the camp-dependent protein kinase pathway to the snfl ... · protein kinase and invertase...
TRANSCRIPT

(:opyright 0 1992 by the Genetics Society of America
Relationship of the CAMP-Dependent Protein Kinase Pathway to the SNFl Protein Kinase and Invertase Expression in Saccharomyces cerevisiae
E. Jane Albert Hubbard, Xiaolu Yang and Marian Carlson
Department of Genetics and Development and Institute of Cancer Research, Columbia University College of Physicians and Surgeons, New York, New York 10032
Manuscript received August 2, 199 1 Accepted for publication September 23, 1991
ABSTRACT The SNFl protein kinase and the associated SNF4 protein are required for release of glucose
repression in Saccharomyces cereuisiae. To identify functionally related proteins, we selected genes that in multicopy suppress the raffinose growth defect of snf4 mutants. Among the nine genes recovered were two genes from the CAMP-dependent protein kinase (cAPK) pathway, MSZl and PDE2. Increased dosage of these genes partially compensates for defects in nutrient utilization and sporulation in snfl and snf4 null mutants, but does not restore invertase expression. These results suggest that SNFl and cAPK affect some of the same cellular responses to nutrients. To examine the role of the cAPK pathway in regulation of invertase, we assayed mutants in which the cAPK is not modulated by CAMP. Expression of invertase was regulated in response to glucose and was dependent on SNFl function. Thus, a CAMP-responsive cAPK is dispensable for regulation of invertase.
T HE yeast Saccharomyces cerevisiae prefers to utilize glucose as a carbon source. When glucose is
plentiful, the cell represses the expression of genes that are involved in utilization of alternate carbon sources, a phenomenon known as glucose repression. This regulatory response is important to the cell and apparently involves many regulatory genes; however, the signaling pathway is not yet understood [for re- view see ENTIAN (1986), GANCEDO and GANCEDO (1 986) and CARLSON ( 1 987)].
One of the key genes is SNFI , which encodes a protein-serine/threonine kinase that is required for release of gene expression from glucose repression (CELENZA and CARLSON 1986). SNFl is the same gene as CAT1 and CCRl (CIRIACY 1977; DENIS 1984; EN- TIAN and ZIMMERMANN 1982; SCHULLER and ENTIAN 1987). Mutations in SNFl cause defects in growth on carbon sources that are less preferred than glucose, general unhealthiness, and defects in sporulation of homozygous diploids (CARLSON, OSMOND and BOT- STEIN 198 1). While the SNFl kinase is clearly essential for the regulatory response to glucose, its exact role remains unclear. The SNFl kinase activity detected in vitro is not affected by the availability of glucose in the culture medium (CELENZA and CARLSON 1989); however, the physiologically relevant targets are as yet unidentified and it remains possible that their phosphorylation is regulated.
Previously, we showed that the SNF4 gene (also known as CAT3; ENTIAN and ZIMMERMANN 1982; SCHULLER and ENTIAN 1988) encodes a protein that is functionally related to the SNFl kinase. The SNF4 protein is physically associated with the SNFl protein
(kwetics 1 3 0 71-80 Uanuary, 1992)
kinase and is required for maximal activity of the kinase in vitro, but SNF4 does not appear to convey regulatory signals (CELENZA and CARLSON 1989; CE- LENZA, ENG and CARLSON 1989; FIELDS and SONG 1989). Mutations in snf4 cause the same array of phenotypes as snfl, but are slightly less severe (NEI- GEBORN and CARLSON 1984; CELENZA, ENG and CARL-
In an effort to further our understanding of the regulatory pathway for the glucose response, we sought to identify other genes that encode proteins that are functionally related to the SNFl protein kinase. We selected for genes that in multicopy sup- press the raffinose growth defect of a snf4 mutant. Although SNFl kinase activity is greatly reduced in a snf4 mutant, genetic and biochemical evidence indi- cates that some residual SNFl kinase activity remains (CELENZA and CARLSON 1989). We selected for suppression of reduced kinase activity, rather than no kinase activity, because we anticipated that a broader range of suppressor genes would be recovered. There are various possible mechanisms by which increased dosage of a gene could compensate for the snf4 defect. For example, the suppressor genes could encode ad- ditional activators of the kinase, substrates of the kinase, activators of a parai!el or partially redundant kinase pathway, or repressors of an antagonistic path- way.
We report here that among the nine genes re- covered using this strategy were two genes from the CAMP-dependent protein kinase (cAPK) pathway, MSII and PDE2. MSZI (also called J U N l ) was previ- ously isolated as a multicopy suppressor of the heat
SON 1989).

72 E. J. A. Hubbard, X. Yang and M. Carlson
shock sensitive phenotype of iral and RAS2Va"9 mu- tants (NIKAWA, SASS and WICLER 1987; RUCCIERI et al. 1989), and PDE2 encodes a high-affinity cAMP phosphodiesterase (SASS et al. 1986; WILSON and TATCHELL 1988).
T h e role of cAMP and cAPK in glucose repression in S. cerevisiae is problematic. cAMP does not function as a direct effector by a mechanism analogous to that in Escherichia coli (MATSUMOTO et al. 1982, 1983; ERASO and GANCEDO 1984). However, a RAS-depend- ent transient elevation in cAMP levels upon addition of glucose to glucose-starved cells has been docu- mented (MBONYI et al. 1990, and references therein). Mutations in CYRl , the gene encoding adenylate cy- clase, reduce expression of invertase and a-D-glucosi- dase (MATSUMOTO, UNO and ISHIKAWA 1984; SCHULTZ and CARLSON 1987). In contrast, mutants defective in BCYl , which encodes the CAMP-respon- sive negative regulatory subunit of cAPK, express invertase, galactokinase, and a-D-glucosidase at wild- type levels or a fewfold higher, and expression is still subject to glucose repression (MATSUMOTO et al. 1983; J. SCHULTZ and M. CARLSON, unpublished results). T h e cAPK pathway is known to affect expression of the glucose repressible gene ADH2 via phosphoryla- tion of the transcriptional activator ADR 1 (BEMIS and DENIS 1988; CHERRY et al. 1989; TAYLOR and YOUNG 1990; THUKRAL et al. 1989); however, the data do not establish that this phosphorylation regulates glu- cose repression of ADH2.
We have examined the ability of increased MSIl and PDE2 gene dosage to suppress various defects in both snf4 and snfl null mutants. We found that these multicopy suppressor genes did not restore invertase expression in response to glucose deprivation, but rather seemed to compensate for defects in nutrient utilization and sporulation. We also examined the effects of the cAPK pathway on regulation of invertase expression by using mutant strains in which cAPK activity is no longer responsive to the levels of CAMP; these strains lack BCYl and carry attenuating muta- tions in the genes encoding the catalytic subunits of cAPK (CAMERON et al. 1988). In these strains, expres- sion of invertase was regulated in response to glucose and still dependent on SNFl function.
MATERIALS AND METHODS
Strains and general genetic methods: Strains of S. cere- visiae used in this study and their sources are listed in Table 1. MCY strains have the S288C genetic background except where other derivation is noted. Standard methods were used for genetic analysis (SHERMAN, FINK and LAWRENCE 1978) and transformation (ITO et al. 1983). Media contained 2% of the carbon source unless otherwise noted. Anaerobic growth was scored after incubation in a GasPak Disposable Anaerobic System (BBL).
Isolation of multicopy suppressor plasmids: Strain MCY 1853 (snf4-A2 ura3) was transformed with a genomic
library on the multicopy vector YEp24 (CARLSON and BOT- STEIN 1982). Approximately 24,000 Ura+ colonies were replica-plated onto supplemented synthetic medium (SHER- MAN, FINK and LAWRENCE 1978) containing raffinose and lacking uracil (SR-Ura). The 476 Rap transformants were retested by spotting cell suspensions onto SR-Ura, and 170 colonies again scored Rap. We selected for further study 90 colonies that grew nearly as well as or better than MCY1853 carrying SNFl on a multicopy plasmid (pCE9; CELENZA and CARLSON 1989). Plasmid DNAs were re- covered by passage through E. coli (HOFFMAN and WINSTON 1987). Five plasmids carrying SNF4 and 33 carrying SNFl were identified by diagnostic restriction digests and South- ern blot analysis. Nine different plasmids, which conferred significant suppression upon retransformation of MCY1853, accounted for 37 of the remaining 52 plasmids.
Plasmid constructions: DNA was manipulated and ana- lyzed using standard methods (MANIATIS, FRITSCH and SAM- BROOK 1982). pJHl0 contains the SalI/EcoRI fragment of pB37 cloned into pUC19 (YANISCH-PERRON, VIEIRA and MESSING 1985). pJH39 carries the 1.4-kb NruI fragment of pB37 cloned into the SmaI site of YEp24 (BOTSTEIN et al. 1979). To construct pJH44, pB37 was digested with XhoI plus BglII, the ends were filled in with Klenow fragment, and the vector-containing fragment was gel-purified and ligated. pJH46 and pJH49 contain the ClaI/SmaI and SalI/ XhoI fragments of pB37 cloned into the BamHI/SmaI and Sal1 sites of YEp24, respectively. pXY 1 contains the Sal1 to XbaI fragment of pB88 ligated to the large SalIINheI frag- ment of YEp24. pXY2 was constructed by cloning the XbaI/ SmaI fragment of pB88 into the NheIISmaI site of YEp24. pXY7 and pXY8 were constructed by cloning the SphI/ScaI and BamHIINheI fragments of pB88 between the SphI/SmaI and BamHIINheI sites of YEp24, respectively.
Sequence analysis and computer methods: Restriction fragments were cloned into M 13 mp 18 or M 1 3 mpl9 (NOR- RANDER, KEMPE and MESSING 1983) and sequenced by the method of SANGER, NICKLEN and COULSON (1977) using Sequenase (U.S. Biochemical) and a 17-mer sequencing primer. Sequences were analyzed using DNA Strider (Com- missariat a I'Energie Atomique-France) and the GCG pack- age (DEVEREUX, HAEBERLI and SMITHIES 1984). Searches of reported sequences were made in the GenBank (BILOFSKY et al. 1986) and EMBL (HAMM and CAMERON 1986) data- bases using the GCG program TFASTA (PEARSON and LIPMAN 1988).
Construction of the msil-Al::URA3 allele: First, the plas- mid pJH5 1 was constructed by ligating the KpnIISphI frag- ment of pB37 to the KpnIISphI fragment of pUCl9. pJH5l was then cut with ClaI and BglII, filled in with Klenow, isolated, and ligated to a 1. l-kb SmaI fragment containing URA3 to obtain pJH52. The KpnI/SphI fragment of pJH52 was used to transform (ROTHSTEIN 1983) the diploid MCY 1093 X MCY 1094 to uracil prototrophy. The presence of the msil-Al::URA3 allele on one homologue of the diploid was confirmed by Southern blot analysis using probes pre- pared from pJH5 1.
Construction of the snfl-l5::LEU2 allele: pJH80 was constructed by inserting the BglII fragment containing the LEU2 gene into the BglII site of the SNFl gene in pCC107 u. CELENZA and M. CARLSON, unpublished results). pCC107 contains SNFl on a HincII- BamHI partial fragment cloned in pUC18. The BamHI/SphI fragment of JH80 was used to transform SP1, TF1 .5prFHR, TF1.5prC H f and MCY2372. This results in a disruption of SNFl at codon 175 (CELENZA and CARLSON 1986).
Southern blot and Northern blot analysis: Standard methods were used for preparation and analysis of genomic

Yeast cAPK Pathway and Invertase 73
TABLE 1
List of S. cerevisiae strains
Strdill Genotype Source
MCY 1093 MCY 1094 MCY 1647 MCY 1845 MCY 1846 MCY 1853 MCY2 17 1 TF1.5prF"' TFl .5prCHs SP 1 MCY2344 MCY2346 MCY2367 MCY2369 MCY2372
MATa his4-539 lys2-801 ura3-52 SUC2 MATa ade2-I01 ura3-52 SUC2 MATa snf4-AI lys2-801 ura3-52 leu2::HIS3 (his3-A200?) SUC2 MATa snfl-AI0 ade2-I01 ura3-52 SUC2 MATa snfl-AI0 lys2-801 ura3-52 SUC2 MATa snf4-A2 his4-539 lys2-801 ura3-52 SUC2 MATa msil-AI::URA3 his4-539 lys2-801 ura3-52 SUC2 MATa t p k P tpkZ::HIS3 tpk3::TRPI bcyl::URA3 his3 leu2 ura3 trpl ade8 (canl?) SUC MATa tpkl" tpk2::HIS3 tpk3::TRPI bcyl::URA3 his3 leu2 ura3 trpl ade8 (canl?) SUC MATa his3 leu2 ura3 trpl ade8 canl SUC Derivative of TF1 .5prCHS with snfl-I5::LEU2 Derivative of TF1 .5prFHR with snfl-I5::LEU2 Derivative of SPl with snfl-l5::LEU2 MATa snfl-I5::LEU2 leu2-3,112 his3-A200 ura3-52 SUC2 MATa his3-A200 leu2-3.1 I2 ura3-52 SUC2
This laboratory This laboratory This laboratory This laboratory This laboratory This laboratory This work M. WICLER M. WICLER M. WICLER This work This work This work This work This laboratorv
DNA (HOFFMAN and WINSTON 1987; MANIATIS, FRITSCH and SAMBROOK 1982). Poly(A)-containing RNAs were iso- lated from glucose-repressed and -derepressed cell cultures as described previously (SAROKIN and CARLSON 1985).
Invertase assays: Glucose-repressed and derepressed cells were prepared and assayed as described previously (NEICE- BORN and CARLSON 1984). Repressed cultures were grown to mid-log phase in 2% glucose, and cells were derepressed by shifting to 0.05% glucose for 2.5 hr when prepared in rich media and 3 hr when synthetic media were used. Cells carrying plasmids were grown in supplemented synthetic medium with selection to maintain the plasmid. Secreted invertase activity was assayed as previously described (GOLD- STEIN and LAMPEN 1975; CELENZA and CARISON 1984).
Analysis of sporulation efficiency: Transformants of MCY1853 carrying pB37, pXY8 or YEp24 were crossed to MCY 1647 to generate diploids homozygous for snf4. MCY1845 transformants carrying pB37, pXY8 or YEp24 were crossed to MCY 1846 to generate diploids homozygous for snfl. MCY 1094 carrying YEp24 was crossed to MCY1093 to generate the wild-type diploid. Single colonies were isolated on supplemented synthetic medium with se- lection for the plasmid and then transferred to solid sporu- lation medium (SHERMAN, FINK and LAWRENCE 1978). After incubation for 8-9 days (6 days for the wild type) at room temperature, at least 500 cells from each colony were ex- amined microscopically.
RESULTS
Isolation of multicopy suppressor plasmids: The snf4-A2 mutant strain MCY1853 was transformed with a yeast genomic library in a multicopy vector, and transformants able to grow on raffinose were identified (see MATERIALS AND METHODS). The snf4 mutant fails to derepress invertase, which catalyzes the extracellular hydrolysis of raffinose. In addition to plasmids carrying SNF4 and SNFI, we recovered nine different plasmids that conferred a range of growth phenotypes (see MATERIALS AND METHODS). One of the plasmids contained the gene MSNI, which was previously isolated as a multicopy suppressor of a snfl-ts mutation (ESTRUCH and CARLSON 1990). Two
other plasmids, which allowed slow growth on raffi- nose, are described here.
Identification of MSZl as a multicopy suppressor of snf4: A plasmid designated pB37 (Figure 1) was recovered from 19 independent transformants. In the first experiment to characterize the insert, the labeled plasmid pJH 10 was hybridized to a Northern blot of poly(A)-containing RNA prepared from glucose-re- pressed and derepressed cells. A 1.7-kb RNA was detected, which was highly abundant in glucose-re- pressed cells and present only at low levels in dere- pressed cells. Sequence analysis starting at the KpnI site (Figure 1) revealed an open reading frame. The partial sequence of 82 codons was compared with sequences in the database (see MATERIALS AND METH- ODS). The sequence showed complete identity with the PGIl gene of S . cerevisiae (TEKAMP-OLSON, NA- JARIAN and BURKE 1988). Comparison of the restric- tion maps indicated that pB37 contains only the 3' end of P G I l .
T o identify the region in pB37 responsible for the suppression phenotype, we constructed several sub- clones in a multicopy vector (pJH39, pJH44, pJH46, and pJH49; Figure 1) and tested their ability to sup- press the R a f phenotype of a snf4 mutant. These data showed that the complementing region lies 3' to PGII . We therefore inspected the sequence 3' to PGIl re- ported by TEKAMP-OLSON, NAJARIAN and BURKE (1988) for the presence of another open reading frame (ORF). An ORF extending more than 900 bp was identified, and comparison to the sequences in the database (see Methods) revealed identity to the gene MSII. The coding sequences for M S I l and P G l l are separated by 356 bp. The two genes were both mapped genetically to the right arm of chromosome 113: MSIl lies -4 cM from tyrl and 33 cM from lys2 (RUGGIERI et al. 1989), and PGIl lies 15 cM from tyrl and 31 cM from lys2 (MAITRA and LOBO 1977).

74 E. J. A. Hubbard, X. Yang and M. Carlson
N EC B SP m m N h Sm II 1 I I I,
MSIl + 1
Multicopy Suppression
+
pJHlO
pJH39
pJH44
pJH46
pJH49 + URA3
pJH52 rnsil-Al::URA3 - 1 kb
FIGURE 1 .-Restriction map of plasmid pB37 and derivatives. Plasmids are described in the text. The open bar indicates the genomic yeast DNA sequence. The shaded area indicates the region that was sequenced. Part of the vector DNA is represented by the thin line. Heavy line indicates the segment of pB37 included in each derivative. Suppression of the snf4A defect in growth on raffinose is shown for each of the multicopy plasmids. Arrows indicate the 5' to 3' direction of the ORFs. Restriction sites: B, BglII; C, ClaI; E, EcoRI; H, HindIII; K , KpnI; N , NruI; Nh, NheI; S, SalI; Sm, SmaI; Sp, SphI; Xb, XbaI; Xh, XhoI. pJH52 was used to construct the chromosomal allele msil-AI::URA3.
MSIl (also calledJUN1) was isolated as a multicopy suppressor of the heat shock-sensitive phenotype of iral and RAS2va"9 mutants and also restores a low level of sporulation to the homozygous diploids (NI- KAWA, SASS and WIGLER 1987; RUGGIERI et al. 1989). The exact relationship of this gene to the cAPK pathway is not clear, although RUGGIERI et al. suggest that MSZl is a negative regulator of the RAS-mediated induction of CAMP. The predicted protein has re- peated sequences homologous to a repeat motif pres- ent in the f i subunit of transducin, the yeast genes TUPl, CDC4, STE4, PRP4 and the Drosophila gene Enhancer of split (RUGGIERI et al. 1989; for review, see SIMON, STRATHMANN and GAUTAM 1991 and WIL- LIAMS and TRUMBLY 1990).
pB37 contains the entire MSIl gene, and MSIl corresponds to the functional region of pB37 (Figure 1). Thus, MSIl is also a multicopy suppressor of snf4.
Multicopy MSZl suppresses pleiotropic defects of snf4 mutants: The snf4 mutation causes a variety of defects in addition to the defect in growth on raffinose (NEIGEBORN and CARLSON 1984; CELENZA, ENG and CARLSON 1989). To determine whether multicopy MSZl suppresses the other defects of snf4 mutants, we tested strain MCY1853 (snf4-A2) carrying pB37 or pJH49 for growth on synthetic medium lacking uracil and containing glucose, sucrose, raffinose, galactose, or glycerol as the carbon source. Plates were incu- bated anaerobically except for those containing glyc- erol. Because copy number can vary, four different transformants were tested. The MSZl multicopy plas- mid improved growth of the snf4 mutant on sucrose, raffinose, galactose, and glycerol relative to the con- trol strain carrying the YEp24 vector, but did not improve growth nearly as well as a multicopy plasmid carrying SNFl (data not shown; see Figure 3).
Diploid strains homozygous for snf4 are also defec- tive in sporulation. T o determine whether increased dosage of MSIl suppresses the sporulation defect, we tested snf4-A2 homozygous diploids carrying pB37 or YEp24. Two diploid colonies carrying each plasmid were induced to sporulate, as described in MATERIALS AND METHODS. A slight increase in the percentage of sporulating cells was observed in diploids carrying multicopy MSIl relative to diploids carrying the vector (Table 2). In addition, some mature asci were ob- served in the strains carrying MSIl in multicopy while no mature asci were observed in strains carrying the vector.
Disruption of the MSZl gene: Previous studies showed that deletion of MSIl did not affect growth rate, heat shock sensitivity, or sporulation of homo- zygous diploids (RUGGIERI et al. 1989). Because MSZl was here isolated as a suppressor of snf4, we con- structed a new disruption and tested for other phe- notypes previously associated with snf4 mutations. Diploids heterozygous for a deletion/substitution mu- tation in the MSIl gene, designated msil-Al::URA3 (Figure l), were constructed as described in Materials and Methods. Tetrad analysis of two diploids yielded four viable spore clones from each ascus. Seven com- plete tetrads (four from one diploid and three from the other) were tested for growth on glucose, raffi- nose, galactose, and glycerol at 30" and on glucose and raffinose at 37 O . All spore clones showed normal growth phenotypes. Invertase activity was also assayed in two complete tetrads, and the mutants were indis- tinguishable from the wild-type spore clones with re- spect to regulation of invertase activity (average de- repressed values 380 and 350, respectively). Thus no new phenotype was identified.
Identification of PDE2 as a suppressor gene: An-

Yeast cAPK Pathway and Invertase 75
TABLE 2
Multicopy MSIl and PDEZ improve sporulation of snfl and snf4 homozygous diploids
Sporulation
of colony
A B
Experiment" Relevant genotype Plasmid-borne ("/.)b
gene
1 snf4/snf4 (pB37) MSIl 7.0 3.4
2 snfl/snfl (pB37) MSll 1.6 2.6
3 snf4/snf4 (pXY8) PDE2 15.3 9.6
snf4/snf4 (YEp24) None 1.4 1.4
snfllsnfl (YEp24) None 0.4 0.2
snf4/snf4 (YEp24) None 4.1 6.0 snfllsnfl (pXY8) PDEZ 5.6 8.0 snfllsnfl (YEp24) None 1.0 0.2 Wild type (YEp24) None 42 43
Strains in a single experiment were grown and sporulated at the same time under identical conditions. Room temperature was generally slightly lower during Experiment 3, and the requirement for SNF4 is known to be less stringent at 23" than at 30" (CELENZA, ENG and CARLSON 1989).
* Percent of sporulating cells was determined as described in Materials and Methods.
other plasmid, designated pB88, was recovered once in this study (Figure 2). pB88 contains a -7.5 kb insert and carries a duplication of part of the 2-pm sequence from the vector. Sequence analysis from the XbaI site yielded a partial sequence of 7 1 codons with complete identity to the sequence of the P R T l gene of S. cerarisiae (HANIC-JOYCE, SINGER andJoHNsToN 1987). As judged by its restriction map, pB88 contains the entire P R T l gene and also the adjacent gene PDE2 (also called SRAS; WILSON and TATCHELL 1988). T o identify the region responsible for the suppression, we constructed and tested a set of subclones (pXY 1, pXY2, pXY7, pXY8; Figure 2). The results show that PDE2 is the relevant gene. PDE2 encodes a high affinity CAMP phosphodiesterase. Increased dosage of PDE2 suppresses defects caused by the RASZVa119 mu- tation, and mutations in PDEB suppress a ras2 muta- tion (CANNON, GIBBS and TATCHELL 1986; SASS et al. 1986; WILSON and TATCHELL 1988).
Four transformants of MCY 1853 (snf4-AZ) carrying
PDE2 on pXY8 were also tested for growth on syn- thetic media lacking uracil and containing sucrose, raffinose, galactose, or glycerol, as above. The plasmid improved growth on all these carbon sources relative to the control strain carrying YEp24 but was not as effective a suppressor as a multicopy plasmid carrying SNFl (data not shown; see Figure 3). Sporulation efficiency was also assessed in snf4/snf4 diploids car- rying multicopy PDE2. An increase in the percentage of sporulating cells was observed in diploids carrying PDE2 relative to diploids carrying the vector (Table 2). In addition, multicopy PDE2, like MSZl, markedly improved the formation of mature asci.
Increased dosage of MSZl and PDEB suppresses defects of snflA mutants: Previous studies indicated that the SNF4 protein is physically associated with the SNFl protein kinase and functions as an activator of the kinase (CELENZA and CARLSON 1989; CELENZA, ENG and CARLSON 1989). One possible mechanism by which increased MSZl or PDEB dosage could suppress snf4 mutant defects is by causing compensatory acti- vation of the SNFl kinase. If this were the mechanism of suppression, we would not expect multicopy MSZl or PDE2 to be able to suppress defects resulting from the complete loss of SNFl function.
T o test whether M U 1 or PDEZ can suppress defects of a snfl deletion mutation, plasmids pJH49 and pXY8 were used to transform the strain MCY 1845 (snf l -AlO). Transformants were tested for growth on raffinose, galactose and glycerol, and both genes im- proved growth on these carbon sources relative to growth of control transformants carrying the vector (Figure 3, A and B, and data not shown). Greater variation in growth phenotype was observed in strains carrying PDE2 plasmids, suggesting that suppression by PDE2 is more dependent on copy number than is suppression by MSZl. Furthermore, plasmids pB37 (MSZl) and pXY8 (PDE2) partially suppressed the sporulation defect of snfl-A10 homozygous diploids (Table 2). These results indicate that the observed suppression does not require functional SNF 1 protein and therefore occurs by a mechanism other than activation of the SNFl kinase.
0
Suppression Multicopy
PW2 + P M
PW8 + %-+
FIGURE 2.-Restriction map of plasmid pB88 and derivatives. Plasmids are described in the text. Symbols are as in Figure 1 . The shaded area indicates the region that was sequenced. Restriction sites: as in Figure 1, plus B, BamHI; Hp, HpaI; Sc, ScaI.
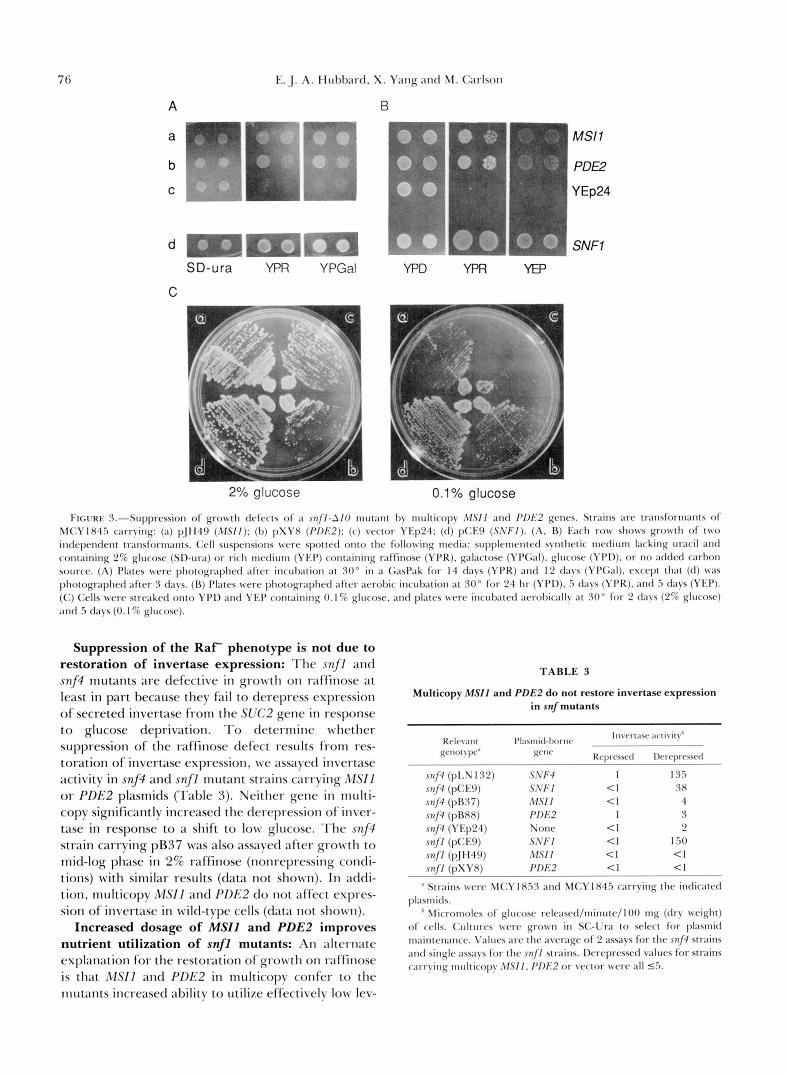
76 E:. J. A. Hut~txtrd, X . Yang and M. Carlson
1'lC;VRE 3.-Suppression of grwvth defc~cts of ;I s n f f - A f 0 nlut;lnt by nlulticopy AISfI ;und f'f)f<2 genes. Strains are transformants of 5 I ( : Y l X 4 5 carrying: (a) pJlH49 (,14Sff): (I)) p S Y X (PDB2): ( c ) vector YEp24; (tl) pCE9 (SNFf). ( A , 13) I21ch row s h o w growth of two independent transIOrm~nts. <;ell suspensions were spotted o n t o the follo\ving metlin: supplemented synthetic medium lacking tnxcil and containing 2% glucose ( S I h r a ) or rich mrtlitlnl (YEP) containing raffinost- (YI'R). gal;lctose (YPCal). g111cose (YI'D), or no atlded carhon soIn-cc. (A) I'l;~tc*s were pI~t)tt~gr~~phed after incul~ation at 30" in ;I G;~sP;tk for 14 days (YPR) and I:! d a y s (YI'Gal). except t h a t (d ) was pllotogr;lphetl alter 3 days. (13) Plates were photogl.;lphed after aerobic incubation a t 30" for 21 hr (YI'D), .5 clays (YI'R). and .5 days (YEP). (C) (:ells were strc.;lked onto YI'I) ;~nd YEP containing 0 . 1 % glucose, and pl;~tes were inculxltetl ; w d ~ i c a l l y a t 30' for 2 thys (2% glucose) ; 1 n d 5 d ; 1 y ( 0 . I % glucose).
Suppression of the R a f phenotype is not due to restoration of invertase expression: The snfl and snf4 mutants are defective i n growth on raffinose at least i n part because they fail to derepress expression of secreted invertase from the SUC2 gene in response to glucose deprivation. T o determine whether suppression of the raffinose defect results from res- toration of invertase expression, we assayed invertase activity in snf4 and snfl mutant strains carrying M S I I or PDE2 plasmids (Table 3). Neither gene i n mul t i - copy significantly increased the derepression of inver- tase in response to a shift to low glucose. The snf4 strain carrying pR37 was also assayed after growth to mid-log phase i n 2% raffinose (nonrepressing condi- tions) with similar results (data not shown). I n addi- tion, multicopy M S I l and PDE2 do not affect expres- sion of invertase in wild-type cells (data not shown).
Increased dosage of MSIl and PDE2 improves nutrient utilization of snfl mutants: An alternate explanation for the restoration of growth on raffinose is that M S I l and PDE2 in multicopy confer to the mutants increased ability to utilize effectively low lev-
TABLE 3
Multicopy MSIl and PDE2 do not restore invertase expression in snfmutants
Krlr\.;tnc l ' l ; ~ s ~ l l i ~ l - l ~ o ~ ~ ~ r 1nwrr;lsr ;trlivicv*
gcI1"cvpr~ gel" K c p r r w d 1)rtt~l)rc~s~c~l
snf4 (p1.N 132) Sh'F4 1 1 35 sn.f4 (pCE9) S,\'Ff < I :3 x
snf4 (pBX8) PIIF2 I 3
snff ( p C I 3 ) S N F f < I I50 snff (pJH49) 114Sf f < I < I s n f f ( p S Y X ) PDB2 < I < I
snf4 (pBJ7) M S I f < I 4
.$?If4 (YF.p24) None < I 2
" Strains were Ml<;Y I X.',? and M<:Y184.', c;rrrying the indicatcd p l ; w d s .
h Siicronloles of glucosr rcle;lsetl/nlin~11(./100 ~ n g (dry weight) of cells. (;ultt~rcs \wre grown in S<;-Ula to sel~ct for plasmid ~ ~ ~ ~ ~ i ~ ~ t ~ ~ ~ l ~ ~ ~ ~ c ~ ~ . \'alt~es arc the ;~vcwge of 2 assays for the snf4 strains ; t n d single assays for the m f f strains. Ikrcpressrd values for strains ravrying nlulticopy AISf f , PDF2 or vector were a 1 1 5.5.

Yeast cAPK Pathway and Invertase 77
els of hexoses and/or other nutrients present in the media. T o test this idea, we first streaked snf l -AI0 mutants carrying MSZl or PDE2 plasmids for single colonies on rich medium (YEP) containing 0.1% glu- cose as the carbon source. Figure 3C shows that both MSIl and PDE2 in multicopy improved colony for- mation relative to the control strain carrying the vec- tor. Colony size was highly variable relative to the control carrying the wild-type SNFl plasmid, and similar variation in colony size was also observed on selective synthetic medium (data not shown); variation in plasmid copy number may be responsible. We also spotted cell suspensions onto YEP medium with no added carbon source, and both MSIl and PDE2 in multicopy improved the growth of the snf l -AI0 mu- tant strain (Figure 3B). Thus, the weak restoration of growth on raffinose and other carbon sources may be attributable to improved ability to utilize low levels of nutrients rather than to suppression of the defects in derepression of specific enzymes such as invertase.
CAMP-responsive cAPK is dispensable for regu- lation of invertase by glucose: The recovery of two genes from the CAMP-dependent protein kinase (cAPK) pathway as suppressors of snf4 prompted us to explore further the relationship of cAPK to glucose regulation. We tested whether this pathway is re- quired for regulation of invertase expression in re- sponse to glucose availability by using two mutant strains, TF1.5prCHS and TF1.5prFHR (Table l), in which cAPK activity is not regulated by cAMP (M. WIGLER, personal communication). These strains carry a bcyl null allele and mutations in the three genes encoding the catalytic subunit of cAPK (TODA et al. 1987b): null mutations in TPK2 and TPK3 and an attenuating mutation in T P K l ( tpk l" ) . These strains, referred to as bcyl tpk" strains, have cAPK activity that is attenuated and independent of the level of cAMP in the cell (see CAMERON et al. 1988). TF1 .5prCHS is heat shock sensitive and TF1 .5prFHR is heat shock resistant due to different degrees of atten- uation of the cAPK activity encoded by the tpkl" allele (M. WICLER, personal communication). We confirmed that our isolates of these strains displayed the expected heat shock phenotypes. We then assayed invertase activity in the two mutant strains and their parent SP1 after growth under glucose repressing conditions (2% glucose) and after a shift to derepressing conditions (0.05% glucose). All three strains showed high level, glucose-repressible invertase activity. Thus, regula- tion of invertase activity in response to glucose does not require modulation of cAPK activity by CAMP.
T o determine if derepression of invertase requires SNFI function in the bcyl tpk" strains, we introduced the snfl-I5::LEU2 mutation into these strains (see MATERIALS AND METHODS). The snfl mutant deriva- tives failed to derepress invertase in response to glu-
TABLE 4
Regulation of invertase expression in bey1 tpk" strains
Invertase activitf
Repressed Derepressed Strain Relevant genotype
SP 1 Wild type 1 220 TF1.5prCHs tpkl" tpk2 tpk3 bcyl 1 170 TF1.5prFHR tpkl" tpk2 tpk3 bcyl 1 140 MCY2367 snf l - I5: :LEU2 1 1 MCY2344 snfl-I5::LEU2 tpkl" 1 <1
MCY2346 snfl-I5::LEU2 tpkl" 1 1 tpk2 tpk3 bcyl
tpk2 tpk3 bcyl
a Micromoles of glucose released/minute/lOO mg (dry weight) of cells. Values are the average of 2-4 assays. For values >1, standard errors were <25%.
cose limitation (Table 4). Therefore, the regulated expression of invertase activity in the bcyl tpk" mutant strains still requires SNFl function.
DISCUSSION
We report here the isolation of two genes related to the cAPK pathway, MSIl and PDE2, as weak mul- ticopy suppressors of the raffinose growth defect of a snf4 null mutant. Increased dosage of MSIl and PDE2 only partially compensates for defects caused by a snf4 mutation, improving nutrient utilization and sporula- tion (which is a response to nutrient limitation) but not restoring invertase expression. A similar pattern of suppression is observed in a snfl null mutant, indicating that the effects of increased dosage of MSIl and PDE2 result from a partial bypass of the require- ment for SNFl kinase activity.
The cAPK pathway is known to affect nutrient utilization and sporulation (see CAMERON et al. 1988 for review), but its role in the regulation of invertase has been less certain. Previously, conditional-lethal cyrl mutations were shown to impair derepression of invertase expression, but a bcyl mutation allowed high-level derepression and normal glucose repression (MATSUMOTO, UNO and ISHIKAWA 1984; MATSUMOTO et al. 1983; SCHULTZ and CARLSON 1987; and J. SCHULTZ and M. CARLSON, unpublished results). Our studies of the bcyl tpk" strains tested the idea that the cAPK pathway mediates regulatory signals that con- trol invertase expression. These strains showed nor- mal regulation of invertase activity in response to glucose deprivation, even though cAPK activity is not modulated in response to cAMP levels. Moreover, derepression of invertase in bcyl tpk" mutant strains is dependent on SNFl function, as is true in wild type. Thus, a CAMP-responsive cAPK is dispensable for regulation of invertase. We cannot exclude that the cAPK pathway contributes to regulation, but we can conclude that an independent regulatory mechanism exists and is sufficient.

78 E. J. A. Hubbard, X. Yang and M. Carlson
It is worth noting that our experiment does not address the possibility that the cAPK pathway affects the kinetics of repression or derepression, because we examined steady-state levels of invertase activity. It also remains possible that glucose-induced changes in cAMP levels (see MBONYI et al. 1990) may be involved in regulation, independent of any modulation of cAPK activity; however, cAMP levels are dramatically elevated in bcyl tpk" strains relative to wild type (NI- KAWA et al. 1987).
Our data do not contradict the idea that cAMP or the cAPK activity may be necessary for invertase expression per se. Rather, these data specifically ex- clude the notion that modulation of cAPK activity by cAMP is required for regulation in response to glu- cose. The idea that proper functioning of the cAPK is necessary for normal levels of invertase expression is, in fact, consistent with the effect of a cyrl (condi- tional lethal) mutation. Moreover, the effects of the cAPK pathway on expression of another glucose-re- pressible gene, ADH2, could be. explained similarly: phosphorylation by cAPK could reduce the intrinsic potency of the transcriptional activator ADRl rather than mediate regulatory signals regarding glucose availability (BEMIS and DENIS 1988; CHERRY et al. 1989; THUKRAL et al., 1989; TAYLOR and YOUNG 1990). We favor the view that the cAPK pathway affects the expression of invertase but not its regula- tion in response to glucose, whereas another pathway including the SNFl protein kinase is responsible for signal transduction.
The invertase gene differs from many genes in- volved in utilization of nonpreferred carbon sources in that its expression is regulated solely by glucose repression, not by substrate induction. The cAPK pathway may indirectly affect the regulation of some genes that are both glucose repressible and inducible, such as the GAL genes, via catabolite inactivation of the cognate sugar transporters. Mutations in the cAPK pathway genes have been shown to alter the catabolite inactivation of the galactose and high affin- ity glucose transporters (RAMOS and CIRILLO 1989). Catabolite inactivation of the transporter could con- tribute to the glucose repression of GAL gene expres- sion by reducing levels of the inducer.
The snfl and snf4 mutations cause pleiotropic phe- notypes that are also associated with mutations that stimulate the cAPK pathway, such as iral , b c y l , and RAS2'""' (TODA et al. 1985, 1987a; SASS et al. 1986; CANNON and TATCHELL 1987; TANAKA, MATSUMOTO and TOH-E 1989). Among these are defects in general health, growth on carbon sources other than glucose, sporulation, thermotolerance and glycogen storage (THOMPSON-JAEGER et al. 199 1). These phenotypes in snf1 mutants may all be related to the failure to respond appropriately to glucose deprivation. [Heat
shock sensitivity could reflect a failure to arrest prop- erly in stationary phase, although snflA mutants show the same percentage of unbudded cells during growth to saturation as wild type (THOMPSON~AEGER et al. 199 1 ; E. J. A. HUBBARD, unpublished results).]
WIGLER and his colleagues have previously sug- gested that responses such as sporulation, glycogen accumulation, and heat shock resistance are under multiple regulatory controls, including CAMP-inde- pendent regulatory mechanisms (CAMERON et al. 1988; TODA et al. 1988). Also, GRANOT and SNYDER (1 99 1) showed that addition of glucose to stationary- phase cells induces CAMP-independent growth-re- lated cellular events. It may be appropriate to consider the SNFl kinase pathway as an important contributor to the general response to nutrient limitation.
THOMPSON-JAEGER et al. (199 1) suggest that the SNFl and cAPK kinases are antagonistic in their effects on thermotolerance and glycogen accumula- tion. Our recovery ofMSIl and PDE2 in this selection, and our finding that their increased dosage suppresses the snfl mutant defects in nutrient utilization and sporulation are consistent with the idea that the two kinase pathways function antagonistically with respect to these phenotypes. We attempted to assess suppres- sion of the heat shock sensitivity of snfl mutants by MSIl and PDE2, but our efforts were hampered by the poor and variable survival of snfl mutant cells during growth to stationary phase in selective syn- thetic media. A similar loss of viability was reported for RAS2"'"' mutants (TODA et al. 1985).
Taken together, these data indicate that SNFl and cAPK affect some of the same cellular functions, perhaps even sharing some common targets. How- ever, these pathways clearly also have distinct func- tions, and we present evidence here that the cAPK pathway does not play an essential role in regulating invertase expression. DENIS and AUDINO (1 99 1) have shown that SNFl and cAPK act independently to control ADH2 expression.
MSIl and PDE2 are two of nine genes recovered by selecting for multicopy suppressors of a snf4 mutant. Several of the other seven genes have also been char- acterized. Increased dosage of the MSNl gene restores high level, regulated invertase expression. MSNl en- codes a nuclear protein that displays weak DNA- binding activity and may function in transcriptional activation; it is a candidate for a target of the SNFl kinase (ESTRUCH and CARLSON 1990). Also recovered in this selection was the YCKl gene, which encodes a protein homologous to rabbit casein kinase I; YCKl and the highly similar gene YCK2 together provide an essential function (L. ROBINSON, E. J. A. HUBBARD, P. ROACH, M. CULBERTSON and M. CARLSON, personal communication). A third suppressor gene encodes a protein containing two zinc-finger motifs (F. ESTRUCH

Yeast cAPK Pathway and Invertase 79
and M. CARLSON, unpublished results). The remaining four genes include two strong suppressors that restore substantial derepression of invertase (E. J. A. HUB- BARD, unpublished results). Further study should elu- cidate the relationships of these multicopy suppressors to the SNFl protein kinase and to the regulatory mechanism for glucose repression.
We thank MICHAEL WICLER for discussion and for generously providing strains. We thank SANDRA THOMPSON-JAEGER and KELLY TATCHELL for communicating unpublished results. This work was supported by U.S. Public Health Service grant GM34095 from the National Institutes of Health, and an American Cancer Society Faculty Research Award to M.C. National Cancer Institute predoc- toral training grant CA09503 provided support for E.J.A.H.
LITERATURE CITED
BEMIS, L. T., and C. L. DENIS, 1988 Identification of functional regions in the yeast transcriptional activator ADRl. Mol. Cell. Biol. 8: 2 125-21 3 1.
BILOFSKY, H. S., C. BURKS, J. W. FICKETT and W. B. GOAD, 1986 The GenBank genetic sequence data bank. Nucleic Acids Res. 14: 1-4.
BOTSTEIN, D., S. C. FALCO, S. E. STEWART, M. BRENNAN, S. SCHERER, D. T. STINCHCOMB, K. STRUHL and R. W. DAVIS, 1979 Sterile host yeast (SHY): a eukaryotic system of biolog- ical containment for recombinant DNA experiments. Gene 8
CAMERON, S., L. LEVIN, M. ZoLLERand M. WIGLER, 1988 CAMP- Independent control of sporulation, glycogen metabolism, and heat shock resistance in S. cerevisiae. Cell 53: 555-566.
CANNON, J. F., J. B. GIB= and K. TATCHRLL, 1986 Suppressors of the ras2 mutation of Saccharomyces cerevisiae. Genetics 113:
CANNON, J. F., and K. TATCHELL, 1987 Characterization of Sac- charomyces cerevisiae genes encoding subunits of cyclic AMP- dependent protein kinase. Mol. Cell. Biol. 7: 2653-2663.
CARLSON, M., 1987 Regulation of sugar utilization in Saccharo- myces species. J. Bacteriol. 169: 4873-4877.
CARLSON, M., and D. BOTSTEIN, 1982 Two differentially regu- lated mRNAs with different 5’ ends encode secreted and intracellular forms of yeast invertase. Cell 28: 145-154.
CARLSON, M., B. C. OSMOND and D. BOTSTEIN, 1981 Mutants of yeast defective in sucrose utilization. Genetics 98: 25-40.
CELENZA, J. L., and M. CARLSON, 1984 Structure and expression of the SNFl gene of Saccharomyces cerevisiae. Mol. Cell. Biol. 4:
CELENZA, J. L., and M. CARLSON, 1986 A yeast gene that is essential for release from glucose repression encodes a protein kinase. Science 233: 1 175-1 180.
CELENZA, J. L., and M. CARLSON, 1989 Mutational analysis of the Saccharomyces cerevisiue SNFl protein kinase and evidence for functional interaction with the SNF4 protein. Mol. Cell. Biol. 9 5034-5044.
CELENZA, J. L., F. J. ENG and M. CARLSON, 1989 Molecular analysis of the SNF4 gene of Saccharomyces cerevisiae: evidence for physical association of the SNF4 protein with the SNFl protein kinase. Mol. Cell, Biol9 5045-5054.
CHERRY, J. R., T. R. JOHNSON, C. DOLLARD, J. R. SCHUSTER and C. L. DENIS, 1989 Cyclic AMP-Dependent protein kinase phosphorylates and inactivates the yeast transcriptional activa- tor ADRl. Cell 5 6 409-4 19.
CIRIACY, M., 1977 Isolation and characterization of yeast mutants defective in intermediary carbon metabolism and in carbon catabolite derepression. Mol. Gen. Genet. 154 213-220.
DENIS, C. L., 1984 Identification of new genes involved in the
17-24.
247-264.
54-60.
regulation of yeast alcohol dehydrogenase 11. Genetics 108 833-844.
DENIS, C., and D. C. AUDINO, 1991 The CCRl (SNF1) and SCH9 protein kinases act independently of CAMP-dependent protein kinase and transcriptional activator ADRl in controlling yeast ADHZ expression. Mol. Gen. Genet. (in press)
DEVEREUX, J., P. HAEBERLI and 0. SMITHIES, 1984 A comprehen- sive set of sequence analysis programs for the VAX. Nucleic Acids Res. 12: 387-395.
ENTIAN, K.-D., 1986 Glucose repression: a complex regulatory system in yeast. Microbiol. Sci. 3: 366.
ENTIAN, K.-D., and F. K. ZIMMERMANN, 1982 New genes involved in carbon catabolite repression and derepression in the yeast Saccharomyces cerevisiae. J. Bacteriol. 151: 1123-1 128.
ERASO, P., and J. M. GANCEDO, 1984 Catabolite repression in yeasts is not associated with low levels of CAMP. Eur. J. Biochem. 141: 195-198.
ESTRUCH, F., and M. CARLSON, 1990 Increased dosage of the MSNZ gene restores invertase expression in yeast mutants defective in the SNFl protein kinase. Nucleic Acids Res. 18:
FIELDS, S., and 0. SONG, 1989 A novel genetic system to detect protein-protein interactions. Nature 340 245-246.
GANCEDO, J. M., and C. GANCEW, 1986 Catabolite repression mutants of yeast. FEMS Microbiol. Rev. 32: 179.
GOLDSTEIN, A., and J. 0. LAMPEN, 1975 @-D-Fructofuranoside fructohydrolase from yeast. Methods Enzymol. 42C: 504-51 1.
GRANOT, D., and M. SNYDER, 1991 Glucose induces CAMP-inde- pendent growth-related changes in stationary-phase cells of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 88: 5724- 5728.
HAMM, G. H., and G. N. CAMERON, 1986 The EMBL data library. Nucleic Acids Res. 14: 5-10.
HANIC-JOYCE, P. J., R. A. SINGER and G. C. JOHNSTON,
1987 Molecular characterization of the yeast PRTl gene in which mutations affect translation initiation and regulation of cell proliferation. J. Biol. Chem. 262: 2845-2851.
HOFFMAN, C. S., and F. WINSTON, 1987 A ten-minute DNA preparation from yeast efficiently releases autonomous plas- mids for transformation of Escherichia coli. Gene 57: 267-272.
ITO, H., Y. FUKUDA, K. MURATA and A. KIMURA, 1983 Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 153: 163-168.
MAITRA, P. K., and 2. LOBO, 1977 Genetic studies with a phos- phoglucose isomerase mutant of Saccharomyces cerevisiae. Mol. Gen. Genet. 156: 55-60.
MANIATIS, T., E. F. FRITSCH and J. SAMBROOK, 1982 Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
MATSUMOTO, K . , I. UNO and T . ISHIKAWA, 1984 Regulation of repressible acid phosphatase by cyclic AMP in Saccharomyces cerevisiae. Genetics 108: 53-66.
MATSUMOTO, K . , I. UNO, A. TOH-E, T. ISHIKAWA and Y. OSHIMA, 1982 Cyclic AMP may not be involved in catabolite repres- sion in Saccharomyces cerevisiae: evidence from mutants capable of utilizing it as an adenine source. J. Bacteriol. 150: 277-285.
MATSUMOTO, K., 1. UNO, T. ISHIKAWA and Y. OSHIMA, 1983 Cyclic AMP may not be involved in catabolite repres- sion in Saccharomyces cerevisiae: evidence from mutants unable to synthesize it. J. Bacteriol. 156: 898-900.
MBONYI, K., L. VAN AELST, J. C. ARGUELLES, A. W. H. JANS and J. M. THEVELEIN, 1990 Glucose-induced hyperaccumulation of cyclic AMP and defective glucose repression in yeast strains with reduced activity of cyclic AMP-dependent protein kinase. Mol. Cell. Biol. 10: 4518-4523.
NEIGEBORN, L., and M. CARLSON, 1984 Genes affecting the reg- ulation of SUCP gene expression by glucose repression in Sac- charomyces cerevisiae. Genetics 108: 845-858.
6959-6964.

80 E. J. A. Hubbard, X. Yang and M. Carlson
NIKAWA, J.-I., P. SASS and M. WIGLER, 1987 Cloning and char- acterization of the low-affinity cyclic AMP phosphodiesterase gene of Saccharomyces cerevisiae. Mol. Cell. Biol. 7: 3629-3636.
NIKAWA, J., S. CAMERON, T. TODA, K. M. FERGUSON and M. WIGLER, 1987 Rigorous feedback control of cAMP levels in Saccharomyces cerevisiae. Genes Dev. 1: 931-937.
NORRANDER, J., T. KEMPE and J. MESSING, 1983 Construction of improved M 13 vectors using oligodeoxynucleotide-directed mutagenesis. Gene 26: 101-1 10.
PEARSON, W. R., and D. J. LIPMAN, 1988 Improved tools for biological sequence analysis. Proc. Natl. Acad. Sci. USA 85:
KAMOS, J., and V. P. CIRILLO, 1989 Role of cyclic-AMP-depend- ent protein kinase in catabolite inactivation of the glucose and galactose transporters in Saccharomyces cerevisiae. J. Bacteriol.
KOTHSTEIN, R. J., 1983 One-step gene disruption in yeast. Meth- ods Enzymol. 101C: 202-210.
RUGGIERI, R., K. TANAKA, M. NAKAFUKU, Y. KAZIRO, A. TOH-E and K. MATSUMOTO, 1989 MSII: a negative regulator of the RAS-CAMP pathway in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 86: 8778-8782.
SANGER, F., S. NICKLEN and A. R. COULSON, 1977 DNA sequenc- ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74: 5463-5467.
SAROKIN, L., and M. CARLSON, 1985 Comparison of two yeast invertase genes: conservation of the upstream regulatory re- gion. Nucleic Acids Res. 13: 6089-6103.
SASS, P., J. FIELD, J. NIKAWA, T. TAKASHI and M. WIGLER, 1986 Cloning and characterization of the high-affinity cAMP phosphodiesterase of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 83: 9303-9307.
SCHULLER, H.-J., and K.-D. ENTIAN, 1987 Isolation and expres- sion analysis of two yeast regulatory genes involved in the derepression of glucose-repressible enzymes. Mol. Gen. Genet.
SCHULLER, H.-J., and K.-D. ENTIAN, 1988 Molecular characteriza- tion of yeast regulatory gene CAT3 necessary for glucose de- repression and nuclear localization of its product. Gene 67:
SCHULTZ, J., and M. CARLSON, 1987 Molecular analysis of SSN6: a gene functionally related to the SNFl protein kinase of Saccharomyces cerevisiae. Mol. Cell. Biol. 7: 3637-3645.
SHERMAN, F., G. R. FINK and C. W. LAWRENCE, 1978 Methods in Yeast Genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
SIMON, M. I., M. P. STRATHMANN and N. GAUTAM, 1991 Diver- sity of G proteins in signal transduction. Science 252: 802- 808.
2444-2448.
171: 3545-3548.
2 0 9 366-373.
247-257.
TANAKA, K., K. MATSUMOTO and A. TOH-E, 1989 IRAl : an inhib- itory regulator of the RAS-cyclic AMP pathway in Saccharomyces cereviviae. Mol. Cell. Biol. 9: 757-768.
TAYLOR, W. E., and E. T. YOUNG, 1990 CAMP-dependent phos- phorylation and inactivation of yeast transcription factor ADR 1 does not affect DNA binding. Proc. Natl. Acad. Sci. USA 87: 4098-4 102.
TEKAMP-OLSON, P., R. NAJARIAN and R. L. BURKE, 1988 The isolation, characterization and nucleotide sequence of the phos- phoglucoisomerase gene of Saccharomyces cerevisiae. Gene 73: 153-161.
THOMPSON-JAEGER, S., J. FRANCOIS, J. P. G A U G H R A N ~ ~ ~ K. TATCH- ELL, 1991 DELETION OF SNFl affects the nutrient response of yeast and resembles mutations which activate the adenylate cyclase pathway. Genetics 129: 697-706.
THUKRAL, S. K., M. A. TAVIANINI, H. BLUMBERG and E. T. YOUNG, 1989 Localization of a minimal binding domain and activa- tion regions in yeast regulatory protein ADRl. Mol. Cell. Biol. 9 2360-2369.
TODA, T., I . UNO, T. ISHIKAWA, S. POWERS, T. KATAOKA, D. BROEK, S. CAMERON, J. BROACH, K. MATSUMOTO and M. WIG- LER, 1985 In yeast, RAS proteins are controlling elements of adenylate cyclase. Cell 4 0 27-36.
TODA, T. , S. CAMERON, P. SASS, M. ZOLLER, J. D. SCOTT, B. MCMULLEN, M. HURWITZ, E. G. KREBS and M. WIGLER, 1987a Cloning and characterization of BCYI: a locus encod- ing a regulatory subunit of the cyclic AMP-dependent pro- tein kinase in Saccharomyces cerevisiae. Mol. Cell. Biol. 7: 1371-1377.
TODA, T., S. CAMERON, P. SASS, M. ZOLLER and M. WIGLER, 1987b Three different genes in S. cerevisiae encode the catalytic subunits of the CAMP-dependent protein kinase. Cell 50: 277-287.
TODA, T., S. CAMERON, P. SASS and M. WIGLER, 1988 SCH9: a gene of Saccharomyces cerevisiae that encodes a protein distinct from, but functionally and structurally related to, CAMP- dependent protein kinase catalytic subunits. Genes Dev. 2: 5 17-527.
WILLIAMS, F. E., and R. J. TRUMBLY, 1990 Characterization of TUPl: a mediator of glucose repression in Saccharomyces cere- visiae. Mol. Cell. Biol. 10: 6500-6511.
WILSON, B. W., and K. TATCHELL, 1988 SRA5 encodes the low- K , cyclic AMP phosphodiesterase of Saccharomyces cerevisiae. Mol. Cell. Biol. 8: 505-510.
YANISCH-PERRON, C., J. VIEIRA and J, MESSING, 1985 Improved M 13 phage cloning vectors and host strains: nucleotide se- quences of the M13 mp18 and pUCl9 vectors. Gene 33: 103-1 19.
Communicating editor: E. W. JONES