synthesis and degradation of surface structures by growing and non-growing bacillus megaterium
TRANSCRIPT

Synthesis and Degradation of Surface Structures by Growing and Non-growing Bacillus megaterium J. CHAL()IYPK. a.
Insti tute of Microbiology, Czechoslovak Academy of Sciences, Prague 4
A bacterial cell growing and multi- plying logarithmically in a defined me- dium is characterized by a distinct size, chemical composition and biochemical activity (Ecker & Schaechter, 1963). DNA, proteins, ribosomal and transfer RNA are mostly stable in growing populations, messenger RNA shows con- tinuous and rapid turnover (see Mandel- stare, 1960, 1963; Neihardt 1963).
A change in the composition of the medium, caused either by an experiment- ing microbiologist or by the micro- organism itself causes a series of changes in the bacterial cell. The rates of synthesis of different cell constituents adapt to the new conditions and the degradation of some cellular components (proteins and RNA) can take place. The controversial data were published about the syn- thesis of structural components in bacteria during this adaptation period (Mandelstam & Halvorson, 1960; Pine, 1965; Schlesinger & Ben-Hamida, 1966).
The presented experiments deal with the study of the synthesis and degrada- tion of surface components of Bacillus megaterium KM -- cell wall mucopeptide and proteins in the membrane fraction -- under different physiological conditions.
We found some years ago that unlike most cellular constituents, the cell wall mucopeptide, is continuously degraded in growing populations and its degrada- tion products can be partially reassimi- luted (Chaloupka, K~e6kov~ & l~ihov~, 1962; Chaloupka, l~ihov~ & K~eSkov~, 1964).
We repeated the experiments using a different technique with similar results (Fig. 1).
The results show that the loss of 14C DAP indicating the degradation of mueo- peptide proceeds by the rate of 10--15 ~o generation which is in agreement with the data obtained by a different tech- nique in previous experiments. A ]urge portion of the degradation products of the cell wall is not reutilized, as the loss of the radioactivity proceeds at a similar rate in the presence or absence of 1~C DAP.
When the culture undergoes shift down (transfer from NH 4 -->NO8 as a source of N) no apparent degradation of mucopeptide was observed during the lag (Fig. 2).
On the contrary, the addition of the mixture of amino acids to the mineral salts -t- glucose medium (shift up) causes an enhanced rate of cell wall degradation.
The differential plot of the loss of the radioactivity against the increase of dry weight (Fig. 3) shows a similar slope in all cases. The lag at the beginning is probably due to a slow equilibration of external and pool DAP.
Unlike the mucopeptide rigid layer the protein component of cytoplasmic membrane fraction is stable in growing culture and does not undergo turnover to a measurable extent (Fig. 4).
During starvation in the nitrogen-free medium the degradation of proteins in Bacillus megaterium proceeds at the rate of approx. 3 per cent/hour and the rate

1967 T U R N O V E t r OF C E L L U L A R S U R F A C E S T R U C T U R E S 265
0.8'
0.7.
0.6,
0.s, "o
~.. 0.4- O
0.3-
0.2
0.1
+'2C DAP OD
J TCA insol.
-800
- 250
- 20O
o u '10O
'50
0 60 120 180 2~0 0 60 120 180 240 T,me(min)
Fig. 1. The cu l ture g rowing in t he mi ne ra l s a l t s + glucose m e d i u m was labe l led du r ing one a n d one h a l f generat ion~ w i th 14C D A P in preser~ce o f x~C lys ine ~nd af te r w a s h i n g t r a n s f e r r e d rote a f resh nonrad ioac t ive m e d i u m con ta in ing l~C D A P + t i c lys ine or x~C lys ine on ly (5 x 10-* M each). T h e r ad ioac t iv i ty of t h e cu l tu re in t h e c o n s t a n t v o l u m e of m e d i a was d e t e r m i n e d b y t h e m e m b r a n e fi l ter t echn ique . � 9 optical dena i ty ; (D - - ho t TCA insoluble coun t s ; CO - - cold TCA insoluble coun t s .
1.40
1.20'
":~ 1,o0, E
0.80'
~ 0.6~-
c~ 0.40,
0.20'
J NO3 l 350
. . . . . . if: 0 ~ 60 ~ 120 IS0 I~ 0 30 60 90 120 150 180
Time(rain) Fig. 2. The cells p re labe l led w i t h 14C D A P in NH4, sa l t s -~- glucose m e d i u m in t h e s ame w a y as in t h e p rev ious e x p e r i m e n t , were t r an s f e r r ed in to t h e m e d i u m con ta in ing ca,~aminoacids (CAA), N H 4 or NO z as a source o f n i t rogen , s u p p l e m e n t e d w i t h m i n e r a l s a l t s + glucose, T h e opt ica l d e n s i t y (OD) sacl r~d ioac t iv i ty in TC& ir~,~oluble fraction w a s de t e rmined .

266 J. CHALOUPKA Vol. 12
of degradation of "membrane proteins" was not greater than that of supernatant proteins (Hafiz & Chaloupka -- prepared for publication). As the synthesis and secretion of the extracellular proteinase
over of proteins in the membrane fraction probably takes place to a measurable extent only in non-growing cells.
The synthesis of structural components was studicd in culture growing in bal-
350
300
E
s
u 250,
t | ! t �9 �9
0.20 0.40 0.60 0.80 1.00 1.20 1.40
Dry weight (mg) Fig. 3. The differential plot of the s4C DAP content in cell walls against the increase in dry weight.
takes place either during growth or during nitrogen starvation (Chaloupka & K~e~kovs 1966), it does not appear likely that the formation of an extra- cellular enzyme would be enabled by the enhanced rate of protein turnover in the cytoplasmic membrane.
To conclude this par t of the work I would like to stress that the two surface layers of the cell probably differ in the metabolic stability. The turnover of mucopeptide in the rigid cell wall seems to be characteristic for the growing populations of Bacillus megaterium. Turn-
anced state as well as in populations under- going shift down or up. The rate of synthesis of cell wall mucopeptide is inversely proportional to the growth rate or the rate of incorporation of 1~C leu into the proteins (Fig. 5). The similar results were obtained with a different method by Sud and Schaechter (1964).
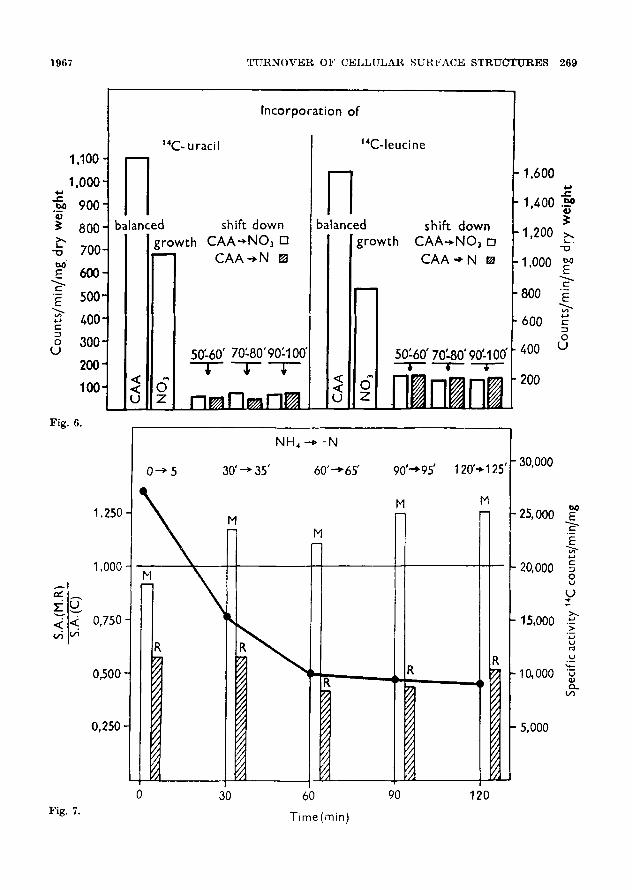
The shift down from CAA to NO3 (or nitrogen free) medium causes the de- crease in the rate of synthesis of protein and R N A to approx 10- -15% of the preshift rate (Fig. 6). The decrease of the rate of synthesis of protein and R NA

1967 T U R N O V E R OF C E L L U L A R S U R F A C E S T R U C T U R E S 267
was similar in nitrogen free as well as in nitrate medium�9 I t is, therefore, posible to conclude that the synthetic activities during the lag phase in the shift down medium are probably exclusively due to the turnover of the respective compo- nents.
To establish the rate of synthesis o f structural proteins the distribution of radioactivi ty in the cells during the lag after shift down t rea tment was followed.
The shift down cells labelled with 10 minutes pulses of 14C leucine were fractionated in order to establish the
1.000 -0.500
0.800
theoretical (no turnover)
0.400
0.600,
0.400
0.200
- 0.200
' ' , , ; ,
0 2 3 4 6 Time (hours)
E
E
~J
t~
Fig. 4. The cells were pre labe l led w i t h =sS m e t h i o n i n e a n d cu l t i va t ed in ~ s h a k e n cu l tu re in t h e n~nera I sai ls ~ glucose -{- *=S m e t h i o n l n e con ta in ing m e d i u m , for more t h a n 3 genera t ions �9 The g rown ceils t a k e n f rom t h e cu l ture were conver t ed to p ro top las t s . The specific r ad ioac t iv i ty of p ro te in in m e m b r a n e f rac t ion ( isolated a f te r d i s rup t ion o f p ro top l a s t s by m e a n s o f t h e osmot ic shock) a n d in t h e s u p e r n a t a n t ( c y ~ p l a s m i c ) f rac t ion was de t e rmined . S.A. - - specific r ad ioac t iv i ty ; t m - - t ime a t wh ich t h e s am p l e was t a k e n ; to - - zero t ime ; C - - cy top la smic f r ac t ion ( s u p e r n a t a n t a t 105,000• g); M - - m e m b r a n e f rac t ion (pellet a t 15 ,000• g); � 9 d r y we igh t (mg/ml) ; O - - t h e o r e - t ica l decrease o f specific r a d i o a c t i v i t y due on ly to t h e d i lu t ion w i th non- labe l l ed proteins �9

268 J . C H A L O U P K A Vol. 12
2,500
2,000
E
E 1,500
r
o U m 1,000
O
500 -
I
0
'"C-leu O CAA
[3 NO3
0 0
T T T
0
0
i !
[]
@
1,000
E
800 F:
e ' -
- 600 o U
<
4O0 C~ d
200
'4C-DAP �9 CAA
�9 NOj
I
2O I i | l i t I I
40 6O 80 1 O0
Time(rain) Fig. 5. T he s amp l e s t a k e n in d i f ferent t ime f rom cul ture g rowing logar i th- mica l ly in CA-& or NO s + mine ra l s a l t s -t- glucose m e d i u m were i ncuba t ed for 10 rain, w i t h e i the r 1'C leu or 1'C D A P . The .specific r ad ioac t iv i ty in h o t TCA insoluble raa ter ia l (in t h e case of 140 DAP) or in p ro te in (in t h e case o f 1'C leucine) was d e t e r m i n e d . C A A - c a s a m i n o acids.
Fig. 6. The cu l tu re growing loga r i t hmica l ly in CA2, -(- s a l t s -k glucose m e d i u m was w a s h e d a n d r e s n s p e n d c d in NO t + sa l t s -b glucose, or in sa l t s + glucose only. The incorpora t ion of 1'C leucine or 140 urac i l into the respec t ive s amp l e s proceeded for 10 m i n u t e s . The r ad ioac t iv i ty of TCA insoluble m a t e r i a l was t h e n d e t e r m i n e d . CA-& - - c a s a m i n o acids ; NO a - - n i t r a t e m e d i u m , N -- ni- t r o g e n free m e d i u m .
Fig. 7. The labe l led c e l l s were conve r t ed to p ro top l a s t s in t h e sucrose m e d i u m con t a in ing chlor amphen i co l a n d an excess of non rad ioac t ive leucine a n d a f te r w a s h i n g lysed b y t h e osmot i c shock" The l y sa t e was cen t r i fuged a t 15 ,000• g for 30 m i n u t e s a n d t h e sup for 90 rain a t 105,000X g. The TCA insoluble ma te r i a l was e x t r a c t e d b y lipophill ic so lven t s a n d t h e n d iges ted w i t h a sma l l a m o u n t (1 r ag /50 - -200 m g prote in) of t r y p s i n to solubilizo t h e r a t h e r insoluble s t r uc tu r a l pro te ins . I n t h e clear so lu t ion t h e specific r ad ioac t iv i ty was de te rmined . S.&. - - specific ac t iv i ty ; M - - m e m b r a n e f rac t ion; 14 - - r i bosoma l f rac t ion; C - - c y t o p l a s m . T he co l umns r e p r e s e n t t he ra t io of s. a. m e m b r a n e (ribo- somal) p ro t e in s to t h a t of so luble pro te ins .
1967 TUI~NOVEI~ OF CELLULAI~ S U R F A C E S T R U C T U R E S 269
'4C- u racil I ,I00
1,0OO
.~ 900
800 balanced
-o 700 t~O B 6OO
-~- 50o
., 400- t -
O 300" u 2OO
100
1,250
Fig. 6.
1,000
<. 0,750
o~soo,
Fig. 7.
Incorporation of
0,250-
growth
<< 2
shift down C A A + N 0 3 []
CAA -~ N
50'-60' 70"80' 90:100' - - . ~ --~-- .-~-.-
'4C-leucine
I balanced shift down
g rowth C A A ~ N O , rn CAA "+ N []
50'-60' 70'-80' 90:100'
NH4 "~ -N
0"-*5 30'--*35'
M
0 30 60
60'-~65" 120"t* 125'
M
90"--~95'
M
R
1
H
R R
1 90 120
Time (rnin)
1,600
1,400 .-~
1,200
b.O 1,000 E C
800 ~
600 = 0
400 U
200
30,000
~,0 25,000 E
20,000 = 0 u
15,000 .~ >
~o, ooo ~
5,000
270 J . C H A L O U P K A Vol. 12
~.ooo -
NO]oNO~
,~, 2,ooo- i
c
~ l . ~ . c
U
$0()-
O ~ m 4.~55'
CAA ~CAA NOpCAA CAA~NOj
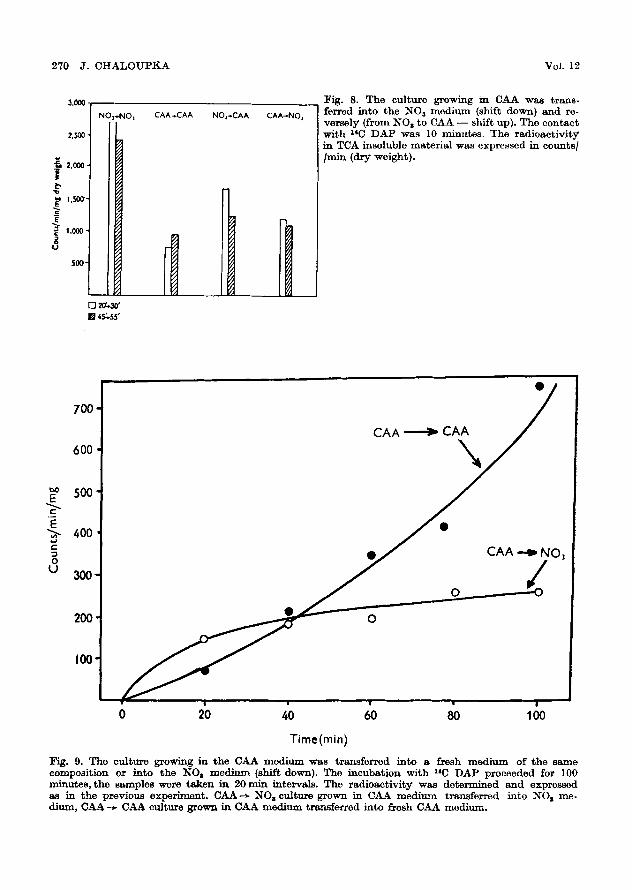
Fig. 8. T h e cu l tu re g rowing in CAA was t r ans - ferred in to t h e NOa m e d i u m (shif t down) a n d re- ve r se ly ( f rom NOs to CAA - - sh i f t up). T h e c o n t a c t with 26C DAP was l0 minutes. The radioactivity in TCA insoluble material was expressed in counts/ /rain (dry weight).
t~O
E e-
E
3 o U
700-
600
500'
400
200 O
C A A ~ C A A
\
O
CAA ~ NO3
�9 / | �9 �9 I
0 20 40 60 80 100
Time(min)
Fig. 9. T h e cu l tu re growing in t h e C AA m e d i u m was t r ans fe r r ed in to a f resh m e d i u m o f t h e s a m e compos i t ion or in to t h e NO s m e d i u m (shif t down), ~r~.o i n c u b a t i o n w i t h 21C D A P proceeded for 100 m i n u t e s , t h e s amp le s were t a k e n in 20 ra in in te rva ls . T he r ad ioac t iv i ty was d e t e r m i n e d a n d expressed as in t h e p rev ious expe r i men t . C A A - ~ N O s cu l tu re g rown in CA_4. m e d i u m t r ans fe r r ed in to •O I me - d ium, CAA -~ CAA cul ture g rown in C A A m e d i u m t rans fe r red in to f resh CAA m e d i u m .

specific radioactivity of proteins in dif- ferent structural components.
Fig. 7 shows that the incorporation into ribosomal fraction was decreased to about 50% in comparison with the
600
down later. The shift down cells thus behave similarly with the cells in the beginning of the stationary phase caused by the amino acid deprival (Shockman, 1963, 1965).
200.
500
F: 400
~> 300. u
100' / . '4C- DAP
o 1o 2'o 3'o 4'0 ;o io
balanced growth
0 C/G �9 C/L
'"C- DAP
, ~ w
,0 10 20 30 40
shift down C//G ~ C/L 0
1967 TURNOVER OF CELLULAR SURFACE STRUCTURES 27I
Time(min)
0.40
0.30
0.20
5- 0.I0 ~:
0.00 ~ O
Fig. 10. The culture grew in balanced state in the synthetic C medium containing mineral salts + glucoso'~ (lactose) - - left side, or was transferred from the glucose into lactose medium (shift down) - - r i g h t side. The incorporation of 14C DAP proceeded for 40 min and the rate of synthesis of mucopeptide was followed by the) membrane filter technique. C/G - - C medium containing glucose; C/L - - C medium containing lactose.
membrane or supernatant (cytoplasmic) proteins. This ratio was approx, constant during 2 hours. In the culture growing logarithmically, the specific radioactivity is the same, in these three fractions.
Unlike the marked decrease of the rate of synthesis of R~TA or proteins after shift down, there was no inhibition of the incorporation of 14G DAP into the cell wall under similar conditions (Fig. 8 and 9).
It seems that the synthesis of the mucopeptide proceeds at a relatively high rate at the beginning of the lag phase of the shift down state, but slows
Almost no incorporation of 14C DAP was found, however, when the shift down~ was performed by the transfer of the cells from mineral salts + glucose into mineral salts -~ lactose medium (Fig. 10)
Under the conditions of the shift up (NO3 ~ CAA - - F i g . 8) the rate of in- corporation of 1~C DAP decreases, which:~ is in agreement with the previous finding that the rate of synthesis of cell wall mucopeptide in Bacillus megaterium is inversely proportional to the rate of synthesis of proteins. The incorporation of 14C leucine into protein in different

272 J . C H A L O U P K A Vol. 12
cell fractions in shift up culture is shown i n Fig. 11.
5,000.
o '4.000-
r~ 3,000-
2,000
1,000-
shift up NO~-CAA
NO3_NO ~ 2Z'12' 10~20'
CMR
Fig. 11. The cu l ture growing in t h e NO s % sa l t s + g lucose m e d i u m was en r i ched w i t h 1 m g / m l o f CAA. T h e two 10 rain pu lses of 1~C leu were ad- mlnis tmred on 2 a n d t0 rain a f t e r t h e add i t ion of CAA. T h e f r ac t iona t ion was p e r f o r m e d by t h e s a m e procedure as m e n t i o n e d in fig. 6 (C, M, R see fig. 7).
R, o f e r : e n c e s
C h a l o u p k a , J . , K~eSkovh, P. , ~ i h o v h , L.: The mucopeptide turnover in the cell walls of growing eulCures of Bacillus megaterium K M . E x p e r i e n t i a 18 : 362, 1962.
C h a l o u p k a , J . , K~eSkovA, P.: Regulation of the formation of protease in Bacillus megaterium. I . Tha influence of ami~m-acids o~ the enzyme fo~mt ion . Fol . microbiol . 11 : 82, 1966.
Cha loupka , J . , ~ i h o v h , I . , K~eSkovg~, P. : Degrada. tion and turnover of bacterial cell wall mucopepbides in growing bacteria. Fol. microbiol . 9 : 9, 1964.
:Ecker, R . E . , Schaechte r , M.: Bacterial growth under conditions of limited nutrition. Ann. N. Y . Aead. Sci; 102 : 549, 1963.
Mar~delstam, d. : The intracellular turnover of protein and nucleic acids and its role in biochemical differentiation. Bacteriol . Rev . 24 : 289, 1960.
The immediate increase in the specific radioactivi ty of ribosomal proteins was expected as the shift up stimulates pre- ferentially the synthesis of ribosomal RNA.
At the present t ime it is not possible to tell whether the increased specific radioactivity in the membrane fraction is due to the synthesis of some structural membrane proteins or to the synthesis of ribosomes firmly a t tached to the membrane (Nielsen & Abrams, 1964).
To summarize the results: The rate of incorporation of 1r DAP
into the mucopeptide is inversely pro- portional to the rate of synthesis of proteins or growth. The shift down caused by limiting the availability of nitrogen does not substantially decrease the synthesis of mucopeptide at the be- ginning of the lag. The decrease of the rate of synthesis of membrane and supernatant proteins equals, the syn- thesis of ribosomal proteins is inhibited to a greater extent. However, if the shift down is due to a change in the carbon source, the incorporation of 14C DAP is immediately inhibited. Under the state of shift up some decrease in the rate of the synthesis of the cell wall was found. The rate of synthesis of ribosomal proteins and protein fraction sedimenting with membranes is enhanced under these conditions.
M a n d o l s t a m , J . : Protein turnover and its function in the economy of the cell. Ann . N.Y. Acad. Sci. 102 : 621, 1963.
M a n d e l s t a m , J . , t I a lvorson , I t . O.: Turnover of protein and nucleic acid in soluble and ribosome fractions of non-growing Eseherichia coli. Biochem. b iophys . A c t s 40 : 43, 1960.
N e i d h a r d t , F. C.: Effects of environment on the com- position of bacterial cells. Ann. Rev . Microbiol. 17 : 61, 1963.
NieLsen, L. , A b r a m s , A.: Rapidly labelled ribosomal R N A associated with membrane ghosts of Strepto- coccus ]aecalis. }3iochcm. b iophys . Res . C o m m u n . 17 : 680, 1964.
P ine , M. J . : Heterogeneity of protein turnover in Escherichia coll. Biochim. b iophys . A c t s 104 : 439, 1965.

1967 T U R N O V E R OF C E L L U L A R S U R F A C E S T R U C T U I 4 E S 273
Sehlesinger , D. , B e n . H a m i d a , F. : Turnover of pro- tein in Escherichia cali starving for nitrogen. Bioch im. b iophys . A c t a 119 : 171, 1966.
S h o c k m a n , G. D.: Amino acid deprivation and bacterial cell wall syntheM~. Trans . ~ . Y . Acad. Sci; 26 : 182, 1963.
S h o c k m a n , C. D. : Symposium on the fine structure and replication of bacteria and their parts. I V . Unbalanced cell.wall synthes~s. Autolysis and cell wall thickening. Bacter io l . R ev . 2 9 : 3 4 5 , 1965.
Sud, I. J . , Schaech te r , .~,L: Dependence el the content of cell envelopes on the growth rate el Bacillus megaterlum. J . Bacter iol . 88 : 1612, 1964.
Discussion Pelzer: One of y o u r first s d e m o n s t r a t e d
t h a t in a chase expt~riment t h e 14C-DAP c o n t e n t of t h e TCA-precip i table m a t e r i a l in ' t h e growing cu l tu re is d imin i shed w i t h t i me . H a v e y o u eve r s t u d i e d t h e chemica l n a t u r e of t h e ~4C-DAP, w h i c h h a s become TCA-soluble in t h i s w a y ?
Chaloupka: No, t h e d e g r a d a t i o n p roduc t s c an however be d iges ted to a m i n o acids, as t h e D A P f rom t h e m u c o p e p t i d e c a n be d e e a r b o x y l a t e d to lys lne.
World: I was n o t sure ff you d id all you r experi- m e n t s in t h e def ined m e d i a w i t h a d d e d lysine. This wou ld p a r t l y e l imina t e derepress ion o f D A P dc- ca rboxy lase on t r ans fe r to s imple r med ia , b u t t he r e
is a poss ib i l i ty t h a t c a s a m i n o acids m a y repress f u r t h e r duo to o t h e r a m i n o acids. For examph~, Dr, ~Vhite found t h a t ty ros ine , t r y p t o p h a n antl pheny l a l an ine repressed D A P dcca rboxy la se i~ Escherichia coll. I d o n ' t k n o w if t h e y wou ld h a v e t h i s effect w i th y o u r s t r a i n o f megaterium. Did yo~t e x a m i n e t h e r ad ioac t iv i t y o f t h e lys ine in yo~tr TCA prec ip i ta tes ?
Ghaloupka: All e x p e r i m e n t s were pe r fo rmed iu t h e presence of non- rad ioac t ive lysine. The dis t r ibu- t ion o f r ad ioac t iv i t y be tween D A P a n d lysine iu TCA prec ip i ta tes was checked by m e a n s o f pape r c h r o m a t o g r a p h y . The cu l tu re g rown o v e r n i g h t i~t t h e presence of ~ C - D A P -~- ~C lysine con t a ined 5 - -10~o of r ad ioac t iv i ty in t h e lys ine spot , 90--95(.)/~ i n t h e D A P .
Sehueffer: The sh i f t -down t r ans f e r f rom N-H a --~ to NOs m e d i u m , w h i c h w i t h t h e (asporogcnouQ K~I s t r a i n of Bacillus megaterium l eads to a re- s u m p t i o n of g r o w t h a f te r a 60 m i n u t e lag, as we h a v e j u s t l ea rned , g ives a ve ry di f ferent resu l t w h e n appl ied to t h e (sporogenous) M a r b u r g strail l of Bacillus subtilis: t h e lag obse rved is v e r y mu , :h longer, t h e popu la t i on is m a s s i v e l y induced t,) sporu la t e a n d a special t y p e o f S p o m u t a n t (th'~ " h y p e r repressed" type) is s e l ec ted for. I t is n o t clear w h e t h e r t h e difference in t h e response of t h e two s t r a i n s is due to t h e k n o w n difference in spor(~- fo rming abi l i ty be tween t h e two s t r a in s , or t~) some o t h e r s t r a i n difference.