuntreated (n � 8) and receiving glucose supplementation prior toharvesting (G) (n � 8) or alprostadil during reperfusion (PG) (n � 8)or the combined treatment (G � PG) (n � 6). Data are expressed asmean � standard error. All groups versus untreated � P � 0.05
treated � P � 0.05.
fatty livers, which was significantly lower and progres-sively decreasing over reperfusion in fasted comparedto fed steatotic organs (Fig. 4A).
Under fed conditions, portal pressure was signifi-cantly higher in fatty than normal livers throughout allof the reperfusion time. Fasting was associated withslightly but not significantly higher levels of portalpressure compared to the fed counterpart in bothgroups (Fig. 5A).
The TBARs concentration, index of lipid peroxida-tion, at the end of reperfusion was significantly higherin fatty than normal livers from fed rats (median: 12.4;range 10.9–14.8 nmol/mg protein versus median: 7.1;range 5.8–9.3 nmol/mg protein; P � 0.05). Fastingexacerbated lipid peroxidation to a similar extent inboth groups (Fig. 6).
Finally, under fed conditions, the level of oxidized
FIG. 6. Thiobarbituric acid-reactive substances (TBARs) concen-tration in liver tissue at 120 min of warm reperfusion after coldpreservation in the different experimental groups (n � 4–6). G:glucose supplementation prior to harvesting; PG: alprostadil duringreperfusion. Fatty versus normal groups � P � 0.05. Fasted versusfed groups � P � 0.05. All fatty fasted groups receiving treatment
FIG. 5. (A) Portal pressure in normal and fatty livers explantedfrom fed (n � 6–8) and fasted (n � 8) rats for 120 min of warmreperfusion after cold preservation. Data are expressed as mean �standard error. Fatty versus normal groups � P � 0.05. (B) Portalpressure in fasted fatty livers untreated and receiving glucose sup-plementation prior to harvesting (G) (n � 8) or continuous alpros-tadil infusion during reperfusion (PG) (n � 8) or the combinedtreatment (G � PG) (n � 6). Data are expressed as mean � standarderror. PG infusion or combined treatment versus untreated � P �0.05
versus fatty fasted untreated � P � 0.05.

194 JOURNAL OF SURGICAL RESEARCH: VOL. 127, NO. 2, AUGUST 2005
protein was similar in fatty and normal livers. How-ever, when rats were subjected to fasting, the increaseof PC concentration at the end of reperfusion was sig-nificantly greater in fatty liver (fed: median 3.1, range2.5–3.9 nmol/mg protein; fasted: median 4.6; range4.2–5.6 nmol/mg protein; P � 0.05) than normal livers(fed: median 2.8; range 1.7–4.6 nmol/mg protein; fast-ed: median 3.2, range 2.6–4.0 nmol/mg protein; P � ns)(Fig. 7).
Effect of Nutritional Supplementation with or withoutProstaglandin Treatment on Preservation-Reperfusion Injury
in Fasted Fatty Liver
Preischemic glucose supplementation greatly re-duced the ALT release observed during the first hour ofreperfusion in fatty livers explanted from fasted rats(after 30 min: 210 � 51 versus 664 � 77 UI/L; P �0.05). PG infusion alone was also able to reduce signif-icantly, although to a lesser extent, the ALT releasecompared to fasted untreated livers (after 30 min: 371� 97 UI/L; P � 0.05). When glucose supplementationwas combined with PG infusion, the ALT release dur-ing reperfusion was almost abolished (after 30 min:117 � 19 UI/L; P � 0.01); however, no addictive effectof the combined treatment could be demonstrated (Fig.3B).
As shown in Fig. 4B, oxygen consumption was quan-titatively and temporally affected by nutritional reple-tion: it increased progressively during reperfusion from174 � 13 to 229 � 13 �mol/g/h in livers explanted fromfasted rats supplemented with glucose, while it de-creased from 187 � 14 to 140 � 16 �mol/g/h in fasteduntreated liver. PG infusion was not associated withany significant effect on oxygen consumption neither ifgiven alone or if given in combination with glucosesupplementation.
FIG. 7. Protein carbonyl (PC) concentration in liver tissue at 120min of warm reperfusion after cold preservation in the differentexperimental groups (n � 4–6). G: glucose supplementation prior toharvesting; PG: alprostadil during reperfusion. Fatty fasted versusfatty fed � P � 0.05. All fatty fasted groups receiving treatmentversus fatty fasted untreated � P � 0.05.
Portal pressure was not influenced by the preisch-
emic glucose supplementation. In contrast, PG infu-sion significantly lowered portal pressure compared tothat measured in fasted untreated rats both whengiven alone or when given in combination with glucose(Fig. 5B).
Finally, all treatments significantly but to a similarextent reduced oxidative injury compared to the fasteduntreated rats (Figs. 6 and 7).
DISCUSSION
Although several surgical and pharmacologicalstrategies have produced promising results in experi-mental models, there are no treatment modalities rou-tinely used in clinical practice to protect fatty liversagainst preservation injury other than minimizing coldand warm ischemic times [1]. This study indicates thatthe preexisting nutritional status is a potential impor-tant and manageable determinant of preservation in-jury in steatotic donor livers. Indeed, a fasting periodlong enough to cause a complete depletion of the gly-cogen stores worsened the reperfusion cell damage,metabolic function, and oxidative stress with a signif-icantly greater extent in fatty than normal rat livers.
Although hypothermia greatly decreases the meta-bolic rate of hepatocytes, a certain amount of adenosinetriphosphate (ATP) is still needed to maintain the cel-lular integrity. Fasting exacerbates ischemic injurymainly by causing depletion of the glycogen stores,which results in a more rapid ATP exhaustion duringischemia, when the oxidative phosphorylation is inhib-ited and glycogen is important to supply glucose forglycolytic ATP generation [9]. After 18 h of fasting, thehepatic glycogen was almost completely depleted ineither fatty or normal livers. This latter finding isapparently against the hypothesis which relates thereduced tolerance of fatty liver to preservation injuryto the quantitative amount of glycogen stores. How-ever, it cannot be excluded that the exhaustion of theglycogen during fasting occurs more rapidly in stea-totic livers because of the initial lower glycogen con-tent, as demonstrated in fed animals [15, 23], thuspredisposing to greater injury during the subsequentpreservation and reperfusion phase.
Food deprivation can also contribute to preservation-reperfusion injury by lowering the antioxidant defensesystems and, thus, making the livers more vulnerableto the free-radical attack. Fatty livers appear to beeven more exposed to the adverse effect of fasting, aswe have previously shown that 18 h of starvation pro-duces a significantly greater oxidative imbalance in thewhole tissue and in the mitochondrial compartment ofsteatotic than normal livers [24, 25]. In the presentexperiments, fasting was associated with a greater con-centration of oxidized lipids and proteins at the end ofreperfusion in fatty than normal livers. Although the
higher TBARs levels could only reflect the lipid accu-
195CARACENI ET AL.: PRESERVATION INJURY IN FATTY LIVER
mulation in the cytosol of hepatocytes, which providesmore substrate for the initiation and subsequent am-plification of the free-radical attack, the concomitanthigher PC level clearly indicates the exacerbation ofthe fasting-induced oxidative damage.
Hepatic oxygen consumption was lowered signifi-cantly by fasting solely in steatotic livers likely reflect-ing a lower metabolic activity of the nutritional-depleted fatty hepatocytes rather than resulting fromKupffer cell inactivation and increased free-radicalgeneration. Although Kupffer cell inactivation hasbeen reported to decrease the oxygen uptake duringreperfusion, it seems unlikely that, in the presentstudy, 18 h of fasting has produced such an effect,which is described only after a much longer period ofstarvation [26]. Furthermore, oxygen serves also forthe generation of free-radical species during reperfu-sion; however, no relation was found between oxygenconsumption and oxidative stress: fasted fatty livershad the lowest oxygen consumption and the highestoxidative damage, while nutritional repletion en-hanced oxygen consumption and reduced the levels ofoxidized lipids and proteins. In contrast, a “no-reflow”phenomenon due to microcirculatory disturbances mayhave also contributed to the lower oxygen consumptionin fatty-fasted livers through a reduced oxygen deliv-ery to the liver cells. The lack of improvement with theinfusion of the vasodilator alprostadil should not betaken as evidence against this event since a number ofother factors, such as adenosine, nitric oxide, and en-dothelins, are involved in the pathogenesis of the mi-crocirculatory alterations [27].
In normal livers, preischemic nutritional repletionby glucose supplementation improves posttransplantrecovery and survival [12–14, 28, 29]. Manipulation ofthe nutritional status is likely to be much more rele-vant in the specific case of livers with fatty infiltration,which present an altered content of energetic sub-strates [15, 23], a defective capacity to synthesize ATPafter transplantation [30], and a closer relationshipbetween nutritional status and preservation injury.
In the present study, it was first demonstrated in thesetting of fatty liver that feeding fasted rats a 40%glucose oral solution produced a significant improve-ment of the cell injury, oxygen consumption, and oxi-dative stress during reperfusion after preservation.This procedure was able to replenish the glycogenstores to approximately 60% of the level seen in fedanimals and it was chosen for the efficacy and theexperimental simplicity already shown in previousstudies [12, 15]. However, several issues have to beclarified before any technique of nutritional support,although relatively easy and safe to perform, can reachthe clinical practice and further studies are needed todefine the best composition and the route and timing of
administration of the energetic substrates taking intoaccount the very peculiar metabolic situation of abrain-death donor with fatty liver.
The PG are effective against ischemia-reperfusioninjury by increasing liver perfusion, by inhibitingwhite blood cell and platelet aggregation to endothe-lium, and probably by having a direct cytoprotectiveeffect [27]. These properties make PG a plausible can-didate for the pharmacological treatment of the micro-circulation impairment in steatotic livers. In thisstudy, infusion of the PGE1 analog, alprostadil, re-duced the serum ALT release and the oxidative stress,improved portal pressure, but had no effect on oxygenconsumption. When glucose supplementation was com-bined with PG infusion, the ALT release was almostabolished even if we could not document a significantaddictive effect compared to the single therapies.
Despite these data confirming the protective effect ofPG against reperfusion injury in fatty liver [4, 5], thelack of a clear synergistic effect on cell injury whenadministered in livers supplemented with glucosecould be apparently discouraging. However, we shouldbear in mind that PG greatly ameliorated liver perfu-sion, the only parameter not significantly affected byglucose supplementation, and that the use of an iso-lated perfused liver model with a cell-free media im-plies some methodological limitations, which may havemasked the full benefit of PG infusion. Indeed, the PGeffect against mechanisms of the inflammatory re-sponse, such as the adherence of circulating neutro-phils and platelets to the endothelium in the initialphase of reperfusion and the neutrophil infiltration inthe liver tissue in the later phases, could not be as-sessed in our experimental model.
In conclusion, this study demonstrates that preser-vation in fatty liver is greatly related to the animalnutritional status. Preischemic supplementation withenergetic substrates may represent a clinically feasibleprotective strategy and the concomitant administra-tion of PG could offer a rational multistep approach toprotect the parenchymal and sinusoidal components,which are both targets of preservation injury in fattylivers.
ACKNOWLEDGMENTS
This study was supported in part by the Ministero dell’Istruzione,Università e Ricerca (M.I.U.R.)–Progetti di Ricerca di Interesse Na-zionale 2001 (Fondi ex-40%), by the University of Bologna (Fondiex-60%), and by the Fondazione Cassa di Risparmio in Bologna.Marco Domenicali was in part sponsored by a Fellowship of theItalian Association for the Study of the Liver (AISF).
REFERENCES
1. Selzner, M., and Clavien, P. A. Fatty liver in liver transplanta-tion and surgery. Semin. Liver Dis. 21: 105, 2001.
2. Teramoto, K., Bowers, J. L., Kruskal, J. B., et al. Hepaticmicrocirculatory changes after reperfusion in fatty and normal
liver transplantation in rat. Transplantation 56: 1076, 1993.
196 JOURNAL OF SURGICAL RESEARCH: VOL. 127, NO. 2, AUGUST 2005
3. Seifalian, A. M., Chidambaram, V., Rolles, K., et al. In vivodemonstration of impaired microcirculation in steatotic humanliver grafts. Liver Transpl. Surg. 4: 71, 1998.
4. Husberg, B. S., Genyk, Y. S., and Klintmalm, G. B. A new ratmodel for studies of the ischemic injury after transplantation offatty livers: improvement after postoperative administration ofprostaglandin. Transplantation 57: 457, 1994.
5. Morioka, D., Kubota, T., Sekido, H., et al. Prostaglandin E1improved the function of transplanted fatty liver in a ratreduced-size-liver transplantation model under conditions ofpermissible cold preservation. Liver Transpl. 9: 79, 2003.
6. Taneja, C., Prescott, L., and Koneru, B. Critical preservationinjury in rat fatty liver is to hepatocytes, not sinusoidal liningcells. Transplantation 65: 167, 1998.
7. Gao, W., Connor, H. D., Leemaster, J. J., et al. Primary non-function of fatty livers produced by alcohol is associated with anew, antioxidant insensitive free radical species. Transplanta-tion 59: 674, 1995.
8. Caraceni, P., Bianchi, C., Domenicali, M., et al. Impairment ofmitochondrial oxidative phosphorylation in rat fatty liver ex-posed to preservation-reperfusion injury. J. Hepatol. 41: 82,2004.
9. Miki, C., Iriyama, K., Mirza, D. F., et al. Postperfusion energymetabolism of steatotic graft and its relation to early graftviability following liver transplantation. Dig. Dis. Sci. 43: 74,1998.
10. Rosser, B. G., and Gores, G. J. Liver cell necrosis: cellularmechanism and clinical implication. Gastroenterology 108: 252,1995.
11. Bradford, B. U., Maotto, M., Lemaster, J. J., et al. New, simplemodels to evaluate zone-specific damage due to hypoxia in theperfused rat liver: time course and effect of nutritional status.J. Pharmacol. Exp. Ther. 236: 263, 1986.
12. Sumimoto, R., Southard, J. H., and Belzer, F. O. Livers fromfasted rats acquire resistance to warm and cold ischemia.Transplantation 55: 728, 1993.
13. Lindell, S. L., Hansen, T., Rankin, M., et al. Donor nutritionalstatus—a determinant of liver preservation injury. Transplan-tation 61: 239, 1996.
14. Cywes, R., Greig, P. D., Morgan, G. R., et al. Rapid donor livernutritional enhancement in a large animal model. Hepatology16: 1271, 1992.
15. Sadamori, H., Tanaka, N., Yagi, T., et al. The effects of nutri-tional repletion on donors for liver transplantation in pigs.Transplantation 60: 317, 1995.
16. Caraceni, P., Nardo, B., Domenicali, M., et al. Ischemia-
reperfusion injury in the rat fatty liver: role of nutritionalstatus. Hepatology 29: 1139, 1999.
17. Shiino, Y., Nakamura, J., Okamoto, T., et al. Improved qualityof fatty liver allografts by starvation in rats. Transplant. Proc.30: 3294, 1998.
18. Hakamada, K., Sasaki, M., Takahashi, K., et al. Sinusoidal flowblock after warm ischemia in rats with diet-induced fatty liver.J. Surg. Res. 70: 12, 1997.
19. Keppler, D., and Decker, K. Glycogen. In Methods of EnzymaticAnalysis, Vol. 6. H. U. Bergmeyer (Ed.), Deerfield Beach, FL:Verlag Chemic, 1986. pp. 11–18.
20. Schneider, W. C., and Greco, A. E. Incorporation of pyrimidinedeoxyribonucleosides into liver lipids and other components.Biochim. Biophys. Acta 228: 610, 1971.
21. Slater, T., and Sawyer, B. The stimulatory effect of carbontetrachloride and other halogenoalkanes on peroxidative reac-tions in rat liver fractions in vitro. Biochem. J. 123: 805, 1971.
22. Levine, R. L., Garland, D., Oliver, C. N., et al. Determination ofcarbonyl content in oxidatively modified proteins. Methods En-zymol. 186: 464, 1990.
23. Lowry, O. H., Rosenbrough, N. J., Farr, A. L., et al. Proteinmeasurement with the Folin phenol reagent. J. Biol. Chem.193: 265, 1951.
24. Caraceni, P., Ryu, H. O., Subbotin, V., et al. Rat hepatocytesisolated from alcohol-induced fatty liver have an increased sen-sitivity to anoxic injury. Hepatology 25: 943, 1997.
25. Grattagliano, I., Vendemiale, G., Caraceni, P., et al. Starvationimpairs antioxidant defense is impaired in fatty livers of ratsfed a choline-deficient diet. J. Nutr. 130: 2131, 2000.
26. Vendemiale, G. L., Grattagliano, I., Caraceni, P., et al. Mito-chondrial oxidative injury and energy metabolism alteration inrat fatty liver: effect of the nutritional status. Hepatology 33:808, 2001.
27. Fusaoka, T., Hunt, K. J., Leemasters, J. J., et al. Evidence thatactivation of Kupffer cells increases oxygen uptake after coldstorage. Transplantation 58: 1067, 1994.
28. Selzner, M., Rudiger, H. A., Graf, R., et al. Protective strategiesagainst ischemic injury of the liver. Gastroenterology 125: 917,2003.
29. Cywes, R., Greig, P. D., Sanabria, J. R., et al. Effect of intra-portal glucose infusion on hepatic glycogen content and degra-dation, and outcome of liver transplantation. Ann. Surg. 216:235, 1992.
30. Sadamori, H., Tanaka, N., Yagi, T., et al. The effects of nutri-tional repletion on donors for liver transplantation in pigs.
Transplantation 60: 317, 1995.