the lipostat, hyperphagia and catch-up growth
TRANSCRIPT

The lipostat, hyperphagia and catch-up growth
M Jobling & S J S Johansen
NFH, University of Tromsù, 9037 Tromsù, Norway
Correspondence: M Jobling, NFH, University of Tromsù, 9037 Tromsù, Norway
Abstract
The hypothesis that long-term energy intake is
matched to energy expenditure arose during the
1950s, and this formed the basis of the lipostatic
model for the control of food intake in mammals.
This gave rise to an additional hypothesis that body
weight varies little over adult life because body fat, a
variable related to body mass, is regulated. There is
now a large body of evidence that adipose tissue
plays a role in the regulation of feeding and body
weight in mammals, and the study of the mechan-
isms by which the brain monitors the signals arising
from the adipose tissue is currently a major area of
research. After a period of nutritional restriction, a
number of compensatory responses are invoked,
and these result in hyperphagia, rapid weight
increase and the repletion of energy reserves.
However, the extent to which animals recover lost
body weight has been reported to vary between
studies. It is hypothesized that the rate at which
animals replete their lipid reserves during catch-up
growth may in¯uence the hyperphagic response
and, hence, whether or not there is complete
recovery of body weight. Preliminary tests carried
out using some data collected in studies of catch-up
growth in salmonids appear to provide support for
the model, but more experimental studies are
needed to provide rigorous testing.
The lipostat
The concept that body weight and adipose mass are
subject to negative feedback control arose about
50 years ago, when it was hypothesized that long-
term (weeks±months) energy intake was actively
matched to energy expenditure. When the energy
balance (the difference between energy intake and
expenditure) of animals was changed experimen-
tally, the changes in body weight induced by such
interventions resulted in the activation of compen-
satory responses that eventually returned body
weight to a value close to the original. For example,
a period of imposed food restriction resulted in
weight loss but, when food was made available
again, the animals overate, i.e. became hyperpha-
gic, until body weight returned to a level similar to
that before restriction. Thus, it was considered that,
although body weight sensu stricto may not be
regulated, a variable, or set of variables, correlated
to body weight must be regulated.
These observations set the stage for Kennedy's
lipostatic model for the control of food intake and
body composition (Kennedy 1953). It was hypothe-
sized that humoral signals generated in proportion
to adipose mass (size of lipid reserves) provide input
(feedback) to the areas of the brain that control
feeding and energy expenditure. In this regulatory
process, there may be alterations in food intake,
spontaneous activity, metabolism, metabolic ef®-
ciency and thermogenesis. Although there is gen-
eral agreement that these responses are co-
ordinated by the brain, the way in which the brain
monitors body adiposity and invokes the compen-
satory responses are still not completely understood,
and a variety of signalling mechanisms have been
proposed (Hervey 1969; Weigle 1994; Cabanac &
Richard 1996; Matson, Wiater & Weigle 1996;
Friedman 1998). In his lipostatic model, Kennedy
(1953) proposed that a change in energy balance
suf®cient to alter adiposity elicited a compensatory
change in food intake as a result of the change in
the amount of negative feedback. Thus, after a
period of food restriction, the negative feedback
signals that inhibit feeding are reduced because of
fat loss. The result is elevated food intake that is
R
# 1999 Blackwell Science Ltd 473
Aquaculture Research, 1999, 30, 473±478

maintained until the level of fat and, hence, the
negative feedback signals return to normal.
Evidence that one such negative feedback signal
was a circulating factor came from parabiosis
studies. In parabiosis, two animals are physically
joined to one another so that they share a cross-
circulation of blood. When normal, lean rats were
joined to rats rendered obese by hypothalamic
lesions, the normal rats reduced their food intake
and would starve to death if not separated from
their obese partners (Hervey 1959). This provided
strong evidence that a circulating, `satiety' factor
from the lesioned, obese rat was affecting the
normal rat. In other words, the obese rat seemed
to secrete a circulating endocrine signal that crossed
over to the normal, lean partner and inhibited its
food intake, but the hypothalamic lesion had
rendered the obese animal unresponsive to the
circulating signal, so that it continued to feed and
accumulate body fat. Later, it was shown that when
rats with diet-induced obesity were parabiosed with
normal, non-obese rats, the food intake of the
normal rats decreased, and there was a reduction in
the body fat of these animals. These changes were
attributed to an energy-stabilizing, satiety factor
carried in the circulation from the obese to the
normal rat (Nishizawa & Bray 1980). Similar
changes were also demonstrated when tube-fed
obese rats were parabiosed with normal partners
(Harris & Martin 1984). The results of these
experiments led to the conclusion that the regula-
tion of both food intake and body energy reserves in
the form of fat was probably mediated by a blood-
borne factor that acted at the level of the
hypothalamus. Evidence is now accumulating in
favour of the hypothesis that an `adipose tissue
factor' (ob protein or leptin) may play a role in the
co-ordination of ingestive behaviour and energy
balance in mammals, but this factor is not seen as
working in isolation (Matson et al. 1996; Blum
1997; Friedman 1998; Hossner 1998; Schwartz &
Seeley 1997).
Catch-up growth
It is now widely accepted that adipose tissue mass
in¯uences ingestive behaviour and food intake in
mammals (Weigle 1994; Matson et al. 1996; Blum
1997; Schwartz & Seeley 1997; Friedman 1998;
Hossner 1998), and there is also evidence that the
size of the body fat stores may have an in¯uence on
feeding in salmonid ®shes (Metcalfe & Thorpe 1992;
Jobling & Miglavs 1993; Shearer, Silverstein &
Plisetskaya 1997). Furthermore, the responses of
®sh after a period of nutritional restriction seem to
resemble those observed in mammals: hyperphagia,
rapid growth and the repletion of energy reserves
(Broekhuizen, Gurney, Jones & Bryant 1994;
Jobling 1994). Thus, ®sh will usually consume
more food and grow faster than expected after a
period of fasting, compensating for lost growth and
returning body mass to a level that approaches that
of conspeci®cs that have had continuous access to
food (Broekhuizen et al. 1994; Jobling 1994). The
extent to which fasted ®sh display hyperphagia on a
return to adequate feeding conditions may be
related to the degree to which their energy reserves
were depleted during fasting, and there may be an
inverse relationship between post-fast food intake
and the size of the lipid depots (Jobling & Miglavs
1993).
Hervey (1969) hypothesized that the long-term
stability of body weight in adult mammals was
mediated through the regulation of a variable
correlated to body mass. Broekhuizen et al. (1994)
extended this hypothesis in a two-compartment
model in which tissues could be broadly divided into
reserves (fat depots and the mobilizable parts of the
musculature) and structural components (skeleton,
circulatory and nervous tissue): it was proposed that
it is the balance between these two components that
is regulated, and that shifts in the balance would
invoke compensatory changes. Thus, it was en-
visaged that the responses observed in animals
undergoing catch-up growth could be related to a
monitoring of the `instantaneous nutritional state',
as re¯ected in the ratio of reserves to structural
tissues. In most animals, the fat depots represent the
easily mobilized energy reserves, and the lean body
mass (LBM) incorporates the structural tissues, so
the ratio fat:LBM may comply with an indicator of
nutritional state (although it must be borne in mind
that LBM will also include a mobilizable compo-
nent).
Among ®sh species, catch-up or compensatory
growth has been reported for salmonids, cyprinids,
pleuronectids, gadoids and centrarchids (e.g.
Weatherley & Gill 1987; Miglavs & Jobling 1989a;
Bejda, Phelan & Studholme 1992; Russell &
Wootton 1992; Broekhuizen et al. 1994; Jobling,
Melùy, dos Santos & Christiansen 1994; Bull &
Metcalfe 1997; Hayward, Noltie & Wang 1997;
Nicieza & Metcalfe 1997), but responses have been
inconsistent. In other words, recovery of lost body
L
474 # 1999 Blackwell Science Ltd, Aquaculture Research, 30, 473±478
Hyperphagia and catch-up growth M Jobling & S J S Johansen Aquaculture Research, 1999, 30, 473±478
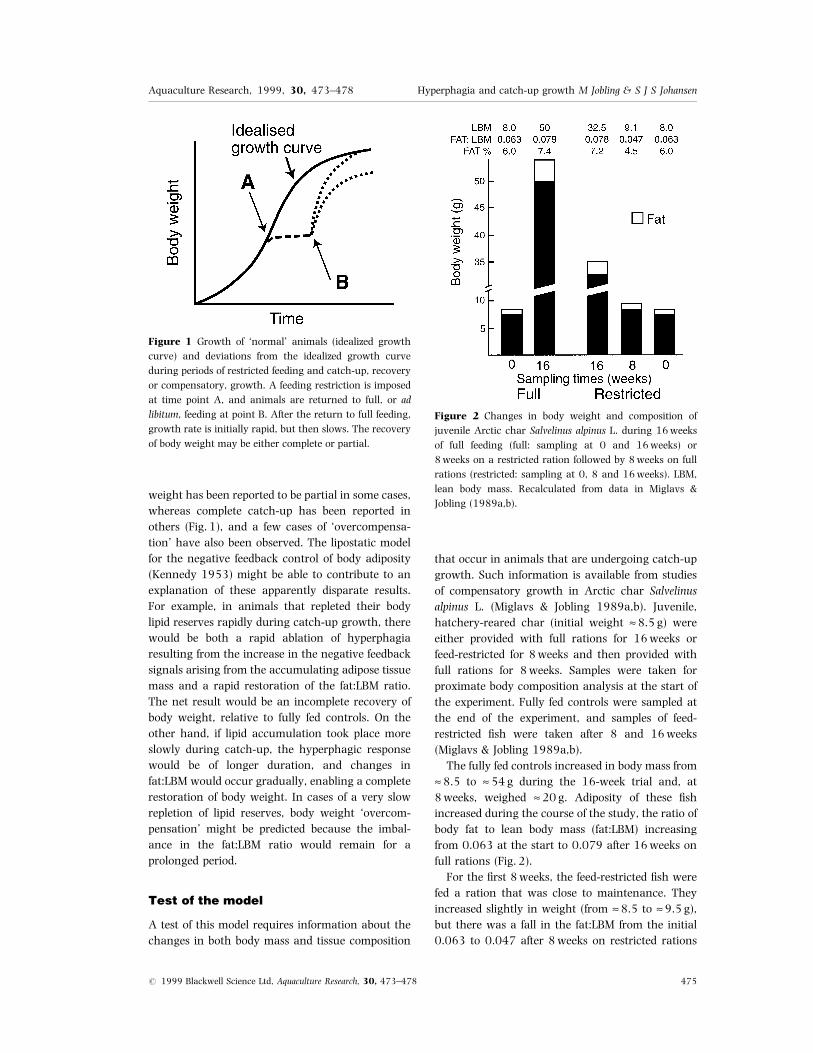
weight has been reported to be partial in some cases,
whereas complete catch-up has been reported in
others (Fig. 1), and a few cases of `overcompensa-
tion' have also been observed. The lipostatic model
for the negative feedback control of body adiposity
(Kennedy 1953) might be able to contribute to an
explanation of these apparently disparate results.
For example, in animals that repleted their body
lipid reserves rapidly during catch-up growth, there
would be both a rapid ablation of hyperphagia
resulting from the increase in the negative feedback
signals arising from the accumulating adipose tissue
mass and a rapid restoration of the fat:LBM ratio.
The net result would be an incomplete recovery of
body weight, relative to fully fed controls. On the
other hand, if lipid accumulation took place more
slowly during catch-up, the hyperphagic response
would be of longer duration, and changes in
fat:LBM would occur gradually, enabling a complete
restoration of body weight. In cases of a very slow
repletion of lipid reserves, body weight `overcom-
pensation' might be predicted because the imbal-
ance in the fat:LBM ratio would remain for a
prolonged period.
Test of the model
A test of this model requires information about the
changes in both body mass and tissue composition
that occur in animals that are undergoing catch-up
growth. Such information is available from studies
of compensatory growth in Arctic char Salvelinus
alpinus L. (Miglavs & Jobling 1989a,b). Juvenile,
hatchery-reared char (initial weight » 8.5 g) were
either provided with full rations for 16 weeks or
feed-restricted for 8 weeks and then provided with
full rations for 8 weeks. Samples were taken for
proximate body composition analysis at the start of
the experiment. Fully fed controls were sampled at
the end of the experiment, and samples of feed-
restricted ®sh were taken after 8 and 16 weeks
(Miglavs & Jobling 1989a,b).
The fully fed controls increased in body mass from
» 8.5 to » 54 g during the 16-week trial and, at
8 weeks, weighed » 20 g. Adiposity of these ®sh
increased during the course of the study, the ratio of
body fat to lean body mass (fat:LBM) increasing
from 0.063 at the start to 0.079 after 16 weeks on
full rations (Fig. 2).
For the ®rst 8 weeks, the feed-restricted ®sh were
fed a ration that was close to maintenance. They
increased slightly in weight (from » 8.5 to » 9.5 g),
but there was a fall in the fat:LBM from the initial
0.063 to 0.047 after 8 weeks on restricted rations
R
Figure 1 Growth of `normal' animals (idealized growth
curve) and deviations from the idealized growth curve
during periods of restricted feeding and catch-up, recovery
or compensatory, growth. A feeding restriction is imposed
at time point A, and animals are returned to full, or ad
libitum, feeding at point B. After the return to full feeding,
growth rate is initially rapid, but then slows. The recovery
of body weight may be either complete or partial.
Figure 2 Changes in body weight and composition of
juvenile Arctic char Salvelinus alpinus L. during 16 weeks
of full feeding (full: sampling at 0 and 16 weeks) or
8 weeks on a restricted ration followed by 8 weeks on full
rations (restricted: sampling at 0, 8 and 16 weeks). LBM,
lean body mass. Recalculated from data in Miglavs &
Jobling (1989a,b).
# 1999 Blackwell Science Ltd, Aquaculture Research, 30, 473±478 475
Aquaculture Research, 1999, 30, 473±478 Hyperphagia and catch-up growth M Jobling & S J S Johansen

(Fig. 2). After the introduction of full feeding, these
®sh became hyperphagic and displayed marked
increases in growth rate (Miglavs & Jobling
1989a). The hyperphagia had ablated by the end
of the trial, and growth rates had also declined to
levels similar to those of the fully fed controls
(Miglavs & Jobling 1989a). However, the body
weights of these ®sh were only » 35 g, so the
compensatory responses had resulted in a partial
recovery of body weight in comparison with the
fully fed controls (Fig. 2).
Comparison of proximate body compositions of
the two groups of ®sh at the end of the trial revealed
similarities in adiposity (fat percentage: 7.4 vs.
7.2%) and fat:LBM (0.079 vs. 0.078) (Fig. 2), which
would have been predicted if lipostatic control of
feed intake were important in governing the growth
of feed-restricted ®sh during the recovery phase.
Additional, albeit circumstantial, evidence for the
idea of the involvement of lipostatic mechanisms in
the regulation of feeding and growth may be
provided by data obtained in a study of seasonal
lipid dynamics of wild, anadromous char
(Jùrgensen, Johansen & Jobling 1997).
Anadromous char undertake annual feeding migra-
tions to the sea during the summer months and
spend 40±50 days in sea water before returning to
fresh water to overwinter.
Char from the north Norwegian population
examined by Jùrgensen et al. (1997) increased
substantially in body weight during the summer
(immature ®sh from about 300 to 600 g; maturing
®sh from 500 to 800 g), and the weight of body lipid
increased approximately ®vefold (Jùrgensen et al.
1997). Fish that were descending to the sea in
spring were lipid depleted (fat:LBM of 0.021 in
males and 0.024 in females) (Fig. 3), and lipid stores
were repleted during the course of the summer.
Analysis of the body composition of char re-entering
fresh water at the end of the summer revealed that
the fat:LBM ratio of both males (0.063) and females
(0.072) (Fig. 3) was within the range of those of
fully fed, hatchery-reared juvenile char (0.063±
0.079) (Fig. 2). Thus, even though circumstantial,
this evidence seems to suggest that the wild,
anadromous char ceased feeding in the sea and re-
entered fresh water once their lipid reserves were
repleted.
Preliminary results of a study carried out on post-
smolt Atlantic salmon Salmo salar L. (initial weight
» 75 g) reveal changes in body composition that
may occur in ®sh undergoing complete compensa-
tion after a period of feed restriction (S J S Johansen,
M Ekli & M Jobling, pers. comm.). Fully fed ®sh were
fed in excess throughout the 16-week trial. For the
®rst 8 weeks, the restricted group was fed half the
ration predicted to support maximum growth and,
during the second half of the trial, these ®sh were
given an unlimited feed supply.
During the restriction phase, the increase in body
weight of the feed-restricted ®sh was only 65% of
that of the fully fed controls. There were differences
between the groups in both LBM (controls 157 g;
restricted 126 g) and proximate chemical composi-
tion (fat:LBM of controls 0.080; restricted 0.066)
(Fig. 4).
By the end of the trial, the ®sh in both groups
weighed » 275 g. In other words, during the
recovery period, the restricted group had increased
growth rate and fully compensated for the poorer
growth during the ®rst half of the trial. The
differences in LBM seen at the end of the feed-
restriction phase had disappeared (LBM controls
252 g; restricted 258 g), and differences in relative
lipid content had been reduced (fat:LBM controls
0.094; restricted 0.087).
Taken together, these observations indicate that
feed restriction results in ®sh with a lower fat
content and suggest that the rate at which body fat
is accumulated may have some regulatory role
during catch-up growth. This seems to be in
accordance with the lipostatic model. On the other
hand, there are some reports that ®sh with fully
L
Figure 3 Body weights and compositions of wild,
anadromous male and female Arctic char Salvelinus
alpinus L. during spring descent to sea water (Descend)
and during the return to fresh water in late summer
(Ascend). LBM, lean body mass. Recalculated from data
in Jùrgensen, Johansen & Jobling (1997).
476 # 1999 Blackwell Science Ltd, Aquaculture Research, 30, 473±478
Hyperphagia and catch-up growth M Jobling & S J S Johansen Aquaculture Research, 1999, 30, 473±478

repleted lipid reserves continue to display catch-up
growth (Bull & Metcalfe 1997; Nicieza & Metcalfe
1997), something that would con¯ict with the
lipostatic model. However, in these latter studies,
lipid content was not monitored directly, but was
estimated using a series of morphometric measure-
ments (Simpson, Metcalfe & Thorpe 1992). When
®sh are undergoing catch-up growth and are
envisaged to be depositing reserves and structural
tissues differently from individuals that are growing
normally, it is open to question whether biometry
will provide an adequate assessment of the size of
the body lipid reserves. Thus, the evidence may be
equivocal, and further tests are required.
Conclusions
The size of the body lipid reserves in¯uences feed
intake in mammals via negative feedback signals to
the central nervous system, and this forms the basis
of the lipostatic model for the regulation of ingestive
behaviour, body weight and body composition. The
size of the body fat stores may have an in¯uence on
feeding in salmonid ®shes, and there is some
evidence for lipostatic involvement in the regulation
of the hyperphagic response seen in ®sh that are
undergoing catch-up growth after a period of
nutritional deprivation. Although a model incorpor-
ating lipostatic regulation would seem to offer a
simple and attractive explanation for the changes in
feed intake and growth observed in ®sh undergoing
catch-up growth, more rigorous testing of the model
is required via studies in which changes in feed
intake, body mass and tissue composition are
examined simultaneously.
References
Bejda A.J., Phelan B.A. & Studholme A.L. (1992) The effect
of dissolved oxygen on the growth of young-of-the-year
winter ¯ounder, Pseudopleuronectes americanus.
Environmental Biology of Fishes 34, 321±327.
Blum W.F. (1997) Leptin: the voice of the adipose tissue.
Hormone Research 48 (Suppl. 4), 2±8.
Broekhuizen N., Gurney W.S.C., Jones A. & Bryant A.D.
(1994) Modelling compensatory growth. Functional
Ecology 8, 770±782.
Bull C.D. & Metcalfe N.B. (1997) Regulation of hyperpha-
gia in response to varying energy de®cits in over-
wintering juvenile Atlantic salmon. Journal of Fish
Biology 50, 498±510.
Cabanac M. & Richard D. (1996) The nature of the
ponderostat: Hervey's hypothesis revived. Appetite 26,
45±54.
Friedman J.M. (1998) Leptin, leptin receptors and the
control of body weight. Nutrition Reviews 56, S38±S46.
Harris R.B.S. & Martin R.J. (1984) Speci®c depletion of
body fat in parabiotic partners of tube-fed obese rats.
American Journal of Physiology 247, R380±R386.
Hayward R.S., Noltie D.B. & Wang N. (1997) Use of
compensatory growth to double hybrid sun®sh growth
rates. Transactions of the American Fisheries Society 126,
316±322.
Hervey G.R. (1959) The effects of lesions in the hypotha-
lamus in parabiotic rats. Journal of Physiology 145,
336±352.
Hervey G.R. (1969) Regulation of energy balance. Nature
223, 629±631.
Hossner K.L. (1998) Cellular, molecular and physiological
aspects of leptin: potential application in animal
production. Canadian Journal of Animal Science 78,
463±472.
Jobling M. (1994) Fish Bioenergetics. Chapman & Hall,
London.
Jobling M. & Miglavs I. (1993) The size of lipid depots ± a
factor contributing to the control of food intake in Arctic
R
Figure 4 Body weight and composition of post-smolt
Atlantic salmon Salmo salar L. during 16 weeks of full
feeding (control: sampling on 23 July and 18 September)
or 8 weeks on a restricted ration followed by 8 weeks on
full rations (restricted: sampling at the end of the
restriction phase on 23 July and on 18 September). LBM,
lean body mass. Unpublished data from S J S Johansen,
M Ekli & M Jobling.
# 1999 Blackwell Science Ltd, Aquaculture Research, 30, 473±478 477
Aquaculture Research, 1999, 30, 473±478 Hyperphagia and catch-up growth M Jobling & S J S Johansen

charr, Salvelinus alpinus? Journal of Fish Biology 43,
487±489.
Jobling M., Melùy O.H., dos Santos J. & Christiansen B.
(1994) The compensatory growth response of the
Atlantic cod: effects of nutritional history. Aquaculture
International 2, 75±90.
Jùrgensen E.H., Johansen S.J.S. & Jobling M. (1997)
Seasonal patterns of growth, lipid deposition and lipid
depletion in anadromous Arctic charr. Journal of Fish
Biology 51, 312±326.
Kennedy G.C. (1953) The role of depot fat in hypothalamic
control of food intake in the rat. Proceedings of the Royal
Society B 140, 578±592.
Matson C.A., Wiater M.F. & Weigle D.S. (1996) Leptin and
the regulation of body adiposity. Diabetes Reviews 4,
488±508.
Metcalfe N.B. & Thorpe J.E. (1992) Anorexia and defended
energy levels in over-wintering juvenile salmon. Journal
of Animal Ecology 61, 175±181.
Miglavs I. & Jobling M. (1989a) Effects of feeding regime on
food consumption, growth rates and tissue nucleic acids
in juvenile Arctic charr, Salvelinus alpinus, with parti-
cular reference to compensatory growth. Journal of Fish
Biology 34, 947±957.
Miglavs I. & Jobling M. (1989b) The effects of feeding
regime on proximate body composition and patterns of
energy deposition in juvenile Arctic charr, Salvelinus
alpinus. Journal of Fish Biology 35, 1±11.
Nicieza A.G. & Metcalfe N.B. (1997) Growth compensation
in juvenile Atlantic salmon: responses to depressed
temperature and food availability. Ecology 78, 2385±
2400.
Nishizawa Y. & Bray G.A. (1980) Evidence for a
circulating ergostatic factor: studies on parabiotic rats.
American Journal of Physiology 239, R344±R351.
Russell N.R. & Wootton R.J. (1992) Appetite and growth
compensation in the European minnow, Phoxinus
phoxinus (Cyprinidae), following short periods of food
restriction. Environmental Biology of Fishes 34, 277±
285.
Schwartz M.W. & Seeley R.J. (1997) The new biology of
weight regulation. Journal of the American Dietetic
Association 97, 54±58.
Shearer K.D., Silverstein J.T. & Plisetskaya E.M. (1997)
Role of adiposity in food intake control of juvenile
chinook salmon (Oncorhynchus tshawytscha).
Comparative Biochemistry and Physiology A 118, 1209±
1215.
Simpson A.L., Metcalfe N.B. & Thorpe J.E. (1992) A simple
non-destructive biometric method for estimating fat
levels in Atlantic salmon, Salmo salar. Aquaculture and
Fisheries Management 23, 23±29.
Weatherley A.H. & Gill H.S. (1987) The Biology of Fish
Growth. Academic Press, London.
Weigle D.S. (1994) Appetite and the regulation of body
composition. FASEB Journal 8, 302±310.
L
478 # 1999 Blackwell Science Ltd, Aquaculture Research, 30, 473±478
Hyperphagia and catch-up growth M Jobling & S J S Johansen Aquaculture Research, 1999, 30, 473±478