the diatoms of timpanogos cave national monument, utah
TRANSCRIPT

The Diatoms of Timpanogos Cave National Monument, UtahAuthor(s): Larry L. St. Clair and Samuel R. RushforthSource: American Journal of Botany, Vol. 63, No. 1 (Jan., 1976), pp. 49-59Published by: Botanical Society of AmericaStable URL: http://www.jstor.org/stable/2441667 .
Accessed: 19/04/2013 08:56
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Botanical Society of America is collaborating with JSTOR to digitize, preserve and extend access to AmericanJournal of Botany.
http://www.jstor.org
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions

Amer. J. Bot. 63(1): 49-59. 1976.
THE DIATOMS OF TIMPANOGOS CAVE NATIONAL MONUMENT, UTAH'
LARRY L. ST. CLAIR AND SAMUEL R. RUSHFORTH Department of Botany and Range Science, Brigham Young University, Provo, Utah 84602
A B S T R A C T The diatom flora of the cavern system of Timpanogos Cave National Monument was in-
vestigated. Diatoms were found throughout the cave system and were distributed according to moisture and natural openings. Twenty-six species were identified and described, including eight not previously collected from caves and four new records for the state of Utah.
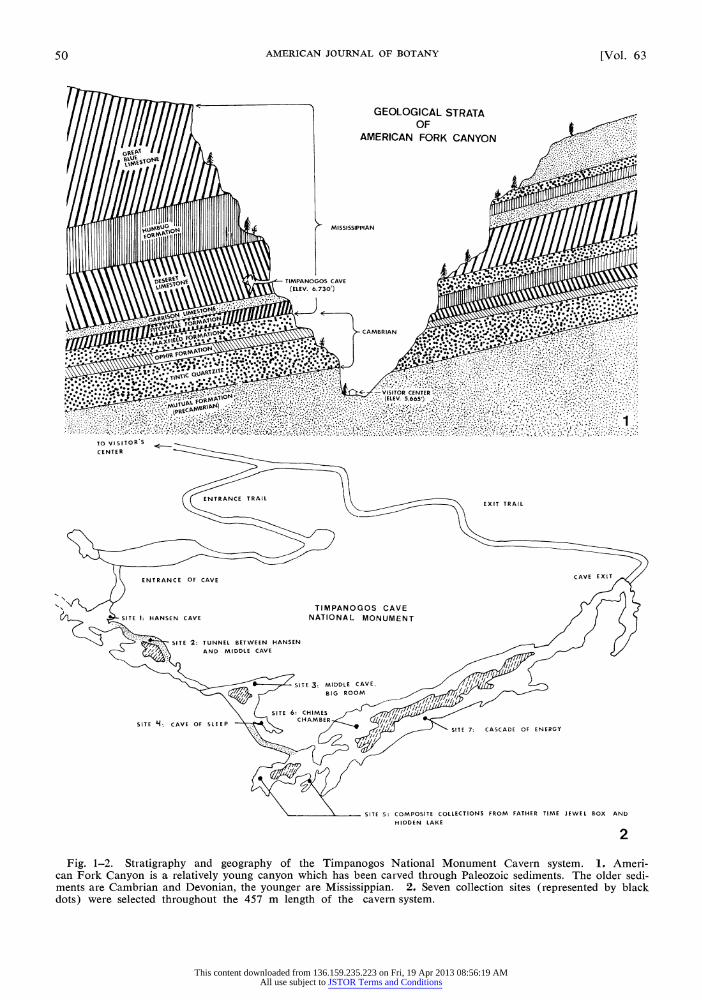
TIMPANOGOS CAVE National Monument, located in American Fork Canyon, Utah County, Utah, was established for a noteworthy limestone cavern system in the Wasatch Mountains. This system is composed of three caves, Hansen, Middle, and Timpanogos, which have been interconnected by short manmade tunnels. The total combined length of the caves and their interconnecting tun- nels is approximately 460 m. These caves pres- ently lie at an elevation of about 2300 m above sea level in a southern exposure of nearly vertical limestone cliffs.
American Fork Canyon is a young, steep-walled canyon carved mostly through Paleozoic sediments (Fig. 1). The lowermost rocks in the canyon are Cambrian and Devonian in age. They have a com- bined exposed thickness of approximately 305 m. Mississippian limestones comprise the remainder of the rocks and are about 458 m thick (Bullock, 1942).
According to Bullock (1942), three distinct geological epochs have resulted in the formation of the cavern system. The first, or faulting epoch, produced fissures which allowed the development of the caves. Two separate faults were involved, and all indications are that faulting was in recent geologic time. The second, or excavational epoch, was a period of dissolution of limestone from the caves. This was apparently accomplished largely by surface streams which were concentrated along the fissures produced during faulting. The final, or depositional epoch, was a period of deposition of sediments in the cave. These sediments were largely deposited in rather flat beds and reach a maximum thickness of 2.4 m.
The Timpanogos Cavern system was first dis- covered in the late 1800's when Hansen Cave was found. Timpanogos Cave was discovered in 1921 and opened for public inspection in 1922. A
1 Received for publication 3 September 1974. This paper was partially supported by Brigham Young
University research grants. The authors wish to thank the personnel at Timpanogos Cave National Monument for their cooperation and help. We also wish to thank Lorna Raty for her help in preparing the illustrations.
permanent lighting system was established by 1925, and the cave system became an important local attraction.
Soon after the lighting system was established, it was noted that several of the important natural formations in the caves began to be discolored. In addition, the areas immediately around the lights began to support thick growths of moss gameto- phytes. The discoloration of certain formations due to moss and algae apparently has become in- tensified during the past few years. As a result of these problems, we began an investigation of several prominent cave formations and other lo- calities in the cave system in order to determine what was growing there and to suggest possible control measures. The present paper treats the diatom species in the Timpanogos Cavern system.
MATERIALS AND METHODS-Thirteen samples were collected from seven sites located in the Han- sen, Middle, and Timpanogos caves in the Tim- panogos Cave National Monument on September 19, 1973. Most samples were taken from areas close to incandescent lights situated throughout the cave system. Samples were collected under sterile conditions, with presterilized instruments and col- lecting vials. Most samples consisted of scrapings from algae-moss associations. However, some samples were taken by scraping dry incrusted al- gae from cave formations, and others were ob- tained by scraping a mucilaginous substance from certain seep walls.
The seven collecting sites were designated as follows (Fig. 2): No. 1, scrapings from cave wall near light in Hansen Cave; No. 2, scrapings from cave floor and wall near light in tunnel between Hansen and Middle caves; No. 3, scrapings from wall in Big Room of Middle Cave; No. 4, scrap- ings from wall near light in Cavern of Sleep; No. 5, scrapings from rocks in Hidden Lake and Father Time Jewel Box (composite sample); No. 6, scrapings from wall near light in Chimes Cham- ber; and No. 7, scrapings of mucilaginous ma- terial from seep wall in Cascade of Energy.
Aqueous preparations of small portions of the 49
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions
50 AMERICAN JOURNAL OF BOTANY [Vol. 63
C ~ GEOLOGICAL STRATA
TO ~ ~ ~ ~ ~ ~ ~ ~ AEIA VISITO CAYO
GRACETRILl
) \ ENTRANCE OF CAVE r v~~ISSSSIPIA
"~~~~~~DS E TIMPANOGOS CAVE) 5
16~~~~~~~~~~~~~~~~~~~~~~
< ~ ~ ~~~~~~~~~~~~~~~ST COMPSIT COLETIN FROME FATHE TIMEON JEE MOOMEN AND(
C~~~~~~~~~Tt,1 auPRVZ11 \2) ( -;> ~~~~~~~~~~~~~~~~~~VSITEO:TINLBTWE ASNXRX
t .(ELEV NDHIDDEN LAKE
Fig. 1~~-2 StaIgah an gerpyofteTimaoo Naioa MonumEn Caver sytem 1. Aeri
To~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~ I I D I N L It K
can Fork Canyon is a relatively young canyon which has been carved through Paleozoic sediments. The older sedi- ments are Cambrian and Devonian, the younger are Mississippian. 2. Seven collection sites (represented by black dots) were selected throughout the 457 m length of the cavern system.
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions

January, 1976] ST. CLAIR AND RUSHFORTH-DIATOMS 51
samples were made upon returning to the labora- tory to determine if viable diatoms were present in the samples. Living specimens were observed in most samples despite their rather low occur- rence.
Standard methods for preparing permanent dia- tom slides were used. Portions of each sample were boiled in concentrated nitric acid and, after thorough washing with distilled water, were dried on cover slips which were then mounted in naph- rax.
Identifications were made with a Zeiss RA re- search microscope provided with Nomarski inter- ference phase-contrast accessories and a 100 x oil-immersion objective. Photomicrographs were taken of each species, and drawings were prepared from these micrographs.
RESULTs-Thirteen genera including twenty- six species of diatoms were identified during this study (Table 1). These were collected from all seven sites .throughout the cavern system. Living diatoms were obtained from each site. These dia- toms are described and discussed on the follow- ing pages.
ORDER Rhizsoleniales, FAMILY Coscinodiscaceae
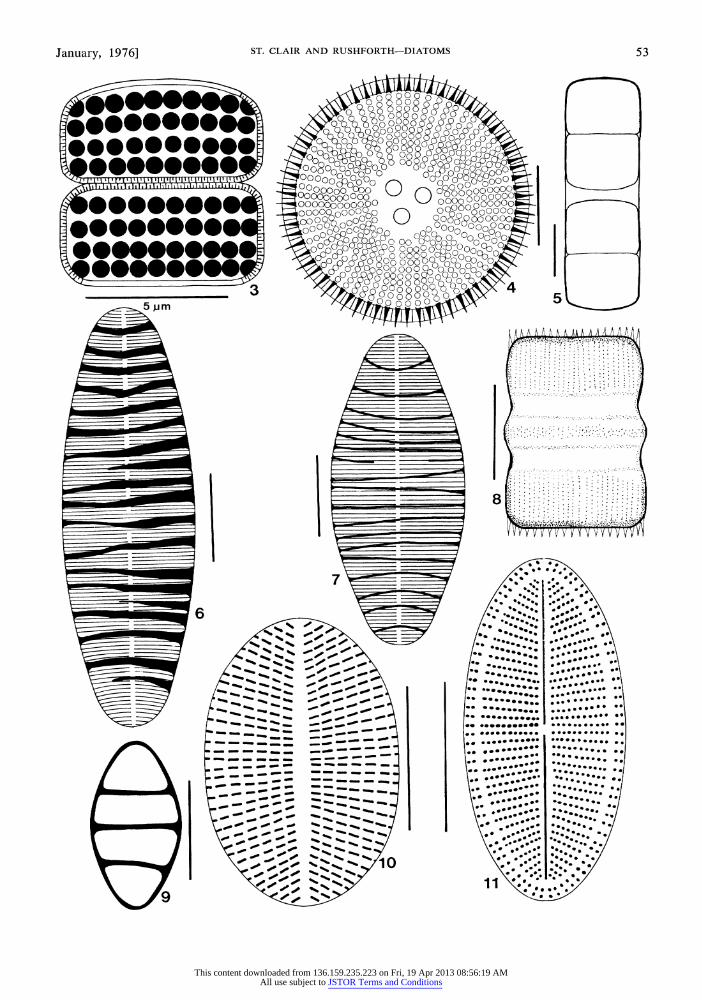
Melosira distans (Ehr.) Kiitzing (Fig. 3): Cells in girdle view 7 ,um long by 4 ,um wide, cylindrical, united into filaments; sulcus present, narrow, spined; valve in valve view punctate; punctae large, aligned to form striae; striae 12 in 10 /m. Collected only from the Big Room of Middle Cave. It was identified from a single specimen.
Melosira roeseana Rabenhorst (Fig. 4, 8): Valve in valve view round, 15-31 ,um in diameter, divided into a central and peripheral zone; cen- tral zone elevated, containing 3-4 large punctae; peripheral zone broad, radially punctate to form striations; striae approximately 10 in 10 ,um; valve in girdle view quadrangular, striate; striae paral- lel, 8 in 10 ,um. Collected from sites 1, 3, and 7. It occurred most commonly in the Big Room of Middle Cave.
Melosira varians Agardh (Fig. 5): Cells cap- sule-like in girdle view; appearing smooth, 23 pum long by 17 ,m wide, united in pairs; punctae in- distinct; sulcus not evident. Observed infrequently in the collection from the Big Room of Middle Cave.
ORDER Fragilariales, FAMILY Fragilariaceae
Diatoma hiemale var. mesodon (Ehr.) Grunow (Fig. 9): Valve 16 ,um long by 9 ,um wide, broadly elliptical; apices rounded; pseudoraphe linear, nar- row; costae 3 in 10 [um; striae fine, approximately 18-20 in 10 ,m. Identified from a single speci-
TABLE 1. Phylogenetic list of diatoms from Timpanogos cavern system
1. Division Bacillariophyta A. Class Bacillariophyceae (Diatomeae)
1. Order Rhizosoleniales a) Family Coscinodiscaceae
Melosira distans (Ehr.) Kiitzing Melosira r oeseana Rabenhorst Melosira varians Agardh
2. Order Fragilariales a) Family Fragilariaceae
Diatoma hiemale var. mesodon (Ehr.) Grunow
Diatoma vulgar-e Bory Fragilaria construens var. binodis (Ehr.)
Grunow Synedra ulna (Nitz.) Ehrenberg
3. Order Achnanthales a) Family Achnantheceae
Achnantlies coar-ctata (Breb. in W. Sm.) Grunow
Achnanthes lanecolata var. dubia Grunow
Cocconeis placentula var. lineata (Ehr.) van Heurck
4. Order Navicuales a) Family Naviculaceae
Navicula atomus (Kiitz.) Grunow Navicula contenta f. biceps Arnott Navicula cry pticephala var. veneta
(Kiitz.) Rabenhorst Navicula murrayii West and West Navicula mutica var. mutica Kuitzing Navicula mutica var. binodis Hustedt Navicula mutica var. undulata (Hilse)
Grunow Navicula permitis Hustedt Navicula radiosa Kuitzing Caloneis bacillum (Grun.) Cleve Pinnular ia intermedia (Lagerst.) Cleve
b) Family Gomphonemaceae Gomphonema olivaceum (Lyngbye)
Kiitzing 5. Order Surirellales
a) Family Nitzschiaceae Hantzschia amphioxys (Ehr.) Grunow Nitzschia fr-ustulum (Rabh.) Grunow Nitzschia linearis (Agardh) Wm. Smith
b) Family Surirellaceae Surir ella ovalis Brebisson
men found in the collection from the Big Room of Middle Cave.
Diatoma vulgare Bory (Fig. 6, 7): Valve 30- 48 Am long by 12-13 pum wide, elliptical-lanceo- late; apices rounded, slightly rostrate; pseudoraphe linear, narrow; costae 6 in 10,um; striae 8-10 in 10,utm. Collected from the Big Room of Middle Cave in moderate numbers.
Fragilaria construens var. binodis (Ehr.) Gru- now (Fig. 13): Valve 19gm long by 5pm wide,
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions

52 AMERICAN JOURNAL OF BOTANY [Vol. 63
elliptical, constricted at midvalve; apices pro- tracted, rostrate-rounded; pseudoraphe narrow, linear; striae parallel throughout, 14 in 10 ,m. Identified from a single specimen found in the col- lection from the Big Room of Middle Cave.
Synedra ulna (Nitz.) Ehrenberg (Fig. 12): Valve 280 1m long by 7 /m wide, linear; apices protracted, rostrate; pseudoraphe narrow, linear; central area quadrangular; striae parallel through- out, 10 in 10,um. Collected from the Big Room of Middle Cave.
ORDER Achnanthales, FAMILY Achnanthaceae
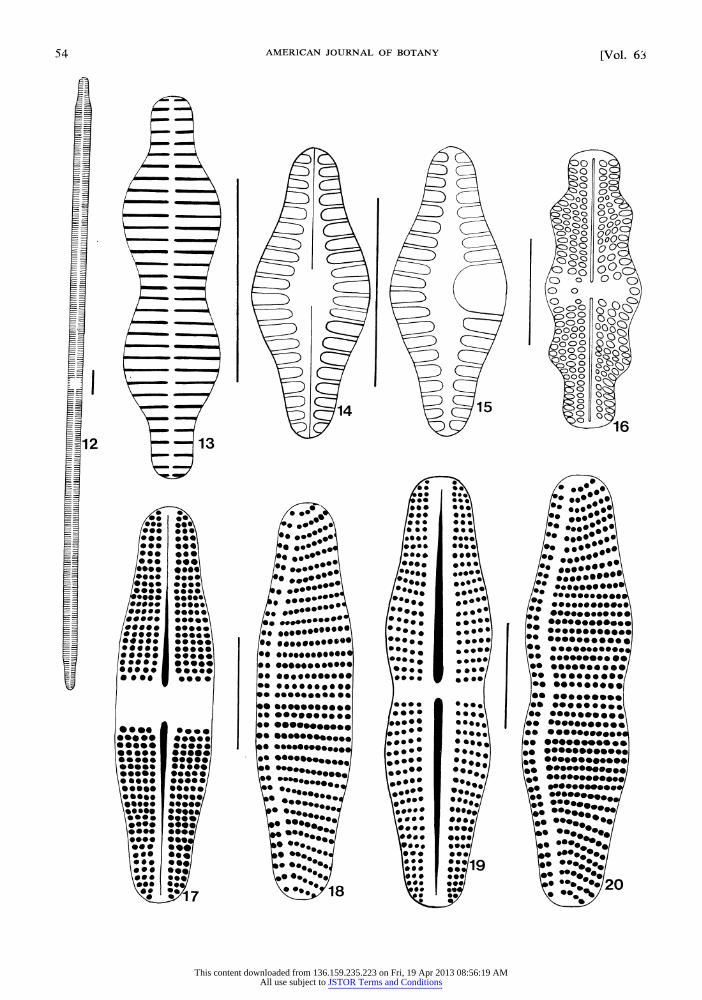
Achnanthes coarctata (Breb. in W. Sm.) Gru- now (Fig. 17-2 0): Valve 3 6-4 5 1um long by 1 0- 12 um wide, linear-elliptical, constricted near cen- ter; apices broadly rounded to truncate; raphe valve with linear, simple raphe; axial area narrow, be- coming wider near central area; central area rect- angular, extending to margins; striae radiate throughout, punctate, 14 in 10 ,m; pseudoraphe valve with narrow, excentric pseudoraphe; central area absent; striae punctate, parallel near middle, becoming radiate near apices, 8-10 in 10 ,um. Collected frequently from the Big Room of Middle Cave. Two general valve shapes were observed. These are illustrated in Fig. 17-20.
Achnanthes lanceolata var. dubia Grunow (Fig. 14-15): Valve 13-15 um long by 4-5prm wide, elliptic-lanceolate; apices protracted, rostrate to slightly capitate; raphe valve with linear, simple raphe; axial area narrow, becoming slightly wider near midvalve; central area rectangular; striae radiate, becoming convergent toward apices, 18 in 10,um; pseudoraphe valve with narrow pseudo- raphe; central area indistinct to small, containing prominent horseshoe-shaped area; striae radiate, 17 in 10 ,m. Collected in moderate numbers from the tunnel connecting Hansen and Middle Caves.
Cocconeis placentula var. lineata (Ehr.) van Heurck (Fig. 10-11): Valve 21-35 1m long by 10,um wide, elliptical to ovoid; apices broadly rounded; raphe valve with linear, filiform raphe; axial area narrow, linear; central area small to ab- sent; striae radiate throughout, 12 to 18 in 10,um, interrupted submarginally; pseudoraphe valve with narrow, linear pseudoraphe; striae radiate through- out, 22 in 10,um. Collected in low numbers from the Big Room of Middle Cave only.
ORDER Naviculales, FAMILY Naviculaceae
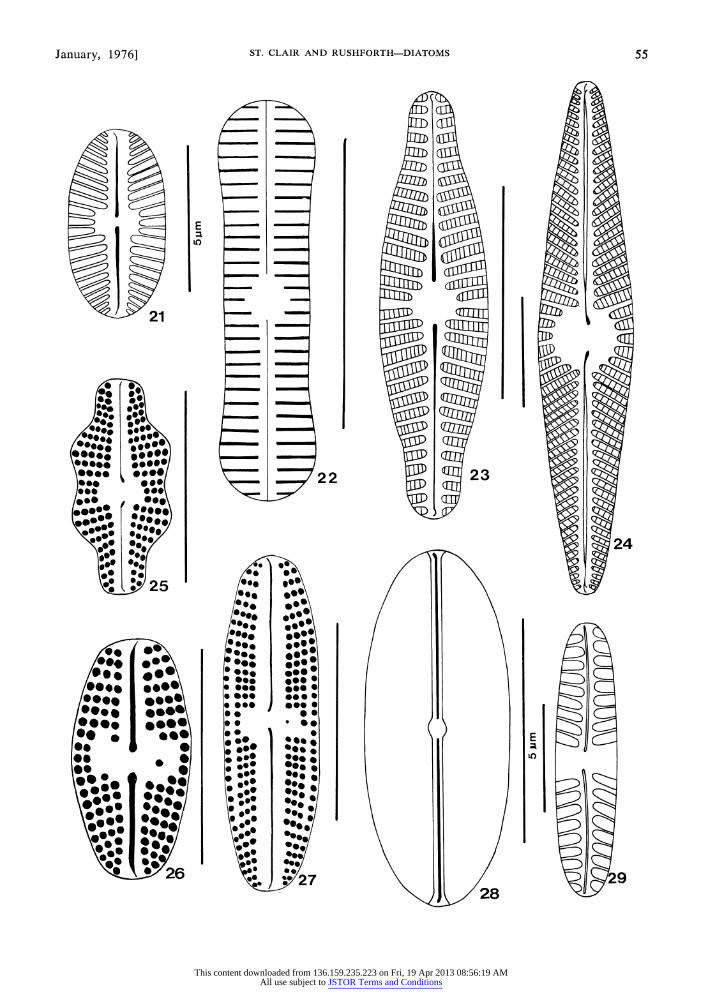
Navicula atomus (Kiitz.) Grunow (Fig. 21): Valve 5-10,um long by 2-4 /m wide, elliptical; apices broadly rounded; raphe linear, filiform; axial area rather broad, widening slightly towards central area; central area narrow, only slightly broader than axial area; striae radiate through- out, approximately 16-18 in 10 /m. Collected in moderate numbers from the Chimes Chamber.
Navicula contenta f. biceps Arnott (Fig. 22): Valve 14-15 1m long by 3-4/Am wide, linear; apices capitate; raphe linear, filiform; axial area narrow, linear, widening toward central area; cen- tral area quadrangular to orbicular; striae parallel throughout, very fine, approximately 30-35 in 10 um. Collected in relatively large numbers from all sites in the cave. This diatom was the most common of all species collected.
Navicula cryptocephala var. veneta (Kiitz.) Rabenhorst (Fig. 23): Valve 20 /m long by 5 /m wide, lanceolate; apices rounded to slightly cuneate; raphe linear; axial area narrow, linear, becoming wider near central area; central area orbicular to rather quadrate; striae radiate, irreg- ular in length at central area, 16 in 10 um. Col- lected in low numbers from the tunnel connecting Hansen and Middle Caves and from the Big Room of Middle Cave.
Fig. 3-11. 3. Melosira distans (Ehr.) Kuitzing (girdle view). 4. Melosira roeseana Rabenhorst (valve view). 5. Melosira varians Agardh (girdle view). 6-7. Diatoma vulgare Bory (valve view). 8. Melosira roeseana Ra- benhorst (girdle view). 9. Diatoma hiemale var. mesodon (Ehr.) Grunow (valve view). 10-11. Cocconeis pla- centula var. lineata (Ehr.) van Heurck. (Fig. 10, pseudoraphe valve; Fig. 11, raphe valve). Each reference line represents 10um except where noted.
Fig. 12-20. 12. Synedra ulna (Nitz.) Ehrenberg (valve view). 13. Fragilaria construens var. binodis (Ehr.) Grunow (valve view). 14-15. Achnanthes lanceolata var. dubia Grunow. (Fig. 14, raphe valve; Fig. 15, pseudo- raphe valve). 16. Navicula mutica var. undulata (Hilse) Grunow. 17-20. Achnanthes coarctata (Breb. in W. Sm.) Grunow. (Figs. 17 & 19, raphe valves; Figs. 18 & 20, pseudoraphe valves). Each reference line represents 10,um.
Fig. 21-29. 21. Navicula atomus (Kiutz.) Grunow (valve view). 22. Navicula contenta f. biceps Arnott (valve view). 23. Navicula ciyptocephala var. veneta (Kiitz.) Rabenhorst (valve view). 24. Navicula radiosa Kiitzing (valve view). 25. Navicula mutica var. binodis Hustedt (valve view). 26-27. Navicula mutica var. mutica Kuitz- ing (valve view). 28. Navicula permitis Hustedt (valve view). 29. Pinnularia intermedia (Lagerst.) Cleve (valve view). Each reference line represents 10 Am except where noted.
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions
January, 1976] ST. CLAIR AND RUSHFORTH-DIATOMS 53
_ f * ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~-
-U0 : 0
f y 9 - -- .. - .~~~~~~~~~0 . - _ E~~~~~~~~~~~~%O60008 ( ... . _ ~~~~ E_ ..... ; , ........... , - ; - .\~~~~~~~~~~~~~~~~~~~~~ 0 C
==O bA b. .: :0 : . ... .::...
e==~~~~~~~ .... a. ;0
_~~~ ~ ~~~~~~~~~~~~~~~~ `O 8c 0 ..U ........0 . 0 -~~~~~~~~~~~~~~~~~~~~~~~~ . _SV)V il 7 TIIII lIIPPi IIIPV0 0 00 n 0
.-~~~~~~~~~~~~~~~~100 000.
-~~~~~~~~~~~~~~~~~~~ 0 ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~0 '9b l - 7 A0 00 0)* ,,
_ ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~6 0, '. ., : :e a
? S g I I ,-0- '''.' SXw @''''''b''Z00 oOo 00
F _ 5o,,',"ss_210~~~~~~~~~~~~~~~~00 R.:'.,:. \ Z 'S,-, ss"S 113', . f~~~~~~~~~~~~~00_'ooOOO /9~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~0 0
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions
54 AMERICAN JOURNAL OF BOTANY [Vol. 63
0 0
000 0 0 00 0
0 0
0 00 000~
00
000 c00 00 00 0 00
12 13
S 0
0~~~~ 0 ~~~0. 000 0
0@ 0 ~~~~~~~~~0 000 0 0 00 00~~~~~~~0 00" 000
0000 0*,000, 01ooo @0 gO 000 000 0. o'o04
000 000 00 0 0 0000000 000 0.00 0@ 0000 0004 0.00*0 0000 00 00000.0oo 0000 00006 00000019000 000 *00 0 .0000 0000 0@00 0.00 0 000 000 0010 *.** 8000 000000060" 000 00. 00'00000 0 00090 00000 0 0.l0.000000 000. 0000 0. 0000 0 000 0 0.0000000 0 00 00000.00000
0000000 0.0000000 00000 0.000 0000 0000 0000 0 00 ~~0000. .00,0 000000.00000 0000 0 000,0000,000
000. 0 0
00 00000000... fee* :00:1000 000000000000 0000. .0000 0 00000 00000
0000000 00000 @000. 0090009090000 000. ..000 0 00000.0000. 0000. 00000 *.0.o.*..00000 0000
s 0000. 00000 000 .000ego, 00 0000000000o
*0 0 o 000000.00000 00000000 *0000 00000 000,0000000o 00 000000.. 0 0 00 *OOOOooe 000 .. , 00000
so 0 0000..g0 00.0o0000.
0000 0.0 0 @00000... ..00 00 0.. 0, *OOOOOoooo 00 0000000 .o0 *0o. 00 0000000000
00000 00000 ~~~~0 00000000.... 0000 000. 0 [email protected] 000 00 0000000 00000 0000. @0 **0*o0000000 00000 000000000000 000 00.000 000o"0.00g :::,e ~ ~ ~ ~ ~ ~ 0 000 00000.0. 00000 0.0 00000.00 0000000. * 0 00000000... 010000 0000 a 00.....0 0 00000 00000 096,4 o
0 0 o00000 040000 00000 000 000 '00. P0 00000 @00 0@~ 00. *00000*o00 00@0 0000 90 * 00o .0000 o00.@. 000000000 0000 0000o ihihoegg 0000 .0000* 0004 000 400 0 ;;;.o..*4 0000 :0000 * 0000000 0
00.0 0 000. 0. 000 .@0 0000 000 0060000 000 0000 000 00 0000 *** ~ O 0. o 0000 *00 o00*0 000 0000 So00. 000 00 0 00,00 @0*
000 0040 *0 000 000 0 0
Soo 0 0 ~*0 **, . , :000.00.2 0 000 0. 00 0s, 00
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions
January, 1976] ST. CLAIR AND RUSHFORTH-DIATOMS 55
E Q LO
6W<
21 111O]ID (ZUDII
0000 0 00004
as* *000d 000 % 0000 22 23 see ed 00 00 Oo 0000 0000 0000 *000 0900
24
25
000 000 **,a 000 000 000 000 40se 000 000
00* 0
E Z LO
000 0
00 'O'e:
*es *00
00
27 9
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions

56 AMERICAN JOURNAL OF BOTANY [Vol. 63
Navicula murrayii West and West (Fig. 36): Valve 20[km long by 6 [m wide, linear lanceolate; apices protracted, rounded to rostrate; raphe linear, simple; axial area narrow, linear through- out; central area rectangular, containing a single isolated punctum; striae radiate throughout, ap- proximately 16 in 10 km. Collected in low num- bers from the Big Room of Middle Cave.
Navicula mutica var. mutica Kiitzing (Fig. 26- 27): Valve 1 1-1 3 ,um long by 5 [km wide, lanceo- late to elliptical; apices rounded to slightly pro- tracted; raphe linear; axial area broad near cen- tral area, narrowing toward apices; central area large, containing a single isolated puncta; striae radiate, distinctly punctate, 16-18 in 10 [m. Col- lected from the Big Room of Middle Cave and from Father Time Jewel Box-Hidden Lake. This diatom displayed a wide variation in form as il- lustrated in Figures 26-27.
Navicula mutica var. binodis Hustedt (Fig. 25): Valve 1 1 ,um long by 5 ,um wide, lanceolate, with biundulate margins; apices protracted, cap- itate; raphe linear; axial area narrow, widened at central area; central area quadrangular; margin at central area somewhat irregular; striae radiate near middle becoming less so toward apices, 18 in 10 um. Collected infrequently from the Big Room of Middle Cave.
Navicula mutica var. undulata (Hilse) Grunow (Fig. 16): Valve 21-26[km long by 9-14[km wide, lanceolate, with triundulate margins; apices protracted, capitate; raphe linear, filiform; axial area narrow, slightly widened at central area; cen- tral area a broad transverse fascia; striae radiate, parallel approaching apices, 17-22 in 10km. Col- lected in moderate numbers from the Big Room of Middle Cave.
Navicula permitis Hustedt (Fig. 28): Valve 9- 11 [m long by 3-4 [m wide, linear-elliptic; apices broadly rounded; raphe linear; axial area narrow, linear; central area small, orbicular to quadrate; striae parallel throughout, extremely fine, approxi- mately 30-35 in 10 km. Collected in rather large numbers from the tunnel between Hansen and Middle Caves, Father Time Jewel Box-Hidden Lake, and the Cascade of Energy.
Navicula radiosa Kiitzing (Fig. 24): Valve 50 /um long by 1 1 ,um wide, lanceolate; apices acutely rounded; raphe linear, simple; axial area linear, narrow, becoming slightly broader toward central
area; central area orbicular; striae radiate through- out, 16 in 10 tm. Identified from a single speci- men collected from the Big Room of Middle Cave.
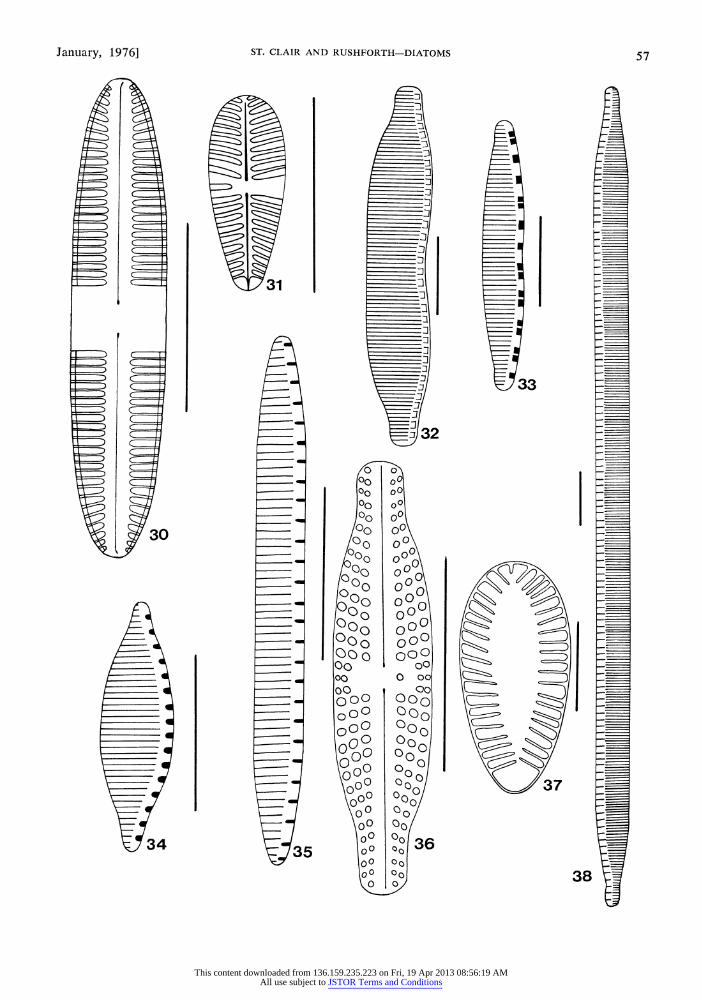
Caloneis bacillum (Grun.) Cleve (Fig. 30): Valve 28 ,m long by 6 ,m wide, linear to slightly lanceolate; apices obtusely rounded; raphe linear, simple; axial area narrow, becoming wider toward central area; central area rectangular; extending to margins; striae parallel, becoming slightly radi- ate toward apices, 22 in 10 tm. Identified from a single specimen collected from the tunnel con- necting Hansen and Middle Caves.
Pinnularia intermedia (Lagerst.) Cleve (Fig. 29): Valve 26 lxm long by 6 jum wide, linear to slightly elliptical, slightly concave at midvalve; apices rounded; raphe linear, simple, proximal ends curved in same direction; axial area narrow, becoming slightly wider near central area; central area a broad transverse fascia; striae radiate, be- coming convergent toward apices, 7 in 10 ,m. Collected in moderate numbers from the Big Room of Middle Cave.
FAMILY Gomphonemaceae Gomphonema olivaceum (Lyngbye) Kiitzing
(Fig. 31): Valve 21kum long by 5[tm wide, clavate; apical pole broadly rounded, 7 [tm wide; basal pole acutely rounded, about 4[tm wide; raphe linear, simple; axial area narrow, linear; central area round, small; striae radiate, becom- ing less so at basal pole, 12 in 10 [m. Collected in low numbers from the Big Room of Middle Cave.
ORDER Surirellales, FAMILY Nitzschiaceae
Hantzschia amphioxys (Ehr.) Grunow (Fig. 32-33): Valve 33-48 tm long by 6-7 [tm wide, linear to slightly arcuate, distinctly constricted at midvalve; apices capitate, bent upwards slightly; keel punctae 6-7 in 10/im; striae slightly radiate at midvalve, becoming parallel toward apices, 20- 21 in 10/im. Collected in low numbers from the Big Room of Middle Cave.
Nitzschia frustulum (Rabh.) Grunow (Fig. 34- 35): Valve 16-30 ktm long by 3-4 ktm wide, linear to linear lanceolate; apices acutely rounded to rostrate or slightly capitate; keel punctae 8 in 10 ,>m; striae parallel throughout, fine, 25-26 in 10 ,>m. Collected infrequently from the tunnel be-
Fig. 30-38. 30. Caloneis bacillum (Grun.) Cleve (valve view). 31. Gomphonema olivaceum (Lyngbye) Kiutz- ing (valve view). 32-33. Hantzschia amphioxys (Ehr.) Grunow (valve view). 34-35. Nitzschia frustulum (Rabh.) Grunow (valve view). 36. Navicula murrayji West and West (valve view). 37. Surirella ovalis Brebisson (valve view). 38. Nitzschia linearis (Agardh) Wm. Smith (valve view). Each reference line represents 10um.
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions
January, 1976] ST. CLAIR AND RUSHFORTH-DIATOMS 57
:3~~~~~~~
:3~ ~~
32
:3
30 0~~~~~~~~~~~~~~~~~~~~~~~: 23 23 23
(3(3 00~~: 31 :323~~
:30 230
C)
] 33
00 0
00 00o
000 0300 00 00
0o 0 00C
0 00 o0 0 0 00 00 000 (0 0
000 00 O0 (00 000 00 000 000
35 00 00
000 000 00 00 3
10 0
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions

58 AMERICAN JOURNAL OF BOTANY [Vol. 63
TABLE 2. Diatom distribution in the Timpanogos Cavern system
Site #1 Site #2 Site #3 Site #4 Site #5 Site #6 Site #7
Tunnel between
Entrance Hansen Big to and Room Cavern Father Time
Hansen Middle Middle of Jewel Box Chimes Cascade of SPECIES Cave Caves Cave Sleep Hidden Lake Chamber Energy
Melosira distans (Ehr.) Kiitzing X Melosira roeseana Rabenhorst Xa x X Melosira varians Agardh X Diatoma hiemale var. mesodon (Ehr.) X
Grunow Diatoma vulgare Bory X Fragilaria construens var. binodis X
(Ehr.) Grunow Synedra ulna (Nitz.) Ehrenberg X Achnanthes coarctata (Breb. in W. X
Sm.) Grunow Achnanthes lanceolata var. dubia X
Grunow Cocconeis placentula var. lineata X
(Ehr.) van Heurck Navicula atomus (Kiitz.) Grunow X Navicula contenta fo. biceps Arnott X X X X X X X Navicula cryptocephala var. veneta X X
(Kutz.) Rabenhorst Navicula murrayii West and West X Navicula mutica var. mutica Kutzing X X Navicula mutica var. binodis Hustedt X Navicula mutica var. undulata (Hilse) X
Grunow Navicula permitis Hustedt X X X Navicula radiosa Kiitzing X Caloneis bacillum (Grun.) Cleve X Pinnularia intermedia (Lagerst.) Cleve X Gomphonema olivaceum (Lyngbye) X
Kiitzing Hantzschia amphioxys (Ehr.) X
Grunow Nitzschia frustulum (Rabh.) Grunow X X Nitzschia linearis (Agardh) Wm. X
Smith Surirella ovalis Brebisson X
TOTAL NUMBER OF SPECIES 2 6 22 1 3 2 3 a X = presence.
tween Hansen and Middle Caves and from the Big Room of Middle Cave.
Nitzschia linearis (Agardh) Wm. Smith (Fig. 38): Valve 125,um long by 7,um wide, linear- lanceolate; slightly constricted at midvalve; apices rostrate to slightly capitate; keel punctae 13 in 10 ,um; striae fine, parallel, 30 in 10,um. Identified from a single specimen collected from the Big Room of Middle Cave.
FAMILY Surirellaceae Surirella ovalis Brebisson (Fig. 37): Valve 27
,um long by 141km wide at widest point, ovate- lanceolate; apical pole broadly rounded, 14 km wide; basal pole acutely rounded, 6 ,m wide;
pseudoraphe narrow, linear; costae submarginal, 5 in 10,um; striae radiate, 20 in 10,um. Collected in moderate numbers from the Big Room of Mid- dle Cave.
DISCUSSION-The distribution of diatoms in the Timpanogos Cavern system is sujmmarized in Table 2. This distribution at first appears to be rather difficult to explain but is apparently deter- mined largely by the amount of moisture, and sec- ondarily by the proximity of natural openings. The main entrance of the cavern system is at the mouth of Hansen Cave. Our site 1 was near this entrance but was comparatively dry. Only two species were found in our collections from that site. Site 2 was only a few meters further into the
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions

January, 1976] ST. CLAIR AND RUSHFORTH-DIATOMS 59
cave but was considerably moister than the first site, and a total of six species were obtained there.
The greatest floristic diversity occurred in the Big Room of Middle Cave where 22 species were identified from our collections. This room was the moistest of our collecting sites. A natural opening occurs at the top of this room approxi- mately 40 m from the floor. When we discovered this fact, we thought that the increased diversity here was likely due to the direct access through the opening of wind, water, and insect-borne in- ocula and perhaps light. However, we later de- termined that no light penetrates this opening to the Big Room since several bends occur in the access tunnel. Likewise, either no or very little water actually enters the cave through this open- ing. However, this room is always moist due to direct seepage of water through the limestone walls. This is particularly noteworthy following precipitation or snow melt outside of the cave. In addition, the opening does cause air currents to be generated, and it is likely that some wind-borne or insect-borne algae could enter the cave at this site. However, such algae could enter the cave just as easily at the other openings, where the in- cidence of diatoms was much lower. This infor- mation leads us to the conclusion that moisture is the most important factor determining the dis- tribution of diatoms in the cavern system.
That the proximity of natural openings is also important to distribution is further supported by the fact that the total occurrence of diatoms dropped to a maximum of three species at sites away from such openings.
Several other researchers including Carter (1971), Claus (1955, 1962a, 1962b, 1964), Friedmann (1955, 1961, 1962), Palik (1960), and Van Landingham (1964, 1966, 1967) have reported on algal floras of caves. Generally, both species diversity and biomass are low in such hab- itats. For instance, until the paper by Carter (1971) the maximum number of diatom species collected from a cave was 16 from both Abaliget Cave in Hungary (Claus, 1962a) and Mammoth Cave in the United States (Van Landingham, 1964). Carter (1971) reported 94 species from the Devil's Hole Cave in Fife, Scotland. How- ever, the samples analyzed by Dr. Carter were collected by Mr. George West in the 1920's and Dr. Carter was not precisely certain from where they were obtained. Due to the extremely high diversity noted in Devil's Hole Cave, it is likely
that at least some samples were not obtained from a typical darkened cavern environment.
Eight taxa from our study have been collected from caves for the first time. These are Melosira roeseana, Melosira distans, Diatoma hiemale var. mesodon, Achnanthes coarctata, Cocconeis pla- centula var. lineata, Navicula murrayii, Gompho- nema olivaceum, and Surirella ovalis. Four taxa are new records from the State of Utah. These are Melosira roeseana, Achnanthes coarctata, Navicula contenta f. biceps, and Navicula mur- rayii. This study has yielded the greatest diversity of cave diatoms thus far recorded from America.
Further investigation into certain aspects of this study will prove useful in answering several puz- zling questions concerning cave diatom distribu- tion and nutrition.
LITERATURE CITED
BULLOCK, KENNETH C. 1942. A study of the geology of the Timpanogos caves. M.A. thesis, Brigham Young University. Provo, Utah.
CARTER, JOHN. 1971. Diatoms from the Devil's Hole Cave, Fife, Scotland. Nova Hedwigia 21: 657- 681.
CLAUS, GEORGE. 1955. Algae and their mode of life in the Baradla Cave at Aggtelek. Acta Bot. Acad. Sci. Hung. 2: 1-26.
1962a. Beitrage zur Kenntnis der Algenflora der Abaligeter Hohle. Hydrobiologia 19: 192-222.
1962b. Data on the ecology of the algae of Peace Cave in Hungary. Nova Hedwigia 4: 55-79.
. 1964. Algae and their mode of life in the Baradla Cave of Aggtelek. Part II. Int. J. Speleol. 1: 13-17.
FRIEDMANN, I. 1955. Geitleria calcar ea n. gen. and n. sp. A new atmophytic lime-incrusting blue-green alga. Bot.Not. 108: 439-445.
. 1961. Chroococcidiopsis kashaii sp. n. and the genus Chroococcidiopsis. Osterr. Bot. Z. 108: 354-367.
1962. The ecology of the atmophytic nitrate- alga Chroococcidiopsis kashaii Friedmann. Arch. Mikrobiol. 42: 42-45.
PALIK, P. 1960. Study into the algal flora of caves. Hidrol. Kozl. 40: 417-422.
VAN, LANDINGHAM, S. 1964. Diatoms from Mam- moth Cave, Kentucky. Int. J. Speleol. 1: 517-539.
. 1966. Three new species of Cymbella from Mammoth Cave, Kentucky. Int. J. Speleol. 2: 133- 136.
1967. A new species of Gomphonema (Bacil- lariophyta) from Mammoth Cave, Kentucky. Int. J. Speleol. 2: 405-406.
This content downloaded from 136.159.235.223 on Fri, 19 Apr 2013 08:56:19 AMAll use subject to JSTOR Terms and Conditions