supplementary,information ... · supplementary,information,,...
TRANSCRIPT

SUPPLEMENTARY INFORMATION Integrated genomic characterization of adrenocortical carcinoma
Guillaume Assié* 1,2,3,4, Eric Letouzé* 5, Martin Fassnacht6,7,8, Anne Jouinot1,2,3, Windy Luscap1,2,3, Olivia Barreau1,2,3,4, Hanin Omeiri1,2,3, Stéphanie Rodriguez1,2,3, Karine Perlemoine1,2,3, Fernande René-‐Corail1,2,3, Nabila Elarouci5, Silviu Sbiera6,7, Matthias Kroiss8, Bruno Allolio7, Jens Waldmann9, Marcus Quinkler10, Massimo Mannelli11, Franco Mantero12, Thomas Papathomas13, Ronald De Krijger13, Antoine Tabarin14,15, Véronique Kerlan15,16, Eric Baudin15,17, Frédérique Tissier1,2,3,18, Bertrand Dousset1,2,3,4,19, Lionel Groussin1,2,3,4, Laurence Amar20, Eric Clauser21, Xavier Bertagna1,2,3,4,15, Bruno Ragazzon1,2,3 ,Felix Beuschlein6, Rossella Libé1,2,3,4,15, Aurélien de Reyniès° 5, Jérôme Bertherat° 1,2,3,4,15. * contributed equally with their respective expertise to this study ° jointly directed this work with their respective expertise 1-‐ Institut National de la Santé et de la Recherche Médicale U1016, Institut Cochin, Paris, France 2-‐ Centre national de la recherche scientifique UMR8104, Paris, France 3-‐ Université Paris Descartes, Sorbonne Paris Cité, Paris, France 4-‐ Center for Rare Adrenal Diseases, Department of Endocrinology, Assistance Publique Hôpitaux de Paris, Hôpital Cochin, Paris, France 5-‐ Programme Cartes d’Identité des Tumeurs (CIT), Ligue Nationale Contre Le Cancer, Paris, France 6-‐ Medizinische Klinik und Poliklinik IV, Klinikum der Universität München, University of Munich, Munich, Germany 7-‐ Department of Internal Medicine I, Endocrine and Diabetes Unit, University Hospital of Würzburg, Würzburg, Germany 8-‐ Comprehensive Cancer Center Mainfranken, University of Würzburg, Würzburg, Germany 9-‐ Department of Surgery, Philipps-‐University Marburg, Marburg, Germany, 10-‐ Department of Clinical Endocrinology, Charité Campus Mitte, Charité University Medicine, Berlin, Germany 11-‐ Department of Experimental and Clinical Biomedical Sciences, University of Florence, Florence, Italy 12-‐ Endocrinology Unit, Department of Medicine, University of Padova, Padova, Italy 13-‐ Department of Pathology, Josephine Nefkens Institute, Erasmus MC-‐University Medical Center, Rotterdam, Netherlands 14-‐ Department of Endocrinology, Diabetes and Metabolic Diseases, University Hospital of Bordeaux, Bordeaux, France 15-‐ Rare Adrenal Cancer Network COMETE-‐, Paris, France 16-‐ Department of Endocrinology, Diabetes and Metabolic Diseases, University Hospital of Brest, Brest, France 17-‐ Department of Nuclear Medicine and Endocrine Oncology, Institut Gustave Roussy, Université Paris-‐Sud, Villejuif 18-‐ Department of Pathology, Assistance Publique Hôpitaux de Paris, Hôpital Pitié-‐Salpétrière, Pierre et Marie Curie University, Paris, France 19-‐ Department of Digestive and Endocrine Surgery, Assistance Publique Hôpitaux de Paris, Hôpital Cochin, Paris, France 20-‐ Hypertension unit, Assistance Publique Hôpitaux de Paris, Hôpital Européen Georges Pompidou, Paris, France 21-‐ Department of Oncogenetics, Assistance Publique Hôpitaux de Paris, Hôpital Cochin, Paris, France
Nature Genetics: doi:10.1038/ng.2953

List of Supplementary Figures and Tables Supplementary Figure 1. SNP patterns frequently observed in ACC. Supplementary Figure 2. Coverage of exome sequencing. Supplementary Figure 3. Mutation rates and types of somatic substitutions in 45 ACC. Supplementary Figure 4. Location of mutations in key genes recurrently mutated in ACC. Supplementary Figure 5. Activation of ß-‐catenin target genes in tumors harboring ZNRF3 inactivating alterations. Supplementary Figure 6. DNA methylation-‐based classification of ACC. Supplementary Figure 7. mRNA expression-‐based classification of ACC. Supplementary Figure 8. Volcano plot analysis of differentially expressed miRNAs in each tumor cluster. Supplementary Figure 9. Expression of MEG3 across 65 normal human tisses. Supplementary Table 1. Clinical data. (a) Summary of clinical data in the discovery and validation cohorts. (b) Sample characteristics and analyses performed. Supplementary Table 2. Significantly recurrent broad (a) and focal (b) copy number changes identified by GISTIC2.0 analysis. Supplementary Table 3. Homozygous deletions (a) and high-‐level amplifications (b) identified by SNP array in 121 ACC. Supplementary Table 4. List of somatic nonsynonymous mutations identified by exome sequencing of 45 ACC (provided as an Excel file). Supplementary Table 5. MutSigCV analysis of mutation significance (top-‐20 genes). Supplementary Table 6. Mutations identified by targeted sequencing in the validation cohort of 77 ACC. Supplementary Table 7. Summary of mutations and copy-‐number alterations affecting candidate driver genes in the entire cohort of 122 ACC. Supplementary Table 8. Comparison of driver genes alteration frequencies in the discovery and validation cohorts.
Nature Genetics: doi:10.1038/ng.2953

Supplementary Table 9. Significantly mutated gene sets. Supplementary Table 10. Analysis of the alternative lengthening of telomeres (ALT) phenotype in 20 ACC. Supplementary Table 11. Multi-‐omics classification of ACC samples. Supplementary Table 12. Significantly deregulated miRNAs in ACC subgroups Mi1 (a), Mi2 (b) and Mi3 (c).
Nature Genetics: doi:10.1038/ng.2953
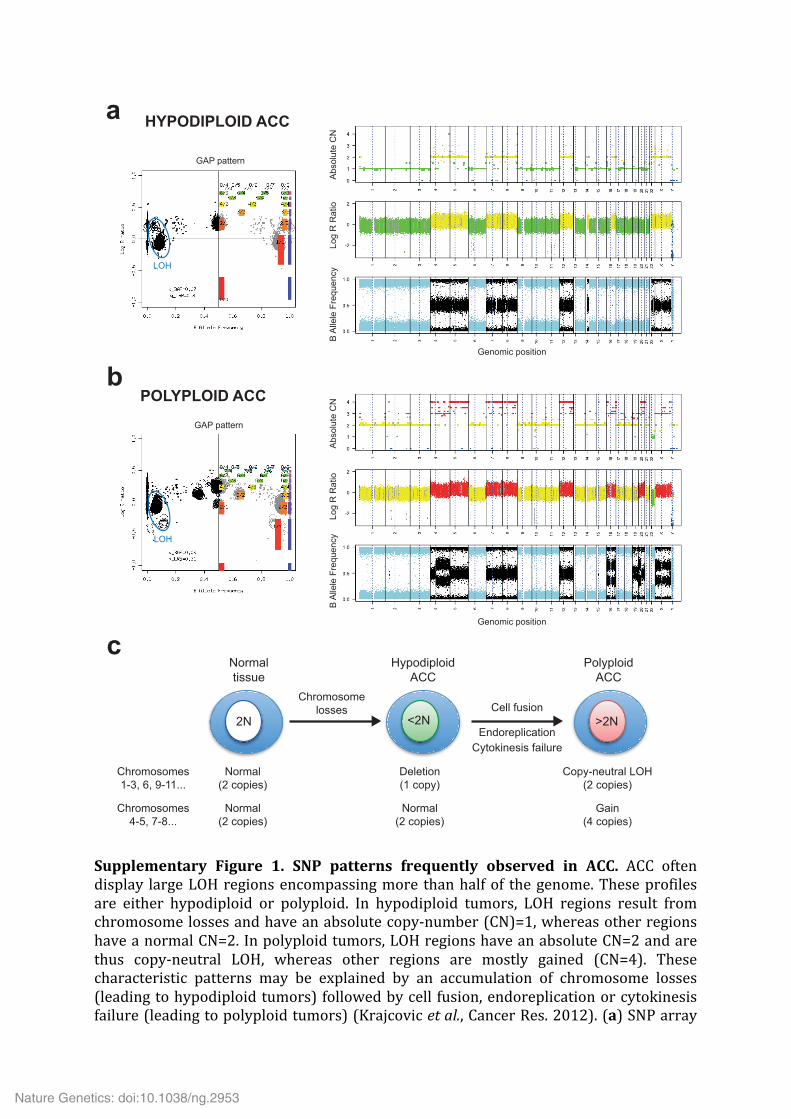
Supplementary Figure 1. SNP patterns frequently observed in ACC. ACC often display large LOH regions encompassing more than half of the genome. These profiles are either hypodiploid or polyploid. In hypodiploid tumors, LOH regions result from chromosome losses and have an absolute copy-‐number (CN)=1, whereas other regions have a normal CN=2. In polyploid tumors, LOH regions have an absolute CN=2 and are thus copy-‐neutral LOH, whereas other regions are mostly gained (CN=4). These characteristic patterns may be explained by an accumulation of chromosome losses (leading to hypodiploid tumors) followed by cell fusion, endoreplication or cytokinesis failure (leading to polyploid tumors) (Krajcovic et al., Cancer Res. 2012). (a) SNP array
GAP pattern
Absolute CN
Log R Ratio
B Allele Frequency
GAP pattern
Absolute CN
Log R Ratio
B Allele Frequency
b
a
POLYPLOID ACC
HYPODIPLOID ACC
cChromosome
losses Cell fusion
EndoreplicationCytokinesis failure
Normaltissue
2N
HypodiploidACC
PolyploidACC
<2N >2N
Chromosomes1-3, 6, 9-11...
Chromosomes4-5, 7-8...
Normal(2 copies)
Normal(2 copies)
Deletion(1 copy)
Normal(2 copies)
Gain(4 copies)
Copy-neutral LOH(2 copies)
Genomic position
Genomic position
LOH
LOH
Nature Genetics: doi:10.1038/ng.2953

pattern of a hypodiploid ACC. The GAP pattern (left) is a sideview projection of segmented log R ratio (LRR, y axis) and B Allele Frequency (BAF, x axis) used to determine the absolute CN and genotype corresponding to each cluster of segments (Popova et al., Genome Biol 2009). Clusters are designated by the ratio of CN to most abundant allele counts (e.g. 3/2 indicates that the segment has a total copy number of 3, with 2 copies of one allele, and 1 of the other). SNP profiles are represented on the right by the absolute CN, LRR and BAF plots. Color codes in the copy number and LRR profiles are as follows: blue, homozygous deletion; green, loss; yellow, normal copy number; red, gain. Blue regions in the BAF plots correspond to regions of LOH. (b) SNP array pattern of a polyploid ACC. (c) Proposed mechanism explaining the existence of hypodiploid ACC with numerous losses and polyploid ACC with numerous copy-‐neutral LOH.
Nature Genetics: doi:10.1038/ng.2953
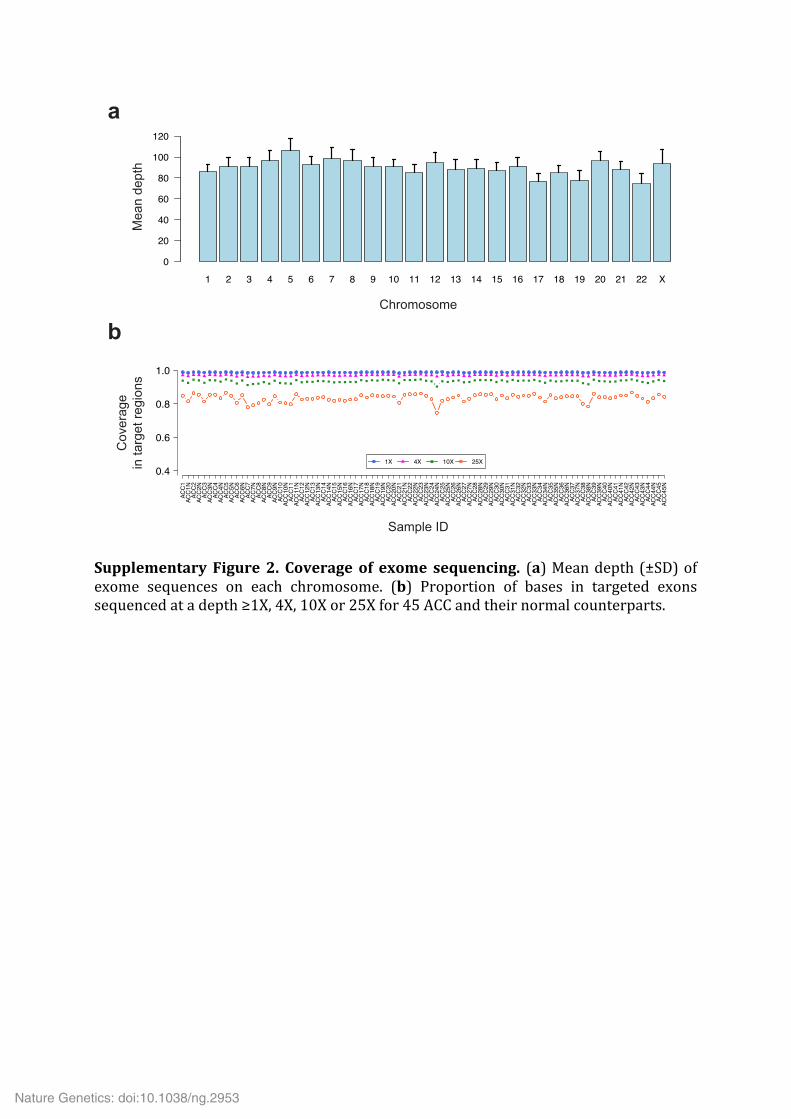
Supplementary Figure 2. Coverage of exome sequencing. (a) Mean depth (±SD) of exome sequences on each chromosome. (b) Proportion of bases in targeted exons sequenced at a depth ≥1X, 4X, 10X or 25X for 45 ACC and their normal counterparts.
! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! ! !
ACC1
ACC1N
ACC2
ACC2N
ACC3
ACC3N
ACC4
ACC4N
ACC5
ACC5N
ACC6
ACC6N
ACC7
ACC7N
ACC8
ACC8N
ACC9
ACC9N
ACC10
ACC10N
ACC11
ACC11N
ACC12
ACC12N
ACC13
ACC13N
ACC14
ACC14N
ACC15
ACC15N
ACC16
ACC16N
ACC17
ACC17N
ACC18
ACC18N
ACC19
ACC19N
ACC20
ACC20N
ACC21
ACC21N
ACC22
ACC22N
ACC23
ACC23N
ACC24
ACC24N
ACC25
ACC25N
ACC26
ACC26N
ACC27
ACC27N
ACC28
ACC28N
ACC29
ACC29N
ACC30
ACC30N
ACC31
ACC31N
ACC32
ACC32N
ACC33
ACC33N
ACC34
ACC34N
ACC35
ACC35N
ACC36
ACC36N
ACC37
ACC37N
ACC38
ACC38N
ACC39
ACC39N
ACC40
ACC40N
ACC41
ACC41N
ACC42
ACC42N
ACC43
ACC43N
ACC44
ACC44N
ACC45
ACC45N
0.4
0.6
0.8
1.0
! 1X 4X 10X 25X
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 X
0
20
40
60
80
100
120
Sample ID
Coverage
in target regions
Mean depth
Chromosome
a
b
Nature Genetics: doi:10.1038/ng.2953
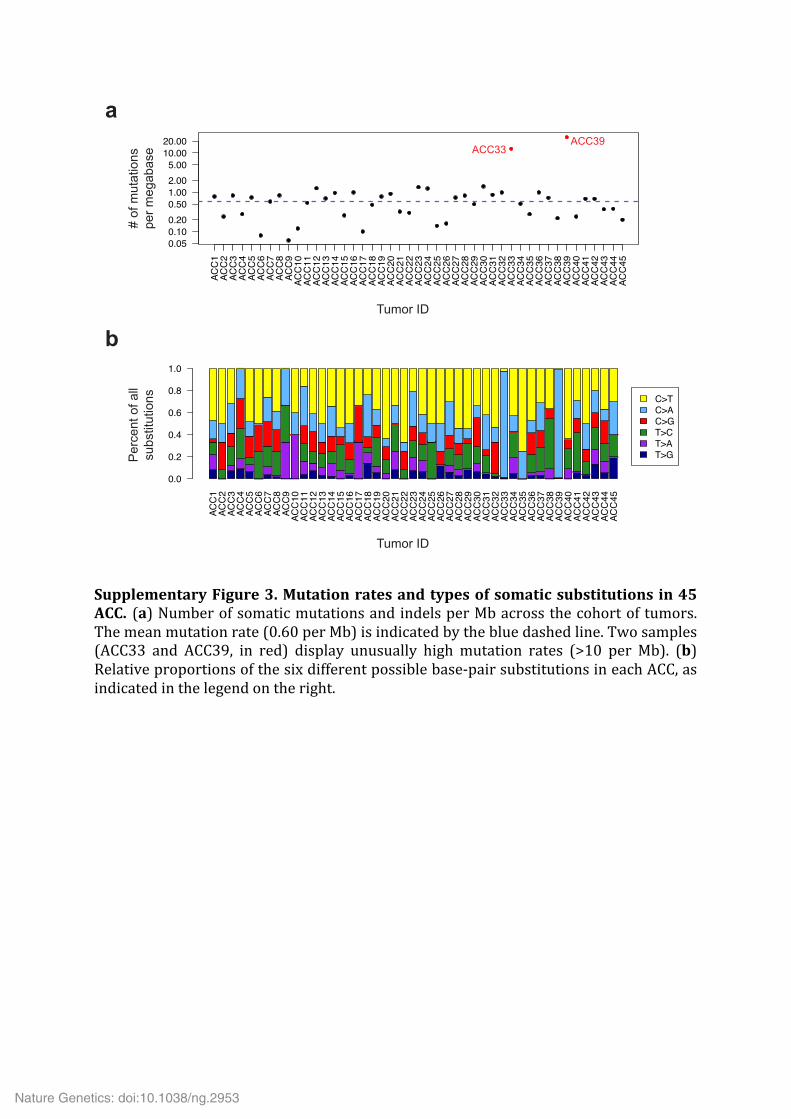
Supplementary Figure 3. Mutation rates and types of somatic substitutions in 45 ACC. (a) Number of somatic mutations and indels per Mb across the cohort of tumors. The mean mutation rate (0.60 per Mb) is indicated by the blue dashed line. Two samples (ACC33 and ACC39, in red) display unusually high mutation rates (>10 per Mb). (b) Relative proportions of the six different possible base-‐pair substitutions in each ACC, as indicated in the legend on the right.
!
!
!
!
!
!
!!
!
!
!
!
!!
!
!
!
!
!!
! !
! !
! !
! !
!
!
! !
!
!
!
!!
!
!
!
! !
! !
!
0.050.100.200.501.002.005.0010.0020.00
ACC1
ACC2
ACC3
ACC4
ACC5
ACC6
ACC7
ACC8
ACC9
ACC10
ACC11
ACC12
ACC13
ACC14
ACC15
ACC16
ACC17
ACC18
ACC19
ACC20
ACC21
ACC22
ACC23
ACC24
ACC25
ACC26
ACC27
ACC28
ACC29
ACC30
ACC31
ACC32
ACC33
ACC34
ACC35
ACC36
ACC37
ACC38
ACC39
ACC40
ACC41
ACC42
ACC43
ACC44
ACC45
Percent of all
substitutions
b
Tumor ID
# of mutations
per megabase
a
Tumor ID
ACC1
ACC2
ACC3
ACC4
ACC5
ACC6
ACC7
ACC8
ACC9
ACC10
ACC11
ACC12
ACC13
ACC14
ACC15
ACC16
ACC17
ACC18
ACC19
ACC20
ACC21
ACC22
ACC23
ACC24
ACC25
ACC26
ACC27
ACC28
ACC29
ACC30
ACC31
ACC32
ACC33
ACC34
ACC35
ACC36
ACC37
ACC38
ACC39
ACC40
ACC41
ACC42
ACC43
ACC44
ACC45
0.0
0.2
0.4
0.6
0.8
1.0
C>TC>AC>GT>CT>AT>G
ACC33ACC39
Nature Genetics: doi:10.1038/ng.2953
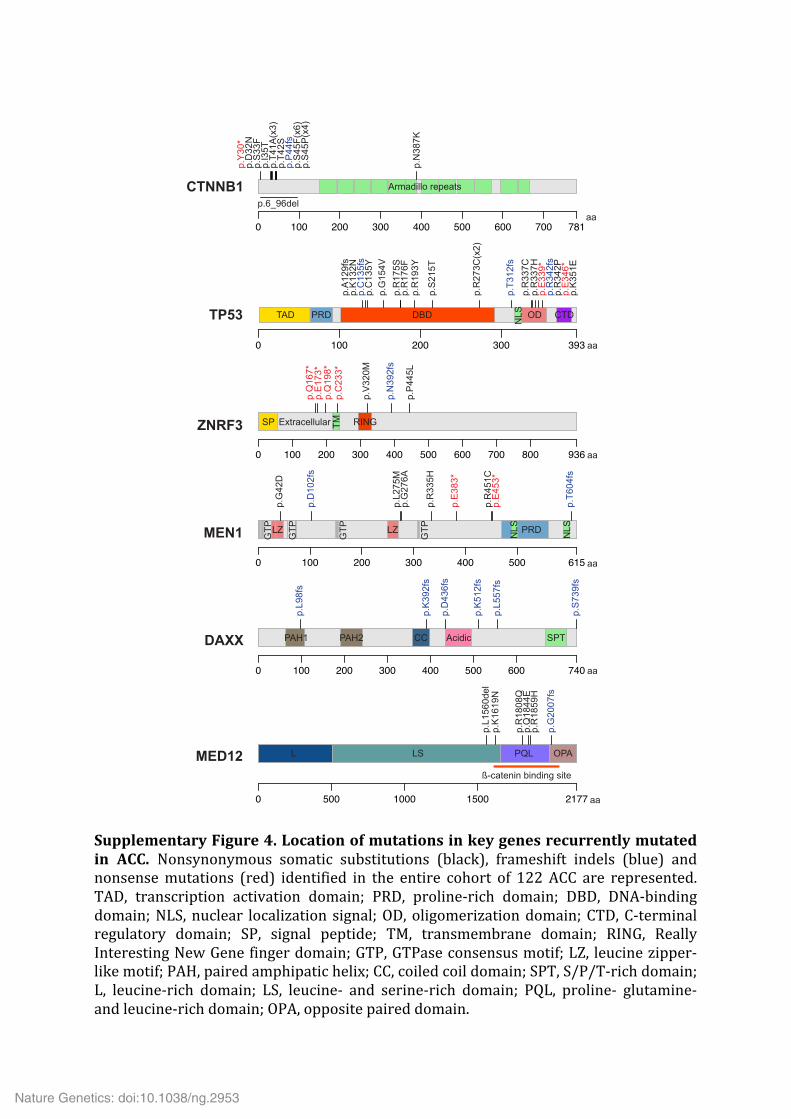
Supplementary Figure 4. Location of mutations in key genes recurrently mutated in ACC. Nonsynonymous somatic substitutions (black), frameshift indels (blue) and nonsense mutations (red) identified in the entire cohort of 122 ACC are represented. TAD, transcription activation domain; PRD, proline-‐rich domain; DBD, DNA-‐binding domain; NLS, nuclear localization signal; OD, oligomerization domain; CTD, C-‐terminal regulatory domain; SP, signal peptide; TM, transmembrane domain; RING, Really Interesting New Gene finger domain; GTP, GTPase consensus motif; LZ, leucine zipper-‐like motif; PAH, paired amphipatic helix; CC, coiled coil domain; SPT, S/P/T-‐rich domain; L, leucine-‐rich domain; LS, leucine-‐ and serine-‐rich domain; PQL, proline-‐ glutamine-‐ and leucine-‐rich domain; OPA, opposite paired domain.
0 100 200 300 400 500 600 700 781
CTNNB1
p.Y30*
p.D32N
p.S33F
p.I35T
p.T41A(x3)
p.T42S
p.S45F(x6)
p.S45P(x4)
p.N387K
Armadillo repeats
aap.6_96del
p.P44fs
0 100 200 300 393
TP53
p.K132N
p.C135Y
p.G154V
p.R175S
p.R176F
p.R193Y
p.S215T
p.R273C(x2)
p.R337C
p.R337H
p.E339*
p.R342fs
p.R342P
p.E346*
p.K351E
TAD PRD DBD
NLS OD CTD
aap.A129fs
p.C135fs
p.T312fs
0 100 200 300 400 500 600 700 800 936
ZNRF3
p.V320M
p.Q167*
p.E173*
p.Q198*
p.C233*
p.P445L
p.N392fs
SP Extracellular TM RING
aa
0 100 200 300 400 500 615
MEN1
p.G42D
p.L275M
p.G276A
p.R335H
p.E383*
p.R451C
p.E453*
LZ LZ PRDNLS
NLSaa
GTP
GTP
GTP
GTP
p.D102fs
p.T604fs
0 100 200 300 400 500 600 740
DAXX
p.L98fs
p.K392fs
p.L557fs
PAH1 PAH2 CC Acidic SPT
aa
p.D436fs
p.K512fs
p.S739fs
0 500 1000 1500 2177
MED12
p.K1619N
p.R1808Q
p.Q1844E
p.R1859H
L LS PQL OPA
aa
ß-catenin binding site
p.L1560del
p.G2007fs
Nature Genetics: doi:10.1038/ng.2953
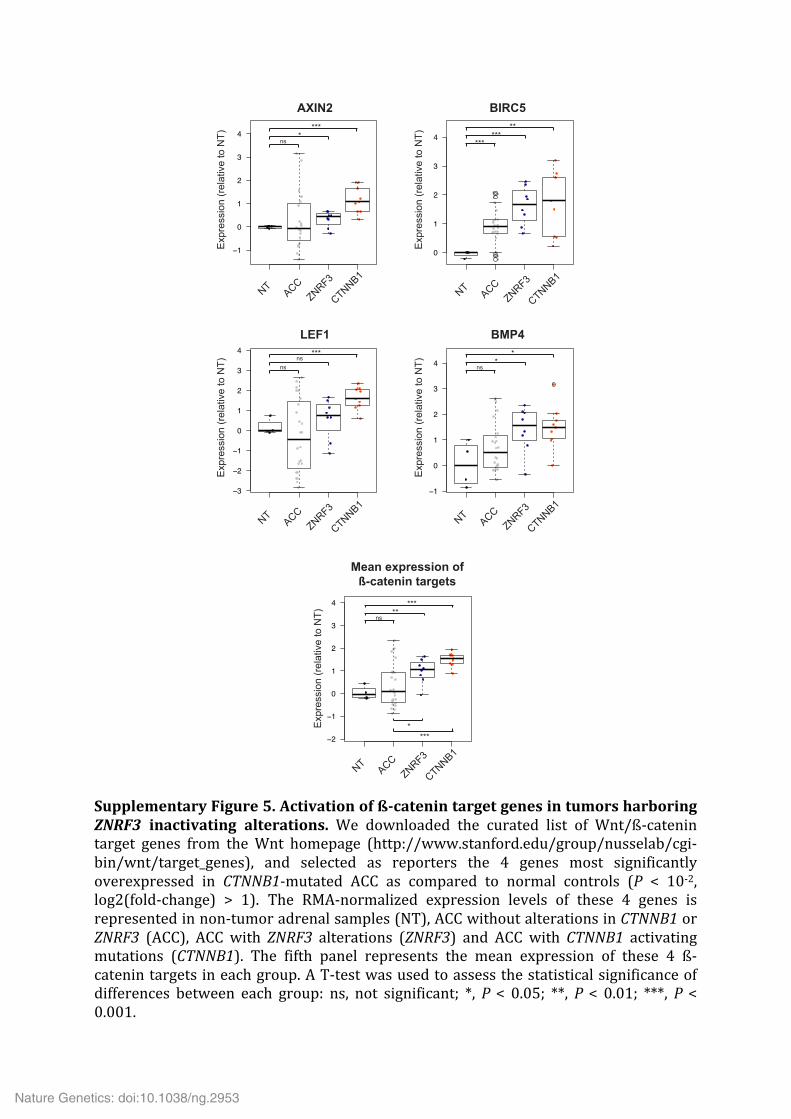
Supplementary Figure 5. Activation of ß-‐catenin target genes in tumors harboring ZNRF3 inactivating alterations. We downloaded the curated list of Wnt/ß-‐catenin target genes from the Wnt homepage (http://www.stanford.edu/group/nusselab/cgi-‐bin/wnt/target_genes), and selected as reporters the 4 genes most significantly overexpressed in CTNNB1-‐mutated ACC as compared to normal controls (P < 10-‐2, log2(fold-‐change) > 1). The RMA-‐normalized expression levels of these 4 genes is represented in non-‐tumor adrenal samples (NT), ACC without alterations in CTNNB1 or ZNRF3 (ACC), ACC with ZNRF3 alterations (ZNRF3) and ACC with CTNNB1 activating mutations (CTNNB1). The fifth panel represents the mean expression of these 4 ß-‐catenin targets in each group. A T-‐test was used to assess the statistical significance of differences between each group: ns, not significant; *, P < 0.05; **, P < 0.01; ***, P < 0.001.
0
2
3
4
!!! !
!
!
!
!
!
!
!
!
!
!
!
!
!!
!
!
!
!
!
!
!
!
!!
!
!
!!
!
!
!
!
!
!
!
!
!
!
!
!
!
0
2
3
4
!
!
!!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!!
!
!
!
!
!
!
!
0
2
3
4
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!!
!
!
!!
! !
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
0
2
3
4
!!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!!
!
!
!
!
!
!
!
!
!
!
!
!!
!
!
!!
!
!!
!
!
0
2
3
4
!
!
!
!
!
!
! !
!
!!
!
!
!
!
!
!
!
!
!
!
!!
!!
!
!
!
!
!
!
!
!
!
!!
!
!
!
!
!
!!
!
!
AXIN2 BIRC5
BMP4LEF1
Mean expression ofß-catenin targets
NT ACCZNRF3
CTNNB1
Expression (relative to NT) ns
****
******
**
nsns
***
ns*
*
ns**
***
****
NT ACCZNRF3
CTNNB1
NT ACCZNRF3
CTNNB1
NT ACCZNRF3
CTNNB1
NT ACCZNRF3
CTNNB1
Expression (relative to NT)
Expression (relative to NT)
Expression (relative to NT)
Expression (relative to NT)
Nature Genetics: doi:10.1038/ng.2953
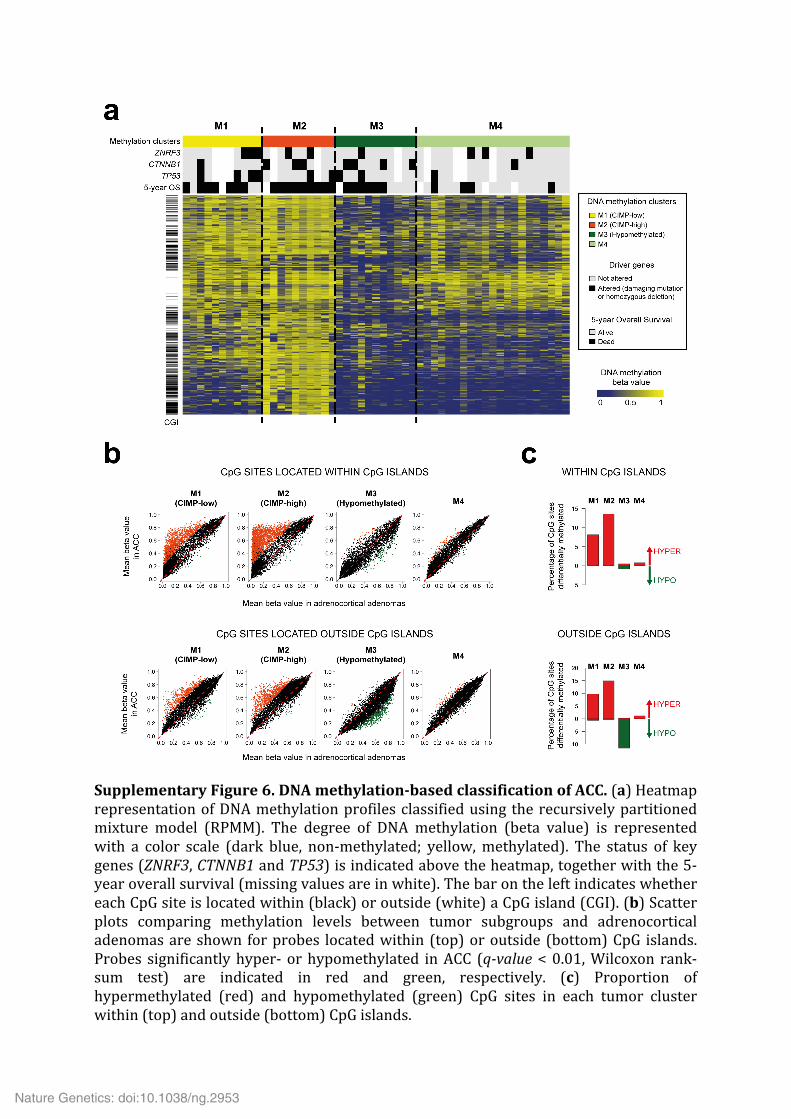
Supplementary Figure 6. DNA methylation-‐based classification of ACC. (a) Heatmap representation of DNA methylation profiles classified using the recursively partitioned mixture model (RPMM). The degree of DNA methylation (beta value) is represented with a color scale (dark blue, non-‐methylated; yellow, methylated). The status of key genes (ZNRF3, CTNNB1 and TP53) is indicated above the heatmap, together with the 5-‐year overall survival (missing values are in white). The bar on the left indicates whether each CpG site is located within (black) or outside (white) a CpG island (CGI). (b) Scatter plots comparing methylation levels between tumor subgroups and adrenocortical adenomas are shown for probes located within (top) or outside (bottom) CpG islands. Probes significantly hyper-‐ or hypomethylated in ACC (q-‐value < 0.01, Wilcoxon rank-‐sum test) are indicated in red and green, respectively. (c) Proportion of hypermethylated (red) and hypomethylated (green) CpG sites in each tumor cluster within (top) and outside (bottom) CpG islands.
Nature Genetics: doi:10.1038/ng.2953
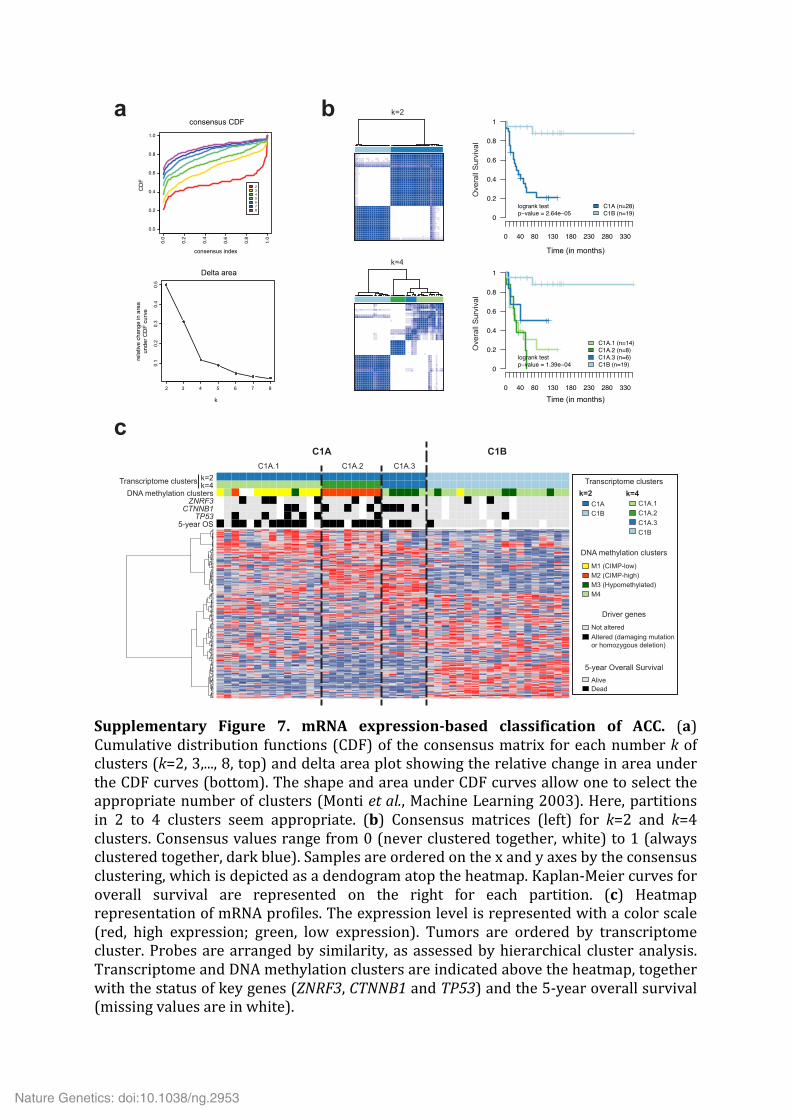
Supplementary Figure 7. mRNA expression-‐based classification of ACC. (a) Cumulative distribution functions (CDF) of the consensus matrix for each number k of clusters (k=2, 3,..., 8, top) and delta area plot showing the relative change in area under the CDF curves (bottom). The shape and area under CDF curves allow one to select the appropriate number of clusters (Monti et al., Machine Learning 2003). Here, partitions in 2 to 4 clusters seem appropriate. (b) Consensus matrices (left) for k=2 and k=4 clusters. Consensus values range from 0 (never clustered together, white) to 1 (always clustered together, dark blue). Samples are ordered on the x and y axes by the consensus clustering, which is depicted as a dendogram atop the heatmap. Kaplan-‐Meier curves for overall survival are represented on the right for each partition. (c) Heatmap representation of mRNA profiles. The expression level is represented with a color scale (red, high expression; green, low expression). Tumors are ordered by transcriptome cluster. Probes are arranged by similarity, as assessed by hierarchical cluster analysis. Transcriptome and DNA methylation clusters are indicated above the heatmap, together with the status of key genes (ZNRF3, CTNNB1 and TP53) and the 5-‐year overall survival (missing values are in white).
0 40 80 130 180 230 280 330
0
0.2
0.4
0.6
0.8
1
C1A (n=28)
C1B (n=19)
logrank test
0.0
0.2
0.4
0.6
0.8
1.0
0.0
0.2
0.4
0.6
0.8
1.0
2
3
4
6
7
8
2 3 4 5 6 7 8
0.1
0.2
0.3
0.4
0.5
k=2
k=4
0 40 80 130 180 230 280 330
0
0.2
0.4
0.6
0.8
1
C1A.1 (n=14)
C1A.2 (n=8)
C1A.3 (n=6)
C1B (n=19)
logrank testrelative change in area
under CDF curve
consensus index
CDF
k
consensus CDF
Delta area
Death
CTNNB1
ZNRF3
METH
EXP
EXP2gk=2k=4
DNA methylation clustersZNRF3
CTNNB1TP53
5-year OS
Overall Survival
Time (in months)
Time (in months)
Overall Survival
a b
c
Transcriptome clusters Transcriptome clustersk=2 k=4
C1AC1B
C1A.1
C1B
C1A.2C1A.3
DNA methylation clusters
M1 (CIMP-low)M2 (CIMP-high)M3 (Hypomethylated)M4
Driver genesNot alteredAltered (damaging mutationor homozygous deletion)
5-year Overall SurvivalAliveDead
C1A C1BC1A.1 C1A.2 C1A.3
Nature Genetics: doi:10.1038/ng.2953
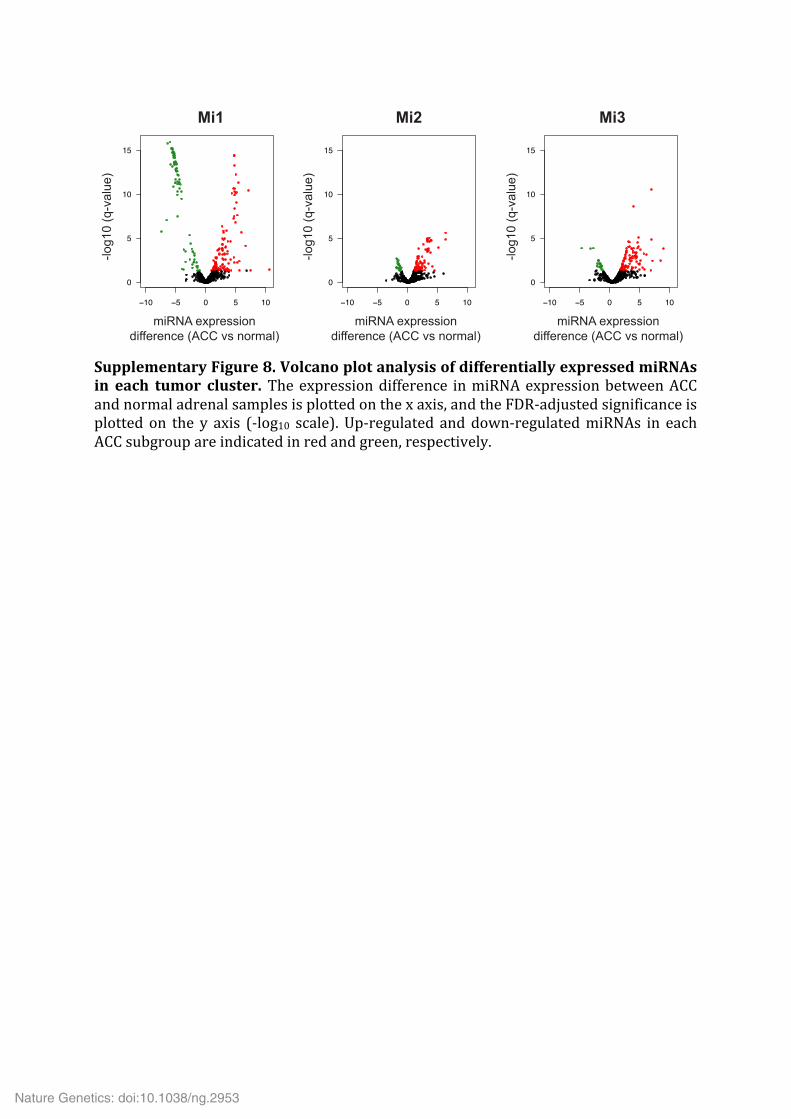
Supplementary Figure 8. Volcano plot analysis of differentially expressed miRNAs in each tumor cluster. The expression difference in miRNA expression between ACC and normal adrenal samples is plotted on the x axis, and the FDR-‐adjusted significance is plotted on the y axis (-‐log10 scale). Up-‐regulated and down-‐regulated miRNAs in each ACC subgroup are indicated in red and green, respectively.
!!!!!
!
!!
!!
!
!
!!!
!!
!!!
!!!
!
!
! !!
!
!!
!!!
!
!
!
!
!
!
!
!!
!
!
!
!
!
!!!
!
!!
!
! !!!
!
!
!
!
!
!!!!!!!
!!
!
!
!!
!
!!!!
!!
!!
!!!!
!!!
!
!
!
!!
!! !
!!
!!!
!!!
!
!
!
!
!!
!
!
!
!
!!
!
!!
! !
!!
!
!!!
!!! !
!
!
!
!
!!
!
!
!
!
!
!!!!!
!
!
!
!!
!!
!
!!!
!
!
!
!!!!!!
!
!
!
!
!!
!!
!
!
!
!!!
!
!!
!
!!
!
!!
!!
!
!
!
!
!!
!!
!!!
!!!
!!!!! !
!
!
!
!
!
!
!!! !
!
!
!! !!
!!
!
!
!
!
!
!
!
! !!!!!!
!
!
!
!!
!!
!
!!
! !
! !!
!! !!
!!
!!
!!!!
!!
!
!!
!!!!
!
!
!
!
!!
!
!!!
!
!!!!
!
!!!!!!!!
!!!!
!!
!!
!
!!!!
!
!
!!
!
!!
!
!!
!!
!
!
!
!!!!
!
!!!!!
!
!
!
!
! !
!
!
!
!!
!!
!!
!
!
!
!
!
!
! !!!!!
!
!!
! !!
!
!!
!
!
!!
! !!!!! !!
!
!
!!!
! !
!!
!
!
!
!
!
!!
!
!
!
!
!
!
!!!!!!!
!
!
!
!
!
!
!
!
!
!
!
!!!
!
!
!
!
!
!
!
!
!!
!
!!
!
!
!
!
!
!
! !!
!
!
!
!
!!!! !
!!
!
!
!
!
!
!
!!!
!!
!
!!!
!!
!!!!!
!
!
!
!!
!!!
!!! !
!
!
!
!!
!
!
!
!
!!
! !
!
!
!!
!!
!!
!!!
!
! !
!
!!! !!
!
!
!! !!!! !!!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!
!!!
!!!!
!
!!!
!!
!
!
!
!!
!
!!
!
!
!
!
!
!
!
!
!
!
!
!
!
!!
!
!
!
!
!
!
!
!
!!
!
!
!
!
!!
!
!
!
!
!
!
!
!!!!!
!!
!!!
!!! !!
!!!
!!!!
!
!!!!! !
!
!!
!
!!
!
!
!!
!!!
!!
!
!
!!
!
!
!!!
!
!!!
!!
!
!!!
!
!
!
!
!
!
!!! !
!
!
!
!
!
!!
!
!
!
!
!!!!
!
!
!
!
!
!!
!
!
!
!
!
!!
!
!
!
!
!
!!!
!!!
!!
!
!
!!
!
!!!! !
!
!!! !
!
-log10 (q-value)
miRNA expressiondifference (ACC vs normal)
Mi1
!!
!!!!
!!!!!!!!!
!!!!!
!!
!!
!! !!!
!!
!!!!
!
!!
!
!
!!
!
!
!
!
!!!
!!
!
!!
!
!!
!
!! !!!!! !!!!!!
!!
!!
!
!
!
!!
!!
!!
!! !
!!!
! !!
!
!
!
!!!
!
!
!!
!!!!
!!
!!
!!!! !
!
!
!!
! !!!! !!!!
!
!
!
!!!! !!
!
!!!
!
!!!
!
!
!
!!!
!!
!! !!
!!
!
!!! !
!!!!
!!!
!
!!
!
!!
!
!!
!!!!!
!
!
!
!!!
!!! !
!
!!
!
!!!
!
!
!!
!!!
!! !!!
!!!!
!
!!
!!
!!!!
!
!!!!!!!
!!
!!!!
!
!!!!!!!
!
!
!!
!
!!!!!!
!!!!!!!!!! !!
!
!!!
!!!!!!
!
!
!!!
!!
! !!! !!!!
!!!!
!! !!!!!!
!!! !!!!! !!!
!
!!
!!! !
!
!!
!!!!
!
!
!!!
!
!
!!!!!
!
!
!
!!
!
! !!
!!
!!
!
!
!!!!
!!
!
!!! !!!
!!
!!!
!
!
!
!
!
!
!
!
!!!!!!
!
!
!
!! !!
!!
!
!!!
!
!!!!
!
!
!
!!
!!!
!!!!
!
!!
!!!!
!
!
!
!
!!!!!!
!
!
!!
!
!!
!!!
!
!!
!
!
!!
!
!!!
!
!
!!!
!!!!
!
!
!
!
!
!
!
!
!!
!!!
! !!
!!!!
!!
!
!
!
!!
!!
!
! !!!
!
!!!!!!
!
!
!!
!!
!
!!
!!
!
!!!!!
!
!!
!
!!
! !!
!
!
!! !!! ! !!!
!
!
!
!
! !!
!
!
!
!
!
!
!
!!!
!
!
!
!
!! !!!
! !!!!!
!
!!!
!!!!!
!
!
!
!!
!
!
!
!
!!!!
!
!!
!!
!! !
!!!
!
!!
!
!
!
!
!!
!
!!
!
!!
!
!!
!!
!!
!
!!!!
!! !! !! !!
!!!!!!! !!
!!
!!!
!!
!
!
!!!!!!!!!!!!
!
!
!!!
!
!!
!! !!
!!
!!!!
!!
!!!
!
!!
!
!
!
!!
!!
!!
!
!
!!
!
!
!!!!
!
! !
!!!!! ! !
!!!
!
!
!
!
!!
!!!!!
! !!
!!
!!
!!!
!!
!
!!!
!
miRNA expressiondifference (ACC vs normal)
! !!!!
! !!
!!!!!!!
!!
!!!!
!
!!
!
!!
!!!!
!!
! !
!
!
! !
! !!
!!! !
!!
!
!!
!!!
!
!!!!
!
!!!
!!!!!!
!!!
!
!
!
!!!
!!!
!
!
!
!!
!!!
!
!
!
!
!!!!!
!!!
!
!!!
!
!
!!
!
!
!
!! !!!
!
!
!
!!
!!
!
!
!!
!!
!!
!!
!!
!!!
!
!!
!
!
!
!
!
!
!
!!
!
!
!
!!!
!
!!
!!
!! !
!
!!
!
! !
!
!!!
!
!!!!
!
!!
!
!! !!
!!
!!
!
!!
!!
!
!
!
!
!
!!!
!!!
!!!
!
!
!!
!
!
!!
!
!
!
!!
!
!
!!
!
!!!
!!!!
!!
!
!
!!!
!!! !!
!
!
!
!
!
! !!!
!!!!!
!
!
!
!!! !!
!
!!
!!!
! !!
!!!!
!!
!
!
!!
!
!!
!!!
!
!!!
!
!
!!
!
!!!
!
!!
!!! !
!
!!!!!!
!
! !!
!
!
!
!
!!
!
!
!!
!
!
! !!!!
!
!
!!!
!
!
!
!
!
!
!
!
!
!!!!
!
!
!!!!
!!
!!
!!
!!! !!
!
!!
! !!!
!! !!
!
!!!!!!
!!!
!!! !!
!
!
!
!
!
!
!!!
!
!
!
!
!
!!!!!
!
!
!
!
!!!
!!!! !!
!!!!
!
!!
!
!!!
!
!!
!
!
!!!
!
!!!
!
!! !
!
!!!!
!!!
!
!
!
!
!
!!
!
!
!!
!
!
!
!
!
!
!!
!
!
!!
!
!
!
!!
!
!
!
!!
!!
! !!
!!
!!
!
!
!!!
!
!
!! !
!
!
!!!
!
!
!!
!
!
!
!!!!
!
!
!
!
!!
!
!
! !!
!
!
!
!
!
!
!
!
!!
!
!
!
!
!!!!
!
!
!
! !!
!
!!!! !!!
!
!
!! !!!!!!
!
!!!!
!
!
!
!! !! !!!!!
!
! !
!!!
!
!
!
!
!
!
!
!!!
!!
!
!
!
!
!
!
!
!
!
!
!!!! !!!!! !!
!!
!!
!
!
!!
!
!
!!
!
!! !! !! !
!!!!
!!!
!!!!!!
!
! !!!!! !!
!
!
!!
! !
!
!
!
!
!
!
!
!!! !!
!
!
!
! !!!
!!
!!
!!
!!!
!
!
!
!
!
!
!
!!!!!
!
!
!
!
!
!
!!
!!
!!
!!
!
!
!
!
!
!!!
!
!
!
!!
!
!!
miRNA expressiondifference (ACC vs normal)
Mi2 Mi3
-log10 (q-value)
-log10 (q-value)
Nature Genetics: doi:10.1038/ng.2953
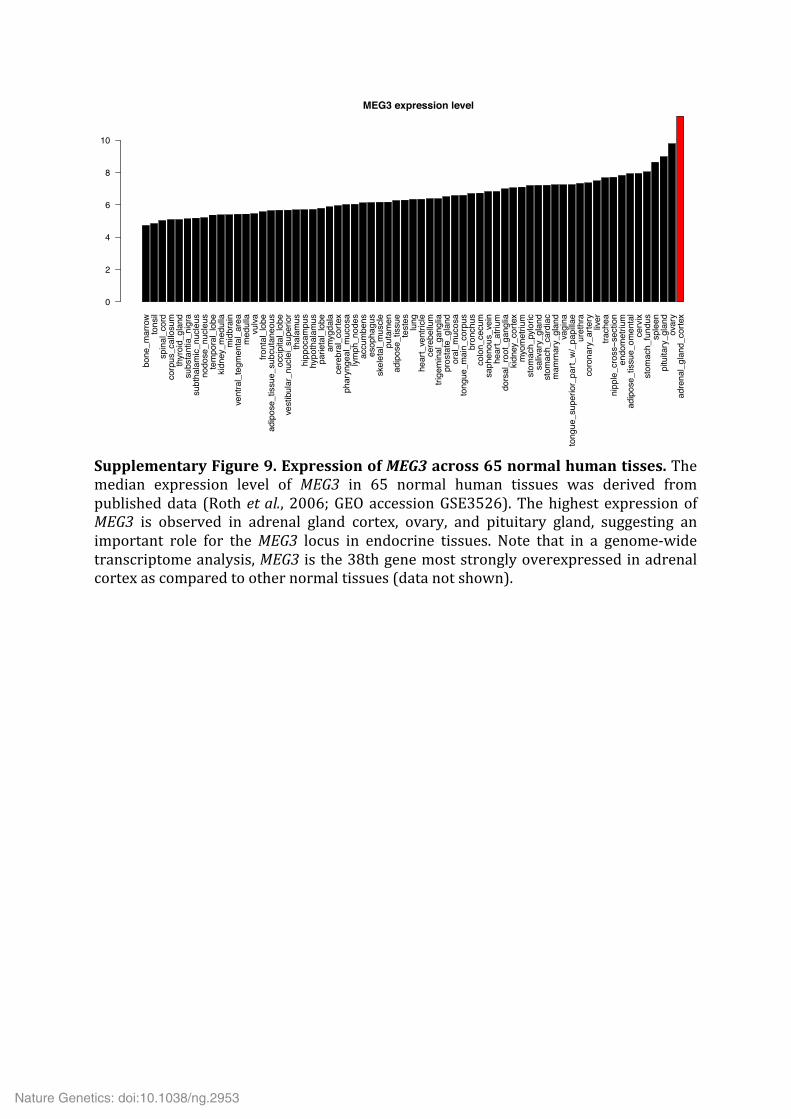
Supplementary Figure 9. Expression of MEG3 across 65 normal human tisses. The median expression level of MEG3 in 65 normal human tissues was derived from published data (Roth et al., 2006; GEO accession GSE3526). The highest expression of MEG3 is observed in adrenal gland cortex, ovary, and pituitary gland, suggesting an important role for the MEG3 locus in endocrine tissues. Note that in a genome-‐wide transcriptome analysis, MEG3 is the 38th gene most strongly overexpressed in adrenal cortex as compared to other normal tissues (data not shown).
bone_m
arrow
tonsil
spinal_cord
corpus_callosum
thyroid_gland
substantia_nigra
subthalamic_nucleus
nodose_nucleus
temporal_lobe
kidney_m
edulla
midbrain
ventral_tegm
ental_area
medulla
vulva
frontal_lobe
adipose_tissue_subcutaneous
occipital_lobe
vestibular_nuclei_superior
thalam
ushippocam
pus
hypothalam
usparietal_lobe
amygdala
cerebral_cortex
pharyngeal_m
ucosa
lymph_nodes
accumbens
esophagus
skeletal_m
uscle
putamen
adipose_tissue
testes
lung
heart_ventricle
cerebellum
trigeminal_ganglia
prostate_gland
oral_m
ucosa
tongue_m
ain_corpus
bronchus
colon_cecum
saphenous_vein
heart_atrium
dorsal_root_ganglia
kidney_cortex
myometrium
stom
ach_pyloric
salivary_gland
stom
ach_cardiac
mam
mary_gland
vagina
tongue_superior_part_w/_papillae
urethra
coronary_artery
liver
trachea
endometrium
adipose_tissue_om
ental
cervix
stom
ach_fundus
spleen
pituitary_gland
ovary
adrenal_gland_cortex
MEG3 expression level
0
2
4
6
8
10
Nature Genetics: doi:10.1038/ng.2953
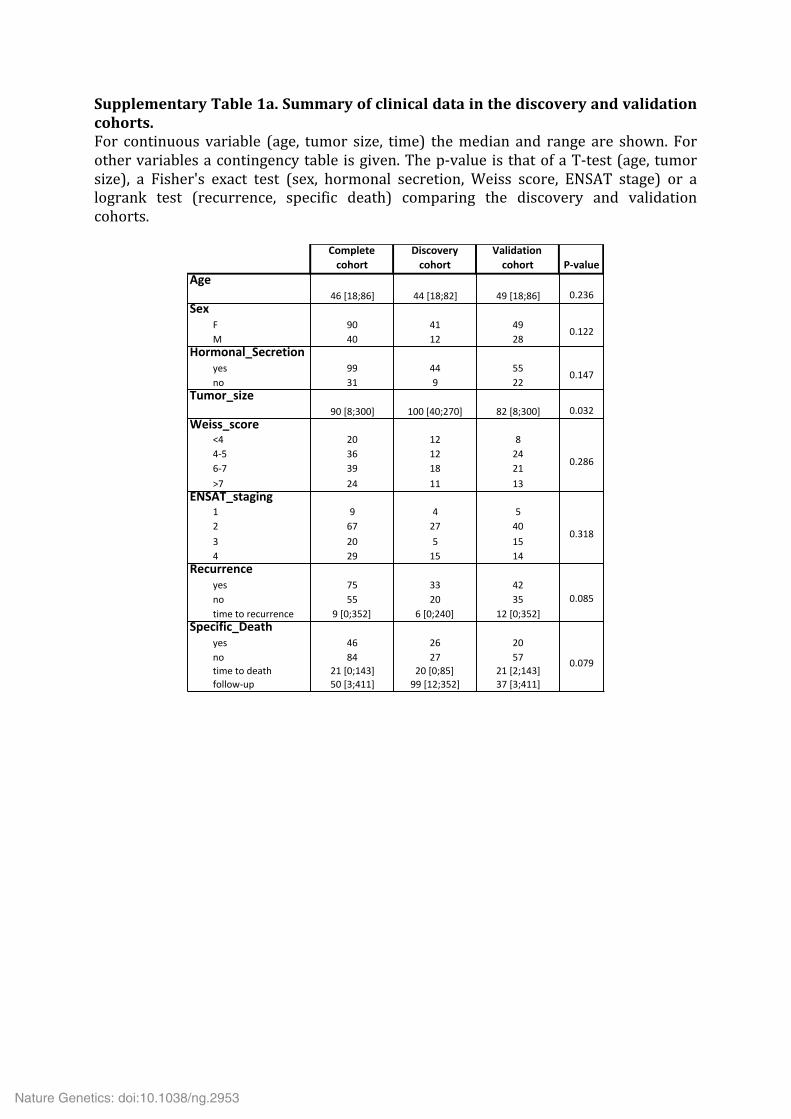
Supplementary Table 1a. Summary of clinical data in the discovery and validation cohorts. For continuous variable (age, tumor size, time) the median and range are shown. For other variables a contingency table is given. The p-‐value is that of a T-‐test (age, tumor size), a Fisher's exact test (sex, hormonal secretion, Weiss score, ENSAT stage) or a logrank test (recurrence, specific death) comparing the discovery and validation cohorts.
!"#$%&'&()"*"+'
,-.)"/&+0()"*"+'
12%-32'-"4()"*"+' 56/2%7&
89&!"#$%&'&"( !!#$%&'&)( !*#$%&'&"( +,)-"
:&;. *+ !% !*/ !+ %) )&
<"+#"42%=:&)+&'-"4012 ** !! 3345 -% * ))
>7#"+=.-?&*+#$&'-++( %++#$!+')6+( &)#$&'-++( +,+-)
@&-..=.)"+&7! )+ %) &!83 -" %) )!"86 -* %& )%96 )! %% %-
AB:8>=.'29-49% * ! 3) "6 )6 !+- )+ 3 %3! )* %3 %!
C&)7++&4)&012 63 -- !)45 33 )+ -3:;<1#:5#=1>?==14>1 *#$+'-3)( "#$+')!+( %)#$+'-3)(
:$&)-D-)=,&2'*012 !" )" )+45 &! )6 36:;<1#:5#@1A:B )%#$+'%!-( )+#$+'&3( )%#$)'%!-(C5DD5E8?F# 3+#$-'!%%( **#$%)'-3)( -6#$-'!%%(
+,%))
+,%!6
+,+6*
+,+&3
+,)&"
+,-%&
Nature Genetics: doi:10.1038/ng.2953

Supplementary Table 1b. Sample characteristics and analyses performed.
Sample Sex Age at diagnosis
Tumor size
(mm)
Tumor side
Hormonal Secretion
Weiss score
ENSAT staging Recurrence
Time to recurrence (months)
Specific Death
Follow up (months)
DNA sequencing SNP array
DNA methylation
array
mRNA expression
array
miRNA sequencing
ACC1 F 70.3 55 left no 4 2 no NA no 151.8 Exome OmniExpress-12v1 X X XACC2 F 25.6 50 left yes 3 1 no NA no 131.1 Exome OmniExpress-12v1 X X XACC3 M 40 70 right no 7 4 yes 0 yes 23 Exome OmniExpress-12v1 XACC4 F 53.3 80 right yes 4 2 no NA no 147.8 Exome OmniExpress-12v1 X X XACC5 M 72.9 120 right yes 8 4 yes 0 yes 0.5 Exome OmniExpress-12v1 X X XACC6 F 18.3 45 right yes 2 1 no NA no 142.8 Exome OmniExpress-12v1 X X XACC7 F 77.5 270 right yes 8 2 yes 0 yes 5.2 Exome OmniExpress-12v1 XACC8 F 50.7 80 right yes 5 2 yes 7.3 yes 36 Exome OmniExpress-12v1 X XACC9 M 63.9 80 right yes 8 4 yes 0 yes 0.6 Exome OmniExpress-12v1 X X XACC10 F 27 130 left yes 7 2 no NA no 73.6 Exome OmniExpress-12v1 X X XACC11 M 29.6 160 right yes 8 4 yes 0 yes 74.4 Exome OmniExpress-12v1 X X XACC12 F 79.3 140 right yes 7 2 no NA no 12 Exome OmniExpress-12v1 X X XACC13 F 46.2 110 left yes 8 4 yes 0 yes 12.7 Exome Human610-Quadv1 X X XACC14 F 43 160 left yes 6 3 yes 6.2 yes 9.5 Exome OmniExpress-12v1 X X XACC15 F 53.9 100 right no 6 2 no NA no 115.1 Exome OmniExpress-12v1 X X XACC16 F 45 100 right yes 7 NA yes 21.8 yes 85.1 Exome OmniExpress-12v1 X X XACC17 F 41 50 left no 3 1 no NA no 59.6 Exome OmniExpress-12v1 X X XACC18 F 37.2 240 left yes 9 4 yes 0 yes 11.3 Exome OmniExpress-12v1 X X XACC19 F 81.6 90 right yes 2 2 no NA no 40.2 Exome Human610-Quadv1 X X XACC20 M 67.5 50 right yes 6 1 yes 19.3 yes 40.2 Exome OmniExpress-12v1 X X XACC21 F 42.3 55 left yes 4 2 no NA no 41.6 Exome OmniExpress-12v1 X X XACC22 F 39.7 85 left no 6 2 no NA no 57.2 Exome OmniExpress-12v1 X X XACC23 F 25.2 110 left yes 5 4 yes 0 yes 48.3 Exome OmniExpress-12v1 X X XACC24 F 41.7 150 left yes 8 2 yes 18 no 35.9 Exome OmniExpress-12v1 X X XACC25 F 37.9 90 left no 3 2 yes 36 no 99.4 Exome OmniExpress-12v1 X X XACC26 F 23.9 95 left yes 9 4 yes 0 yes 9.4 Exome OmniExpress-12v1 X X XACC27 F 59.5 110 left yes 5 2 no NA no 40.9 Exome OmniExpress-12v1 X XACC28 F 75.5 170 right yes 5 2 yes 10 no 28 Exome OmniExpress-12v1 X X XACC29 F 37.6 90 left yes 6 2 yes 56.8 no 81.8 Exome OmniExpress-12v1 X X XACC30 F 34.1 90 right yes 7 3 yes 7.1 yes 11.7 Exome OmniExpress-12v1 XACC31 M 26.1 140 right yes 7 4 yes 0 yes 30.2 Exome Human610-Quadv1 X XACC32 F 26.5 80 left yes 9 4 yes 0 yes 2 Exome OmniExpress-12v1 X X XACC33 F 48.4 150 left yes 6 4 yes 11.8 yes 19.8 Exome Human610-Quadv1 X X XACC34 F 58.8 110 right yes 3 2 yes 31 yes 34.3 Exome OmniExpress-12v1 XACC35 F 49.6 70 right yes 2 2 no NA no 129.8 Exome OmniExpress-12v1 X X XACC36 M 54.3 160 left yes 7 3 yes 29.3 yes 57.9 Exome Human610-Quadv1 X X XACC37 F 79.6 90 right yes 6 4 yes 0 yes 7.6 Exome Human610-Quadv1 X X XACC38 F 29 70 right yes 2 2 no NA no 118 Exome OmniExpress-12v1 X X XACC39 F 44.5 220 right yes 4 2 no NA no 119.5 Exome OmniExpress-12v1 X X XACC40 F 28.5 65 left yes 3 2 no NA no 144.3 Exome OmniExpress-12v1 X X XACC41 F 68.9 120 right no 6 3 yes 7 yes 19 Exome OmniExpress-12v1 XACC42 M 28.9 150 left yes 4 4 yes 0 yes 21 Exome Human610-Quadv1 X X XACC43 M 52.4 200 right yes 8 4 yes 0 yes 24.3 Exome OmniExpress-12v1 X X XACC44 F 30 75 left yes 5 2 no NA no 111.5 Exome OmniExpress-12v1 X X XACC45 F 46.3 100 right yes 3 2 no NA no 97.6 Exome OmniExpress-12v1 X X XACC46 F 59.4 160 left yes 6 2 yes 11 no 12.9 X X XACC47 F 18.6 40 right yes 6 2 yes 18.4 yes 55.4 X X XACC48 M 39.6 110 right yes 3 2 yes 17.3 yes 26 X X XACC49 F 40.2 60 right yes 2 2 no NA no 158.8 X X XACC50 F 53.8 150 left yes 6 4 yes 0 no 154.2 X X XACC51 M 30.6 250 right no 5 2 no NA no 85.1 X XACC52 F 44.6 NA Abdo no 8 NA yes 240 no 351.7 X XACC55 M 52.9 NA left yes 5 3 yes 4.5 yes 11.4 X XACC56 F 53 70 right yes NA 3 no NA no 122.78 Targeted HumanCore-12v1ACC57 F 40 58 right yes 9 4 yes 22.31 no 28.85 Targeted HumanCore-12v1ACC58 M 60 125 left yes 7 2 no NA no 20.83 Targeted HumanCore-12v1ACC59 F 42 105 left yes 5 2 yes 50.27 no 60.62 Targeted HumanCore-12v1ACC60 M 18 300 right yes 8 3 yes 9.43 yes 29.44 Targeted HumanCore-12v1ACC61 M 46 76 left yes NA 3 no NA no 36.83 Targeted HumanCore-12v1ACC62 F 79 70 right yes NA 2 yes 14.69 no 28.06 Targeted HumanCore-12v1ACC63 F 53 NA left no 7 1 yes 8.21 yes 47.7 Targeted HumanCore-12v1ACC64 F 41 160 right yes NA 2 no NA no 21.36 Targeted HumanCore-12v1ACC65 M 50 70 left yes NA 3 yes 70.97 no 129.87 Targeted HumanCore-12v1ACC66 F 24 100 left yes 3 4 yes 0 yes 10.32 Targeted HumanCore-12v1ACC67 M 42 50 left no 6 4 yes 13.96 yes 26.41 Targeted HumanCore-12v1ACC68 M 72 140 right yes 6 2 yes 48.03 no 151.56 Targeted HumanCore-12v1ACC69 F 75 50 right yes 4 1 no NA no 47.87 Targeted HumanCore-12v1ACC70 M 59 55 right yes 5 4 yes 0 yes 39.56 Targeted HumanCore-12v1ACC71 F 66 130 right yes 7 4 yes 0 yes 4.24 Targeted HumanCore-12v1ACC72 F 37 130 left yes NA 2 no NA no 121.4 Targeted HumanCore-12v1ACC73 M 48 140 left no NA 2 yes 189.5 no 272.49 Targeted HumanCore-12v1ACC74 M 60 120 left no 4 3 yes 9.59 yes 80.95 Targeted HumanCore-12v1ACC75 M 69 95 left yes 4 2 no NA no 40.9 Targeted HumanCore-12v1ACC76 F 45 130 right yes 9 3 yes 8.41 yes 12.39 Targeted HumanCore-12v1ACC77 F 49 80 left yes 9 3 yes 5.06 yes 26.28 TargetedACC78 F 68 8 left yes 7 2 yes 18.56 yes 23.79 Targeted HumanCore-12v1ACC79 F 68 60 left yes 6 4 yes 0 yes 21.32 Targeted HumanCore-12v1ACC80 F 65 120 right no 8 2 yes 38.6 no 93.27 Targeted HumanCore-12v1ACC81 F 79 160 left yes 8 2 yes 23.52 yes 28.94 Targeted HumanCore-12v1ACC82 M 31 82 left no 3 2 no NA no 76.35 Targeted HumanCore-12v1ACC83 F 29 38 left no 3 2 no NA no 14.59 Targeted HumanCore-12v1ACC84 M 63 8.2 right no 4 2 no NA no 71.49 Targeted HumanCore-12v1ACC85 F 67 51 right no 7 2 no NA no 3.25 Targeted HumanCore-12v1ACC86 F 43 80 right no 8 NA yes 98.89 yes 143.31 Targeted HumanCore-12v1ACC87 M 37 20 left yes 5 1 yes 6.37 no 66.69 Targeted HumanCore-12v1ACC88 F 76 100 right yes 7 2 yes 10.81 yes NA Targeted HumanCore-12v1ACC89 F 47 160 right no 4 2 no NA no 67.91 Targeted HumanCore-12v1ACC90 M 24 120 right yes 7 2 no NA no 60.25 Targeted HumanCore-12v1ACC91 F 31 83 right no 6 2 no NA no 63.77 Targeted HumanCore-12v1ACC92 M 37 75 right yes 6 2 no NA no 24.48 Targeted HumanCore-12v1ACC93 F 25 50 left yes 4 NA yes 352 no 411.17 Targeted HumanCore-12v1ACC94 M 42 40 right yes 4 2 yes 58.97 no 102.8 Targeted HumanCore-12v1ACC95 F 46 90 left yes 6 2 yes 13.27 no 54.77 Targeted HumanCore-12v1ACC96 F 33 70 left no 3 2 no NA no 23.2 Targeted HumanCore-12v1ACC97 F 30 230 right no 6 4 yes 16.72 no 49.38 Targeted HumanCore-12v1ACC98 F 49 13.5 left yes 4 3 no NA no 43.1 Targeted HumanCore-12v1ACC99 F 48 70 left yes 5 2 no NA no 35.29 Targeted HumanCore-12v1ACC100 F 41 10.5 right yes 8 3 yes 7.06 no 36.86 Targeted HumanCore-12v1ACC101 F 42 55 right yes 5 2 no NA no 14.62 Targeted HumanCore-12v1ACC102 M 45 50 right no 4 2 no NA no 11.86 Targeted HumanCore-12v1ACC103 F 24 65 right no 7 NA yes 21.03 no 56.64 Targeted HumanCore-12v1ACC104 F 64 80 left yes 3 2 yes 35.98 no 76.91 Targeted HumanCore-12v1ACC105 M 49 180 left no 5 2 no NA no 30.55 Targeted HumanCore-12v1ACC106 F 22 120 right yes 4 2 no NA no 14.98 Targeted HumanCore-12v1ACC107 M 34 170 right yes 9 4 yes 0 yes 1.81 Targeted HumanCore-12v1ACC108 F 57 70 right yes 7 3 no NA no 29.14 Targeted HumanCore-12v1ACC109 F 61 20 right yes 6 1 no NA no 36.9 Targeted HumanCore-12v1ACC110 M 26 140 right yes 7 2 no NA no 25.49 Targeted HumanCore-12v1ACC111 F 58 70 right no 8 3 yes 0 no 50 Targeted HumanCore-12v1ACC112 M 53 37 right no 3 1 no NA no 27.56 Targeted HumanCore-12v1ACC113 F 53 80 left no 4 2 no NA no 11.1 Targeted HumanCore-12v1ACC114 F 37 60 right yes 4 4 no NA no 25.43 Targeted HumanCore-12v1ACC115 F 53 105 right yes 3 2 no NA no 21.78 Targeted HumanCore-12v1ACC116 F 72 80 left yes 3 4 yes 0 yes 4.7 Targeted HumanCore-12v1ACC117 M 63 140 left no 5 2 yes 37.06 no 50.86 Targeted HumanCore-12v1ACC118 M 46 95 right yes 9 3 yes 3.42 no 50.46 Targeted HumanCore-12v1ACC119 F 69 8.3 left yes 4 4 yes 0 yes 2.37 Targeted HumanCore-12v1ACC120 M 22 220 left yes NA 2 yes 9.59 no 36.99 Targeted HumanCore-12v1ACC121 F 86 93 right yes NA 3 no NA no 14.59 Targeted HumanCore-12v1ACC122 F 49 110 right yes 8 2 yes 18.96 no 24.71 Targeted HumanCore-12v1ACC123 F 79 65 left yes 5 3 no NA no 3.81 Targeted HumanCore-12v1ACC124 F 49 110 right yes 7 3 no NA no 19.02 Targeted HumanCore-12v1ACC125 M 44 170 left yes 9 4 yes 0 no 5.75 Targeted HumanCore-12v1ACC126 M 60 100 NA yes 4 2 no NA no 5.62 Targeted HumanCore-12v1ACC127 M 72 90 left no NA 2 no NA no 60.62 Targeted HumanCore-12v1ACC128 M 46 90 right yes 6 4 yes 0 yes 5.98 Targeted HumanCore-12v1ACC129 F 37 90 left yes NA 2 yes 22.57 no 59.14 Targeted HumanCore-12v1ACC130 F 51 110 left yes 4 2 yes 12.81 yes 16.3 Targeted HumanCore-12v1ACC131 F 43 170 right yes 7 4 yes 0 yes 14.36 Targeted HumanCore-12v1ACC132 F 63 76 left yes 5 2 no NA no 21.78 Targeted HumanCore-12v1
DIS
CO
VERY
CO
HO
RT
VALI
DAT
ION
CO
HO
RT
CLINICAL DATA ANALYSES PERFORMED
Nature Genetics: doi:10.1038/ng.2953
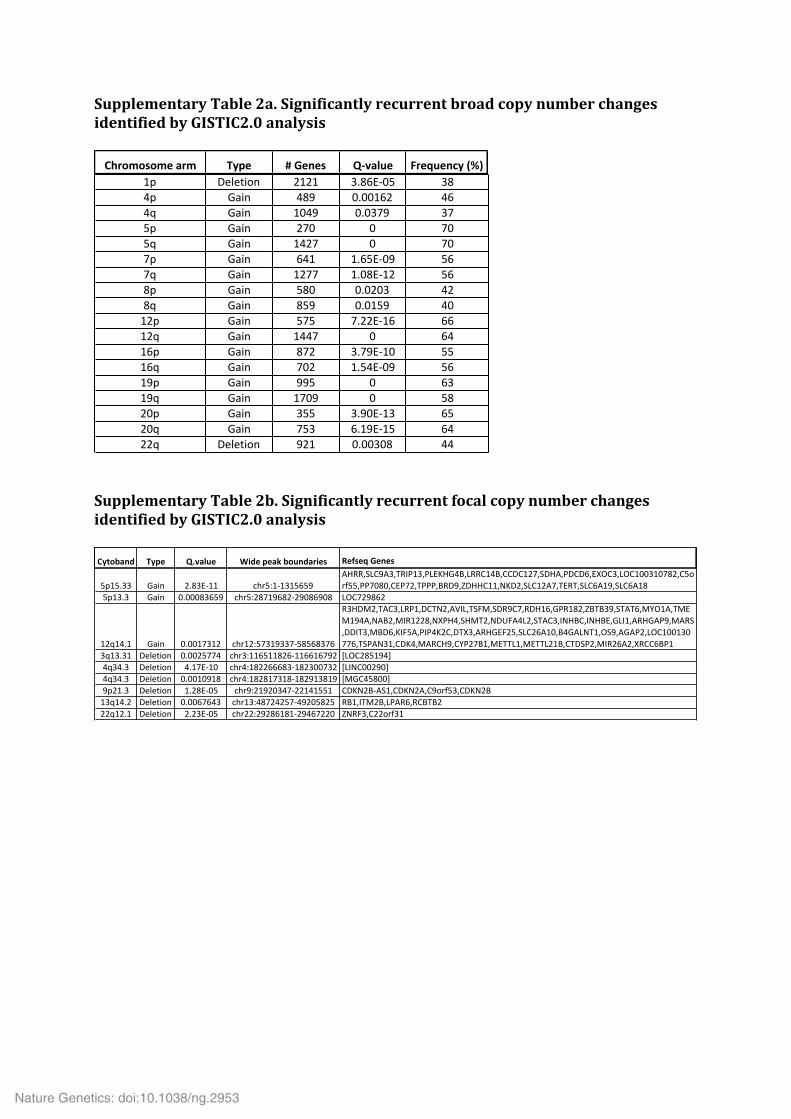
Supplementary Table 2a. Significantly recurrent broad copy number changes identified by GISTIC2.0 analysis
Supplementary Table 2b. Significantly recurrent focal copy number changes identified by GISTIC2.0 analysis
!"#$%$&$%'()#% *+,' -(.'/'& 012)34' 5#'64'/7+(89:!" #$%$&'() *!*! +,-./012 +-3" 45') 3-6 1,11!.* 3.37 45') !136 1,1+86 +82" 45') *81 1 8127 45') !3*8 1 818" 45') .3! !,.2/016 2.87 45') !*88 !,1-/0!* 2.-" 45') 2-1 1,1*1+ 3*-7 45') -26 1,1!26 31!*" 45') 282 8,**/0!. ..!*7 45') !338 1 .3!." 45') -8* +,86/0!1 22!.7 45') 81* !,23/016 2.!6" 45') 662 1 .+!67 45') !816 1 2-*1" 45') +22 +,61/0!+ .2*17 45') 82+ .,!6/0!2 .3**7 #$%$&'() 6*! 1,11+1- 33
!"#$%&'( )"*+ ,-.&/0+ 12(+3*+&43%$0'(&52+6 7+86+93:+'+6
!"#!$%% &'() *$+%,-## ./0!1#-#%#!2!34566789:34%7;6<=#%7=9,>5&?@7966:#?@7::A:#*B78A547=A:A27,CD:%79D:#EE%#EB+*7:!F0G!!7==BE+E7:,=B*7;===7@6A37HA55:##7I>A*789:#*4B7;,6;789:24#3789:24#+
!"#%$% &'() E$EEE+%2!3 ./0!1*+B#32+*-*3E+23E+ 9D:B*3+2*
#*J#?$# &'() E$EE#B%#* ./0#*1!B%#3%%B-!+!2+%B2
6%5AK*7;4:%796=#7A:;I*74L<97;8MK78A63:B76A5#27&=6#+*7H@;@%378;4;27KND#47;K,K#3?47I4@*7K<6#**+7IC=5?785K;*7IAOM4?9*78;4:%7<I5@:7<I5@,7&9<#7465&4=37K4687AA<;%7K@A27><M!47=<=?>*:7A;C%7465&,M*!789:*24#E7@?&49I;#7D8374&4=*79D:#EE#%EBB27;8=4I%#7:A>?7K46:537:N=*B@#7K,;;9#7K,;;9*#@7:;A8=*7K<6*24*7C6::2@=#
%J#%$%# APQPR(F) E$EE*!BB? ./0%1##2!##+*2-##22#2B3* S9D:*+!#3?T?J%?$% APQPR(F) ?$#B,-#E ./0?1#+**222+%-#+*%EEB%* S9<I:EE*3ET?J%?$% APQPR(F) E$EE#E3#+ ./0?1#+*+#B%#+-#+*3#%+#3 SK&:?!+EET3"*#$% APQPR(F) #$*+,-E! ./031*#3*E%?B-**#?#!!# :A>I*@-48#7:A>I*47:3F0G!%7:A>I*@#%J#?$* APQPR(F) E$EE2B2?% ./0#%1?+B*?*!B-?3*E!+*! 6@#7<;K*@79=46276:@;@***J#*$# APQPR(F) *$*%,-E! ./0**1*3*+2#+#-*3?2B**E HI6M%7:**F0G%#
Nature Genetics: doi:10.1038/ng.2953
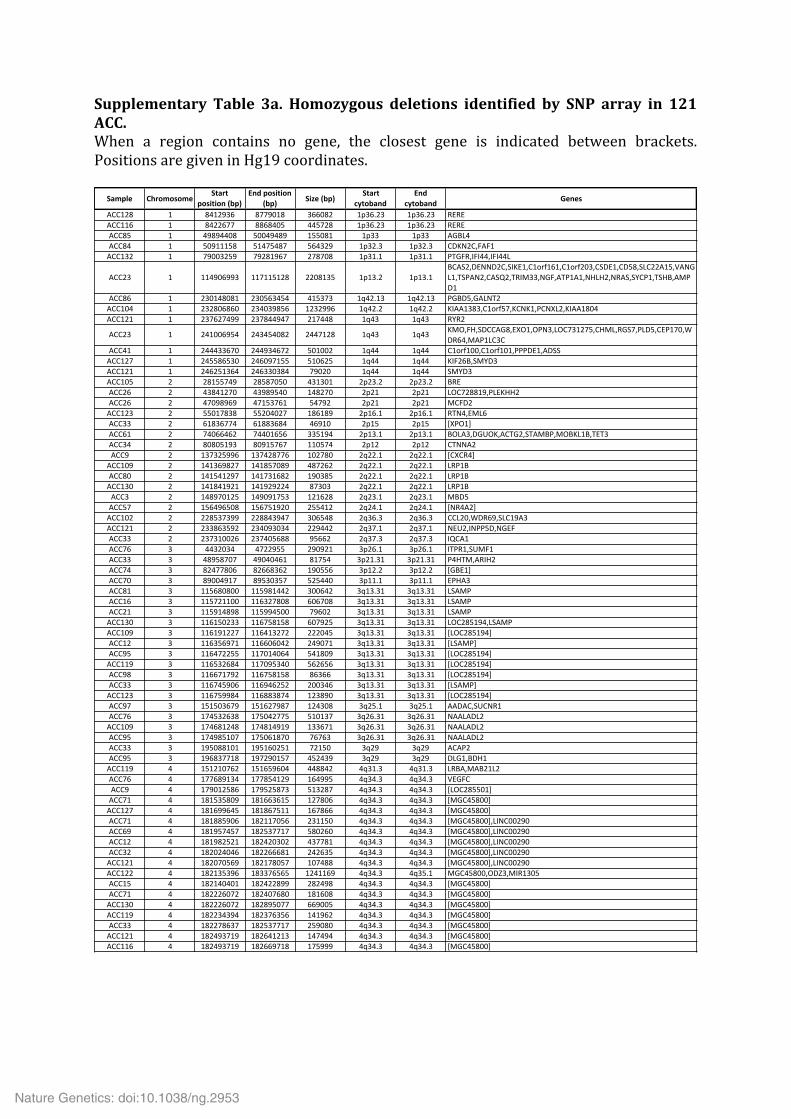
Supplementary Table 3a. Homozygous deletions identified by SNP array in 121 ACC. When a region contains no gene, the closest gene is indicated between brackets. Positions are given in Hg19 coordinates.
!"#$%& '()*#*+*#&!,"),-
$*+.,.*/-01$23/4-$*+.,.*/-
01$2 !.5&-01$2!,"),-
67,*1"/43/4-
67,*1"/4 8&/&+
!""#$% # %&#$'() %**'+#% ())+%$ #,()-$( #,()-$( ././!""##) # %&$$)** %%)%&+0 &&0*$% #,()-$( #,()-$( ././!""%0 # &'%'&&+% 0++&'&%' #00+%# #,(( #,(( !123&!""%& # 0+'###0% 0#&*0&%* 0)&($' #,($-( #,($-( "456$"78!8#!""#($ # *'++($0' *'$%#')* $*%*+% #,(#-# #,(#-# 9:18.7;8;&&7;8;&&3
!""$( # ##&'+)''( ##*##0#$% $$+%#(0 #,#(-$ #,#(-#2"!<$74/664$"7<;5/#7"#=>?#)#7"#=>?$+(7"<4/#7"40%7<3"$$!#07@!613#7:<9!6$7"!<A$7:.;B((76187!:9#!#76C3C$76.!<7<D"9#7:<C27!B94#
!""%) # $(+#&%+%# $(+0)(&0& �(*( #E&$-#( #E&$-#( 9124071!36:$!""#+& # $($%+)%)+ $(&+('%0) #$($'') #E&$-$ #E&$-$ 5;!!#(%(7"#=>?0*75"65#79"6F3$75;!!#%+&!""#$# # $(*)$*&'' $(*%&&'&* $#*&&% #E&( #E&( .D.$
!""$( # $&#++)'0& $&(&0&+%$ $&&*#$% #E&( #E&( 5BG78C7<4""!1%7/FG#7G96(73G"*(#$*07"CB37.1<*793407"/9#*+7H4.)&7B!9#3"("
!""&# # $&&&(()*+ $&&'(&)*$ 0+#++$ #E&& #E&& "#=>?#++7"#=>?#+#79994/#7!4<<!""#$* # $&00%)0(+ $&)+'*#00 0#+)$0 #E&& #E&& 5;8$)27<BD4(!""#$# # $&)$0#()& $&)((+(%& *'+$+ #E&& #E&& <BD4(!""#+0 $ $%#00*&' $%0%*+0+ &(#(+# $,$(-$ $,$(-$ 2./!""$) $ &(%&#$*+ &('%'0&+ #&%$*+ $,$# $,$# 3G"*$%%#'793/5CC$!""$) $ &*+'%')' &*#0(*)# 0&*'$ $,$# $,$# B"84$!""#$( $ 00+#*%(% 00$+&+$* #%)#%' $,#)-# $,#)-# .:6&7/B3)!""(( $ )#%()**& )#%%()%& &)'#+ $,#0 $,#0 IF9G#J!"")# $ *&+))&)$ *&&+#)0) ((0#'& $,#(-# $,#(-# 2G3!(741KG57!":1$7<:!B297BG253#27:/:(!""(& $ %+%+0#'( %+'#0*)* ##+0*& $,#$ $,#$ ":66!$!""' $ #(*($0'') #(*&$%**) #+$*%+ $E$$-# $E$$-# I"F".&J!""#+' $ #&#()'%$* #&#%0*+%' &%*$)$ $E$$-# $E$$-# 3.9#2!""%+ $ #�&#$'* #&#*(#)%$ #'+(%0 $E$$-# $E$$-# 3.9#2!""#(+ $ #&#%&#'$# #&#'$'$$& %*(+( $E$$-# $E$$-# 3.9#2!""( $ #&%'*+#$0 #&'+'#*0( #$#)$% $E$(-# $E$(-# B240!""0* $ #0)&')0+% #0)*0#'$+ $00&#$ $E$&-# $E$&-# I6.&!$J!""#+$ $ $$%0(*('' $$%%&('&* (+)0&% $E()-( $E()-( ""3$+7H4.)'7<3"#'!(!""#$# $ $((%)(0'$ $(&+'(+(& $$'&&$ $E(*-# $E(*-# 6/K$7;69904761/8!""(( $ $(*(#++$) $(*&+0)%% '0))$ $E(*-( $E(*-( ;A"!#!""*) ( &&($+(& &*$$'00 $'+'$# (,$)-# (,$)-# ;:9.#7<KB8#!""(( ( &%'0%*+* &'+&+&)# %#*0& (,$#-(# (,$#-(# 9&C:B7!.;C$!""*& ( %$&**%+) %$))%()$ #'+00) (,#$-$ (,#$-$ I12/#J!""*+ ( %'++&'#* %'0(+(0* 0$0&&+ (,##-# (,##-# /9C!(!""%# ( ##0)%+%++ ##0'%#&&$ (++)&$ (E#(-(# (E#(-(# 3<!B9!""#) ( ##0*$##++ ##)($*%+% )+)*+% (E#(-(# (E#(-(# 3<!B9!""$# ( ##0'#&%'% ##0''&0++ *')+$ (E#(-(# (E#(-(# 3<!B9!""#(+ ( ##)#0+$(( ##)*0%#0% )+*'$0 (E#(-(# (E#(-(# 3G"$%0#'&73<!B9!""#+' ( ##)#'#$$* ##)&#($*$ $$$+&0 (E#(-(# (E#(-(# I3G"$%0#'&J!""#$ ( ##)(0)'*# ##))+)+&$ $&'+*# (E#(-(# (E#(-(# I3<!B9J!""'0 ( ##)&*$$00 ##*+#&+)& 0&#%+' (E#(-(# (E#(-(# I3G"$%0#'&J!""##' ( ##)0($)%& ##*+'0(&+ 0)$)0) (E#(-(# (E#(-(# I3G"$%0#'&J!""'% ( ##))*#*'$ ##)*0%#0% %)()) (E#(-(# (E#(-(# I3G"$%0#'&J!""(( ( ##)*&0'+) ##)'&)$0$ $++(&) (E#(-(# (E#(-(# I3<!B9J!""#$( ( ##)*0''%& ##)%%(%*& #$(%'+ (E#(-(# (E#(-(# I3G"$%0#'&J!""'* ( #0#0+()*' #0#)$*'%* #$&(+% (E$0-# (E$0-# !!4!"7<K"6.#!""*) ( #*&0($)(% #*0+&$**0 0#+#(* (E$)-(# (E$)-(# 6!!3!43$!""#+' ( #*&)%#$&% #*&%#&'#' #(()*# (E$)-(# (E$)-(# 6!!3!43$!""'0 ( #*&'%0#+* #*0+)#%*+ *)*)( (E$)-(# (E$)-(# 6!!3!43$!""(( ( #'0+%%#+# #'0#)+$0# *$#0+ (E$' (E$' !"!9$!""'0 ( #')%(**#% #'*$'+#0* &0$&(' (E$' (E$' 431#724C#!""##' & #0#$#+*)$ #0#)0')+& &&%%&$ &E(#-( &E(#-( 3.2!7B!2$#3$!""*) & #**)%'#(& #**%0&#$' #)&''0 &E(&-( &E(&-( @/18"!""' & #*'+#$0%) #*'0$0%*( 0#($%* &E(&-( &E(&-( I3G"$%00+#J!""*# & #%#0(0%+' #%#))()#0 #$*%+) &E(&-( &E(&-( IB1"&0%++J!""#$* & #%#)'')&0 #%#%)*0## #)*%)) &E(&-( &E(&-( IB1"&0%++J!""*# & #%#%%0'+) #%$##*+0) $(##0+ &E(&-( &E(&-( IB1"&0%++J73;6"++$'+!"")' & #%#'0*&0* #%$0(**#* 0%+$)+ &E(&-( &E(&-( IB1"&0%++J73;6"++$'+!""#$ & #%#'%$0$# #%$&$+(+$ &(**%# &E(&-( &E(&-( IB1"&0%++J73;6"++$'+!""($ & #%$+$&+&) #%$$)))%# $&$)(0 &E(&-( &E(&-( IB1"&0%++J73;6"++$'+!""#$# & #%$+*+0)' #%$#*%+0* #+*&%% &E(&-( &E(&-( IB1"&0%++J73;6"++$'+!""#$$ & #%$#(0(') #%((*)0)0 #$&##)' &E(&-( &E(0-# B1"&0%++7G4L(7B;.#(+0!""#0 & #%$#&+&+# #%$&$$%'' $%$&'% &E(&-( &E(&-( IB1"&0%++J!""*# & #%$$$)+*$ #%$&+*)%+ #%#)+% &E(&-( &E(&-( IB1"&0%++J!""#(+ & #%$$$)+*$ #%$%'0+** ))'++0 &E(&-( &E(&-( IB1"&0%++J!""##' & #%$$(&('& #%$(*)(0) #&#')$ &E(&-( &E(&-( IB1"&0%++J!""(( & #%$$*%)(* #%$0(**#* $0'+%+ &E(&-( &E(&-( IB1"&0%++J!""#$# & #%$&'(*#' #%$)&#$#( #&*&'& &E(&-( &E(&-( IB1"&0%++J!""##) & #%$&'(*#' #%$))'*#% #*0''' &E(&-( &E(&-( IB1"&0%++J
Nature Genetics: doi:10.1038/ng.2953
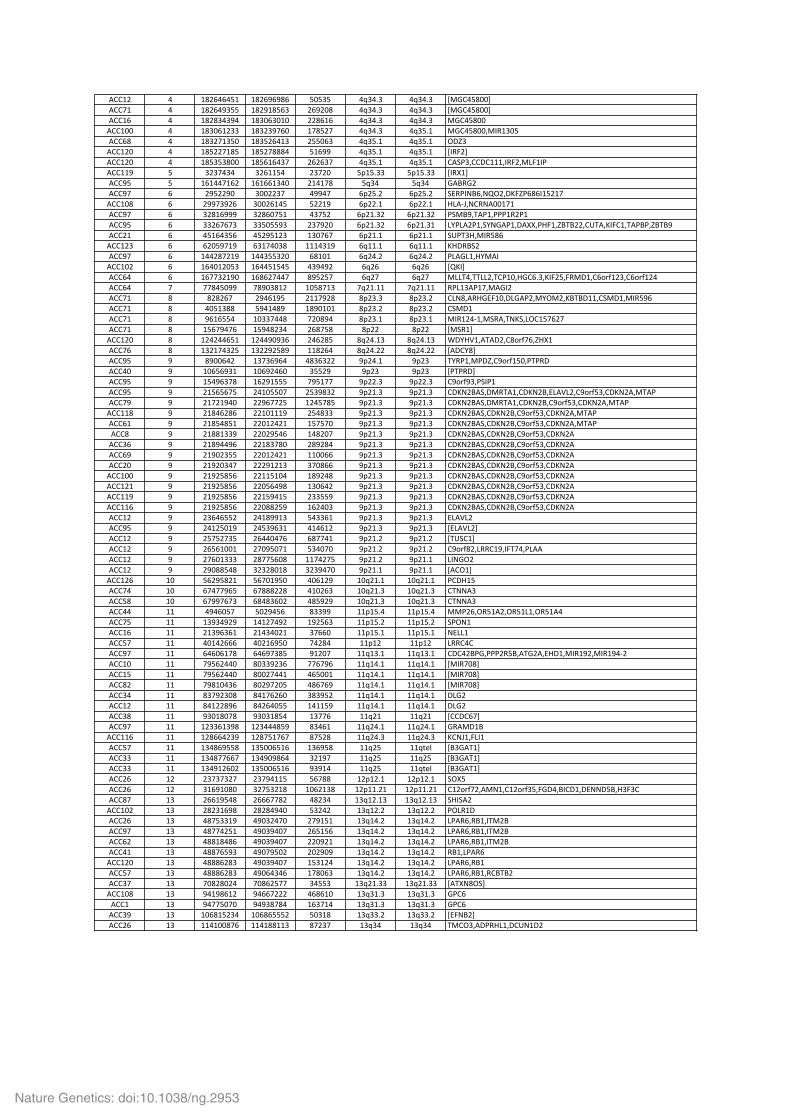
!""#$ % #&$'%'%(# #&$')')&' (*(+( %,+%-+ %,+%-+ ./0"%(&**1!""2# % #&$'%)+(( #&$)#&('+ $')$*& %,+%-+ %,+%-+ ./0"%(&**1!""#' % #&$&+%+)% #&+*'+*#* $$&'#' %,+%-+ %,+%-+ /0"%(&**!""#** % #&+*'#$++ #&+$+)2'* #2&($2 %,+%-+ %,+(-# /0"%(&**3/45#+*(!""'& % #&+$2#+(* #&+($'%#+ $((*'+ %,+(-# %,+(-# 678+!""#$* % #&($$2#&( #&($2&&&% (#')) %,+(-# %,+(-# .459$1!""#$* % #&(+(+&** #&('#'%+2 $'$'+2 %,+(-# %,+(-# "!:;+3""7"###3459$3/<9#4;!""##) ( +$+2%+% +$'##(% $+2$* (=#(-++ (=#(-++ .45>#1!"")( ( #'#%%2#'$ #'#''#+%* $#%#2& (,+% (,+% 0!?50$!"")2 ' $)($$)* +**$$+2 %))%2 '=$(-$ '=$(-$ :@5;4A?'3AB6$37C98;'&'4#($#2!""#*& ' $))2+)$' +**$'#%( ($$#) '=$$-# '=$$-# D<!EF3A"5A!**#2#!"")2 ' +$&#'))) +$&'*2(# %+2($ '=$#-+$ '=$#-+$ ;:/?)3G!;#3;;;#5$;#!"")( ' ++$'2'2+ ++(*(()+ $+2)$* '=$#-+$ '=$#-+# <H;<!$;#3:HA0!;#37!>>3;D9#38?G?$$3"IG!3C49"#3G!;?;38?G?)!""$# ' %(#'%+(' %($)(#$+ #+*2'2 '=$#-# '=$#-# :I;G+D3/45(&'!""#$+ ' '$*()2#) '+#2%*+& ###%+#) ',##-# ',##-# CD75?:$!"")2 ' #%%$&2$#) #%%+((+$* '&#*# ',$%-$ ',$%-$ ;<!0<#3DH/!4!""#*$ ' #'%*#$*(+ #'%%(#(%( %+)%)$ ',$' ',$' .BC41!""'% ' #'22+$#)* #'&'$2%%2 &)($(2 ',$2 ',$2 /<<G%3GG<<$3G";#*3D0"'-+3C49$(395/7#3"'JKL#$+3"'JKL#$%!""'% 2 22&%(*)) 2&)*+&#$ #*(&2#+ 2,$#-## 2,$#-## 5;<#+!;#23/!04$!""2# & &$&$'2 $)%'#)( $##2)$& &=$+-+ &=$+-$ "<A&3!5D0@9#*37<0!;$3/H6/$3C?G?7##3":/7#3/45()'!""2# & %*(#+&& ()%#%&) #&)*#*# &=$+-$ &=$+-$ ":/7#!""2# & )'#'((% #*++2%%& 2$*&)% &=$+-# &=$+-# /45#$%E#3/:5!3GAC:3<6"#(2'$2!""2# & #('2)%2' #()%&$+% $'&2(& &=$$ &=$$ ./:5#1!""#$* & #$%$%%'(# #$%%)*)+' $%'$&( &,$%-#+ &,$%-#+ M7HDN#3!G!7$3"&JKL2'38D>#!""2' & #+$#2%+$( #+$$)$(&) ##&$'% &,$%-$$ &,$%-$$ .!7"H&1!"")( ) &)**'%$ #+2+')'% %&+'+$$ )=$%-# )=$+ GH5;#3/;783")JKL#(*3;G;57!""%* ) #*'(')+# #*')$%'* +(($) )=$+ )=$+ .;G;571!"")( ) #(%)'+2& #'$)#((( 2)(#22 )=$$-+ )=$$-+ ")JKL)+3;:4;#!"")( ) $#('('2( $%#*((*2 $(+)&+$ )=$#-+ )=$#-+ "7CA$?!:37/5G!#3"7CA$?3@<!N<$3")JKL(+3"7CA$!3/G!;!""2) ) $#2$#)%* $$)'22$( #$%(2&( )=$#-+ )=$#-+ "7CA$?!:37/5G!#3"7CA$?3")JKL(+3"7CA$!3/G!;!""##& ) $#&%'$&' $$#*###) $(%&++ )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!3/G!;!""'# ) $#&(%&(# $$*#$%$# #(2(2* )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!3/G!;!""& ) $#&&#++) $$*$)(%' #%&$*2 )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!!""+' ) $#&)%%)' $$#&+2&* $&)$&% )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!!""') ) $#)*$+(( $$*#$%$# ##**'' )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!!""$* ) $#)$*+%2 $$$)#$#+ +2*&'' )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!!""#** ) $#)$(&(' $$##(#*% #&)$%& )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!!""#$# ) $#)$(&(' $$*('%)& #+*'%$ )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!!""##) ) $#)$(&(' $$#()%#( $++(() )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!!""##' ) $#)$(&(' $$*&&$() #'$%*+ )=$#-+ )=$#-+ "7CA$?!:3"7CA$?3")JKL(+3"7CA$!!""#$ ) $+'%'(($ $%#&))#+ (%++'# )=$#-+ )=$#-+ @<!N<$!"")( ) $%#$(*#) $%(+)'+# %#%'#$ )=$#-+ )=$#-+ .@<!N<$1!""#$ ) $(2($2+( $'%%*%2' '&22%# )=$#-$ )=$#-$ .GI:"#1!""#$ ) $'('#**# $2*)(*2# (+%*2* )=$#-$ )=$#-$ ")JKL&$3<55"#)349G2%3;<!!!""#$ ) $2'*#+++ $&22('*& ##2%$2( )=$#-$ )=$#-# <4A06$!""#$ ) $)*&&(%& +$+$&*#& +$+)%2* )=$#-# )=$#-# .!"6#1!""#$' #* ('$)(&$# ('2*#)(* %*'#$) #*,$#-# #*,$#-# ;"7D#(!""2% #* '2%22)'( '2&&&$$& %#*$'+ #*,$#-+ #*,$#-+ "GAA!+!""(& #* '2))2'2+ '&%&+'*$ %&()$) #*,$#-+ #*,$#-+ "GAA!+!""%% ## %)%'*(2 (*$)%(' &++)) ##=#(-% ##=#(-% //;$'365(#!$365(#<#365(#!%!""2( ## #+)+%)$) #%#$2%)$ #)$('+ ##=#(-$ ##=#(-$ :;6A#!""#' ## $#+)'+'# $#%+%*$# +2''* ##=#(-# ##=#(-# A@<<#!""(2 ## %*#%$''' %*$#')(* 2%$&% ##=#$ ##=#$ <55"%"!"")2 ## '%'*'#2& '%')2+&( )#$*2 ##,#+-# ##,#+-# "7"%$?;03;;;$5(?3!G0$!3@D7#3/45#)$3/45#)%E$!""#* ## 2)('$%%* &*++)$+' 22'2)' ##,#%-# ##,#%-# ./452*&1!""#( ## 2)('$%%* &**$2%%# %'(**# ##,#%-# ##,#%-# ./452*&1!""&$ ## 2)&#*%+' &*$)2$*( %&'2') ##,#%-# ##,#%-# ./452*&1!""+% ## &+2)$+*& &%#2'$'* +&+)($ ##,#%-# ##,#%-# 7<0$!""#$ ## &%#$$&)' &%$'%*(( #%##() ##,#%-# ##,#%-# 7<0$!""+& ## )+*#&*2& )+*+#&(% #+22' ##,$# ##,$# .""7"'21!"")2 ## #$++'#+)& #$+%%%&() &+%'# ##,$%-# ##,$%-# 05!/7#?!""##' ## #$&''%$+) #$&2(#2'2 &2($& ##,$%-+ ##,$%-+ C"AF#39<4#!""(2 ## #+%&')((& #+(**'(#' #+')(& ##,$( ##,OPQ .?+0!G#1!""++ ## #+%&22''2 #+%)*)&'% +$#)2 ##,$( ##,$( .?+0!G#1!""++ ## #+%)#$'*$ #+(**'(#' )+)#% ##,$( ##,OPQ .?+0!G#1!""$' #$ $+2+2+$2 $+2)%##( ('2&& #$=#$-# #$=#$-# :6>(!""$' #$ +#')#*&* +$2(+$#& #*'$#+& #$=##-$# #$=##-$# "#$JKL2$3!/A#3"#$JKL+(3907%3?4"7#37@AA7(?3D+9+"!""&2 #+ $''#)(%& $'''22&$ %&$+% #+,#$-#+ #+,#$-#+ :D4:!$!""#*$ #+ $&$+#')& $&$&%)%* (+$%$ #+,#$-$ #+,#$-$ ;6<5#7!""$' #+ %&2(++#) %)*+$%2* $2)#(# #+,#%-$ #+,#%-$ <;!5'35?#34G/$?!"")2 #+ %&22%$(# %)*+)%*2 $'(#(' #+,#%-$ #+,#%-$ <;!5'35?#34G/$?!""'$ #+ %&&#&%&' %)*+)%*2 $$*)$# #+,#%-$ #+,#%-$ <;!5'35?#34G/$?!""%# #+ %&&2'()+ %)*2)(*$ $*$)*) #+,#%-$ #+,#%-$ 5?#3<;!5'!""#$* #+ %&&&'$&+ %)*+)%*2 #(+#$% #+,#%-$ #+,#%-$ <;!5'35?#!""(2 #+ %&&&'$&+ %)*'%+%' #2&*'+ #+,#%-$ #+,#%-$ <;!5'35?#35"?G?$!""+2 #+ 2*&$&*$% 2*&'$(22 +%((+ #+,$#-++ #+,$#-++ .!G>A&6:1!""#*& #+ )%#)&'#$ )%''2$$$ %'&'#* #+,+#-+ #+,+#-+ 0;"'!""# #+ )%22(*2* )%)+&2&% #'+2#% #+,+#-+ #+,+#-+ 0;"'!""+) #+ #*'&#($+% #*'&'((($ (*+#& #+,++-$ #+,++-$ .@9A?$1!""$' #+ ##%#**&2' ##%#&&##+ &2$+2 #+,+% #+,+% G/"6+3!7;5D<#37"IA#7$
Nature Genetics: doi:10.1038/ng.2953
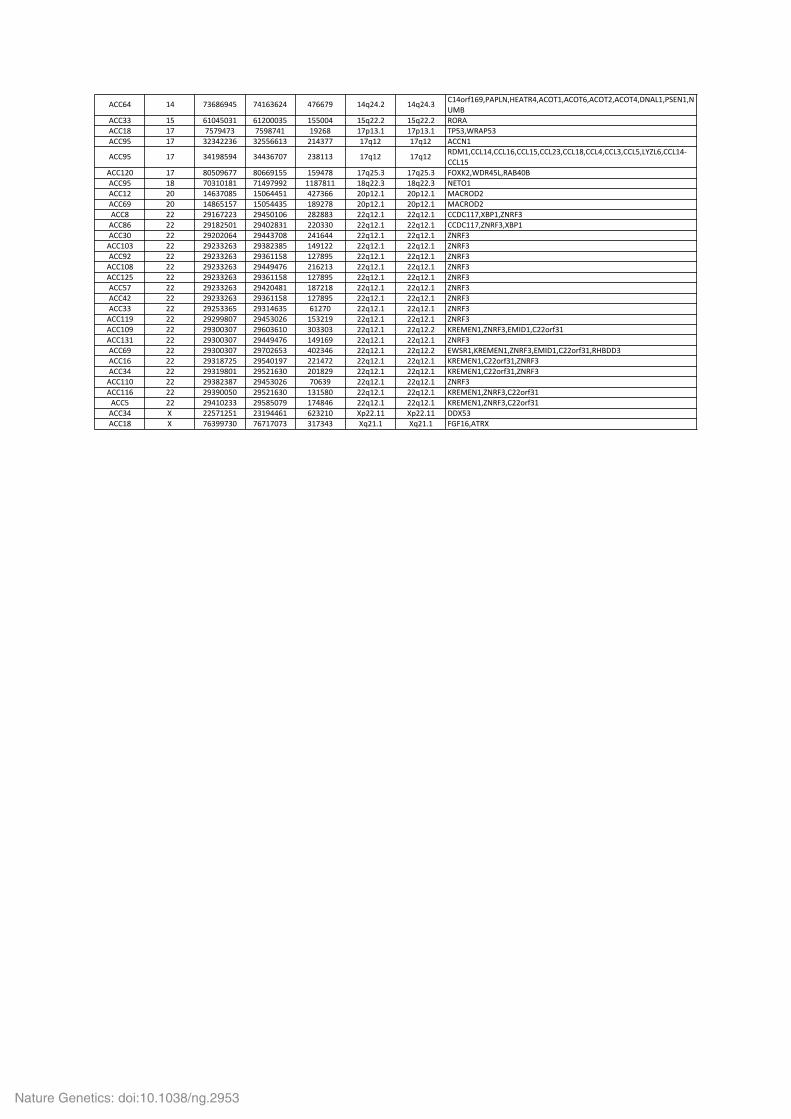
!""#$ %$ &'#(#)$* &$%#'#+$ $&##&) %$,+$-+ %$,+$-' "%$./0%#)12!234156!78$1!"97%1!"97#1!"97+1!"97$1:4!3%12;64%14<=>
!""'' %* #%?$*?'% #%+???'* %**??$ %*,++-+ %*,++-+ 898!!""%( %& &*&)$&' &*)(&$% %)+#( %&@%'-% %&@%'-% 72*'1A8!2*'!"")* %& '+'$++'# '+**##%' +%$'&& %&,%+ %&,%+ !""4%
!"")* %& '$%)(*)$ '$$'#&?& +'(%%' %&,%+ %&,%+ 8:=%1""3%$1""3%#1""3%*1""3+'1""3%(1""3$1""3'1""3*13BC3#1""3%$D""3%*
!""%+? %& (?*?)#&& (?##)%** %*)$&( %&,+*-' %&,+*-' E9FG+1A:8$*318!>$?>!"")* %( &?'%?%(% &%$)&))+ %%(&(%% %(,++-' %(,++-' 4679%!""%+ +? %$#'&?(* %*?#$$*% $+&'## +?@%+-% +?@%+-% =!"89:+!""#) +? %$(#*%*& %*?*$$'* %()+&( +?@%+-% +?@%+-% =!"89:+!""( ++ +)%#&++' +)$*?%?# +(+((' ++,%+-% ++,%+-% "":"%%&1F>2%1C48E'!""(# ++ +)%(+*?% +)$?+('% ++?''? ++,%+-% ++,%+-% "":"%%&1C48E'1F>2%!""'? ++ +)+?+?#$ +)$$'&?( +$%#$$ ++,%+-% ++,%+-% C48E'!""%?' ++ +)+''+#' +)'(+'(* %$)%++ ++,%+-% ++,%+-% C48E'!"")+ ++ +)+''+#' +)'#%%*( %+&()* ++,%+-% ++,%+-% C48E'!""%?( ++ +)+''+#' +)$$)$&# +%#+%' ++,%+-% ++,%+-% C48E'!""%+* ++ +)+''+#' +)'#%%*( %+&()* ++,%+-% ++,%+-% C48E'!""*& ++ +)+''+#' +)$+?$(% %(&+%( ++,%+-% ++,%+-% C48E'!""$+ ++ +)+''+#' +)'#%%*( %+&()* ++,%+-% ++,%+-% C48E'!""'' ++ +)+*''#* +)'%$#'* #%+&? ++,%+-% ++,%+-% C48E'!""%%) ++ +)+))(?& +)$*'?+# %*'+%) ++,%+-% ++,%+-% C48E'!""%?) ++ +)'??'?& +)#?'#%? '?''?' ++,%+-% ++,%+-+ G86=64%1C48E'16=H:%1"++./0'%!""%'% ++ +)'??'?& +)$$)$&# %$)%#) ++,%+-% ++,%+-% C48E'!""#) ++ +)'??'?& +)&?+#*' $?+'$# ++,%+-% ++,%+-+ 6A;8%1G86=64%1C48E'16=H:%1"++./0'%185>::'!""%# ++ +)'%(&+* +)*$?%)& ++%$&+ ++,%+-% ++,%+-% G86=64%1"++./0'%1C48E'!""'$ ++ +)'%)(?% +)*+%#'? +?%(+) ++,%+-% ++,%+-% G86=64%1"++./0'%1C48E'!""%%? ++ +)'(+'(& +)$*'?+# &?#') ++,%+-% ++,%+-% C48E'!""%%# ++ +)')??*? +)*+%#'? %'%*(? ++,%+-% ++,%+-% G86=64%1C48E'1"++./0'%!""* ++ +)$%?+'' +)*(*?&) %&$($# ++,%+-% ++,%+-% G86=64%1C48E'1"++./0'%!""'$ F ++*&%+*% +'%)$$#% #+'+%? F@++-%% F@++-%% ::F*'!""%( F &#'))&'? &#&%&?&' '%&'$' F,+%-% F,+%-% EIE%#1!78F
Nature Genetics: doi:10.1038/ng.2953
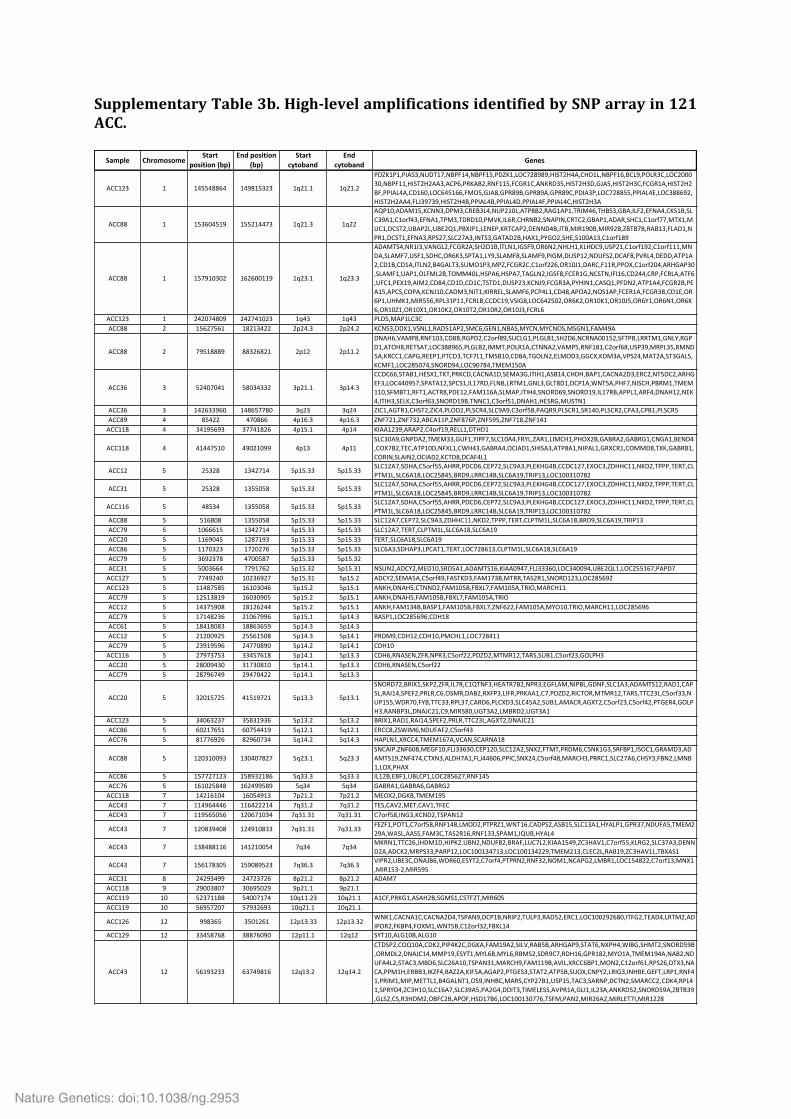
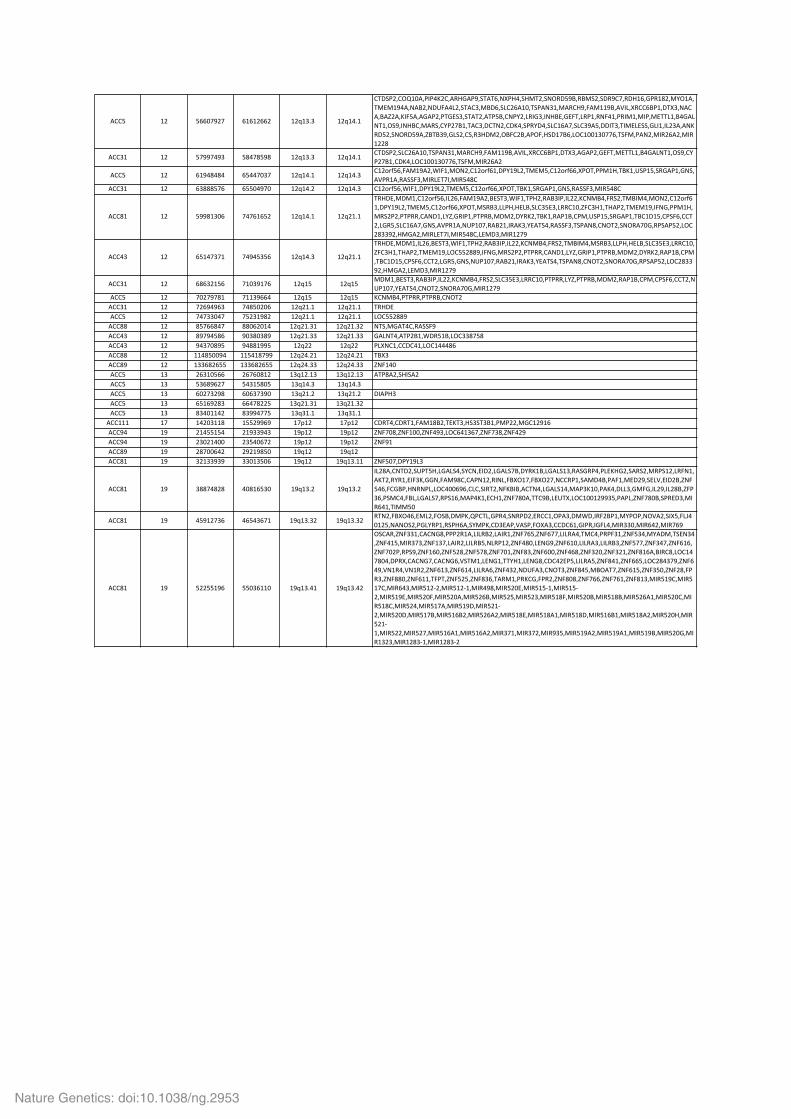
Supplementary Table 3b. High-‐level amplifications identified by SNP array in 121 ACC.
!"#$%& '()*#*+*#&!,"),-
$*+.,.*/-01$23/4-$*+.,.*/-
01$2!,"),-
56,*1"/43/4-
56,*1"/4 7&/&+
!""#$% # #&''&(()& #&*(#'%$% #+$#,# #+$#,$
-./0#-#1-2!3%145.6#7148-9#&148-9#'1-./0#1:;"7$(*(*1<236$<&!1"<.#:148-9#)18":*1-;:=%"1:;"$>>>%>148-9##1<236$<$!!%1!"-)1-=0!8$1=49##'19"?=#"1!40=.%'1<236$<%.1?@!'1<236$<%"19"?=#!1<236$<$891--2!:&!1".#)>1:;")&'#))19A;'1?@!(1?-=(*81?-=(*!1?-=(*"1-.2!%-1:;"7$((''1--2!:&B1:;"%(()*$1<236$<$!!&19:@%*7%*1<236$<&81--2!:&81--2!:&.1--2!:&91--2!:&"1<236$<%!
!""(( # #'%)>&'#* #''$#&&7% #+$#,% #+$$
!C-#>1!.!A#'10"44%1.-A%1"=B8%:&145-$#>:1!6-(8$1=!?#!-#16=2A&)16<83%1?8!12:9$1B94!&1"03#813:"%*!#1"#DEF&%1B94!#16-A%16.=.#>1-AG012:)=1"<=48$134!-241"=6"$1?8!-#1!.!=13<"#1"#DEF771A6H#1A5"#1."36$158!-$:158B$C#1-8H2-#1:B4B-10=6"!-$1.B44.&81@681A2=#*>81A2=*$81/868781=!8#%19:!.#14-=#1."36#1B94!%1=-3$713:"$7!%12463%1?!6!.$81<!H#1-I?;$13<B13#>>!#%1"#DEF#(*
!""(( # #'7*#>%>$ #)$)>>##* #+$%,# #+$%,%
!.!A63&14=#2%1G!4?:$19"?=$!13<$.#8126:4#12?39*1;=)4$14<:<#10:<."*153-$#1"#DEF#*$1"#DEF###1A4.!13:!A971539#13.<"1;=)0%13-6!#1:I*13:!A9(13:!A9*1-2?A1.53-#$14.593$1."!9(1-G=:&1.B..1!6-#!$1".#81".#!126:4$18&?!:6%135A;#-%1A-/19"?=$"1"#DEF$$)1;=#>@#1.!="19##=1--;H1"#DEF$>&1!=<?!-%>13:!A9#15!-#1;:9A:$816;AA&>:1<3-!)1<3-!716!?:4$12?39(19"B=#?14"3641292#)1".$&&1"=-19"=:!1!69)159"#1-BH#*1!2A$1".(&1".#.1".#"1636.#1.53-$%10"4@*19"?=%!1-I<24#1"!3C#1-9.4$1!6-#!&19"?=$81-B!#'1!-"31";-!10"4@#>1"!.A%1426#102==B:13:!A9)1-"-&:#1".&(1!-;!$14;3#!-19"B=#!19"?=%81".#B1;=)-#15<A0#1A2='')1=-:%#-##19"=:81""."#*1G32?(1:;")&$'>$1;=)0$1;=#>0#1;=#>@'1;=)I#1;=)4#1;=)0)1;=#>/#1;=#>H#1;=#>0$1;=#>6$1;=#>=$1;=#>@%19"=:)
!""#$% # $&$>7&(>* $&$7&#>$% #+&% #+&% -:.'1A!-#:"%"!""(( $ #')$7')# #($#%&$$ $J$&,% $J$&,$ 0"43%1..H#1G34:#1=!.'#!-$13A")1?B4#148!31AI"41AI"4;31A3?4#19!A&*!
!""(( $ 7*'#(((* ((%$)($# $J#$ $J##,$
.4!<)1G!A-(1=49#>%1".(81=?-.$1"$DEF(*135":?#1-:?:8#13<$.)14"=4!>>#'$1396-81:==6A#1?4:I1=?-
.#1!6;<(1=B63!61:;"%((*)'1-:?:8$12AA61-;:=#!1"644!$1G!A-'1=49#(#1"$DEF)(153-%*1A=-:%'1=A4.'!10=""#1"!-?1=BB-#1-6".%16"97:#16A38#>1".(!16?;:4$1B:A;.%1??"H10.A%!1G-3$&1A!6$!136%?!:'10"A9#1:;"$('>7&134;=.*&1:;"*>7(&16ABA#'>!
!""%) % '$&>7>&# '(>%&%%$ %J$#,# %J#&,%
""."))136!8#1<B3H#16061-=0".1"!"4!#.13BA!%?1262<#1!38#&1"<.<18!-#1"!"4!$.%1B="$146'."$1!=<?B9%1:;"&&>*'713-!6!#$13-"3#12:#7=.19:481:=6A#1?4:%1?:6(.#1."-#!1K46'!1-<971423"<1-8=A#16ABA##>139A86#1=96#1!"6=(1-.B#$19!A##)!13:A!-1262<&134;=.)*134;=.#*12:#7=81!--:#1!=9&1.4!<#$14B0&1262<%13B:01"%DEF)%134;=.#*81644"#1"%DEF'#1.4!<#1<B3=?1A5364#
!""%) % #&$)%%*)> #&()'77(> %+$% %+$& /2"#1!?6=#1"<36$1/2"&1-:;.$1-:3"=&13:"*!*1"%DEF'(1-!C=*1-:3"=#13=#&>1-:3"=$1"-!%1"-8#1-:3"='!""(* & ('&$$ &7>()) &J#),% &J#),% /497$#1/497%$1!8"!##-1/49(7)-1/49'*'1/497#(1/49#&#!""##( & %&#*')*% %77&#($) &J#',# &J#& 02!!#$%*1!=!-$1"&DEF#*1=B::#1.6<.#
!""##( & &#&&7'#> &*>$#>** &J#% &J##3:"%>!*1?4-.!$16ABA%%1?59#1I2-9713:"#>!&19=I:1/!=#1:2A"<#1-<;H$81?!8=!$1?!8=?#1"4?!#18B4.&1";H78$16B"1!6-#>.149H:#1"K<&%1?!8=!&1;"2!.#13<23!%1!6-(!#142-!:#1?=H"=#1";AA.(16H01?!8=8#1";=2413:!24$1;"2!.$10"6.(1."!9&:#
!""#$ ' $'%$( #%&$7#& 'J#',%% 'J#',%% 3:"#$!713.<!1"'DEF''1!<==1-.".)1"B-7$13:"*!%1-:B0<?&81""."#$71BH;"%1/.<<"##140.$16---16B=61":-6A#:13:")!#(1:;"$'(&'18=.*1:=="#&813:")!#*16=2-#%1:;"#>>%#>7($
!""%# ' $'%$( #%''>'( 'J#',%% 'J#',%% 3:"#$!713.<!1"'DEF''1!<==1-.".)1"B-7$13:"*!%1-:B0<?&81""."#$71BH;"%1/.<<"##140.$16---16B=61":-6A#:13:")!#(1:;"$'(&'18=.*1:=="#&813:")!#*16=2-#%1:;"#>>%#>7($
!""##) ' &('%& #%''>'( 'J#',%% 'J#',%% 3:"#$!713.<!1"'DEF''1!<==1-.".)1"B-7$13:"*!%1-:B0<?&81""."#$71BH;"%1/.<<"##140.$16---16B=61":-6A#:13:")!#(1:;"$'(&'18=.*1:=="#&813:")!#*16=2-#%1:;"#>>%#>7($
!""(( ' '#)(>( #%''>'( 'J#',%% 'J#',%% 3:"#$!71"B-7$13:"*!%1/.<<"##140.$16---16B=61":-6A#:13:")!#(18=.*13:")!#*16=2-#%!""7* ' #>)))#' #%&$7#& 'J#',%% 'J#',%% 3:"#$!716B=61":-6A#:13:")!#(13:")!#*!""$> ' ##)*>&' #$(7#*% 'J#',%% 'J#',%% 6B=613:")!#(13:")!#*!""() ' ##7>%$% #7$>$7) 'J#',%% 'J#',%% 3:")!%13.<!-%1:-"!6#16B=61:;"7$()#%1":-6A#:13:")!#(13:")!#*!""7* ' %)*$%7( &7>>'(7 'J#',%% 'J#',%$!""%# ' '>>%))& 77*#7)$ 'J#',%$ 'J#',%# 4354$1!."I$1AB.#>13=.'!#1!.!A63#)102!!>*&719:@%%%)>1:;"%&>>*&158B$C:#1:;"$''#)71-!-.7!""#$7 ' 77&*$&> #>$%)*$7 'J#',%# 'J#',$ !."I$13BA!'!1"'DEF&*19!360.%19!A#7%81A6==16!3$=#134;=.#$%1:;"$(')*$!""#$% ' ##&(7'(' #)#>%>&) 'J#',$ 'J#',# !40<1.4!<'1"644.$19!A#>'8198H:719!A#>'!16=2;1A!="<##!""7* ' #$'#%(#* #)>%>*>' 'J#',$ 'J#',# !40<1.4!<'19!A#>'8198H:719!A#>'!16=2;!""#$ ' #&%7'*>( #(#$)$&& 'J#',$ 'J#',# !40<19!A#%&818!3-#19!A#>'8198H:71/49)$$19!A#>'!1AI;#>16=2;1A!="<##1:;"$(')*)!""7* ' #7#&($%) $#>)7**) 'J#',# 'J#&,% 8!3-#1:;"$(')*)1".<#(!"")# ' #(&#(>(% #(()%)'* 'J#&,% 'J#&,%!""#$ ' $#$>>*$' $'')#'>( 'J#&,% 'J#&,# -=.A*1".<#$1".<#>1-A"<:#1:;"7$(&##!""7* ' $%*#*'*) $&77>(*> 'J#&,$ 'J#&,# ".<#>!""##) ' $7*7%7'% %%&'7)#( 'J#&,# 'J#%,% ".<)1=4!3B41/9=14-=%1"'DEF$$1-./.$1A6A=#$16!=31358#1"'DEF$%1?;:-<%!""$> ' $(>>*&%> %#7%>(#> 'J#&,# 'J#%,% ".<)1=4!3B41"'DEF$$!""7* ' $(7*)7&* $*&7>&$$ 'J#&,# 'J#%,%
!""$> ' %$>#'7$' &#'#*7$# 'J#%,% 'J#%,#
34;=.7$18=2H#130-$1/9=12:7=1"#C649%1<B!6=78$14-=%1B?9:!A142-8:1?.4913:"#!%1!.!A63#$1=!.#1"!-3:1=!2#&13-B9$1-=:=1")1;3A=1.!8$1=H9-%1:29=1-=0!!#1"71-./.$1=2"6;=1A6A=#$16!=3166"$%:1"'DEF%%145-#''1K.=7>19I8166"%%1=-:%71"!=.)1-:"H.%13:"&'!$1358#1!A!"=1!?H6$1"'DEF$%1"'DEF&$1-6?B=&1?;:-<%1=!48-%:1.4!@"$#1"*1A2='(>15?6%!$1:A8=.$15?6%!#
!""#$% ' %&>)%$%7 %'(%#*%) 'J#%,$ 'J#%,$ 8=2H#1=!.#1=!2#&13-B9$1-=:=166"$%:1!?H6$1.4!@"$#!""() ' )>$#7)'# )>7'&&#* '+#$,# '+#$,# B=""(1/3K2A)14.59!9$1"'DEF&%!""7) ' (#77)*$) ($*)>7%& '+#&,$ '+#&,% <!-:4#1H=""&16ABA#)7!1G"!413"!=4!#(
!""(( ' #$>%#>>*% #%>&>7($7 '+$%,# '+$%,%34"!2-1/49)>(1AB?9#>19:@%%)%>1"B-#$>13:"#$!$134H$196A61-=.A)1"340#?%13=98-#123;"#1?=!A.%1!.!A63#*1/49&7&1"6H4%1!:.<7!#19:@&&)>)1--2"134H$&1"'DEF&(1A!="<%1-=="#13:"$7!)1"<3I%1984$1:A48#1:;H1-<!H
!""() ' #'77$7#$% #'(*%$#() '+%%,% '+%%,% 2:#$81B89#158:"-#1:;"$(')$71=49#&'!""7) ' #)#>$'(&( #)$&**'(* '+%& '+%& ?!8=!#1?!8=!)1?!8=?$!""##( 7 #&$#)#>& #)>'&*#% 7J$#,$ 7J$#,$ AB;H$1.?0816ABA#*'!""&% 7 ##&*)&&&) ##)&$$$#& 7+%#,$ 7+%#,$ 6B31"!G$1AB61"!G#169B"!""&% 7 ##*')'>') #$>)7#>%& 7+%#,%# 7+%#,%# "7DEF'(124?%10"4.$163-!4#$
!""&% 7 #$>(%*&>( #$&*#>(%% 7+%#,%# 7+%#,%% 9B/9#1-;6#1"7DEF'(1=49#&(1:A;.$1-6-=/#1K46#)1"!.-3$1!38#'13:"#%!#1<I!:-#1?-=%714.59!'16ABA$$*!1K!3:1!!3319!A%"16!3$=#)1=49#%%13-!A#12C581<I!:&
!""&% 7 #%(&((##) #&#$#>>'& 7+%& 7+%& A0=4#166"$)1@<.A#.1<2-0$1584$14.598$18=!91:5"7:$102!!#'&*1/"%<!G#1"7DEF''10:=?$13:"%7!%1.B44.$!1!."0$1A=-3%%1-!=-#$1:;"#>>#%&7#%1:;"#>>#%&$$*16ABA$#%1":B"$:1=!8#*1/"%<!G#:168H!3#
!""&% 7 #')#7(%>' #'*>(*'$% 7+%),% 7+%),% G2-=$158B%"1.4!@8)1K.=)>1B3I6$1"7DEF&1-6-=4$1=49%$14;A#14"!-?$1:A8=#1:;"#'&($$1"7DEF#%1A4H#1A2=#'%L$1A2='*'
!""%# ( $&$*%&** $&7$%7$) (J$#,$ (J$#,$ !.!A7!""##( * $*>>%(>7 %>)*'>$* *J$#,# *J$#,#!""##* #> '$%7##(( '&>>7#7& #>+##,$% #>+$#,# !#"91-=0?#1!3!<$813?A3#1"369$61A2=)>'!""##* #> ')*'7$>7 '7*%$)*% #>+$#,# #>+$#,#
!""#$) #$ **(%)' %'>#$)# #$J#%,%% #$J#%,%$ K40#1"!"4!#"1"!"4!$.&163-!4*1."-#814=2-$165:-%1=!.'$1B="#1:;"#>>$*$)(>1269?$16B!.&1:=6A$1!.2-;=$1908-&19;HA#1K46'81"#$DEF%$198H:#&
!""#$* #$ %%&'(7)( %((7)>*> #$J##,# #$+#$ 3I6#>1!:?#>81!:?#>
!""&% #$ ')#*%$%% )%7&*(#) #$+#%,$ #$+#&,$
"6.3-$1";C#>!1".0$1-2-&0$"1.?0!19!A#*!$132:G1=!8'81!=<?!-*136!6)14H-<&1K28?13<A6$134;=.'*81;=A.:$1.4!@"#&1AA-#*1B3I6#1AI:)81AI:)1=8A3$13.=*"71=.<#)1?-=#($1AI;#!16ABA#*&!14!8$14.59!&:$136!"%1A8.)13:"$)!#>163-!4%#1A!="<*19!A##*81!G2:1H="")8-#1A;4$1"#$DEF)#1=-3$)1.6H%14!"!1--A#<1B=88%120/9&18!/$!1029'!1!?!-$1-6?B3%136!6$1!6-'8135;H1"4-I$1:=2?%124<8B1?B961:=-#1=49-=2A#1A2-1AB66:#18&?!:46#1;3*124<8"1A!=31"I-$78#153-#'16!"%13!=4-1."64$13A!=""$1".0&1=-:
-=I.&1/"%<#>13:"#)!713:"%*!'1-!$?&1..26%162AB:B331!G-=#!1?:2#12:$%!1!40=.'$134;=.'*!1/868%*1?:3$1"31=%<.A$1;89"$81!-;91<3.#78)1:;"#>>#%>77)1639A1-!4$1A2=$)!$1A2=:B6721A2=#$$(
Nature Genetics: doi:10.1038/ng.2953
!""# $% #&&'()%( &$&$%&&% $%*$+,+ $%*$-,$
"./01%2"34$'!2151-6%"2!789!1)20.!.&2:;18-208<.%20:37/#)=27=<0%20/7)"(27/8$&2917$>%2<?3$!2.<@<$)-!2:!=%2:/AB!-C%20.!"+2<=/&20C"%&!$'2.01!:+$2<!7"8)2B!<$$)=2!D5C2;7""&=1$2/.;+2:!"!2=!E%!265B#!2!9!1%21.9@0+20.!.%2!.1#=2":1?%2C759+25:8=@[email protected]$27:B-$2175<$2<512<@..C$2=-9!C:.$230)25:8="2<!702"?1%(=$2.!"+2/".:%2"/6-2017?/-20C"$&!(20C"+)!#2//5.+2.5<@C@0029C5$25C%+!2!:67/#%20:37/#)!2E=.=+)29C0%2"027+8/<%23=B"%=2!13B280/$(=&2C3"$''$+'((&2.0B<21!:%2<57%&!%2<57$%%>
!""+$ $% #())(-)+ #>-(>#)> $%*$+,+ $%*$-,$ "./01%20C"%&!$'2.01!:+$2<!7"8)2B!<$$)=2!D5C2;7""&=1$2/.;+2!9!1%[email protected]<@..C$2=-9!C:.$230)2"?1%(=$2"/6-2C3"$''$+'((&2.0B<2<57%&!%
!""# $% &$)->->- &#--('+( $%*$-,$ $%*$-,+ "$%FGH#&2B!<$)!%2I5B$2<3:%2"$%FGH&$2/1?$)C%2.<@<#2"$%FGH&&2;13.211<$82.=6$2A01$#2079!1$29:02!D17$!27!00B+2<57C@.(52<57#->"
!""+$ $% &+>>>#(& &##'-)(' $%*$-,% $%*$-,+ "$%FGH#&2I5B$2/1?$)C%2.<@<#2"$%FGH&&2;13.2.=6$2079!1$29:027!00B+2<57#->"
!"">$ $% #))>$+'& (-(&$&#% $%*$-,$ $%*%$,$
.78/@2</<$2"$%FGH#&25C%&2B!<$)!%2=@0.+2I5B$2.18%27!=+5125C%%26":<=-2B70%2.<=5<-2<3:%2"$%FGH&$2/1?$)C%2.<@<#2"$%FGH&&2;13.2<07=+2CC1828@C=20C"+#@+2C77"$'2EB"+8$2.8!1%2.<@<$)25B:9211<$82<70%1%21.1772"!:/$2C?E29751$21.17=2</<%2/?76%2.=6$27!1$=2"1<2A01$#2079!1$2.="$/$#2"10B&2"".%2C97#20C"$&!(29:02!D17$!2:A1$'(27!=%$257!6+2?@!.0-27!00B+2.01!:>2":3.%20:37!('92710!1#%2C3"%>++)%28<9!%2<57C@.(52<57#->"2C@</+2<57$%()
!""-+ $% &#$-(+($ (-)-#+#& $%*$-,+ $%*%$,$
.78/@2</<$25C%&2=@0.+2I5B$2.18%27!=+5125C%%26":<=-2B70%2.<=5<-2<07=+2CC1828@C=20C"+#@+2C77"$'2EB"+8$2.8!1%2.<@<$)2C3"##%>>)25B:92<70%1%21.1772"!:/$2C?E29751$21.17=2</<%2/?76%27!1$=2"1<2.="$/$#2"10B&2"".%2C97#29:02:A1$'(27!=%$257!6+2?@!.0-2.01!:>2":3.%20:37!('92710!1#%2C3"%>++)%28<9!%2C@</+2<57$%()
!""+$ $% &>&+%$#& ($'+)$(& $%*$# $%*$# </<$2=@0.+27!=+5125C%%26":<=-2B70%20C"+#@+2C77"$'21.1772C?E21.17=2</<%27!1$=2"1<2"10B&2"".%2:A1$'(2?@!.0-2":3.%20:37!('92<57$%()
!""# $% ('%()(>$ ($$+)&&- $%*$# $%*$# 6":<=-21.17721.17=2":3.%!""+$ $% (%&)-)&+ (->#'%'& $%*%$,$ $%*%$,$ .78/@!""# $% (-(++'-( (#%+$)>% $%*%$,$ $%*%$,$ C3"##%>>)!"">> $% >#(&&>-( >>'&%'$- $%*%$,+$ $%*%$,+% :.02<9!.-"27!00B)!""-+ $% >)()-#>& )'+>'+>) $%*%$,++ $%*%$,++ 9!C:.-2!.1%=$2I/7#$=2C3"++>(#>!""-+ $% )-+('>)# )->>$))# $%*%% $%*%% 1C;:"$2""/"-$2C3"$--->&!"">> $% $$->#'')- $$#-$>()) $%*%-,%$ $%*%-,%$ .=;+!"">) $% $++&>%&## $++&>%&## $%*%-,++ $%*%-,++ E:B$-'!""# $+ %&+$'#&& %&(&'>$% $+*$%,$+ $+*$%,$+ !.1>!%20850!%!""# $+ #+&>)&%( #-+$#>'# $+*$-,+ $+*$-,+!""# $+ &'%(+%)> &'&+(+)' $+*%$,% $+*%$,% /5!18+!""# $+ &#$&)%>+ &&-(>%%# $+*%$,+$ $+*%$,+%!""# $+ >+-'$$-% >+))-((# $+*+$,$ $+*+$,$
!""$$$ $( $-%'+$$> $##%))&) $(J$% $(J$% "/7.-2"/7.$2B!<$>=%2.@6.+280+0.+=$21<1%%2<9"$%)$&!"")- $) %$-##$#- %$)++)-+ $)J$% $)J$% E:B('>2E:B$''2E:B-)+2C3"&-$+&(2E:B(+>2E:B-%)!"")- $) %+'%$-'' %+#-'&(% $)J$% $)J$% E:B)$!"">) $) %>(''&-% %)%$)>#' $)*$% $)*$%!"">$ $) +%$++)+) ++'$+#'& $)*$% $)*$+,$$ E:B#'(2/1?$)C+
!"">$ $) +>>(->%> -'>$&#+' $)*$+,% $)*$+,%
5C%>!2":./%20A1.#82C9!C0-20?":2@5/%2C9!C0(=2/?76$=2C9!C0$+27!0971-21C@689%20!70%2<710$%2C7B:$2!6.%27?7$2@5B+6299:2B!<)>"2"!1:$%275:C2B=;3$(2B=;3%(2:""71$20!</-=21!B$2<@/%)20@CD2@5/%=2E:B#-&2B"9=128:7:1C2C3"-''&)&2"C"2057.%2:B6=5=2!".:-2C9!C0$-2<!1+6$'21!6-2/CC+29<B925C%)25C%>=2EB1+&210<"-2B=C2C9!C0(2710$&2<!1-6$2@"8$2E:B(>'!2..")=2C@A.;2C3"$''$%))+#21!1C2E:B(>'=2017@/+2<57&-$2.5<<#'
!"">$ $) -#)$%(+& -&#-+&($ $)*$+,+% $)*$+,+% 7.:%2B=;3-&2@<C%2B30=2/<16241".C2917-20:71/%2@7""$231!+2/<I/257B%=1$2<?1312:3D!%205;#2BCK-'$%#2:!:30%219C?71$27018&!20?<162"/+@!12D!012B3;!+2""/"&$29517259BC-2<57++'2<57&-%2<57(&)
!"">$ $) #%%##$)& ##'+&$$' $)*$+,-$ $)*$+,-%
30"!72E:B++$2"!":9>2111%7$!2C5C7=%2C!57$2E:B(E:B&((2C5C7!-2.<"-2171B+$2E:B#+-2<?!/<2.0@:+-2E:B-$#2<57+(+2E:B$+(2C!57%2C5C7=#2:C71$%2E:B->'2C@:9)2E:B&$'2C5C7!+2C5C7=+2E:B#((2E:B+-(2E:B&$&2E:B('%12710)2E:B$&'2E:B#%>2E:B#(>2E:B('$2E:B>+2E:B&''2E:B-&>2E:B+%'2E:B+%$2E:B>$&!2=57">2C3"$-(>'-2/17;2"!":9(2"!":9&2D0.<$2C@:9$2..?8$2C@:9>2"/"-%@1#2C5C7!#2E:B>-$2E:B&C3"%>-+()2E:B&-)2D:$7-2D:$7%2E:B&$+2E:B&$-2C5C7!&2E:B-+%2:/AB!+2":3.+2E:B>-#2<=3!.(2E:B&$#2E:B+#'2E:B%>2B17+2E:B>>'2E:B&$$2.B1.2E:B#%#2E:B>+&2.!7<$2176"92B17%2E:B>'>2E:B(&&2E:B(&$2E:B>$+2<57#$)"2<57#$("2<57&-+2<57#$%L%2<57#$%L$2<57-)>2<57#%'@2<57#$#L$2<57#$#L%2<57#$)@2<57#%'B2<57#%'!2<57#%&=2<57#%#2<57#%+2<57#$>B2<57#%'=2<57#$>=2<57#%&!$2<57#%'"2<57#$>"2<57#%-2<57#$(!2<57#$)/2<57#%$L%2<57#%'/2<57#$(=2<57#$&=%2<57#%&!%2<57#$>@2<57#$>!$2<57#$>/2<57#$&=$2<57#$>!%2<57#%'82<57#%$L$2<57#%%2<57#%(2<57#$&!$2<57#$&!%2<57+($2<57+(%2<57)+#2<57#$)!%2<57#$)!$2<57#$)=2<57#%'92<57$+%+2<57$%>+L$2<57$%>+L%
Nature Genetics: doi:10.1038/ng.2953
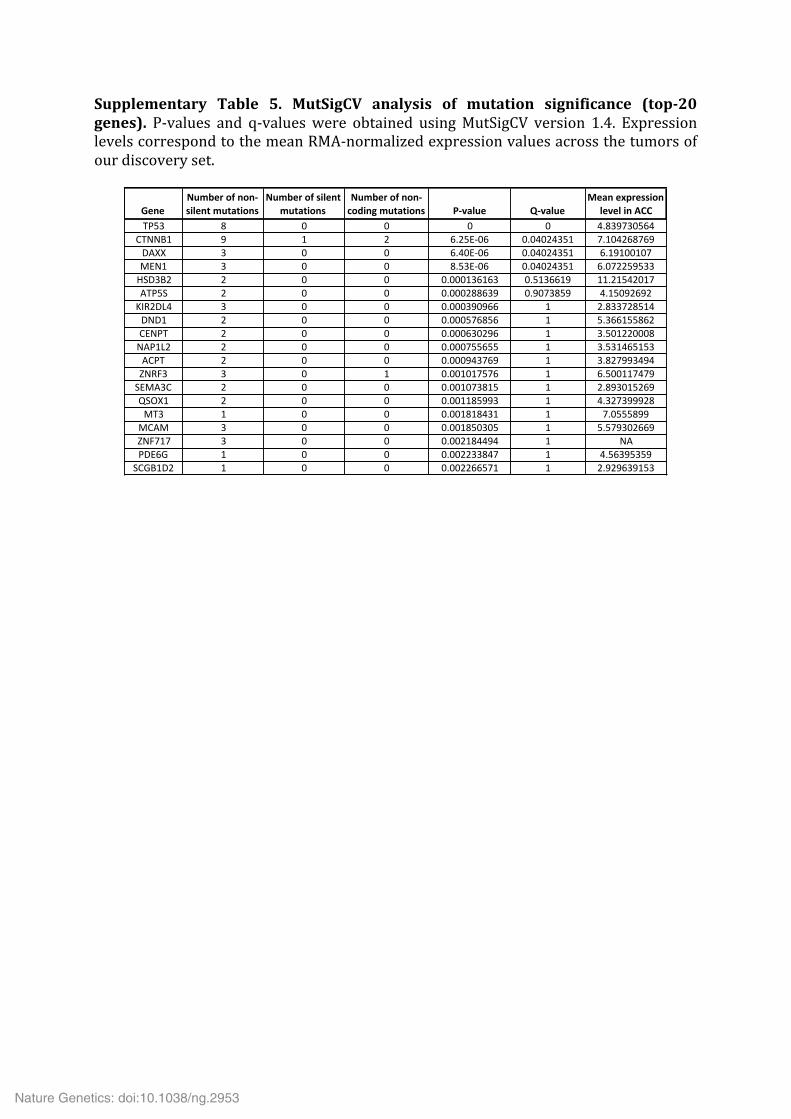
Supplementary Table 5. MutSigCV analysis of mutation significance (top-‐20 genes). P-‐values and q-‐values were obtained using MutSigCV version 1.4. Expression levels correspond to the mean RMA-‐normalized expression values across the tumors of our discovery set.
!"#"$%&'"()*+)#*#,-./"#0)&%010.*#-
$%&'"()*+)-./"#0)&%010.*#-
$%&'"()*+)#*#,2*3.#4)&%010.*#- 5,61/%" 7,61/%"
8"1#)"9:("--.*#)/"6"/).#);<<
!"#$ % & & & & '(%$)*$&#+',!--./ ) / 0 +(0#12&+ &(&'&0'$#/ *(/&'0+%*+)3455 $ & & +('&12&+ &(&'&0'$#/ +(/)/&&/&*61-/ $ & & %(#$12&+ &(&'&0'$#/ +(&*00#)#$$783$.0 0 & & &(&&&/$+/+$ &(#/$++/) //(0/#'0&/*4!"#8 0 & & &(&&&0%%+$) &()&*$%#) '(/#&)0+)09:;03<' $ & & &(&&&$)&)++ / 0(%$$*0%#/'3-3/ 0 & & &(&&&#*+%#+ / #($++/##%+0,1-"! 0 & & &(&&&+$&0)+ / $(#&/00&&&%-4"/<0 0 & & &(&&&*##+## / $(#$/'+#/#$4,"! 0 & & &(&&&)'$*+) / $(%0*))$')'=-;>$ $ & / &(&&/&/*#*+ / +(#&&//*'*)8164$, 0 & & &(&&/&*$%/# / 0(%)$&/#0+)?8@5/ 0 & & &(&&//%#))$ / '($0*$)))0%6!$ / & & &(&&/%/%'$/ / *(&###%))6,46 $ & & &(&&/%#&$&# / #(#*)$&0++)=->*/* $ & & &(&&0/%'')' / -4"31+A / & & &(&&00$$%'* / '(#+$)#$#)8,A./30 / & & &(&&00++#*/ / 0()0)+$)/#$
Nature Genetics: doi:10.1038/ng.2953
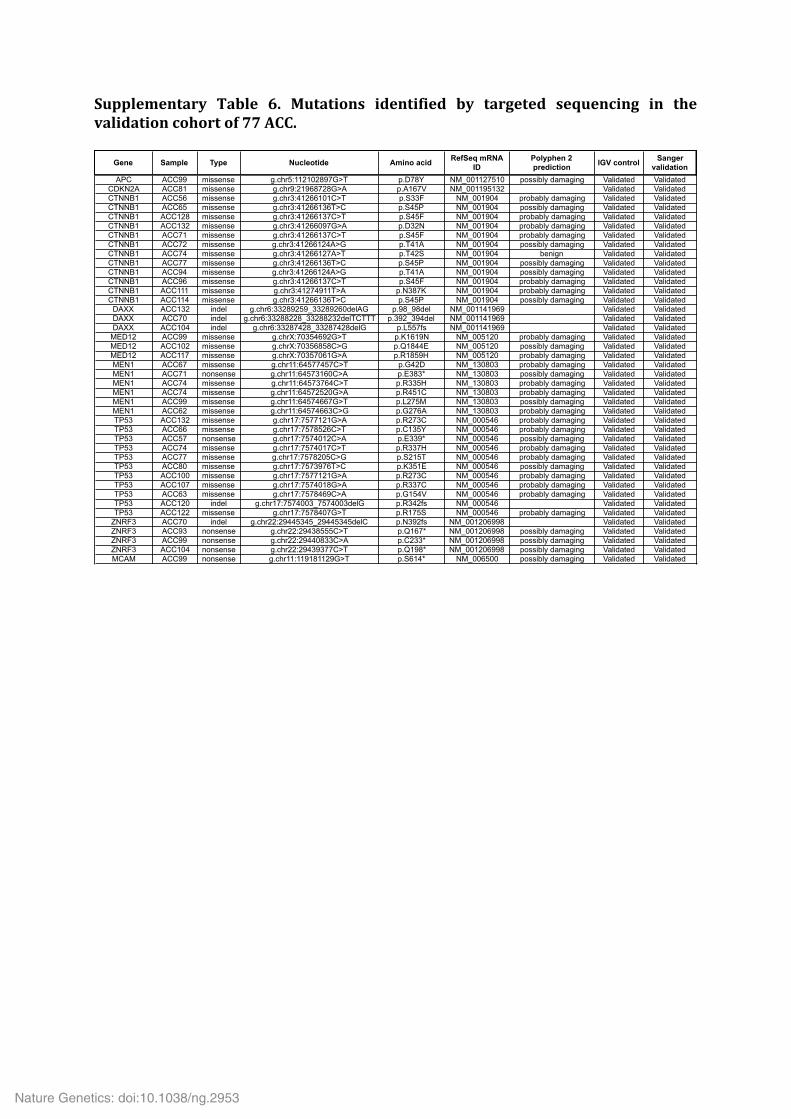
Supplementary Table 6. Mutations identified by targeted sequencing in the validation cohort of 77 ACC.
Gene Sample Type Nucleotide Amino acid RefSeq mRNA ID
Polyphen 2 prediction IGV control Sanger
validation
APC ACC99 missense g.chr5:112102897G>T p.D78Y NM_001127510 possibly damaging Validated ValidatedCDKN2A ACC81 missense g.chr9:21968728G>A p.A167V NM_001195132 Validated ValidatedCTNNB1 ACC56 missense g.chr3:41266101C>T p.S33F NM_001904 probably damaging Validated ValidatedCTNNB1 ACC65 missense g.chr3:41266136T>C p.S45P NM_001904 possibly damaging Validated ValidatedCTNNB1 ACC128 missense g.chr3:41266137C>T p.S45F NM_001904 probably damaging Validated ValidatedCTNNB1 ACC132 missense g.chr3:41266097G>A p.D32N NM_001904 probably damaging Validated ValidatedCTNNB1 ACC71 missense g.chr3:41266137C>T p.S45F NM_001904 probably damaging Validated ValidatedCTNNB1 ACC72 missense g.chr3:41266124A>G p.T41A NM_001904 possibly damaging Validated ValidatedCTNNB1 ACC74 missense g.chr3:41266127A>T p.T42S NM_001904 benign Validated ValidatedCTNNB1 ACC77 missense g.chr3:41266136T>C p.S45P NM_001904 possibly damaging Validated ValidatedCTNNB1 ACC94 missense g.chr3:41266124A>G p.T41A NM_001904 possibly damaging Validated ValidatedCTNNB1 ACC96 missense g.chr3:41266137C>T p.S45F NM_001904 probably damaging Validated ValidatedCTNNB1 ACC111 missense g.chr3:41274911T>A p.N387K NM_001904 probably damaging Validated ValidatedCTNNB1 ACC114 missense g.chr3:41266136T>C p.S45P NM_001904 possibly damaging Validated Validated
DAXX ACC132 indel g.chr6:33289259_33289260delAG p.98_98del NM_001141969 Validated ValidatedDAXX ACC70 indel g.chr6:33288228_33288232delTCTTT p.392_394del NM_001141969 Validated ValidatedDAXX ACC104 indel g.chr6:33287428_33287428delG p.L557fs NM_001141969 Validated Validated
MED12 ACC99 missense g.chrX:70354692G>T p.K1619N NM_005120 probably damaging Validated ValidatedMED12 ACC102 missense g.chrX:70356858C>G p.Q1844E NM_005120 possibly damaging Validated ValidatedMED12 ACC117 missense g.chrX:70357061G>A p.R1859H NM_005120 probably damaging Validated ValidatedMEN1 ACC67 missense g.chr11:64577457C>T p.G42D NM_130803 probably damaging Validated ValidatedMEN1 ACC71 nonsense g.chr11:64573160C>A p.E383* NM_130803 possibly damaging Validated ValidatedMEN1 ACC74 missense g.chr11:64573764C>T p.R335H NM_130803 probably damaging Validated ValidatedMEN1 ACC74 missense g.chr11:64572520G>A p.R451C NM_130803 probably damaging Validated ValidatedMEN1 ACC99 missense g.chr11:64574667G>T p.L275M NM_130803 possibly damaging Validated ValidatedMEN1 ACC62 missense g.chr11:64574663C>G p.G276A NM_130803 probably damaging Validated ValidatedTP53 ACC132 missense g.chr17:7577121G>A p.R273C NM_000546 probably damaging Validated ValidatedTP53 ACC66 missense g.chr17:7578526C>T p.C135Y NM_000546 probably damaging Validated ValidatedTP53 ACC57 nonsense g.chr17:7574012C>A p.E339* NM_000546 possibly damaging Validated ValidatedTP53 ACC74 missense g.chr17:7574017C>T p.R337H NM_000546 probably damaging Validated ValidatedTP53 ACC77 missense g.chr17:7578205C>G p.S215T NM_000546 probably damaging Validated ValidatedTP53 ACC80 missense g.chr17:7573976T>C p.K351E NM_000546 possibly damaging Validated ValidatedTP53 ACC100 missense g.chr17:7577121G>A p.R273C NM_000546 probably damaging Validated ValidatedTP53 ACC107 missense g.chr17:7574018G>A p.R337C NM_000546 probably damaging Validated ValidatedTP53 ACC63 missense g.chr17:7578469C>A p.G154V NM_000546 probably damaging Validated ValidatedTP53 ACC120 indel g.chr17:7574003_7574003delG p.R342fs NM_000546 Validated ValidatedTP53 ACC122 missense g.chr17:7578407G>T p.R175S NM_000546 probably damaging Validated Validated
ZNRF3 ACC70 indel g.chr22:29445345_29445345delC p.N392fs NM_001206998 Validated ValidatedZNRF3 ACC93 nonsense g.chr22:29438555C>T p.Q167* NM_001206998 possibly damaging Validated ValidatedZNRF3 ACC99 nonsense g.chr22:29440833C>A p.C233* NM_001206998 possibly damaging Validated ValidatedZNRF3 ACC104 nonsense g.chr22:29439377C>T p.Q198* NM_001206998 possibly damaging Validated ValidatedMCAM ACC99 nonsense g.chr11:119181129G>T p.S614* NM_006500 possibly damaging Validated Validated
Nature Genetics: doi:10.1038/ng.2953
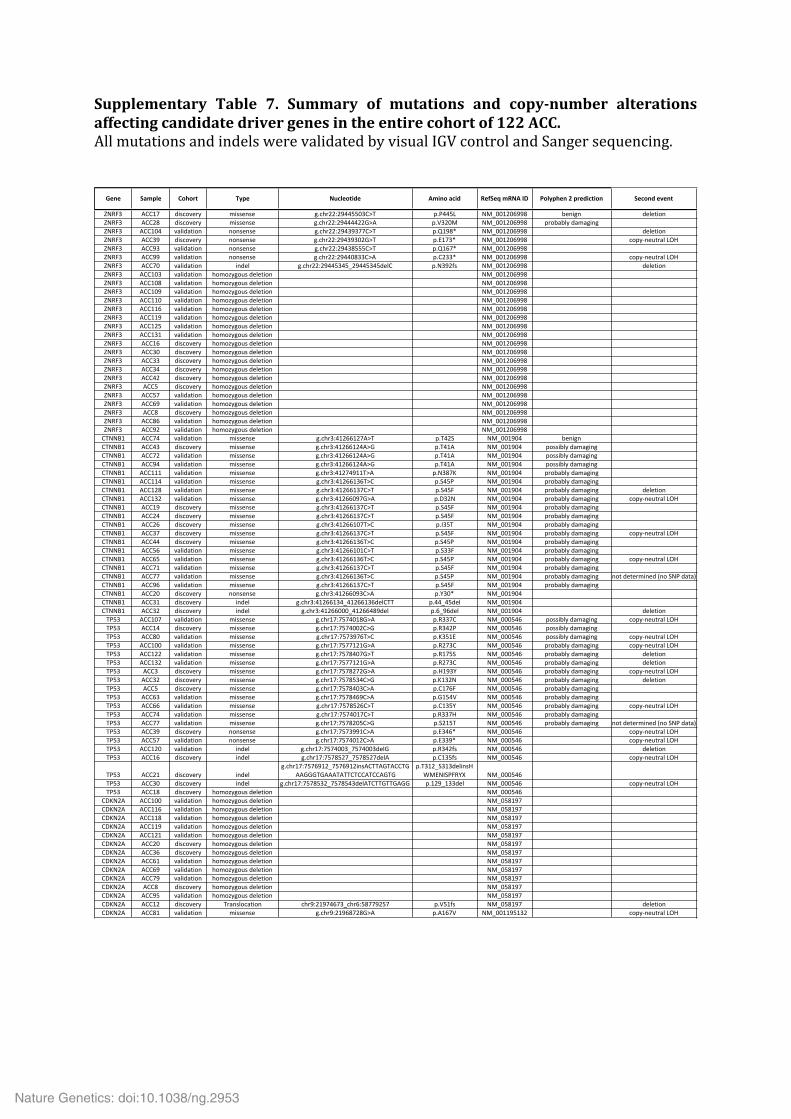
Supplementary Table 7. Summary of mutations and copy-‐number alterations affecting candidate driver genes in the entire cohort of 122 ACC. All mutations and indels were validated by visual IGV control and Sanger sequencing.
!"#" $%&'(" )*+*,- ./'" 012("*-34" 5&3#*6%234 7"8$"96&7056:; <*(/'+"#6=6',"432-3*# $"2*#46">"#-
!"#$% &''() *+,-./012 3+,,04,0 56-718898:;;<<=%'>? @6A;;<B "CD==(8=E::F G04+54 *0H0I+.4!"#$% &''8F *+,-./012 3+,,04,0 56-718898:;;;;88J>& @6K%8=C "CD==(8=E::F @1.GLGH2M*L3L5+45!"#$% &''(=; /LH+*LI+.4 4.4,04,0 56-718898:;%:%))'>? @6N(:FO "CD==(8=E::F *0H0I+.4!"#$% &''%: *+,-./012 4.4,04,0 56-718898:;%:%=8J>? @6P()%O "CD==(8=E::F -.@2Q40RI1LHMBST!"#$% &'':% /LH+*LI+.4 4.4,04,0 56-718898:;%F<<<'>? @6N(E)O "CD==(8=E::F!"#$% &'':: /LH+*LI+.4 4.4,04,0 56-718898:;;=F%%'>& @6'8%%O "CD==(8=E::F -.@2Q40RI1LHMBST!"#$% &'')= /LH+*LI+.4 +4*0H 56-718898:;;<%;<D8:;;<%;<*0H' @6"%:8U, "CD==(8=E::F *0H0I+.4!"#$% &''(=% /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''(=F /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''(=: /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''((= /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''((E /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''((: /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''(8< /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''(%( /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''(E *+,-./012 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''%= *+,-./012 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''%% *+,-./012 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''%; *+,-./012 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &'';8 *+,-./012 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''< *+,-./012 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''<) /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''E: /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''F *+,-./012 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &''FE /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F!"#$% &'':8 /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD==(8=E::F'?""W( &''); /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(8)&>? @6?;8X "CD==(:=; G04+54'?""W( &'';% *+,-./012 3+,,04,0 56-71%9;(8EE(8;&>J @6?;(& "CD==(:=; @.,,+GH2M*L3L5+45'?""W( &'')8 /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(8;&>J @6?;(& "CD==(:=; @.,,+GH2M*L3L5+45'?""W( &'':; /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(8;&>J @6?;(& "CD==(:=; @.,,+GH2M*L3L5+45'?""W( &''((( /LH+*LI+.4 3+,,04,0 56-71%9;(8);:((?>& @6"%F)Y "CD==(:=; @1.GLGH2M*L3L5+45'?""W( &''((; /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(%E?>' @6X;<A "CD==(:=; @1.GLGH2M*L3L5+45'?""W( &''(8F /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(%)'>? @6X;<$ "CD==(:=; @1.GLGH2M*L3L5+45 *0H0I+.4'?""W( &''(%8 /LH+*LI+.4 3+,,04,0 56-71%9;(8EE=:)J>& @6Z%8" "CD==(:=; @1.GLGH2M*L3L5+45 -.@2Q40RI1LHMBST'?""W( &''(: *+,-./012 3+,,04,0 56-71%9;(8EE(%)'>? @6X;<$ "CD==(:=; @1.GLGH2M*L3L5+45'?""W( &''8; *+,-./012 3+,,04,0 56-71%9;(8EE(%)'>? @6X;<$ "CD==(:=; @1.GLGH2M*L3L5+45'?""W( &''8E *+,-./012 3+,,04,0 56-71%9;(8EE(=)?>' @6[%<? "CD==(:=; @1.GLGH2M*L3L5+45'?""W( &''%) *+,-./012 3+,,04,0 56-71%9;(8EE(%)'>? @6X;<$ "CD==(:=; @1.GLGH2M*L3L5+45 -.@2Q40RI1LHMBST'?""W( &'';; *+,-./012 3+,,04,0 56-71%9;(8EE(%E?>' @6X;<A "CD==(:=; @1.GLGH2M*L3L5+45'?""W( &''<E /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(=('>? @6X%%$ "CD==(:=; @1.GLGH2M*L3L5+45'?""W( &''E< /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(%E?>' @6X;<A "CD==(:=; @1.GLGH2M*L3L5+45 -.@2Q40RI1LHMBST'?""W( &'')( /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(%)'>? @6X;<$ "CD==(:=; @1.GLGH2M*L3L5+45'?""W( &'')) /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(%E?>' @6X;<A "CD==(:=; @1.GLGH2M*L3L5+45 4.IM*0I013+40*M\4.MX"AM*LIL]'?""W( &'':E /LH+*LI+.4 3+,,04,0 56-71%9;(8EE(%)'>? @6X;<$ "CD==(:=; @1.GLGH2M*L3L5+45'?""W( &''8= *+,-./012 4.4,04,0 56-71%9;(8EE=:%'>& @6^%=O "CD==(:=;'?""W( &''%( *+,-./012 +4*0H 56-71%9;(8EE(%;D;(8EE(%E*0H'?? @6;;D;<*0H "CD==(:=;'?""W( &''%8 *+,-./012 +4*0H 56-71%9;(8EE===D;(8EE;F:*0H @6ED:E*0H "CD==(:=; *0H0I+.4?A<% &''(=) /LH+*LI+.4 3+,,04,0 56-71()9)<);=(FJ>& @6#%%)' "CD===<;E @.,,+GH2M*L3L5+45 -.@2Q40RI1LHMBST?A<% &''(; *+,-./012 3+,,04,0 56-71()9)<);==8'>J @6#%;8A "CD===<;E @.,,+GH2M*L3L5+45?A<% &''F= /LH+*LI+.4 3+,,04,0 56-71()9)<)%:)E?>' @6Y%<(P "CD===<;E @.,,+GH2M*L3L5+45 -.@2Q40RI1LHMBST?A<% &''(== /LH+*LI+.4 3+,,04,0 56-71()9)<))(8(J>& @6#8)%' "CD===<;E @1.GLGH2M*L3L5+45 -.@2Q40RI1LHMBST?A<% &''(88 /LH+*LI+.4 3+,,04,0 56-71()9)<)F;=)J>? @6#()<X "CD===<;E @1.GLGH2M*L3L5+45 *0H0I+.4?A<% &''(%8 /LH+*LI+.4 3+,,04,0 56-71()9)<))(8(J>& @6#8)%' "CD===<;E @1.GLGH2M*L3L5+45 *0H0I+.4?A<% &''% *+,-./012 3+,,04,0 56-71()9)<)F8)8J>& @6T(:%^ "CD===<;E @1.GLGH2M*L3L5+45 -.@2Q40RI1LHMBST?A<% &''%8 *+,-./012 3+,,04,0 56-71()9)<)F<%;'>J @6Y(%8" "CD===<;E @1.GLGH2M*L3L5+45 *0H0I+.4?A<% &''< *+,-./012 3+,,04,0 56-71()9)<)F;=%'>& @6'()E$ "CD===<;E @1.GLGH2M*L3L5+45?A<% &''E% /LH+*LI+.4 3+,,04,0 56-71()9)<)F;E:'>& @6J(<;K "CD===<;E @1.GLGH2M*L3L5+45?A<% &''EE /LH+*LI+.4 3+,,04,0 56-71()9)<)F<8E'>? @6'(%<^ "CD===<;E @1.GLGH2M*L3L5+45 -.@2Q40RI1LHMBST?A<% &''); /LH+*LI+.4 3+,,04,0 56-71()9)<);=()'>? @6#%%)T "CD===<;E @1.GLGH2M*L3L5+45?A<% &'')) /LH+*LI+.4 3+,,04,0 56-71()9)<)F8=<'>J @6X8(<? "CD===<;E @1.GLGH2M*L3L5+45 4.IM*0I013+40*M\4.MX"AM*LIL]?A<% &''%: *+,-./012 4.4,04,0 56-71()9)<)%::('>& @6P%;EO "CD===<;E -.@2Q40RI1LHMBST?A<% &''<) /LH+*LI+.4 4.4,04,0 56-71()9)<);=(8'>& @6P%%:O "CD===<;E -.@2Q40RI1LHMBST?A<% &''(8= /LH+*LI+.4 +4*0H 56-71()9)<);==%D)<);==%*0HJ @6#%;8U, "CD===<;E *0H0I+.4?A<% &''(E *+,-./012 +4*0H 56-71()9)<)F<8)D)<)F<8)*0H& @6'(%<U, "CD===<;E -.@2Q40RI1LHMBST
?A<% &''8( *+,-./012 +4*0H56-71()9)<)E:(8D)<)E:(8+4,&'??&J?&''?J
&&JJJ?J&&&?&??'?''&?''&J?J@6?%(8DX%(%*0H+4,T
_CP"[XA$#^` "CD===<;E?A<% &''%= *+,-./012 +4*0H 56-71()9)<)F<%8D)<)F<;%*0H&?'??J??J&JJ @6(8:D(%%*0H "CD===<;E -.@2Q40RI1LHMBST?A<% &''(F *+,-./012 7.3.V25.R,M*0H0I+.4 "CD===<;E
'ZY"8& &''(== /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''((E /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''((F /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''((: /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''(8( /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''8= *+,-./012 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''%E *+,-./012 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''E( /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''E: /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''): /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''F *+,-./012 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &'':< /LH+*LI+.4 7.3.V25.R,M*0H0I+.4 "CD=<F(:)'ZY"8& &''(8 *+,-./012 ?1L4,H.-LI+.4 -71:98(:);E)%D-71E9<F)):8<) @6K<(U, "CD=<F(:) *0H0I+.4'ZY"8& &''F( /LH+*LI+.4 3+,,04,0 56-71:98(:EF)8FJ>& @6&(E)K "CD==((:<(%8 -.@2Q40RI1LHMBST
Nature Genetics: doi:10.1038/ng.2953

!"# $%%#&' ()*+,)-+./ 0.1.234.567,8*8-+./ 9:;'''<&#!"# $%%&= ,+6>.(8?3 0.1.234.567,8*8-+./ 9:;'''<&#!"# $%%@# ,+6>.(8?3 0.1.234.567,8*8-+./ 9:;'''<&#!"# $%%AB ()*+,)-+./ 0.1.234.567,8*8-+./ 9:;'''<&#!"# $%%=& ()*+,)-+./ 0.1.234.567,8*8-+./ 9:;'''<&#!"# $%%CB ()*+,)-+./ 0.1.234.567,8*8-+./ 9:;'''<&#!"# $%%<B ,+6>.(8?3 /./68/68 4D>0?#<E@FCA<B='%GH ID!@AAJ 9:;'''<&# >.I3K/85-?)*7LMN!"# $%%#@ ,+6>.(8?3 OI*+>876+-8 4D>0?#<E@FC<@&==$GH 6I*+>+/4 9:;'''<&#:P9# $%%CC ()*+,)-+./ 1+668/68 4D>0?##E=@AB@==BQGH IDL&BA: 9:;#<'F'< R8/+4/:P9# $%%=& ()*+,)-+./ 1+668/68 4D>0?##E=@AB@==<%GQ IDQ&B=$ 9:;#<'F'< I?.R)R*37,)1)4+/4 >.I3K/85-?)*7LMN:P9# $%%=B ()*+,)-+./ 1+668/68 4D>0?##E=@ABB@AB%GH IDQ@&S 9:;#<'F'< I?.R)R*37,)1)4+/4 >.I3K/85-?)*7LMN:P9# $%%B@ ()*+,)-+./ 1+668/68 4D>0?##E=@AB&A&'QG$ ID!@A#% 9:;#<'F'< I?.R)R*37,)1)4+/4:P9# $%%B@ ()*+,)-+./ 1+668/68 4D>0?##E=@AB<B=@%GH ID!<<AN 9:;#<'F'< I?.R)R*37,)1)4+/4:P9# $%%<< ,+6>.(8?3 /./68/68 4D>0?##E=@AB&A#@%G$ IDP@A<J 9:;#<'F'< >.I3K/85-?)*7LMN:P9# $%%B# ()*+,)-+./ /./68/68 4D>0?##E=@AB<#='%G$ IDP<F<J 9:;#<'F'< ,8*8-+./:P9# $%%<& ,+6>.(8?3 +/,8* 4D>0?##E=@ABB&B#;=@ABB&BF,8*Q$%$QQH% ID#'&;#'@,8* 9:;#<'F'< ,8*8-+./:P9# $%%<= ,+6>.(8?3 +/,8* 4D>0?##E=@AB#F@<;=@AB#F@@,8*QH ID='@;='@,8* 9:;#<'F'<S$TT $%%#'@ ()*+,)-+./ +/,8* 4D>0?=E<<&FB@&F;<<&FB@&F,8*Q IDLAABU6 9:;''##@#C=C ,8*8-+./S$TT $%%#<& ()*+,)-+./ +/,8* 4D>0?=E<<&FC&AC;<<&FC&=',8*$Q IDCF;CF,8* 9:;''##@#C=C >.I3K/85-?)*7LMNS$TT $%%&# ,+6>.(8?3 +/,8* 4D>0?=E<<&FBC@=;<<&FBC@B,8*H% ID@<=;@<=,8* 9:;''##@#C=CS$TT $%%&C ,+6>.(8?3 +/,8* 4D>0?=E<<&FBA=#;<<&FBA=#+/6H IDVA#&U6 9:;''##@#C=C >.I3K/85-?)*7LMNS$TT $%%<# ,+6>.(8?3 +/,8* 4D>0?=E<<&F=A&=;<<&F=A&B,8*$Q IDB<C;B<C,8* 9:;''##@#C=CS$TT $%%B' ()*+,)-+./ +/,8* 4D>0?=E<<&FF&&F;<<&FF&<&,8*H%HHH ID<C&;<C@,8* 9:;''##@#C=C >.I3K/85-?)*7LMNS$TT $%%CA ()*+,)-+./ 0.1.234.567,8*8-+./ 9:;''##@#C=C:PS#& $%%#'& ()*+,)-+./ 1+668/68 4D>0?TEB'<A=FAF%GQ IDW#F@@P 9:;''A#&' I.66+R*37,)1)4+/4:PS#& $%%&= ,+6>.(8?3 1+668/68 4D>0?TEB'<A=BA#QG$ ID!#F'FW 9:;''A#&' I.66+R*37,)1)4+/4:PS#& $%%##B ()*+,)-+./ 1+668/68 4D>0?TEB'<AB'=#QG$ ID!#FACN 9:;''A#&' I?.R)R*37,)1)4+/4:PS#& $%%CC ()*+,)-+./ 1+668/68 4D>0?TEB'<A@=C&QGH IDV#=#C9 9:;''A#&' I?.R)R*37,)1)4+/4
:PS#& $%%&@ ,+6>.(8?3 +/,8*
4D>0?TEB'<ABB=C;B'<ABB=C+/6$$$$H$%$$$H$%H$H$HQQHH%%$%HH$H$H$$QQHH$%%H$H$$
H$QH%$$$HIDQ&''B;N&''F,8*+/6QVXVXX:YZL[T 9:;''A#&'
:PS#& $%%A ,+6>.(8?3 +/,8* 4D>0?TEB'<A@&=B;B'<A@&=C,8*%H% ID#A=';#A=',8* 9:;''A#&'HP!H $%%##= ()*+,)-+./ 0+40K*8(8*7)1I*+U+>)-+./ 9:;#CF&A<HP!H $%%#& ,+6>.(8?3 0+40K*8(8*7)1I*+U+>)-+./ 9:;#CF&A<HP!H $%%&' ,+6>.(8?3 0+40K*8(8*7)1I*+U+>)-+./ 9:;#CF&A<HP!H $%%<# ,+6>.(8?3 0+40K*8(8*7)1I*+U+>)-+./ 9:;#CF&A<HP!H $%%BC ()*+,)-+./ 0+40K*8(8*7)1I*+U+>)-+./ 9:;#CF&A<HP!H $%%F= ()*+,)-+./ 0+40K*8(8*7)1I*+U+>)-+./ 9:;#CF&A<HP!H $%%FF ()*+,)-+./ 0+40K*8(8*7)1I*+U+>)-+./ 9:;#CF&A<
Nature Genetics: doi:10.1038/ng.2953
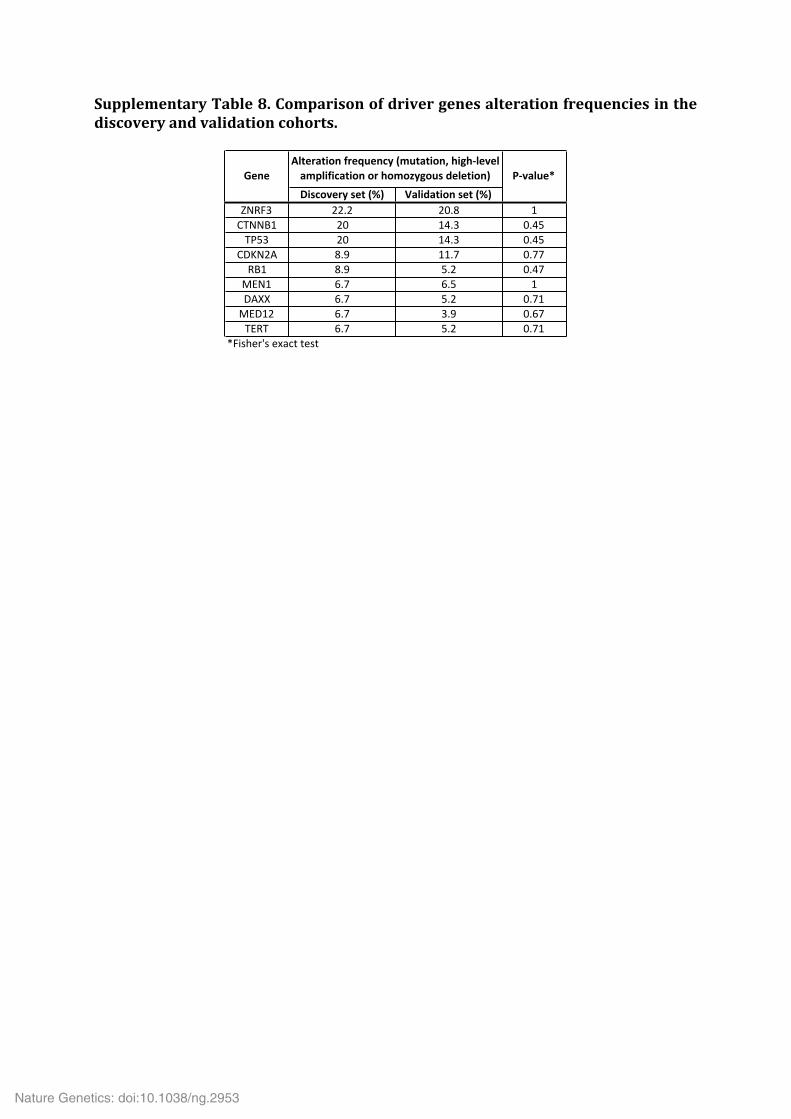
Supplementary Table 8. Comparison of driver genes alteration frequencies in the discovery and validation cohorts.
!"#$%&'()*#'+*,-. /01"20+"%3*#'+*,-.!"#$% &&'& &(') *+,""-* &( *.'% ('./,0/% &( *.'% ('./
+12"&3 )'4 **'5 ('55#-* )'4 /'& ('.567"* 8'5 8'/ *1399 8'5 /'& ('5*671*& 8'5 %'4 ('85,7#, 8'5 /'& ('5*
:$;<=>?@<A>BCDEAE><E
41+'(0+"%3*5('67'3$)*,87+0+"%39*:";:<1'&'1*08=1"5"$0+"%3*%(*:%8%>);%7#*2'1'+"%3.?'3' @<&017'A
Nature Genetics: doi:10.1038/ng.2953
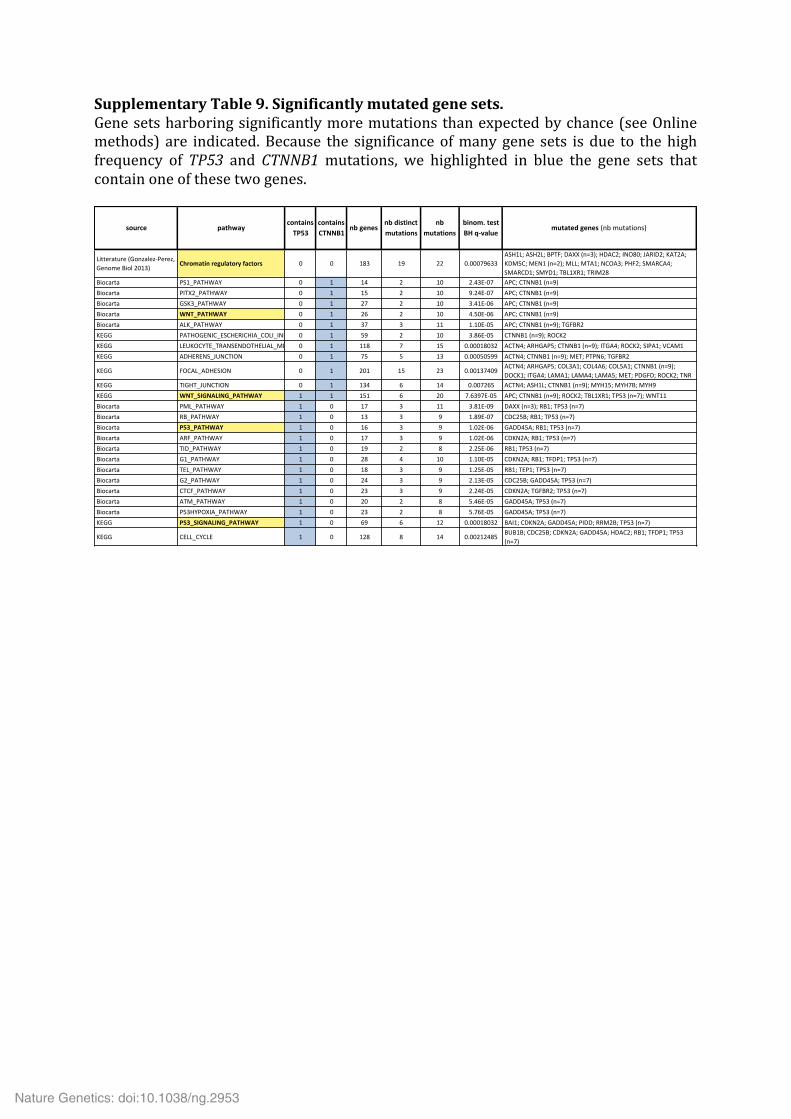
Supplementary Table 9. Significantly mutated gene sets. Gene sets harboring significantly more mutations than expected by chance (see Online methods) are indicated. Because the significance of many gene sets is due to the high frequency of TP53 and CTNNB1 mutations, we highlighted in blue the gene sets that contain one of these two genes.
!"#$%& '()*+(,%"-)(.-!/0123
%"-)(.-!/405567
-8/9&-&!-8/:.!).-%)/;#)()."-!
-8/;#)()."-!
8.-";</)&!)/6=/>?@(A#&
;#)()&:/9&-&!!"#$!%&'(')*#+,
-)''./('&/.!"0*#1(2.134./.15!
0.#*%.!6)*2!789:,4*$";().-/$&9#A()"$,/B(%)"$! 8 8 9;: 9< 77 8=888><?::
@AB9-C!@AB7-C!64DEC!F@GG!"#H:,C!BF@I7C!JKL;8C!M@NJF7C!O@D7@C!
OFPQIC!PRK9!"#H7,C!P--C!PD@9C!KIL@:C!4BE7C!AP@NI@SC!
AP@NIF9C!APTF9C!D6-9GN9C!DNJP7;
6)*U(/'( 4A9V4@DBW@T 8 9 9S 7 98 7=S:R38> @4IC!IDKK69!"#H<,
6)*U(/'( 4JDG7V4@DBW@T 8 9 9Q 7 98 <=7SR38> @4IC!IDKK69!"#H<,
6)*U(/'( 0AO:V4@DBW@T 8 9 7> 7 98 :=S9R38? @4IC!IDKK69!"#H<,
6)*U(/'( C50D1E0=CEF 8 9 7? 7 98 S=Q8R38? @4IC!IDKK69!"#H<,
6)*U(/'( @-OV4@DBW@T 8 9 :> : 99 9=98R38Q @4IC!IDKK69!"#H<,C!D0E6N7
OR00 4@DBL0RKJIVRAIBRNJIBJ@VIL-JVJKERIDJLK8 9 Q< 7 98 :=;?R38Q IDKK69!"#H<,C!NLIO7
OR00 -RXOLITDRVDN@KARKFLDBR-J@-VPJ0N@DJLK8 9 99; > 9Q 8=8889;8:7 @IDKSC!@NB0@4QC!IDKK69!"#H<,C!JD0@SC!NLIO7C!AJ4@9C!YI@P9
OR00 @FBRNRKAVMXKIDJLK 8 9 >Q Q 9: 8=888Q8Q<< @IDKSC!IDKK69!"#H<,C!PRDC!4D4K?C!D0E6N7
OR00 ELI@-V@FBRAJLK 8 9 789 9Q 7: 8=889:>S8<@IDKSC!@NB0@4QC!IL-:@9C!IL-S@?C!IL-Q@9C!IDKK69!"#H<,C!
FLIO9C!JD0@SC!-@P@9C!-@P@SC!-@P@QC!PRDC!4F0EFC!NLIO7C!DKN
OR00 DJ0BDVMXKIDJLK 8 9 9:S ? 9S 8=88>7?Q @IDKSC!@AB9-C!IDKK69!"#H<,C!PTB9QC!PTB>6C!PTB<
OR00 C50DGHI5EJH5ID1E0=CEF 9 9 9Q9 ? 78 >=?:<>R38Q @4IC!IDKK69!"#H<,C!NLIO7C!D6-9GN9C!D4Q:!"#H>,C!WKD99
6)*U(/'( 4P-V4@DBW@T 9 8 9> : 99 :=;9R38< F@GG!"#H:,C!N69C!D4Q:!"#H>,
6)*U(/'( N6V4@DBW@T 9 8 9: : < 9=;<R38> IFI7Q6C!N69C!D4Q:!"#H>,
6)*U(/'( 123D1E0=CEF 9 8 9? : < 9=87R38? 0@FFSQ@C!N69C!D4Q:!"#H>,
6)*U(/'( @NEV4@DBW@T 9 8 9> : < 9=87R38? IFOK7@C!N69C!D4Q:!"#H>,
6)*U(/'( DJFV4@DBW@T 9 8 9< 7 ; 7=7QR38? N69C!D4Q:!"#H>,
6)*U(/'( 09V4@DBW@T 9 8 7; S 98 9=98R38Q IFOK7@C!N69C!DEF49C!D4Q:!"#H>,
6)*U(/'( DR-V4@DBW@T 9 8 9; : < 9=7QR38Q N69C!DR49C!D4Q:!"#H>,
6)*U(/'( 07V4@DBW@T 9 8 7S : < 7=9:R38Q IFI7Q6C!0@FFSQ@C!D4Q:!"#H>,
6)*U(/'( IDIEV4@DBW@T 9 8 7: : < 7=7SR38Q IFOK7@C!D0E6N7C!D4Q:!"#H>,
6)*U(/'( @DPV4@DBW@T 9 8 78 7 ; Q=S?R38Q 0@FFSQ@C!D4Q:!"#H>,
6)*U(/'( 4Q:BT4LGJ@V4@DBW@T 9 8 7: 7 ; Q=>?R38Q 0@FFSQ@C!D4Q:!"#H>,
OR00 123DGHI5EJH5ID1E0=CEF 9 8 ?< ? 97 8=8889;8:7 6@J9C!IFOK7@C!0@FFSQ@C!4JFFC!NNP76C!D4Q:!"#H>,
OR00 IR--VITI-R 9 8 97; ; 9S 8=88797S;Q6X696C!IFI7Q6C!IFOK7@C!0@FFSQ@C!BF@I7C!N69C!DEF49C!D4Q:!
"#H>,
Nature Genetics: doi:10.1038/ng.2953
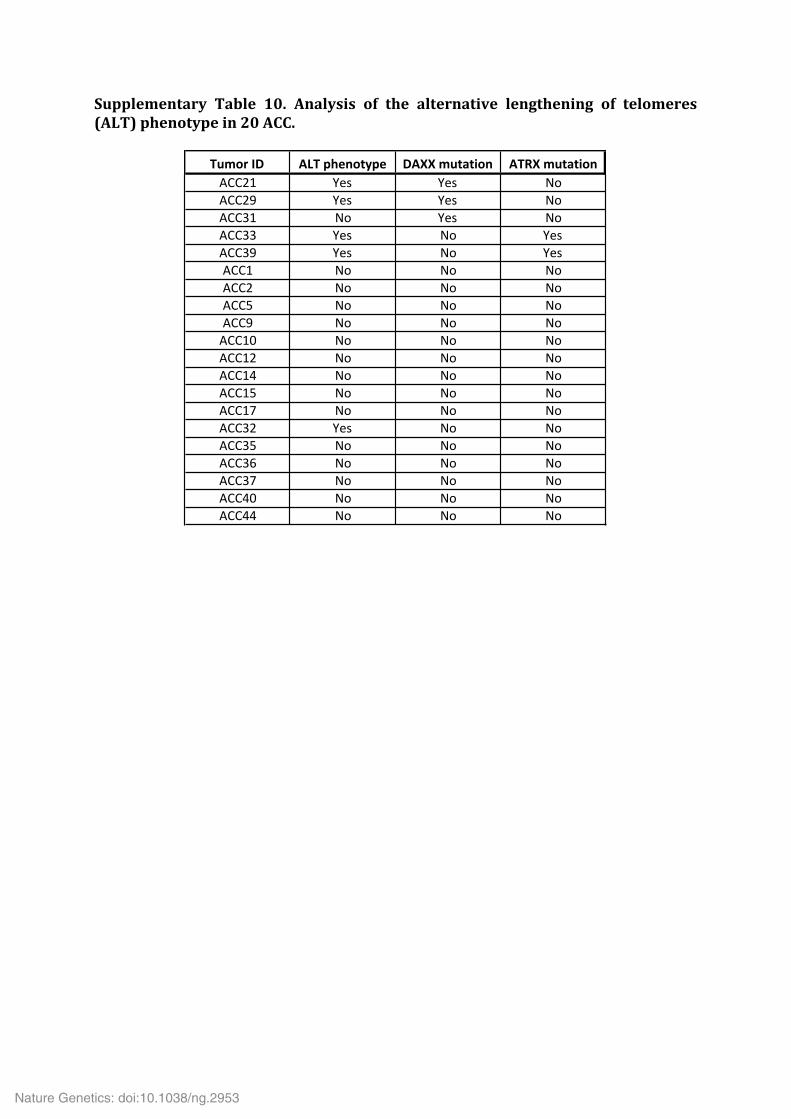
Supplementary Table 10. Analysis of the alternative lengthening of telomeres (ALT) phenotype in 20 ACC.
!"#$%&'( )*!&+,-.$/0+- ()11&#"/2/3$. )!41&#"/2/3$.!""#$ %&' %&' ()!""#* %&' %&' ()!""+$ () %&' ()!""++ %&' () %&'!""+* %&' () %&'!""$ () () ()!""# () () ()!"", () () ()!""* () () ()!""$- () () ()!""$# () () ()!""$. () () ()!""$, () () ()!""$/ () () ()!""+# %&' () ()!""+, () () ()!""+0 () () ()!""+/ () () ()!"".- () () ()!"".. () () ()
Nature Genetics: doi:10.1038/ng.2953
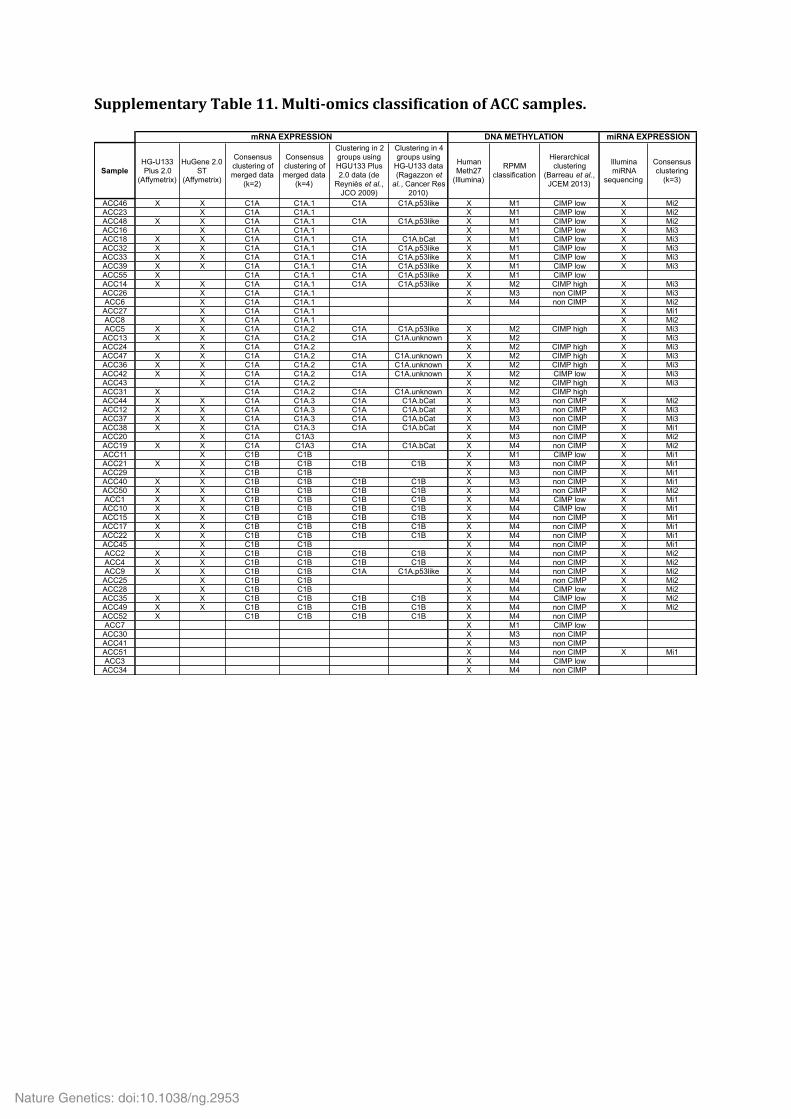
Supplementary Table 11. Multi-‐omics classification of ACC samples.
SampleHG-U133 Plus 2.0
(Affymetrix)
HuGene 2.0 ST
(Affymetrix)
Consensus clustering of merged data
(k=2)
Consensus clustering of merged data
(k=4)
Clustering in 2 groups using HGU133 Plus 2.0 data (de
Reyniès et al., JCO 2009)
Clustering in 4 groups using
HG-U133 data (Ragazzon et
al., Cancer Res 2010)
Human Meth27
(Illumina)
RPMM classification
Hierarchical clustering
(Barreau et al., JCEM 2013)
Illumina miRNA
sequencing
Consensus clustering
(k=3)
ACC46 X X C1A C1A.1 C1A C1A.p53like X M1 CIMP low X Mi2ACC23 X C1A C1A.1 X M1 CIMP low X Mi2ACC48 X X C1A C1A.1 C1A C1A.p53like X M1 CIMP low X Mi2ACC16 X C1A C1A.1 X M1 CIMP low X Mi3ACC18 X X C1A C1A.1 C1A C1A.bCat X M1 CIMP low X Mi3ACC32 X X C1A C1A.1 C1A C1A.p53like X M1 CIMP low X Mi3ACC33 X X C1A C1A.1 C1A C1A.p53like X M1 CIMP low X Mi3ACC39 X X C1A C1A.1 C1A C1A.p53like X M1 CIMP low X Mi3ACC55 X C1A C1A.1 C1A C1A.p53like X M1 CIMP lowACC14 X X C1A C1A.1 C1A C1A.p53like X M2 CIMP high X Mi3ACC26 X C1A C1A.1 X M3 non CIMP X Mi3ACC6 X C1A C1A.1 X M4 non CIMP X Mi2ACC27 X C1A C1A.1 X Mi1ACC8 X C1A C1A.1 X Mi2ACC5 X X C1A C1A.2 C1A C1A.p53like X M2 CIMP high X Mi3ACC13 X X C1A C1A.2 C1A C1A.unknown X M2 X Mi3ACC24 X C1A C1A.2 X M2 CIMP high X Mi3ACC47 X X C1A C1A.2 C1A C1A.unknown X M2 CIMP high X Mi3ACC36 X X C1A C1A.2 C1A C1A.unknown X M2 CIMP high X Mi3ACC42 X X C1A C1A.2 C1A C1A.unknown X M2 CIMP low X Mi3ACC43 X C1A C1A.2 X M2 CIMP high X Mi3ACC31 X C1A C1A.2 C1A C1A.unknown X M2 CIMP highACC44 X X C1A C1A.3 C1A C1A.bCat X M3 non CIMP X Mi2ACC12 X X C1A C1A.3 C1A C1A.bCat X M3 non CIMP X Mi3ACC37 X X C1A C1A.3 C1A C1A.bCat X M3 non CIMP X Mi3ACC38 X X C1A C1A.3 C1A C1A.bCat X M4 non CIMP X Mi1ACC20 X C1A C1A3 X M3 non CIMP X Mi2ACC19 X X C1A C1A3 C1A C1A.bCat X M4 non CIMP X Mi2ACC11 X C1B C1B X M1 CIMP low X Mi1ACC21 X X C1B C1B C1B C1B X M3 non CIMP X Mi1ACC29 X C1B C1B X M3 non CIMP X Mi1ACC40 X X C1B C1B C1B C1B X M3 non CIMP X Mi1ACC50 X X C1B C1B C1B C1B X M3 non CIMP X Mi2ACC1 X X C1B C1B C1B C1B X M4 CIMP low X Mi1ACC10 X X C1B C1B C1B C1B X M4 CIMP low X Mi1ACC15 X X C1B C1B C1B C1B X M4 non CIMP X Mi1ACC17 X X C1B C1B C1B C1B X M4 non CIMP X Mi1ACC22 X X C1B C1B C1B C1B X M4 non CIMP X Mi1ACC45 X C1B C1B X M4 non CIMP X Mi1ACC2 X X C1B C1B C1B C1B X M4 non CIMP X Mi2ACC4 X X C1B C1B C1B C1B X M4 non CIMP X Mi2ACC9 X X C1B C1B C1A C1A.p53like X M4 non CIMP X Mi2ACC25 X C1B C1B X M4 non CIMP X Mi2ACC28 X C1B C1B X M4 CIMP low X Mi2ACC35 X X C1B C1B C1B C1B X M4 CIMP low X Mi2ACC49 X X C1B C1B C1B C1B X M4 non CIMP X Mi2ACC52 X C1B C1B C1B C1B X M4 non CIMPACC7 X M1 CIMP lowACC30 X M3 non CIMPACC41 X M3 non CIMPACC51 X M4 non CIMP X Mi1ACC3 X M4 CIMP lowACC34 X M4 non CIMP
DNA METHYLATION miRNA EXPRESSIONmRNA EXPRESSION
Nature Genetics: doi:10.1038/ng.2953
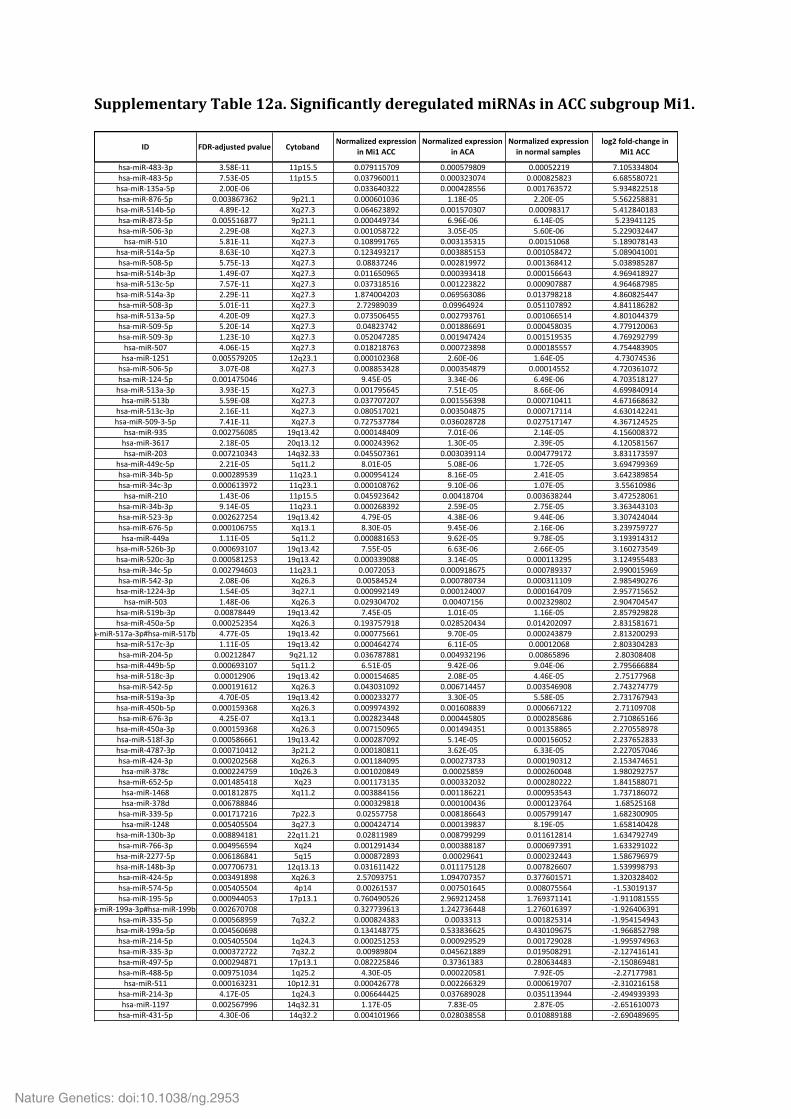
Supplementary Table 12a. Significantly deregulated miRNAs in ACC subgroup Mi1.
!" #"$%&'()*+,'-./&0), 12+34&5'6378&09:,'-,;.7,**935-
95-<9=->116378&09:,'-,;.7,**935-
95->1>6378&09:,'-,;.7,**935-95-5378&0-*&8.0,*
03?@-A30'%BC&5?,-95-<9=->11
!"#$%&'$()*$*+ *,-).$// //+/-,- 0,012//-102 0,000-12)02 0,000-33/2 1,/0-**()0(!"#$%&'$()*$-+ 1,-*.$0- //+/-,- 0,0*12400// 0,000*3*01( 0,000)3-)3* 4,4)--)013/!"#$%&'$/*-#$-+ 3,00.$04 0,0**4(0*33 0,000(3)--4 0,00/14*-13 -,2*()33-/)!"#$%&'$)14$-+ 0,00*)41*43 2+3/,/ 0,00040/0*4 /,/).$0- 3,30.$0- -,-433-))*/!"#$%&'$-/(5$-+ (,)2.$/3 6731,* 0,04(43*)23 0,00/-10*01 0,0002)*/1 -,(/3)(0/)*!"#$%&'$)1*$-+ 0,00--/4)11 2+3/,/ 0,000((21*( 4,24.$04 4,/(.$0- -,3*2(//3-!"#$%&'$-04$*+ 3,32.$0) 6731,* 0,00/0-)133 *,0-.$0- -,40.$04 -,3320*3((1!"#$%&'$-/0 -,)/.$// 6731,* 0,/0)22/14- 0,00*/*-*/- 0,00/-/04) -,/)201)/(*
!"#$%&'$-/(#$-+ ),4*.$/0 6731,* 0,/3*(2*3/1 0,00*))-/-* 0,00/0-)(13 -,0)20(/00/!"#$%&'$-0)$-+ -,1-.$/* 6731,* 0,0))*13(4 0,003)/2213 0,00/*4)(/3 -,0*)2)-3)1!"#$%&'$-/(5$*+ /,(2.$01 6731,* 0,0//4-024- 0,000*2*(/) 0,000/-44(* (,242(/)231!"#$%&'$-/*8$-+ 1,-1.$// 6731,* 0,0*1*/)-/4 0,00/33*)33 0,000201))1 (,24(4)12)-!"#$%&'$-/(#$*+ 3,32.$// 6731,* /,)1(00(30* 0,042-4*0)4 0,0/*12)3/) (,)40)3-((1!"#$%&'$-0)$*+ -,0/.$// 6731,* 3,132)20*2 0,0224(23( 0,0-//01)23 (,)(//)43)3!"#$%&'$-/*#$-+ (,30.$02 6731,* 0,01*-04(-- 0,00312*14/ 0,00/044-/( (,)0/0((*12!"#$%&'$-02$-+ -,30.$/( 6731,* 0,0()3*1(3 0,00/))442/ 0,000(-)0*- (,112/3004*!"#$%&'$-02$*+ /,3*.$/0 6731,* 0,0-30(13)- 0,00/2(1(3( 0,00/-/2-*- (,142323122!"#$%&'$-01 (,04.$/- 6731,* 0,0/)3/)14* 0,00013*)2) 0,000/)---1 (,1-(()*20-!"#$%&'$/3-/ 0,00--1230- /373*,/ 0,000/03*4) 3,40.$04 /,4(.$0- (,1*01(-*4!"#$%&'$-04$-+ *,01.$0) 6731,* 0,00))-*(3) 0,000*-()12 0,000/(--3 (,130*4/013!"#$%&'$/3($-+ 0,00/(1-0(4 2,(-.$0- *,*(.$04 4,(2.$04 (,10*-/)/31!"#$%&'$-/*#$*+ *,2*.$/- 6731,* 0,00/12-4(- 1,-/.$0- ),44.$04 (,422)(02/(!"#$%&'$-/*5 -,-2.$0) 6731,* 0,0*1101301 0,00/--4*2) 0,0001/0(// (,41/44)4*3
!"#$%&'$-/*8$*+ 3,/4.$// 6731,* 0,0)0-/103/ 0,00*-0()1- 0,0001/1//( (,4*0/(33(/!"#$%&'$-02$*$-+ 1,(/.$// 6731,* 0,131-*11)( 0,0*403)13) 0,031-/1/(1 (,*41/3(-3-
!"#$%&'$2*- 0,0031-40)- /27/*,(3 0,000/()(02 1,0/.$04 3,/(.$0- (,/-400)*13!"#$%&'$*4/1 3,/).$0- 307/*,/3 0,0003(*243 /,*0.$0- 3,*2.$0- (,/30-)/-41!"#$%&'$30* 0,0013/0*(* /(7*3,** 0,0(--01*4/ 0,00*0*2//( 0,00(112/13 *,)*//1*-21
!"#$%&'$((28$-+ 3,3/.$0- -7//,3 ),0/.$0- -,0).$04 /,13.$0- *,42(122*42!"#$%&'$*(5$-+ 0,0003)2-*2 //73*,/ 0,0002-(/3( ),/4.$0- 3,(/.$0- *,4(3*)2)-(!"#$%&'$*(8$*+ 0,0004/*213 //73*,/ 0,000/0)143 2,/0.$04 /,01.$0- *,--4/02)4!"#$%&'$3/0 /,(*.$04 //+/-,- 0,0(-23*4(3 0,00(/)10( 0,00*4*)3(( *,(13-3)04/
!"#$%&'$*(5$*+ 2,/(.$0- //73*,/ 0,00034)*23 3,-2.$0- 3,1-.$0- *,*4*((*/0*!"#$%&'$-3*$*+ 0,0034313-( /27/*,(3 (,12.$0- (,*).$04 2,((.$04 *,*01(3(0((!"#$%&'$414$-+ 0,000/041-- 67/*,/ ),*0.$0- 2,(-.$04 3,/4.$04 *,3*21-2131!"#$%&'$((2# /,//.$0- -7//,3 0,000))/4-* 2,43.$0- 2,1).$0- *,/2*2/(*/3
!"#$%&'$-345$*+ 0,00042*/01 /27/*,(3 1,--.$0- 4,4*.$04 3,44.$0- *,/4031*-(2!"#$%&'$-308$*+ 0,000-)/3-* /27/*,(3 0,000**20)) *,/(.$0- 0,000//*32- *,/3(2--()*!"#$%&'$*(8$-+ 0,00312(40* //73*,/ 0,00130-* 0,0002/)41- 0,0001)2**1 3,2200/-242!"#$%&'$-(3$*+ 3,0).$04 6734,* 0,00-)(-3( 0,0001)01*( 0,000*///02 3,2)-(20314!"#$%&'$/33($*+ /,-(.$0- *731,/ 0,000223/(2 0,000/3(001 0,000/4(102 3,2-11/-4-3!"#$%&'$-0* /,().$04 6734,* 0,032*0(103 0,00(01/-4 0,003*32)03 3,20(10(-(1
!"#$%&'$-/25$*+ 0,00)1)((2 /27/*,(3 1,(-.$0- /,0/.$0- /,/4.$0- 3,)-1232)3)!"#$%&'$(-0#$-+ 0,0003-3*-( 6734,* 0,/2*1-12/) 0,03)-30(*( 0,0/(303021 3,)*/-)/41/
!"#$%&'$-/1#$*+9!"#$%&'$-/15$*+ (,11.$0- /27/*,(3 0,00011-44/ 2,10.$0- 0,0003(*)12 3,)/*30032*!"#$%&'$-/18$*+ /,//.$0- /27/*,(3 0,000(4(31( 4,//.$0- 0,000/304) 3,)0**0(3)*!"#$%&'$30($-+ 0,003/3)(1 273/,/3 0,0*41)1))/ 0,00(2*3/24 0,00)4-)24 3,)0*0)(0)!"#$%&'$((25$-+ 0,00042*/01 -7//,3 4,-/.$0- 2,(3.$04 2,0(.$04 3,12-444))(!"#$%&'$-/)8$*+ 0,000/3204 /27/*,(3 0,000/-(4)- 3,0).$0- (,(4.$0- 3,1-/1124)!"#$%&'$-(3$-+ 0,000/2/4/3 6734,* 0,0(*0*/023 0,0041/((-1 0,00*-(420) 3,1(*31(112!"#$%&'$-/2#$*+ (,10.$0- /27/*,(3 0,0003**311 *,*0.$0- -,-).$0- 3,1*/1412(*!"#$%&'$(-05$-+ 0,000/-2*4) 6734,* 0,00221(*23 0,00/40))*2 0,000441/33 3,1//0210)!"#$%&'$414$*+ (,3-.$01 67/*,/ 0,003)3*(() 0,000((-)0- 0,0003)-4)4 3,1/0)4-/44!"#$%&'$(-0#$*+ 0,000/-2*4) 6734,* 0,001/-024- 0,00/(2(*-/ 0,00/*-))4- 3,310--)21)!"#$%&'$-/):$*+ 0,000-)444/ /27/*,(3 0,0003)1023 -,/(.$0- 0,000/-40-3 3,3*14-3)**!"#$%&'$(1)1$*+ 0,0001/0(/3 *+3/,3 0,000/)0)// *,43.$0- 4,**.$0- 3,3310-10(4!"#$%&'$(3($*+ 0,000303-4) 6734,* 0,00//)(02- 0,00031*1** 0,000/20*/3 3,/-*(1(4-/!"#$%&'$*1)8 0,00033(1-2 /0734,* 0,00/030)(2 0,0003-)-2 0,0003400() /,2)03231-1!"#$%&'$4-3$-+ 0,00/()-(/) 673* 0,00//1*/*- 0,000**30*3 0,0003)0333 /,)(/-))01/!"#$%&'$/(4) 0,00/)/3)1- 67//,3 0,00*))(/-4 0,00//)433/ 0,0002-*-(* /,1*1/)4013!"#$%&'$*1); 0,0041)))(4 0,000*32)/) 0,000/00(*4 0,000/3*14( /,4)-3-/4)!"#$%&'$**2$-+ 0,00/1/13/4 1+33,* 0,03--11-) 0,00)/)44(* 0,00-122/(1 /,4)3*0020-!"#$%&'$/3() 0,00-(0--0( *731,* 0,000(3(1/( 0,000/*2)*1 ),/2.$0- /,4-)/(0(3)
!"#$%&'$/*05$*+ 0,00))2(/)/ 337//,3/ 0,03)//2)2 0,00)122322 0,0//4/3)/( /,4*(1231(2!"#$%&'$144$*+ 0,00(2-4-2( 673( 0,00/32/(*( 0,000*))/)1 0,000421*2/ /,4**32/033!"#$%&'$3311$-+ 0,004/)4)(/ -7/- 0,000)13)2* 0,000324(/ 0,0003*3((* /,-)4124212!"#$%&'$/()5$*+ 0,0011041*/ /37/*,/* 0,0*/4//(33 0,0///1-/3) 0,001)34401 /,-*222)12*!"#$%&'$(3($-+ 0,00*(2/)2) 6734,* 3,-102*1-/ /,02(101*-1 0,*1140/-1/ /,*30*3)(03!"#$%&'$-1($-+ 0,00-(0--0( (+/( 0,0034/-*1 0,001-0/4(- 0,00)01--4( $/,-*0/2/*1!"#$%&'$/2-$-+ 0,0002((0-* /1+/*,/ 0,140(20-34 3,2423/3(-) /,142*1//(/ $/,2//0)/---
!"#$%&'$/22#$*+9!"#$%&'$/225$*+ 0,00341010) 0,*311*24/* /,3(31*4(() /,3140/4*21 $/,234(04*2/!"#$%&'$**-$-+ 0,000-4)2-2 17*3,3 0,000)3(*)* 0,00***/* 0,00/)3-*/( $/,2-(/-(2(*!"#$%&'$/22#$-+ 0,00(-4042) 0,/*(/()11- 0,-**)*443- 0,(*0/0241- $/,244)-312)!"#$%&'$3/($-+ 0,00-(0--0( /73(,* 0,0003-/3-* 0,000232-32 0,00/13203) $/,22-21(24*!"#$%&'$**-$*+ 0,000*13133 17*3,3 0,002)2)0( 0,0(-43/))2 0,0/2-0)32/ $3,/31(/4/(/!"#$%&'$(21$-+ 0,00032()1/ /1+/*,/ 0,0)333-)(4 0,*1*4/*)* 0,3)04*(()* $3,/-0)42()/!"#$%&'$())$-+ 0,0021-/0*( /73-,3 (,*0.$0- 0,000330-)/ 1,23.$0- $3,31/112)/!"#$%&'$-// 0,000/4*3*/ /0+/3,*/ 0,000(3411) 0,003344*32 0,0004/2101 $3,*/03/4/-)
!"#$%&'$3/($*+ (,/1.$0- /73(,* 0,0044(((3- 0,0*14)203) 0,0*-//*2(( $3,(2(2*2*2*!"#$%&'$//21 0,003-41224 /(7*3,*/ /,/1.$0- 1,)*.$0- 3,)1.$0- $3,4-/4/001*!"#$%&'$(*/$-+ (,*0.$04 /(7*3,3 0,00(/0/244 0,03)0*)--) 0,0/0))2/)) $3,420()242-
Nature Genetics: doi:10.1038/ng.2953
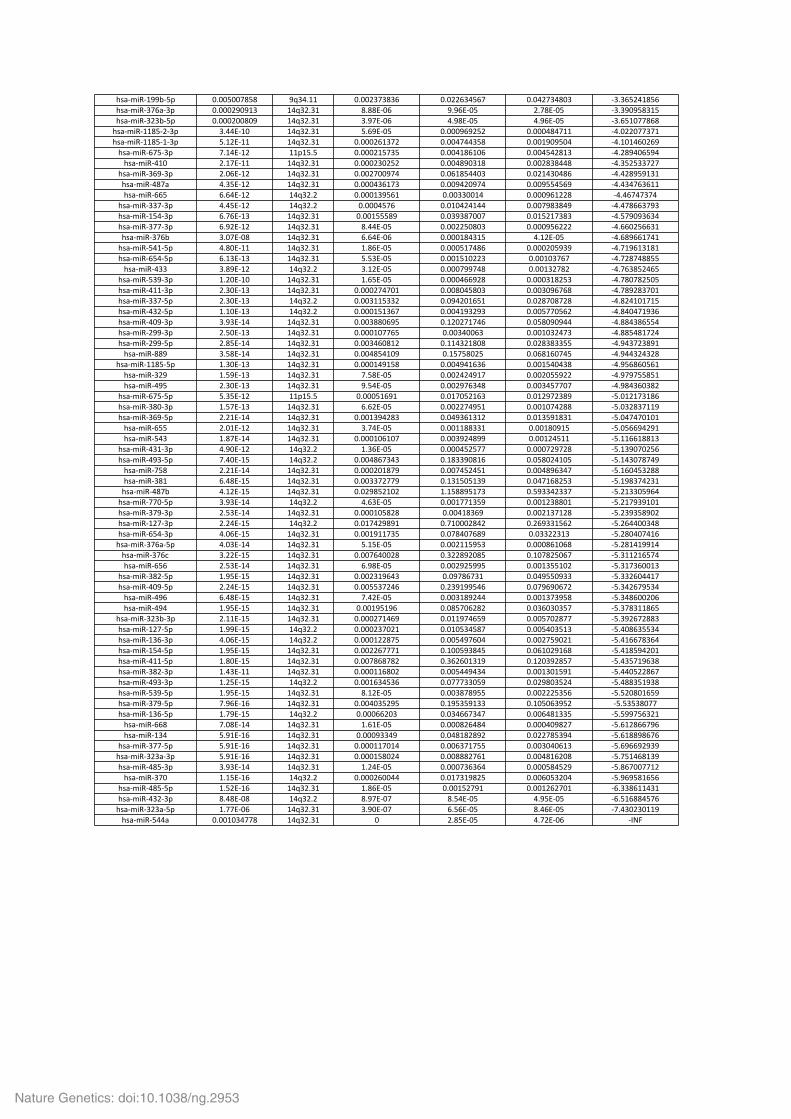
!"#$%&'$())*$+, -.--+--/0+0 )123.(( -.--42/2025 -.-44523+5/ -.-34/230-2 $2.25+43(0+5!"#$%&'$2/5#$2, -.---4)-)(2 (3124.2( 0.006$-5 ).)56$-+ 4./06$-+ $2.2)-)+02(+!"#$%&'$242*$+, -.---4--0-) (3124.2( 2.)/6$-5 3.)06$-+ 3.)56$-+ $2.5+(-//050!"#$%&'$((0+$4$2, 2.336$(- (3124.2( +.5)6$-+ -.---)5)4+4 -.---303/(( $3.-44-//2/(!"#$%&'$((0+$($2, +.(46$(( (3124.2( -.---45(2/4 -.--3/332+0 -.--()-)+-3 $3.(-(35-45)!"#$%&'$5/+$2, /.(36$(4 ((,(+.+ -.---4(+/2+ -.--3(05(-5 -.--3+340(2 $3.40)3-5+)3!"#$%&'$3(- 4.(/6$(( (3124.2( -.---42-4+4 -.--30)-2(0 -.--4020330 $3.2+4+22/4/
!"#$%&'$25)$2, 4.-56$(4 (3124.2( -.--4/--)/3 -.-5(0+33-2 -.-4(32-305 $3.340)+)(2(!"#$%&'$30/# 3.2+6$(4 (3124.2( -.---325(/2 -.--)34-)/3 -.--)++3+5) $3.323/525((!"#$%&'$55+ 5.536$(4 (3124.4 -.---(2)+5( -.--22--(3 -.---)5(440 $3.35/3/2/3
!"#$%&'$22/$2, 3.3+6$(4 (3124.4 -.---3+/5 -.-(-343(33 -.--/)0203) $3.3/0552/)2!"#$%&'$(+3$2, 5./56$(2 (3124.2( -.--(+++0) -.-2)20/--/ -.-(+4(/202 $3.+/)-)2523!"#$%&'$2//$2, 5.)46$(4 (3124.2( 0.336$-+ -.--44+-0-2 -.---)+5444 $3.55-4+552(!"#$%&'$2/5* 2.-/6$-0 (3124.2( 5.536$-5 -.---(032(+ 3.(46$-+ $3.50)55(/3(!"#$%&'$+3($+, 3.0-6$(( (3124.2( (.056$-+ -.---+(/305 -.---4-+)2) $3./()5(2(0(!"#$%&'$5+3$+, 5.(26$(2 (3124.2( +.+26$-+ -.--(+(-442 -.--(-2/5/ $3./40/300++!"#$%&'$322 2.0)6$(4 (3124.4 2.(46$-+ -.---/))/30 -.--(24/04 $3./520+435+
!"#$%&'$+2)$2, (.4-6$(- (3124.2( (.5+6$-+ -.---355)40 -.---2(04+2 $3./0-/04+-+!"#$%&'$3(($2, 4.2-6$(2 (3124.2( -.---4/3/-( -.--0-3+0-2 -.--2-)5/50 $3./0)402/-(!"#$%&'$22/$+, 4.2-6$(2 (3124.4 -.--2((+224 -.-)34-(5+( -.-40/-0/40 $3.043(-(/(+!"#$%&'$324$+, (.(-6$(2 (3124.4 -.---(+(25/ -.--3()24)2 -.--+//-+54 $3.03-3/()25!"#$%&'$3-)$2, 2.)26$(3 (3124.2( -.--200-5)+ -.(4-4/(/35 -.-+0-)-)33 $3.003205++3!"#$%&'$4))$2, 4.+-6$(2 (3124.2( -.---(-//5+ -.--23--52 -.--(-243/2 $3.00+30(/43!"#$%&'$4))$+, 4.0+6$(3 (3124.2( -.--235-0(4 -.((324(0-0 -.-402022++ $3.)32/420)(!"#$%&'$00) 2.+06$(3 (3124.2( -.--30+3(-) -.(+/+0-4+ -.-50(5-/3+ $3.)33243240
!"#$%&'$((0+$+, (.2-6$(2 (3124.2( -.---(3)(+0 -.--3)3(525 -.--(+3-320 $3.)+505-+5(!"#$%&'$24) (.+)6$(2 (3124.2( /.+06$-+ -.--4343)(/ -.--4-++)44 $3.)/)/++0+(!"#$%&'$3)+ 4.2-6$(2 (3124.2( ).+36$-+ -.--4)/5230 -.--23+//-/ $3.)0325-204
!"#$%&'$5/+$+, +.2+6$(4 ((,(+.+ -.---+(5)( -.-(/-+4(52 -.-(4)/420) $+.-(4(/2(05!"#$%&'$20-$2, (.+/6$(2 (3124.2( 5.546$-+ -.--44/3)+( -.--(-/3400 $+.-2402/(()!"#$%&'$25)$+, 4.4(6$(3 (3124.2( -.--(2)3402 -.-3)25(2(4 -.-(2+)(02( $+.-3/3/-(-(!"#$%&'$5++ 4.-(6$(4 (3124.2( 2./36$-+ -.--((0022( -.--(0-)(+ $+.-+55)34)(!"#$%&'$+32 (.0/6$(3 (3124.2( -.---(-5(-/ -.--2)430)) -.--(43+(( $+.((55(00(2
!"#$%&'$32($2, 3.)-6$(4 (3124.4 (.256$-+ -.---3+4+// -.---/4)/40 $+.(2)-/-4+5!"#$%&'$3)2$+, /.3-6$(+ (3124.4 -.--305/232 -.(022)-0(5 -.-+0-43(-+ $+.(32-/0/3)!"#$%&'$/+0 4.4(6$(3 (3124.2( -.---4-(0/) -.--/3+43+( -.--30)523/ $+.(5-3+2400!"#$%&'$20( 5.306$(+ (3124.2( -.--22/4//) -.(2(+-+(2) -.-3/(504+2 $+.()02/342(!"#$%&'$30/* 3.(46$(+ (3124.2( -.-4)0+4(-4 (.(+00)+(/2 -.+)223422/ $+.4(22-+)53!"#$%&'$//-$+, 2.)26$(3 (3124.4 3.526$-+ -.--(//(2+) -.--(4200-( $+.4(/)2)(-(!"#$%&'$2/)$2, 4.+26$(3 (3124.2( -.---(-+040 -.--3(025) -.--4(2/(40 $+.42)2+0)-4!"#$%&'$(4/$2, 4.436$(+ (3124.4 -.-(/34)0)( -./(---4034 -.45)22(+54 $+.4533--230!"#$%&'$5+3$2, 3.-56$(+ (3124.2( -.--()((/2+ -.-/03-/50) -.-22442(2 $+.40-3-/3(5!"#$%&'$2/5#$+, 3.-26$(3 (3124.2( +.(+6$-+ -.--4((+)+2 -.---05(-50 $+.40(3())(3!"#$%&'$2/57 2.446$(+ (3124.2( -.--/53--40 -.2440)4-0+ -.(-/04+-5/ $+.2((4(5+/3!"#$%&'$5+5 4.+26$(3 (3124.2( 5.)06$-+ -.--4)4+))+ -.--(2++(-4 $+.2(/25--(2
!"#$%&'$204$+, (.)+6$(+ (3124.2( -.--42()532 -.-)/05/2( -.-3)++-)22 $+.2245-33(/!"#$%&'$3-)$+, 4.436$(+ (3124.2( -.--++2/435 -.42)())+35 -.-/)5)-5/4 $+.2345/)+23!"#$%&'$3)5 5.306$(+ (3124.2( /.346$-+ -.--2(0)433 -.--(2/2)+0 $+.2305--4-5!"#$%&'$3)3 (.)+6$(+ (3124.2( -.--()+()5 -.-0+/-5404 -.-25-2-2+/ $+.2/02((05+
!"#$%&'$242*$2, 4.((6$(+ (3124.2( -.---4/(35) -.-(()/35+) -.--+/-40// $+.2)45/4002!"#$%&'$(4/$+, (.))6$(+ (3124.4 -.---42/-4( -.-(-+23+0/ -.--+3-2+(2 $+.3-052++23!"#$%&'$(25$2, 3.-56$(+ (3124.4 -.---(440/+ -.--+3)/5-3 -.--4/+)-4( $+.3(55/0253!"#$%&'$(+3$+, (.)+6$(+ (3124.2( -.--445///( -.(--+)203+ -.-5(-4)(50 $+.3(0+)34-(!"#$%&'$3(($+, (.0-6$(+ (3124.2( -.--/050/04 -.2545-(2() -.(4-2)40+/ $+.32+/()520!"#$%&'$204$2, (.326$(( (3124.2( -.---((50-4 -.--+33)323 -.--(2-(+)( $+.33-+4405/!"#$%&'$3)2$2, (.4+6$(+ (3124.4 -.--(523+25 -.-///22-+) -.-4)0-2+43 $+.3002+()20!"#$%&'$+2)$+, (.)+6$(+ (3124.2( 0.(46$-+ -.--20/0)++ -.--444+2+5 $+.+4-0-(5+)!"#$%&'$2/)$+, /.)56$(5 (3124.2( -.--3-2+4)+ -.()+2+)(22 -.(-+-52)+4 $+.+2+20-//!"#$%&'$(25$+, (./)6$(+ (3124.4 -.---554-2 -.-2355/23/ -.--530(22+ $+.+))/+524(!"#$%&'$550 /.-06$(3 (3124.2( (.5(6$-+ -.---045303 -.---3-)04/ $+.5(4055/)5!"#$%&'$(23 +.)(6$(5 (3124.2( -.---)223) -.-30(040)4 -.-44/0+2)3 $+.5(00)05/5
!"#$%&'$2//$+, +.)(6$(5 (3124.2( -.---((/-(3 -.--52/(/++ -.--2-3-5(2 $+.5)55)4)2)!"#$%&'$242#$2, +.)(6$(5 (3124.2( -.---(+0-43 -.--0004/5( -.--30(54-0 $+./+(350(2)!"#$%&'$30+$2, 2.)26$(3 (3124.2( (.436$-+ -.---/25253 -.---+03+4) $+.05/--//(4!"#$%&'$2/- (.(+6$(5 (3124.4 -.---45--33 -.-(/2()04+ -.--5-+24-3 $+.)5)+0(5+5
!"#$%&'$30+$+, (.+46$(5 (3124.2( (.056$-+ -.--(+4/)( -.--(454/-( $5.2205((32(!"#$%&'$324$2, 0.306$-0 (3124.4 0.)/6$-/ 0.+36$-+ 3.)+6$-+ $5.+(5003+/5!"#$%&'$242#$+, (.//6$-5 (3124.2( 2.)-6$-/ 5.+56$-+ 0.356$-+ $/.32-42-(()!"#$%&'$+33# -.--(-23//0 (3124.2( - 4.0+6$-+ 3./46$-5 $89:
Nature Genetics: doi:10.1038/ng.2953
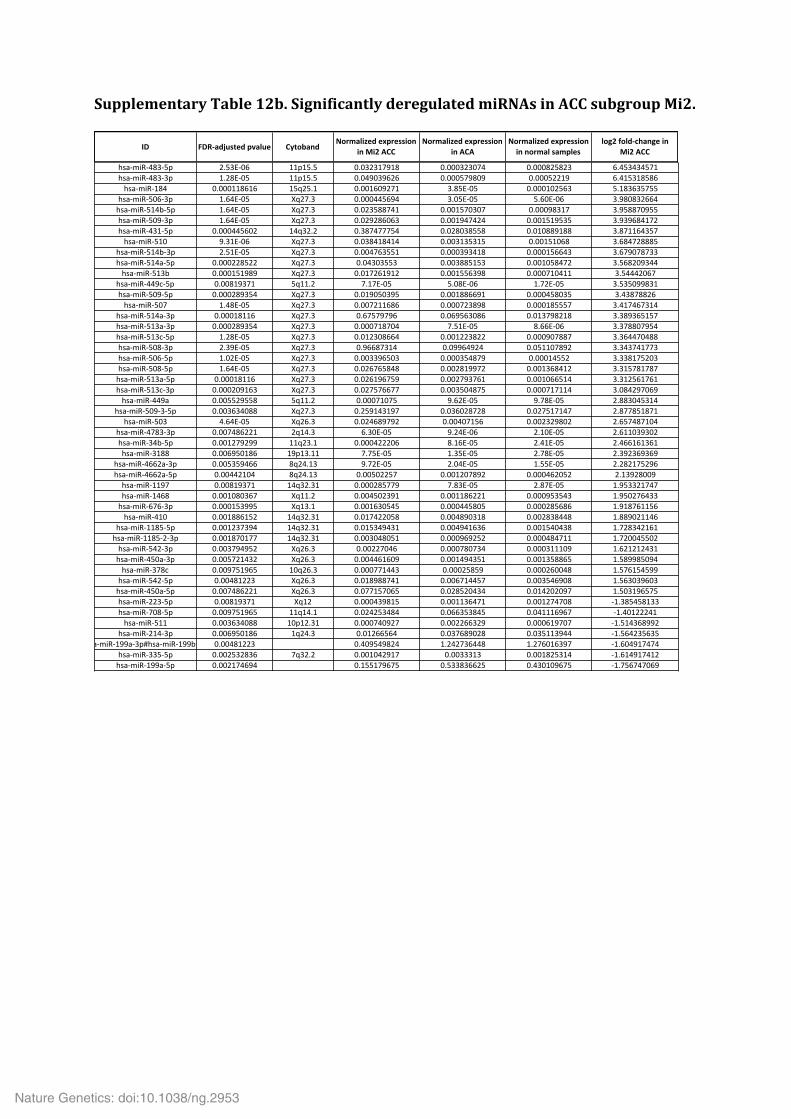
Supplementary Table 12b. Significantly deregulated miRNAs in ACC subgroup Mi2.
!" #"$%&'()*+,'-./&0), 12+34&5'6378&09:,'-,;.7,**935-
95-<9=->116378&09:,'-,;.7,**935-
95->1>6378&09:,'-,;.7,**935-95-5378&0-*&8.0,*
03?=-@30'%AB&5?,-95-<9=->11
!"#$%&'$()*$+, -.+*/$01 22,2+.+ 0.0*-*2342) 0.000*-*03( 0.000)-+)-* 1.(+*(*(+32!"#$%&'$()*$*, 2.-)/$0+ 22,2+.+ 0.0(40*41-1 0.000+34)04 0.000+--24 1.(2+*2)+)1!"#$%&'$2)( 0.00022)121 2+5-+.2 0.002104-32 *.)+/$0+ 0.00020-+1* +.2)*1*+3++
!"#$%&'$+01$*, 2.1(/$0+ 65-3.* 0.000((+14( *.0+/$0+ +.10/$01 *.4)0)*-11(!"#$%&'$+2(7$+, 2.1(/$0+ 65-3.* 0.0-*+))3(2 0.002+30*03 0.0004)*23 *.4+))304++!"#$%&'$+04$*, 2.1(/$0+ 65-3.* 0.0-4-)101* 0.0024(3(-( 0.002+24+*+ *.4*41)(23-!"#$%&'$(*2$+, 0.000((+10- 2(5*-.- 0.*)3(333+( 0.0-)0*)++) 0.020))42)) *.)3221(*+3!"#$%&'$+20 4.*2/$01 65-3.* 0.0*)(2)(2( 0.00*2*+*2+ 0.002+201) *.1)(3-)))+
!"#$%&'$+2(7$*, -.+2/$0+ 65-3.* 0.00(31*++2 0.000*4*(2) 0.0002+11(* *.13403)3**!"#$%&'$+2(#$+, 0.000--)+-- 65-3.* 0.0(*0*++* 0.00*))+2+* 0.0020+)(3- *.+1)-04*((!"#$%&'$+2*7 0.0002+24)4 65-3.* 0.023-1242- 0.002++1*4) 0.000320(22 *.+(((-013
!"#$%&'$((48$+, 0.00)24*32 +522.- 3.23/$0+ +.0)/$01 2.3-/$0+ *.+*+044)*2!"#$%&'$+04$+, 0.000-)4*+( 65-3.* 0.0240+0*4+ 0.002))1142 0.000(+)0*+ *.(*)3))-1!"#$%&'$+03 2.()/$0+ 65-3.* 0.003-221)1 0.0003-*)4) 0.0002)+++3 *.(23(13*2(
!"#$%&'$+2(#$*, 0.0002)221 65-3.* 0.13+34341 0.014+1*0)1 0.02*34)-2) *.*)4*1+2+3!"#$%&'$+2*#$*, 0.000-)4*+( 65-3.* 0.00032)30( 3.+2/$0+ ).11/$01 *.*3))034+(!"#$%&'$+2*8$+, 2.-)/$0+ 65-3.* 0.02-*0)11( 0.002--*)-- 0.000403))3 *.*1((30())!"#$%&'$+0)$*, -.*4/$0+ 65-3.* 0.411)3*2( 0.0441(4-( 0.0+2203)4- *.*(*3(233*!"#$%&'$+01$+, 2.0-/$0+ 65-3.* 0.00**41+0* 0.000*+()34 0.0002(++- *.**)23+-0*!"#$%&'$+0)$+, 2.1(/$0+ 65-3.* 0.0-131+)() 0.00-)2443- 0.002*1)(2- *.*2+3)23)3!"#$%&'$+2*#$+, 0.0002)221 65-3.* 0.0-12413+4 0.00-34*312 0.002011+2( *.*2-+12312!"#$%&'$+2*8$*, 0.000-0421* 65-3.* 0.0-3+31133 0.00*+0()3+ 0.00032322( *.0)(-43014!"#$%&'$((4# 0.00++-4++) +522.- 0.0003203+ 4.1-/$0+ 4.3)/$0+ -.))*0(+*2(
!"#$%&'$+04$*$+, 0.00*1*(0)) 65-3.* 0.-+42(*243 0.0*10-)3-) 0.0-3+232(3 -.)33)+2)32!"#$%&'$+0* (.1(/$0+ 65-1.* 0.0-(1)434- 0.00(032+1 0.00-*-4)0- -.1+3()320(
!"#$%&'$(3)*$*, 0.003()1--2 -52(.* 1.*0/$0+ 4.-(/$01 -.20/$0+ -.1220*4*0-!"#$%&'$*(7$+, 0.002-34-44 225-*.2 0.000(---01 ).21/$0+ -.(2/$0+ -.(11212*12!"#$%&'$*2)) 0.0014+02)1 24,2*.22 3.3+/$0+ 2.*+/$0+ -.3)/$0+ -.*4-*14*14
!"#$%&'$(11-#$*, 0.00+*+4(11 )5-(.2* 4.3-/$0+ -.0(/$0+ 2.++/$0+ -.-)-23+-41!"#$%&'$(11-#$+, 0.00((-20( )5-(.2* 0.00+0--+3 0.002-03)4- 0.000(1-0+- -.2*4-)004!"#$%&'$2243 0.00)24*32 2(5*-.*2 0.000-)+334 3.)*/$0+ -.)3/$0+ 2.4+**-23(3!"#$%&'$2(1) 0.0020)0*13 6522.- 0.00(+0-*42 0.0022)1--2 0.0004+*+(* 2.4+0-31(**!"#$%&'$131$*, 0.0002+*44+ 652*.2 0.0021*0+(+ 0.000((+)0+ 0.000-)+1)1 2.42)3122+1!"#$%&'$(20 0.002))12+- 2(5*-.*2 0.023(--0+) 0.00()40*2) 0.00-)*)(() 2.))40-22(1
!"#$%&'$22)+$+, 0.002-*3*4( 2(5*-.*2 0.02+*(4(*2 0.00(4(21*1 0.002+(0(*) 2.3-)*(-212!"#$%&'$22)+$-$*, 0.002)30233 2(5*-.*2 0.00*0()0+2 0.000414-+- 0.000()(322 2.3-00(++0-!"#$%&'$+(-$*, 0.00*34(4+- 65-1.* 0.00--30(1 0.0003)03*( 0.000*22204 2.1-2-2-(*2!"#$%&'$(+0#$*, 0.00+3-2(*- 65-1.* 0.00((12104 0.002(4(*+2 0.002*+))1+ 2.+)44)+04(!"#$%&'$*3)8 0.0043+241+ 205-1.* 0.000332((* 0.000-+)+4 0.000-100() 2.+312+(+44!"#$%&'$+(-$+, 0.00()2--* 65-1.* 0.02)4))3(2 0.00132((+3 0.00*+(140) 2.+1*0*410*!"#$%&'$(+0#$+, 0.003()1--2 65-1.* 0.0332+301+ 0.0-)+-0(*( 0.02(-0-043 2.+0*241+3+!"#$%&'$--*$+, 0.00)24*32 652- 0.000(*4)2+ 0.0022*1(32 0.002-3(30) $2.*)+(+)2**!"#$%&'$30)$+, 0.0043+241+ 2252(.2 0.0-(-+*()( 0.011*+*)(+ 0.0(2221413 $2.(02---(2!"#$%&'$+22 0.00*1*(0)) 20,2-.*2 0.0003(04-3 0.00--11*-4 0.000124303 $2.+2(*1)44-
!"#$%&'$-2($*, 0.0014+02)1 25-(.* 0.02-11+1( 0.0*31)40-) 0.0*+22*4(( $2.+1(-*+1*+!"#$%&'$244#$*,9!"#$%&'$2447$*, 0.00()2--* 0.(04+(4)-( 2.-(-3*1(() 2.-31021*43 $2.10(423(3(
!"#$%&'$**+$+, 0.00-+*-)*1 35*-.- 0.0020(-423 0.00***2* 0.002)-+*2( $2.12(423(2-!"#$%&'$244#$+, 0.00-23(14( 0.2++23413+ 0.+**)*11-+ 0.(*020413+ $2.3+13(3014
Nature Genetics: doi:10.1038/ng.2953
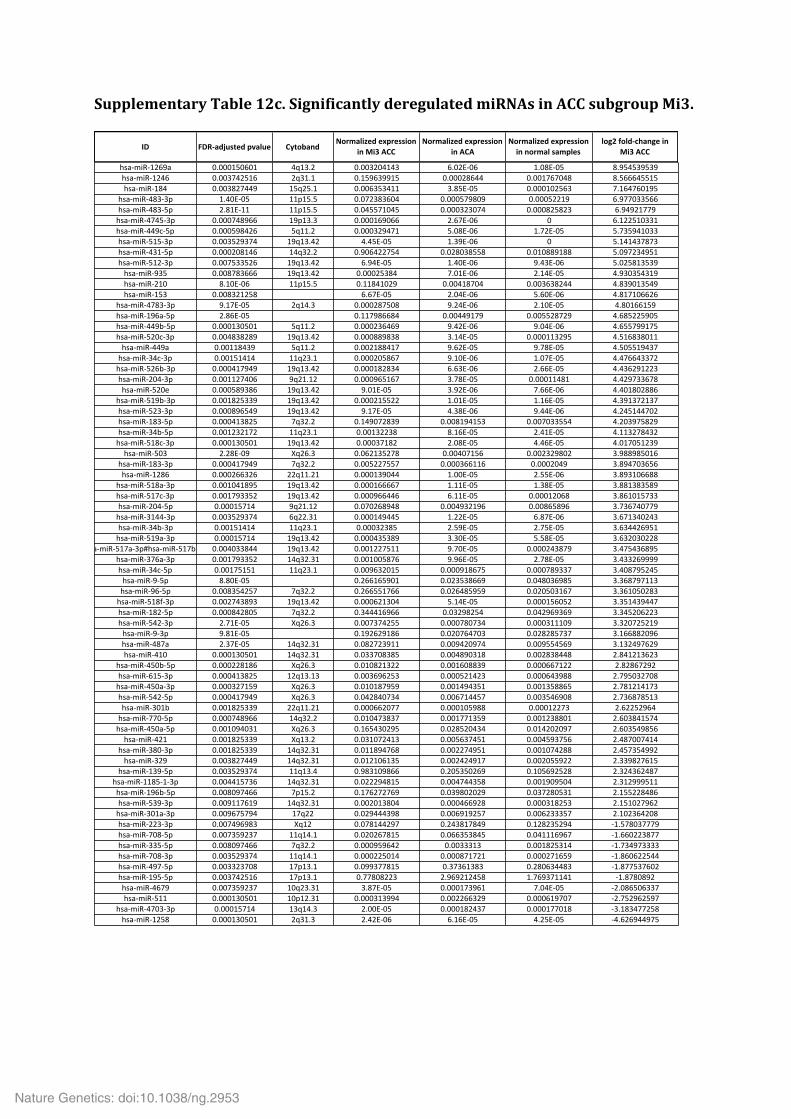
Supplementary Table 12c. Significantly deregulated miRNAs in ACC subgroup Mi3.
!" #"$%&'()*+,'-./&0), 12+34&5'6378&09:,'-,;.7,**935-
95-<9=->116378&09:,'-,;.7,**935-
95->1>6378&09:,'-,;.7,**935-95-5378&0-*&8.0,*
03?@-A30'%BC&5?,-95-<9=->11
!"#$%&'$()*+# ,-,,,(.,*,( /0(1-) ,-,,1),/(/1 *-,)2$,* (-,32$,. 3-+./.1+.1+!"#$%&'$()/* ,-,,14/).(* )01(-( ,-(.+*1++(. ,-,,,)3*// ,-,,(4*4,/3 3-.***/..(.!"#$%&'$(3/ ,-,,13)4//+ (.0).-( ,-,,*1.1/(( 1-3.2$,. ,-,,,(,).*1 4-(*/4*,(+.
!"#$%&'$/31$15 (-/,2$,. ((5(.-. ,-,4)131*,/ ,-,,,.4+3,+ ,-,,,.))(+ *-+44,11.**!"#$%&'$/31$.5 )-3(2$(( ((5(.-. ,-,/..4(,/. ,-,,,1)1,4/ ,-,,,3).3)1 *-+/+)(44+!"#$%&'$/4/.$15 ,-,,,4/3+** (+5(1-1 ,-,,,(*+,** )-*42$,* , *-()).(,11(!"#$%&'$//+6$.5 ,-,,,.+3/)* .0((-) ,-,,,1)+/4( .-,32$,* (-4)2$,. .-41.+/(,11!"#$%&'$.(.$15 ,-,,1.)+14/ (+0(1-/) /-/.2$,. (-1+2$,* , .-(/(/14341!"#$%&'$/1($.5 ,-,,,),3(/* (/01)-) ,-+,*/))4./ ,-,)3,13..3 ,-,(,33+(33 .-,+4)1/+.(!"#$%&'$.()$15 ,-,,4.11.)* (+0(1-/) *-+/2$,. (-/,2$,* +-/12$,* .-,).3(1.1+!"#$%&'$+1. ,-,,3431*** (+0(1-/) ,-,,,).13/ 4-,(2$,* )-(/2$,. /-+1,1./1(+!"#$%&'$)(, 3-(,2$,* ((5(.-. ,-((3/(,)+ ,-,,/(34,/ ,-,,1*13)// /-31+,(1./+!"#$%&'$(.1 ,-,,31)().3 *-*42$,. )-,/2$,* .-*,2$,* /-3(4(,**)*
!"#$%&'$/431$15 +-(42$,. )0(/-1 ,-,,,)34.,3 +-)/2$,* )-(,2$,. /-3,(**(.+!"#$%&'$(+*#$.5 )-3*2$,. ,-((4+3**3/ ,-,,//+(4+ ,-,,..)34)+ /-*3.)).+,.!"#$%&'$//+7$.5 ,-,,,(1,.,( .0((-) ,-,,,)1*/*+ +-/)2$,* +-,/2$,* /-*..4++(4.!"#$%&'$.),6$15 ,-,,/313)3+ (+0(1-/) ,-,,,33+313 1-(/2$,. ,-,,,((1)+. /-.(*313,((!"#$%&'$//+# ,-,,((3/1+ .0((-) ,-,,)(33/(4 +-*)2$,. +-432$,. /-.,..(+/14!"#$%&'$1/6$15 ,-,,(.(/(/ ((0)1-( ,-,,,),.3*4 +-(,2$,* (-,42$,. /-/4**/114)!"#$%&'$.)*7$15 ,-,,,/(4+/+ (+0(1-/) ,-,,,(3)31/ *-*12$,* )-**2$,. /-/1*)+())1!"#$%&'$),/$15 ,-,,(()4/,* +0)(-() ,-,,,+*.(*4 1-432$,. ,-,,,((/3( /-/)+411*43!"#$%&'$.),8 ,-,,,.3+13* (+0(1-/) +-,(2$,. 1-+)2$,* 4-**2$,* /-/,(3,)33*
!"#$%&'$.(+7$15 ,-,,(3).11+ (+0(1-/) ,-,,,)(..)) (-,(2$,. (-(*2$,. /-1+(14)(14!"#$%&'$.)1$15 ,-,,,3+*./+ (+0(1-/) +-(42$,. /-132$,* +-//2$,* /-)/.(//4,)!"#$%&'$(31$.5 ,-,,,/(13). 401)-) ,-(/+,4)31+ ,-,,3(+/(.1 ,-,,4,11../ /-),1+4.3)+!"#$%&'$1/7$.5 ,-,,()1)(4) ((0)1-( ,-,,(1))13 3-(*2$,. )-/(2$,. /-((1)43/1)!"#$%&'$.(36$15 ,-,,,(1,.,( (+0(1-/) ,-,,,14(3) )-,32$,. /-/*2$,. /-,(4,.()1+!"#$%&'$.,1 )-)32$,+ 90)*-1 ,-,*)(1.)43 ,-,,/,4(.* ,-,,)1)+3,) 1-+33+3.,(*
!"#$%&'$(31$15 ,-,,,/(4+/+ 401)-) ,-,,.))4..4 ,-,,,1**((* ,-,,,),/+ 1-3+/4,1*.*!"#$%&'$()3* ,-,,,)**1)* ))0((-)( ,-,,,(1+,// (-,,2$,. )-..2$,* 1-3+1(,**33
!"#$%&'$.(3#$15 ,-,,(,/(3+. (+0(1-/) ,-,,,(****4 (-((2$,. (-132$,. 1-33(131.3+!"#$%&'$.(46$15 ,-,,(4+11.) (+0(1-/) ,-,,,+**//* *-((2$,. ,-,,,(),*3 1-3*(,(.411!"#$%&'$),/$.5 ,-,,,(.4(/ +0)(-() ,-,4,)*3+/3 ,-,,/+1)(+* ,-,,3*.3+* 1-41*4/,44+!"#$%&'$1(//$15 ,-,,1.)+14/ *0))-1( ,-,,,(/+//. (-))2$,. *-342$,* 1-*4(1/,)/1!"#$%&'$1/7$15 ,-,,(.(/(/ ((0)1-( ,-,,,1)13. )-.+2$,. )-4.2$,. 1-*1//)*+.(!"#$%&'$.(+#$15 ,-,,,(.4(/ (+0(1-/) ,-,,,/1.13+ 1-1,2$,. .-.32$,. 1-*1),1,))3
!"#$%&'$.(4#$15:!"#$%&'$.(47$15 ,-,,/,113// (+0(1-/) ,-,,())4.(( +-4,2$,. ,-,,,)/134+ 1-/4./1*3+.!"#$%&'$14*#$15 ,-,,(4+11.) (/01)-1( ,-,,(,,.34* +-+*2$,. )-432$,. 1-/11)*++++!"#$%&'$1/6$.5 ,-,,(4.(.( ((0)1-( ,-,,+*1),(. ,-,,,+(3*4. ,-,,,43+114 1-/,34+.)/.!"#$%&'$+$.5 3-3,2$,. ,-)**(*.+,( ,-,)1.13**+ ,-,/3,1*+3. 1-1*34+4((1!"#$%&'$+*$.5 ,-,,31./).4 401)-) ,-)**..(4** ,-,)*/3.+.+ ,-,),.,1(*4 1-1*(,.,)31!"#$%&'$.(3;$15 ,-,,)4/13+1 (+0(1-/) ,-,,,*)(1,/ .-(/2$,. ,-,,,(.*,.) 1-1.(/1+//4!"#$%&'$(3)$.5 ,-,,,3/)3,. 401)-) ,-1///(*+** ,-,1)+3)./ ,-,/)+*+1*+ 1-1/.),*))1!"#$%&'$./)$15 )-4(2$,. 90)*-1 ,-,,414/).. ,-,,,43,41/ ,-,,,1(((,+ 1-1),4).)(+!"#$%&'$+$15 +-3(2$,. ,-(+)*)+(3* ,-,),4*/4,1 ,-,)3)3.414 1-(**33),+*!"#$%&'$/34# )-142$,. (/01)-1( ,-,3)4)1+(( ,-,,+/),+4/ ,-,,+../.*+ 1-(1)/+4*)+!"#$%&'$/(, ,-,,,(1,.,( (/01)-1( ,-,114,313. ,-,,/3+,1(3 ,-,,)313//3 )-3/()(1*)1
!"#$%&'$/.,7$.5 ,-,,,))3(3* 90)*-1 ,-,(,3)(1)) ,-,,(*,331+ ,-,,,**4()) )-3)3*4)+)!"#$%&'$*(.$15 ,-,,,/(13). ()0(1-(1 ,-,,1*+*).1 ,-,,,.)(/)1 ,-,,,*/1+33 )-4+.,1)4,3!"#$%&'$/.,#$15 ,-,,,1)4(.+ 90)*-1 ,-,(,(34+.+ ,-,,(/+/1.( ,-,,(1.33*. )-43()(/(41!"#$%&'$./)$.5 ,-,,,/(4+/+ 90)*-1 ,-,/)3/,41/ ,-,,*4(//.4 ,-,,1./*+,3 )-41*343.(1!"#$%&'$1,(7 ,-,,(3).11+ ))0((-)( ,-,,,**),44 ,-,,,(,.+33 ,-,,,())41 )-*)).)+*/!"#$%&'$44,$.5 ,-,,,4/3+** (/01)-) ,-,(,/41314 ,-,,(44(1.+ ,-,,()133,( )-*,13/(.4/!"#$%&'$/.,#$.5 ,-,,(,+/,1( 90)*-1 ,-(*./1,)+. ,-,)3.),/1/ ,-,(/),),+4 )-*,1./+3.*!"#$%&'$/)( ,-,,(3).11+ 90(1-) ,-,1(,4)/(1 ,-,,.*14/.( ,-,,/.+14.* )-/34,,4/(/
!"#$%&'$13,$15 ,-,,(3).11+ (/01)-1( ,-,((3+/4*3 ,-,,))4/+.( ,-,,(,4/)33 )-/.41./++)!"#$%&'$1)+ ,-,,13)4//+ (/01)-1( ,-,()(,*(1. ,-,,)/)/+(4 ,-,,),..+)) )-11+3)4*(.
!"#$%&'$(1+$.5 ,-,,1.)+14/ ((0(1-/ ,-+31(,+3** ,-),.1.,)*+ ,-(,.*+).)3 )-1)/1*)/34!"#$%&'$((3.$($15 ,-,,//(.41* (/01)-1( ,-,)))+/3(. ,-,,/4//1.3 ,-,,(+,+.,/ )-1()+++.((!"#$%&'$(+*7$.5 ,-,,3,+4/** 45(.-) ,-(4*)4)4*+ ,-,1+3,),)+ ,-,14)3,.1( )-(..))3/3*!"#$%&'$.1+$15 ,-,,+((4*(+ (/01)-1( ,-,,),(13,/ ,-,,,/**+)3 ,-,,,1(3).1 )-(.(,)4+*)!"#$%&'$1,(#$15 ,-,,+*4.4+/ (40)) ,-,)+///1+3 ,-,,*+(+).4 ,-,,*)111.4 )-(,)1*/),3!"#$%&'$))1$15 ,-,,4/+*+31 90() ,-,43(//)+4 ,-)/13(43/+ ,-()3)1.)+/ $(-.43,1444+!"#$%&'$4,3$.5 ,-,,41.+)14 ((0(/-( ,-,),)*43(. ,-,**1.13/. ,-,/(((*+*4 $(-**,))1344!"#$%&'$11.$.5 ,-,,3,+4/** 401)-) ,-,,,+.+*/) ,-,,111(1 ,-,,(3).1(/ $(-41/+41111!"#$%&'$4,3$15 ,-,,1.)+14/ ((0(/-( ,-,,,)).,(/ ,-,,,34(4)( ,-,,,)4(*.+ $(-3*,*)).//!"#$%&'$/+4$.5 ,-,,11)14,3 (45(1-( ,-,++1443(. ,-141*(131 ,-)3,*1//31 $(-344.14*,)!"#$%&'$(+.$.5 ,-,,14/).(* (45(1-( ,-443,3))1 )-+*+)()/.3 (-4*+14((/( $(-343,3+)!"#$%&'$/*4+ ,-,,41.+)14 (,0)1-1( 1-342$,. ,-,,,(41+*( 4-,/2$,. $)-,3*.,*114!"#$%&'$.(( ,-,,,(1,.,( (,5()-1( ,-,,,1(1++/ ,-,,))**1)+ ,-,,,*(+4,4 $)-4.)+*).+4
!"#$%&'$/4,1$15 ,-,,,(.4(/ (10(/-1 )-,,2$,. ,-,,,(3)/14 ,-,,,(44,(3 $1-(31/44).3!"#$%&'$().3 ,-,,,(1,.,( )01(-1 )-/)2$,* *-(*2$,. /-).2$,. $/-*)*+//+4.
Nature Genetics: doi:10.1038/ng.2953