the fate of embryonal-carcinoma cells in mouse blastocysts
TRANSCRIPT

Diffcrentiation (1987) 33 : 247-253 Differentiation 0 Springer-Verlag 1987
The fate of embryonal-carcinoma cells in mouse blastocysts G. Barry Pierce*, Juan Arechaga**, Alan Jones, Andrea Lewellyn, and Robert S. Wells Department of Patholoav. Universitv of Colorado School of Medicine. 4200 East Ninth Avenue. Denver, Colorado 80262, USA
Abstract. Double-labeled embryonal-carcinoma (ECa) cells were injected into blastocysts or incorporated into blasto- cysts by aggregation, and their fate after various periods of time in culture was investigated. ECa-247 cells labeled with fluorescent microscopheres werc easily identified in whole blastocysts. These blastocysts were embedded in plas- tic, serially sectioned, and prepared for autoradiography. The 'H-thymidine label on the embryonalcarcinoma cells allowed precise localization of the cancer-derived cells. ECa-247 cells preferentially localized in the mural trophec- toderm, with a few being seen in primitive endoderm and, even more rarely, in the inner cell mass. Selected autoradio- grams were reembedded and thin sectioned for transmis- sion electron microscopy. The cancer-derived cells were found to have differentiated in accordance with their local- iza t ion.
Introduction
Chimeric mice have been produced by injecting embryonal- carcinoma cells into the blastocysts of mice and then placing the injected blastocysts into the uteri of pseudopregnant animals [3, 13, 151. Chimeric offspring were recognized first by coat color [3] and later by isoenzyme analyses which indicated apparently normal cancer-derived cells in virtually all organs [2, 10, 11. 13, 15, 16, 231. There are strain differ- ences in the behavior of embryonal-carcinoma cells injected into blastocysts. As already mentioned, some form chimer- as, some form chimeras in extra-embryonic tissue only, some form chimeras that also have embryonal carcinomas, and some do not respond to the blastocyst [2, 3, 10, 11, 13, 15, 16, 231. It is concluded that the blastocyst is able to regulate some embryonal-carcinoma cells (even aneup- loid ones [20]), so that they and their offspring respond to normal physiologic controls. In this sense, cancer-derived cells may be considered to be normal.
Assays have been developed to analyze the mechanism of the regulation of cancer cells by the blastocyst (20, 241. In a tumor assay, the process proved to be tissue specific: of the five types of tumor tested in the blastocyst, only embryonal carcinoma was regulated and failed to produce tumors in the expected numbers [20, 211. In a colony assay,
* To whom offprint requests should be sent ** Present address: Catedra de Histologia y Embriologia. Universi- dad de Granada, Granada, Spain
embryonal-carcinoma cells were responsive to the regula- tion of colony formation by the blastocyst at between 4 and 5 h after mitosis [24]. Contact with trophectoderm in the presence of blastocele fluid was required for regulation
Such tumor and colony assays do not indicate the fate of the embryonal-carcinoma cells that fail to grow. The assumption that the cancer cells that had failed to produce tumors or colonies had differentiated was logical on the basis of the known ability of embryonal carcinomas to make chimeras, but this remained unproven in the in vitro assays. The situation was further complicated by the obser- vation that the embryonalcarcinoma line employed, i.e., ECa-247, was regulated in the tumor and colony assays but did not form injection chimeras in vivo (unpublished results). The loss of tumor- and colony-forming ability may be one of the early steps in the long and complex process of chimera formation.
The present study describes experiments in which the fate of labeled ECa-247 cells injected into blastocysts was determined in vitro. Most labeled cells were incorporated into trophectoderm, a type of differentiation not usually attributed to egg-derived teratocarcinomas.
[22].
Methods
The cell line employed in these experiments was ECa 247, this being a subline of OTT6050, and egg-derived teratocar- cinoma (121. This line grows well in vitro. As already men- tioned, it does not form viable injection chimeras, yet the tumor forming ability and colony-forming ability of these cells are abrogated after sojourn in the blastocyst [20, 24).
ECa-247 cells were labeled with fluorescent micro- spheres by culturing them overnight in plastic Petri dishes at a density of 2 x 10' cells/ml in Eagle's minimal essential medium containing 10% fetal calf serum (MEM + 10) and fluorescent carboxylated microspheres (0.7 pm; CX Green Covaspheres; Covalent Technology Corporation, Ann Ar- bor, Mich) at a concentration of 5.5 x lo* beads/ml [20]. The labeled cells were washed three times in MEM + 10, and they cloned as efficently as unlabeled cells. The time of the cell cycle was not changed.
Double-labeled ECa-247 cells were produced by adding 'H-methylthymidine (specific activity, 27 Ci/mM at a con- centration of 0.25 pCi/ml to the medium for labeling with microspheres). The toxicity of the 'H-thymidine depended less on the duration of labeling than on the concentration

248
of reagcnt. Higher concentrations of 'H-thymidine drasti- cally reduced cloning efficiency, but with the amount em- ployed, the cloning efficiency was reduced by not more than 20%.
Blastocysts were flushed from the uteri of CD-1 females (Charles River Company) on the 3rd day of gestation, and eight cell eggs were flushed from the oviducts on the 2nd day of gestation. The cells were placed in a drop of MEM + 10 medium under pre-equilibrated paraffin oil in 35-mm plastic dishes [8], and were kept in an incubator at 100% humidity in an atmosphere consisting of 5% CO, in air at 37" C until used.
The method used for injecting embryonalcarcinoma cells into blastocysts was as described previously [20]. The method used for making aggregation chimeras was a modi- fied version of that described by Pedersen and Spindle [17]. This involved removing the zona pellucida by incubating the eightcell eggs in 0.5% pronase (Sigma) in 0.5% PVP-40 (Sigma) in phophate-buffered salihe (PBS) at 37" C. After removal of the pronase, pairs of eightcell embryos were placed in prewamed microdrops of MEM + 1 % fetal calf serum containing 0.1 YO phytohemagglutinin and Hepes buffer under washed mineral oil. Clusters of five to nine labeled ECa-247 cells were placed between the two embryos using an orally controlled pipette. The cells were incubated for about 10 min at 37" C, rinsed twice in PBS, and cultured for 48 h to the blastocyst stage in MEM + 10 at 100% hu- midity in an atmosphere consisting of 5% CO, in air at 37" C. In later experiments, two eightcell embryos and five to nine embryonal-carcinoma cells were aggregated in capil- lary tubes as illustrated in Fig. 1 a-g. Orally controlled pi- pettes were used to place eightcell embryos (Fig. 1 b) in small drops of MEM + 10 under oil (Fig. 1 a). The embryos and media were aspirated into siliconized glass capillary tubes, and the tips of the tubes were plugged by aspirating a drop of washed mineral oil into the orifice (Fig. 1 c). The tubes were incubated in the vertical position (Fig. 1 d), and the embryos were aggregated by gravity. After aggregation and the development of blastocysts, the oil droplet was aspirated into the body of the pipette (Fig. le), and the tip was broken (Fig. 1 f) in order to make an orifice large enough to expel the newly formed blastocyst without injury (Fig. lg). This procedure proved to be an efficient and quick way of producing a high percentage of chimeric blas- tocysts.
Blastocysts containing labeled cells (as determined by fluorescence microscopy) were immediately fixed in 2% glu- taraldehyde in 0.2 M PBS, postfixed in 1 YO osmium tetrox- ide, dehydrated, and embedded in epoxy resin. The prepara- tions were serially sectioned. The 5-pm thick sections were mounted on glass slides and dipped in NTB-2 emulsion (Kodak) for autoradiography as described previously [18]. In these autoradiograms, the labeled cells were identified and tabulated according to their localization. Selected sec- tions were then reembedded in epoxy resin by inverting a gelatin capsule filled with Epon over the 5-pm-thick sec- tions. After reembedding, these sections were recut for ul- tramicroscopy .
Results In the first experiment, blastocysts were injected with fluo- rescent-bead-labeled ECa-247 cells, cultured for the times indicated in Table 1, and examined as intact specimens
I I
Fig. 1. This method for mass-producing chimeric blastocysts by aggregation is eflicient and reproducible. For a description of the procedure. see Methods (1)
Table 1. Localization in blastocysts of ECa-247 cells labeled with fluorescent microspheres after various periods of culture
~
Culture Number of Number of Number of time (h) blastocysts blastocysts with labeled cells
labeled cells per blastocyst
24 53 39 (73%) 1.7 (88/53) 48 38 33 (86%) 3.4 (130/38) 72 33 28 (86%) 5.4 (177/33)
under a fluorescence microscope. Irrespective of the amount of washing, labeled cells had a tendency to release micro- spheres which were engulfed by other cells. Against this background of scattered single microspheres, labeled em- bryonalcarcinoma cells could be identified by focal collec- tions of microspheres. As noted in Table 1, labeled embryo- nal-carcinoma cells were found in 73%-86% of blastocysts, and each blastocyst contained an average of 1.7, 3.4, and 5.4 labeled cells after 24.48, and 72 h of incubation, respec- tively. The foci of beads were difficult to identify by 72 h because of dilution of the label at mitosis. Although the injected embryonalcarcinoma cells were placed on the in- ner cell mass (ICM), most of them were observed in associa- tion with trophectoderm after incubation. The resolution in some of the whole mounts made it difficult to determine whether the labeled cells were in or on trophectderm.
Some of the injected blastocysts that had been incubated for either 48 or 72 h in vitro were prepared for electron
249
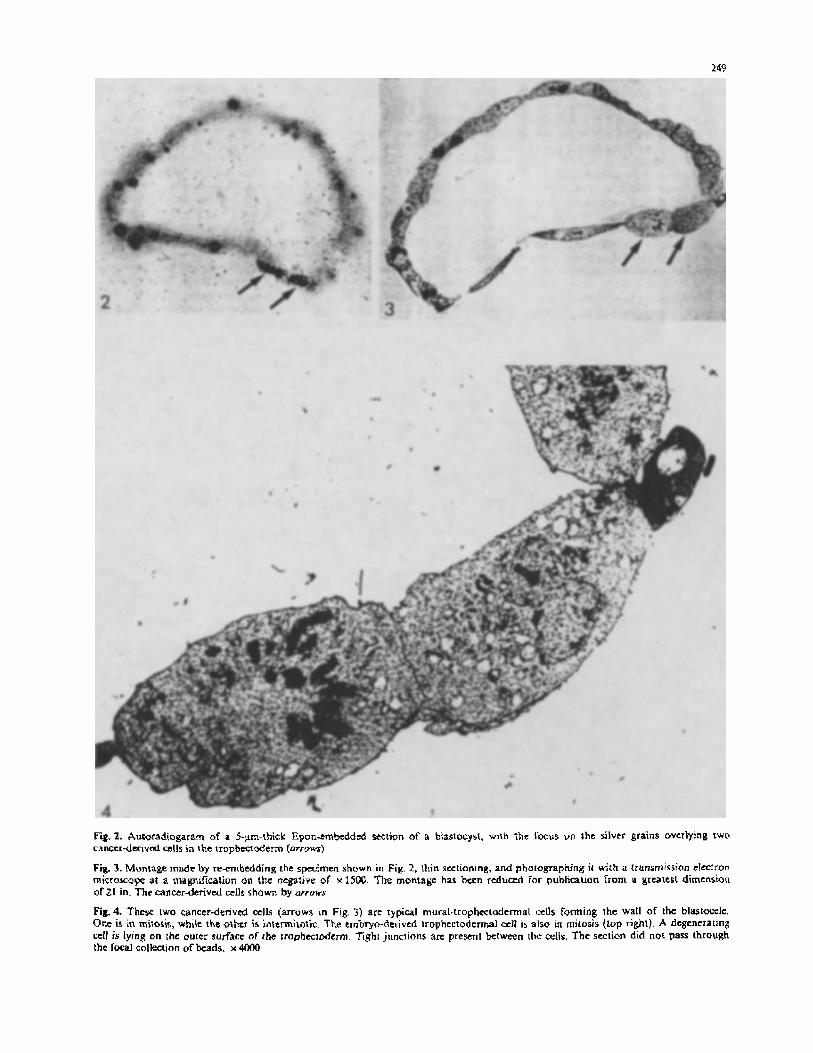
Fig.2. Autoradiogaram of a 5-p-thick Epm-trnkddcd Kclion of a biastocyst, with rht lbcus vn the silver grains overlying two canccr-derivcd cells in the tropbecrderm (urruws)
Fig. 3. Montage made by reernbedding the spcimen shown in Fig, 2, lhin sectioning. and photographing ir with a transmission cIcclron miCKOKOpe at a magnification on the ntgafive of x 15W. The montage has bcen rtduccd for publication from a greatest dimension or21 in. The cancerderived cells shown by urruws
Fig.4. These IWO cancer-derived cells (arrows in Fig. 3) arc typical rnuralrtrophcctadtrmal cells forming 1he wall of the bla$tofelc. One IS in mitoris, whik the othtr is initmilotic. The embryo-denvcd trophtctodermal cell is also in mitosis (lop righl). A degenerating cctl is lying on the outer surface of the rrophecioderm. Tight juncrions are present between the cells. The section did n ~ t pass through the f@xal collection of beads. x 4OOO

250
microscopy. Thick sections were cut until bead-labeled cells were encountered, at which point thin sections were pre- pared. Specimens after 48 h of incubation proved more use- ful than those after 72 h of incubation, because many of the latter had hatched from the zona pellucida and attached to the plastic of the dish, so that the normal embryological relationships were difficult to discern.
When viewed under an electron microscope, labeled cells in 48-h blastocysts were usually found in the trophecto- dermal layer. This confirmed the light-microscope observa- tions, but these was still the possibility of artefacts. Injected blastocysts collapse with the outflow of fluid through the wound made in the mural trophectoderm. Conceivably, the embryonalcarcinoma cells might have floated off the ICM and become trapped in the puncture wounds. It seemed reasonable to suspect this artefact, because egg-derived ter- atocarcinomas rarely spontaneously differentiate into tro- phectoderm [6]. As a consequence, fusion chimeras were made containing double-labeled embryonal-carcinoma cells to avoid the possibility of this artefact.
Of the blastocysts made by the fusion of two eight-cell eggs with five to nine double-labeled ECa-247 cells, 84 con- tained focal areas of fluorescent beads, indicating the suc- cessful incorporation of some of the labeled cancer cells. Many of the cancer cells attached to the plastic Petri dishes, however, and were lost. The 84 blastocysts containing fluo- rescent cells were serially sectioned at a thickness of 5 pm, and the labeled cells were identified by autoradiography using a light microscope. Surprisingly, only 12 labeled cells were observed in or on the ICM. Three of these cells were in an ICM, suggesting that, with further development, this blastocyst might have produced a chimeric mouse. The other 9 cells were incorporated in the endodermal layer of the blastocyst.
One-hundred-and-ten labeled cells were found on the external surface of the trophectoderm. Some appeared inti- mately attached to the trophectoderm, but others merely lay adjacent to it. There was no evidence that any of these cells were in the process of being incorporated into the trophectodenn.
Eighty-five cells were found in the trophectoderm, most of them being in the mural trophectuderm. Only 4 were in the polar trophectoderm. Fourteen cells were found on the inner surface of the trophectoderm, and most of these. were degenerating.
Five thick sections (each from a different blastmyst) were reembedded, and thin sections were prepared for
transmission electron microscopy in order to define the fine structure of the double-labeled cells. Due to the way in which the sections were reembedded, it was possible to see the silver grains of the light autoradiograms in the blocks. This facilitated identification of the labeled cells during sectioning. Montages were prepared at low magnifi- cation, and higher-magnification pictures were then taken of the labeled cells identified in the montages.
The first 5-pm-thick section selected for re-embedding contained two labeled cells, one being densely labeled, the other lightly labeled. They lay adjacent to each other in the mural trophectoderm (Fig. 2). These cells were easily recognized in the montage (Fig. 3, arrows). The densely labeled cancer-derived cell was in mitosis, but the lightly labeled cell adjacent to it was intermitotic. The lightly la- beled cell may have recently undergone mitosis, which would account for the dilution of the label. Both had the appearance of trophectodermal cells [4, 7. 8, 141, and mem- brane differentiations typical of trophectoderm were ob- served when embryo and cancer-derived cells abutted (Fig. 4 [7, 81). There were tight and gap junctions, but the plane of sectioning did not allow high-resolution microsco- py. The cytoplasm of the cancer-derived and embryo-de- rived cells was similar. Attached to the outer surface of the trophectoderm, and not labeled either by beads or by autoradiography. was a cell with a very dense granular cyto- plasm and a vesiculated nucleus (Figs. 3, 4). This was con- strued as being a degenerating cell. An embryo-derived mural-trophectodenn cell, which lay adjacent to the degen- erating cell, was also in mitosis. It was concluded that two canerderived cells had been incorporated in the trophecto- derm, one of them being in the process of division, the other being intermitotic. They both had the appearance of mural trophectoderm.
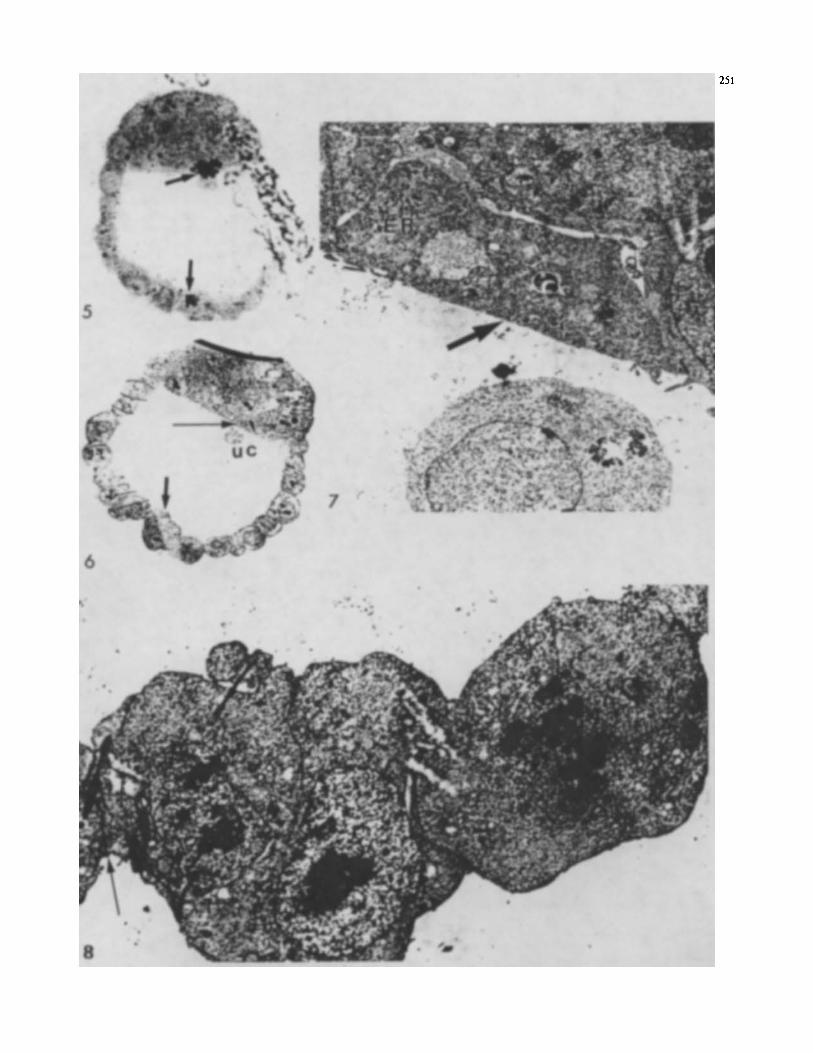
The second reembedded specimen was chosen because one labeled cell was found in the mural trophectoderm op- posite the ICM, and another was found on the Mastocele surface of the ICM (Fig. 5). From the montage, it appeared that the latter cell was in the primitive endodermal layer of the blastocyst (Fig. 6). The cytoplasm of the labeled cell in the trophectoderm was indistinguishable from the adja- cent trophectodermal cells, and even contained the glycogen that accumulates with the culturing of trophectoderm (Fig. 8). Typical membrane differentiations were present. The labeled cell of the ICM lay in the endodermal layer (Fig. 7). had a better-developed rough endoplasmic reticu- lum (RER) than is typical for ICM cells, and closely resem-
Fig. 5. Autoradiogram with the focus on the silver grains overlying one cancer-derived cell in the endodem and another in the mural trophectoderm (arrows)
fig. 6. Montage made by re-embedding the section shown in Fig. 5 and processing it as described in the legend to Fig. 3. The montage has been reduced from a greatest dimension of 16 in. An unlabeled a l l (uc) lies in the blastoccle and serves as a landmark for locating the canerderived a l l in the endoderm (long arrow) and the cancer-derived cell in the mural trophectodem (short arrow)
fig. 7. The landmark a l l in the blastoccle (see Figs. 5.6) is visible at the lower edge of this electron micrograph, and the cancer-derived cell is shown by the arrow. It is a typical endodermal cell with abundant rough endoplasmic reticulum (ER). Note that this ultrathin section docs not include the nucleus (which lay deeper in the specimen) or beads (localized in clumps and not always present in such sections) x 8000
fig. 8. The canctr-derived trophectodennal cell shown in Figs. 5 and 6 is visible on the lefr; Note the membrane differentiations (arrows) and the similarity of the cytoplasm to that of normal trophectodennal cells. The embryo-derived trophectodermal cell on the righr is in mitosis. x 8400
251

252
bled primitive endoderm [19]. Thus, from its position and morphologic appearance, it was concluded that this cell had, in fact, differentiated into endoderm.
The third specimen had two labeled cells attached to the inner surface of trophectoderm as well as a labeled cell apparently incorporated in the trophectoderm. These cells were easily identified in the montage. The cell incorpo- rated in the trophectoderm had the appearance of a typical mural trophectodemal cell. Tight and gap junctions were identified in the interconnecting membranes of this cancer- derived cell and of the embryo-derived trophectoderm. At the light-autographic level, two labeled cells were identified on the inner surface of the trophectoderm. At the electron- microscope level, one was necrotic, while the other ap- peared to be viable and was intimately attached to the inner surface of the trophectoderm. With the resolution attained, no membrane differentiations were found between the latter cell and the embryo-derived trophectoderm, and its ultimate fate, i.e., to be incorporated into the trophectoderm or to degenerate, could not be determined.
The fourth montage had two labeled cells attached to the inner surface of trophectoderm and one labeled cell incorporated in the trophectoderm. The labeled cell in the trophectoderm had a typical trophectoderm appearance, having appropriate membrane differentiations and a signifi- cant concentration of glycogen in its cytoplasm. The labeled cells on the inner surface of the trophectoderm were necrot- ic. The final montage was from a blastocyst in which a labeled cell was present on the blastocele surface of the ICM. This cell was easily identified in the montage and exhibited the cytoplasmic differentiations characteristic of early endo- derm. Of the 66 embryo-derived mural-trophectodermal nuclei encountered in the five montages, 3 were in mitosis (4.5%); of the 5 cancer-derived trophectodermal cells, 1 was in mitosis (20%).
Discussion
Egg-derived teratocarcinomas seldom, if ever, spontaneous- ly differentiate into trophectoderm [a. Consequently, when labeled ECa cells were injected into the blastocyst, and, after 24 h of incubation, seemed to have been incorporated into the mural-trophectodermal layer, an artefact was sus- pected. It seemed possible that the cancer cells might have floated off the ICM as the blastaccle collapsed, and with the outflow of fluid, have become trapped in the puncture wound made in the mural trophectoderm by the injection pipette.
To obviate this possible artefact, aggregation chimeras were made using double-labeled ECa-247 cells. The result- ing data confirmed the impressions obtained with the injec- tion chimeras. The fluorescent label allowed easy selection of blastocysts that had incorporated embryonalkarcinoma cells, and 'H-thymidine labcling made precise localization of the cells possible. Of the 97 ECa-247 cells incorporated in blastocysts, 85 were in trophectoderm. with most of the rest being in the primitive endoderm on the blastocele sur- face of the ICM. Only 3 cells were in a single ICM. The lack of incorporation of ECa-247 cells into the ICM pre- sumably accounts for the lack of success in producing chi- meras when one to five ECa-247 cells were injected into blastocysts (unpublished data).
Five 5-pm-thick sections (each representing a blastocyst) were subjected to light-microscope autoradiography and were then re-embedded and examined using an electron mi- croscope. Montages were made of each. The labeled cancer- derived cells in the trophectodermal layer resembled tro- phectoderm [4, 71. They had the tight junctions, gap junc- tion's, and desmosomes typical of trophectodermal mem- branes [7,8]. Their cytoplasm closely resembled that of nor- mal trophectoderm. It has to be acknowledged that there are no specific markers for trophectoderm, but the appear- ance of these cells was compatible with trophoblast differen- tiation. One of the cancer-derived cells was in mitosis (Fig. 3), while another had just completed mitosis. From the data shown in Table 1, it is clear that cancer-derived mural-trophectodermal cells divide once or twice before be- coming postmitotic. This is a higher rate of cell division than Copp measured in mural trophectoderm [S]. He sug- gested, in accordance with the observations of Gardner [9], that the ICM stimulates cell division in polar trophecto- derm. Mural trophectoderm has a low rate of cell division, but it grows due to a contribution of cells from the polar trophectoderm. Possibly, a memory of the rapid prolifera- tion of the neoplastic state carries over after cancer-derived cells have been incorporated in the trophectoderm. Copp has suggested a memory of the contact of polar trophecto- derm with ICM as an explanation for the higher rates of mitosis in mural trophectoderm that lies near the ICM IS]. Possibly, a round or more of mitosis is required to fix the differentiation. It is concluded, from their position and ap- pearance, that the cancer-derived cells in the trophectoderm had differentiated into trophectoderm. Papaioannou et al. [15] found progeny of their C86 embryonal-carcinoma lines in the trophectodermal fractions of two out of three chimer- as studied. Neither the PSA-1jNG-2 nor PSA-4flG12 tera- tocarcinoma lines colonized the trophectoderm in aggrega- tion chimeras [18]. Evidence has been obtained that cells derived from the C145 embryonal-carcinoma line preferen- tially populate the extra-embryonic endoderm [ 161. Thus, there appear to be strain differences in the ability of embry- onal carcinomas to colonize embryos [l 1).
Many labeled ECa-247 cells were observed on the outer surface of the trophectoderm. We believe that they could not be incorporated into the blastocyst because, in the col- ony assay, ECa-247 cells cloned equally well in the perivitte- line space (the space between the trophectoderm and the zona pellucida) as on plastic 1241. The majority of labeled cells on the inner surface of the trophectoderm were necrot- ic. The viable-appearing ones were in close opposition to trophectoderm. This observation might suggest that, in order to survive, ECa-247 cells must be integrated into the structure of the blastocyst quickly. It is of some interest that Chinese-hamster ovary cells die quickly in the blasto- cele [U]. In addition, ECa-247 cells cultured in rabbit blas- tocele fluid die quickly (unpublished finding). It seems likely the rabbit blastocyst, which has an abundance of blastocele fluid, would be an ideal model for studying, the mechanism by which ECa-247 cells become incorporated into and dif- ferentiate into trophectoderm.
Acknowledgemnts. We gratefully acknowledge the excellent techni- cal work of J. White. and the editorial assistance of V. Starbuck. This work was supported in part by a gift from R.J.R. Nabisco. and grants from the National Institutes of Health (CA-35367 and CA-36069).

253
References
1. Arechaga J , Wells R , Pierce G B (1 985) Aggregation chimaeras in capillary tubes. An efficient new technique for clonal analysis of development in mammals. J Cell Biochrm Suppl 9B:267
2. Bradley A. Evans M, Kaufman M, Robertson E (1984) Forma- tion of germ-line chimeras from embryo-derived teratocarcin- oma cell lines. Nature 309: 25S2.57
3. Brinster RL (1974) Effect of cells transferred into the mouse blastocyst on subsequent development. J Exp Med
4. Calarco PG, Brown EH (1969) An ultrastructural and cytologi- cal study of preirnplantation development of the mouse. J Exp Zoo1 171 :253-283
5. Copp AJ (1978) Interaction between inner cell mass and tro- phectoderm of the mouse blastocyst. I. A study of cell prolifera- tion. J Embryol Exp Morphol48: 109-125
6. Damjanov I , Solter D (1974) Experimental teratoma. Curr Top Pathol 59:69-130
7. Ducibella T, Anderson E (1979) The effects of calcium defi- ciency on the formation of the zonula occludens and blastocele in the mouse embryo. Dev Biol 73:4658
8. Ducibella T. Abertini DF, Anderson E, Biggin J D (1975) The preimplantation mammalian embryo: Characterization of in- tercellular junctions and their appearance during development. Dev Biol45:231-250
9. Cardner RL (1972) An investigation of inner cell mass and trophoblast tissues following their isolation from the mouse blastocyst. J Embryol Exp Morphol 28:279-312
10. Hanaoka K, Kato Y, Noguchi T (1986) Comparative study on the ability of various teratocarcinomas to form chimeric mouse embryos. Dev Growth Differ 28:223-231
1 I . Illmensee K (1978) Reversion of malignancy and normalized differentiation of teratocarcinoma cells in chimeric mice. In: Russell LB (ed) Genetic mosaics and chimeras in mammals. Plenum, New York, pp 3-25
12. Lehman JM, Speers WC, Swartzendruber DE, Pierce GB (1 974) Neoplastic differentiation : Characteristics of cell lines
140: 1049-1056
derived from a nxrine teratocarcinoma. J Cell Physiol
13. Mintr B, Illmensee K (1975) Normal genetically mosaic mice produced from malignant teratocarcinoma cells. Proc Natl Acad Sci USA 72:258>3589
14. Nadijcka M. Hillman N (1974) Ultrastructural studies of the mouse blastocyst substages. J Embryol Exp Morphol 32: 675-695
15. Papaioannou VE, hlcBurney MW, Gardner RL, Evans RL (1975) Fate of teratoarcinoma cells injected into early mouse embryos. Nature 258 :70-73
16. Papaioannou VE, Evans EP, Gardner RL, Graham C F (1979) Growth and differentiation of an embryonal carcinoma cell line (C145). J Embryol Exp Morphol 54:277-295
17. Pedenen RA, Spindle A1 (1980) Role of the blastocele microen- vironment in early mouse differentiation. Nature 284: 550-552
18. Pierce GB, Wallace C (1971) Differentiation of malignant to benign cells. Cancer Res 31 : 127-1 34
19. Pierce GB. Midgley AR, Feldman JD. Sri Ram, J (1962) Parie- tal yolk sac carcinoma. Clue to the histogencsis of Reichert's membrane of the mouse embryo. Am J Pathol41: 549-566
20. Pierce GB, Lewis SH. Miller G, Moritz E, Miller P (1979) Tumorigenicity of embryonal carcinoma as an assay to study control of malignancy be the murine blastocyst. Proc Natl Acad Sci USA 76:6649-6651
21. Pierce GB. Pantazis CG. Caldwell JE, Wells RS (1982) Specific- ity of control of tumor formation by the blastocyst. Cancer
22. Pierce GB, Aguilar 13, Hood G . Wells RS (1984) Trophecto- derm in control of niurine embryonal carcinoma. Cancer Res
23. Stewart CL (1982) Formation of viable chimeras by aggrega- tion between teratocarcinomas and preimplantation mouse em- bryos. J Embryol Exp Morphol 67:167-179
24. Wells RS (1982) An in vitro assay for regulation of embryonal carcinoma by the blastocyst. Cancer Res 42: 27362741
84: 1 3-28
R a 42:1082-1087
44:3987-3996
Received August 1986 / Accepted in revised form January 20, 1987