the influence of postnatal handling on adult neuroendocrine and behavioural stress reactivity
TRANSCRIPT

Journal of Neuroendocrinology, 1999, Vol. 11, 925–933
The Influence of Postnatal Handling on Adult Neuroendocrine andBehavioural Stress Reactivity
P. Meerlo,* K. M. Horvath,*† G. M. Nagy,† B. Bohus* and J. M. Koolhaas**Department of Animal Physiology, University of Groningen, the Netherlands.†Neuroendocrine Research Laboratory, Department of Human Morphology and Developmental Biology, Semmelweis University of Medicine,Budapest, Hungary.
Key words: neonatal handling, stress, coping, anxiety, catecholamines, prolactin, corticosterone.
Abstract
Environmental stimuli during early stages of life can influence the development of an organism andmay result in permanent changes in adult behaviour and physiology. In the present study weinvestigated the influence of early postnatal handling on adult neuroendocrine and behaviouralstress reactivity in Wistar rats. Pups were subjected to handling from postnatal day 1–21. The youngwere taken from the nest every day for 15 min and each of the pups was handled separately.Control nests were left undisturbed. When the animals had reached an adult age of 3–4 monthsthey were individually housed and subjected to a series of tests to measure their stress reactivity. Inthe first experiment we established adult behavioural coping with stressors and anxiety in thefollowing series of tests: open field test, shock prod defensive burying test, elevated plus maze andconditioned fear test. Collectively, the data clearly indicate that handled animals are characterizedby a lower stress-induced anxiety. Yet, handled and control animals do not differ in their generalway of coping with stressors. Although the lower anxiety in handled animals is often reflected in ahigher activity, they are not more active per se. In a second experiment, animals were provided witha permanent jugular vein canula for repeated blood sampling to determine stress hormones:noradrenaline, adrenaline, prolactin and corticosterone. Animals were subjected to a novelty testand a conditioned fear test. The neuroendocrine response profile is consistent with the conclusionthat handled animals are less anxious than controls but are not different in their general strategy ofcoping with stressors. The handled animals showed an attenuated adrenaline, prolactin andcorticosterone response. Yet, in neither of the two tests there was a difference in noradrenalineresponse, a typical marker for an active coping strategy. Interestingly, the differences inneuroendocrine reactivity already appeared in response to a mild novelty challenge when there wereno clear behavioural differences yet. The neuroendocrine measures are in line with the behaviouraldata but more sensitively reflect the differences between handled and control animals.
The stress response that enables an organism to cope with of the HPA axis results in release of cortisol or corticosteronefrom the adrenal cortex. The increase in catecholaminergicthreatening and challenging environmental stimuli consists of
a complex set of autonomic, endocrine, and behavioural activity and HPA axis activity, in a complex interplay withvarious other neuroendocrine systems, regulates cardio-components. Two of the main neuroendocrine systems
involved in the integrated response to stressors are the vascular function and energy turnover, facilitates an adequatebehavioural response to the stressor, and serves to re-establishautonomic sympathetic nervous system and the hypothal-
amus-pituitary-adrenal (HPA) axis (1, 2). Activation of the homeostasis afterwards.It is well known that the physiological and behaviouralsympathetic nervous system results in release of noradrenaline
from sympathetic nerve terminals throughout the body and stress response to a given stimulus varies considerablybetween individuals. There seems to be a characteristicsecretion of adrenaline from the adrenal medulla. Activation
Correspondence to: Peter Meerlo, Department of Neurobiology and Physiology, Northwestern University, 2153 North Campus Drive, Evanston,IL 60208, USA (e-mail: [email protected]).
© 1999 Blackwell Science Ltd

926 Postnatal handling and adult stress reactivity
individuality in appraisal of the environment and the response conditioned fear test. The open field test was performed tomeasure general explorative and locomotor activity into stressful situations. Such interindividual variation may
originate from genetic factors (3, 4) but, in addition, there is response to novelty. The shock-prod defensive burying testis especially suitable to differentiate between animals with angrowing evidence that the adult stress reactivity can be
influenced by early developmental factors as well (5, 6). active or passive coping strategy. The elevated plus mazeand the conditioned fear test were used to measure non-In mammals, the period around birth is characterized
by dynamic developmental processes in brain and body. conditioned and conditioned anxiety response. In the secondexperiment we measured the noradrenaline, adrenaline, pro-Environmental stimuli during the early stages of life can
influence these processes and may result in physiological and lactin and corticosterone response in a novelty test and aconditioned fear test.behavioural changes that persist into adulthood. Research
concerning developmental influences on stress reactivity hasstarted decades ago with reports showing that adult rats
Materials and methodshandled during the first few weeks of life exhibit a markedly
Postnatal handlingreduced HPA axis response to a variety of stressors (seeThe day after birth, nests were mixed to randomize genetic variation and alle.g. (7–9)). Compared to non-handled controls, postnatallynests were reduced to eight pups (six males and two females). Subsequently,handled animals have a lower adult corticotropin-releasinghalf of the newly arranged nests were left undisturbed except for cage cleaninghormone (CRH) and adrenocorticotropin (ACTH) response,once a week (control nests) and half of the nests underwent handling treatment
they secrete less corticosterone and corticosterone levels from postnatal day 1–21. The handling procedure started with taking thereturn faster to baseline (6, 10). There is evidence that rat mother from the nest and temporarily placing her in an empty cage. Then,
all pups were individually placed in glass jars lined with paper towel in apups are especially sensitive to developmental influences onwarm water bath (32–34 °C) for 15 min. During this 15 min each pup wasthe HPA axis during the first few weeks after birth (11). Inhandled for 3 min. After 15 min the pups were returned to their home cage,
contrast to the well-established effect of postnatal handling followed by the mother. Daily handling took place at the end of the lighton HPA axis reactivity, surprisingly little, if any, attention phase. On day 24 the animals were weaned. The male pups remained group-
housed, six/cage, until testing at adult age. When the animals reached an agehas been given to sympathetic reactivity.of 3–4 months they were individually housed and subjected to a series ofOn the level of the behavioural response to stressors,tests to measure their neuroendocrine and behavioural stress reactivity.several studies have reported changes in handled animals thatExperiment 1: Behavioural stress reactivity and anxietysuggest lower anxiety. For instance, it was found that handled
animals show a decreased emotional reactivity to being In the first experiment, the rats were subjected to a series of tests to measuretheir anxiety and behavioural coping with stress. The experiment was per-captured and handled (12), explore more and defecate lessformed with 12 handled animals from six different nests and 10 controlwhen placed in a novel environment (9) and spent more timeanimals from six different nests. At an age of 14 weeks the animals wereon the open arms of an elevated plus (13). individually housed in perspex cages (40×25×15 cm) with ad lib. access to
In many tests, lower anxiety is associated with a higher food and water. They were allowed 1 week of adaptation to the new housingcondition. Subsequently, they were subjected to open field tests, shock prodbehavioural activity. Differences in activity, however, maydefensive burying test, elevated plus test and a conditioned fear test. The testsnot only be related to anxiety but may also reflect differenceswere separated by at least a week and were performed during the light phase.in the general way of coping with environmental challenges.
Therefore, in addition to a lower stress-induced anxiety, the Open field testhigher activity of handled animals might be due to a more The open field is widely used to measure general explorative and locomotor
activity in response to novelty. Sometimes it is also used as a very mildactive coping strategy. Yet, the question of whether earlyanxiety test (17). The animals were subjected to a 5-min open field test threeneonatal influences alter the way of coping with stressors hastimes on consecutive days. We chose to do the test three times because otherreceived little attention (14). This question is particularly studies have suggested that differences in general explorative activity may
interesting since differences in behavioural coping with stres- only show up after repeated testing (9). The open field consisted of a roundarena divided in two imaginary concentric circles with diameters of 60 cmsors are associated with specific differences in neuroendocrineand 120 cm, respectively: a central zone and an outer zone. Locomotion ofprofile. A more active coping style is associated with anthe animals was recorded and analysed with a computerized imaging analysisincreased sympathetic tone and reactivity as reflected insystem (Ethovision, Noldus Information Technology, Wageningen, the
higher basal levels of noradrenaline and a higher noradrena- Netherlands). The following parameters were determined: total traveledline response to stressors (15, 16). In addition, a more active distance, number of visits to the central zone and time spend in the central
zone. The open field arena was thoroughly cleaned before each test.mode of response is often accompanied by a lower HPA axisresponse, although this association between behavioural
Shock-prod defensive burying testcoping and corticosterone release is less clear (4, 16). The shock-prod defensive burying is often used to establish whether animalsIn the present study we investigated the effects of early have an active or passive coping strategy in responding to stressors (16, 18).
The animals were brought to a separate test room and were exposed in theirpostnatal handling on adult stress reactivity in rats. One aimhome cage to an electrified shock-prod (7 cm long, 1 cm diameter). The prodwas to see whether postnatal handling alters adult sympath-was inserted through a small hole in the cage wall, a few cm above theetic reactivity, in addition to the well-established attenuationbedding. Whenever the animals touched the prod a constant current of 2 mA
of HPA axis reactivity. A further aim was to establish whether was delivered. Generally, animals quickly approach and explore the shock-postnatal handling alters the general coping style of the prod and receive a brief shock. After this, many animals start burying the
prod with the bedding material in their cage. The behaviour of the animalsanimals. In two experiments, a broad neuroendocrine andwas manually scored by means of a keyboard for 10-min following firstbehavioural profile was measured, which together are indicat-contact with the shock-prod. The following behaviours were recorded: (1)ive of anxiety and coping with stressors. In the first experiment burying (digging and pushing sawdust towards the prod); (2) sniffing at the
we measured the behaviour in an open field test, a shock prod (stretching and sniffing closely around the prod); (3) exploration( locomotor activity and explorative activity directed at the floor or the cageprod defensive burying test, an elevated plus maze, and a
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 925–933

Postnatal handling and adult stress reactivity 927
walls); (4) rearing (raising on hind limbs in a vertical position); (5) grooming; immunoassay. The sensitivity was 1 ng/ml plasma and values were expressedin terms of rat prolactin reference RP-3. The intra- and interassay coefficientsand (6) immobility. In addition, the latency time to the first contact (shock)
and the latency time to burying behaviour was recorded. One day after the of variation were 5% and 10%, respectively.first exposure, the uncharged prod was reintroduced in the cage to measurethe conditioned response. Novelty stress and conditioned fear response
In the first test the animals were subjected to a mild novelty stress. One byone, the rats were carried in their home cage to a separate room and placedElevated plus mazein a large unfamiliar cage (40×40×40) for 5 min and then returned to theirThe elevated plus maze is a widely used non-conditioned anxiety tests (19).own cage and their own room. The second test, a conditioned fear test, wasThe elevated plus consisted of two open arms and two closed arms 55 cmsimilar to that described in experiment 1. Animals were placed in a shockabove the floor. Each arm was 45 cm long and 10 cm wide. The open armsbox (40×40×40) and immediately received a foot shock of 0.8 mA for 3 s.had rims of about 1 cm. The closed arms had 50 cm high walls with a closedThe animals were left in the cage for 1 min and were then returned to theirtop. At the start of the test the animals were placed on the centre of the plushome cage and their own room. The next day the animals were reintroducedand the experimenter left the room. The test lasted 5 min and the behaviourin the box, this time for 5 min without receiving a shock. The conditionedof the animals was recorded on video for later analysis. The followingneuroendocrine response was measured. In both the novelty and conditionedparameters were determined: (1) time spent in the closed arms and on thefear test, blood samples were taken at t=0 (baseline before the test), t=5 (atopen arms, and time in the centre between the arms; and (2) number of visitsthe end of the test), and at t=15, 30 and 60 (respectively 10, 25 and 55 minto the closed and open arms. The plus maze was thoroughly cleaned beforeafter the test).each test.
Conditioned fear responseResultsIn addition to the non-conditioned elevated plus anxiety test, we measured a
conditioned fear response in a fourth behavioural test. On day 1 of the testthe animals were introduced to a large cage (40×40×40 cm) and immediately Experiment 1: Behavioural stress reactivity and anxietyreceived an inescapable foot shock of 0.8 mA for 3 s delivered through thegrid floor. The animals were left in the shock box for 1 min and then returned Open field testto their home cage and their own room. Twenty-four hours later the animals Table 1 shows the results of the open field tests that werewere reintroduced into the box, this time for 5 min without receiving a shock. performed on 3 consecutive days. There were no significantGenerally, animals show a clear conditioned immobility response upon forced
differences between handled and control animals in the totalre-exposure to the shock box. The behaviour of the animals was manuallyrecorded by means of keyboard. The following behaviours were recorded: (1) distance they covered during the 5-min test, the time theylocomotion ( locomotor and explorative activity); (2) rearing (raising on hind spent in the centre, or the number of times they entered thelimbs in a vertical position); (3) sniffing (explorative activity directed at the centre. The open field test thus did not indicate majorcage floor); (4) grooming; and (5) immobility. The shock-box was thoroughly
differences between handled and control animals in generalcleaned before each test.locomotor and explorative activity.
Experiment 2: Neuroendocrine stress reactivity
In a second series of experiments, animals were provided with a permanent Shock-prod defensive burying testheart catheter for repeated blood sampling to determine their neuroendocrine
In the defensive burying test, all animals approached thereactivity. Experiment 2 was performed with new animals: 11 handled animalsprod and received a brief shock. There was no significantfrom five different nests and 10 control animals from five different nests. At
an age of 16 weeks the animals were individually housed in Plexiglas cages difference between the handled and control animals in the(25×25×30 cm) with ad lib. access to food and water. After surgery and at latency to the first contact with the prod: 15.4 s (SEM 4.3)least 10 days of recovery, they were subjected to two challenge tests that in control animals and 7.4 s (SEM 1.4) in handled animals.resembled tests performed in experiment 1: a novelty challenge (a sort of
Soon after getting a shock most animals started burying thesmall-scale open field test) and a conditioned fear test. In both tests theprod with the bedding material in their cage. There was noimmobility of the animals was scored and blood samples were taken at several
time points to determine the catecholamine, prolactin and corticosterone difference in the bury latency between handled and controlresponse. The two tests were separated by 4 days and were both performed animals: 40.6 s (SEM 5.9) in control animals and 45.2 s (SEMbetween the third and ninth hour of the light phase.
6.7) in handled animals. Table 2 shows the behaviour of theanimals for a 10-min period after the first shock. There were
Surgery, blood sampling and chemical determinations no significant differences between handled and control ratsAnimals were provided with a permanent heart catheter allowing frequent
in the time spent burying the probe after they had received ablood sampling in undisturbed, freely moving rats (20). Under deep halothaneshock. After the first shock, animals sometimes very carefullyanaesthesia, a silicon catheter (0.5 mm ID, 1.0 mm OD) was inserted in the
right atrium via the jugular vein. The other end of the catheter was fixed on approached the prod again and start sniffing at it but theythe skull of the rat with stainless steel screws and dental cement. Animals seldom got another shock. Also this prod-directed explorationwere allowed to recover from surgery for at least 10 days before starting the did not differ between handled and control animals. Allexperiments. During the last days of the recovery period the animals were
together, the handled and control animals did not seem tohabituated to the procedure of blood sampling by connecting them to apolyethylene sampling tube at least three times for several hours. On experi- differ in their way of coping with the threatening prod. Onemental days, the animals were connected to the sampling tube 2 h before the day later, the uncharged prod was reintroduced in the cagestart of the test. During the experiment, blood samples of 0.6 ml were taken to measure the conditioned response (Table 2). The animalsand immediately transferred to chilled (0 °C) centrifuge tubes containing 10 ml
clearly remembered what had happened the day before andheparin (500 U/ml ) as anticoagulant and 0.1% EDTA as antioxidant. Bloodwithout touching the prod all of them started burying again.was centrifuged at 4 °C for 10 min at 2600 g and the supernatant was stored
at −80 °C for measurement of catecholamines, prolactin and corticosterone. This conditioned burying response did not differ betweenDetermination of plasma catecholamine was performed by high-performance the handled and control animals. However, the handledliquid chromatography (HPLC) in combination with electrochemical detec- animals on average spent a little more time sniffing at thetion, with minor modifications of the method by Smedes et al. (21).
prod (t-test, T1,20=−2.27, P<0.05), probably reflecting aCorticosterone concentrations were determined by HPLC and ultraviolet lightdetection (22). Plasma concentrations of prolactin were measured by radio- slightly lower anxiety.
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 925–933
928 Postnatal handling and adult stress reactivity
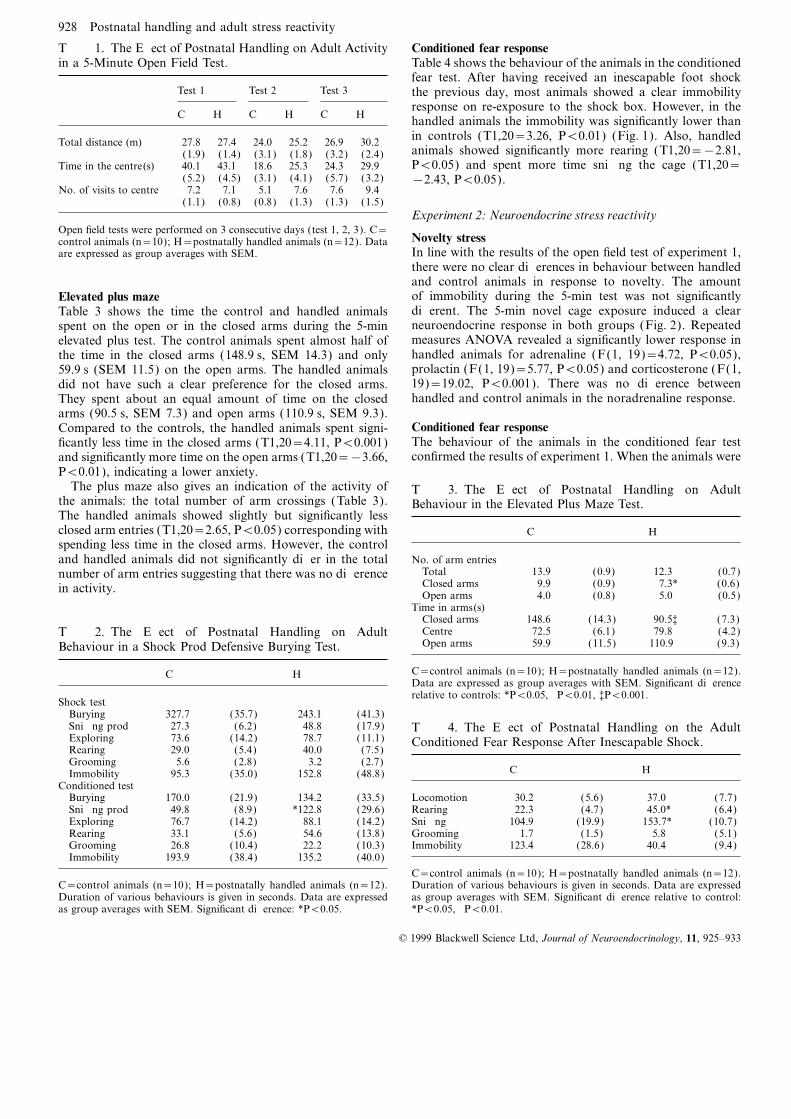
T 1. The Effect of Postnatal Handling on Adult Activity Conditioned fear responseTable 4 shows the behaviour of the animals in the conditionedin a 5-Minute Open Field Test.fear test. After having received an inescapable foot shock
Test 1 Test 2 Test 3 the previous day, most animals showed a clear immobilityresponse on re-exposure to the shock box. However, in the
C H C H C H handled animals the immobility was significantly lower thanin controls (T1,20=3.26, P<0.01) (Fig. 1). Also, handledTotal distance (m) 27.8 27.4 24.0 25.2 26.9 30.2animals showed significantly more rearing (T1,20=−2.81,(1.9) (1.4) (3.1) (1.8) (3.2) (2.4)P<0.05) and spent more time sniffing the cage (T1,20=Time in the centre(s) 40.1 43.1 18.6 25.3 24.3 29.9
(5.2) (4.5) (3.1) (4.1) (5.7) (3.2) −2.43, P<0.05).No. of visits to centre 7.2 7.1 5.1 7.6 7.6 9.4
(1.1) (0.8) (0.8) (1.3) (1.3) (1.5)Experiment 2: Neuroendocrine stress reactivity
Open field tests were performed on 3 consecutive days (test 1, 2, 3). C=Novelty stresscontrol animals (n=10); H=postnatally handled animals (n=12). DataIn line with the results of the open field test of experiment 1,are expressed as group averages with SEM.there were no clear differences in behaviour between handledand control animals in response to novelty. The amountof immobility during the 5-min test was not significantlyElevated plus mazedifferent. The 5-min novel cage exposure induced a clearTable 3 shows the time the control and handled animalsneuroendocrine response in both groups (Fig. 2). Repeatedspent on the open or in the closed arms during the 5-minmeasures ANOVA revealed a significantly lower response inelevated plus test. The control animals spent almost half ofhandled animals for adrenaline (F(1, 19)=4.72, P<0.05),the time in the closed arms (148.9 s, SEM 14.3) and onlyprolactin (F(1, 19)=5.77, P<0.05) and corticosterone (F(1,59.9 s (SEM 11.5) on the open arms. The handled animals19)=19.02, P<0.001). There was no difference betweendid not have such a clear preference for the closed arms.handled and control animals in the noradrenaline response.They spent about an equal amount of time on the closed
arms (90.5 s, SEM 7.3) and open arms (110.9 s, SEM 9.3).Conditioned fear responseCompared to the controls, the handled animals spent signi-The behaviour of the animals in the conditioned fear testficantly less time in the closed arms (T1,20=4.11, P<0.001)confirmed the results of experiment 1. When the animals wereand significantly more time on the open arms (T1,20=−3.66,
P<0.01), indicating a lower anxiety.The plus maze also gives an indication of the activity of T 3. The Effect of Postnatal Handling on Adult
the animals: the total number of arm crossings (Table 3). Behaviour in the Elevated Plus Maze Test.The handled animals showed slightly but significantly lessclosed arm entries (T1,20=2.65, P<0.05) corresponding with C Hspending less time in the closed arms. However, the control
No. of arm entriesand handled animals did not significantly differ in the totalTotal 13.9 (0.9) 12.3 (0.7)number of arm entries suggesting that there was no differenceClosed arms 9.9 (0.9) 7.3* (0.6)in activity.Open arms 4.0 (0.8) 5.0 (0.5)
Time in arms(s)Closed arms 148.6 (14.3) 90.5‡ (7.3)Centre 72.5 (6.1) 79.8 (4.2)T 2. The Effect of Postnatal Handling on AdultOpen arms 59.9 (11.5) 110.9† (9.3)Behaviour in a Shock Prod Defensive Burying Test.
C=control animals (n=10); H=postnatally handled animals (n=12).C HData are expressed as group averages with SEM. Significant differencerelative to controls: *P<0.05, †P<0.01, ‡P<0.001.
Shock testBurying 327.7 (35.7) 243.1 (41.3)Sniffing prod 27.3 (6.2) 48.8 (17.9) T 4. The Effect of Postnatal Handling on the AdultExploring 73.6 (14.2) 78.7 (11.1) Conditioned Fear Response After Inescapable Shock.Rearing 29.0 (5.4) 40.0 (7.5)Grooming 5.6 (2.8) 3.2 (2.7)
C HImmobility 95.3 (35.0) 152.8 (48.8)Conditioned test
Burying 170.0 (21.9) 134.2 (33.5) Locomotion 30.2 (5.6) 37.0 (7.7)Rearing 22.3 (4.7) 45.0* (6.4)Sniffing prod 49.8 (8.9) *122.8 (29.6)
Exploring 76.7 (14.2) 88.1 (14.2) Sniffing 104.9 (19.9) 153.7* (10.7)Grooming 1.7 (1.5) 5.8 (5.1)Rearing 33.1 (5.6) 54.6 (13.8)
Grooming 26.8 (10.4) 22.2 (10.3) Immobility 123.4 (28.6) 40.4† (9.4)Immobility 193.9 (38.4) 135.2 (40.0)
C=control animals (n=10); H=postnatally handled animals (n=12).Duration of various behaviours is given in seconds. Data are expressedC=control animals (n=10); H=postnatally handled animals (n=12).
Duration of various behaviours is given in seconds. Data are expressed as group averages with SEM. Significant difference relative to control:*P<0.05, †P<0.01.as group averages with SEM. Significant difference: *P<0.05.
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 925–933
Postnatal handling and adult stress reactivity 929
Behavioural reactivity
Several observations justify the conclusion that handledanimals do not have a more active coping strategy. First, inthe open field handled and control animals did not differ intheir general explorative and locomotor activity. Second, inthe defensive burying test there were no differences betweenhandled and control rats in the time they spent burying theshock-prod or in the amount of immobility. Third, in theelevated plus maze test the handled and control animals didnot differ in the total number of arm crossings. Especially,the shock-prod defensive burying test is very informative.This test allows the animals to deal with the potential threatof the nearby shock prod by adopting either a passive strategy(immobility) or an active strategy (burying). Since the threatis to a certain degree controllable by either one of thesestrategies, the test is only mildly stressful. The response ofclassical stress hormones like corticosterone is moderate andmuch lower than in the case, for instance, an inescapableshock (23). Yet, at the same time the test is extremely suitableto differentiate between active or passive copers (16, 18).Experiment 1 Experiment 2
Imm
ob
ility
(s)
250
200
150
100
50
0
Control
Handled
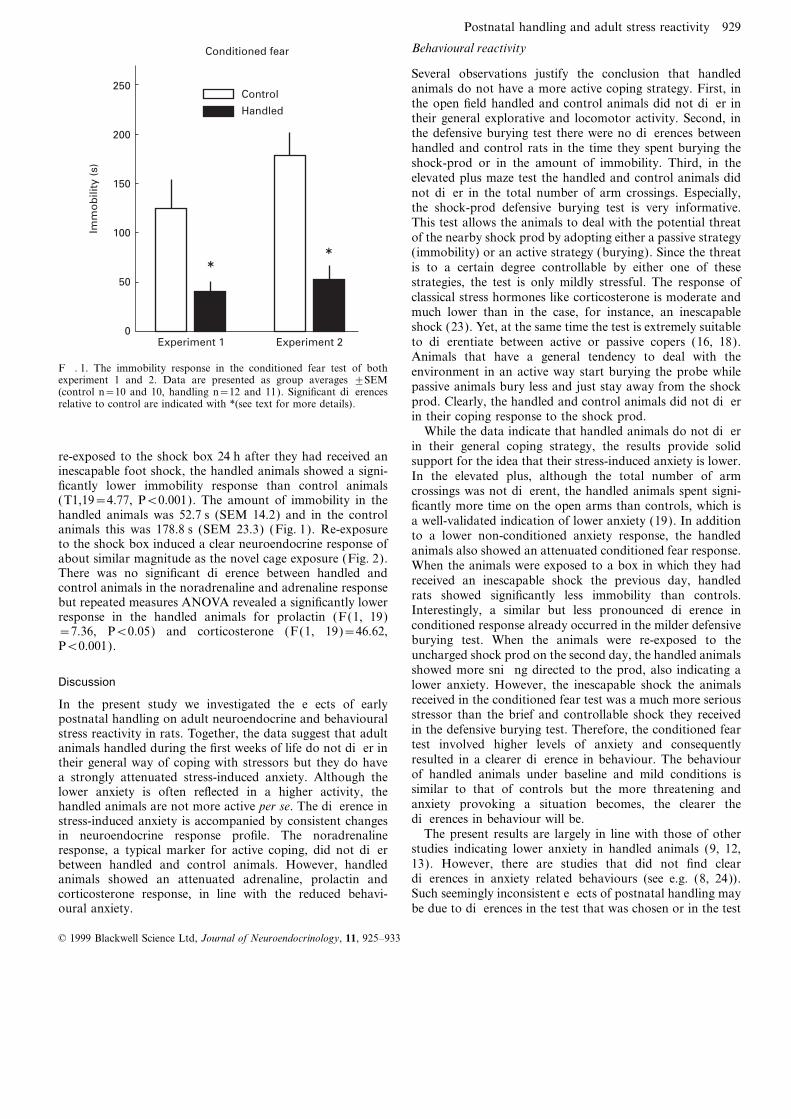
Conditioned fear
Animals that have a general tendency to deal with theF. 1. The immobility response in the conditioned fear test of both environment in an active way start burying the probe whileexperiment 1 and 2. Data are presented as group averages ±SEM passive animals bury less and just stay away from the shock(control n=10 and 10, handling n=12 and 11). Significant differences
prod. Clearly, the handled and control animals did not differrelative to control are indicated with *(see text for more details).in their coping response to the shock prod.
While the data indicate that handled animals do not differin their general coping strategy, the results provide solid
re-exposed to the shock box 24 h after they had received an support for the idea that their stress-induced anxiety is lower.inescapable foot shock, the handled animals showed a signi- In the elevated plus, although the total number of armficantly lower immobility response than control animals crossings was not different, the handled animals spent signi-(T1,19=4.77, P<0.001). The amount of immobility in the ficantly more time on the open arms than controls, which ishandled animals was 52.7 s (SEM 14.2) and in the control a well-validated indication of lower anxiety (19). In additionanimals this was 178.8 s (SEM 23.3) (Fig. 1). Re-exposure to a lower non-conditioned anxiety response, the handledto the shock box induced a clear neuroendocrine response of animals also showed an attenuated conditioned fear response.about similar magnitude as the novel cage exposure (Fig. 2). When the animals were exposed to a box in which they hadThere was no significant difference between handled and
received an inescapable shock the previous day, handledcontrol animals in the noradrenaline and adrenaline responserats showed significantly less immobility than controls.but repeated measures ANOVA revealed a significantly lowerInterestingly, a similar but less pronounced difference inresponse in the handled animals for prolactin (F(1, 19)conditioned response already occurred in the milder defensive=7.36, P<0.05) and corticosterone (F(1, 19)=46.62,burying test. When the animals were re-exposed to theP<0.001).uncharged shock prod on the second day, the handled animalsshowed more sniffing directed to the prod, also indicating a
Discussion lower anxiety. However, the inescapable shock the animalsreceived in the conditioned fear test was a much more seriousIn the present study we investigated the effects of earlystressor than the brief and controllable shock they receivedpostnatal handling on adult neuroendocrine and behaviouralin the defensive burying test. Therefore, the conditioned fearstress reactivity in rats. Together, the data suggest that adulttest involved higher levels of anxiety and consequentlyanimals handled during the first weeks of life do not differ inresulted in a clearer difference in behaviour. The behaviourtheir general way of coping with stressors but they do haveof handled animals under baseline and mild conditions isa strongly attenuated stress-induced anxiety. Although thesimilar to that of controls but the more threatening andlower anxiety is often reflected in a higher activity, theanxiety provoking a situation becomes, the clearer thehandled animals are not more active per se. The difference indifferences in behaviour will be.stress-induced anxiety is accompanied by consistent changes
The present results are largely in line with those of otherin neuroendocrine response profile. The noradrenalinestudies indicating lower anxiety in handled animals (9, 12,response, a typical marker for active coping, did not differ13). However, there are studies that did not find clearbetween handled and control animals. However, handleddifferences in anxiety related behaviours (see e.g. (8, 24)).animals showed an attenuated adrenaline, prolactin andSuch seemingly inconsistent effects of postnatal handling maycorticosterone response, in line with the reduced behavi-
oural anxiety. be due to differences in the test that was chosen or in the test
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 925–933
930 Postnatal handling and adult stress reactivity
Time (min)
0 20 50
Co
rtic
ost
ero
ne
(µg
/100
ml)
0
10
20
30
40
10 30 6040
p<0.001
Time (min)
0 20 500
10
20
30
40
10 30 6040
p<0.001
Pro
lact
in (
ng
/ml)
0
4
8
16
20 p<0.05
0
4
8
16
20 p<0.05
Ad
ren
alin
e (p
g/m
l)
0
40
80
120
200 p<0.05
0
40
80
120
200 n.s.
No
rad
ren
alin
e (p
g/m
l)
0
400
800
1200
1600n.s.
0
400
800
1200
1600n.s.
160 160
ControlHandled
ControlHandled
12 12
Novelty Conditioned fear
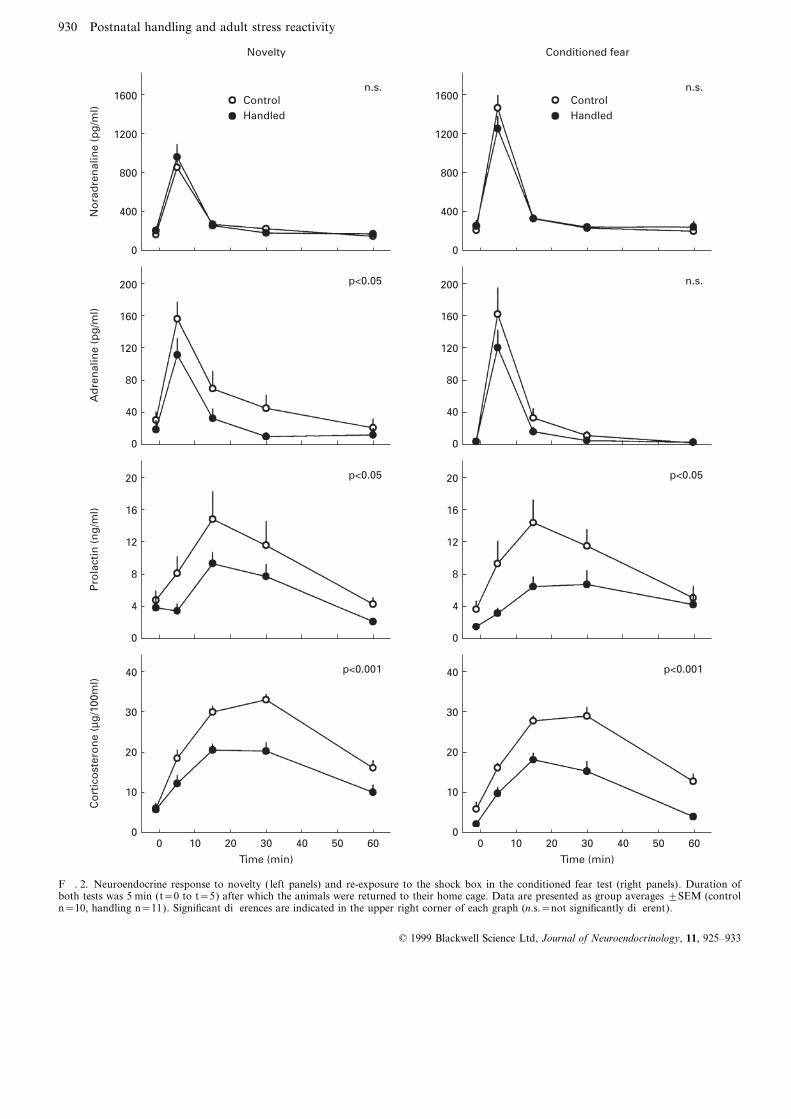
F. 2. Neuroendocrine response to novelty ( left panels) and re-exposure to the shock box in the conditioned fear test (right panels). Duration ofboth tests was 5 min (t=0 to t=5) after which the animals were returned to their home cage. Data are presented as group averages ±SEM (controln=10, handling n=11). Significant differences are indicated in the upper right corner of each graph (n.s.=not significantly different).
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 925–933

Postnatal handling and adult stress reactivity 931
circumstances. As our results suggest, the more anxiety- Neurobiological mechanisminducing the circumstances are the more likely one is to find
The crucial factor or factors in the handling proceduredifferences in behaviour. Moreover, several studies haveinducing the developmental changes, as well as the neuro-suggested a subtle interplay between genetic and ontogeneticbiological mechanism underlying the alterations in adultfactors (5, 14). The developmental effects of neonatal stimula-endocrine and behavioural reactivity are topics of intensivetion may strongly depend on the genetic background uponstudy. Handling of the pups, or associated stimuli such aswhich they are superimposed. Obviously, the anxiety-reducingmaternal licking and grooming, triggers the release of thyroideffects of postnatal handling will be less in animals thathormones and this in turn increases serotonin release inalready have a low innate emotionality (5).several brain areas (6, 25, 26). This in itself does not seemto persist until adulthood but it may affect changes in thedevelopment of various other systems. The present study, inaddition to others, shows that postnatal handling induces
Neuroendocrine reactivity changes in adult anxiety and neuroendocrine reactivity.Several studies have implicated a lower CRH response asThe results of the neuroendocrine measurements are in line
a possible central feature underlying the various physiologicalwith the behavioural data and consistent with the interpreta-and behavioural differences in adult handled rats (6, 10).tion that handled animals are less anxious but do not have aCRH plays an important role in the integrated stress responsegeneral tendency for more active coping than control animals.and has a wide variety of neuroendocrine and behaviouralAnimals with a more active coping strategy generally have aeffects, many of which occur directly within the centralhigher sympathetic tone and reactivity as reflected in highernervous system (for reviews see (2, 27)). CRH is an importantbasal levels of noradrenaline and a higher noradrenalinemediator of anxiety and, in addition, it interacts with theresponse to stressors (15, 16). Clearly this was not the caseautonomic nervous system via which it can influence catechol-in the handled animals. Also, a more active coping withamine release (28–30). The effects of CRH on prolactinstressors is often associated with a higher adrenaline responserelease are complex and sex-dependent. Yet, in males at leastto stressors (15, 16). Contrary to this, handled animals had
a lower adrenaline response to the mild novelty challenge, in it appears to have a stimulatory or permissive role (31, 32).fact, supporting the conclusion that handled animals are less And, lastly, CRH is well known as the pre- eminent regulatoranxious. Also in the conditioned fear test the adrenaline of the corticosterone response to stress (2, 27). A lower CRHresponse in handled animals on average was lower but this release in handled animals in response to stressors may thusdifference did not reach statistical significance. This may have explain many of the behavioural and neuroendocrine differ-been partly due to the somewhat higher variability in the ences with control animals. The attenuated corticosteroneresponse and the faster return to baseline levels after the response, in addition, appears to be partly the consequenceanimals had been placed back in their own cage. Also, the of an increased negative feedback due to higher concentrationeffect of lower anxiety in handled animals may in this case of corticosteroid receptors in the hippocampus (6, 33).have been partly opposed by their higher behavioural activity. Although many studies so far have focused on changes in
In both the novelty and the conditioned fear test the the regulation of CRH and the HPA axis, not all of thehandled animals had a much lower corticosterone response. neuroendocrine and behavioural changes in handled animalsThis would be consistent with a more active coping style are necessarily related to a difference in a single neuro-since it is often suggested that a more active mode of coping transmitter system or receptor type. Other systems that haveis associated with a lower activation of the HPA axis. On the been specifically implicated in the attenuated anxiety inother hand, the lower corticosterone and prolactin responses postnatally handled rats are the GABA-ergic system (34) andalso support the conclusion that handled animals are less the benzodiazepine system (35). Both of these systems haveanxious. Thus, all together, the neuroendocrine data, too, are an anxiety reducing effect and both GABA receptor bindingconsistent with the interpretation that handled animals have and benzodiazepine receptor numbers are increased ina lower stress-induced anxiety but are not more active copers. handled animals.
Interestingly, differences in neuroendocrine reactivityalready appeared in response to a mild novelty challengewhen there were no clear behavioural differences. Whilebehavioural differences became apparent only in more Conclusionsthreatening situations, the neuroendocrine response of the
Brief daily handling strongly influences the development ofhandled animals was already lower under conditions of veryrat pups resulting in an attenuated adult anxiety and neuro-mild stress and little anxiety, such as a novel cage. Theendocrine reactivity. Other kinds of stimuli or manipulationsneuroendocrine measures thus more sensitively reflected theduring the early life of neonates of course may have otherdifferences between the handled and control animals.effects. In fact, there is some evidence suggesting that moreMoreover, the present study shows that in addition to theserious intrusions may have opposite effects compared towell-established effect of postnatal handling on adult HPAhandling and ultimately may result in an increased stressaxis reactivity, handled animals also have a lower adrenalineresponse in the adult offspring (5). This seems to be case, forand prolactin reactivity. Neonatal handling thus appears toexample, when pups are subjected to maternal deprivation,have a broad effect on a wide variety of neuroendocrine
systems. i.e. taking them away from the nest for prolonged periods of
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 925–933

932 Postnatal handling and adult stress reactivity
9 Levine S, Haltmeyer GC, Karas GG, Denenberg VH. Physiological and5 or 6 h every day (36, 37). An increased HPA axis reactivitybehavioural effects of infantile stimulation. Physiol Behav 1967; 2: 55–59.was also found in adult rats that had been made ill as pups
10 Plotsky PM, Meaney MJ. Early, postnatal experience alters hypothalamicby endotoxin administration (38). corticotropin-releasing factor (CRF) mRNA, median eminence CRF
Clearly, neonatal development is a dynamic process. content and stress-induced release in adult rats. Mol Brain Res 1993;18: 195–200.Environmental influences at this early stage of life can have
11 Meaney MJ, Aitken DH. The effects of early postnatal handling on thea large impact and, for better or worse, result in changes thatdevelopment of hippocampal glucocorticoid receptors: temporal para-appear to last a lifetime. Effects of infantile stimulation even meters. Dev Brain Res 1985; 22: 301–304.
have a potential impact on the adult vulnerability to stress- 12 Nunez JF, Ferre P, Escorihuela RM, Tobena Fernandez-Teruel A. Effectof postnatal handling of rats on emotional, HPA-axis and prolactinrelated disorders. The stress response is a functional adapta-reactivity to novelty and conflict. Physiol Behav 1996; 60: 1355–1359.tion that enables animals and humans to cope with acute
13 Vallee M, Mayo W, Dellu F, Le Moal M, Simon H, Maccari S. Prenatalthreats, however, frequent or chronic activation of the stressstress induces high anxiety and postnatal handling induces low anxiety
systems also has its costs. Many of the hormones involved in in adult offspring: correlation with stress-induced corticosterone secre-acute stress-responses can promote the development of vari- tion. J Neuroscience 1997; 17: 2626–2636.
14 Steimer T, Escorihuela RM, Fernandez-Teruel A, Driscoll P. Long-termous maladaptations ranging from autoimmune diseases tobehavioural and neuroendocrine changes in Roman high-(RHA/Verh)mental disorders (39). In the case of neonatally handledand low-(RLA/Verh) avoidance rats following neonatal handling. Intanimals, there is some evidence that the attenuated corticos-J Devl Neuroscience 1998; 16: 165–174.
terone response may protect them from age-related hippo- 15 Fokkema DS, Smit K, Van der Gugten J, Koolhaas JM. A coher-campal cell-loss and cognitive impairment (40). Yet, since ent pattern among social behavior, blood pressure, corticosterone and
catecholamine measures in individual male rats. Physiol Behav 1988;postnatal handling affects various other neuroendocrine sys-42: 485–489.tems in addition to the HPA axis, it is difficult to generalize
16 Sgoifo A, De Boer SF, Haller J, Koolhaas JM. Individual differences inabout the possible consequences for health and disease. The plasma catecholamine and corticosterone responses of wild-type rats:alterations induced by infantile stimulation may decrease the relationship with aggression. Physiol Behav 1996; 60: 1403–1407.
17 Meerlo P, Overkamp GJF, Benning MA, Koolhaas JM, Van denvulnerability to one disorder but increase the sensitivity toHoofdakker RH. Long-term changes in open field behaviour followinganother. Further study is needed to elucidate the full impacta single social defeat in rats can be reversed by sleep deprivation. Physiolof early neonatal influences on adult stress reactivity andBehav 1996; 60: 115–119.
vulnerability to stress-related disorders. 18 De Boer SF, Slangen JL, Van der Gugten J. Plasma catecholamine andcorticosterone levels during active and passive shock-prod avoidancebehaviour in rats: effects of chlordiazepoxide. Physiol Behav 1990; 47:1089–1098.Acknowledgements
19 Pellow S, Chopin P, File SE, Briley M. Validation of open: closed armentries in an elevated plus maze as a measure of anxiety in the rat.The authors thank Stephan Bouman, Jan Bruggink, and Debbie Otjens forJ Neurosci Methods 1985; 14: 149–167.technical assistance and catecholamine analysis (Department of Animals
20 Steffens AB. A method for frequent sampling of blood and continuousPhysiology, University of Groningen), and Bela Toth for prolactin analysisinfusions of fluids in the rat without disturbing the animals. Physiol(Department of Human Morphology and Developmental Biology,Behav 1969; 4: 833–836.Semmelweis University of Medicine, Budapest). This study was supported by
21 Smedes F, Kraak JC, Poppe H. Simple and fast solvent extraction systemthe Netherlands Organization for Scientific Research (NWO-SLW, projectfor selective and quantitative isolation of adrenaline, noradrenaline and805–30–181).dopamine from plasma and urine. J Chromatogr 1982; 231: 25–39.
22 Dawson R, Kontur P, Monjan A. High-performance liquid chromato-Accepted 22 June 1999 graphic (HPLC) separation and quantitation of endogenous gluco-
corticoids after solid-phase extraction from plasma. Horm Res 1984;20: 89–94.
23 Koolhaas JM, De Boer SF, De Ruiter AJH, Meerlo P, Sgoifo A. SocialReferencesstress in rats and mice. Acta Physiol Scand 1997; 161: 69–72.
24 Hilakivi-Clarke LA, Turka J, Lister RG, Linnoila M. Effects of early1 Axelrod J, Reisine TD. Stress hormones: their interaction and regulation.postnatal handling on brain (-adrenoreceptors and behavior in testsScience 1984; 224: 452–459.related to stress. Brain Res 1991; 542: 286–292.2 Johnson EO, Kamilaris TC, Chrousos GP, Gold PW. Mechanisms of
25 Liu D, Dioro, Tannenbaum Caldji C, Francis D, Freedman A, Sharmastress: a dynamic overview of hormonal and behavioral homeostasis.S, Pearson D, Plotsky P, Meaney MJ.Maternal care, hippocampalNeurosci Biobehav Rev 1992; 16: 115–130.glucocorticoid receptors, and hypothalamic-pituitary-adrenal responses3 Benus RF, Bohus B, Koolhaas JM, Van Oortmerssen GA. Heritableto stress. Science 1997; 277: 1659–1662.variation for aggression as a reflection of individual coping styles.
26 Smythe JW, Rowe WB, Meaney MJ. Neonatal handling alters serotoninExperientia 1991; 47: 1008–1019.(5-HT) turnover and 5–HT2 receptor binding in selected brain regions:4 Castanon H, Mormede P. Psychobiogenetics: adapted tools for the studyrelationship to the handling effect on glucocorticoid receptor expression.of the coupling between behavioral and neuroendocrine traits of emo-Dev Brain Res 1994; 80: 183–189.tional reactivity. Psychoneuroendocrinology 1994; 19: 257–282.
27 Dunn AJ, Berridge CW. Physiological and behavioral responses to5 Anisman H, Zaharia MD, Meaney MJ, Merali Z. Do early life eventscorticotropin-releasing factor administration: is CRF a mediator ofpermanently alter behavioral and hormonal responses to stressors? Intanxiety or stress responses? Brain Res Rev 1990; 15: 71–100.J Devl Neuroscience 1998; 16: 149–164.
28 Brown MR, Fisher LA, Spiess J, Rivier C, Rivier J, Vale W.6 Meaney MJ, Mitchell JB, Aitken DH, Bhatnagar S, Bodnoff SR, InyCorticotropin-releasing factor: actions on the sympathetic nervous systemLJ, Sarrieau A. The effect of neonatal handling on the development ofand metabolism. Endocrinology 1982; 111: 928–931.adrenocortical response to stress: implications for neuropathology and
29 Swiergiel AH, Takahashi LK, Rubin WH, Kalin NH. Antagonism ofcognitive deficits in later life. Psychoneuroendocrinology 1991; 16: 85–103.corticotropin-releasing factor receptors in the locus coeruleus attenuates7 Ader R. The effect of early experience on the adrenocortical response toshock-induced freezing in rats. Brain Res 1992; 587: 263–268.different magnitudes of stimulation. Physiol Behav 1970; 5: 837–840.
30 Swiergiel AH, Takahashi LK, Kalin NH. Attenuation of stress-induced8 Hess JL, Deneberg VH, Zarrow MX, Pfeifer WD. Modification of thebehavior by antagonism of corticotropin-releasing factor receptors in thecorticosterone response curve as a function of handling in infancy.
Physiol Behav 1969; 4: 109–112. central amygdala in the rat. Brain Res 1993; 623: 229–234.
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 925–933

Postnatal handling and adult stress reactivity 933
31 Morel G, Enjalbert A, Proulx L, Pelletier G, Barden N, Grossard F, 36 Lad CO, Owens MJ, Nemeroff CB. Persistent changes in corticotropin-releasing factor neuronal systems induce by maternal deprivation.Dubois PM. Effect of corticotropin-releasing factor on the release and
synthesis of prolactin. Neuroendocrinology 1989; 49: 669–675. Endocrinology 1996; 137: 1212–1218.37 Biagini G, Pich EM, Carani C, Marrama P, Agnati F. Postnatal maternal32 Akema T, Chiba A, Oshida M, Kimura F, Toyoda J. Permissive role of
corticotropin-releasing factor in the acute stress-induced prolactin release separation during the stress hyporesponsive period enhances the adreno-cortical response to novelty in adult rats by affecting feedback regulationin female rats. Neurosci Lett 1995; 198: 146–148.
33 Meaney MJ, Aitken DH, Bodnoff SR, Iny LJ, Tatarewicz JE, Sapolsky in the CA1 hippocampal field. Int J Devl Neuroscience 1998; 16: 187–197.38 Shanks N, Larocque S, Meaney MJ. Neonatal endotoxin exposure altersRM. Early postnatal handling alters glucocorticoid receptor concentra-
tions in selected brain regions. Behav Neurosci 1985; 99: 765–770. the development of the hypothalamic-pituitary-adrenal axis: early illnessand later responsiveness to stress. J Neuroscience 1995; 15: 376–384.34 Bolden SW, Hambley JW, Johnston GAR, Rogers LJ. Neonatal stress
and long-term modulation of GABA receptors in the rat brain. Neurosci 39 McEwen BS. Protective and damaging effects of stress mediators. N EnglJ Med 1998; 338: 171–179.Lett 1990; 111: 258–262.
35 Bodnoff SR, Suranyi-Cadotte B, Quirion R, Meaney MJ. Postnatal 40 Meaney MJ, Aitken DH, Berkel CV, Bhatnagar S, Sapolsky RM. Effectof neonatal handling on age-related impairments associated with thehandling reduces novelty-induced fear and increases [3H ]flunitrazepam
binding in rat brain. Eur J Pharmacology 1987; 144: 105–107. hippocampus. Science 1988; 239: 766–768.
© 1999 Blackwell Science Ltd, Journal of Neuroendocrinology, 11, 925–933