the leydig cell in larval amphibian epidermis. fine structure and function
TRANSCRIPT

The Leydig Cell in Larval Amphibian Epidermis FINE STRUCTURE AND FUNCTION
DOUGLAS E. KELLY Department of Biological Structure, University of Washington, Seattle, Washington
ABSTRACT The large, presumably mucus and/or fluid secreting cells of Leydig have been studied in the epidermis of the newt, Taricha torosa, by light and electron microscopy. Mitotic activity in visibly differentiated Leydig cells results in their in- creasing numbers so that by mid-larval stages they comprise the major cellulax com- ponent of tailfin skin. Subsequently, the number of these cells diminishes as the epidermis thickens; their final disappearance coincides with metamorphosis and epi- dermal cornification. During the course of larval life, clear vesicles within Leydig cell cytoplasm accumulate granular material and, in time, assume the morphology of typical mucous aggregates. Concomitantly, the cytoplasm progressively becomes clear in appearance and nearly devoid of organelles.
The surface of larval skin is normally provided with a thin mucous coat by con- tinual production on the part of apical epidermal cells. When this surface is exposed to air, observation over a period of time reveals that the outer mucous coat hardens rapidly. There is, no indication that additional fluid or mucus is provided to the drying surface from Leydig cells. Rather, light and electron micrographs of epidermis after various degrees of desiccation suggest that Leydig cells do respond to drying, but do so by adding fluid and perhaps mucous material to subsurface, extracellular compartments of the epidermis. The concept that Leydig cells provide an internal fluid reserve is discussed in relaticn to epidermal development and prevailing phys- iological and structural evidence bearing on intercellular compartments.
Leydig cells are conspicuous components of the epidermis in larval amphibians. Since their first description (Leydig, 1857), these cells have attracted attention be- cause of their large diameter (often ex- ceeding 25 LI), their clear watery cytoplasm with presumed mucous inclusions, and their unique peripheral supportive network (the net of Langerhans, 1873) (see review of Dawson, '20).
In the single prior consideration of Leydig cells based on electron microscop- ical evidence, Hay ('61) was able to show that in the skin of larval salamanders, -4mblystoma punctatum and A. opacunz, the dense bands comprising Langerhans' net are in reality bundles of much finer ( ca . 50 A ) filaments. Hay noted the simi- larity of these filaments to tonofilaments of surrounding epidermal cells, and described the convergence of filaments from the net toward desrnosomes, facts which suggested a supportive role for the net. She was also able to demonstrate in methacrylate sec- tions the periodic acid-Schiff positivity of the large cytoplasmic granules of Leydig cells, thus strengthening the supposition
ANAT. REC., 154: 685-700
of light microscopists (e.g., Dawson, '20) that these cells are involved in mucus pro- duction.
Despite this information, i t has not been ascertained that Leydig cells actually do release mucus or other materials in any appreciable or regular fashion. Nor is in- formation available as to what factors might evoke Leydig cell secretion, or what direction such secretion might take.
On the assumption that Leydig cell func- tion in the newt, Taricha torosa, might relate to an epidermal fluid reserve, ex- periments were designed to test the re- sponse of these cells to varying degrees of dehydration. The experimental data are discussed in this report in relation to a number of fine structural and develop- mental aspects.
METHODS AND MATERIALS
Larvae of the west coast newt, Taricha torosa, were obtained from breeding sites near Stanford, California. While Leydig cells are contained in larval skin from most body regions, lateral surfaces of the dorsal tailfin midway between vent and
685

686 DOUGLAS E. KELLY
tailtip were the routine source of epidermis for this study. Tailfin samples for light and electron microscopy were excised from MS-222 anesthetized (Tricaine methane- sulfonate, Sandoz Pharmaceuticals) newt larvae and fixed as small blocks in ice-cold 3.75% osmium tetroxide in 0.05 M s-col- lidine buffer (Bennett and Luft, '59) for 1-2 hours. Other blocks were fixed in 0.5% glu- taraldehyde or 0.25% acrolein plus 0.25% glutaraldehyde in a half strength Niu- Twitty solution (Niu and Twitty, '53) fol- lowed by postosmication using the same solvent. These aldehyde fixation media have a pH of 7.3-7.5 and a milliosmolarity of 112-132. After dehydration in an ethanol series, the blocks were embedded in Epon 812 (Luft, '61), and sectioned on a Porter-Blum MT-2 ultramicrotome using glass and diamond knives. Thick (1-2 w) sections were stained with methylene blue- azure I1 (Richardson et al., '60) and examined and photographed by light mi- croscopy. Thin sections for electron micros- copy were stained with half-saturated aqueous uranyl acetate, 8-10 minutes at 55"C, followed by 8-10 minutes treatment with alkaline lead citrate (Reynolds, '63). Electron micrographs were taken on a Siemens Elmiskop I operating at 60 KV with voltage alignment and compensated lateral and rotational chromatic aberra- tions (Luft, '63).
Examinations were made on normal epi- dermis at various ages of larval and post- metamorphic life (hatching to 20 weeks). In addition, light and electron microscop- ical observations were made on larval epi- dermis subjected to drying for various periods of time (1-20 minutes). This was accomplished by placing an anesthetized larva in a drop of pond water on a wax plate so that the tailfin floated on the sur- face of the drop. This exposed one lateral surface of the tailfin to air (at about 37°C) and left the other surface immersed. The exposed surface could be observed with a dissecting microscope during the course of drying and the viscosity of materials on the drying surface could be estimated by stroking the surface with a fine hairloop. When fixed after given periods of this treatment, the "wet" and "dry" sides of a given tailfin could be compared micro-
scopically to each other and to normaI, un- treated epidermis of the same age.
OBSERVATIONS
Normal fine structure and development In histological sections, Leydig cells are
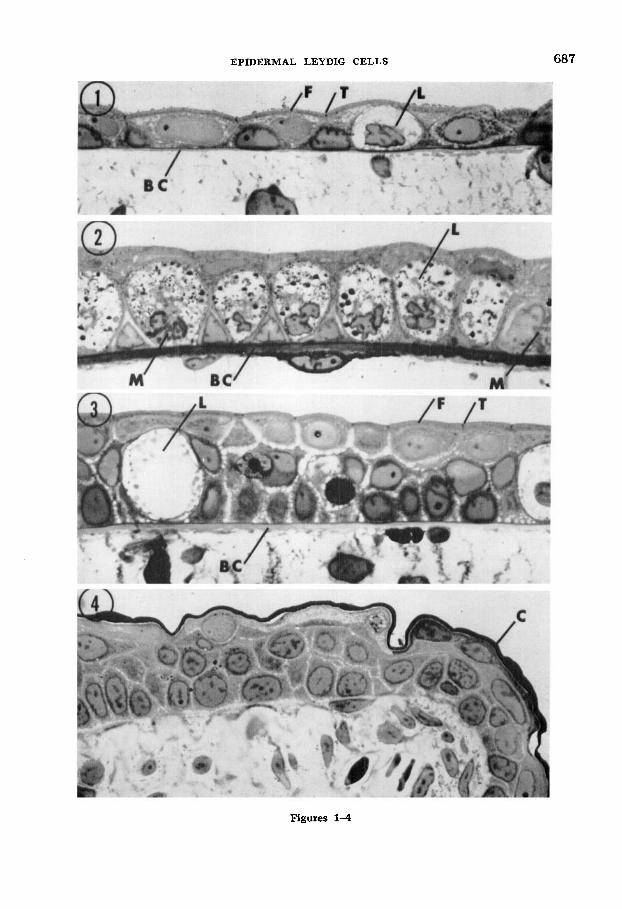
readily recognizable at all larval ages by their size and relative lack of cytoplasmic structure. However, in the tailfin there is a notable differential in the number of these cells per unit area of skin at differ- ent ages (figs. 1-3). From hatching stages until three weeks thereafter, the lateral tailfin epidermis is essentially two cell layers thick and it contains few, widely scattered Leydig cells sandwiched between the main layers. Subsequently, Leydig cells proliferate rapidly so that by mid- larval stages (8-12 weeks) they form a nearly continuous layer between the apical and basal epidermal cells. This seems to be the peak density of Leydig cells, for as the larva approaches metamorphosis (nor- mally 14-18 weeks) and the epidermis thickens, the Leydig cells again become widely scattered, After metamorphosis and keratinization of outer epidermal layers, Leydig cells are lacking (fig. 4).
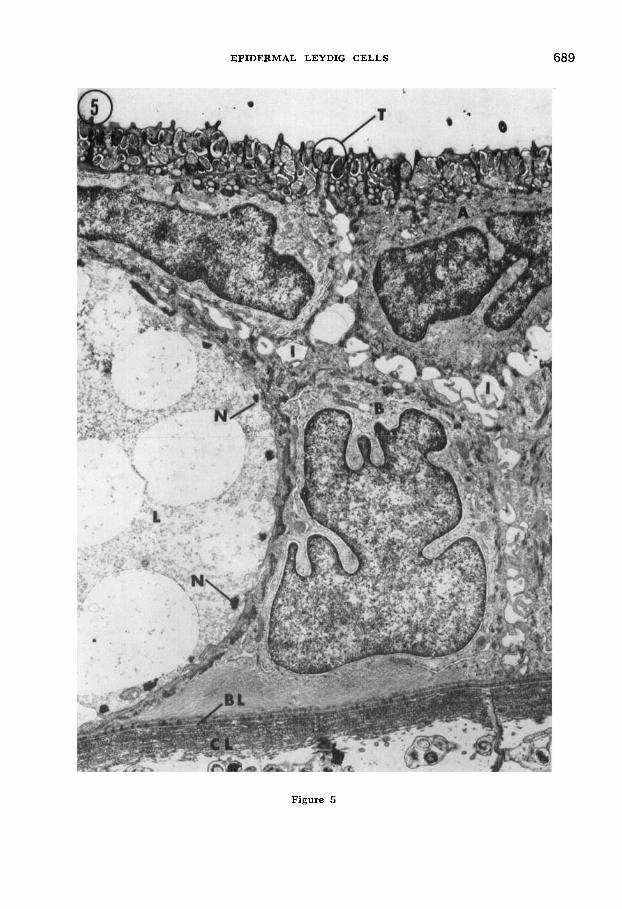
Figure 5 is a survey electron micrograph of a section of epidermis from a six-week- old larva illustrating the main cell types and their relationships. A portion of a Leydig cell (L) displays the relatively structureless, yet flocculent cytoplasm which is typical of Leydig cells at 3-6 weeks. Large, clear, membrane-bounded vesicles occupy much of the cytoplasm, and the dense filament bundles of the
Figs. 1-4 Light micrographs of Richardson- stained, Epon-embedded epidermis sections from the dorsal tailfins of newts after three (fig. l ) , 10 (fig. 2) , and 16 (fig. 3) weeks of larval life, and one month past metamorphosis (fig. 4). Frequency of Leydig cells (L) increases markedly during mid-larval stages when mitotic figures ( M ) are common in both Leydig and other epi- dermal cells. At later larval stages, Leydig cells are sparse and the epidermis thickened. After metamorphosis and cornification ( C ) of outer epidermal cells, Leydig cells are lacking. Note the basement membrane complex ( B C ) which separates epidermis and mesenchymal compo- nents, and which increases in thickness by mid- larval stages. Terminal bars (T) and the mu- cinogenic free border ( F ) are visible along larval apical epidermal cells. All sections osmium-fixed and 500 x.
EPIDERMAL LEYDIG CELLS 687
Figures 1 4

688 DOUGLAS E. KELLY
Langerhans net ( N ) are visible along the periphery of the cell. An apical epidermal cell ( A ) which displays a mucinogenic free border packed with mucous vesicles, and a basal epidermal cell ( B ) lie adjacent to the Leydig cell. The basal cell directly overlies a complex composed of an adepi- dermal globular layer, a basal lamina, and some twenty or more collagenous lamellae (all of which collectively comprise the light microscopists' basement membrane in this system). The various cells are tied to- gether and to the adepidermal complex by a system of tight and intermediate junc- tions, desmosomes, and hemidesmosomes. Apical epidermal cells are attached to each other along the outer surface of the skin by tight and intermediate junctions (com- ponents of the terminal bar [T in fig. 51 or junctional complex [Farquhar and Palade, '63, '651 ) . Hemidesmosomes are limited to basal epidermal cells where these latter border the adepidermal complex (fig. 8). A detailed analysis of these attachment mechanisms and adepidermal layers is the subject of a separate report (Kelly, '66).
So far as the present study can disclose, Leydig cells are always completely sur- rounded by apical and basal epidermal cells (figs. 1-7). In no case have Leydig cells been observed to possess a free, ex- posed apical border or a basal surface abutting directly on adepidermal compo- nents. Even when the number of Leydig cells is at its highest, the apical and basal surfaces of these cells are always sepa- rated from basal lamina on the one side, and the outer environment on the other, by thin extensions of apical and basal cells (figs. 6, 7). Since apical cell extensions at the skin surface seem always to be ad- joined by tight junctions, the possibility for surface exposure of Leydig cells is re- mote and has not been observed. The only junctional mechanisms in which Leydig cells themselves participate are the desmo- somes which join them to neighboring apical and basal cells. Terminal bars or hemidesmosomes have not been observed associated with Leydig cells.
Hay ('61) has already outlined the pre- sumably supportive association between the desmosomes and the filaments com- prising the Langerhans net. It can be added from the present study that the
dense cytoplasmic plaques and other components of Leydig cell desmosomes have the same general arrangement found in other epidermal desmosomes. Most of the filaments approaching a plaque from the Langerhans net arch or loop past the plaque and return into the net. This course is best described as a filament ex- cursion in and out of the plaque vicinity rather than a termination or ending of a filament at the plaque. Significance of this type of desmosome architecture is dis- cussed elsewhere (Kelly, '66).
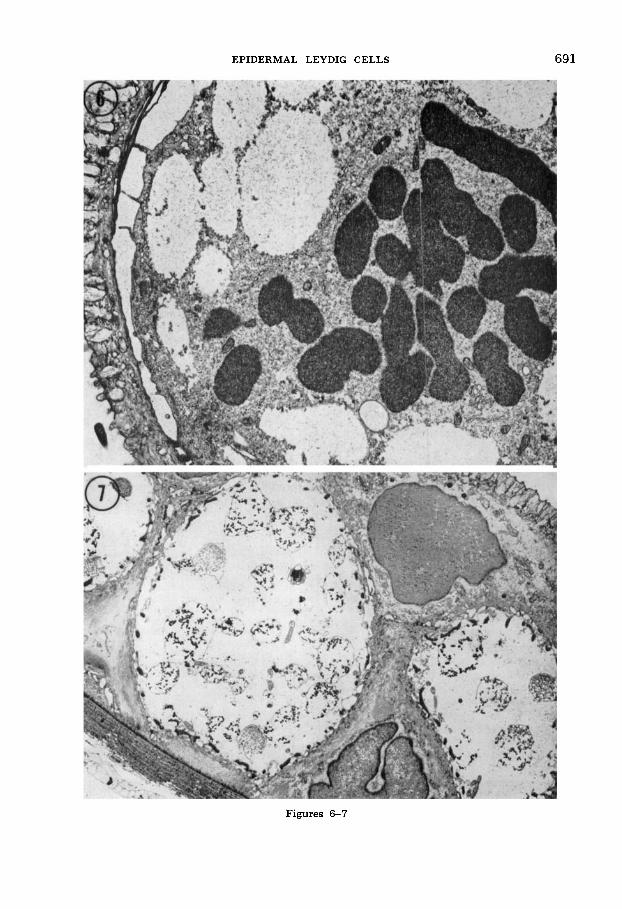
Leydig cells commonly display mitotic figures, as do apical and basal epidermal cells ( M in fig. 2), a fact noted also by previous workers (see review of Dawson, '20). Mitotic Leydig cells are particularly abundant during young and mid-larval stages when the number of Leydig cells increases markedly. The mitotic figures occupy cytoplasm which possesses all the ultrastructural properties of large differ- entiated Leydig cells (flocculent cytoplasm, Langerhans net, vesicles, desmosomes, etc.) (fig. 6). There is no indication of cytoplasmic dedifferentiation during Ley- dig cell mitosis, or that an undifferentiated stem cell is the source of new Leydig cells.
The fine structure of Leydig cells changes markedly and progressively dur- ing the course of larval life. Between hatching stages and 4-6 weeks, the cyto- plasm is rather densely flocculent and con- tains a few cisternae, ribosomes, and other organelles (figs. 5, 6). The numerous large membrane-bounded vesicles are virtually clear. Subsequently, during mid-larval stages, the relative contrast between cyto- plasm and vesicles reverses so that the
Fig. 5 Low magnification electron micrograph of a thin section of normal four-week larval tailfin epidermis. Apical ( A ) and basal ( B ) epi- dermal cells are visible along with a portion of a Leydig cell (L) . The latter displays clear, sparsely granular vesicles and a flocculent cyto- plasm at this age. Portions of the Langerhans net ( N ) are also visible. Note the vesicular mucinogenic free border of the apical cells (top) and the collagenous lamellae (CL) which, along with a basal lamina ( B L ) composes the basement membrane complex (bottom). T denotes a region of terminal bar components between adjacent apical cells. Relatively wide intercellular com- partments ( I ) are bridged by desmosome-bearing cell processes. Acrolein/glutaraldehyde fixation with uranyl acetate/lead stain. 5,600 x.
EPIDERMAL LEYDIG CELLS 689
Figure 5

690 DOUGLAS E. KELLY
cytoplasm becomes nearly clear and only lightly flocculent while the contents of the vesicles gain added density (figs. 7, 8). Moreover, mid-larval vesicle contents dis- play a variety of forms ranging from coarse, dense, loosely aggregated granular material in some vesicles to closely packed, fine, ma- terial in others (fig. 8). Often the surround- ing membranes of the vesicles are broken, probably as a preparative artifact, and the granular contents are scattered through the cell cytoplasm. The coarse, dense granules appear to be common in younger mid-lar- Val stages (fig. 7), while the finer, closely packed variety becomes predominant toward late larval life (fig. 9).
In older stages the vesicles closely re- semble mucous vesicles or granules known from other systems (e.g., mucous goblet cells), and they are the periodic acid- Schiff positive components which previous workers have noted (see Hay, '61). Mem- branes around the presumed mucous in- clusions are usually disrupted. At this stage only that part of the cytoplasm near the Leydig cell nucleus contains an appre- ciable number of organelles ( a scant amount of Golgi membranes and endo- plasmic reticulum, plus widely scattered mitochondria). The ground substance of the cytoplasm appears clear and watery, containing only scattered flocculent mate- rial. This material matches in its distribu- tion and density a similar or identical sub- stance in extracellular spaces. Indeed, late larval Leydig cell ground cytoplasm is virtually indistinguishable from the extra- cellular fluid matrix, a fact noted previ- ously by Hay ('61).
Leydig cell response to d y i n g
visible. The layer is quite sticky when touched with a hairloop and is probably derived from continual production on the part of the mucinogenic border of apical epidermis cells. Within the first minute of exposure to air, the surface becomes vis- cous and adhesive to objects coming into contact with it, but after five minutes of drying the surface is hard and no longer sticky. A hairloop may be brushed freely over the surface with no tendency for adhe- sion. With continued drying the surface of the skin gradually becomes furrowed in a vaguely hexagonal pattern so that after ten minutes it is visibly dry and wrinkled, and after 20 minutes it is best described as grossly shriveled. These changes occur even though the contralateral side of the tailfin has remained immersed in pond water.
At no point during these observations is there any visible evidence of the release of moist or mucoid materials onto the sur- face of the skin beyond the thin mucous layer initially present when environmental water recedes.
Light and electron microscopical exam- ination of skin subjected to drying reveals the profound morphological effects of even relatively brief exposure to such treatment. Figure 10 is a photomicrograph of a sec- tion through the tailfin of a four-week-old larva. One side of the tailfin has been dried for five minutes. The dry side (bot- tom) is thin, quite densely stained, and displays Leydig cells ( L ) whose nuclei and cytoplasmic volumes are markedly reduced.
Fig. 6 A portion of a Leydig cell from the epidermis of a four-week larva. The cell contains mitotic chromosomes plus the clear vesicles, floc- culent cvtodasm, and elements of a Laneerhans
If one that mucous products net (lower ieft) which are typical of Leydig cells at this age. Note that a thin partition of vesicu-
from Leydig are destined for lar apical cell cytoplasm (left) separates the as a protective Coat onto the Outer Surface Leydig cell from the outside environment. Extra- of the skin (Dawson, '20) , such release cellular compartments are relatively clear, but might be induced by exposure of the &in contain flocculent material similar in appearance
to that of Leydig cell vesicles. Osmium fixation surface to air. This possibility was tested with uranyl acetate/lead stain. 5,000 x. by visual, light microscopic, and electron
Fig. 7 Low magnification electron micrograph microscopic observation. of the epidermis from a ten-week larva. At this age Leydig cells form a massive, though not
of a mid-larval newt is observed with a continuous, layer between apical (upper right) dissecting microscope, an inevitable series and basal cells. Leydig cell cytoplasm is clear by of events can be discerned. As water re- comparison to earlier ages, and most Leydig cell
vesicles contain coarse, dense, granular material. cedes from the surface, a thin, glistening, Osmium fixation with uranyl acetate/lead stain. previously existent mucous layer becomes
When an area Of
1,850 X.
EPIDERMAL LEYDIG CELLS
Figures 6-7
691

692 DOUGLAS E. KELLY
Leydig cells of the opposite, wet side ap- pear relatively normal with the exception that some are noticeably vacuolated (center). As will be seen, the vacuoles are, in fact, extracellular spaces (E). The rest of the wet epidermis appears normal.
The increased density and shrinkage oc- curring in air-exposed epidermis becomes evident shortly after initial exposure. In the present series of examinations, some shrunken, dense, individual apical cells could be discerned in epidermis exposed to air for as little as one minute. Leydig cells, however, generally appear unaffected by this short interval of drying. By five min- utes (fig. l o ) , virtually all the cells, in- cluding Leydig cells, are involved to the extent that total thickness of the drying epidermis is less than half that of the op- posite side. By ten minutes, the shrinkage reaches maximum. Leydig cells are no longer distinguishable in thick sections, and the drying is beginning to affect visibly the morphology of even the epidermal cells of the wet contralateral side.
Figure 11 is a light micrograph of a tailfin section after ten minutes exposure of one surface (bottom) to air. The ef- fects of drying are manifest not only in the density and lack of recognizable Leydig cells in the dried surface, but also in the appearance of a similar density and shrink- age especially in the basal cells of the op- posite, wet side of the tailfin. Moreover, drastic changes have taken place in the Ley- dig cells (L) of the wet side. They appear as only thin silhouettes occupying wide, round spaces in the epidermis. Figures 12 and 13 show portions of these same epidermal surfaces in adjacent thin sections exam- ined by electron microscopy. The dried side (fig. 12) displays a dense mucous coat ( M L ) over its exposed surface. The density of this coat probably corresponds to the hardened character noted for it in vis- ual examination of dried surfaces. All cells of the dried epidermis are distinctly con- densed; cytoplasmic ground substance is lacking and membranous components are compressed into a minimal cellular space. Nuclei are also condensed in appearance, and Leydig cells (L) are barely detectable by their relative position between apical and basal cells. The cytoplasm of Leydig cells is minimal in volume and virtually
indistinguishable from that of surrounding cells. Mesenchymal cells beneath the epi- dermis show similar effects of desiccation.
The Leydig cells of the opposite, wet side (fig. 13) are particularly interesting be- cause they display a phase of response which seems to pass very quickly on the rapidly drying exposed side. While the cytoplasmic volume of these cells has di- minished giving each a compressed, strand-like image in the section, the surrounding extracellular space (E) has enlarged in a complementary fashion. The large, round profile of the space oc- cupied by the Leydig cell has largely been retained, but it is filled by apparent extra- cellular fluid. This latter material is floc- culent in the fixed specimen. The Leydig cell proper is left with a compact nucleus toward which the cell membrane has pressed. Cytoplasmic organelles, including Golgi membranes, mitochondria, and ribo- somes, have remained within the cell mem- brane, indicating its integrity. A few, ap- parently mucus-containing vesicles (M) also remain in the cytoplasmic compart- ment.
Various degrees of this response by Leydig cells to drying have been seen in the present study; the configuration in fig- ure 13 seems to represent a mid-point in the progression of this response. Extended
~~
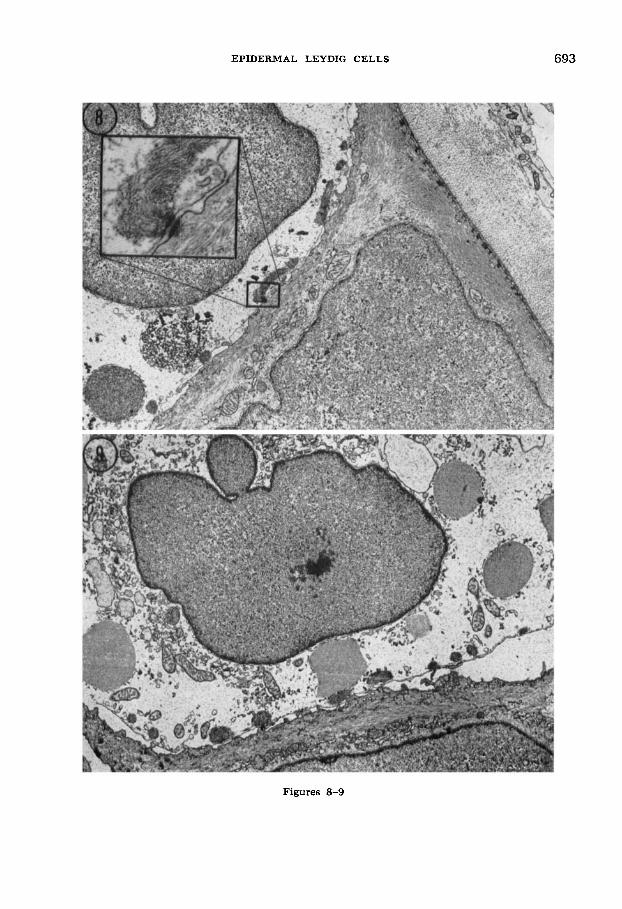
Fig. 8 Basal region of epidermis from the tailfin of a 12-week newt larva. Portions of a Leydig cell (top left) and a basal epidermal cell (bottom) are visible as well as the basal lamina and underlying collagenous lamellae (upper right). Both coarse and finely granular vesicles are present in the Leydig cell a t this stage. The participation of filaments from the Langerhans net with a desmosome joining the Leydig cell to the adjacent basal cell is shown in the inset. Note the clear, flocculent cytoplasm of the Leydig cell by comparison to the filamentous appearance of the basal cell. Hemidesmosomes are visible in the basal cell where the latter borders adepi- dermal components. Osmium fixation with uranyl acetate/lead stain. 6,200 X.
Fig. 9 The nucleus-containing portion (ca. 1/6 the cross-sectional area) of a Leydig cell from the tailfin epidermis of a 16-week larva. Vesicles within the Leydig cell are predominantly finely granular and presumably correspond to mucous granules. Note the scant rough and smooth endoplasmic reticulum near the nucleus, and smaller, less densely packed vesicles. Leydig cell background cytoplasm is virtually indistinguish- able from extracellular material. Osmium fixa- tion with uranyl acetate/lead stain. 5,300 X.
EPIDERMAL LEYDIG CELLS
Figures 8-9
693
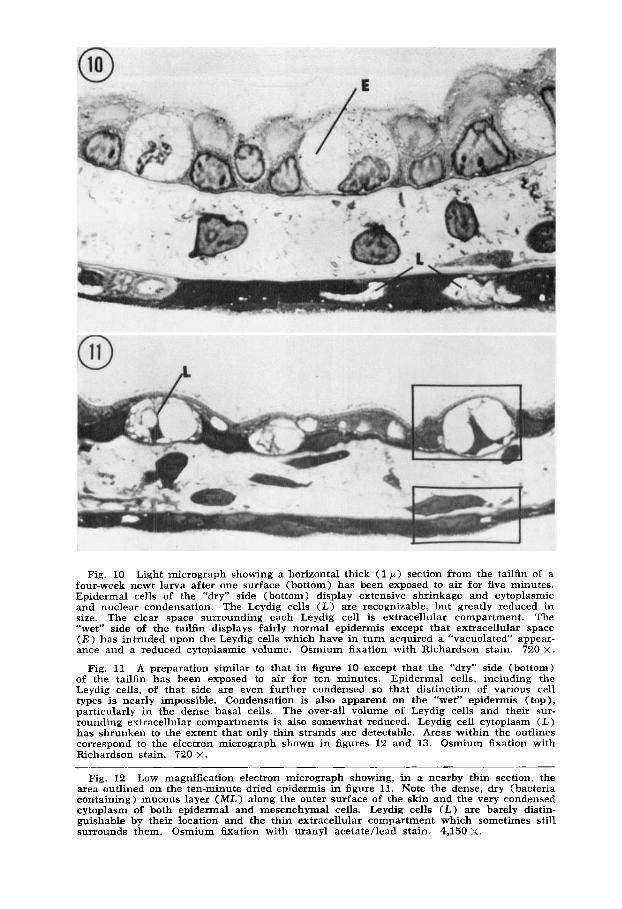
Fig. 10 Light micrograph showing a horizontal thick (1 p ) section from the tailfin of a four-week newt larva after one surface (bottom) has been exposed to air for five minutes. Epidermal cells of the “dry” side (bottom) display extensive shrinkage and cytoplasmic and nuclear condensation. The Leydig cells (L) are recognizable, but greatly reduced in size. The clear space surrounding each Leydig cell is extracellular compartment. The “wet” side of the tailfin displays fairly normal epidermis except that extracellular space ( E ) has intruded upon the Leydig cells which have in turn acquired a “vacuolated” appear- ance and a reduced cytoplasmic volume. Osmium fixation with Richardson stain. 720 X.
A preparation similar to that in figure 10 except that the “dry” side (bottom) of the tailfin has been exposed to air for ten minutes. Epidermal cells, including the Leydig cells, of that side are even further condensed so that distinction of various cell types is nearly impossible. Condensation is also apparent on the “wet” epidermis (top), particularly in the dense basal cells. The over-all volume of Leydig cells and their sur- rounding extracellular compartments is also somewhat reduced. Leydig cell cytoplasm ( L ) has shrunken to the extent that only thin strands are detectable. Areas within the outlines correspond to the electron micrograph shown in figures 12 and 13. Osmium fixation with Richardson stain. 720 X.
Fig. 11
Fig. 12 Low magnification electron micrograph showing, in a nearby thin section, the area outlined on the ten-minute dried epidermis in figure 11. Note the dense, dry (bacteria containing) mucous layer (ML) along the outer surface of the skin and the very condensed cytoplasm of both epidermal and mesenchymal cells. Leydig cells (L) are barely distin- guishable by their location and the thin extracellular compartment which sometimes still surrounds them. Osmium h a t i o n with uranyl acetate/lead stain. 4,150 X.
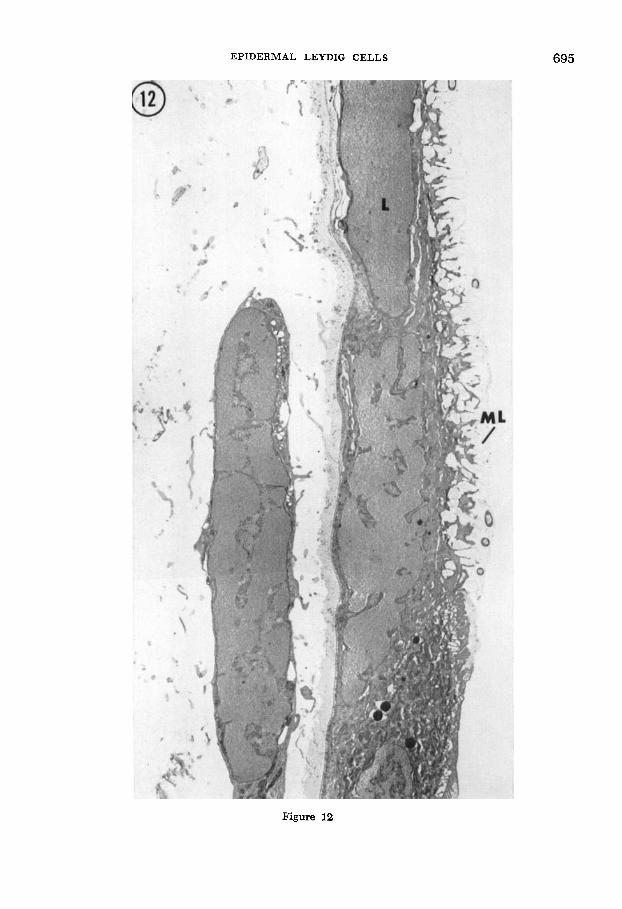
EPIDERMAL LEYDIG CELLS 695
Figure 12
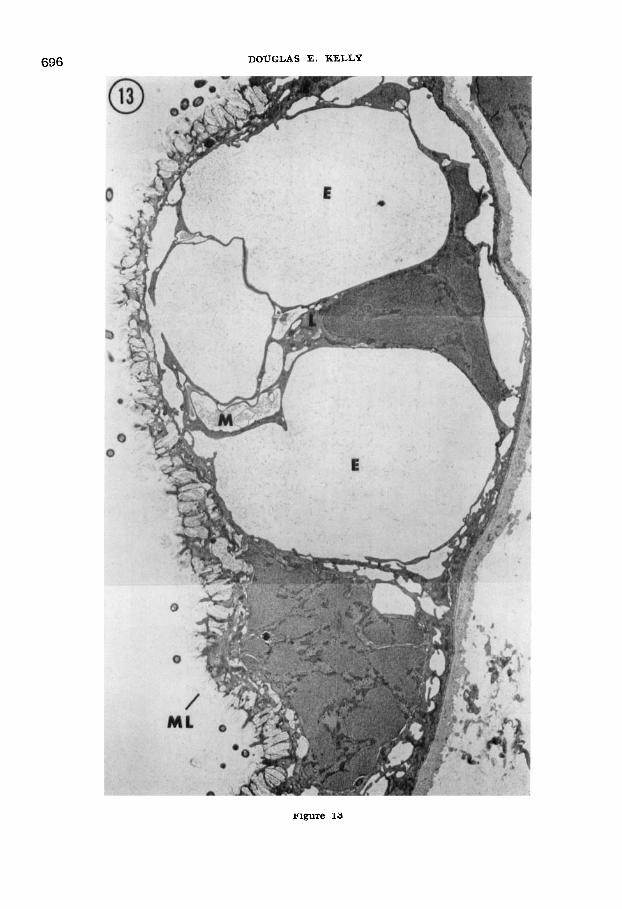
696 DOUGLAS E. KELLY
rigure 13

EPIDERMAL LEYDIG CELLS 697
drying (e.g., Leydig cells of the dry side after ten minutes [fig. 121, or those on the wet side after 20 or more minutes) results not only in collapse of the Leydig cell proper, but also collapse of the surrounding fluid-filled extracellular spaces. Micro- graphs of sections taken after lesser amounts of desiccating trauma reveal Leydig cells which display partial evacua- tion of apparent cytoplasmic fluid into the extracellular spaces (e.g., the “vacuoles” in
There is no evidence from any of the micrographs taken of epidermis from either wet or exposed sides of drying tail- fins to indicate that Leydig cells ever come into continuity with the surface of the skin. Neither is there evidence for the release of materials from the Leydig cells to the sur- face. In all epidermal surfaces which have been examined under normal or traumatic conditions, the apical epidermal cells, with their mucous vesicles and mucous coat, form a continuous cellular barrier inter- posed between the Leydig cells and the exterior environment.
fig. 10).
DISCUSSION
The observations reported above lend support to the concept (Dawson, ’20) that Leydig cells are uniquely differentiated ele- ments of larval skin which probably play a significant and heretofore unrecognized physiological role. Morphological evidence presently available suggests that these cells are specialized for production and storage of fluid and mucous materials. The extent of this specialization is realized in the large
Fig. 13 Low magnification electron micro- graph showing, in a thin section, the area out- lined on the “wet” side of the ten-minute dried tailfin in figure 11. The mucous layer ( M L ) covering the epidermal surface is dispersed rather than dry and dense as in figure 12. Cytoplasm of the Leydig cell ( L ) has clearly responded to drying of the opposite side of the tailfin by con- tracting into fine, organelle-containing strands. Large extracellular compartments (E) occupy the space formerly held by Leydig cell cytoplasm. These are mled with a sparsely flocculent ap- parent fluid similar to that in other remaining extracellular compartments, and similar also to the contents of normal, undried Leydig cell cyto- plasm and vesicles at this age (see figs. 5 and 6). A few vesicles, perhaps containing mucus (MI, remain within the Leydig cell cytoplasm. Osmium fixation with uranyl acetate/lead stain. 4,150 X.
size which these cells attain, their relative lack of cytoplasmic components other than fluid or mucous materials, and their char- acteristic peripheral supportive skeleton, the Langerhans net (Hay, ’61 ).
The quantity of Leydig cells and the morphology of their vesicles change rapidly during the progress of larval life and the changes are in apparent coordination with other aspects of skin differentiation within the same period. When their number in- creases in the skin, Leydig cells are per- haps remarkable for the degree to which they display cytoplasmic specialization while at the same time retaining a high capacity for mitotic proliferation. This mitotic capacity is, incidentally, manifest not only in the frequency of mitotic figures one sees in these cells, but also in the rapidity with which Leydig cell nuclei have been seen in this and other laboratories (Hay, personal communication) to incor- porate tritiated thymidine. Throughout mid-larval stages (from 6-14 weeks) the Leydig cell population assumes the appear- ance of a massive intermediate layer sand- wiched between apical and basal epider- mal cells (fig. 2), although it is seldom that one Leydig cell will directly abut an- other. Cytoplasmic volume of the Leydig cell population probably comprises the major epidermal component at this time. Moreover, the population is a proliferative one, adding new cells to the apparent layer concomitant with the acquisition of the new apical and basal cells, and in pace with general epidermal growth. Ulti- mately, however, the pace is not main- tained, and during the last few weeks of larval life, Leydig cells become increas- ingly scarce. Their decline appears to co- incide with the acquisition of additional epidermal cell layers beyond the two which characterize young to mid-larval stages. Final disappearance of Leydig cells occurs at the time of first cornification of the outermost layer and the onset of many other epidermal and adepidermal meta- morphic changes (Kelly, ’66).
The frequency with which Leydig cells have been referred to as “mucous cells” (or by equivalent terminology) (Langer- hans, 1873; Dawson, ’20; Hay, ’61) seems to imply a belief that these cells should be

698 DOUGLAS E. KELLY
engaged in a significant external release of mucoid material. Dawson (’20) who stud- ied these cells (which he termed “club cells”) in neotenous adult Necturus skin was inclined to favor a discharge directly onto the epidermal surface. Yet he also listed several alternate possibilities derived from his own observations and those in the older literature. These included : dis- charge coincident with and perhaps nec- essary to molting; discharge of a poison- ous secretion upon injury to the cell; or production of “intercellular secretions of value in keeping the skin in healthy con- dition” (as first suggested by Pfitzner, 1879).
It should be pointed out here that the Leydig cells seen in this newt species do contain probable mucous granules, but the number of these inclusions is scant by com- parison to those of known goblet cells or the mucous skin cells found in the ante- rior head region of the same larvae (Kelly, ’66). Perhaps a more characteristic fea- ture of Leydig cells is the clear, “watery” nature of their cytoplasm. This property is first manifest in the contents of the nu- merous large cytoplasmic vesicles. Later, as the vesicle contents gradually assume the morphology of mucous granules, the general, massive cytoplasm becomes clear and nearly devoid of organelles. A counter argument to the mucous secretion concept might be that the accumulation of electron lucent, presumably aqueous, material is related to the prime function of these cells, and the increasingly dense vesicles are a by-product.
A concept for Leydig cell function in- volving mucus or fluid release onto the outer surface of the skin cannot be sup- ported by the evidence presented in this study. Light and electron microscopical examination of larval skin at various stages and under normal or desiccating conditions have failed to disclose a single image of a Leydig cell having unobstructed access to the surface of the skin. The mucous layer normally coating the surface appears to be a product of the mucinogenic free border of the apical epidermal cells. It is a product which is rapidly depleted and hardened upon exposure to air. Judging from visual observation of drying skin, the
hardening of surface mucus commences within a minute after environmental water is withdrawn, and there is never eruption on the surface of mucous or fluid exudates such as might be expected from the deple- tion of Leydig cells.
Leydig cells, do, however, respond to the stimulus of drying, and light and electron microscopical observations disclose that this response occurs concomitant with the hardening of the outer mucous coat. It appears that the fluid, and perhaps mu- cous, contents of these cells are released into extracellular compartments beneath the epidermal surface. The subsequent re- duction in Leydig cell cytoplasmic volume occurs prior to the time when condensa- tion and shrinkage of the epidermis as a whole impinges upon the Leydig cell and its immediate extracellular environment. The result during early phases of drying is the progressive increase of extracellular volume around Leydig cells as they, in turn, contract. Fluid reserve provided by Leydig cells is not unlimited, however, and continued desiccation rapidly results in what appears to be destructive condensa- tion of all epidermal cells.
The above interpretations mean that a newt larva has a very limited capability to withstand prolonged exposure to air. But, perhaps contrary to expectation, this limited capability is not dependent upon the release of mucus or fluid in copious amounts over exposed body surfaces. In- stead, the physiological significance of Ley- dig cells seems to be in the provision of fluid reserve for the subsurface intercellu- lar pool. The larval newt appears to combat desiccation by providing the skin surface with a hardened, perhaps impervious, coat beneath which the fluid reserve is mobil- ized. After metamorphosis, a permanent cornified outer coat is maintained, and Leydig cells no longer exist as epidermal components.
The maintenance of the subsurface ex- tracellular fluid pool may well be of prime importance in functional regulation of vital activities performed by the skin. Various lines of evidence (see Farquhar and Palade, ’63, ’65; Ussing, ’63-’64) point to the compartmentalization of the epider- mis into intra- and extracellular pools as

EPIDERMAL LEYDIG CELLS 699
a basis for selective permeability and trans- port properties which amphibian skin is acknowledged to possess. Integrity of the extracellular pool appears dependent upon apical tight junctions which seal the pool from the external environment, and upon active “ion pumps” associated with cell membranes which border the extracellular compartments. In view of these facts, it is perhaps not surprising that a larval newt should respond to air exposure by acquiring a hardened mucoid “sealer” over his body surface while at the same time mobilizing Leydig cell products to assure, temporarily, the fluid requirements of the internal extracellular pool.
Plausibility of this proposal must be tempered by the realization that speaking of Leydig cell function in terms of the pro- vision of fluid or mucoid products is prob- ably a gross oversimplification. Fluid and mucous products for these cells have been interpreted from morphological images alone while in actuality little biochemical information is yet available. The accuracy of the proposals concerning Leydig cell function discussed in this report is depend- ent upon further confirmation along bio- chemical as well as structural lines.
ACKNOWLEDGMENT
This investigation was supported by a re- search grant (GB-1592) from the National Science Foundation. The author acknowl- edges with thanks the able technical as- sistance of Miss Anne-Marie Lindgren, the comments of Drs. Daniel Szollosi and George Odland who critically reviewed the manuscript, and the secretarial help of Mrs. Lucille Klein and Mrs. Doris Ringer.
LITERATURE CITED Bennett, H. S., and J. H. Luft 1959 s-Collidine
as a basis for buffering fixatives. J. Biophysic. Biochem. Cytol., 6: 113-114.
Dawson, A. B. 1920 The integument of Nec- turus maculosus. J. Morph., 34: 487-589.
Farquhar, M. G., and G. E. Palade 1963 Junc- tional complexes in various epithelia. J. Cell Biol., 17: 375412.
1965 Cell junctions in amphibian skin. J. Cell Biol., 26: 263-293.
Hay, E. D. 1961 Fine structure of an unusual intracellular supporting network in the Leydig cells of Amblystoma epidermis. J. Biophysic. Biochem. Cytol., 10: 457-463.
Kelly, D. E. 1966 Fine structure of desmo- somes, hemidesmosomes, and an adepidermal globular layer in developing newt epidermis. J. Cell Biol., 28: 51-72.
Langerhans, P. 1873 Ueber die Haut der Lame von Salamandra maculosa. Arch. Mikr. Anat., 9: 745-752.
Leydig, F. 1857 Lehrbuch der Histologie des Menschen und der Thiere, Hamm, G. Grotesche Buchhandlung (C. Miiller).
Luft, J. H. 1961 Improvements in epoxy resin embedding methods. J. Biophysic. Biochem. Cytol., 9: 409-414.
1963 Appearance of lateral chromatic aberration with metal-stained thin sections, and the compensation of this field error. J. Appl. Physics, 34: 2513.
Niu, M. C., and V. C. Twitty 1953 The differ- entiation of gastrula ectoderm in medium con- ditioned by axial mesoderm. Proc. Nat. Acad. Sci., 39: 985-989.
Pfitzner, W. 1879 Die Leydig’schen Schleim- zellen in der Epidermis der Larve von Sala- mandra macufosa. Dissertation, Kiel.
Reynolds, E. S. 1963 The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol., 17: 208-213.
Richardson, K. C., L. Jarett and E. H. Finke 1960 Embedding in epoxy resins for ultrathin sectioning in electron microscopy. Stain Tech- nol., 35: 313-325.
Ussing, H. H. 1963-64 Transport of electrolytes and water across epithelia. Harvey Lectures, Series 59: 1-30, Academic Press, New York and London.