systems biology i: bistability macromolecules jonathan weissman
TRANSCRIPT

Systems Biology I: Bistability
Macromolecules
Jonathan Weissman

Systems Biology
How does complex biological behavior emerge from the organization of proteins into pathways and networks?
Common properties:1. Bistability
• convert continuous stimulus into discrete response2. Hysteresis
• “memory” of stimulus long after it is withdrawn

Outline for this lecture:
Use simple graphical method to describe conditions thatlead to bistability
Discuss two biological examples of bistability:Xenopus oocyte maturation - MAPK Lac operon
Go through experiments that demonstrate system is bistableUnderstand what the origin of bistability isDescribe the physiological consequences of bistability

Cellular responses to stimuli: switches versus dimmers

Ak1S
k−1I⏐ →⏐ ⏐← ⏐⏐ ⏐ A*
forward rate=k1S A[ ] =k1S Atot[ ] −k1S A*[ ]Atot =A+A*
reverse rate=k−1I A*[ ]
At steady state:
k −1I A*[ ] =k1S Atot[ ] −k1S A*[ ]
A ⏐ →⏐← ⏐⏐ A*
stimulus
inactivation
Michaelian system - no feedback
k1S k −1I
System is monostablePerturb [A*/Atot], return to ss
Assume S, I far fromsaturation

Michaelian system - no feedback, cont.
At steady state:
k −1I A*[ ] =k1S Atot[ ] −k1S A*[ ]A*[ ]Atot[ ]
=S
k−1I / k1+S, when S=k−1I / k1,
A*[ ]Atot[ ]
=0.5
A*[ ]Atot[ ]
=S
EC50 +S, EC50 =S req to produce 50% response
increase S
Response levels off becausethe larger S gets, the less inactive A there is for the kinase to act on, and the moreA* there is for the p’tase
Ak1S
k−1I⏐ →⏐ ⏐← ⏐⏐ ⏐ A*

Michaelian system with linear feedback
forward rate =basal rate + feedback rate
forward rate=k1S A[ ] + k2 A*[ ] A[ ]forward rate=(k1S+ k2 A*[ ])( Atot[ ] − A*[ ])
Extremes:With no [A*] feedback will be 0
When [A*]/[Atot]=1 and [A]=0 sofeedback also equals 0
In between:Will be maximum when [A*]=[A]
feedback rate = k2 A*[ ] A[ ]
e.g. A* increases activity of Sor phosphorylates A in trans

forward rate =basal rate + feedback rate
forward rate=k1S A[ ] + k2 A*[ ] A[ ]
Consider simple case wherek1S is small so basal rateis negligible
Michaelian system with linear feedback, cont.

Michaelian system with linear feedback, cont.
strong feedback
weak feedback
k2 Atot[ ] > k−1I
k2 Atot[ ] < k−1I
stable ON state butunstable OFF state
stable OFF statebut no ON state
Two steady states:
One steady state:

How can we stabilize the OFF state?
1. Make feedback sigmoidal function of A*
• three steady-states, one unstable (threshold) and two stable
• at threshold, if decrease A*, back reaction is faster than forward reaction and system is driven to OFF state
• at threshold, if increase A*, forward reaction is faster than back reaction and system is driven to ON state

Making feedback sigmoidal function of A*, cont.
i. cooperativity
feedback ∝ A*[ ]n
ii. zero-order ultrasensitivity
Zero-order conditionsrate independent of [substrate]
rate ∝[substrate]First-order conditions
e.g. A* activates S and takes n molecules of A* to activate S

Making feedback sigmoidal function of A*, cont.
ii. zero-order ultrasensitivity, cont.
Opposing reactions with the [substrate] >> Km for the modifying enzymes
First-order
Zero-order
Atot =0.01×Km
Atot =100×Km
First-order: changing p’tase by 1.5x has smalleffect on steady state
Zero-order: changing p’taseby 1.5x changes ss from >90%A* to >90% A
A A*kinase
phosphatase

Making feedback sigmoidal function of A*, cont.
ii. zero-order ultrasensitivity, cont.
• small changes in enzyme activity lead to large changes in modified protein, steady state
• low abundance, high affinity inhibitor of S
• once inhibitor is saturated, S will be effective at causing increases in [A*] (same as S-P below)
iii. inhibitor ultrasensitivity
- inhibitor: 1/9 EC50 causes 10% response90% response at 9 x EC50
+ inhibitor: 10/9 units kinase causes 10% response 90% response at 10 units kinase
81-fold change in kinase 10 to 90%
9-fold change in kinase 10 to 90%

2. Saturate back reaction with A* before feedback saturates
How can we stabilize the OFF state?
• back reaction saturates as A* accumulates
• back reaction can more than keep up with first bit of feedback, but then is overwhelmed as feedback continues to rise

How can we stabilize the OFF state?
2. Saturate back reaction with A* before feedback saturates
1. Make feedback a sigmoidal function of A*
i. cooperativity
ii. zero-order ultrasensitivity
iii. inhibitor ultrasensitivity
Systems are bistable only for a limited range of kinetic parameters

Switching between the OFF and ON states
• continuously increase feedback-independent stimulus S
• bistable system converts continuous change in S into discontinuous change in output (A*)
forward rate =basal rate + feedback rate
basal rate=k1S A[ ]
increase S - increase slope of basal rate

Switching between the OFF and ON states
What happens when we lower the stimulus?
OFF state reappears when S is lowered to 3, but no driving force to leaveON state
HYSTERESIS/IRREVERSIBILITY

Hysteresis and irreversibility
Hysteresis
Any bistable system will exhibit some degree of hysteresis
Path from ON to OFF is different than that from OFF to ON
Irreversibility
System stays in ON state indefinitely after S is removed
Occurs when feedback is strong
Decreases chattering between ON and OFF states when S is near threshold
Mechanism for biochemical memory - unless positive feedback is broken system will remain in ON state - can remember a stimulus long after itis removed

Example 1: Xenopus oocyte maturation
immature
G2 arrested
mature
metaphase arrest,meiosis II
progesterone
MAPK cascade
How does this system turn a continuously varying stimulus (progesterone) into an all-or-none response?
meiosis IGVBD

Xenopus oocyte maturation, cont.
Measure response of oocytes to different [progesterone] by measuringMAPK phosphorylation (activation)
Response of population - treat oocytes with give [progesterone], makeextract, measure MAPK phosphorylation and tabulate % activation
Response isgraded - Michaelian!

Xenopus oocyte maturation, cont.
How do we interpret these observations?
[progesterone] [progesterone]
OR
Answer: Look at the response of individuals

Xenopus oocyte maturation, cont.
Response is all-or-none - no intermediate levels of MAPK activationBISTABLE
Where does this behavior come from?
Clue: Microinjection of activated Mos gives same result (bistability),indicating it comes from pathway downstream of Mos (MAPK cascade)

Xenopus oocyte maturation, cont.
What are the functional consequences of bistability for this system?
Small change in stimulus near threshold throws switch
At low stimulus buffered a bit from change
No intermediate states

Xenopus oocyte maturation, cont.
Can quantitate the sensitivity of the response - for a given change in stimulus, how much change In output (MAPK phosphorylation) results
Michaelian - requires 81-fold change in [ligand] to drive system from 10% to 90% on
Ultrasensitive - requires <81-fold change in [ligand] to drive system from 10% to 90% on
Subsensitive - requires >81-fold change in [ligand] to drive system from 10% to 90% on
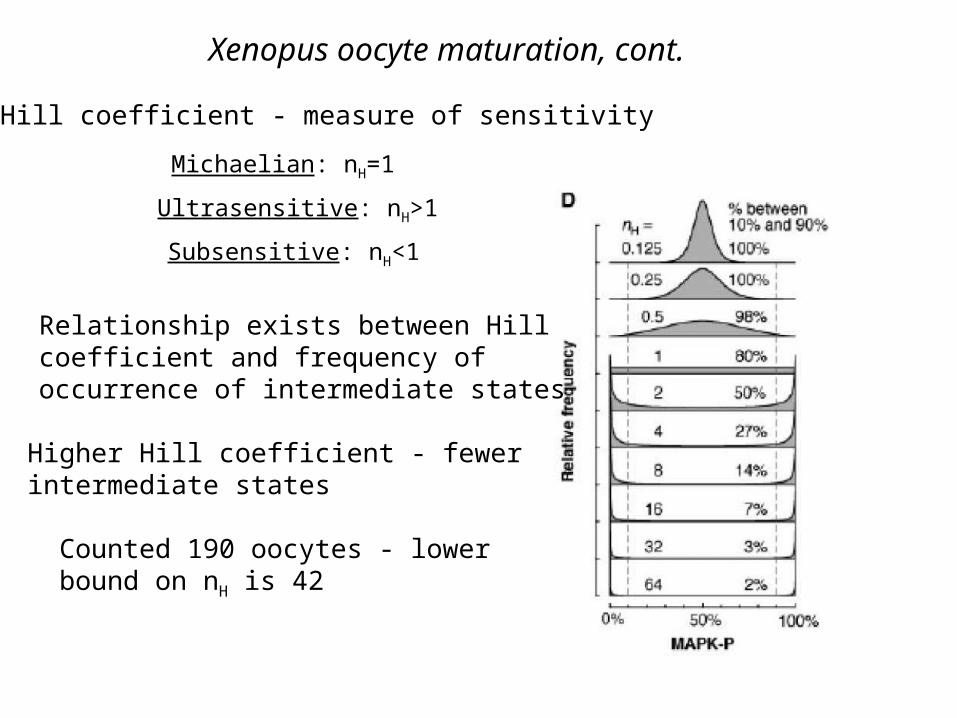
Xenopus oocyte maturation, cont.
Hill coefficient - measure of sensitivity
Michaelian: nH=1
Ultrasensitive: nH>1
Subsensitive: nH<1
Relationship exists between Hillcoefficient and frequency of occurrence of intermediate states
Higher Hill coefficient - fewerintermediate states
Counted 190 oocytes - lowerbound on nH is 42

Xenopus oocyte maturation, cont.
What is the origin of the bistability?
1. Multistep ultrasensitivity - stabilizing the off state
MAPKKMAPKKKà Üà à à àá àà à à à MAPKK-P
MAPKKKà Üà à à àá àà à à à MAPKK-PP
MAPKMAPKKà Üà à à àá àà à à à MAPK-P
MAPKKà Üà à à àá àà à à à MAPK-PP
Two phosphorylation events required for MAPK and MAPKK activation
Activating kinase must dissociate in between phosphorylationevents (to load ATP)
Rate will vary as [upstream kinase]2
Two steps - effects are multiplicative
REMEMBER: Bistability arises from positive feedback anda mechanism to stabilize the off state

Xenopus oocyte maturation, cont.
1. Multistep ultrasensitivity - stabilizing the off state
• forward rate (slope) varies as [kinase]2
• 1 unit of kinase gives 50% S-P; double kinase and rate increases 4-fold
Michaelian Multistep ultrasensitivity
• S-P must increase to higher level than in Michaelian case for rates to balance and reach new steady state (80% vs. 67%)
• leads to sigmoidal stimulus-response curve

1. Multistep ultrasensitivity, cont. - stabilizing the off state
Xenopus oocyte maturation, cont.
Measure dependence of MAPK activation on Mos input in vitro in oocyteextracts
nH ~ 5
~2.5 fold change in MAPKKKchanges MAPK from 10%to 90% activity

2. Zero-order ultrasensitivity - stabilizing the off state
Xenopus oocyte maturation, cont.
• no clear data that reactions in MAPK cascade are operating in zero-order conditions, but concentrations and kinetic properties that have been measured are consistent with this idea
3. Positive feedback
Mos ⏐ →⏐ Mos* ⏐ →⏐ MEK ⏐ →⏐ MAPK
+
• protein synthesis-dependent feedback from active MAPK to Mos
• small change in activity leads to large change in ss position

Xenopus oocyte maturation, cont.
Is the biochemistry underlying oocyte maturation irreversible?
• Is the [progesterone] required to activate MAPK and Cdc2 different from the [progesterone] required to maintain activities?
Induction: incubate withprogesterone, wait untilmaturation plateaus
Maintenance: incubate with600 nM progesterone, waituntil GVBD plateaus, washfor 10 hr, incubate withdifferent [progesterone]
Maturation is irreversible - “memory” of progesterone

Xenopus oocyte maturation, cont.
• based on what we learned about bistability, postulate it is the strength of the feedback that gives rise to irreversibility
• predict that if feedback is disrupted, system should lose irreversibility or “memory”
+E2=MAPK pathway activated
+E2, wash=MAPK pathway activated then 16 hr wash
Disrupt feedback withCHX, Mos antisense
“Memory” requires positive feedback

• also predict that disrupting positive feedback should alter bistability - make less ultrasensitive
• quantitate response of individual oocytes to microinjected Mos in presence and absence of CHX
• observe oocytes with intermediate amounts of phosphorylation - loss of bistability
Xenopus oocyte maturation, cont.
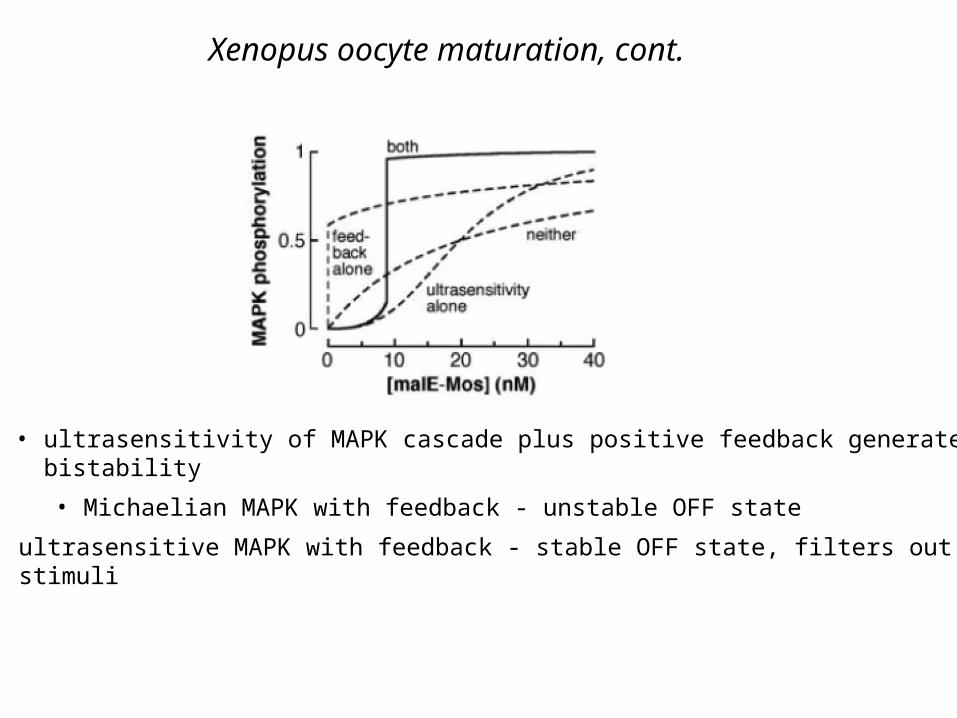
Xenopus oocyte maturation, cont.
• ultrasensitivity of MAPK cascade plus positive feedback generates bistability
• Michaelian MAPK with feedback - unstable OFF state
• ultrasensitive MAPK with feedback - stable OFF state, filters out small stimuli

Lac operon
• beta-galactosidase is formed by bacterial cells grown in the presence of lactose - beta-gal is necessary for metabolism of lactose
• known that non-metabolizable galactosides (e.g. TMG; thiomethyl-galactoside) induce beta-gal formation
• add high [TMG], see induction of beta-gal at maximal induction rate - under these conditions cells respond uniformly
• at lower [TMG], see dose-dependent rate of induction of beta-gal
• cells that have been exposed to a high [TMG] and then shifted to lower [TMG] (maintenance concentrations) continue to synthesize beta-gal at the maximum rate
Phenotypic change following transient signal -memory of inducer!

Lac operon, cont.
Is the Lac system bistable? Cells growing in [TMG]that gives intermediate levelof beta-gal synthesis (e.g. 30%)
Dilute to maintenance [TMG] togive ~1 bacterium/10 tubes
• 10% of the tubes developed bacterial populations
• 30% of the cultures from single bacteria had maximal beta-gal levels; 70% had only small amounts
Population at intermediate [TMG] consists of cells that are fullyinduced and those that are uninduced - BISTABLE

Lac operon, cont.
LacI = Lac repressor
LacY = Lac permease
TMG
TMG
LacI LacY
LacZ
LacZ = beta-galactosidase
• autocatalytic positive feedback (double negative)
• if there is non-linearity in response, might expect bistability in TMG, LacI activity, LacY and LacZ expression

Lac operon, cont.
net rate =d TMG[ ]
dt=vin −vout
vout =vbind +vcatab =kbind TMG[ ] + 0 =kbind TMG[ ]For vin, know ∝ TMG[ ]eBut TMG also indirectly activates entry of TMG through LacI
Response=TMG[ ]i
K + TMG[ ]in
vin = TMG[ ]e k0 +TMG[ ]i
K + TMG[ ]in
⎛
⎝⎜
⎞
⎠⎟
Need k0 parameter because if there is no permease the firstmolecule of TMG cannot enter the cell, independent of the externalconcentration - either operon is not totally repressed or TMG canslowly diffuse
Could come from TMG bindingLacI, relationship between activeLacI and LacY production…

Lac operon, cont.
[TMG]
[TMG]
Three steady-states:ss1, 3 stabless2 unstable
Bistable
vout =kbind TMG[ ]
vin = TMG[ ]e k0 +TMG[ ]i
K + TMG[ ]in
⎛
⎝⎜
⎞
⎠⎟

Lac operon, cont.
Can see bistability and hysteresis in single cell experiments
Take cells expressing Lac promoter-GFP and grow in different [TMG] - either initially uninduced or induced
Uninduced then grown20 hr in 18 M TMG
Bistability and hysteresis
start uninduced
start induced
[maintenance]

Lac operon, cont.
When inducer is added, permease synthesis will be initiated ata rate determined by inducer in medium
At low [inducer] rate of permease synthesis may be so low that probability of making a permease molecule during lifetime of bacteriumis small
Once bacterium has permease, [inducer] inside cell will increase, whichwill increase probability that second molecule of permease will be formed
Two permeases further increases rate of permease - autocatalytic rise tomaximal permease
Once have maximal permease, bacterium and progeny will be inducedindefinitely since [inducer] inside is above maintenance concentration
Maintenance can be explained if the number of permease molecules is largerelative to threshold, there is high probability that each daughter cell willreceive sufficient permease to insure induction
Transfer to less than maintenance - observe exponential decrease in activity,equivalent to constant chance of becoming uninduced - partitioning betweendaughter cells - some will not get enough to insure induction
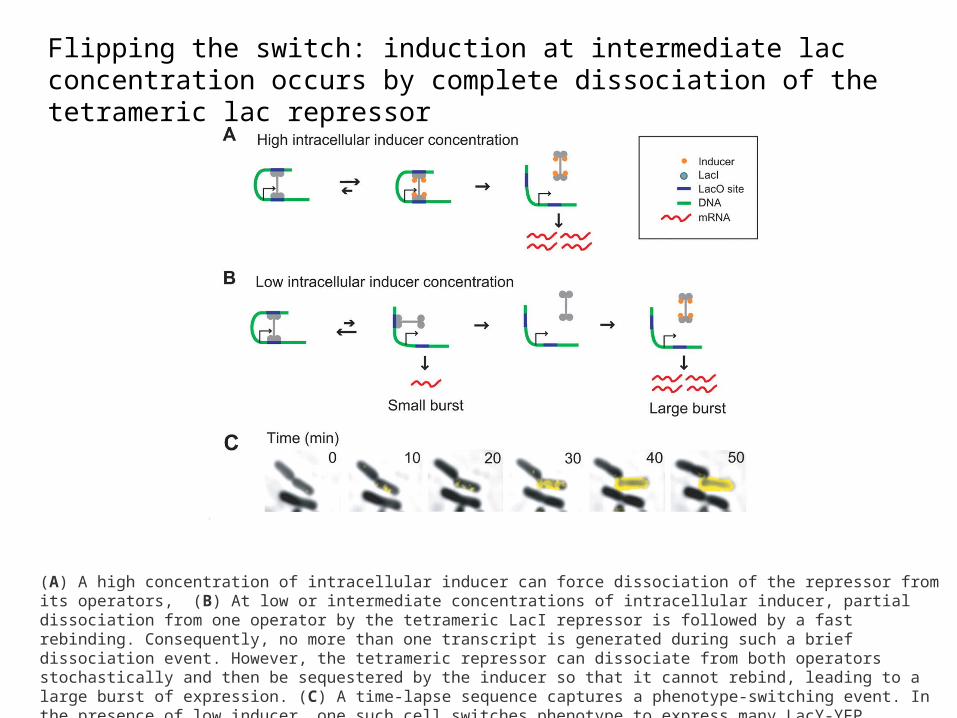
Flipping the switch: induction at intermediate lac concentration occurs by complete dissociation of the tetrameric lac repressor
(A) A high concentration of intracellular inducer can force dissociation of the repressor from its operators, (B) At low or intermediate concentrations of intracellular inducer, partial dissociation from one operator by the tetrameric LacI repressor is followed by a fast rebinding. Consequently, no more than one transcript is generated during such a brief dissociation event. However, the tetrameric repressor can dissociate from both operators stochastically and then be sequestered by the inducer so that it cannot rebind, leading to a large burst of expression. (C) A time-lapse sequence captures a phenotype-switching event. In the presence of low inducer, one such cell switches phenotype to express many LacY-YFP molecules (yellow fluorescence overlay) whereas the other daughter cell does not

Bistability Biobliography Reviews & Comments on Research Articles:
Ferrell, J. E., Jr. (1996). Tripping the switch fantastic: how a protein kinase cascade can convert graded inputs into switch-like outputs. Trends Biochem Sci 21, 460-6.
Koshland, D. E., Jr. (1998). The era of pathway quantification. Science 280, 852-3.
Koshland, D. E., Jr., Goldbeter, A., and Stock, J. B. (1982). Amplification and adaptation in regulatory and sensory systems. Science 217, 220-5.
Laurent, M. and Kellershohn, N. (1999) Multistability: a major means of differentiation and evolution in biological systems. Trends Biochem. Sci. 24, 418-422. Research articles:
Novick, A. and Weiner, M. (1957). Enzyme induction as an all-or-none phenomenon. Proc. Natl. Acad. Sci. U S A 43, 553-566.
Cohn, M. and Horibata, K. (1959). Analysis of the differentiation and of the heterogeneity within a population of Escherichia coli undergoing induced beta-galactosidase synthesis. J. Bacteriology 78, 613-633. Ferrell, J. E., Jr., and Machleder, E. M. (1998). The biochemical basis of an all-or-none cell fate switch in Xenopus oocytes [see comments]. Science 280, 895-8.
Goldbeter, A., and Koshland, D. E., Jr. (1981). An amplified sensitivity arising from covalent modification in biological systems. Proc Natl Acad Sci U S A 78, 6840-4.
Goldbeter, A., and Koshland, D. E., Jr. (1984). Ultrasensitivity in biochemical systems controlled by covalent modification. J. Biol. Chem. 259, 14441-14447.
Huang, C. Y., and Ferrell, J. E., Jr. (1996). Ultrasensitivity in the mitogen-activated protein kinase cascade. Proc Natl Acad Sci U S A 93, 10078-83.
Xiong, W. and Ferrell, J. E., Jr. (2003). A positive-feedback-based bistable 'memory module' that governs a cell fate decision. Nature 426, 460-465.
Ozbudak, E. M., Thattai, M., Lim, H. H., Shraiman, B. and van Oudenaarden, A. Multistability in the lactose utilization network of Eschericia coli. Nature 427, 737-740. Tanaka M, Collins S, Toyama B, Weissman JS. (2006). The physical basis of how prion conformations determine strain phenotypes. Nature 442, 585-589.
Choi PJ, Cai l,Freda K, Xie XS (2008) A stochastic single gene event triggers switching of a bacterial celll. Science 332. 442-446.