the antagonistic effect of kt and dihydro-ouabain on the
TRANSCRIPT

Jourutal of Ph1tysiology (1995), 484.3, pp. 617-628
The antagonistic effect of Kt and dihydro-ouabain on theNa+ pump current of single rat and guinea-pig cardiac cells
A. N. Hermans, H. G. Glitsch*t and F. Verdonek
Interdisciplinary Research Centre, Catholic University of Leuven, Campus Kortrijk,B-8500 Kortrijk, Belgium and *Department of Cell Physiology, Ruhr-University,
D-44 780 Bochunm, Germany
1. The antagonistic effect of extracellular potassium ions (K+) and dihydro-ouabain (DHO) on
the Na+-K+ pump current (Ip) was studied in isolated ventricular cells.
2. The mvocytes were isolated from rats and guinea-pigs, two species with differentsensitivity towards cardiac glycosides. Ip measurements were performed at 32-34 °C bymeans of whole-cell recording. The membrane potential was held at -20 mV throughout.
3. The DHO concentration ([DHO]) required for half-maximal Ip inhibition (apparent KDvalue, KD') amounted to 2-4 x 10-3 and 1P4 x 10`5 M for rat and guinea-pig myocytes,respectively, at 54 mm Kt+.
4. The data suggest one-to-one binding of DHO to the Na+-K+ pump and a smallerassociation rate constant, as well as a larger dissociation rate constant, for binding of DHOin the rat cells.
5. Ip activation by Ko+ was nearly identical in myocytes of both species and was measured tobe half-maximal at -1 mm K+. Half-maximal IP activation by K+ remained essentiallyunchanged, but Ip decreased in media containing [DHO] near the respective KD' at 5 4 mmK+.
6. The concentration-response curve of Ip inhibition by DHO was shifted to higher [DHO] athigher [K+]O. KD' increased correspondingly. The slope of the currve was unaffected.
7. lp and KD' display ed a similar dependence on [K+]O.8. KD' was larger in Na+-free than in Na+-containing media under conditions in which the
activation of Ip by K+ was nearly the same.
9. It is concluded that the antagonism between K+ and DHO, with regard to the activation ofIp, is non-competitive. A possible mechanism of the antagonism is discussed. Themechanism implies binding of K+ and DHO to different conformational states of theNa+-K+ pump which are temporarily exposed to the external face of the sarcolenmma in thepump cycle. The DHO-bound states do not participate in the generation of Ip.
The pharmacological action of cardiac glycosides isantagonized by external K+. The antagonism is due toopposite effects on the activity of the sarcolemmalNa+-K+ pump (reviews: Akera & Brody, 1978; Eisner &Smith, 1992). However, the mechanism underlying theantagonism is not completely understood. Evidence frombiochemical and pharmacological work suggests thatcardiotonic steroids bind to a site of the Na+-K+ pumpfacing the extracellular medium (reviews: Schwartz,Lindenmayer & Allen, 1975; Robinson & Flashner, 1979).This binding site represents a defined conformationalstate (probably E2 P(Na2); see Stiirmer & Apell, 1992) ofthe Na+-K+-ATPase in the pump cycle. Modulation of
cardiac glycoside binding to the Na+-K+ pump probablyimplies a change in the steady-state concentration of thepuinp conformation representing the glycoside receptor(see Schwartz et at. 1975).
Intracellular Na+ and extracellular K+ modulate thepump activity and may, therefore, alter the steady-stateconcentration of the conformational state required forglycoside binding. In fact, experimental conditionsknown to increase the internal Na+ concentration ([Na+]i)enhance both Na+-K+ pumping and glycoside binding incardiac multicellular preparations (e.g. Yamamnoto, Akera& Brody, 1979; Herzig & Mlohr, 1985). Furthermore,
t To ,hom correspondence slhould b)e addressed.
3485- 617

A. N. Hermanrs, H. G. Glitsch and F Verdonck
voltage clamp measurements (Stimers, Lobaugh, Liu,Shigeto & Lieberman, 1990) and whole-cell recording(Hainill, Alarty, Neher, Sakmann & Sigworth, 1981) of thepumpl current (In), an indicator of Na+-K+ pump activity,in isolated cardiac cells at different intracellular [Na+]idemonstrated an increase in the apparent affinity of theNa+-K+ pump towards glycosides wNith increasing [Na+]i(Stimers, Liu & Lieberman, 1991; Bielen, Glitsch &Verdonck, 1992). Corresponding experiments at variousextracellular KI+ concentrations ([K+]O) revealed a
diminished inhibition of Ip by cardiac glycosides withincreasing [K+]o and tlhus a decrease in the apparentaffinitv of the Na+-K+ pump for the drugs (Stimers et al.1991; Bielen et al. 1992). According to the latter authorsthe antagonistic effect of K+ on the inhibition of Ip bydilhydro-ouabain (DHO) in rabbit Purkinje cells is almostentirely due to a reduction of the apparent DHOassociation rate constant, w7hich is in agreement with an
earlier study on a multicellular guinea-pig ventricularpreparation (Daut, 1983).
In this paper Ne describe experiments which investigatethe antagonistic effects of K+ and DHO on Ip in isolatedrat and guinea-pig ventricular cells. Glycoside-sensitive(guinea-pig) and -insensitive (rat) species were chosen inorder to look for possible differences in K+-DHOantagonism. The measurements Mwere carried out bymeans of whole-cell recording. This technique offers
adequate control of membrane potential and ioniccomposition on both sides of the sarcolemma. The resultsof the experiments support the view that variations in[K+]0 alter the probability of the glycoside binding to a
defined conformational state of the Na+-K+ pump whichrepresents the drug receptor.
METHODSIsolation of single cellsAdult Wistar rats (-250 g) were decapitated under etheranaesthesia. Young guinea-pigs (250-300 g) were killed blcervical dislocation after being anaesthetized with chloroform.Single ventricular, cells wvere isolated from the heart by an
enzy-matic treatment dur ing a Langendorff perfusion ith Ca2+-poor solutions at 35 OC. The details of the procedure have beendlescribed earlier (Bielen, Glitsch & erdonck, 1991 a, 1992).Following the Langendorff perfusion the v-entricles were cut intopieces and gently stirr ed in a Ca2+-deficient medium. Dissociated,rodl-shaped ventricular cells wsere transferred to culture dishes(diameter, 3-6 cm) in wvhich the Ca2+ concentration of the solutionwas increased stepwise at room temperature to 1 mat for ratmyocytes and 1 8 mnlI for guinea-pig cells.
SolutionsTwo different media were used for internal perfusion via thepatch pipettes. Patch pipette solution A contained (mM): 110caesium aspartate; 40 NaOH; 10 EGTA; 40 Hepes; 5 MAgC.l12; 5glucose; 5 MgATP; 5 sodium creatine phosphate (adjusted topH 7 3 at 32-34°C with HCl; free AMg2+ -2 inm). Thecomposition of patch pipette solution B was (mm): 100 NaCI; 20CsCl; 5 EGTA; 0-15 CaC(I12; 6 AMgCI2; 40 Hepes; 5 glucose; 5
MgATP (adjusted to pH 7-3 with CsOH; free Ca2+ .10-8 M, freeMgg2+ -4 mm). High Na+ concentrations of 50 (solution A) or100 imn (solution B) were chosen in order to minimize the effectsof possible subsarcolemmal Na' depletion during strongactivation of the Na+-K+ pump (Bielen, Glitsch & Verdonck,1991 b). The results obtained with either medium were identical.Most of the experiments were carried out with internalsolution A. Solution B was only used for some cells to verify theresults obtained with solution A (data included in Figs 3, 4 and6). The standard (external) superfusion medium contained (mM):144 NaCl; 0-10-8 KCl; 015gCl2; 1 8 CaCl2; 10 Hepes; 10glucose (adjusted to pH 7 3 at 32-34°C with NaOH). Themedium also contained 2 mm BaCG2 and 5 mm NiC12 in order todiminish sarcolemmal K+ ancl Ca2+ conductances and Na+-Ca2+exchange. In Na+-free solutions choline chloride (plus 5 x 10- Matropine sulphate; adjusted to pH 7-3 with LiOH) replaced NaClin equimolar concentration. In the absence of Na+, DHO wasadded to the K+-free solution in order to check for the real zero 4level.
DrugsDihydro-ouabain (DHO; Sigma, Deisenhofen, Germany) wNasadded to the superfusion media either directly or from anaqueous stock solution (10-2 m). The final [DHO] of the mediavaried betwN-een 10-6 and 5 x 10-3 M.
Experimental procedure and whole-cell recordingA culture dish containing ventricular cells showing clear cross-striation wsas installed on the stage of an inverted microscope(IM 35; Zeiss, Oberkochen, Germany) where the wNhole-cellrecording was performed. A plastic ring was pressed down to thebottom of the dish in order to reduce its volume to - 0 3 ml. Thedish was then perfused with prewarmed (32-34 °C) superfusionmedium at a rate of about 2 ml min-'. Test solutions wereapplied close to the cell under study via a multibarrelled pipette(tip diameter, 150 ,um). Gravitational release of the media wascontrolled by a command valve unit. Solution change at the cellsurface was complete in less than 200 ms. During superfusionthe temperature in the immediate vicinity of the cell decreasedby about 1 0-1 5 'C. Whole-cell recording was carried out at amembrane potential of -20 mV throughout. The initialresistance of the patch pipettes filled with either pipette solutionvaried between 2 and 4 MIQ. The clamped membrane potentialand the corresponding membrane current were measured byimeans of an Axoclamp 2A voltage clamp amplifier (AxonInstruments, Burlingame, CA, USA) and registered on a penrecorder (Multicorder; WVatanabe, Tokyo). I, was identified asthe outward current which could be activated b) K+ and blockedby DHO. The cell surface was derived from the capacitive chargeflowing during small hyperpolarizing voltage pulses. Assuming acell capacitance of 1 ,uF cm2 the surface area of rat cells -Wasestimated to be (113 + 3) x 10-6 cm2 (n = 130), and that ofguinea-pig myocytes (125 + 3) x 10-6 cm2 (a1 = 189).
StatisticsData are presented as means + S.E.M. where possible, and S.E.M.is indicated in the figures only the size of the data points isexceeded.
RESULTSIp is activated by K+ and inhibited by DHOUnder physiological conditions 3 Na+ are extruded fromthe cell and 2 K+ are taken up during each pump cycle.
J. Physiol. 484.3618
Dihydro-oua bain,
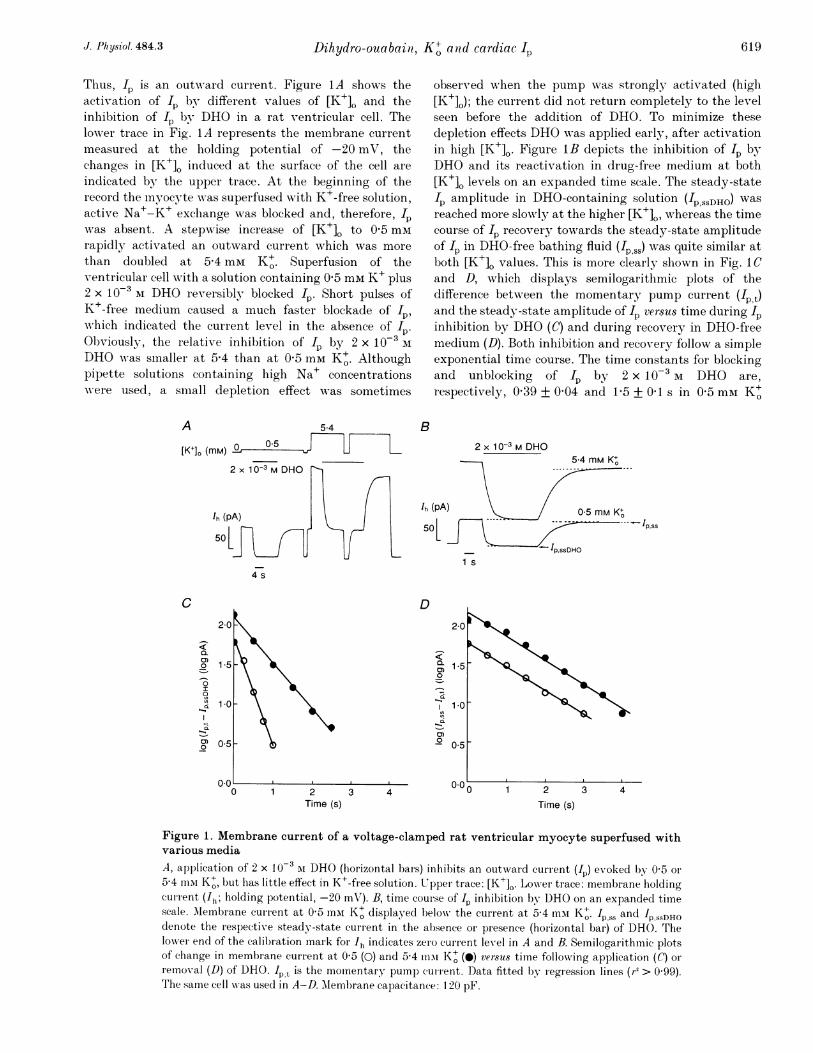
Thus, I. is an outwTard current. Figure 1 A shows theactivation of I by different values of [K+]. and theinhibition of I b)Ny DHO in a rat ventricular cell. Thelower trace in Fig. 1A represents the membrane currentmeasured at the holding potential of -20 mV, thechanges in [K+]. induced at the surface of the cell are
indicated by the upper trace. At the beginning of therecord the myocyte was superfused with K+-free solution,active Na+-K+ exchange was blocked and, therefore, Ipwas absent. A stepwise increase of [K+]. to 0 5 mmrapidly activated an outward current which was more
than doubled at 5A4 mM K+. Superfusion of theventricular cell wNith a solution containing 0 5 mm K+ plus2 x 1 0-3 M DHO reversibly blocked Ip. Short pulses ofK+-free medium caused a much faster blockade of I,
which indicated the current level in the absence of I4.Obviously, the relative inhibition of I by 2 x 10-3 AlDHO Aas smaller at 5-4 than at 0 5 mM K+. Althoughpipette solutions containing high Na+ concentrationswNere used, a smiiall depletion effect was sometimes
A 54
[K+]. (mM) 0 05
2 x 10-3M DHO [_
K+ and cardiac I 619
observed wrhen the pump was strongly activated (higlh[K+]O); the current did not return completely to the levelseen before the addition of DHO. To minimize thesedepletion effects DHO wvas applied early, after activationin high [K+]0. Figure lB depicts the inhibition of Ip byDHO and its reactivation in drug-free medium at both[K+]o levels on an expanded time scale. The steady-stateI. amplitude in DHO-containing solution ('PSSDHO) wasreached more slowly at the higher [K+]0, whereas the timecourse of Ip recovery- towards the steady-state amplitudeof Ip in DHO-free bathing fluid (Ip s) was quite similar atboth [K+], values. This is more clearly shown in Fig. 1 Cand D, which displays semilogarithmic plots of thedifference between the momentary pump current ('pt)and the steady-state amplitude of Ip versus time during Ipinhibition by DHO (C) and during recovery in DHO-freemedium (D). Both inhibition and recovery follow a simpleexponential time course. The time constants for blockingand unblocking of Ip by 2 x 10-3 M DHO are,respectively, 0 39 + 0 04 and 1P5 + 0 1 s in 0 5 mm K+
B2 x 1i03 M DHO
54 mM Ko
Ih (pA) 05 mM Ko
50 -IPss
_ 'p,ssDHO
1 s
4 s
C
0) 1a:
0oi 11-I
0-
!Zi
) 0.0
0 1 2Time (s)
D
06
-2
-1
Iluo
!Z0)
0
3 4Time (s)
Figure 1. Membrane current of a voltage-clamped rat ventricular myocyte superfused withvarious media
A, application of 2 x I0-3Oi DHO (horizontal bams) inhibits an outward current (IP) evoked by 0 5 or
5-4 mni K+, but has little effect in K+-free solution. Upper trace: [K+]0. LowTer trace: membrane holdingcurient (Ih; holding potenitial, -20 mV). B, time course of Ip inhibition by DHO on an expanded timescale. Alembrane curient at 0 5 min K+ displayed below the current at 5-4 mai K+. Is and Ip,ssDHOdenote the respective steady-state current in the absence or presence (horizontal bar) of DHO. Thelower end of the calibration mark for Ih indicates zero current level in A and B. Semilogarithmic plotsof change in membrane current at 0 5 (0) and 5 4 mnii K+ (A) versus time followNing application (C or
removal (D) of DHO. Ip t is the momentary pump currenit. Data fitted by regiession lines (r2> 0 99).The same cell was used in A-D. AMembrane capacitance: 120 pF.
J. Physiol. 484.3
A. N. Hermans, H. G. Glitsch and F Verdonck
A B
5.4 1 5
0.
[K+], (mM) 0 0*5 _)2 x10-5 M DHO °
i
Ih (pA) I_C4L
L
40 s0 10 20 30 40 50
Time (s)
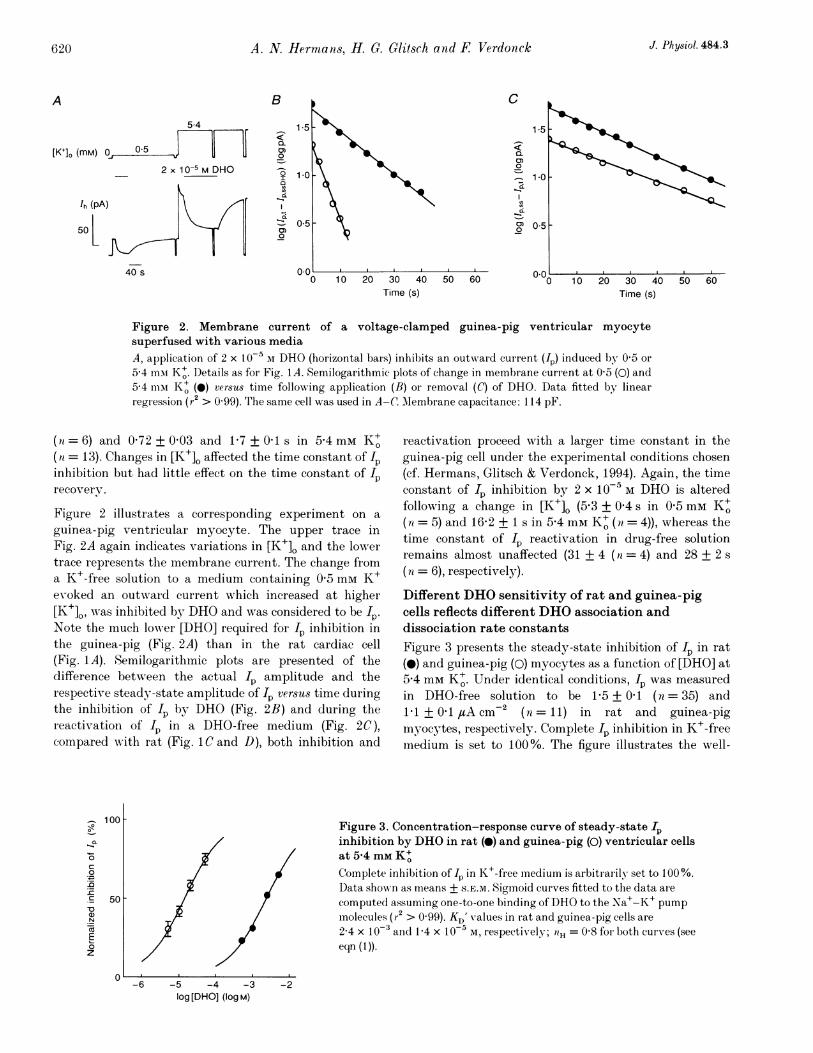
Figure 2. Membrane current of a voltage-clamped guinea-pig ventricular myocytesuperfused with various mediaA, application of 2 x l0`5 AI DHO (horizontal bars) inhibits an outward current (In) induced by 0 5 or5-4 mui K+. Details as for Fig. 1A. Semilogarithmic plots of change in membrane current at 0 5 (0) and5-4 mixt K+ (0) versus time following application (B) or removal (C of DHO. Data fitted by linearregression (r2 > 0-99). The same cell was used in A-C. Membrane capacitance: 114 pF.
(a =6) and 072 + 003 and 1P7+01ls in 5'4 mm K+(a = 13). Changes in [K+]O affected the time constant of Ipinhibition but had little effect on the time constant of IPrecovery.
Figure 2 illustrates a corresponding experiment on aguinea-pig ventricular myocyte. The upper trace inFig. 2A again indicates variations in [K+]. and the lowertrace represents the membrane current. The change froma K+-free solution to a medium containing 0 5 mM K+evoked an outward current which increased at higher[K+]O, was inhibited by DHO and was considered to be Ip.Note the much lower [DHO] required for I inhibition inthe guinea-pig (Fig. 2A) than in the rat cardiac cell(Fig. 1A). Semilogarithmic plots are presented of thedifference between the actual Ip amplitude and therespective steady-state amplitude of Ip versus time duringthe inhibition of Ip by DHO (Fig. 2B) and during thereactivation of Ip in a DHO-free medium (Fig. 2C),compared with rat (Fig. lCand D), both inhibition and
100 FI-O
0c0
z.-'D
a)
z
reactivation proceed with a larger time constant in theguinea-pig cell under the experimental conditions chosen(cf. Hermans, Glitsch & Verdonck, 1994). Again, the timeconstant of Ip inhibition by 2 x 10' M DHO is alteredfollowing a change in [K+]. (5 3 + 0G s in 0 5 mmK+i(a = 5) and 16-2 + 1 s in 5-4 mm K+ (71 = 4)), whereas thetime constant of Ip reactivation in drug-free solutionremains almost unaffected (31 + 4 (a = 4) and 28 + 2 s(n = 6), respectively).
Different DHO sensitivity of rat and guinea-pigcells reflects different DHO association anddissociation rate constantsFigure 3 presents the steady-state inhibition of Ip in rat(0) and guinea-pig (0) myocytes as a function of [DHO] at5-4 mm K+. Under identical conditions, Ip was measuredin DHO-free solution to be 1P5 + 0 1 (n= 35) and1P1 + 0-1 A cm2 (n = 11) in rat and guinea-pigmyocytes, respectively. Complete Ip inhibition in K+-freemedium is set to 100%. The figure illustrates the well-
Figure 3. Concentration-response curve of steady-state Ipinhibition by DHO in rat (0) and guinea-pig (0) ventricular cellsat 5-4 mM K+Complete inbibition of Ip in K+-free medium is arbitrarily set to 100%.Data shown as means + S.E.M. Sigmoid cuIrves fitted to the data arecomputed assuming one-to-one binding ofDHO to the Na+-K+ pumpmolecules (r2 > 0 99). KD' values in rat and guinea-pig cells are2-4 x 10-3 and 1P4 x 10-5 m, respectively; tiH = 0-8 for both curves (seeeqn (1)).
50 F
-2
C
0)Q
1--
0)
-
Iq
OL
60Time (s)
-6 -5 -4 -3log [DHO] (log M)
J. Physiol. 484.3620
I

J .ihydiro-ouabairi, K+ atrid cardiac 62
Table 1. Kinetics of Ip inhibition by DHO in rat and guinea-pig myocytes at various [K+]O:comparison between calculated and measured KD' values
k'1( 103 -1 S-1)13 0*2 (9)0 9 + 0 2 (9)0 4 + 0-03 (13)0 4 + 0 09 (8)
Rat myocytes
k'2(s-')
07 +004(12)0 8 + 0 08 (8)0 6 + 0 04 (13)07 007 (8)
Guinea-pig myocytes
k/2/kil(10-3 Al)
0 540-911 471 75
Kl' i'1
( 10-3 1 ) (t1 03 M-1 S-1)0.59 10 1 + 0 95(9)082 60±049(28)236 26+035(17)202 24+059(8)
k'2(s'l)
0 04 + 0 004 (9)0 04 + 0 003 (17)0 05 + 0 007 (13)0 03 + 0 004 (8)
kl2/k'l KD'(I 0-5 I) (10 5M)0 370651 78
1P37
0 360501 401 87
k'1, apparent association rate constant; k'2, apparent dissociation rate constant; k'2/k',, calculated KD' value;KD', apparent KD value derived frOm111easuIred steady-state IP inhibition. it values are given in par entheses.
known fact that the rat cardiac Na+-K+ pump exhibits a
much lower glycoside sensitivity than that of guinea-pigcells. Ip in guinea-pig myocytes is nearly blocked by10-4 M DHO whereas that in rat ventricular cells is onlyreduced by about 10%. The sigmoid curves fitted to thedata represent Hill equations of the form:
100 x [DHO] IHPercentage Ip inhibition = (1)
(KD/)lH + [DUO]""Æ
These were obtained by least-squares non-linear regression.The adequate Hill coefficient (nH) turned out to bebetween 0 80 and 1 2 for all relevant data sets analysedhere. The numbers suggest one-to-one binding of DHO tothe receptor conformnation of the Na+-K+ pump. Thefitting procedure yielded KD' values of 2A4 x 10-3 M DHOfor rat cells (n = 11-35) and 1 4 x 10-5 M DHO for guinea-pig myocytes (71 = 6-7). These DHO concentrationscaused half-maximal IP inhibition (KD') in the respectivecells. KD' was about 150 times larger for rat than forguinea-pig cells. If one-to-one binding is the basis of theDHO-Na+-K+ pump interaction the followingrelationship slhould apply:
k'
ki'Iwhere KD' denotes the apparent equilibrium dissociationconstant of DHO binding to the Na+-K+ pump and k'l
Figure 4. Activation of Ip by K+ in rat (0) and guinea-pig (0)ventricular cells
I,h amplitudes normalized to the respective ampitude at 5-4 mii Kt,hich was arbitrarily- set to 100%. The curXve fitted to the data obey s
the Hill equation:
Ip tmax [K+]ot@}l1p-=
(Ko.5)"It + [K+]O H
wx'here KO.5, the [lK+] for half-maximal activation is 1 miar and tH is 15
(1 = 5-13).
and k'2 signify the apparent association rate constant andthe apparent dissociation rate constant, respectively. Theapparent rate constants were derived from the bindingand unbinding of DHO in experiments similar to thosedepicted in Figs 1 and 2. The procedure has beendescribed earlier in detail (Bielen et al. 1992). Table 1
gives the kinetic parameters and calculated KD' values at5-4 mm K+ (and other [K+]. values). It is evident fromthese results that the lower glycoside sensitivity of ratcardiac cells is due to a combination of a smaller k'1 (meanratio: 0-15) and a higher k'2 value (mean ratio: 18) ifcompared with the corresponding numbers for guinea-pigcells. The calculated KD' values (according to eqn (2)) are inreasonable agreement with the values determined fromsteady-state inhibition as shown in Fig. 3 (see Table 1).This finding favours the proposed one-to-one binding.
Activation of 4p by K+ is nearly identical in rat andguinea-pig myocytesIn view; of the K+-DHO antagonism it may be expectedthat the different sensitivity to cardiac glycosides of ratand guinea-pig ventricular minocytes would beaccompanied with a corresponding difference in theactivation of Ip by K+. This expectation was checked bymeasuring Ip as a function of [K+]O in ventricular cells ofboth species under identical experimental conditions. Themeasurements were performed as shown in the first halfof the traces shown in Figs 1A and 2A. Starting with a
-
a)N
coE0
z
2 3 4 5 6 7 8[K+]. (mm)
[K+]o(11111)
0 51.05.4
10 8
621J. I'Ihysi'ol. 484.3
A. N. Herma as, H. G. Glitsch atudi F Verdonck
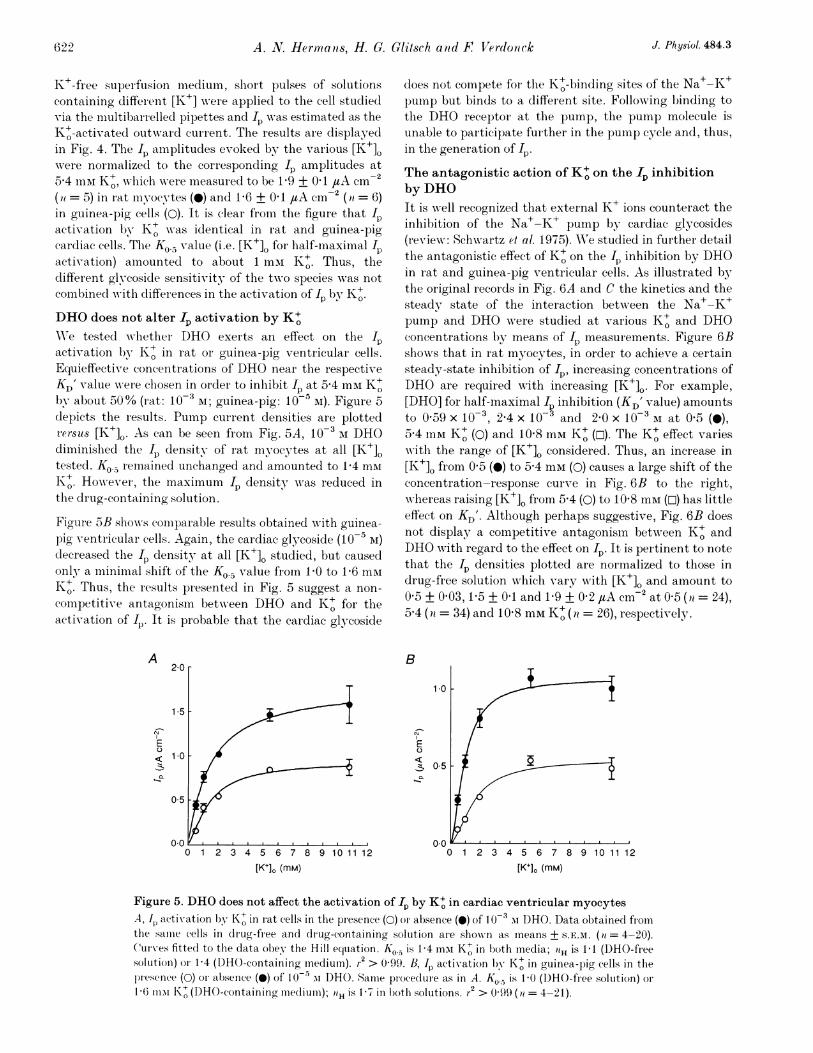
K+-free supei fusion mediumn, sbort pulses of solutionscontainino different [K+] w-ere appliecl to the cell studiedvia the multibarrelled pipettes and wp\ras estimated as theK+-activated outward current. The results are displayedin Fig. 4. Tbe I- amplitudes evoked by the various [K+].were normalized to the corresponding Ip amplitudes at5-4 mM K+, wNhich wrere measured to be 1P9 + 0 1 /tA cm-(it = 5) in rat miyocytes (0) and 1P6 + 0 1 /tA cm2 (ai = 6)in guinea-pig cells (0). It is clear from the figure that I4activation by K+ was identical in rat and guinea-pigcardiac cells. The K0.5 value (i.e. [K+]0 for half-maximal Ipactivation) amounted to about 1 mmI K+. Thus, thedifferent glycoside sensitivity of the two species was notcombinecd with differences in the activation of Ip by K+O.DHO does not alter Ip activation by K+WVe tested whether DHO exerts an effect on the I)activation by K+ in rat or guinea-pig ventricular cells.Equieffective concentrations of DHO near the respectiveKD' value wN-ere chosen in order to inlhibit IP at 54 mM K+Oby about 50% (rat: 10-3 AI; guinea-pig: 10 5 M). Figure 5depicts the results. Pump current densities are plotted2ersus [K+]o. As can be seen from Fig. 5A, 10-3 M DHOdiminished the Ip density of rat myocytes at all [K+]Otested. Ko., reemained unchanged and amounted to 1P4 mmK+. However, thie maximnum Ip density wNras reduced inthe drug-containing solution.
Figure 5B shows comparable results obtained with guinea-pig ventricular cells. Again, the cardiac glycoside (10-5 lI)decreased the Ip density at all [K+]O studied, but causedonly a minimial shift of the Ko.5 value fiom G10 to 1P6 miK+. Thus, the results presented in Fig. 5 suggest a non-competitive antagonism betw-een DHO and K+ for theactivation of I,. It is probable that the cardiac glycoside
A 2.0
1.5
E< 10
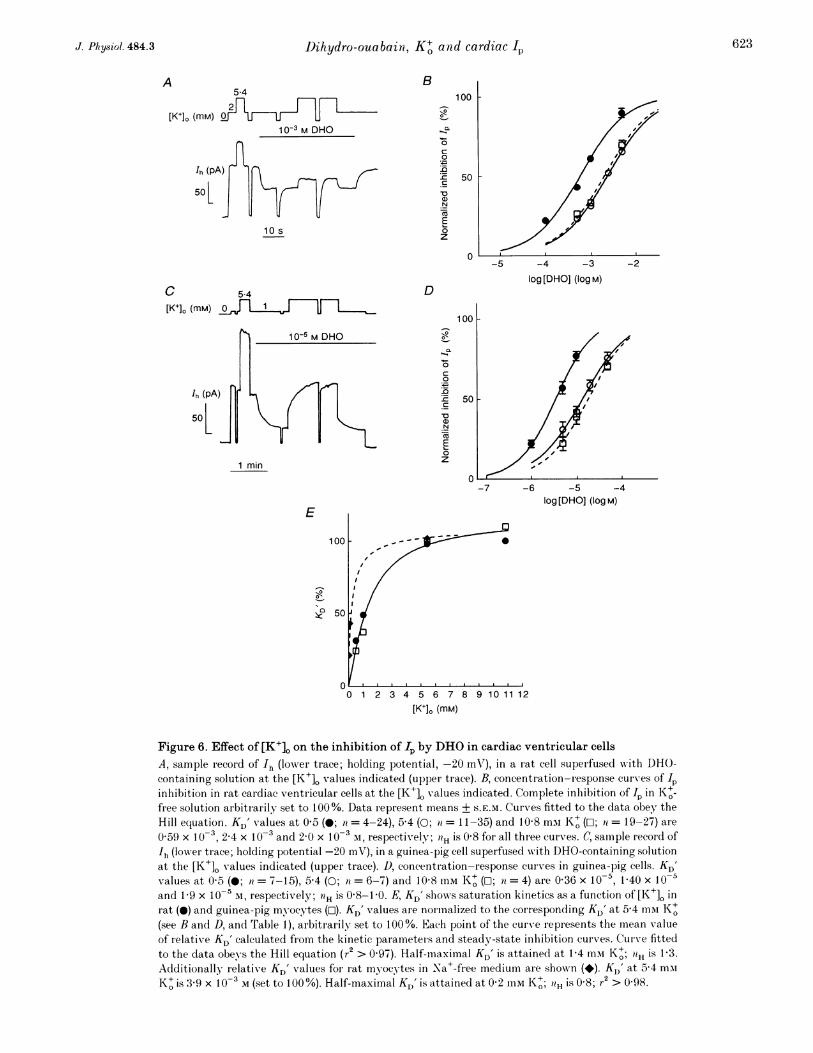
does not compete for the K+-binding sites of the Na+-K+pump but binds to a different site. Following l)inding tothe DHO receptor at the pump, the pump molecule isunable to participate further in the pump cycle and, thus,in the generation of IP.The antagonistic action of K+ on the 1p inhibitionby DHOIt is well recognized that external K+ ions counteract theinhibition of the Na+-K+ pump by cardiac glycosides(review: Schwartz et al. 1975). WVe studied in further detailthe antagonistic effect of K+ on the Ip inhibition by DHOin rat and guinea-pig ventricular cells. As illustrated bythe original records in Fig. 6A and C the kinetics and thesteady state of the interaction betwreen the Na+-K+pump and DHO were studied at various K+ and DHOconcentrations by means of IP measurements. Figure 6Bshows that in rat mnyocytes, in order to achieve a certainsteady-state inhibition of Ip, increasing concentrations ofDHO are required with increasing [K+]o. For example,[DHO] for lhalf-maximal I inhibition (KD' value) amountsto 0 59 x 103, 2A4 x 10 N and 2-0 x 10-3 M at 0 5 (0),5@4 mM K+ (0) and 10-8 mi K+ (w). The K' effect varieswith the range of [K+]. considered. Thus, an increase in[K+]o from 0 5 (0) to 5-4 mml (0) causes a large shift of theconcentration-response curve in Fig. 6B to the right,wAhereas raising [K+]. from 5-4 (0) to 10-8 mmvi (O) has littleeffect on KD'. Altlhough perhaps suggestive, Fig. 6B doesnot display a competitive antagonism between K+ andDHO with regard to the effect on Ip. It is pertinent to notethat the Ip densities plotted are normalized to those indrug-free solution which vary with [K+]O and amount to0 5 + 0 03, 1 5 + 0 1 and 1 9 + 0 2 uA cm-2 at 0 5 (n = 24),5 4 (it = 34) and 10 8 mm K+ (7l = 26), respectively.
B
1*0
E0
: 05
00
[K+], (mM)0 1 2 3 4 5 6 7 8 9 10 11 12
[K+], (mM)
Figure 5. DHO does not affect the activation of Ip by K+ in cardiac ventricular myocytesA, 1I activation by K+ in iat cells in the presence (0) oI absence (0) of 10-3 mI DHO. Data obtained fiomthe samiie cells in drug-free and drug-containing solution aIe showsn as means+ S.E.M. (a = 4-20).CuIrves fittecl to the data obey the Hill equiation. Ko. is 1-4 mI K+ in both media; 0H is 1-1 (DHO-freesolutioni) or) 1-4 (DHO-containing mnedium). r2 > 0-99. B, Ip activxation by K+ in guinea-pig cells in theplesenlce (0) or absence (O) of 10`- m DHO. Same procecluie as in A. Ko.5 is 10 (DHO-free solution) oi.16 mmi K1+ (DHO-containing meediumii); iiH is 1 7 in both solutions. r2 > 0 99 (it = 4-21).
J. Physiol. 484.3622
JDihydro-ouabain, K+ and cardiac Ip
B5.4
2
[K+]. (mM)10-3 M DHO
Ih (pA)-
501
10 s
C 5-4
[K+]o (mM)
C
0
0n
c
'D_4aN
0
z
100
50
0
D
10-5 M DHO
Ih (pA)
500
1 min
E
00-I
0.c
a
1-
0)0
-c'D
log[DHO] (10gM)
-7 -6 -5 -4log[DHO] (10gM)
0 1 2 3 4 5 6 7 8 9 10 11 12
[K+], (mM)
Figure 6. Effect of [K+]. on the inhibition of Lp by DHO in cardiac ventricular cellsA, sample record of 'h (lower trace; holding potential, -20 mV), in a rat cell superfused wvith DHO-containing solution at the [K+]. values indicated (upper trace). B, concentration-response curves of Ipinhibition in rat cardiac ventricular cells at the [K+], values indicated. Complete inhibition of fp in Ko+-free solution arbitrarily set to 100%. Data represent means + S.E.M. Curves fitted to the data obey theHill equation. KD' values at 0 5 (0; it = 4-24), 5-4 (0; ii = 11-35) and 10 8 mmI K+ (O; 71 = 19-27) are
0 59 x 10-3, 2-4 x 10-3 and 2-0 x 10-3 M, respectively; 0H is 0-8 for all three curves. C, samnple recor d ofIh (lowver trace; holding potential -20 mV), in a guinea-pig cell superfused with DHO-containing solutionat the [K+]O values indicated (upper trace). D, concentration-response curves in guinea-pig cells. KD'values at 0 5 (0; ta = 7-15), 5-4 (0; ai = 6-7) and 10-8 mM K+ (0; n = 4) are 0-36 x 10-5, 1-40 x 10-5and 1-9 x 10-5 m, respectively; '?H is 0 8-1 0. E, KD' shows saturation kinetics as a function of [K+]O inrat (@) and guinea-pig myocytes (O-1). KD' values are nor malizecl to the corresponding KD' at 5 4 miiK+m(see B and D, and Table 1), arbitrarily set to 100 %. Each point of the curve represents the mnean valueof relativxe KD' calculated from the kinetic parameteris and steady-state inhibition curves. Cur ve fittedto the data obeys the Hill equation (r2 > 0 97). Half-maximal KD' is attained at 1-4 mm K+; aH is 1-3.Additionally relative KD' values for rat myocytes in Na+-free medium are shown (*). K,)' at 5*4 mmK+ is 3-9 x 10-3 M (set to 100%). Half-maximcal KD' is attained at 0 2 mm KI+; "H is 0 8; r2 > 0-98.
A
623J. Phy8iol. 484.3
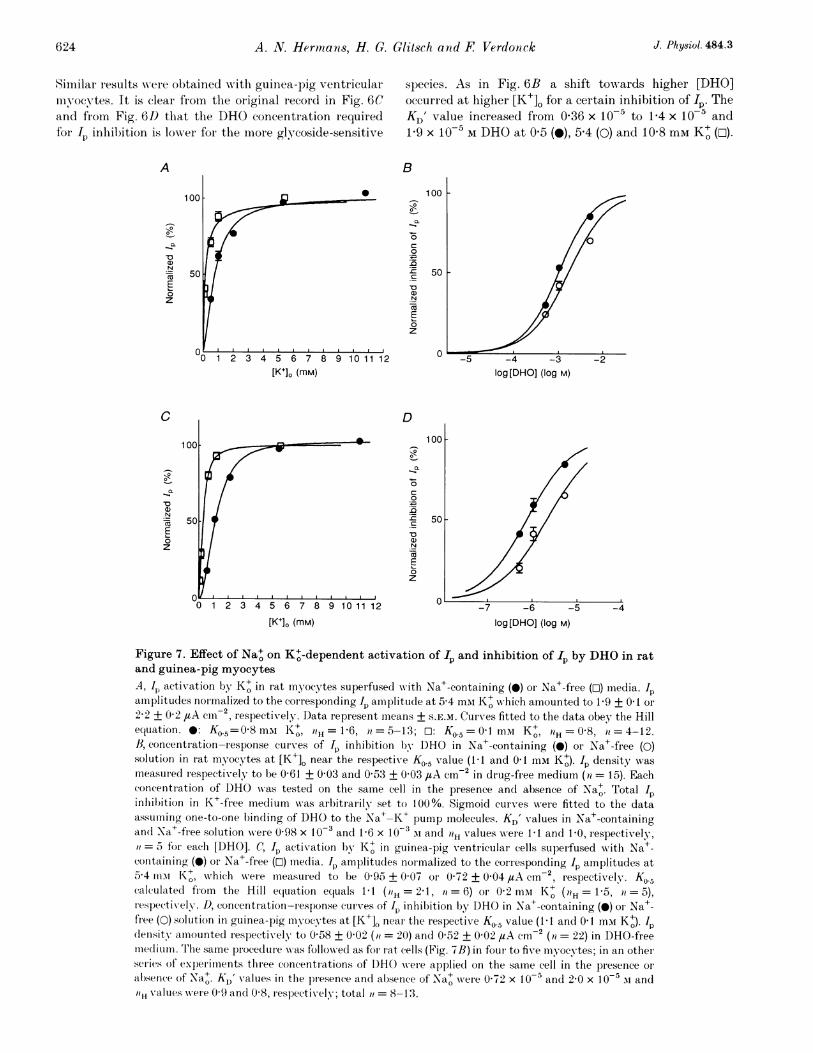
Similar results were obtained wvith guinea-pig ventricularmyocytes. It is clear from the original recor(l in Fig. 6Cand from Fig. 6D that the DHO concentration requiredfor Ip inhibition is lowrer for the more glycoside-sensitive
A
100| L) 0
-
a>N
Ca
0z
501
species. As in Fig. 6B a shift towards higher [DHO]occurred at higher [K+]L for a certain inhibition of Ip. TheKD' value increased from 0-36 x 10-5 to 1P4 x 10`5 and1.9 X 10-5 M DHO at 0 5 (0), 5-4 (0) and 10-8 mm K+ (O).
B
100
0~~~~~~0
50a)N
E0z
0
[K+]. (mM)
C
0-
a)
E0z
D
log [DHO] (log M)
100 --
0-
QVa)N
c
0
._
c
_4
E0z
50-
01 2 3 4 5 6 7 8 9 10 11 12
[K+], (mM)
Figure 7. Effect of Na+ on K+-dependent activation of 1I and inhibition of Ip by DHO in ratand guinea-pig myocytesA, Jp activation by K+ in rat myocytes superfused with Na+-containing (0) or Na+-free (o) media. 1pamiiplitudes normalized to the corresponding Ip amplitucle at 5 4 mAi K+ which amounted to 1-9 + 0O1 or
2-2 + 0 2 #tA cm-2, respectively. Data represent means + S.E.M. CuIves fitted to the data obey the Hillequation. *: K0.5=0-8 mMi Kt, 1H= 1-6, it = 5-13; T: K0.5 = l mNi Kt, 0-8,iH = 4-12.1B, concentration-response curves of II inhibition lb DHO in Na+-containing (0) or Na+-free (0)solution in rat myocytes at [K+]O near the respective KO.5 value (1 1 and 01 mMi K+). Ip density was
measur ed respectively to be 0 61 + 0 03 and 0 53 + 003 ,uA cm-2 in drug-free medium (ii = 15). Eachconcentration of DHO was tested on the same cell in the presence and absence of Na+. Total Ip
0~inhilbition in K+-free mediumn was arbitrarily set to 100%. Sigmoid curv-es were fitted to the dataassuminhg one-to-one binding of DHO to the Na+-K+ pumnp molecules. KD' values in Na+-containingan(d Na+-free solution were 0 98 x 10-3 and 1-6 x 10-3 si and iH values were 1 1 and 1-0, respectively,=5 for each [DHO]. C, Ip activation by K+ in guinea-pig ventiicular cells superfused wxith Na+-
conitaining (0) or Na+-free (O) media. Ip amplitudes normalized to the corresponding Ip amplitudes at5-4 iiisa K+, which were measur ed to be 0 95 + 0 07 or 0 72 + 0 04 #uA cml , respectively. Ko.5ccalculatecd fiom the Hill e(luation equals 1 1 (H = 21, i = 6) or 0a2 ms Kt (7H = 15, = 5),
respectWivelv. D, concentration-res)onse curves of Jp inhibition bY DHO in Naa+-containing (0) or Na+-free (0) solution in guinea-pig yocytes at [K+]O near the respective Ko.5 value (1 1 and 0 I mim K+). Ipdlensitv amounted respectively to 0 58 + 0 02 (it = 20) and 0 52 + 0 02 1tA cm2 (a = 22) in DHO-freemiiecliumn. T'lhe same proceduLre was followed as for Irat cells (Fig. 7B) in four to five myocytes; in an otherseries of experiments three concentrations of DHO were appliecl on the samne cell in the presence or
absenice of Na+. KD' values in the presence and absence of Na+ wrere 0 72 x 10-5 and 2 0 x 10-5 ui andi? \values were 0 9 and 0 8, reslectikely; total it = 8-13.
624 A. N. Hermans, H. 0. Glitsch and F Verdonck J. Physiol. 484.3
-7 -6 -5log [DHO] (log M)
-4

Dihydlro-ouabaiui, K+ and cardiac I6
Again, augmnenting [K+]. from 0 5 (0) to 5 4 miI (0)exertedI a mnuch stronger effect on the concentration-response curve than raising [K+]O from 5-4 to 10-8 mM K+(D). The I, densities to which the Ip amplitudes in Fig. 6Dare normalized were measured to be 0 33 + 0 03,11 +007, 1P13+009/LAcm.2 at 05 (ra=10), 5-4(71 = 11) and 10-8 miMi K+ (n = 5), respectively.
Figure 6E depicts a plot of normalized KD' values for IPinhibition by DHO in rat (@) and guinea-pig (o) cellsversus [K+]O. The DHO concentrations wrhich caused 50%Ip inhibition at the various [KI+]L were normalized to thecorresponding KD' value at 5-4 imMi K+. These v-aluesamounted to 2 4 x 10-3 M DHO for rat myocytes and1.4 x 10-5 M DHO for guinea-pig ventricular cells.Interestingly, the plots exhibit saturation kinetics. TheKD' values increased in a range of [K+]O (0 2-5 4 mM) inw-hich the Na+-K+ pump was increasingly activated withhiigher [K+]o, whereas KD' remained essentiallyunchanged at K' concentrations (> 5 4 inm) which causedalmost inaxiinal IP activation. A comparison with Fig. 4n- veals a similar dependence of normalized KD' valuesan; Ip densities on [K+]O. Half-naximal values of bothpararuieters were reached at about 1 umiI K+. In theabsence of Na+ the curve was markedly shifted to the left(*; rat miuvocytes). The half-maximal KD' value wasalready attained at 0-2 mai K+. Similarly, Ip activationby- K+ was shifted towards lower [K+]O (see below).
The kinetics of the Na+-K+ pump DHO interaction,studied at various K+ and DHO concentrations rangingfrom 5 x 10-4 to 5 x 10-3 M for rat myocytes and from5 x10a to i04 M for guinea-pig ventricular cells, aresummarized in Table 1. Obviously, an increase in [K+]Oclecreased the apparent association rate constant, k' , ofDHO binding but had no unequivocal effect on theapparent dissociation rate constant, k'2, in cells of bothspecies. According to these kinetic data extracellular K+ions clearly hamper the binding of DHO to the Na+-K+pump but do not strongly affect the dissociation of DHOfrom the pump molecules.
Na+ alters the antagonistic effect of K+ and DHO onIp inhibitionExtracellular Na ions are known to interfer competitivelywith the binding of K+ to the Na+-K+ pump. This Nasfirst demonstrated in erythrocytes where the Ko.5 valuefor the activation of the pump by K+ was found to benarkedly lower in Na+-free than in Na+-containingmedia (Post, MIerrit, Kinsolving & Albright, 1960).Similarly, Ip activation by K+ is facilitated in cardiacventricular mnyocytes (Nakao & Gadsby, 1989) andPurkinje cells (Bielen, Glitsch & Verdonck, 1993)superfused with Na+-free solution. Since the resultspresented so far revealed that activation of the Na+-K+pUInp by K+-binding affects the interaction between DHOand the pumimp, we thought it miglht be interesting tostudly the possiible effect of Na+ on the DHO-K+p 0 ~~~~~~~~~~~~~~~0
antagonism. Due to the increase in affinity towards K+ bythe omission of Na+, a distinct Ip amplitude can be evokedby a much lower [K+]0 in Na+-free solution. If competitiveantagonismn between K+ and DHO for a common bindingsite were to exist, DHO binding should be promoted by areduction in K+. To check this idea, inhibition of thepumnp by DHO was investigated in media of different[K+]. (and [Na+]0), but which caused similar Ip activation.
In principle the experiments w-ere carried out as illustratedin Figs 1A, B, and 2A but Na+-free, choline-containingsuperfusion mnedia wAere applied to cells as well. Figure 7Adisplays normalized IP amplitudes of rat ventricularm)yocytes as a function of [K+]. in Na+-free (O) and Na+-containing (0) solution. The Ip amplitudes recorded at thevarious [KI+]0 were normalized to the respective IPamplitude at 5 4 mMIvK+ which was measured to be1.9 + 01 ,/tA cm-2 (>l = 5) in Na+-containing mediumand 2-2 + 0-2 ,tA cm2 (it = 12) in Na+-free choline-containing solution. Half-maximnal IP activation is evokedby 0-8 or 041 inm K+ in Na+-containing oI Na+-freesolution, respectively. Figure 7B slhoNws the result of Ipinhlibition by DHO at 1.1 mAi K+ in the presence of Na+(0) and 041 mM K+ in the absence of Na+ (0). Thesedifferent K+ concentrations allowed the pump to activateto about the same extent. Ip density in the absence ofDHO was almost the same in both solutions (0-61 + 0 03and 0 53 + 0 03 ,tA cm2, respectively). In the absence ofNa+, the concentration-response curve was shifted tohigher [DHO], resulting in an increased KD' (from0 98 x 10-3 to 1.6 x 10-3 M). The slope of the curveremained almost unchanged. This figure suggests that Ipactivation by K+ binding to the Na+-K+ pumnp ratherthan [K+]0 per se is a determinant in the antagonisticaction of K+ and DHO on cardiac Ip activation.
Corresponding results were obtained with the moreglycoside-sensitive guinea-pig ventricular mnyocytes.Figure 7C displays the relative IP amplitudes as afunction of [KI+] in Na+-containing and Na+-freesolution, respectively. The IP amplitudes were normalizedto the respective Ip amplitude at 5 4 mM K+ whichamounted to 0 95 + 0 07 utA cm-2 (a = 6) in Na+-containing medium and to 0-72 + 0 04 /tA cm-2 (71 = 11)in Na-free solution. Again, IP activation by low [K+]0 isstrengtlhened in the Na+-free medium (O). Half-maximalIp activation is evoked by 1.1 or 0'2 mm K+ in Na+-containing or Na+-free solution, respectively. Figure 7Dsummarizes the effect of Na+ on DHO affinity when K+was varied 10-fold. For an almost identical Ip activation(0058 + 0-02 versus 0-52 + 0-02 #tA cml2 in the presence(0) and absence (0) of Na+) DHO affinity did not increasewAhen [K+]0 was reduced. On the contrary, as for ratmnyocytes, KD' was increased (from 0-72 x 10-5 to2 0 X 10` M).At external K+ concentrations of 0 05 and 01 umM and inthe absence of Na+, the KD' values for inhibition by DHO
J. Physicol. 484.3 625

A. N. Hermnans, H. G. Glitsch anid F Verdonck
in rat myocytes were, respectively, 21 and 43% of the KD'found at maximal pump activation wN-ithi 5-4 mi'A K+ (seeFig. 6E: *). These values fit rather well with relative KD'values of 31 and 49% at 0 5 and 1 imi K+, respectiv-ely, inthe presence of Na+ (Fig. 6E: 0). Although the [K+]0differed by a factor of ten, the relative activation of IPwas nearly the same in the Na+-free and Na+-containingmedia and, thus, the relative KD' values are also similar.
It is evident from these results that K+ influences KD' byactivation of the pump. This is true in the presence, aswiell as in the absence, of Na+. Compared with superfusionwith Na+-containing solutions, application of Na+-freesolutions results in a lower affinityT of the pumnp for DHOw-hen the pump is activated to a similar degree, althoughthere is a 10-fold low!er [K+]..
DISCUSSIONThe techinique of wN-hole-cell recording (Hainill et al. 1981)offers the opportunity to measure II in single cardiacmyocytes. The current is considered to be a reliableindicator of active Na+-K+ exchange under a variety ofexperimental conditions (for references see Bielen et al.1991 a). We used wNhole-cell recording in order toinvestigate in some detail the interaction betwNeenNa+-K+ pump activity (indicated by Ip), K+ and thecardiac glycoside DHO in cardiac ventricular cells fromntwo species with different glycoside sensitivities.
Ip is generated by Na+-K+ pump moleculesexhibiting low glycoside sensitivityThe inhibition of Ip by DHO is well described for cellsfrom both species by simple saturation kinetics,stuggesting a single population of pump molecules and aone-to-one binding of DHO to the Na+-K+ pump. This istrue for a range between about 15 and up to 95% of Ipinhibition (Figs 3 and 6). In contrast to our findings,AMogul, Rasmussen, Singer & Ten Eick (1989) suggestedthe existence of two IP components of different DHOaffinity in guinea-pig ventricular cells. The high affinity,component (KD', 50 ni'. at 5-4 mM K+) amiounted to about40% of the total Ip. Similarly, Berlin, Fielding & Ishizuka(1992) argued in favour of twAo IP components displayingdifferent sensitivities toA-ards ouabain in rat ventricularmiyocytes. The high affinity component (KD', 1/1tM at15tmvim K) contributed up to 32% of the total Ip. At5-4 miA K+ we could finid only a very small fraction of Ip ofrat mvocytes inhibited by DHO at concentrations lowNerthan 10-5 MI; 1i-6 and 10`5 Ai DHO suppressed Ip by1P9 + 0 4 (; = 15) and 4-9 + 0-8% (ti = 8), respectivelySimilarly, no consistent Ip inhibition by <2 x 10-6 Al
DHO at 5 4 mni K+ was observed in guinea-pig ventricularcells. HowN,ever, the presence of the highly ouabain-sensitive (KD 20 nm) a2-isoform of the Na+-K+-ATPaseseemns to be established in rat ventricular cells (-25% of
the total ouabain-sensitive ATPase activity; Lucchesi &Sw-eadner, 1991), but is under debate in guinea-pigventricular myocytes (in favour: Guede, Gerbi,Charlemagne & Lelievre, 1991; no evidence: Sweadner,Herrera, Amato, Moellmann, Gibbons & Repke, 1994). Ifat all functional under our experimental conditions thex2-isoformn contributed little to Ip. Thus, Ip described hereprobably reflects in both species the Na+-K+-ATPaseactivity of the cx,-isoform which showN-s a lower glycosideaffinity than a2.
Properties of Ip, in rat and guinea-pig ventricularmyocytesAs can be seen from Fig. 3, much higher DHOconcentrations are required for inhibition of 1p in rat thanin guinea-pig cells. The different glycoside sensitivity ofthe inyocytes is characterized by different association anddissociation rate constants for the interaction betweenDHO and the Na+-K+ pump (Table 1). It is notaccompanied by a different activation of 1p by K+ (Fig. 4).In our experiments the [K+]0 observed for half-maximalIP amplitudes (KO.5) agree reasonably well with Ko.5values published by other authors (rat: Berlin et al. 1992;Kinard, Liu, Liu & Stimers, 1994; guinea-pig: Nakao &GadsbY, 1989).
A hypothetical mechanism underlying theantagonistic effect ofDHO and K+ on Lp activationThe [K+]± required for half-maximal IP activation remainsunchanged in DHO-containing media, whereas Ipdecreases (Fig. 5). This non-competitive antagonismbetween DHO and K+ with regard to 1p activationstrongly suggests different binding sites for K+ and DHOat the pump molecules and may be illustrated by thefollowing hypothetical scheme of a part of the pumpcycle:
2 Na+ 2 K+
-E2-P(Na2) J- --% E2.P - E2-P(K2)
DHO DHO
k',7 2
E2-P(Na2)DHO E2 PDHO - E2 P(K2)DHO
2 Na+ 2 K+
DHO and other cardiac glycosides bind to a conformationalstate of the Na+-K+ pump which is temporarily exposedto the extracellular medium during the pump cycle. Thisconformational state seems to be E2 P(Na2) (Stiirmer &Apell, 1992). The state normally releases 2 Na+ and binds2 K+ in order to form E2P(K2) which is essential for theactive Na+-K+ exchange measured as Jp. Followingbinding of DHO to E2 P(Na2) the conformational stateE2 P(Na2)DHO is obtained wlhich releases 2 Na+ and binds2 K+ with a similar affinity to that of E2P. However
626 J. Physiol. 484.3

J. Physiol. 484.3 Dihydro-ouabaiH, K+ and cardiac Ip 6270~~~~~
E2 P(K2)DHO cannot participate in the Na+-K+ exchangethrough the pump. As a consequence Ip decreases but Ko.5for the activation of IJ by K+ remains unaltered in DHO-containing solution. If the K+ concentration of a mediumcontaining thie drug is increased, more E2*P(K2) is formedand, consequently, active Na+-K+ transport and IP areenhanced. At the same time less E2 P(Na2) is available forDHO binding and, therefore, k'l decreases, whereas i'2 islittle affected (Table 1). The rate constants are consideredto be rate limiting in the lower branch of the scheme. Anew equilibrium between E2 P(Na2) and E2 P(Na2)DHO isestablished which is characterized by a new KD' value(Fig. 6B and D). According to the scheme, Ip is expected toincrease in two phases upon an increase in [K+]concentration in DHO-containing solution. A rapidcomponent should reflect the higher [E2-P(K2)] at higher[K+]O. It should be followed by a slower phase due to theredistribution between E2 P(Na2)DHO and E2 P(Na2)which also will finally produce an increase in [E2 P(K2)]and thereby in Ip. In fact, both components of Ip increasecan be observed in the records from guinea-pig myocytes(Fig. 6C), but are not as well resolved in the Ipmeasurements on rat cells (Fig. 6A), probably because ofthe higher speed of the 'slow' Ip increase in thesemyocytes under the experimental conditions chosen (cf.Hermans et al. 1994). The scheme proposed also illustrateswhy the KD' value of DHO binding to the Na+-K+ pumpand the activation of Ip display a similar dependence on[K+]0. KD' increases at higher [K+]O because k', declines,whereas k'2 remains nearly unchanged (Table 1). The shiftof KD' directly reflects the occupancy of E2 P by K+ sincethe slope of the concentration-response curves in Fig. 6Band D is unaltered (nH 1) at the various K+concentrations. A simnilar relationship betwveen ouabaiirbinding, activation of the Na+-K+ pump and K+ hadalreadv been observed by Baker & Willis (1970) on HeLaand Girardi (cardiac) cells. Figure 7A-D emphasizes thatpump activation by K+ binding is the important factor forthe value of KD'. At a 10-fold lower [K+]O but withmaintainance of the same Ip in the absence of Na+,affinity of the pump for DHO was decreased. Thisobservation can be easily explained by state-dependentbinding of DHO. \VNhen wve assume that the Na+-boundconformational state E2 P(Na2) favours binding of DHO,the omission of Na+ will promote release of Na+,(lecreasing [E2 P(Na2)] and the probability that DHObinds to the pump mnolecule.
Concluding noteIn conclusion the experiments reported above show thatthe antagonistic effect of DHO and K+ on the IPamplitude of rat and guinea-pig cardiac myocytes is non-competitive in nature. DHO and K+ bind to differentconformational states of the Na+-K+ pump, which aretransientl- exposed to the external side of the sarcolemma.
Upon K+ binding the state is remnoved from the extra-cellular side and, therefore, becomes unaccessible forDHO. The E2 P(K2) state participates in the generation ofIp. The DHO-bouncl state E2 P(Na2)DHO binds K+ as theDHO-free state but remains at the external side of the cellmembrane and does not contribute to IP.
AKERA, T. & BRODY, T. I. (1978). The role of Na+ K+-ATPase in theinotropic action of cligitalis. Pharmnacological Reviews 29, 187-220.
BAKER, 1P. F. & \VILLIS, J. S. (1970). Potassium ions ancl the binclingof cardiac glycosicles to mammalian cells. Nature 226, 521-523.
BERLIN, J. B., FIEI,DIN(o, A. J. & ISHIZUKA, N. (1992). Identificationof low and hiigh affinity ouabain-sensitive Na pump current involtage-clamped rat cardiac mniocytes. Aii nals of the New YorkAcademty of Sciences 671, 440-442.
BIEILEN, F. VT., GLITSCH, H. G. & VERDONCK, F. (1991 a). Dependenceof Na+ pump current on external monovalent cations andmemhrane potential in ral)bit cardiac u'trlkinje cells. Journial ofPhlysiology 442, 169-189.
BIELEN, F. V., GLITSCH, H. G. & VERDONCK, F. (1991 b). Chaniges ofthe subsarcolemmal Na' concentration in internally perfusedcardliac cells. Biochimaica et Biophysica Acta 1065, 269-271.
BIELEN, F. VT., GLITSCH, H. G. & VERDONCK, F. (1992). The kineticsof the inhibition by- dihNdroouabain of the sodium pump currentin single rabbit carcliac Purkinje cells. N\au yn -,Schmniedeberg sA rchives of Pharm7acology 345, 100-107.
BIELEN, F. V., GLITSCH, H. G. & VERDONCK, F. (1993). Na' pumpcurrent-voltage relationships of rahbit cardiac Purkinje cells inNa+-free solution. Journal of Physiology 465, 699-714.
DAUT, J. (1983). Inhihition of the sodiuim pump in guinea-pigventricular muscle bh dihlvdroouabain: effects of externalpotassium and sodium. Journal of Physiology 339, 643-662.
EISNER, D. A. & SMITH, T. WV. (1992). TIhle Na-K pump and itseffectors in cardiac muscle. In The Heart and CardiovasclltarSystemii, 2nd edn, vol. 1, ecl. FOZZARD, H. A., HABER, E., JENNINGS,R. B., KATZ, A. Al. & AMORCGAN, H. E., pp. 863-902. Raven Press,New York.
G,UEDE, F. G., GERBI, A., CHARLEIMAG1NE, 1). & LELIEVRE, L. G.(1991.) Two functional Na+/K+-ATPase isoformns in the leftventricle of guinea-pig heart. European Journial of Biochemtistry196,129-134.
HA,MILL, 0. P., MIARTY, A., NEHER, E., SAK.MANN, B. & SIGWORTH,F. J. (1981). Improved p)atch-clamp techniques for high-resolutioncuirrent recording friom cells ancl cell-filee membrane p1atches.Pfliigers Archiv 391, 85-100.
HERNIANS, A. N., GLITSCH, H. G. & VERDONCK, F. (1994). The effectof carcliac glvrcosides on the Na+ pump current-voltagerelationship of isolated lat and guinea-pig heart cells. Journal ofPhysiology, 481, 279-291.
HERZIG, S. & MOHR, K. (1985). Sodliulml load and higlh affinityoual)ain binding in rat and guinea-pig carlciac tissue. BritishJournial of Pharmaacology 84, 685-688.
KINARD, T. A., LIu, X.N-., LIU, S. & STINIERS,J. R. (1994). Effect ofNa Pip on KO activation of the Na-K PI1)P in adult rat camdiacmnvocvtes. Americani Journal of Physiology 266, C37-41.
LUCCHESI, P. A. & SWEADINER, K. J. (1991). Postnatal clhanges inNa,K9-ATPase isoforml expression in mat cardiac ventricle. Journalaof Biological fhem7istry 266, 9327/-9331 .

A. N. Hermans, H. G. Glitsch and F Verdonck
OIoGUL, D. J., RASMUSSEN, H. H., SINGER, D. H. & TEN EICK, R. E.(1989). Inhibition of Na-K pump current in guinea pig ventricularmyocytes by dihyclroouabain occurs at high- and low-affinitysites. Circulationi Research 64,1063-1069.
NAKAO, Al. & GADSBY, D. C. (1989). [Na] and [K] dependence of theNa/K pump current-voltage relationship in guinea pigventricular m) ocytes. Journial of General Physiology 94, 539-565.
POST, R. L., M\ERRIT, C. R., KINSOLVING, C. R. & ALBRIGHT, C. D.(1960). Mlembrane adenosine triphosplhatase as a participant in theactive transport of sodium and potassium in the humanerythrocyte. Journal of Biological Chemistry 235, 1796-1802.
ROBINSON, J. D. & FLASHNER, M. S. (1979). The (Na+ + K )-activated ATPase. Biochimica et Biophysica Acta 549,145-176.
SCHWARTZ, A., LINDENMAYER, G. E. & ALLEN, J. C. (1975). Thesodium-potassium adenosine triphosphatase: Pharmacological,physiological and biochemical aspects. Pharmacological Reviews27, 3-134.
STIMERS, J. R., LOBAUGH, L. A., Liu, S., SHIGETO, N. & LIEBERMAN,Al. (1990). Intracellular sodium affects ouabain interaction withthe Na/K pump in cultured chick cardiac myocytes. Journal ofGenieral Physiology 95, 77-95.
STIMERS, J. R., Liu, S. & LIEBERMAN, M. (1991). Apparent affinity ofthe Na/K pump for ouabain in cultured chick cardiac myocytes.Journal of Genieral Physiology 98, 815-833.
ST{RMER, WV. & APELL, H.-J. (1992). Fluorescence study on cardiacglycoside binding to the Na,K-pump. FEBS Letters 300, 1-4.
SWEADNER, K. J., HERRERA, V. L. M., AMATO, S., MOELLMANN, A.,GIBBONS, D. K. & REPKE, K. R. H. (1994). Immunologicidentification of Na+,K+-ATPase isoforms in myocardium.Circulation Research 74, 669-678.
YAMAMOTO, S., AKERA, T. & BRODY, T. M. (1979). Sodium influx rateand ouabain-sensitive rubidium uptake in isolated guinea pigatrium. Biochimica et Biophysica Acta 555, 270-284.
AcknowledgementsThe authors wish to thank Professors A. Pappano and WV.Schwarz for stimulating comments on an earlier draft of thepaper.
Received 14 June 1994; accepted 11 October 1994.
628 J. Physiol. 484.3