taasr1 , a transcription factor gene in wheat, confers drought stress tolerance in...
TRANSCRIPT

TaASR1, a transcription factor gene in wheat, confersdrought stress tolerance in transgenic tobacco
WEI HU*, CHAO HUANG*, XIAOMIN DENG, SHIYI ZHOU, LIHONG CHEN, YIN LI, CHENG WANG, ZHANBING MA,QIANQIAN YUAN, YAN WANG, RUI CAI, XIAOYU LIANG, GUANGXIAO YANG & GUANGYUAN HE
The Genetic Engineering International Cooperation Base of Chinese Ministry of Science and Technology, Chinese NationalCenter of Plant Gene Research (Wuhan) HUST Part, Key Laboratory of Molecular Biophysics of Chinese Ministry ofEducation, College of Life Science and Technology, Huazhong University of Science & Technology (HUST), Wuhan 430074,China
ABSTRACT
Abscisic acid (ABA)-, stress-, and ripening-induced (ASR)proteins are reported to be involved in abiotic stresses.However, it is not known whether ASR genes confer droughtstress tolerance by utilizing the antioxidant system. In thisstudy, a wheat ASR gene, TaASR1, was cloned and character-ized. TaASR1 transcripts increased after treatments withPEG6000, ABA and H2O2. Overexpression of TaASR1 intobacco resulted in increased drought/osmotic tolerance,which was demonstrated that transgenic lines had lessermalondialdehyde (MDA), ion leakage (IL) and reactiveoxygen species (ROS), but higher relative water content(RWC) and superoxide dismutase (SOD) and catalase (CAT)activities than wild type (WT) under drought stress. Overex-pression of TaASR1 in tobacco also enhanced the expressionof ROS-related and stress-responsive genes under osmoticstress. In addition, transgenic lines exhibited improved toler-ance to oxidative stress by retaining more effective antioxi-dant system. Finally, TaASR1 was localized in the cell nucleusand functioned as a transcriptional activator. Taken together,our results showed that TaASR1 functions as a positive factorunder drought/osmotic stress, involved in the regulation ofROS homeostasis by activating antioxidant system and tran-scription of stress-associated genes.
Key-words: abscisic acid-, stress-, and ripening-induced(ASR) protein; antioxidant system; reactive oxygen species(ROS); stress-responsive genes.
INTRODUCTION
Plants are frequently challenged by various harsh environ-mental factors, among which drought stress is adversebecause it limits plant growth, development and crop produc-tivity (Umezawa et al. 2006). To combat such environmentalstress factors, plants deploy a variety of complex mechanismsfor their protection. The major response of plants to droughtstress include perception of stress signals and their transduc-tion that activate various stress-related genes and synthesis
of proteins with diverse functions resulting in physiologicaland metabolic responses (Zhu 2002; Seki et al. 2003;Shinozaki, Yamaguchi-Shinozaki & Seki 2003; Hu et al.2006). Well-characterized proteins involved in drought stressinclude molecular chaperons, osmotic adjustment proteins(Tamura et al. 2003), ion channels (Ward & Schroeder 1994),transporters (Klein et al. 2004) and antioxidation or detoxi-fication proteins (Bartels 2001). Expression of these func-tional proteins is largely regulated by specific transcriptionfactors (Singh, Foley & Onate-Sanchez 2002; Zhu 2002; Huet al. 2006).
Abscisic acid (ABA)-, stress-, and ripening-induced (ASR)proteins have low molecular weight and are heat-stable andhighly hydrophilic observed specifically in plants. ASR geneswere first reported in cultivated tomato (Iusem et al. 1993)and at least 30 ASR genes were identified in various speciesof dicotyledonous and monocotyledonous plants (Carrari,Fernie & Iusem 2004; Frankel et al. 2006; Saumonneau et al.2012) with no evidence for the gene in Arabidopsis (Carrariet al. 2004). Although the precise physiological function ofASRs remains unknown, they are presumed to have func-tional duality in plants. Firstly, ASRs are classified to group 7in the late embryogenesis abundant (LEA) protein familywith high hydrophilicity, which correlates to the high glycinecontent for direct plant protection (Garay-Arroyo et al. 2000;Battaglia et al. 2008; Saumonneau et al. 2012). Secondly,ASRs have been suggested to be transcriptional regulatorsbecause of their ability to bind DNA in a sequence-specificand Zn2+-dependent manner during the transition from adisordered to an ordered state (Cakir et al. 2003; Kalifa et al.2004a; Goldgur et al. 2007; Maskin et al. 2007; Shkolnik &Bar-Zvi 2008). Finally, the preferential subcellular localiza-tion of the ASRs is not clear-cut between the cytoplasm andthe nucleus (Cakir et al. 2003; Wang et al. 2003; Kalifa et al.2004a; Urtasun et al. 2010; Chen et al. 2011). As suggestedinitially by Maskin et al. (2001), ASR proteins may act asdownstream components of a common signal transductionpathway involved in plant cell response to environmentalfactors.
ASR proteins are reported to be involved in plant devel-opment, senescence and fruit ripening (Huang, Lin & Wang2000; Wang et al. 2003; Maskin et al. 2007; Chen et al. 2011).Evidence also indicates that a large number of ASR genesrespond to ABA and abiotic stresses such as water deficit,
Correspondence: G. Yang or G. He, Fax: +0086 27 87792272; e-mail:[email protected]; [email protected]
*These authors contributed equally to this work.
Plant, Cell and Environment (2013) doi: 10.1111/pce.12074
bs_bs_banner
© 2013 Blackwell Publishing Ltd 1

salt, cold, aluminium and limited light (Schneider, Salamini &Gebhardt 1997; Maskin et al. 2001; Hong et al. 2002; Jeanneauet al. 2002; Cakir et al. 2003; Kalifa et al. 2004a,b; Yang et al.2005; Saumonneau et al. 2008, 2012; Shkolnik & Bar-Zvi2008; Kim et al. 2009; Liu et al. 2010; Hsu et al. 2011; Arenhartet al. 2013; Kim, Kim & Yoon 2012). Overexpression of ASRgenes from lily, tomato, rice and plantain conferred toleranceto abiotic stresses (Kalifa et al. 2004b; Yang et al. 2005; Kimet al. 2009; Liu et al. 2010; Hsu et al. 2011).The responsivenessof ASR genes to distinct environmental cues such as drought,cold and osmotic stress was studied mainly by RNA analysis(Saumonneau et al. 2012). Interestingly, a recent studyshowed that rice ASR protein, OsASR1, could function as aneffective reactive oxygen species (ROS) scavenger and itsexpression in yeast cells enhanced acquired tolerance ofROS-induced oxidative stress through induction of variouscell rescue proteins (Kim et al. 2012). However, whetherASRs confer drought stress tolerance by regulating ROShomeostasis through the antioxidant system is yet to beelucidated in plants.
As an international staple crop, wheat production is con-strained by multiple environmental stresses, such as drought,salinity and extreme temperatures. Therefore, understandingthe molecular mechanisms of abiotic stress responses is nec-essary for genetic improvement of stress resistance in wheat.Although ASR genes respond to various stresses, their exactroles in abiotic stresses tolerance are not well known in wheat.In this study, we characterized the first wheat ASR gene,TaASR1, which is an ortholog of OsASR5 (Os11g06720)(Philippe et al. 2010).Our results showed that TaASR1 confersdrought/osmotic stress tolerance by activating the antioxidantsystem and regulating the expression of stress-responsive andantioxidant defence-associated genes.
MATERIALS AND METHODS
Plant materials and treatments
Wheat (Triticum aestivum L. cv. Chinese Spring) seeds weresurface-sterilized with 75% (v/v) ethanol for 2 min and 1%(v/v) mercury chloride for 10 min and washed three times insterile water. Sterilized seeds were germinated on sterilewater, grown in Murashige and Skoog (MS) liquid mediumand cultured in growth chambers (16 h light/8 h dark cyclewith a light intensity of 200 mmol m-2 s-1 at 25 °C). ForPEG6000 treatment, 10-day-old seedlings were transferredinto Petri dishes containing 20% PEG6000 solution and incu-bated under light for 24 h. For treatment with signalling mol-ecules, 10-day-old seedlings were sprayed with 100 mm ABA,100 mm methyl jasmonate (MeJA), 2 mM salicylic acid (SA),50 mm auxin and 10 mM H2O2 each and plants were incu-bated under light for 24 h. For inhibitor or scavenger treat-ment, the plants were pretreated with 1 mM tungstate and100 mm fluridone for 6 h, respectively, and then exposed to20% PEG6000 for 6 h, respectively. The plants were pre-treated with 5 mM dimethyl thiourea (DMTU) and 10 mMimidazole for 6 h, respectively, and then exposed to 20%PEG6000 for 6 h, respectively. The treatment of tungstate,
fluridone, DMTU or imidazole alone was also used as con-trols in the experiment. In all these treatments wheat seed-lings with similar state of growth were used and untreatedwheat seedlings were used as controls. For differentialexpression analysis, organs including roots, stems and leaveswere cut from sterile seedlings while pistils, stamens andlemmas were obtained from wheat plants in the field.Samples from treated or control plants were frozen in liquidnitrogen and stored at -80 °C until extraction of total RNAand qRT-PCR assay.
Cloning and bioinformatics analysis of TaASR1
About 16 espressed sequence tag (EST) sequences belongingto the ASR gene family were acquired from the Dana-FarberCancer Institute (DFCI) wheat gene index database (http://compbio.dfci.harvard.edu/cgi-bin/tgi/gimain.pl?gudb=wheat).These ESTs were divided into seven groups based onsequence similarity with each other and BLASTX analysiswith other ASRs (Supporting InformationTable S1).Since theESTs from group 1 did not show a clear 3′-end, rapid amplifi-cation of cDNA ends (RACE) protocol was employed todetermine the 3′ end of the ASR gene. Amplification wasperformed using the SMART RACE cDNA amplification kit(Clontech, Mountain View, CA, USA) with primer P1 (Sup-porting Information Table S2) and double-stranded mixedcDNA template obtained from leaves of wheat seedlingstreated with 200 mM NaCl, 20% PEG6000 or cold (4 °C) for2 h. The full-length cDNA sequence was identified usingDNAMAN software and amplified from wheat Poly (A)+
mRNA by PCR using primer P2 (Supporting InformationTable S2). All amplified products were cloned into thepMD18-T vector (TaKaRa, Kyoto, Japan) and sequenced onan ABI PRISM310 Genetic Analyzer (Perkin Elmer AppliedBiosystems, Foster City, CA, USA) using the BigDye Termi-nation Cycle Sequencing Ready Reaction Kit (Perkin Elmer).All sequences were analysed using BLAST (http://ncbi.nlm.nih.gov/blast) in the NCBI database.
RT-PCR
RT-PCR was used to determine the expression of specificESTs in the ASR gene family after treating wheat seedlingswith PEG and the expression of TaASR1 in the transgenictobacco lines. Primers (P4, P5, P20, P22-25; Supporting Infor-mation Table S2) used in RT-PCR had high specificity asdetermined by agarose gel electrophoresis and was also con-firmed by sequencing PCR products amplified by all primerpairs. All reactions were performed using TaKaRa DNApolymerase for 30 cycles. Expression levels of target geneswere normalized using TaActin or NtUbiquitin as internalcontrols.
qRT-PCR
Expression of TaASR1 in wheat tissues and leaves aftervarious treatments and stress-associated genes in tobaccowere analysed by qRT-PCR using the fluorescent DNA
2 W. Hu et al.
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

intercalating dye SYBRGreen (ToYoBo, Osaka, Japan) on aMJ research opticon 2 detection system. About 200 ngPoly(A)+ mRNA from each treatment was converted intocDNA using AMV Reverse Transcriptase (Promega,Madison, WI, USA) at 42 °C in a 20 mL reaction volume thatsubsequently served as the template for qRT-PCR. Primers(P3-P15, P26-28; Supporting Information Table S2) used inqRT-PCR were designed based on amino acid sequence afterexcluding the highly conserved protein domains. Theseprimers had high efficiency and specificity, which were deter-mined by the melting curve analysis using the Opticonmonitor 2 qRT-PCR software and agarose gel electrophore-sis. PCR products amplified by the primer pairs weresequenced to confirm specificity. In all experiments, appro-priate negative controls without template were included todetect primer dimers and/or any possible contamination.Prior to the experiments, qRT-PCR was performed on aseries of template and primer dilutions to determine theoptimal template and primer concentrations for maximumamplification of target genes. Amplification efficiencies forthe internal control and target genes were between 0.9 and1.1. Each sample had four replications and the data wereanalysed using the Opticon monitor 2 qRT-PCR software.TaActin or NtUbiquitin was used as internal controls thatserved as a benchmark for normalization. mRNA fold differ-ence was calculated relative to the calibrator [samples withdistilled water treatment or wild-type (WT) samples undernormal conditions] and relative expression level of genes wascalculated using the 2-DDCt formula (Livak & Schmittgen2001).
Subcellular localization of TaASR1 protein
The complete coding sequence of TaASR1 was amplified byPCR using primer (P16; Supporting Information Table S2)containing NcoI/SpeI restriction sites. After sequence confir-mation, the PCR product was digested with NcoI/SpeI andcloned into the pCAMBIA1304 vector so that the gene isunder control of Cauliflower mosaic virus 35S (CaMV 35S)promoter to create a fusion construct. Recombinant plasmidspCAMBIA1304-TaASR1-GFP and pCAMBIA1304-GFP(used as control) were separately transferred into onion epi-dermal cells using a gene gun (PDS-1000; BIO-RAD, Her-cules, CA, USA) according to the instruction manual.Transformed cells were examined by laser-scanning confocalmicroscopy (LSM510: Karl Zeiss, Jena, Germany) after incu-bation at 25 °C for 24 h on MS medium. For the subcellularlocalization of TaASR1 in transgenic lines, root samples weresliced with razor blades and mounted in water between glassslide and cover slip. Fluorescence of the GFP fusion proteinswas observed with scanning microscopy.
Analysis of transcriptional activationin yeast cells
Transcription activation assay was performed in the yeaststrain AH109 with LacZ and His reporter genes. The com-plete coding sequence of TaASR1, N-terminal (TaASR1-N)
and C-terminal (TaASR1-C) fragments were amplified byPCR using primers P17, P18 and P19 (Supporting Informa-tion Table S2), respectively. The PCR products were clonedinto EcoRI/BamHI sites in the pGBKT7 vector to createpGBKT7-TaASR1, pGBKT7-TaASR1-N and pGBKT7-TaASR1-C. The three constructs along with the negativecontrol pGBKT7 plasmid were used to transform AH109yeast strain according to the manufacturer’s protocol (Clon-tech, Palo Alto, CA, USA). Transformed strains were con-firmed by PCR and then plated on SD/Trp- or SD/His-plates.Transcription activation was evaluated according to thegrowth status of yeast cells after incubating plates for 3 d with5-bromo-4-chloro-3-indolyl-b-D-galacto- pyranoside (X-gal).
Plant transformation and generation oftransgenic plants
The recombinant plasmids pCAMBIA1304-TaASR1-GFPunder the control of the CaMV 35S promoter was introducedinto Agrobacterium tumefaciens strain LBA4404.Transforma-tion of tobacco was performed using an Agrobacterium-mediated leaf disc transformation, as described by Horschet al. (1985).Seeds from the transgenic plants were selected onMS medium containing 40 mg L-1 of hygromycin. Thehygromycin-resistant T1 seedlings were confirmed by PCRanalysis using primers to amplify TaASR1 (P20; SupportingInformation Table S2) and GFP (P21; Supporting Informa-tion Table S2) genes. Three independent transgenic T2 lineseedlings (OE2, OE5 and OE12) that almost all survived onMS medium containing 40 mg L-1 of hygromycin were used inthe experiments. Expression of TaASR1 in the three inde-pendent T2 lines was investigated by semi-quantitativeRT-PCR analysis using primers P20 and P5 (Supporting Infor-mationTable S2) and using NtUbiquitin as an internal control.
Stress tolerance of the WT, vector control (VC)and transgenic lines
For drought stress tolerance assay, the WT,VC and transgeniclines were cultured on MS medium under a 16 h light/8 hdark cycle at 25 °C for 1 week. These plants were then trans-planted in containers filled with a mixture of soil and sand(3:1) and were watered regularly for 2 weeks.Three-week-oldplants similar in growth state were subjected to water with-holding for 30 d. Thirty seedlings from each line or controlwere used to detect survival rate after withholding water for30 days. After withholding water for 15 or 30 d, leaves werecollected from plants to measure relative water content(RWC), ion leakage (IL), contents of malonaldehyde(MDA), H2O2 and activities of superoxide dismutase (SOD;EC 1.15.1.1), catalase (CAT; EC 1.11.1.6) and peroxidase(POD; EC 1.11.1.7). For osmotic stress tolerance assay, earlyseedlings from the WT,VC and transgenic lines were culturedin MS medium under a 16 h light/8 h dark cycle at 25 °C for1 week. Seedlings were then transplanted to MS with orwithout 150 or 300 mM mannitol for 1 week.Whole seedlingswere used in the physiological experiments to measure rootlength, ROS accumulation, and SOD and CAT activities. One
The role of TaASR1 in drought stress 3
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

hundred and fifty surface-sterilized seeds from each trans-genic line or control were sown on MS with or without 150 or300 mM mannitol for 8 d to detect germination rate. Foroxidative stress assay, WT, VC and transgenic lines were cul-tured in MS medium under a 16 h light/8 h dark cycle at 25 °Cfor 1 week. The plants were then transplanted to containersfilled with a mixture of soil and sand (3:1) and were wateredregularly for 4 weeks. Five-week-old tobacco plants in similargrowth state were subjected to 30 mm MV treatment for 2weeks. Leaves from WT, VC and transgenic tobacco lineswere used as samples to detect chlorophyll, H2O2 and activi-ties of SOD, CAT and POD.The WT,VC and transgenic lineswere cultured in MS medium under a 16 h light/8 h dark cycleat 25 °C for 1 week and then transplanted to MS suppliedwith 30 mm MV for 1 week. Whole seedlings were used todetect the expression of SOD, CAT and POD.
Analysis of RWC, IL, MDA, ROS accumulationand antioxidant enzyme activity
RWC was measured according to Barrs & Weatherley (1962).Fresh weight (FW) of tobacco leaves was recorded followedby soaking the leaves for 4 h in distilled water at room tem-perature with constant light. After this treatment the turgidweight (TW) was recorded. The leaves were then dried for24 h at 80 °C to obtain the total dry weight (DW). RWC wascalculated from the equation: RWC (%) = [ (FW - DW)/(TW - DW) ] ¥ 100. IL was measured based on the methoddescribed by Jiang & Zhang (2001) with slight modifications.Leaves were cut into strips and incubated in 10 mL distilledwater at room temperature for 12 h. Initial conductivity (C1)was measured with a DDBJ-350 conductivity meter (Shang-hai Precision & Scientific Instrument Co., Ltd., Shanghai,China) followed by boiling the samples for 30 min tomeasure complete ion leakage (I). The leaves were thencooled to room temperature to measure the electrolyte con-ductivity (C2). Ion leakage (I) was calculated according tothe equation: I (%) = C1/C2 ¥ 100. MDA content was deter-mined by the thiobarbituric acid (TBA)-based colorimetricmethod as described by Heath & Packer (1968).
The measurement of H2O2 content was carried out asdescribed in the previous study (Jiang & Zhang 2001). His-tochemical detection of H2O2 and O2
- was performed accord-ing to Pan et al. (2012). To measure H2O2, seedlings wereincubated with 5 mg mL-1 DAB at pH 3.8 for 20 h and todetect O2
- samples were incubated in 0.5 mg mL-1 NBT for20 h in the dark. Seedlings were then decolorized by boilingin 96% ethanol for 10 min followed by cooling and sampleextraction with fresh ethanol at room temperature and pho-tographed. The activities of SOD, POD and CAT were spec-trophotometrically measured. SOD, POD and CAT wereextracted from about 0.5 g of samples homogenized in 5 mLextraction buffer containing 0.05 M phosphate buffer(pH 7.8) and 1% polyvinylpyrolidone. The homogenate wasthen centrifuged at 10 000 g for 10 min at 4 °C and the result-ing supernatant was used to assay enzyme activities. SODand CAT activities were measured using SOD and CATDetection Kit (A001 and A007; Jiancheng, Nanjing, China)
according to the manufacturer’s instruction.Total POD activ-ity was measured as a change in absorbance at 470 nm afterguaiacol oxidation according to the method described previ-ously (Polle, Otter & Seifert 1994).
Sequence analysis
Amino acid sequences were aligned by ClusterX (http://www.clustal.org/) and phylogenetic tree was constructed using Clus-terX and Mega (http://www.megasoftware.net/) softwares.
Statistical analysis
Statistical analysis was performed using Microsoft Excel andStatistical Package for the Social Sciences (Chicago, IL,USA). Analysis of variance was used to compare the statis-tical difference based on Student’s t-test (n = 4). For themeasurement of RWC, IL, MDA, H2O2, SOD and CAT in 3-or 5-week-old tobacco plants, each sample contained fourreplicates (each replicate contained three seedlings). For themeasurement of H2O2, SOD and CAT in 1-week-old tobaccoplants, each sample contained four replicates (each replicatecontained five seedlings).
RESULTS
Cloning and sequence analysis of the full-lengthTaASR1 cDNA
From the DFCI wheat gene index database, 16 ASR ESTsequences were acquired. Based on sequence similarityamong them and with other ASRs (Supporting InformationTable S1), they were divided into seven groups. Members ofone group may be homologs of the same ASR gene as theyexhibited significantly high sequence similarity. Althoughgroup 1 shared no similarity with ASRs from rice, they exhib-ited high similarity with ASRs from other species such asBrachypodium distachyon, Solanum lycopersicum and Musaacuminate. Groups 2–6 showed high identity with OsASR1,OsASR1, OsASR3, OsASR3 and OsASR4, respectively. Thegroup 7 did not have any identity with ASRs from otherspecies based on BLASTX analysis. RT-PCR results showedthat expression of BE414974 (group 1), TC418737 (group 2),CJ554788 (group 3) and CK154393 (group 6) was induced byPEG treatment using primers P22, P23, P24 and P25, respec-tively (Supporting Information Table S2) (Supporting Infor-mation Fig. S1). Genes of groups 4 and 5 were not amplifiedin the bread wheat (Chinese Spring) despite using multipleprimer combinations, which may due to sequence variation indifferent varieties and the allohexaploid nature of itsgenome. Notably, group 1 exhibited higher expression innormal conditions and was strongly up-regulated after PEGtreatment. Therefore group 1 was selected as a candidate forfunctional characterization.
Employing RACE-PCR the full-length cDNA of ASR gene,designated as TaASR1 (GenBank Accession No.: HQ287799),was isolated from the leaves of wheat seedlings. TaASR1cDNA comprised of 544 bp with a 414 bp open reading frame
4 W. Hu et al.
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

(ORF). The deduced TaASR1 protein contained 137 aminoacid residues with a predicted molecular mass of 15.3 kDa.BLASTX analysis showed that TaASR1 shared a high degreeof sequence identity with ASRs from other plant species:86% sequence identity with BdASR3 from B. distachyon,80% with ZmASR2 from Zea mays, 72% with MaASR fromMusa ABB Group and 70% with OsASR from Oryza sativaJaponica Group. Based on amino acid sequence alignment,two highly conserved regions were observed in TaASR1: asmall N-terminal sequence with 18–20 amino acids anda longer C-terminal region with at least 80 amino acids(Supporting Information Fig. S2). The N-terminal con-sensus sequence has a stretch of six His residues in a 10amino acid sequence that is typical for Zn-binding. In theC-terminal region, there are two Ala-rich regions, one sitefor N-myristoylation, a putative nuclear targeting signal(KKQDHKEAKEASGEKKHHHFG) and an ABA/WDSdomain that is described in ABA-, water stress- and ripening-induced proteins (Supporting Information Fig. S2). A phylo-genetic tree was constructed based on the amino acidsequence alignment of TaASR1 and 15 other plant ASRsfrom GenBank (Supporting Information Fig. S3). On an evo-lutionary timescale, these 16 ASR proteins branch out intoseparate monocot and dicotyledon groups with TaASR1 veryclose to OsASR and ZmASR2 in monocot group, implyingthat ASR family is conserved in monocot and dicotyledongroups. These results suggested that TaASR1 obtained in thisstudy is a novel member of the ASR family in wheat.
TaASR1 transcript is induced by PEG6000, ABAand H2O2 treatments
To determine the expression patterns of TaASR1 in differentwheat tissue, qRT-PCR was carried out with mRNA fromdifferent tissues. TaASR1 was expressed in all tissues includ-ing root, stem, leaf, stamen, pistil and lemma with higherexpression in root, stem, leaf and lemma (Fig. 1a). To inves-tigate the response of TaASR1 to osmotic stress, PEG6000treatment was applied. Results showed that TaASR1 expres-sion increased 2.2-fold after 2 h of PEG treatment andreached the highest level (5.7-fold) after 6 h followed by adecrease (Fig. 1b). Because osmotic stress is known to induceaccumulation of various signal molecules, the effect of ABA,MeJA, SA, auxin and H2O2 on TaASR1 transcription was alsoexamined. After ABA treatment, expression of TaASR1 wasinduced to the highest level (4.0-fold) at 2 h (Fig. 1c) andafter H2O2 treatment TaASR1 levels were induced 2.9-fold at2 h and peaked at 6 h (14.7-fold) (Fig. 1d). TaASR1 transcriptinduction by MeJA, SA and auxin was marginal and occurredmuch later (Supporting Information Fig. S4). These resultssuggested that TaASR1 transcript was strongly induced byPEG6000, ABA and H2O2.
ABA and H2O2 signalling are involved in theinduction of TaASR1 by PEG6000 treatment
To explore whether the up-regulation of TaASR1 underPEG6000 treatment involves ABA signalling, tungstate and
fluridone were chosen as the inhibitors of ABA biosynthesis(Jiang & Zhang 2002; Zhang et al. 2011a). Plants were pre-treated with tungstate and fluridone for 6 h followed by PEGtreatment. The results clearly showed that TaASR1 wasup-regulated at 6 h after the treatment with PEG (5.1-fold)and ABA (3.1-fold) (Fig. 2), consistent with the results inFig. 1. Treatment with fluridone and tungstate reduced thefold increase to 1.8-fold and 1.2-fold at 6 h after the PEGtreatment (Fig. 2a). There was no obvious difference for the
Figure 1. Expression patterns of TaASR1 in different organs andin wheat leaves after PEG6000, ABA, and H2O2 treatments byqRT-PCR analysis. (a) Different organs (R: root; S: stem; L: leaf;ST: stamen; P: pistil; LE: lemma); (b) 20% PEG6000 treatment;(c) 100 mm ABA treatment; (d) 10 mM H2O2 treatment. Verticalbars indicate � SE of four replicates on one sample. Threebiological experiments were performed, which produced similarresults.
The role of TaASR1 in drought stress 5
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

expression of TaASR1 with the treatment of tungstate andfluridone (Fig. 2a). In addition, TaSAG3 induced by ABA andTaSnRK2.7 not induced by ABA were chosen as markergenes to evaluate the effects of ABA biosynthesis inhibitortreatments (Zhang et al. 2011b; Zhao et al. 2012). The resultssuggested that TaSAG3 exhibited a similar expressionpattern with TaASR1 after various treatments and that treat-ment with tungstate and fluridone had no effect on theup-regulation of TaSnRK2.7 under PEG treatment (Fig. 2a).These results implied that up-regulation of TaASR1 by PEGpossibly involves ABA signalling.
To detect whether the up-regulation of TaASR1 underPEG treatment involves H2O2 signalling, DMTU andimidazole were chosen as the inhibitors and scavenger ofH2O2, respectively (Zhang et al. 2006). Wheat seedlings werepretreated with DMTU and imidazole for 6 h to stop theproduction of H2O2, followed by exposure to PEG6000 treat-ment for 6 h. Figure 2b showed that TaASR1 was induced byPEG (4.93-fold) and H2O2 (10.3-fold) treatment, which is inline with the results in Fig. 1. Pretreatment with inhibitors orscavenger of H2O2 inhibited the up-regulation of TaASR1 inthe PEG6000-treated wheat seedlings (Fig. 2b). The treat-ment of DMTU and imidazole had no effect on the expres-sion of TaASR1 (Fig. 2b). Moreover, TaHO1 up-regulated byH2O2 was employed as a positive control gene (Xu et al.2011), which displayed a similar expression profile withTaASR1 under various treatments (Fig. 2b). These resultssuggested that the up-regulation of TaASR1 by PEG treat-ment possibly involves H2O2 signalling.
Generation of transgenic tobaccooverexpressing TaASR1
To further investigate the role of TaASR1 in drought/osmoticstress tolerance, transgenic tobacco plants overexpressingTaASR1 under the control of CaMV 35S promoter weregenerated. A total of 14 transgenic lines (T1) were identifiedby hygromycin-resistance analysis and PCR using primersspecific to TaASR1 and GFP (data not shown).Among the T1
lines, three (OE2, OE5 and OE12) segregated at a ratio of 3:1based on hygromycin resistance. Moreover, seedlings from allthree transgenic T2 lines grew well on MS medium containing40 mg L-1 of hygromycin. In this experiment, tobacco plantstransformed with the vector alone were also subjected tosimilar analysis. Expression of TaASR1 was also determinedby RT-PCR analysis, which showed the presence of TaASR1mRNA in all three transgenic lines but not in the WT (Sup-porting Information Fig. S5). Among the lines OE5 andOE12 had higher TaASR1 expression levels.
Overexpression of TaASR1 enhances droughttolerance in transgenic tobacco
To characterize the performance of TaASR1 transgenic linesunder drought stress in soil, all three transgenic lines and con-trols were tested at the seedling stage. Three-week-old seed-lings of transgenic, WT and VC tobacco plants were subjectedto water stress. After withholding water for 20 d, all leaves inWT andVC plants were severely curled,while only some of the
Figure 2. Effects of inhibitors or scavengers of ABA and H2O2 on the TaASR1 transcripts under PEG treatments. (a) Effects ofpretreatment with inhibitors of ABA biosynthesis on the expression of TaASR1 in the leaves of wheat seedlings exposed to PEG6000;(b) Effects of pretreatment with scavenger and inhibitor of H2O2 on the expression of TaASR1 in the leaves of wheat seedlings exposedto PEG6000. Vertical bars indicate �SE of three replicates on one sample. Three biological experiments were performed, which producedsimilar results.
6 W. Hu et al.
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment
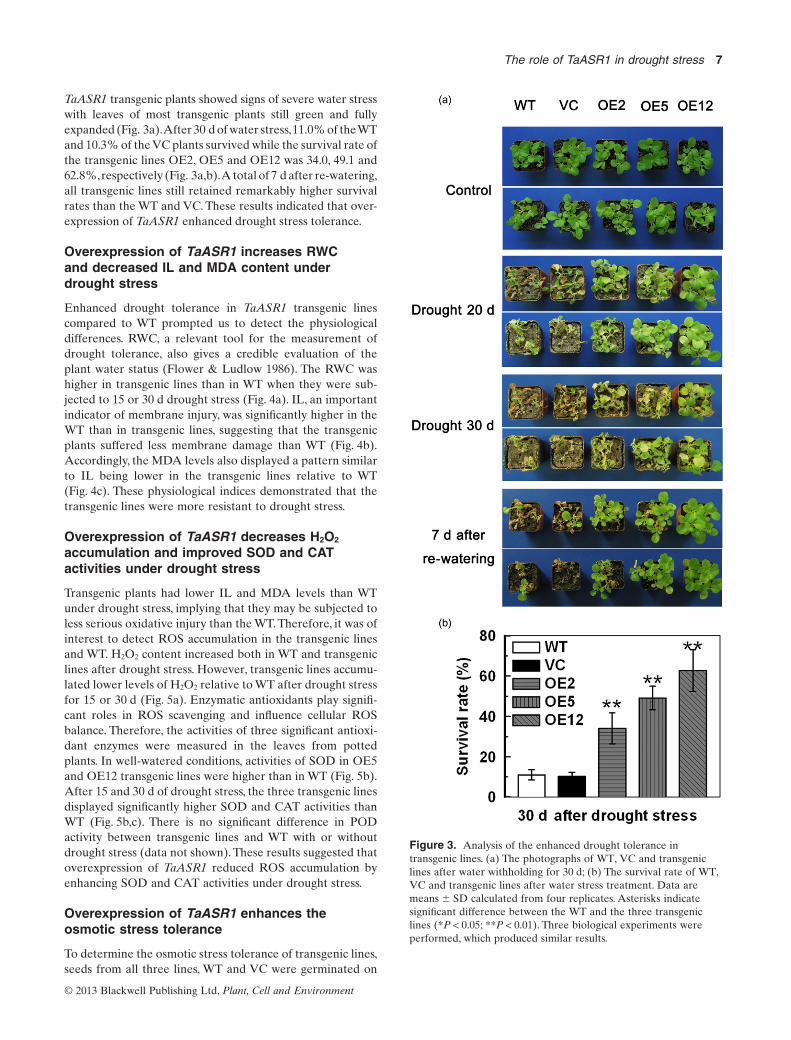
TaASR1 transgenic plants showed signs of severe water stresswith leaves of most transgenic plants still green and fullyexpanded (Fig. 3a).After 30 d of water stress,11.0% of theWTand 10.3% of the VC plants survived while the survival rate ofthe transgenic lines OE2, OE5 and OE12 was 34.0, 49.1 and62.8%,respectively (Fig. 3a,b).A total of 7 d after re-watering,all transgenic lines still retained remarkably higher survivalrates than the WT and VC. These results indicated that over-expression of TaASR1 enhanced drought stress tolerance.
Overexpression of TaASR1 increases RWCand decreased IL and MDA content underdrought stress
Enhanced drought tolerance in TaASR1 transgenic linescompared to WT prompted us to detect the physiologicaldifferences. RWC, a relevant tool for the measurement ofdrought tolerance, also gives a credible evaluation of theplant water status (Flower & Ludlow 1986). The RWC washigher in transgenic lines than in WT when they were sub-jected to 15 or 30 d drought stress (Fig. 4a). IL, an importantindicator of membrane injury, was significantly higher in theWT than in transgenic lines, suggesting that the transgenicplants suffered less membrane damage than WT (Fig. 4b).Accordingly, the MDA levels also displayed a pattern similarto IL being lower in the transgenic lines relative to WT(Fig. 4c). These physiological indices demonstrated that thetransgenic lines were more resistant to drought stress.
Overexpression of TaASR1 decreases H2O2
accumulation and improved SOD and CATactivities under drought stress
Transgenic plants had lower IL and MDA levels than WTunder drought stress, implying that they may be subjected toless serious oxidative injury than the WT. Therefore, it was ofinterest to detect ROS accumulation in the transgenic linesand WT. H2O2 content increased both in WT and transgeniclines after drought stress. However, transgenic lines accumu-lated lower levels of H2O2 relative to WT after drought stressfor 15 or 30 d (Fig. 5a). Enzymatic antioxidants play signifi-cant roles in ROS scavenging and influence cellular ROSbalance. Therefore, the activities of three significant antioxi-dant enzymes were measured in the leaves from pottedplants. In well-watered conditions, activities of SOD in OE5and OE12 transgenic lines were higher than in WT (Fig. 5b).After 15 and 30 d of drought stress, the three transgenic linesdisplayed significantly higher SOD and CAT activities thanWT (Fig. 5b,c). There is no significant difference in PODactivity between transgenic lines and WT with or withoutdrought stress (data not shown). These results suggested thatoverexpression of TaASR1 reduced ROS accumulation byenhancing SOD and CAT activities under drought stress.
Overexpression of TaASR1 enhances theosmotic stress tolerance
To determine the osmotic stress tolerance of transgenic lines,seeds from all three lines, WT and VC were germinated on
Figure 3. Analysis of the enhanced drought tolerance intransgenic lines. (a) The photographs of WT, VC and transgeniclines after water withholding for 30 d; (b) The survival rate of WT,VC and transgenic lines after water stress treatment. Data aremeans � SD calculated from four replicates. Asterisks indicatesignificant difference between the WT and the three transgeniclines (*P < 0.05; **P < 0.01). Three biological experiments wereperformed, which produced similar results.
The role of TaASR1 in drought stress 7
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

MS or MS supplemented with 150 or 300 mM mannitol for8 d (Fig. 6).When sowing on MS medium containing 150 mMor 300 mM mannitol, the WT and VC germinated much laterthan those of transgenic seeds. Germination rate was clearlyhigher in transgenic lines than in WT and VC after 150 mMmannitol treatment for 2–8 d (Fig. 6B b). Furthermore, ger-mination rate was significantly higher in transgenic lines thanin WT and VC after 300 mM mannitol treatment for 4–8 d(Fig. 6C c). In addition, little difference was observedbetween the plants under normal conditions (Fig. 6A a). In asecond experiment, seedlings grown on MS medium for 7 dwere transferred to MS or MS supplemented with 150 or300 mM mannitol for 7 d followed by measurement of rootlength (Fig. 6). On MS medium alone, all transgenic plantsshowed little difference in growth compared to controls(Fig. 6D,G). On MS containing 150 mM mannitol, transgenic
lines displayed longer root length than WT and VC(Fig. 6E,G). Moreover, on MS containing 300 mM mannitol,transgenic lines also exhibited longer root length than WTand VC (Fig. 6F,G). These results indicated that overexpres-sion of TaASR1 increased the acclimatization to osmoticstress during seed germination and root elongation in trans-genic tobacco plants.
Overexpression of TaASR1 decreases ROSaccumulation and improved SOD and CATactivities under osmotic stress
To further confirm the ability of transgenic tobacco plants toscavenge ROS, the ROS accumulation and activities of anti-oxidant enzymes were determined under osmotic stress.Seedlings grown on MS for 7 d were transferred to MS or MSsupplemented with 300 mM mannitol for 7 d. Transgenic
Figure 4. Analysis of RWC (a), IL (b) and MDA (c) in WT andtransgenic lines under normal and drought conditions.Three-week-old tobacco plants were deprived of water for 30 d.After withholding water for 15 or 30 d, tobacco leaves werecollected to measure RWC, IL, contents of MDA. Data aremeans � SD calculated from four replicates. Asterisks indicatesignificant difference between the WT and the three transgeniclines (*P < 0.05; **P < 0.01). Three biological experiments wereperformed, which produced similar results.
Figure 5. Analysis of H2O2 content (a), SOD (b) and CAT(c) activities in the WT and transgenic lines under normal anddrought conditions. Three-week-old tobacco plants were deprivedof water for 30 d. After withholding water for 15 or 30 d, tobaccoleaves were collected to detect H2O2 content, SOD and CATactivities. Data are means � SD calculated from four replicates.Asterisks indicate significant difference between the WT and thethree transgenic lines (*P < 0.05; **P < 0.01). Three biologicalexperiments were performed, which produced similar results.
8 W. Hu et al.
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

lines treated with mannitol showed lower accumulation ofH2O2 and O2
- than WT in the shoot apex, and cotyledonwhich was evident from the brown (DAB staining) and blue(NBT staining) pigments (Fig. 7a). Under normal growthconditions, there was no difference in H2O2 or O2
- accumu-lation between WT and transgenic seedlings (data notshown). H2O2 content, SOD and CAT activities were meas-ured in whole seedlings from tobacco plants with or withoutosmotic stress. In normal growth conditions, CAT activity inthe transgenic lines was slightly higher than in WT (Fig. 7d).After 7 days of osmotic stress, transgenic lines containedsignificantly lower H2O2 and higher SOD and CAT activitiesthan WT (Fig. 7b–d). These results suggested that overex-pression of TaASR1 reduced ROS accumulation by enhanc-ing the SOD and CAT activities under osmotic stress.
TaASR1 regulates ROS-related andstress-responsive genes underosmotic treatment
To gain deeper understanding of TaASR1 function in drought/osmotic stress tolerance, transcript levels of ROS scavenging-related and stress-responsive genes were detected in WT andthe transgenic line OE12 under normal and osmotic stressconditions (Fig. 8). Genes selected for this analysis includeNtSOD, NtCAT and NtPOX involved in ROS detoxification,NtNCED1 involved in ABA biosynthesis, NtERD10C,NtERD10D and NtLEA5 related to stress defence, the regu-latory gene NtDREB3 and lipid-transfer protein genesNtLTP1 and TobLTP1. Under normal conditions, expressionof NtERD10C, NtERD10D and NtNCED1 were higher inOE12 than in WT. Although expression levels of all tested
genes were up-regulated by osmotic stress, they were higher intransgenic plants than in WT except for the NtPOX underosmotic stress.These results suggested that overexpression ofTaASR1 in tobacco enhances the expression of ROS-relatedand stress-responsive genes under osmotic stress.
Overexpression of TaASR1 enhances oxidativestress tolerance in transgenic tobacco
To further confirm the role of TaASR1 in regulating anti-oxidant mechanisms, the relationship between TaASR1 andoxidative stress was investigated. To induce oxidative stress,1- and 1-week-old tobacco plants were subjected to 30 mmmethyl viologen treatment. This resulted in more severecotyledon bleaching or chlorosis in WT and VC plants thantransgenic plants (Fig. 9a–c). In addition, transgenic linesdisplayed higher chlorophyll and lower H2O2 contents thanWT and VC (Fig. 9d,e).Thus, the transgenic plants were moreresistant to oxidative stress than the controls. To determinewhether overexpression of TaASR1 affects the antioxidativeenzymes under oxidative stress, activities of SOD, CAT andPOD were measured (Fig. 10a,b). These activities increasedin WT,VC and the three lines after oxidative stress. However,transgenic lines had improved CAT and POD activitiesunder oxidative stress relative to WT and VC. We also exam-ined the expression of NtCAT and NtPOX, which werehigher in transgenic lines than in WT under oxidative stress(Fig. 10c,d). There was no obvious difference in activity andexpression of SOD between the transgenic lines and WT(data not shown). These results suggested that overexpres-sion of TaASR1 reduced H2O2 accumulation by enhancingantioxidant system under oxidative stress.
(a) (b) (c)
(A) (B) (C) (D)
(E)
(F)
(G)
Figure 6. Enhanced osmotic stress tolerance of TaASR1-overexpressing plants. Transgenic lines, WT or VC germinated on MS mediumcontaining 0 (A, a) 150 mM (B, b) or 300 mM (C, c) mannitol for 8 d, and the germination rate was calculated. A, B and C are photos of thefirst 8 d after germination on mediums. One-week-old tobacco seedlings were transplanted to MS or MS supplied with 150 or 300 mMmannitol for 1 week. The photographs were taken (D, E, F) and root length was calculated (G). Vertical bars indicate � SD calculated fromfour replicates. Four biological experiments were performed, which produced similar results.
The role of TaASR1 in drought stress 9
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

TaASR1 locates in the cellular nucleus
Cellular localization of the TaASR1 protein was investi-gated in a transient expression assay with 35S::TaASR1-GFP (pCAMBIA1304-TaASR1-GFP) translational fusionin onion epidermal cells using particle bombardment.
Fluorescence of the 35S::TaASR1-GFP chimera wasassociated with cellular nucleus in onion epidermal cells, sug-gesting a nuclear localization (Fig. 11a). In contrast, fluo-rescence of cells transformed with the control 35S::GFP(pCAMBIA1304-GFP) was distributed throughout thecell including the nucleus, cytoplasm and cytomembrane(Fig. 11a).In addition,subcellular localization of theTaASR1-GFP fusion proteins was determined in young roots fromtransgenic tobacco seedlings under a microscope (Fig. 11b).Inthe root cells of plants transformed with TaASR1-GFP, fluo-rescence was observed in the nuclei. In contrast, fluorescencewas found in cell nuclei, cytoplasm and plasma membrane inthe root cells of GFP-expressing plants.Thus,TaASR1 proteinwas confirmed to be localized in the cellular nucleus, which isconsistent with the characteristics of transcription factors.
N-terminal domain of TaASR1 showstranscriptional activity
Transcriptional activity of the TaASR1 protein was examinedusing a yeast expression system. Yeast strain AH109 wastransformed with the fusion plasmids pGBKT7-TaASR1,pGBKT7-TaASR1-N, pGBKT7-TaASR1-C and pGBKT7(Control) (Fig. 12a) and the growth status of transformantswas evaluated. Yeast cells containing pGBKT7-TaASR1 and
Figure 7. Analysis of ROS accumulation and the activities ofSOD and CAT in WT and the transgenic lines under normal andosmotic conditions. One-week-old seedlings after mannitoltreatment were used to detect H2O2 and O2
- by DAB (upper) andNBT (lower) staining (a). H2O2 content (b), the activities of SOD(c) and CAT (d) were measured in WT and transgenic lines undernormal and osmotic conditions. Data are means � SD calculatedfrom four replicates. Asterisks indicate significant differencebetween the WT and the three transgenic lines (*P < 0.05;**P < 0.01). Three biological experiments were performed, whichproduced similar results.
Figure 8. Analysis of expression levels of ROS-related andstress-responsive genes in the WT and the transgenic line (OE12)by qRT-PCR under normal and osmotic conditions. One-week-oldtobacco seedlings were transplanted to MS or MS supplied with300 mM mannitol for 1 week. The whole seedlings were used todetect gene expression. Vertical bars indicate � SE of fourreplicates on one sample. Three biological experiments wereperformed, which produced similar results.
10 W. Hu et al.
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

pGBKT7-TaASR1-N grew well in SD medium lacking histi-dine, whereas cells containing pGBKT7-TaASR1-C andpGBKT7 did not grow (Fig. 12b). In the presence of X-gal,yeast cells that grew well on the SD medium without histi-dine turned blue (Fig. 12b), suggesting the activation ofLacZ, a second reporter gene. In addition, all yeast cells grewwell on SD medium lacking tryptophan (Fig. 12b). Theseresults demonstrated the presence of transcriptional activityin the full-length TaASR1 protein and N-terminal domainand its absence in the C-terminal domain.
DISCUSSION
Some ASR genes have been reported to play a role inresponse to abiotic stresses and ABA (Kalifa et al. 2004b;Yang et al. 2005; Kim et al. 2009; Liu et al. 2010; Hsu et al.2011; Saumonneau et al. 2012). However, there is no reporton ASR genes in wheat which is an important staple crop. Inthis study we report the presence of ASR gene family inwheat based on the limited ESTs available in DFCI (wheatgene index database) and the up-regulation of ASR genetranscripts in wheat plants subjected to PEG treatment.Further investigation indicated that TaASR1 transcriptswere also induced by ABA and H2O2 treatments and thatthe up-regulation of TaASR1 by PEG treatment possiblyinvolves ABA and H2O2 signalling. Although the response ofASR genes to abiotic stresses and ABA has been confirmedin some plant species, their exact roles in abiotic stress toler-ance are unclear. In particular the correlation between ASRgenes and ROS under drought stress is not known. Ingeneral, the association of ROS homeostasis to oxidative
stress is well known. When the level of ROS is low in cells,they act as signal molecules. However, under severe bioticand abiotic stress, ROS can be overproduced and causedamage to cell growth by oxidizing proteins, lipids and DNA(Polle 2001; Mittler et al. 2004; Ning et al. 2010). Therefore,plants have developed antioxidant systems to protect them-selves against oxidative stress caused by abiotic stressesthrough adjusting ROS homeostasis. Thus, in the presentstudy, we were interested in detecting the correlationbetween TaASR1 and ROS under drought/osmotic stress,which improved the understanding of ASR function duringabiotic stresses.
On the basis of physiological and biochemical analysis, wehave shown that the enhanced tolerance of transgenic lines todrought stress is related to the maintenance of better waterstatus, less IL and MDA content. MDA is the product of lipidperoxidation caused by ROS and in general is used to evalu-ate ROS-mediated injuries in plants (Moore & Roberts1998). Thus, this study implies that lipid peroxidation causedby ROS damage was relatively relieved in transgenic plantsunder drought stress. To minimize cellular damage caused bydrought stress plants maintain their ROS balance (Mittleret al. 2004; Miller et al. 2010). Transgenic tobacco lines accu-mulated a lower level of H2O2 and O2
- under drought/osmotic stress relative to WT, which implies that ROSscavenging systems in transgenic plants might work moreeffectively compared to WT. To detoxify ROS-induced stress,plants have evolved a complex antioxidant system thatincludes several enzymes, which scavenge ROS and protectcells against oxidative stress (Jaleel et al. 2009; Huang, Liu &Chen 2010; Miller et al. 2010). Among these enzymes, SOD
(d)(a)
(e)
(b)
(c)
Figure 9. Analysis of the enhanced oxidative stress tolerance in transgenic lines. One-week-old tobacco plants were subjected to MVtreatment for 1 week, and the photographs were taken (a). Five-week-old tobacco plants were subjected to MV treatment for 2 weeks, thephotographs were taken (b, c). Tobacco leaves were sampled to detect chlorophyll (d) and H2O2 (e) content. Data are means � SDcalculated from four replicates. Asterisks indicate significant difference between the WT and the three transgenic lines (*P < 0.05;**P < 0.01). Three biological experiments were performed, which produced similar results.
The role of TaASR1 in drought stress 11
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

provides the first line of defence against ROS by catalysingthe dismutation of O2
- to oxygen and H2O2, which are thenscavenged by the coordinated actions of CAT and POD(Blokhina, Virolainen & Fagerstedt 2003; Huang et al. 2010).Analysis of the antioxidant enzymes indicated that the trans-genic lines displayed significantly higher SOD and CATactivities than WT under drought/osmotic stress in 1- and3-week-old tobacco plants. These suggested that overexpres-sion of TaASR1 gene enhanced the activation of antioxidantdefence system, which in turn protected transgenic linesagainst ROS-mediated injury under drought/osmotic stress.Moreover, transgenic plants had lower H2O2 content andimproved antioxidant system than controls under MV treat-ment. Recent studies showed that overexpression of OsASR1enhanced cold tolerance in transgenic rice plants (Kimet al. 2009). Subsequently, OsASR1 was expressed in yeast,where OsASR1 protein improved tolerance to H2O2-inducedoxidative stress by activating redox homeostasis and prote-ostasis, and up-regulating a variety of antioxidant proteins(Kim et al. 2012). Furthermore, Arenhart et al. (2013)
reported that putative superoxide dismutase (gi|108708142),and cytosolic ascorbate peroxidases -OsAPx1 and OsAPx2(LOC_Os03g17690) (gi|115452337) were reduced in ASR5-RNAi rice plants when compared to non-transformed plantsunder normal conditions. In addition, ASR5-silenced riceplants are sensitive to aluminium and drought stresses. Thesestudies implied that antioxidant system may be involved inASRs conferring abiotic stresses tolerance. In the presentstudy, we have provided the physiological evidence thatTaASR1 confers drought/osmotic and oxidative stress toler-ance by enhancing antioxidant system and thereby scaveng-ing ROS in vivo.
To gain further insights into the function of TaASR1 indrought/osmotic stress tolerance at molecular level, theexpression of stress-responsive and antioxidant defence-associated genes was investigated. The phytohormone ABAplays an important role in responses to abiotic stresses. LilyASR was reported to be involved in ABA signalling and inconferring drought and salt resistance in Arabidopsis. In theleaves of transgenic Arabidopsis, ABA levels were markedly
Figure 10. Analysis of activities and expression of CAT and POD in the WT and transgenic lines under normal conditions and MVtreatment. Five-week-old tobacco plants under normal conditions and MV treatment were sampled to detect CAT (a) and POD (b)activities. One-week-old tobacco seedlings under normal conditions and MV treatment were used to detect the expression of CAT (c) andPOD (d). Data are means � SD calculated from four replicates. Asterisks indicate significant difference between the WT and transgenic lines(*P < 0.05; **P < 0.01). Three biological experiments were performed, which produced similar results.
12 W. Hu et al.
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

induced under water-deficit conditions, resulting in stomatalclosure (Yang et al. 2005). Biochemical and genetic evidencerevealed that ABA biosynthesis under stress is largely regu-lated by the rate-limiting enzyme, 9-cis-epoxycarotenoiddioxygenase (NCED) (Qin & Zeevaart 1999; Huang et al.2010). Expression of NtNCED1 was up-regulated inTaASR1-overexpressing plants compared to WT, implyingthat ABA biosynthesis may be promoted in transgenic plants.Dehydration-responsive element-binding proteins (DREBs)also play important roles in regulating stress responses. Over-expression of DREB genes has been shown to enhancedrought tolerance in plants (Umezawa et al. 2006). GrapeASR (VvMSA) was reported to interact with a DREB tran-scription factor in the nucleus (Saumonneau et al. 2008). Inthis study, NtDREB3, a DREB family member, was inducedin transgenic plants under osmotic stress, which implied theactivation of DREB-mediated stress defence. NtERD10(C/D) and NtLEA5 encode groups 2 and 5 LEA proteins thatplay a role in binding water, stabilizing labile enzymes, andprotecting cellular and macromolecular structures during tol-erance to dehydration (Hundertmark & Hincha 2008; Liuet al. 2009). Stronger induction of NtERD10C, NtERD10Dand NtLEA5 suggested that more LEA proteins may besynthesized in transgenic plants. This resulted in less destruc-tion to the transgenic plants, which are also supported bygreater water retention and lesser membrane damage underdrought stress. NtLTP1 and TobLTP1 belong to the lipid-transfer protein family that is involved in plants’ responseto ABA, cold-, drought- and salt-stress (Dunn et al. 1991;Torres-Schumann, Godoy & Pintor-Toro 1992; Ouvrard et al.1996; Trevino & OConnell 1998; Zhang et al. 2005). Overex-pression of TERF1 in tobacco enhanced drought tolerance,ABA sensitivity and the expression of NtLTP1 and TobLTP1with ABA treatment (Zhang et al. 2005). Cutin monomerelicited the synthesis of H2O2 in rice leaves and induced theexpression of lipid-transfer protein gene OsLTP5 (Kim et al.2008). In the present study, transgenic plants showedincreased expression of NtLTP1 and TobLTP1 underosmotic stress, suggesting that TaASR1 affects the expressionof some lipid-transfer protein genes that are responsive toABA, H2O2 and drought stress. Lipid-transfer protein wasalso shown to be induced in tobacco plants overexpressingtomato ASR1 (Kalifa et al. 2004b). In addition, increasedexpression of NtSOD and NtCAT in transgenic lines maycontribute to the enhanced ability to scavenge ROS underdrought/osmotic stress.
Zinc is an essential cofactor that is bound to proteins bycoordination of the amino acids His, Asp, Glu and Cys, andwater molecules (Rom et al. 2006). Zinc-binding sites have
Figure 11. Subcellular localization of TaASR1 fused with GFP.The fusion protein 35S::TaASR1-GFP (pCAMBIA1304-TaASR1-GFP) and 35 S::GFP (pCAMBIA1304-GFP, control)were transiently expressed in onion epidermal cells and visualizedwith laser-scanning confocal microscopy after bombardment 24 h(a). Microscopy images of tobacco root cortical cells transformedwith GFP alone as a control or TaASR1 (b). Three biologicalexperiments were performed, which produced similar results.
The role of TaASR1 in drought stress 13
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

been classified depending on the ability of these residues toform part of a catalytic domain or stabilize the tertiary struc-ture. His is the most common and important residue forzinc-binding in catalytic sites (Rom et al. 2006). TaASR1protein contains a characteristic stretch of six His residues atthe N-terminal consensus, implying that TaASR1 may act asa transcriptional regulator and bind to DNA in a Zn2+-dependent manner during the transition from a disordered toan ordered state (Cakir et al. 2003; Kalifa et al. 2004a;Goldgur et al. 2007; Maskin et al. 2007; Shkolnik & Bar-Zvi2008). Although the zinc-dependent DNA-binding of tomatoASR1 was mapped to the central/carboxy-terminal domain,two zinc ions that are bound to the amino-terminal domainwere also observed and the complete ASR1 protein exhib-ited maximal DNA-binding capacity (Rom et al. 2006). Inthis study, TaASR1 could activate transcription in yeast cellsand the activity was mapped to a variable N-terminal region,which is consistent with the recent study on the strawberryASR gene (Chen et al. 2011). Subcellular localization indi-cated that the TaASR1-GFP fusion protein was located innucleus, which is in line with previous studies on ASR genesfrom lily and grape (Cakir et al. 2003; Wang et al. 2003).Thus,it is speculated that TaASR1 functions as a transcriptionfactor and plays a role in regulating the expression of stressdefence genes.
In conclusion, the findings of this study demonstratedTaASR1 function in drought/osmotic stress tolerance.TaASR1 conferred drought/osmotic stress tolerance throughregulating the expression of stress- and defence-associated
genes and enhancing the antioxidant system, thus preventingplants from oxidative damage. Further studies are required toidentify the targets of TaASR1. This will enhance characteri-zation of the ROS scavenging-mediated drought tolerance.
ACKNOWLEDGMENTS
This work was supported by International S & T CooperationKey Projects of MoST (Grant no. 2009DFB30340), NationalGenetically Modified New Varieties of Major Projects ofChina (2011ZX08002-004, 2011ZX08010-004), Key Projectsof S & T Research of MoE of China (Grant no. 109105) andWuhan Municipal S & T research project (Grant no.201120922286). We thank Analytical and Testing Center ofHuazhong University of Science and Technology (HUST) forsupplying the MJ Research Opticon 2 qRT-PCR Machine.
REFERENCES
Arenhart R.A., Lima1 J.C., Pedron M., et al. (2013) Involvement of ASR genesin aluminum tolerance mechanisms in rice. Plant, Cell & Environment 36,52–67.
Barrs H.D. & Weatherley P.E. (1962) A reexamination of the relative turgiditytechnique for estimating water deficit in leaves. Australian Journal of Bio-logical Sciences 15, 413–428.
Bartels D. (2001) Targeting detoxification pathways: an efficient approach toobtain plants with multiple stress tolerance? Trends in Plant Science 6,284–286.
Battaglia M., Olvera-Carrillo Y., Garciarrubio A., Campos F. & CovarrubiasA.A. (2008) The enigmatic LEA proteins and other hydrophilins. PlantPhysiology 148, 6–24.
Figure 12. Transactivation activity of the TaASR1 protein in yeast. (a) A schematic diagram illustrating the TaASR1 cDNA fragmentsencoding different portions of TaASR1 that were fused to DNA sequences encoding the GAL4 DNA binding domain in the yeast vectorpGBKT7. (b) Transactivation analysis of TaASR1 in yeast. Fusion proteins of pGBKT7-TaASR1, pGBKT7-TaASR1-N, pGBKT7-TaASR1-Cand pGBKT7 were expressed in yeast strain AH109. The transformants were streaked on the SD/Trp- and SD/His- medium. The plates wereincubated for 3 d and subjected to X-gal assay. Three biological experiments were performed, which produced similar results.
14 W. Hu et al.
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

Blokhina O., Virolainen E. & Fagerstedt K.V. (2003) Antioxidants, oxidativedamage and oxygen deprivation stress: a review. Annals of Botany 91, 179–194.
Cakir B., Agasse A., Gaillard C., Saumonneau A., Delrot S. & Atanassova R.(2003) A grape ASR protein involved in sugar and abscisic acid signaling.The Plant Cell 15, 2165–2180.
Carrari F., Fernie A.R. & Iusem N.D. (2004) Heard it through the grapevine?ABA and sugar cross-talk: the ASR story. Trends in Plant Science 9, 57–59.
Chen J.Y., Liu D.J., Jiang Y.M., Zhao M.L., Shan W., Kuang J.F. & Lu W.J.(2011) Molecular characterization of a strawberry FaASR gene in relation tofruit ripening. PLoS ONE 6, e24649.
Dunn M.A., Hughes M.A., Zhang L., Pearce R.S., Quigley A.S. & Jack P.L.(1991) Nucleotide sequence and molecular analysis of the low temperatureinduced cereal gene. BLT4. Molecular and General Genetics 229, 389–394.
Flower D.J. & Ludlow M.M. (1986) Contribution of osmotic adjustment to thedehydration tolerance of water-stressed pigeon pea (Cajanus cajan (L.)millsp.) leaves. Plant, Cell & Environment 9, 33–40.
Frankel N., Carrari F., Hasson E. & Iusem N.D. (2006) Evolutionary history ofthe Asr gene family. Gene 15, 74–83.
Garay-Arroyo A., Colmenero-Flores J.M., Garciarrubio A. & CovarrubiasA.A. (2000) Highly hydrophilic proteins in prokaryotes and eukaryotes arecommon during conditions of water deficit. Journal of Biological Chemistry275, 5668–5674.
Goldgur Y., Rom S., Ghirlando R., Shkolnik D., Shadrin N., Konrad Z. &Bar-Zvi D. (2007) Desiccation and zinc binding induce transition of tomatoabscisic acid stress ripening 1, a water stress- and salt stress-regulated plant-specific protein, from unfolded to folded state. Plant Physiology 143, 617–628.
Heath R.L. & Packer L. (1968) Photoperoxidation in isolated chloroplasts. I.Kinetics and stoichiometry of fatty acid peroxidation. Archives in Biochem-istry and Biophysics 125, 189–198.
Hong S.H., Kim I.J., Yang D.C. & Chung W.I. (2002) Characterization of anabscisic acid responsive gene homologue from Cucumis melo. Journal ofExperimental Botany 53, 2271–2272.
Horsch R.B., Fry J.E., Hoffmann N.L., Eichholtz D., Rogers S.C. & Fraley R.T.(1985) A simple and general method for transferring genes into plants.Science 227, 1229–1231.
Hsu Y.F., Yu S.C., Yang C.Y. & Wang C.S. (2011) Lily ASR protein-conferredcold and freezing resistance in Arabidopsis. Plant Physiolology and Bio-chememistry 49, 937–945.
Hu H.H., Dai M.Q., Yao J.L., Xiao B.Z., Li X.H., Zhang Q.F. & Xiong L.Z.(2006) Overexpressing a NAM,ATAF, and CUC (NAC) transcription factorenhances drought resistance and salt tolerance in rice. Proceedings of theNational Academy of Sciences of the United States of America 103, 12987–12992.
Huang J.C., Lin S.M. & Wang C.S. (2000) A pollen-specific and desiccation-associated transcript in Lilium longiflorum during development and stress.Plant & Cell Physiology 41, 477–485.
Huang X.S., Liu J.H. & Chen X.J. (2010) Overexpression of PtrABF gene, abZIP transcription factor isolated from Poncirus trifoliata, enhances dehy-dration and drought tolerance in tobacco via scavenging ROS and modulat-ing expression of stress-responsive genes. BMC plant Biology 10, 230.
Hundertmark M. & Hincha D.K. (2008) LEA (late embryogenesis abundant)proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics9, 118.
Iusem N.D., Bartholomew D.M., Hitz W.D. & Scolnik P.A. (1993) Tomato(Lycopersicon esculentum) transcript induced by water deficit and ripening.Plant Physiology 102, 1353–1354.
Jaleel C.A., Riadh K., Gopi R., Manivannan P., Inès J., Al-Juburi H.J., ZhaoC.X., Shao H.B. & Panneerselvam R. (2009) Antioxidant defense response:physiological plasticity in higher plants under abiotic constraints. ActaPhysiologiae Plantarum 31, 427–436.
Jeanneau M., Gerentes D., Foueillassar X., Zivy M., Vidal J., Toppan A. &Perez P. (2002) Improvement of drought tolerance in maize: towards thefunctional validation of the Zm-Asr1 gene and increase of water use effi-ciency by over-expressing C4-PEPC. Biochimie 84, 1127–1135.
Jiang M.Y. & Zhang J.H. (2001) Effect of abscisic acid on active oxygenspecies, antioxidative defence system and oxidative damage in leaves ofmaize seedlings. Plant & Cell Physiology 42, 1265–1273.
Jiang M.Y. & Zhang J.H. (2002) Water stress-induced abscisic acid accumula-tion triggers the increased generation of reactive oxygen species andup-regulates the activities of antioxidant enzymes in maize leaves. Journal ofExperimental Botany 53, 2401–2410.
Kalifa Y., Gilad A., Konrad Z., Zaccai M., Scolnik P.A. & Bar-Zvi D. (2004a)The water- and salt-stress-regulated Asr1 (abscisic acid stress ripening) geneencodes a zinc-dependent DNA-binding protein. Biochemistry Journal 381,373–378.
Kalifa Y., Perlson E., Gilad A., Konrad Z., Scolnik P.A. & Bar-Zvi D. (2004b)Over-expression of the water and salt stress-regulated Asr1 gene confers anincreased salt tolerance. Plant, Cell & Environment 27, 1459–1468.
Kim I.S., Kim Y.S. & Yoon H.S. (2012) Rice ASR1 protein with reactive oxygenspecies scavenging and chaperone-like activities enhances acquired toler-ance to abiotic stresses in Saccharomyces cerevisiae. Molecules and Cells 33,285–293.
Kim S.J., Lee S.C., Hong S.K., An K., An G. & Kim S.R. (2009) Ectopicexpression of a cold-responsive OsAsr1 cDNA gives enhanced cold toler-ance in transgenic rice plants. Molecules and Cells 27, 449–458.
Kim T.H., Park J.H., Kim M.C. & Cho S.H. (2008) Cutin monomer inducesexpression of the rice OsLTP5 lipid transfer protein gene. Journal of PlantPhysiology 165, 345–349.
Klein M., Geisler M., Suh S.J., et al. (2004) Disruption of AtMRP4, a guard cellplasma membrane ABCC-type ABC transporter, leads to deregulation ofstomatal opening and increased drought susceptibility. The Plant Journal 39,219–236.
Liu H.Y., Dai J.R., Feng D.R., Liu B., Wang H.B. & Wang J.F. (2010) Charac-terization of a novel plantain Asr gene, MpAsr, that is regulated in responseto infection of Fusarium oxysporum f. sp. cubense and abiotic stresses.Journal of Integrative Plant Biology 52, 315–323.
Liu X., Wang Z., Wang L., Wu R., Phillips J. & Deng X. (2009) LEA 4 groupgenes from the resurrection plant Boea hygrometrica confer dehydrationtolerance in transgenic tobacco. Plant Science 176, 90–98.
Livak K.J. & Schmittgen T.D. (2001) Analysis of relative gene expression datausing real-time quantitative PCR and the 2-DDCt method. Methods 25, 402–408.
Maskin L., Gubesblat G.E., Moreno J.E., Carrari F.O., Frankel N., Sambade A.,Rossi M. & Iusem N.D. (2001) Differential expression of the members of theAsr gene family in tomato (Lycopersicon esculentum). Plant Science 161,739–746.
Maskin L., Frankel N., Gudesblat G., Demergasso M., Pietrasanta L. & IusemN.D. (2007) Dimerization and DNA-binding of ASR1, a small hydrophilicprotein abundant in plant tissues suffering from water loss. Biochemical andBiophysical Research Communications 352, 831–835.
Miller G., Suzuki N., Ciftci-Yilmaz S. & Mittler R. (2010) Reactive oxygenspecies homeostasis and signaling during drought and salinity stresses. Plant,Cell & Environment 33, 453–457.
Mittler R., Vanderauwera S., Gollery M. & Van Breusegem F. (2004) Reactiveoxygen gene network of plants. Trends in Plant Science 9, 490–498.
Moore K. & Roberts L.J. II (1998) Measurement of lipid peroxidation. FreeRadical Research 28, 659–671.
Ning J., Li X., Hicks L. & Xiong L. (2010) A Raf-Like MAPKKK gene DSM1mediates drought resistance through reactive oxygen species scavenging inrice. Plant Physiology 152, 876–890.
Ouvrard O., Cellier F., Ferrare K., Tousch D., Lamaze T., Dupuis J.M. &Casse-Delbart F. (1996) Identification and expression of water stress- andabscisic acid-regulated genes in a drought-tolerant sunflower genotype.Plant Molecular Biology 31, 819–829.
Pan J., Zhang M., Kong X., Xing X., Liu Y., Zhou Y., Liu Y., Sun L. & Li D.(2012) ZmMPK17, a novel maize group D MAP kinase gene, is involved inmultiple stress responses. Planta 235, 661–676.
Philippe R., Courtois B., McNally K.L., et al. (2010) Structure, allelic diversityand selection of Asr genes, candidate for drought tolerance, in Oryza sativaL. and wild relatives. Theoretical and Applied Genetics 121, 769–787.
Polle A. (2001) Dissection the superoxide dismutase-ascorbate-glutathionepathway by metabolic modeling: computer analysis as a step towards fluxanalysis. Plant Physiology 126, 445–462.
Polle A., Otter T. & Seifert F. (1994) Apoplastic peroxidases and lignificationin needles of Norway spruce (Picea abies L.). Plant Physiology 106, 53–60.
Qin X. & Zeevaart J.A.D. (1999) The 9-cis-epoxycarotenoid cleavage reactionis the key regulatory step of abscisic acid biosynthesis in water-stressed bean.Proceedings of the National Academy of Sciences of the United States ofAmerica 96, 15354–15361.
Rom S., Gilad A., Kalifa Y., Konrad Z., Karpasas M.M., Goldgur Y. & Bar-ZviD. (2006) Mapping the DNA- and zinc-binding domains of ASR1 (abscisicacid stress ripening), an abiotic-stress regulated plant specific protein. Bio-chimie 88, 621–628.
Saumonneau A., Agasse A., Bidoyen M.T., Lallemand M., Cantereau A.,Medici A., Laloi M. & Atanassova R. (2008) Interaction of grape ASR
The role of TaASR1 in drought stress 15
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment

proteins with a DREB transcription factor in the nucleus. FEBS Letters 582,3281–3287.
Saumonneau A., Laloi M., Lallemand M., Rabot A. & Atanassova R. (2012)Dissection of the transcriptional regulation of grape ASR and response toglucose and abscisic acid. Journal of Experimental Botany 63, 1495–1510.
Schneider A., Salamini F. & Gebhardt C. (1997) Expression patterns andpromoter activity of the cold-regulated gene ci21A of potato. Plant Physi-ology 113, 335–345.
Seki M., Kamei A., Yamaguchi-Shinozaki K. & Shinozaki K. (2003) Molecularresponses to drought, salinity and frost: common and different paths forplant protection. Current Opinion in Biotechnology 14, 194–199.
Shinozaki K., Yamaguchi-Shinozaki K. & Seki M. (2003) Regulatory networkof gene expression in the drought and cold stress responses. Current Opinionin Plant Biology 6, 410–417.
Shkolnik D. & Bar-Zvi D. (2008) Tomato ASR1 abrogates the response toabscisic acid and glucose in Arabidopsis by competing with ABI4 for DNAbinding. Plant Biotechnology Journal 6, 368–378.
Singh K., Foley R.C. & Onate-Sanchez L. (2002) Transcription factors in plantdefense and stress responses. Current Opinion in Plant Biology 5, 430–436.
Tamura T., Hara K., Yamaguchi Y., Koizumi N. & Sano H. (2003) Osmoticstress tolerance of transgenic tobacco expressing a gene encoding amembrane-located receptor-like protein from tobacco plants. Plant Physi-ology 131, 454–462.
Torres-Schumann S., Godoy J.A. & Pintor-Toro J.A. (1992) A probable lipidtransfer protein gene is induced by NaCl in stems of tomato plants. PlantMolecular Biology 18, 749–757.
Trevino M.B. & OConnell M.A. (1998) Three drought-responsive members ofthe nonspecific lipid-transfer protein gene family in Lycopersicon pennelliishow different developmental patterns of expression. Plant Physiology 116,1461–1468.
Umezawa T., Fujita M., Fujita Y., Yamaguchi-Shinozaki K. & Shinozaki K.(2006) Engineering drought tolerance in plants: discovering and tailoringgenes unlock the future. Current Opinion in Biotechnology 17, 113–122.
Urtasun N., Correa Garcia S., Iusem N. & Moretti B. (2010) Predominantlycytoplasmic localization in yeast of ASR1, a non-receptor transcriptionfactor from plants. Open Biochemistry Journal 4, 68–71.
Wang H.J., Jauh G.Y., Hsu Y.H. & Wang C.S. (2003) The nuclear localizationsignal of a pollen-specific, desiccation-associated protein of a lily is necessaryand sufficient for nuclear targeting. Botanical Bulletin of Academia Sinica 44,123–128.
Ward J.M. & Schroeder J.I. (1994) Calcium-activated K+ channels and calcium-induced calcium release by slow vacuolar ion channels in guard cell vacuolesimplicated in the control of stomatal closure. The Plant Cell 6, 669–683.
Xu D.K., Jin Q.J., Xie Y.J., Liu Y.H., Lin Y.T., Shen W.B. & Zhou Y.J. (2011)Characterization of a wheat heme oxygenase-1 gene and its responses todifferent abiotic stresses. International Journal of Molecular Sciences 12,7692–7707.
Yang C.Y., Chen Y.C., Jauh G.Y. & Wang C.S. (2005) A lily ASR proteininvolves abscisic acid signaling and confers drought and salt resistance inArabidopsis. Plant Physiology 139, 836–846.
Zhang A., Jiang M., Zhang J., Tan M. & Hu X. (2006) Mitogen-activatedprotein kinase is involved in abscisic acid-induced antioxidant defense andacts downstream of reactive oxygen species production in leaves of maizeplants. Plant Physiology 141, 475–487.
Zhang A., Zhang J., Zhang J., Ye N., Zhang H., Tan M. & Jiang M. (2011a)Nitric oxide mediates brassinosteroid-induced ABA biosynthesis involved inoxidative stress tolerance in maize leaves. Plant & Cell Physiology 52, 181–192.
Zhang H., Mao X., Jing R., Chang X. & Xie H. (2011b) Characterization of acommon wheat (Triticum aestivum L.) TaSnRK2.7 gene involved in abioticstress responses. Journal of Experimental Botany 62, 975–988.
Zhang X., Zhang Z., Chen J., Chen Q., Wang X.C. & Huang R. (2005) Express-ing TERF1 in tobacco enhances drought tolerance and abscisic acid sensi-tivity during seedling development. Planta 222, 494–501.
Zhao L., Zhang H., Zhang B., Bai X. & Zhou C. (2012) Physiological andmolecular changes of detached wheat leaves in responding to various treat-ments. Journal of Integrative Plant Biology 54, 567–576.
Zhu J.K. (2002) Salt and drought stress signal transduction in plants. AnnualReview of Plant Biology 53, 247–273.
Received 19 June 2012; received in revised form 15 January 2013;accepted for publication 22 January 2013
SUPPORTING INFORMATION
Additional Supporting Information may be found in theonline version of this article at the publisher’s web-site:
Figure S1. Expression of ESTs from ASR gene family inwheat. Ten-day-old wheat seedlings after 20% PEG6000treatment were sampled to extract RNA for RT-PCR analy-sis using TaActin as an internal control. Three biologicalexperiments were performed, which produced similar results.Figure S2. Comparison of TaASR1 with other known ASRproteins.Amino acid sequences are aligned by ClusterX soft-ware.ABA/WDS motifs are shown in the box.Letters markedwith double transverse lines refer to a putative nuclear target-ing signal. One site for N-myristoylation is marked with aster-isk. One Zn-binding His-rich region and two Ala-rich regionsare underlined. The accession numbers of these known pro-teins in GenBank are as follows: OsASR (AAX92999.1) fromOryza sativa, ZmASR2 (NP_001147703.1) from Zea mays,MaASR (ACZ60132.1) from Musa, LlASR (AAM51877.1)from Lilium longiflorum, RcASR (XP_002524296.1) fromRicinus communis.Figure S3. Phylogenetic relationship of TaASR1 withZmASR2 (NP_001147703.1) from Zea mays, OsASR(AAX92999.1) from Oryza sativa, MaASR (ACZ60132.1)from Musa, GbASR (AAR23420.1) from Ginkgo biloba,LlASR (AAM51877.1) from Lilium longiflorum, RcASR(XP_002524296.1) from Ricinus communis, SlASR4(AAY98026.1) from Solanum lycopersicum, GmASR(AAR26524.1) from Glycine max, ScASR4 (AAY98030.1)from Solanum corneliomuelleri, SlASR2 (ABY84857.1)from Solanum lycopersicum, SpASR2 (ABY84856.1) fromSolanum pimpinellifolium, LpASR2 (ABS19518.1)from Lycopersicon peruvianum, PpASR (BAA96451.1)from Pyrus pyrifolia, VpASR (ABC86744.1) from Vitis pseu-doreticulata and NbASR (ACV52581.1) from Nicotianabenthamiana. The accession numbers of these known pro-teins in GenBank are given in parentheses.Figure S4. Expression profiles of TaASR1 after MeJA (A),SA (B) and auxin (C) treatments in wheat by QPCR analysis.Vertical bars indicate � SE of four replicates on one sample.Three biological experiments were performed, which pro-duced similar results.Figure S5. Expression of TaASR1 in transgenic lines.The WTand transgenic lines were cultured in MS medium for twoweeks and the whole seedlings were used to extract RNA todetect gene expression using NtUbiquitin as an internalcontrol.Three biological experiments were performed, whichproduced similar results.Table S1. Identities of ESTs in wheat ASR gene family withrice ASR gene family by using BLASTX analysis.Table S2. Primers used for PCR analysis.
16 W. Hu et al.
© 2013 Blackwell Publishing Ltd, Plant, Cell and Environment