the apical-basal cell polarity determinant crumbs regulates hippo ... · the apical-basal cell...
TRANSCRIPT

The apical-basal cell polarity determinant Crumbsregulates Hippo signaling in DrosophilaChiao-Lin Chena,b, Kathleen M. Gajewskic, Fisun Hamaratoglua,d,e, Wouter Bossuyta, Leticia Sansores-Garciaa,Chunyao Taoa, and Georg Haldera,b,d,1
aDepartment of Biochemistry and Molecular Biology, bProgram in Genes and Development, and cDepartment of Systems Biology, University of TexasM. D. Anderson Cancer Center, Houston, TX 77030; dProgram in Developmental Biology, Baylor College of Medicine, Houston, TX 77030; and eGrowthand Development, Biozentrum, University of Basel, CH-4056 Basel, Switzerland
Edited* by Sean B. Carroll, University of Wisconsin, Madison, WI, and approved August 2, 2010 (received for review March 26, 2010)
Defects in apical-basal cell polarity and abnormal expression of cellpolarity determinants are often associated with cancer in verte-brates. In Drosophila, abnormal expression of apical-basal determi-nants can cause neoplastic phenotypes, including loss of cell polarityand overproliferation. However, the pathways through which api-cal-basal polarity determinants affect growth are poorly under-stood. Here, we investigated the mechanism by which the apicaldeterminant Crumbs (Crb) affects growth in Drosophila imaginaldiscs. Overexpression of Crb causes severe overproliferation, andwe found that loss of Crb similarly results in overgrowth of imaginaldiscs. Crb gain and loss of function causeddefects inHippo signaling,a key signaling pathway that controls tissue growth in Drosophilaand mammals. Manipulation of Crb levels caused the up-regulationof Hippo target genes, genetically interacted with known Hippopathway components, and required Yorkie, a transcriptional coacti-vator that acts downstream in the Hippo pathway, for target geneinduction and overgrowth. Interestingly, Crb regulates growth andcell polarity through different motifs in its intracellular domain. Ajuxtamembrane FERM domain-binding motif is responsible forgrowth regulation and induction of Hippo target gene expression,whereas Crb uses a PDZ-bindingmotif to form a complexwith otherpolarity factors. The Hippo pathway component Expanded, an api-cally localized adaptor protein, is mislocalized in both crb mutantcells and Crb overexpressing tissues, whereas the other Hippo path-way components, Fat and Merlin, are unaffected. Taken together,our data show that Crb regulates growth through Hippo signaling,and thus identify Crb as a previously undescribed upstream inputinto the Hippo pathway.
organ size control | tumor suppressor | cell proliferation | imaginal discs
Proper establishment and maintenance of apical-basal cell po-larity in epithelial tissues is essential for animal development.
Epithelial cell polarity is established and maintained throughthe concerted actions of three conserved polarity modules, theCrumbs (Crb), atypical protein kinase C (aPKC), and Discs large(Dlg) polarity modules (1–4). The Crb complex, composed ofCrb, PALS1-associated tight junction protein (Patj), and Stardust(Sdt), and the aPKC complex, composed of aPKC, Par6, and Ba-zooka, localize to the subapical region and are important for theestablishment and maintenance of the apical domain (1–4). Dis-ruption of either apical complex in Drosophila embryos, for ex-ample, causes loss of apical-basal cell polarity, loss of apical mark-ers, and expansion of the basolateral domain. The Dlg modulecontainsDlg, Lethal giant larvae (Lgl), and Scribble (Scrib). Theseproteins localize to the basolateralmembrane and are required foreach other’s localization, and perturbation of the Dlg complexcauses loss of basolateral markers and expansion of apical markers(1–4). Interestingly, altering the expression of genes involved inthe regulation of apicobasal polarity is often associated with thedevelopment of cancer in vertebrates and can lead to the de-velopment of neoplastic tumors inDrosophila imaginal discs (2, 4,5). Imaginal discs are epithelial precursors of adult fly tissues anda widely used model system for the study of tissue growth and
patterning. Imaginal discs that are homozygous mutant for scrib,dlg, or lgl lose their apical-basal polarity and severely overgrow (1–4). Similarly, overexpression of the apical determinant Crb causesovergrowth of Drosophila imaginal discs in addition to causingdefects in cell polarity and expansion of apical domain markers tothe basolateral domain (6, 7). Each of these situations thus pro-motes an overabundance of the apical domain. Several mechanismshave been suggested to explain the observed phenotypes. For ex-ample, defects in apical-basal polarity may cause overgrowth asa resultofderegulationofmany signalingpathways as a consequenceof mistrafficking of receptors, or the polarity complex proteins mayspecifically modulate one or more growth-controlling pathways (5,8). However, although abnormal expression of apicobasal determi-nants can cause dramatic overgrowth phenotypes, the mechanismsthrough which they affect growth are poorly understood.Here, we provide evidence that Crb acts through the Hippo
pathway to regulate tissue size. Crb is a transmembrane domainprotein with a large extracellular domain and a short intracel-lular domain (9). The extracellular domain contains 28 epidermalgrowth factor-like repeats and 3 laminin G-like repeats, and theintracellular domain contains two conserved motifs, a juxtamem-brane motif (JM) and a PDZ-binding motif (PBM) (10). TheHippo pathway has emerged as a key signaling pathway that reg-ulates growth of imaginal discs (11–14). Hippo signaling limits cellproliferation in imaginal discs, and flies that lack Hippo pathwayactivity have severely overgrown discs and corresponding adultstructures. Several components of the pathway have been discov-ered, anda signal transductionpathway fromtheplasmamembraneinto the nucleus has emerged. Central to the Hippo pathway is akinase cascade involving theHippo (Hpo) andWarts (Wts) kinasesand their adaptor proteins Salvador and Mats. Active Hpo phos-phorylates and activates Wts, which then phosphorylates and in-hibits the activity of the transcriptional coactivator Yorkie (Yki).Active Yki translocates to the nucleus, where it forms a complexwith the transcription factor Scalloped to induce the expression oftarget genes that drive cell proliferation and cell survival, such ascyclin E, diap1, and the bantam miRNA. Thus, when active, HpoandWts suppress cell proliferation by restraining the activity ofYki.Several components are known to act upstream of Hpo and
Wts. The atypical cadherin Fat was identified as a receptor thatregulates the Hippo pathway (11–14). It was proposed that Fattransduces signals from Dachsous (Ds), an atypical cadherinrelated to Fat, and Four-jointed, an extracellular kinase thatphosphorylates Fat and Ds (12, 14, 15). Fat signal transductioninvolves an unconventional myosin, Dachs (D), the kinase Discs
Author contributions: C.-L.C., F.H., and G.H. designed research; C.-L.C., K.M.G., F.H.,L.S.-G., and C.T. performed research; C.-L.C., K.M.G., F.H., W.B., and G.H. analyzed data;and C.-L.C., W.B., and G.H. wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1004060107/-/DCSupplemental.
15810–15815 | PNAS | September 7, 2010 | vol. 107 | no. 36 www.pnas.org/cgi/doi/10.1073/pnas.1004060107
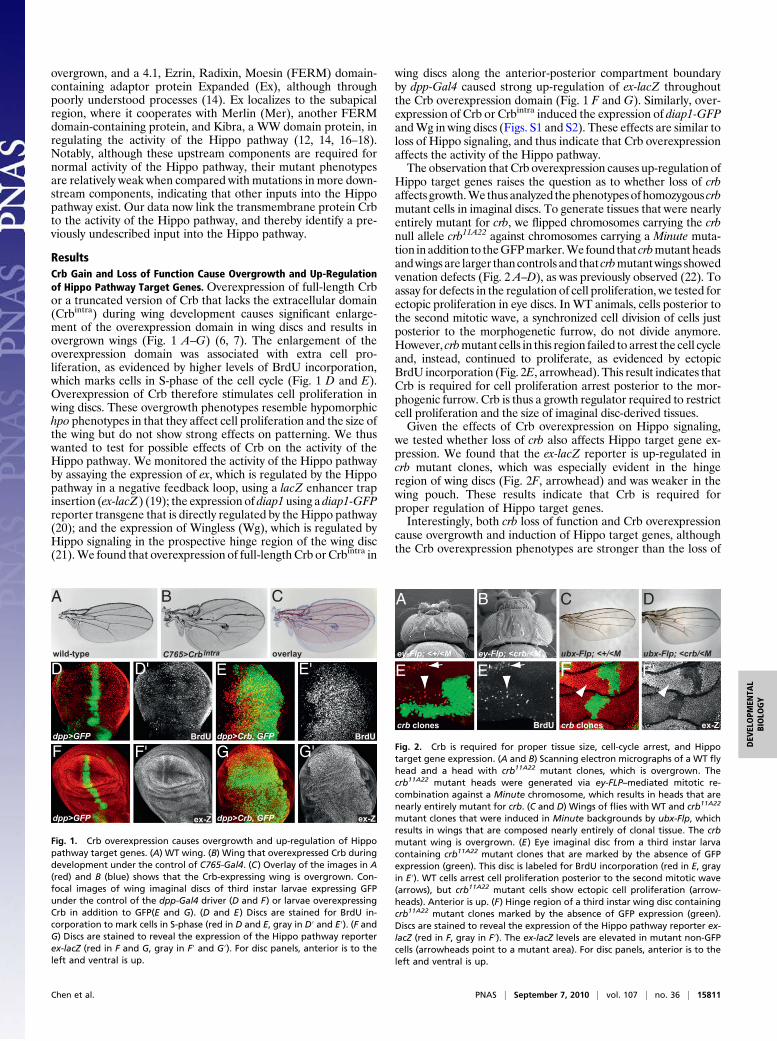
overgrown, and a 4.1, Ezrin, Radixin, Moesin (FERM) domain-containing adaptor protein Expanded (Ex), although throughpoorly understood processes (14). Ex localizes to the subapicalregion, where it cooperates with Merlin (Mer), another FERMdomain-containing protein, and Kibra, a WW domain protein, inregulating the activity of the Hippo pathway (12, 14, 16–18).Notably, although these upstream components are required fornormal activity of the Hippo pathway, their mutant phenotypesare relatively weak when compared with mutations in more down-stream components, indicating that other inputs into the Hippopathway exist. Our data now link the transmembrane protein Crbto the activity of the Hippo pathway, and thereby identify a pre-viously undescribed input into the Hippo pathway.
ResultsCrb Gain and Loss of Function Cause Overgrowth and Up-Regulationof Hippo Pathway Target Genes. Overexpression of full-length Crbor a truncated version of Crb that lacks the extracellular domain(Crbintra) during wing development causes significant enlarge-ment of the overexpression domain in wing discs and results inovergrown wings (Fig. 1 A–G) (6, 7). The enlargement of theoverexpression domain was associated with extra cell pro-liferation, as evidenced by higher levels of BrdU incorporation,which marks cells in S-phase of the cell cycle (Fig. 1 D and E).Overexpression of Crb therefore stimulates cell proliferation inwing discs. These overgrowth phenotypes resemble hypomorphichpo phenotypes in that they affect cell proliferation and the size ofthe wing but do not show strong effects on patterning. We thuswanted to test for possible effects of Crb on the activity of theHippo pathway. We monitored the activity of the Hippo pathwayby assaying the expression of ex, which is regulated by the Hippopathway in a negative feedback loop, using a lacZ enhancer trapinsertion (ex-lacZ) (19); the expression of diap1 using a diap1-GFPreporter transgene that is directly regulated by the Hippo pathway(20); and the expression of Wingless (Wg), which is regulated byHippo signaling in the prospective hinge region of the wing disc(21).We found that overexpression of full-length Crb or Crbintra in
wing discs along the anterior-posterior compartment boundaryby dpp-Gal4 caused strong up-regulation of ex-lacZ throughoutthe Crb overexpression domain (Fig. 1 F and G). Similarly, over-expression of Crb or Crbintra induced the expression of diap1-GFPandWg in wing discs (Figs. S1 and S2). These effects are similar toloss of Hippo signaling, and thus indicate that Crb overexpressionaffects the activity of the Hippo pathway.The observation that Crb overexpression causes up-regulation of
Hippo target genes raises the question as to whether loss of crbaffects growth.Wethus analyzed thephenotypesofhomozygous crbmutant cells in imaginal discs. To generate tissues that were nearlyentirely mutant for crb, we flipped chromosomes carrying the crbnull allele crb11A22 against chromosomes carrying a Minute muta-tion in addition to theGFPmarker.We found that crbmutant headsandwings are larger thancontrols and that crbmutantwings showedvenation defects (Fig. 2 A–D), as was previously observed (22). Toassay for defects in the regulation of cell proliferation, we tested forectopic proliferation in eye discs. In WT animals, cells posterior tothe second mitotic wave, a synchronized cell division of cells justposterior to the morphogenetic furrow, do not divide anymore.However, crbmutant cells in this region failed to arrest the cell cycleand, instead, continued to proliferate, as evidenced by ectopicBrdU incorporation (Fig. 2E, arrowhead). This result indicates thatCrb is required for cell proliferation arrest posterior to the mor-phogenic furrow. Crb is thus a growth regulator required to restrictcell proliferation and the size of imaginal disc-derived tissues.Given the effects of Crb overexpression on Hippo signaling,
we tested whether loss of crb also affects Hippo target gene ex-pression. We found that the ex-lacZ reporter is up-regulated incrb mutant clones, which was especially evident in the hingeregion of wing discs (Fig. 2F, arrowhead) and was weaker in thewing pouch. These results indicate that Crb is required forproper regulation of Hippo target genes.Interestingly, both crb loss of function and Crb overexpression
cause overgrowth and induction of Hippo target genes, althoughthe Crb overexpression phenotypes are stronger than the loss of
D'D
BA
E
C
G'GF'F
E'wild-type C765>Crbintra overlay
BrdU
ex-Z
BrdU
ex-Z dpp>Crb, GFP
dpp>Crb, GFP dpp>GFP
dpp>GFP
Fig. 1. Crb overexpression causes overgrowth and up-regulation of Hippopathway target genes. (A) WT wing. (B) Wing that overexpressed Crb duringdevelopment under the control of C765-Gal4. (C) Overlay of the images in A(red) and B (blue) shows that the Crb-expressing wing is overgrown. Con-focal images of wing imaginal discs of third instar larvae expressing GFPunder the control of the dpp-Gal4 driver (D and F) or larvae overexpressingCrb in addition to GFP(E and G). (D and E) Discs are stained for BrdU in-corporation to mark cells in S-phase (red in D and E, gray in D′ and E′). (F andG) Discs are stained to reveal the expression of the Hippo pathway reporterex-lacZ (red in F and G, gray in F′ and G′). For disc panels, anterior is to theleft and ventral is up.
CA B D
F'FE'E
crb clonesBrdU
ubx-Flp; <+/<M ubx-Flp; <crb/<Mey-Flp; <+/<M ey-Flp; <crb/<M
crb clones ex-Z
Fig. 2. Crb is required for proper tissue size, cell-cycle arrest, and Hippotarget gene expression. (A and B) Scanning electron micrographs of a WT flyhead and a head with crb11A22 mutant clones, which is overgrown. Thecrb11A22 mutant heads were generated via ey-FLP–mediated mitotic re-combination against a Minute chromosome, which results in heads that arenearly entirely mutant for crb. (C and D) Wings of flies with WT and crb11A22
mutant clones that were induced in Minute backgrounds by ubx-Flp, whichresults in wings that are composed nearly entirely of clonal tissue. The crbmutant wing is overgrown. (E) Eye imaginal disc from a third instar larvacontaining crb11A22 mutant clones that are marked by the absence of GFPexpression (green). This disc is labeled for BrdU incorporation (red in E, grayin E′). WT cells arrest cell proliferation posterior to the second mitotic wave(arrows), but crb11A22 mutant cells show ectopic cell proliferation (arrow-heads). Anterior is up. (F) Hinge region of a third instar wing disc containingcrb11A22 mutant clones marked by the absence of GFP expression (green).Discs are stained to reveal the expression of the Hippo pathway reporter ex-lacZ (red in F, gray in F′). The ex-lacZ levels are elevated in mutant non-GFPcells (arrowheads point to a mutant area). For disc panels, anterior is to theleft and ventral is up.
Chen et al. PNAS | September 7, 2010 | vol. 107 | no. 36 | 15811
DEV
ELOPM
ENTA
LBIOLO
GY
function phenotypes. The similarity of the loss and over-expression phenotypes indicates that WT levels of Crb are es-sential for normal functioning of the Hippo pathway.
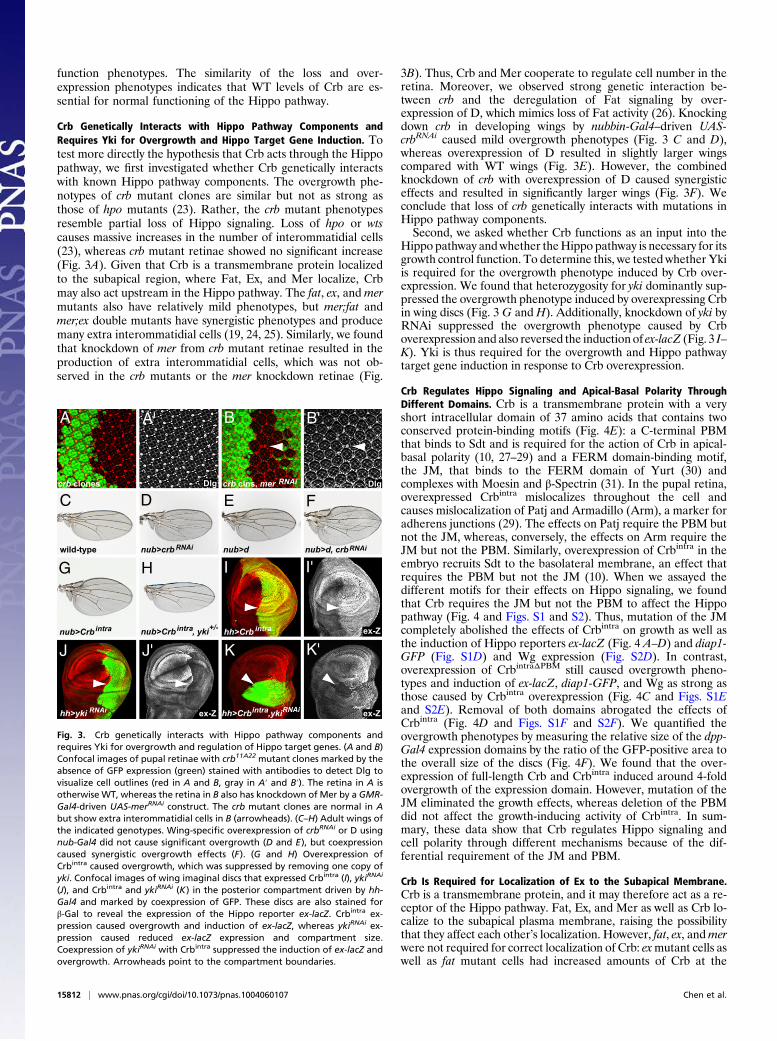
Crb Genetically Interacts with Hippo Pathway Components andRequires Yki for Overgrowth and Hippo Target Gene Induction. Totest more directly the hypothesis that Crb acts through the Hippopathway, we first investigated whether Crb genetically interactswith known Hippo pathway components. The overgrowth phe-notypes of crb mutant clones are similar but not as strong asthose of hpo mutants (23). Rather, the crb mutant phenotypesresemble partial loss of Hippo signaling. Loss of hpo or wtscauses massive increases in the number of interommatidial cells(23), whereas crb mutant retinae showed no significant increase(Fig. 3A). Given that Crb is a transmembrane protein localizedto the subapical region, where Fat, Ex, and Mer localize, Crbmay also act upstream in the Hippo pathway. The fat, ex, and mermutants also have relatively mild phenotypes, but mer;fat andmer;ex double mutants have synergistic phenotypes and producemany extra interommatidial cells (19, 24, 25). Similarly, we foundthat knockdown of mer from crb mutant retinae resulted in theproduction of extra interommatidial cells, which was not ob-served in the crb mutants or the mer knockdown retinae (Fig.
3B). Thus, Crb and Mer cooperate to regulate cell number in theretina. Moreover, we observed strong genetic interaction be-tween crb and the deregulation of Fat signaling by over-expression of D, which mimics loss of Fat activity (26). Knockingdown crb in developing wings by nubbin-Gal4–driven UAS-crbRNAi caused mild overgrowth phenotypes (Fig. 3 C and D),whereas overexpression of D resulted in slightly larger wingscompared with WT wings (Fig. 3E). However, the combinedknockdown of crb with overexpression of D caused synergisticeffects and resulted in significantly larger wings (Fig. 3F). Weconclude that loss of crb genetically interacts with mutations inHippo pathway components.Second, we asked whether Crb functions as an input into the
Hippo pathway andwhether theHippo pathway is necessary for itsgrowth control function. To determine this, we tested whether Ykiis required for the overgrowth phenotype induced by Crb over-expression. We found that heterozygosity for yki dominantly sup-pressed the overgrowth phenotype induced by overexpressing Crbin wing discs (Fig. 3 G and H). Additionally, knockdown of yki byRNAi suppressed the overgrowth phenotype caused by Crboverexpression and also reversed the induction of ex-lacZ (Fig. 3 I–K). Yki is thus required for the overgrowth and Hippo pathwaytarget gene induction in response to Crb overexpression.
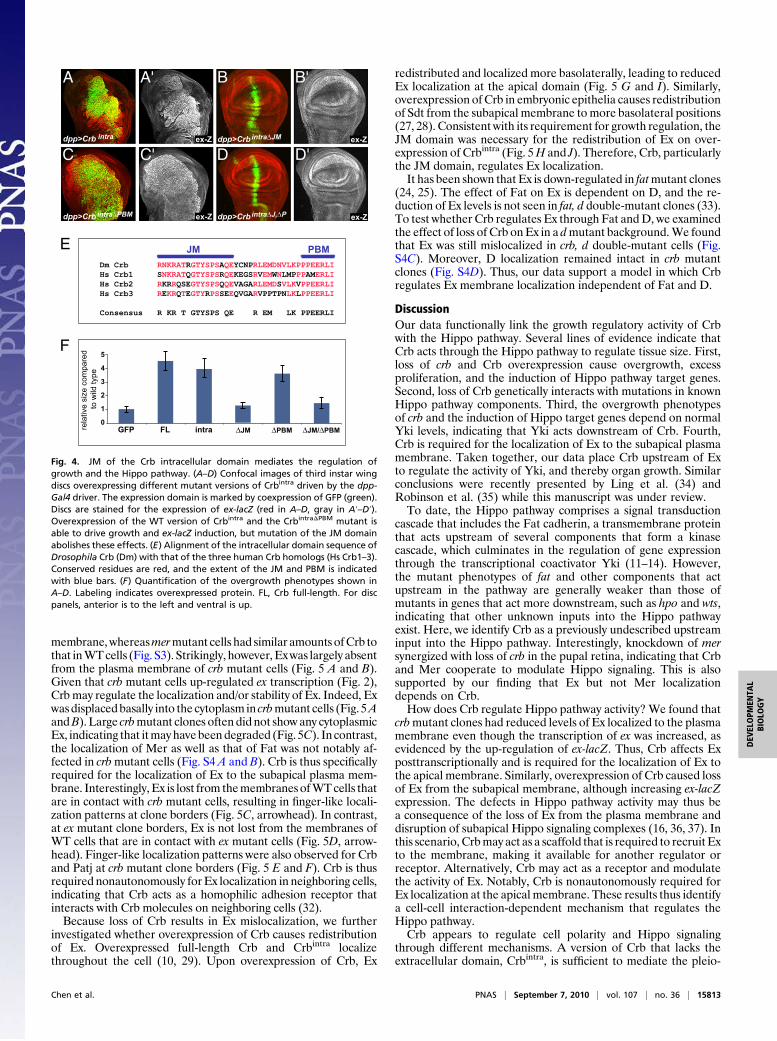
Crb Regulates Hippo Signaling and Apical-Basal Polarity ThroughDifferent Domains. Crb is a transmembrane protein with a veryshort intracellular domain of 37 amino acids that contains twoconserved protein-binding motifs (Fig. 4E): a C-terminal PBMthat binds to Sdt and is required for the action of Crb in apical-basal polarity (10, 27–29) and a FERM domain-binding motif,the JM, that binds to the FERM domain of Yurt (30) andcomplexes with Moesin and β-Spectrin (31). In the pupal retina,overexpressed Crbintra mislocalizes throughout the cell andcauses mislocalization of Patj and Armadillo (Arm), a marker foradherens junctions (29). The effects on Patj require the PBM butnot the JM, whereas, conversely, the effects on Arm require theJM but not the PBM. Similarly, overexpression of Crbintra in theembryo recruits Sdt to the basolateral membrane, an effect thatrequires the PBM but not the JM (10). When we assayed thedifferent motifs for their effects on Hippo signaling, we foundthat Crb requires the JM but not the PBM to affect the Hippopathway (Fig. 4 and Figs. S1 and S2). Thus, mutation of the JMcompletely abolished the effects of Crbintra on growth as well asthe induction of Hippo reporters ex-lacZ (Fig. 4 A–D) and diap1-GFP (Fig. S1D) and Wg expression (Fig. S2D). In contrast,overexpression of CrbintraΔPBM still caused overgrowth pheno-types and induction of ex-lacZ, diap1-GFP, and Wg as strong asthose caused by Crbintra overexpression (Fig. 4C and Figs. S1Eand S2E). Removal of both domains abrogated the effects ofCrbintra (Fig. 4D and Figs. S1F and S2F). We quantified theovergrowth phenotypes by measuring the relative size of the dpp-Gal4 expression domains by the ratio of the GFP-positive area tothe overall size of the discs (Fig. 4F). We found that the over-expression of full-length Crb and Crbintra induced around 4-foldovergrowth of the expression domain. However, mutation of theJM eliminated the growth effects, whereas deletion of the PBMdid not affect the growth-inducing activity of Crbintra. In sum-mary, these data show that Crb regulates Hippo signaling andcell polarity through different mechanisms because of the dif-ferential requirement of the JM and PBM.
Crb Is Required for Localization of Ex to the Subapical Membrane.Crb is a transmembrane protein, and it may therefore act as a re-ceptor of the Hippo pathway. Fat, Ex, and Mer as well as Crb lo-calize to the subapical plasma membrane, raising the possibilitythat they affect each other’s localization. However, fat, ex, andmerwere not required for correct localization of Crb: exmutant cells aswell as fat mutant cells had increased amounts of Crb at the
DC
BA
E
A' B'
Fcrb clones Dlg crb clns, mer
I'IHG
K'KJ'J
DlgRNAi
wild-type nub>crb RNAi nub>d, crbRNAinub>d
nub>Crbintra nub>Crb , ykiintra +/- hh>Crb intra
hh>yki RNAi hh>Crb ,ykiintra RNAi ex-Zex-Z
ex-Z
Fig. 3. Crb genetically interacts with Hippo pathway components andrequires Yki for overgrowth and regulation of Hippo target genes. (A and B)Confocal images of pupal retinae with crb11A22 mutant clones marked by theabsence of GFP expression (green) stained with antibodies to detect Dlg tovisualize cell outlines (red in A and B, gray in A′ and B′). The retina in A isotherwise WT, whereas the retina in B also has knockdown of Mer by a GMR-Gal4-driven UAS-merRNAi construct. The crb mutant clones are normal in Abut show extra interommatidial cells in B (arrowheads). (C–H) Adult wings ofthe indicated genotypes. Wing-specific overexpression of crbRNAi or D usingnub-Gal4 did not cause significant overgrowth (D and E), but coexpressioncaused synergistic overgrowth effects (F). (G and H) Overexpression ofCrbintra caused overgrowth, which was suppressed by removing one copy ofyki. Confocal images of wing imaginal discs that expressed Crbintra (I), ykiRNAi
(J), and Crbintra and ykiRNAi (K) in the posterior compartment driven by hh-Gal4 and marked by coexpression of GFP. These discs are also stained forβ-Gal to reveal the expression of the Hippo reporter ex-lacZ. Crbintra ex-pression caused overgrowth and induction of ex-lacZ, whereas ykiRNAi ex-pression caused reduced ex-lacZ expression and compartment size.Coexpression of ykiRNAi with Crbintra suppressed the induction of ex-lacZ andovergrowth. Arrowheads point to the compartment boundaries.
15812 | www.pnas.org/cgi/doi/10.1073/pnas.1004060107 Chen et al.
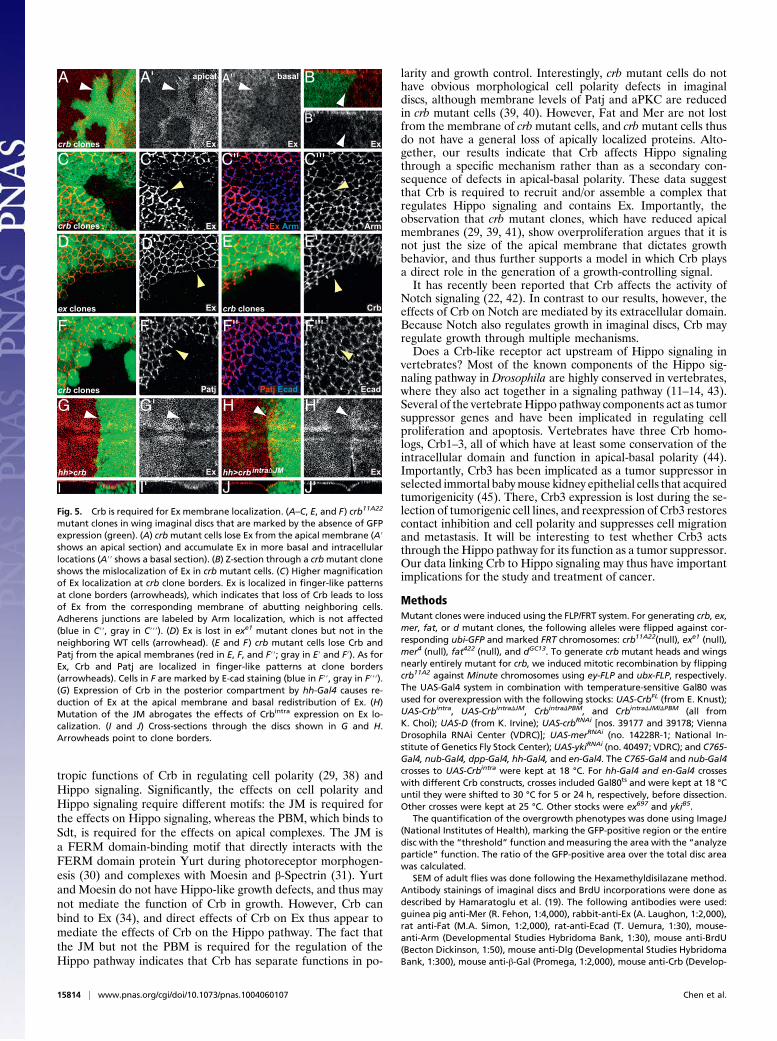
membrane,whereasmermutant cells had similar amounts ofCrb tothat inWTcells (Fig. S3). Strikingly, however, Exwas largely absentfrom the plasma membrane of crb mutant cells (Fig. 5 A and B).Given that crb mutant cells up-regulated ex transcription (Fig. 2),Crb may regulate the localization and/or stability of Ex. Indeed, Exwasdisplacedbasally into the cytoplasm in crbmutant cells (Fig. 5AandB). Large crbmutant clones often did not show any cytoplasmicEx, indicating that itmayhave beendegraded (Fig. 5C). In contrast,the localization of Mer as well as that of Fat was not notably af-fected in crbmutant cells (Fig. S4 A and B). Crb is thus specificallyrequired for the localization of Ex to the subapical plasma mem-brane. Interestingly,Ex is lost from themembranes ofWTcells thatare in contact with crb mutant cells, resulting in finger-like locali-zation patterns at clone borders (Fig. 5C, arrowhead). In contrast,at ex mutant clone borders, Ex is not lost from the membranes ofWT cells that are in contact with ex mutant cells (Fig. 5D, arrow-head). Finger-like localization patterns were also observed for Crband Patj at crb mutant clone borders (Fig. 5 E and F). Crb is thusrequired nonautonomously for Ex localization in neighboring cells,indicating that Crb acts as a homophilic adhesion receptor thatinteracts with Crb molecules on neighboring cells (32).Because loss of Crb results in Ex mislocalization, we further
investigated whether overexpression of Crb causes redistributionof Ex. Overexpressed full-length Crb and Crbintra localizethroughout the cell (10, 29). Upon overexpression of Crb, Ex
redistributed and localized more basolaterally, leading to reducedEx localization at the apical domain (Fig. 5 G and I). Similarly,overexpression of Crb in embryonic epithelia causes redistributionof Sdt from the subapical membrane tomore basolateral positions(27, 28). Consistent with its requirement for growth regulation, theJM domain was necessary for the redistribution of Ex on over-expression of Crbintra (Fig. 5H and J). Therefore, Crb, particularlythe JM domain, regulates Ex localization.It has been shown that Ex is down-regulated in fatmutant clones
(24, 25). The effect of Fat on Ex is dependent on D, and the re-duction of Ex levels is not seen in fat, d double-mutant clones (33).To test whether Crb regulates Ex through Fat andD, we examinedthe effect of loss of Crb onEx in a dmutant background.We foundthat Ex was still mislocalized in crb, d double-mutant cells (Fig.S4C). Moreover, D localization remained intact in crb mutantclones (Fig. S4D). Thus, our data support a model in which Crbregulates Ex membrane localization independent of Fat and D.
DiscussionOur data functionally link the growth regulatory activity of Crbwith the Hippo pathway. Several lines of evidence indicate thatCrb acts through the Hippo pathway to regulate tissue size. First,loss of crb and Crb overexpression cause overgrowth, excessproliferation, and the induction of Hippo pathway target genes.Second, loss of Crb genetically interacts with mutations in knownHippo pathway components. Third, the overgrowth phenotypesof crb and the induction of Hippo target genes depend on normalYki levels, indicating that Yki acts downstream of Crb. Fourth,Crb is required for the localization of Ex to the subapical plasmamembrane. Taken together, our data place Crb upstream of Exto regulate the activity of Yki, and thereby organ growth. Similarconclusions were recently presented by Ling et al. (34) andRobinson et al. (35) while this manuscript was under review.To date, the Hippo pathway comprises a signal transduction
cascade that includes the Fat cadherin, a transmembrane proteinthat acts upstream of several components that form a kinasecascade, which culminates in the regulation of gene expressionthrough the transcriptional coactivator Yki (11–14). However,the mutant phenotypes of fat and other components that actupstream in the pathway are generally weaker than those ofmutants in genes that act more downstream, such as hpo and wts,indicating that other unknown inputs into the Hippo pathwayexist. Here, we identify Crb as a previously undescribed upstreaminput into the Hippo pathway. Interestingly, knockdown of mersynergized with loss of crb in the pupal retina, indicating that Crband Mer cooperate to modulate Hippo signaling. This is alsosupported by our finding that Ex but not Mer localizationdepends on Crb.How does Crb regulate Hippo pathway activity? We found that
crbmutant clones had reduced levels of Ex localized to the plasmamembrane even though the transcription of ex was increased, asevidenced by the up-regulation of ex-lacZ. Thus, Crb affects Exposttranscriptionally and is required for the localization of Ex tothe apical membrane. Similarly, overexpression of Crb caused lossof Ex from the subapical membrane, although increasing ex-lacZexpression. The defects in Hippo pathway activity may thus bea consequence of the loss of Ex from the plasma membrane anddisruption of subapical Hippo signaling complexes (16, 36, 37). Inthis scenario, Crbmay act as a scaffold that is required to recruit Exto the membrane, making it available for another regulator orreceptor. Alternatively, Crb may act as a receptor and modulatethe activity of Ex. Notably, Crb is nonautonomously required forEx localization at the apical membrane. These results thus identifya cell-cell interaction-dependent mechanism that regulates theHippo pathway.Crb appears to regulate cell polarity and Hippo signaling
through different mechanisms. A version of Crb that lacks theextracellular domain, Crbintra, is sufficient to mediate the pleio-
C'C
BA
D
A' B'
E
D'ex-Z ex-Z
ex-Z ex-Z
dpp>Crb intra
dpp>Crb intraΔPBM
dpp>Crb intraΔJM
dpp>Crb intraΔJ,ΔP
Dm Crb RNKRATRGTYSPSAQEYCNPRLEMDNVLKPPPEERLIHs Crb1 SNKRATQGTYSPSRQEKEGSRVEMWNLMPPPAMERLIHs Crb2 RKRRQSEGTYSPSQQEVAGARLEMDSVLKVPPEERLIHs Crb3 REKRQTEGTYRPSSEEQVGARVPPTPNLKLPPEERLI
Consensus R KR T GTYSPS QE R EM LK PPEERLI
JM PBM
F
rela
tive
size
com
pare
d to
wild
type
intraGFP ΔPBMΔJM ΔJM/ΔPBM0
1
2
3
4
5
FL
Fig. 4. JM of the Crb intracellular domain mediates the regulation ofgrowth and the Hippo pathway. (A–D) Confocal images of third instar wingdiscs overexpressing different mutant versions of Crbintra driven by the dpp-Gal4 driver. The expression domain is marked by coexpression of GFP (green).Discs are stained for the expression of ex-lacZ (red in A–D, gray in A′–D′).Overexpression of the WT version of Crbintra and the CrbintraΔPBM mutant isable to drive growth and ex-lacZ induction, but mutation of the JM domainabolishes these effects. (E) Alignment of the intracellular domain sequence ofDrosophila Crb (Dm) with that of the three human Crb homologs (Hs Crb1–3).Conserved residues are red, and the extent of the JM and PBM is indicatedwith blue bars. (F) Quantification of the overgrowth phenotypes shown inA–D. Labeling indicates overexpressed protein. FL, Crb full-length. For discpanels, anterior is to the left and ventral is up.
Chen et al. PNAS | September 7, 2010 | vol. 107 | no. 36 | 15813
DEV
ELOPM
ENTA
LBIOLO
GY
tropic functions of Crb in regulating cell polarity (29, 38) andHippo signaling. Significantly, the effects on cell polarity andHippo signaling require different motifs: the JM is required forthe effects on Hippo signaling, whereas the PBM, which binds toSdt, is required for the effects on apical complexes. The JM isa FERM domain-binding motif that directly interacts with theFERM domain protein Yurt during photoreceptor morphogen-esis (30) and complexes with Moesin and β-Spectrin (31). Yurtand Moesin do not have Hippo-like growth defects, and thus maynot mediate the function of Crb in growth. However, Crb canbind to Ex (34), and direct effects of Crb on Ex thus appear tomediate the effects of Crb on the Hippo pathway. The fact thatthe JM but not the PBM is required for the regulation of theHippo pathway indicates that Crb has separate functions in po-
larity and growth control. Interestingly, crb mutant cells do nothave obvious morphological cell polarity defects in imaginaldiscs, although membrane levels of Patj and aPKC are reducedin crb mutant cells (39, 40). However, Fat and Mer are not lostfrom the membrane of crbmutant cells, and crb mutant cells thusdo not have a general loss of apically localized proteins. Alto-gether, our results indicate that Crb affects Hippo signalingthrough a specific mechanism rather than as a secondary con-sequence of defects in apical-basal polarity. These data suggestthat Crb is required to recruit and/or assemble a complex thatregulates Hippo signaling and contains Ex. Importantly, theobservation that crb mutant clones, which have reduced apicalmembranes (29, 39, 41), show overproliferation argues that it isnot just the size of the apical membrane that dictates growthbehavior, and thus further supports a model in which Crb playsa direct role in the generation of a growth-controlling signal.It has recently been reported that Crb affects the activity of
Notch signaling (22, 42). In contrast to our results, however, theeffects of Crb on Notch are mediated by its extracellular domain.Because Notch also regulates growth in imaginal discs, Crb mayregulate growth through multiple mechanisms.Does a Crb-like receptor act upstream of Hippo signaling in
vertebrates? Most of the known components of the Hippo sig-naling pathway in Drosophila are highly conserved in vertebrates,where they also act together in a signaling pathway (11–14, 43).Several of the vertebrateHippo pathway components act as tumorsuppressor genes and have been implicated in regulating cellproliferation and apoptosis. Vertebrates have three Crb homo-logs, Crb1–3, all of which have at least some conservation of theintracellular domain and function in apical-basal polarity (44).Importantly, Crb3 has been implicated as a tumor suppressor inselected immortal babymouse kidney epithelial cells that acquiredtumorigenicity (45). There, Crb3 expression is lost during the se-lection of tumorigenic cell lines, and reexpression of Crb3 restorescontact inhibition and cell polarity and suppresses cell migrationand metastasis. It will be interesting to test whether Crb3 actsthrough the Hippo pathway for its function as a tumor suppressor.Our data linking Crb to Hippo signaling may thus have importantimplications for the study and treatment of cancer.
MethodsMutant clones were induced using the FLP/FRT system. For generating crb, ex,mer, fat, or d mutant clones, the following alleles were flipped against cor-responding ubi-GFP and marked FRT chromosomes: crb11A22(null), exe1 (null),mer4 (null), fat422 (null), and dGC13. To generate crb mutant heads and wingsnearly entirely mutant for crb, we induced mitotic recombination by flippingcrb11A2 against Minute chromosomes using ey-FLP and ubx-FLP, respectively.The UAS-Gal4 system in combination with temperature-sensitive Gal80 wasused for overexpression with the following stocks: UAS-CrbFL (from E. Knust);UAS-Crbintra, UAS-CrbintraΔJM, CrbintraΔPBM, and CrbintraΔJM/ΔPBM (all fromK. Choi); UAS-D (from K. Irvine); UAS-crbRNAi [nos. 39177 and 39178; ViennaDrosophila RNAi Center (VDRC)]; UAS-merRNAi (no. 14228R-1; National In-stitute of Genetics Fly Stock Center); UAS-ykiRNAi (no. 40497; VDRC); and C765-Gal4, nub-Gal4, dpp-Gal4, hh-Gal4, and en-Gal4. The C765-Gal4 and nub-Gal4crosses to UAS-Crbintra were kept at 18 °C. For hh-Gal4 and en-Gal4 crosseswith different Crb constructs, crosses included Gal80ts and were kept at 18 °Cuntil they were shifted to 30 °C for 5 or 24 h, respectively, before dissection.Other crosses were kept at 25 °C. Other stocks were ex697 and ykiB5.
The quantification of the overgrowth phenotypes was done using ImageJ(National Institutes of Health), marking the GFP-positive region or the entiredisc with the “threshold” function and measuring the area with the “analyzeparticle” function. The ratio of the GFP-positive area over the total disc areawas calculated.
SEM of adult flies was done following the Hexamethyldisilazane method.Antibody stainings of imaginal discs and BrdU incorporations were done asdescribed by Hamaratoglu et al. (19). The following antibodies were used:guinea pig anti-Mer (R. Fehon, 1:4,000), rabbit-anti-Ex (A. Laughon, 1:2,000),rat anti-Fat (M.A. Simon, 1:2,000), rat-anti-Ecad (T. Uemura, 1:30), mouse-anti-Arm (Developmental Studies Hybridoma Bank, 1:30), mouse anti-BrdU(Becton Dickinson, 1:50), mouse anti-Dlg (Developmental Studies HybridomaBank, 1:300), mouse anti-β-Gal (Promega, 1:2,000), mouse anti-Crb (Develop-
C'C
A''A
C''
A' B
E'ED'D
C'''crb clones Ex
Patj
Crb
F'''F''F'F
hh>crb hh>crb
H'HG'G
crb clones
ex clones crb clones
intraΔJM
B'
Ex
Ex Ex
Ex
J'JI'I
Ex ArmEx Arm
EcadPatj Ecad
Ex
crb clones
apical basal
Fig. 5. Crb is required for Ex membrane localization. (A–C, E, and F) crb11A22
mutant clones in wing imaginal discs that are marked by the absence of GFPexpression (green). (A) crbmutant cells lose Ex from the apical membrane (A′shows an apical section) and accumulate Ex in more basal and intracellularlocations (A′′ shows a basal section). (B) Z-section through a crbmutant cloneshows the mislocalization of Ex in crb mutant cells. (C) Higher magnificationof Ex localization at crb clone borders. Ex is localized in finger-like patternsat clone borders (arrowheads), which indicates that loss of Crb leads to lossof Ex from the corresponding membrane of abutting neighboring cells.Adherens junctions are labeled by Arm localization, which is not affected(blue in C′′, gray in C′′′). (D) Ex is lost in exe1 mutant clones but not in theneighboring WT cells (arrowhead). (E and F) crb mutant cells lose Crb andPatj from the apical membranes (red in E, F, and F′′; gray in E′ and F′). As forEx, Crb and Patj are localized in finger-like patterns at clone borders(arrowheads). Cells in F are marked by E-cad staining (blue in F′′, gray in F′′′).(G) Expression of Crb in the posterior compartment by hh-Gal4 causes re-duction of Ex at the apical membrane and basal redistribution of Ex. (H)Mutation of the JM abrogates the effects of Crbintra expression on Ex lo-calization. (I and J) Cross-sections through the discs shown in G and H.Arrowheads point to clone borders.
15814 | www.pnas.org/cgi/doi/10.1073/pnas.1004060107 Chen et al.

mental Studies Hybridoma Bank, 1:50), mouse anti-Patj (H. Bellen, 1:500),and mouse anti-V5 (Invitrogen, 1:200).
ACKNOWLEDGMENTS. We thank Elisabeth Knust (Max Planck Institute ofMolecular Cell Biology, Dresden, Germany), Hugo Bellen (Baylor Collegeof Medicine, Houston, TX), Kwang-Wook Choi (Korea Advanced Institute ofScience and Technology, Daejon, Korea), Duojia Pan (The Johns HopkinsUniversity, Baltimore, MD), Richard G. Fehon (University of Chicago, Chicago,IL), Allen Laughon (University of Wisconsin, Madison, WI), Kenneth D. Irvine(Rutgers, The StateUniversity ofNewJersey, Piscataway,NJ),MichaelA. Simon
(Stanford University, Stanford, CA), Tadashi Uemura (Kyoto University, Kyoto,Japan), the BloomingtonDrosophila Stock Center, the Developmental StudiesHybridoma Bank (University of Iowa), the National Institute of Genetics FlyStock Center, and the Vienna Drosophila RNAi Center for fly stocks and anti-bodies.We also thankMarlese Pisegna for technical help andmembers of theG.H. laboratory for discussions. We thank Kenneth Dunner, Jr., for technicalhelp with the SEM analysis, which was done at M. D. Anderson core facilitiessupported by National Cancer Institute Cancer Center Core Grant CA16672.Thisworkwas supportedby aNational Institutes ofHealthgrant (toG.H.), andF.H. is a fellow of the Human Frontier Science Program.
1. Tepass U, Tanentzapf G, Ward R, Fehon R (2001) Epithelial cell polarity and celljunctions in Drosophila. Annu Rev Genet 35:747–784.
2. Dow LE, Humbert PO (2007) Polarity regulators and the control of epithelialarchitecture, cell migration, and tumorigenesis. Int Rev Cytol 262:253–302.
3. Assémat E, Bazellières E, Pallesi-Pocachard E, Le Bivic A, Massey-Harroche D (2008)Polarity complex proteins. Biochim Biophys Acta 1778:614–630.
4. Humbert PO, et al. (2008) Control of tumourigenesis by the Scribble/Dlg/Lgl polaritymodule. Oncogene 27:6888–6907.
5. Hariharan IK, Bilder D (2006) Regulation of imaginal disc growth by tumor-suppressorgenes in Drosophila. Annu Rev Genet 40:335–361.
6. Lu H, Bilder D (2005) Endocytic control of epithelial polarity and proliferation inDrosophila. Nat Cell Biol 7:1232–1239.
7. Sotillos S, Díaz-Meco MT, Caminero E, Moscat J, Campuzano S (2004) DaPKC-dependent phosphorylation of Crumbs is required for epithelial cell polarity inDrosophila. J Cell Biol 166:549–557.
8. Vaccari T, Bilder D (2005) The Drosophila tumor suppressor vps25 prevents non-autonomous overproliferation by regulating notch trafficking. Dev Cell 9:687–698.
9. Tepass U, Theres C, Knust E (1990) crumbs encodes an EGF-like protein expressed onapical membranes of Drosophila epithelial cells and required for organization ofepithelia. Cell 61:787–799.
10. Klebes A, Knust E (2000) A conserved motif in Crumbs is required for E-cadherinlocalisation and zonula adherens formation in Drosophila. Curr Biol 10:76–85.
11. Harvey K, Tapon N (2007) The Salvador-Warts-Hippo pathway—An emerging tumour-suppressor network. Nat Rev Cancer 7:182–191.
12. Reddy BV, Irvine KD (2008) The Fat and Warts signaling pathways: New insights intotheir regulation, mechanism and conservation. Development 135:2827–2838.
13. Pan D (2007) Hippo signaling in organ size control. Genes Dev 21:886–897.14. Badouel C, Garg A, McNeill H (2009) Herding Hippos: Regulating growth in flies and
man. Curr Opin Cell Biol 21:837–843.15. Willecke M, Hamaratoglu F, Sansores-Garcia L, Tao C, Halder G (2008) Boundaries of
Dachsous Cadherin activity modulate the Hippo signaling pathway to induce cellproliferation. Proc Natl Acad Sci USA 105:14897–14902.
16. Yu J, et al. (2010) Kibra functions as a tumor suppressor protein that regulates Hipposignaling in conjunction with Merlin and Expanded. Dev Cell 18:288–299.
17. Baumgartner R, Poernbacher I, Buser N, Hafen E, Stocker H (2010) The WW domainprotein Kibra acts upstream of Hippo in Drosophila. Dev Cell 18:309–316.
18. Genevet A, Wehr MC, Brain R, Thompson BJ, Tapon N (2010) Kibra is a regulator ofthe Salvador/Warts/Hippo signaling network. Dev Cell 18:300–308.
19. Hamaratoglu F, et al. (2006) The tumour-suppressor genes NF2/Merlin and Expandedact through Hippo signalling to regulate cell proliferation and apoptosis. Nat Cell Biol8:27–36.
20. Zhang L, et al. (2008) The TEAD/TEF family of transcription factor Scalloped mediatesHippo signaling in organ size control. Dev Cell 14:377–387.
21. Cho E, et al. (2006) Delineation of a Fat tumor suppressor pathway. Nat Genet 38:1142–1150.
22. Richardson EC, Pichaud F (2010) Crumbs is required to achieve proper organ sizecontrol during Drosophila head development. Development 137:641–650.
23. Udan RS, Kango-Singh M, Nolo R, Tao C, Halder G (2003) Hippo promotesproliferation arrest and apoptosis in the Salvador/Warts pathway. Nat Cell Biol 5:914–920.
24. Willecke M, et al. (2006) The fat cadherin acts through the hippo tumor-suppressorpathway to regulate tissue size. Curr Biol 16:2090–2100.
25. Silva E, Tsatskis Y, Gardano L, Tapon N, McNeill H (2006) The tumor-suppressor genefat controls tissue growth upstream of expanded in the hippo signaling pathway.Curr Biol 16:2081–2089.
26. Mao Y, et al. (2006) Dachs: An unconventional myosin that functions downstream ofFat to regulate growth, affinity and gene expression in Drosophila. Development 133:2539–2551.
27. BachmannA, SchneiderM,TheilenbergE,GraweF,KnustE (2001)Drosophila Stardust isa partner of Crumbs in the control of epithelial cell polarity. Nature 414:638–643.
28. Hong Y, Stronach B, Perrimon N, Jan LY, Jan YN (2001) Drosophila Stardust interactswith Crumbs to control polarity of epithelia but not neuroblasts. Nature 414:634–638.
29. Izaddoost S, Nam SC, Bhat MA, Bellen HJ, Choi KW (2002) Drosophila Crumbs isa positional cue in photoreceptor adherens junctions and rhabdomeres. Nature 416:178–183.
30. Laprise P, et al. (2006) The FERM protein Yurt is a negative regulatory component ofthe Crumbs complex that controls epithelial polarity and apical membrane size. DevCell 11:363–374.
31. Médina E, et al. (2002) Crumbs interacts with moesin and beta(Heavy)-spectrin in theapical membrane skeleton of Drosophila. J Cell Biol 158:941–951.
32. Tanentzapf G, Smith C, McGlade J, Tepass U (2000) Apical, lateral, and basalpolarization cues contribute to the development of the follicular epithelium duringDrosophila oogenesis. J Cell Biol 151:891–904.
33. Feng Y, Irvine KD (2007) Fat and expanded act in parallel to regulate growth throughwarts. Proc Natl Acad Sci USA 104:20362–20367.
34. Ling C, et al. (2010) The apical transmembrane protein Crumbs functions as a tumorsuppressor that regulates Hippo signaling by binding to Expanded. Proc Natl Acad SciUSA 107:10532–10537.
35. Robinson BS, Huang J, Hong Y, Moberg KH (2010) Crumbs regulates Salvador/Warts/Hippo signaling in Drosophila via the FERM-domain protein Expanded. Curr Biol 20:582–590.
36. Badouel C, et al. (2009) The FERM-domain protein Expanded regulates Hippopathway activity via direct interactions with the transcriptional activator Yorkie. DevCell 16:411–420.
37. Ho LL, Wei X, Shimizu T, Lai ZC (2010) Mob as tumor suppressor is activated at the cellmembrane to control tissue growth and organ size in Drosophila. Dev Biol 337:274–283.
38. Wodarz A, Hinz U, Engelbert M, Knust E (1995) Expression of crumbs confers apicalcharacter on plasma membrane domains of ectodermal epithelia of Drosophila. Cell82:67–76.
39. Hamaratoglu F, et al. (2009) The Hippo tumor-suppressor pathway regulates apical-domain size in parallel to tissue growth. J Cell Sci 122:2351–2359.
40. Genevet A, et al. (2009) The Hippo pathway regulates apical-domain sizeindependently of its growth-control function. J Cell Sci 122:2360–2370.
41. Pellikka M, et al. (2002) Crumbs, the Drosophila homologue of human CRB1/RP12, isessential for photoreceptor morphogenesis. Nature 416:143–149.
42. Herranz H, Stamataki E, Feiguin F, Milán M (2006) Self-refinement of Notch activitythrough the transmembrane protein Crumbs: Modulation of gamma-secretaseactivity. EMBO Rep 7:297–302.
43. Zhao B, Li L, Lei Q, Guan KL (2010) The Hippo-YAP pathway in organ size control andtumorigenesis: An updated version. Genes Dev 24:862–874.
44. Bazellieres E, Assemat E, Arsanto JP, Le Bivic A, Massey-Harroche D (2009) Crumbsproteins in epithelial morphogenesis. Front Biosci 14:2149–2169.
45. Karp CM, et al. (2008) Role of the polarity determinant crumbs in suppressingmammalian epithelial tumor progression. Cancer Res 68:4105–4115.
Chen et al. PNAS | September 7, 2010 | vol. 107 | no. 36 | 15815
DEV
ELOPM
ENTA
LBIOLO
GY