suplemento/supplement comportamento do … · comportamento do triatoma sordida em condiÇÕes de...
TRANSCRIPT

SUPLEMENTO/SUPPLEMENT
COMPORTAMENTO DO TRIATOMA SORDIDAEM CONDIÇÕES DE LABORATÓRIO
Edmundo JuarezEunice Pinho de Castro Silva
Departamento de Epidemiologia da
Faculdade de Saúde PúblicaUniversidade de São Paulo
Universidade de São Paulo
Faculdade de Saúde Públ ica
São Paulo
1 9 8 2

COMPORTAMENTO DO TRIATOMA SORDIDA EM CONDIÇÕESDE LABORATÓRIO*
Edmundo JuarezEunice Pinho de Castro Silva
Foi observado o comportamento do Triatoma sordida, mantidoem condições de laboratório em estufas com umidade relativa do ar de60-70% e temperatura de 25°C e 30°C. A temperatura mais alta determinouum encurtamento da duração das fases de evolução. Os machos tiveram aduração do 5.° estádio maior do que a das fêmeas. A duração das fêmeasadultas foi maior do que a dos machos. Em relação ao sangue ingeridoobservaram-se médias maiores à temperatura de 25°C, exceto no 5.° estádio,bem como, nos 4.° e 5.° estádios, as fêmeas tiveram médias maiores. O númerode respostas à temperatura de 25.°C foi maior nos 1.°, 2.° e 3.° estádios; no4.° ocorreu interação temperatura-sexo; no 5.° não houve diferença significan-te; o número de repastos nos machos foi maior do que o das fêmeas, apenasno 5.° estádio. O peso inicial dos insetos foi maior, à temperatura de 25°C,em todos os estádios, exceto no 1.°; as fêmeas, à temperatura de 25.°C, tiverampeso maior do que o dos machos nos 4.° e 5.° estádios e, à temperatura de30°C, apenas no 5.° estádio. Foi observado o efeito da temperatura e dosexo sobre a resistência ao jejum do T. sordida que, após a muda para osestádios consecutivos, não foram mais manipulados e nem receberam qual-quer quantidade de alimento. Observou-se que a resistência média ao jejum,medida em dias, foi maior em todos os estádios do ciclo evolutivo, porémna fase adulta ocorreu interação temperatura-sexo. A correlação entre aresistência ao jejum e o último peso do estádio imediatamente anterior foia única que se mostrou sistematicamente positiva, sendo significante emapenas oito das doze possíveis, considerados todos os estádios.
UNITERMOS: Triatoma sordida. Triatomíneos, comportamento em labo-ratório.
*Síntese da tese de livre-docência apresentada à Faculdade de Saúde Pública da USP em 1979.
INTRODUÇÃO
Forattini e col.11,12,13,14,15,16,17 (1971, 1973,1974, 1975, 1977, 1979) estudando os as-pectos ecológicos da tripanossomíase ameri-cana no Estado de São Paulo, mostraramalguns fatos importantes relacionados aocomportamento do Triatoma sordida no seuambiente natural, procurando com istoestimar o potencial que este triatomíneo temcomo vetor do Tripanosoma cruzi e as
possibilidades de ocupar um lugar de des-taque na transmissão da doença de Chagas.Segundo esses autores, o T. sordida temvalência ecológica apreciável, habitandoecótopos artificiais como conseqüência dadispersão ativa de que é capaz. É atraídoe se adapta facilmente aos ecótopos arti-ficiais sendo prevista a invasão de domi-cílios. A sua densidade aumenta quandodesaparecem os inimigos naturais, sendo,

inclusive, capaz de sobreviver às queimadas.Os adultos são encontrados com maior fre-qüência no primeiro semestre do ano, dimi-nuindo em seguida, pela tendência à dis-persão e ocupação de novos ecótopos. Essese outros aspectos ecológicos estudados poresses autores permitem supor que o T.sordida, no Estado de São Paulo, poderávir a desempenhar papel importante na trans-missão do T. cruzi.
A capacidade de resistir ao je jum de dife-rentes espécies de triatomíneos tem desper-tado a curiosidade de pesquisadores, queprocuraram determinar o tempo de sobre-vida desses insetos, quando submetidos acondições de ausência de alimentação. Estascondições, no entanto, foram bastante dife-rentes, segundo o autor e a espécie estu-dada.
Tobar 32 (1952), Pellegrino2 6 (1952),Hack 20 (1955), estudando o T. infestans,Rickmann 30 (1962) estudando o Triatomaprotracta, Dias9 (1965) estudando o T.infestans, o Rhodnius prolixas e o Panstron-gylus megistus e P ipp in 2 9 (1970) estudandoo Triatoma sanguisuga e o Triatoma gers-taeckeri utilizaram diferentes técnicas parasubmeter os triatomíneos ao jejum, as quaispermitiram, inclusive, que ocorressem mudas,além de permitir sobrevivências prolongadas.Isto porque o j e jum a que os submeteramnão era absoluto, isto é, o estado alimentardos insetos após a muda para o estádio aser observado, ou não era bem conhecidoou foram oferecidas pequenas quantidadesde sangue, insuficientes para o pleno desen-volvimento do inseto.
Perlowagora-Szumlevics 28 (l969), Zele-don35 (1970), Costa5 (1972) e Castanho4
(1972), trabalhando respectivamente com T.infestans, Triatoma dimidiata, Triatomabrasiliensis e P. megistus, estudaram ocomportamento dos insetos frente ao jejumabsoluto após a muda, porém utilizaramdiferentes condições de temperatura.
Os trabalhos de Forattini e col.11-17 (1971,1973, 1974, 1975, 1977, 1979) com referênciaao T. sordida no seu ambiente natural,mostram que a valência ecológica deste
triatomíneo é apreciável, bem como é grandea sua capacidade de sobrevivência emambientes adversos tais como as queimadas.Estes fatos, ao lado do grande ecletismoalimentar dos triatomíneos, permitem espe-cular sobre o comportamento do T. sordidano seu ambiente natural e mesmo artificialno que diz respeito à sua capacidade deresistir às dificuldades de encontrar ali-mentos.
O conhecimento do comportamento desseinseto em condições de laboratório, aindaque totalmente artificiais, poderá trazersubsídios de alguma importância para setentar explicar o que ocorre no ambientenatural.
Com o objetivo de contribuir para o conhe-cimento do comportamento do T. sordidaem condições de laboratório, a duas tempe-ratura, e, na expectativa de permitir aformulação da hipótese sobre o seu compor-tamento no ambiente natural, nesta publi-cação são apresentados os resultados obtidosna observação da: 1- duração da fase em-brionária, do ciclo evolutivo e da faseadulta; 2 - necessidade alimentar, número derepastos necessários para obtenção do ali-mento e o seu crescimento ponderai; 3 - re-sistência ao jejum.
MATERIAL E MÉTODOS
Os insetos estudados foram obtidos deuma colônia de T. sordida ao Departamentode Epidemiologia da Faculdade de SaúdePública - USP, iniciada em dezembro de1968, a partir de insetos capturados noambiente domiciliar, procedente de CampoGrande, Estado de Mato Grosso.
Durante seis dias consecutivos, foramrecolhidos todos os ovos dessa colônia eseparados, diariamente, em dois lotes domesmo tamanho, destinados a tratamentodiferente quanto à temperatura, o que foifeito por sorteio.
Para ambos os lotes, observou-se o mesmoprocedimento, até o f inal do estudo, dife-rindo apenas a condição de temperatura.

Um foi submetido à temperatura de 25°Ce o outro à de 30°C.
Diriamente, após a coleta dos ovos edistribuição em dois lotes, os ovos foramcolocados individualmente em frascos deBorrel, numerados seqüencialmente, com ofundo revestido por papel de fil tro. Apósa eclosão, os frascos foram munidos comsuporte de cartolina e fechados com telade nylon de malha f ina , presa com elástico.
Ao f ina l do sexto dia, cada lote estavaconstituído por 346 ovos, resultante dacontribuição diária de 68, 62, 58, 62, 47 e49 ovos.
Em ambas as estufas, manteve-se aumidade relativa do ar entre 60 e 70%,utilizando-se para tanto tiras de papel def i l t ro com uma das extremidades mergulhadaem água contida em cristalizadores.
A alimentação dos insetos foi realizada emcamundongos albinos, imobilizados em telade arame e apoiados sobre o frasco deBorrel, de tal forma que ficassem em íntimocontacto com a tela de vedação, permitindoque o inseto pudesse efetuar o repasto.
Diariamente os insetos foram observados,individualmente, anotando-se as ocorrênciasde interesse para o estudo, atividade reali-zada em intervalos regulares de 24 horas.Em seguida era oferecido o repasto, apenasnos dias predeterminados, isto é, os insetosda estufa mantida a 25°C foram alimentadosem um dia determinado da semana enquantoque os da estufa a 30°C o foram no diaseguinte.
O tempo de oferecimento da fonte aoinseto foi de 40 min, e se constitui nodobro do tempo observado por Dias 8 (1956),que verificou a média de 20 min para otempo de sucção do T. sordida. Por analogiacom o trabalho de Juarez22 (1970), tomou--se o dobro do valor dessa média parapermitir que todos os insetos tivessemoportunidade de sugar o sangue necessário.
Imediatamente antes e após o repasto,os insetos foram pesados em balança analí-tica com precisão de ± 0,1 mg, sendo aquantidade de sangue ingerida determinada
pela diferença entre o peso inicial e o pesofina l nesta pesagem.
As anotações dos dados de interesseforam feitas em protocolos de laboratório,para cada inseto individualmente. Foramanotadas as seguintes informações: data dapostura, data da exclosão, data da muda,data da morte, pesos inicial e final em cadapesagem e quantidade de sangue ingerido.
Aplicou-se o processo de amostragemcasual simples, sempre que se selecionaramamostras destinadas ao estudo do jejum.Entre os ovos foram sorteados 30, e apósa eclosão as correspondentes ninfas doprimeiro estádio permaneceram em jejumabsoluto até a morte. As demais ninfas doprimeiro estádio foram alimentadas e, entreestas, foram sorteadas 30 que deveriam sersubmetidas a je jum absoluto no segundoestádio após a muda. O mesmo procedi-mento foi observado com relação aos demaisestádios, sejam ninfas, sejam de formaadulta.
Era de se desejar que, para a seleçãode uma amostra de ninfas de um determi-nado estádio destinada a jejum no estádioseguinte, fosse aplicada amostragem estrati-f icada em que os critérios de estratificaçãoseriam os correspondentes às variáveis obser-vadas até aquele estádio. Desta forma, paraseleção de uma amostra de ninfas deprimeiro estádio a serem destinadas ao je jumno segundo, deveríamos estratificar as ninfasalimentadas do primeiro estádio, segundoos critérios: tempo de incubação, duração,sangue ingerido, número de repastos e pesoinicial do primeiro estádio. Dada, entre-tanto, a natureza e o número de variáveiscada vez maior às quais correspondiam oscritérios de estratificação, tornou-se inviávela aplicação de tal processo na seleção dasamostras destinadas ao je jum.
O número de insetos destinado ao estudode resistência ao je jum foi diferente de 30em alguns estádios, em decorrência dasperdas verificadas e pela desigualdade naproporção de machos e fêmeas que evoluíramaté a forma adulta.

Os insetos destinados ao estudo de resis-tência ao je jum, após a muda para o estádiocorrespondente não mais foram manipuladosa fim de ser evitado que qualquer trauma-tismo pudesse abreviar a sua sobrevida.
À exceção do procedimento relativo aoj e j u m , a metodologia acima descrita foiutilizada por Jua rez 2 2 (1970) estudando aevolução do ciclo evolutivo do T. infestanssubmetido a diferentes condições de tempe-ratura, 25°C e 30°C, alimentação emcamundongos albinos e galinha e infecçãopelo T. cruzi, em estufas mantidas comumidade relativa do ar entre 60 e 70%.
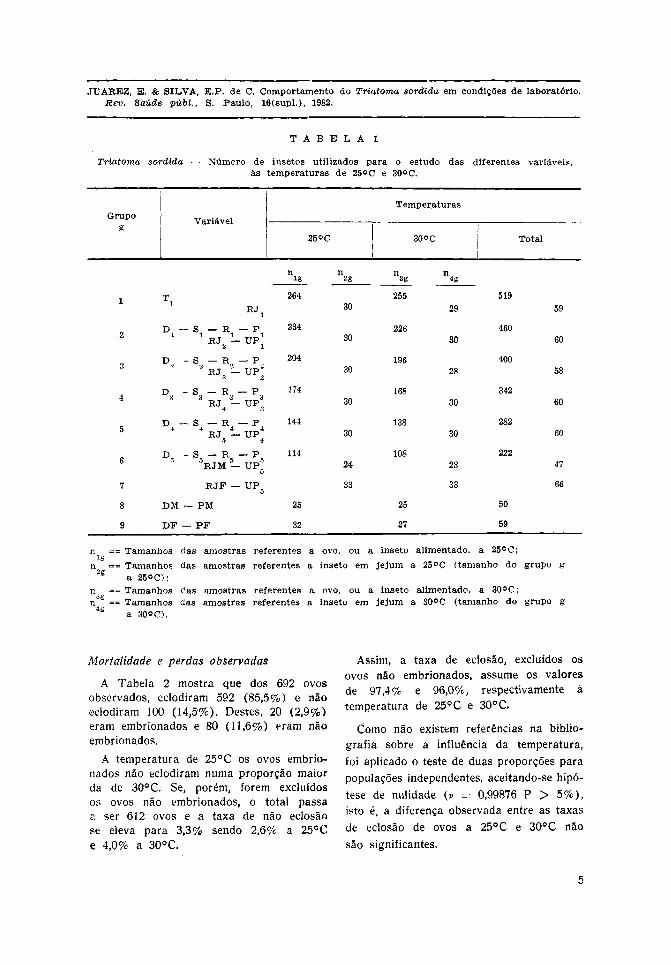
De acordo com a metodologia apresen-tada, para cada temperatura estudada, foramsorteados 9 grupos com diferentes objetivos:
Grupo 1- estudo do je jum da ninfa do1° estádio
Grupo 2 - estudo do je jum da ninfa do2.° estádio
Grupo 3 - estudo do j e jum da ninfa do3.° estádio
Grupo 4 - estudo do je jum da ninfa do4.° estádio
Grupo 5 - estudo do j e j u m da n infa do5.° estádio
Grupo 6 - estudo do j e jum do adultomacho
Grupo 7 - estudo do j e j u m do adultofêmea
Grupo 8 - do adultomacho alimentado
Grupo 9 - estudo da sobrevida do adultofêmea alimentado.
Análise estatística
As características populacionais, média,desvio padrão, mediana, valor máximo, valormínimo, referentes às diversas variáveisestudadas do T. sordida, foram estimadaspelos valores amostrais correspondentesobservados nas amostras de ovos, ninfasdos diferentes estádios e adultos.
A comparação das médias referentes àsvariáveis de estudo e correspondentes àsduas temperaturas foi feita empregando-se
análise de variância a dois critérios, modelofixo, hierárquico ou com classificaçãocruzada e também com o teste de duasmédias.
Na comparação das médias do sexomasculino com as do sexo feminino, bemcomo das quatro médias definidas porsexo e temperatura, empregou-se análisede variância modelo fixo a dois critérioscruzados, teste de Duncan, teste de Schefféou teste de duas médias.
Na Tabela 1 encontra-se a distribuiçãodos insetos segundo as variáveis de estudo,onde é utilizada a seguinte notação: D(duração) , S (sangue ingerido), R (númerode repastos), P (peso inicial do inseto emum determinado estádio), TI (duração dafase embrionária), RJ (resistência ao j e j u m ) ,UP (úl t imo peso do inseto em um determi-nado estádio), M (macho), F (fêmea).
RESULTADOS E COMENTÁRIOS
A fim de verif icar o efeito das tempera-turas e da constituição dos grupos de insetospara o estudo das variáveis, foi realizadaanálise de variância modelo f ixo hierárquico.
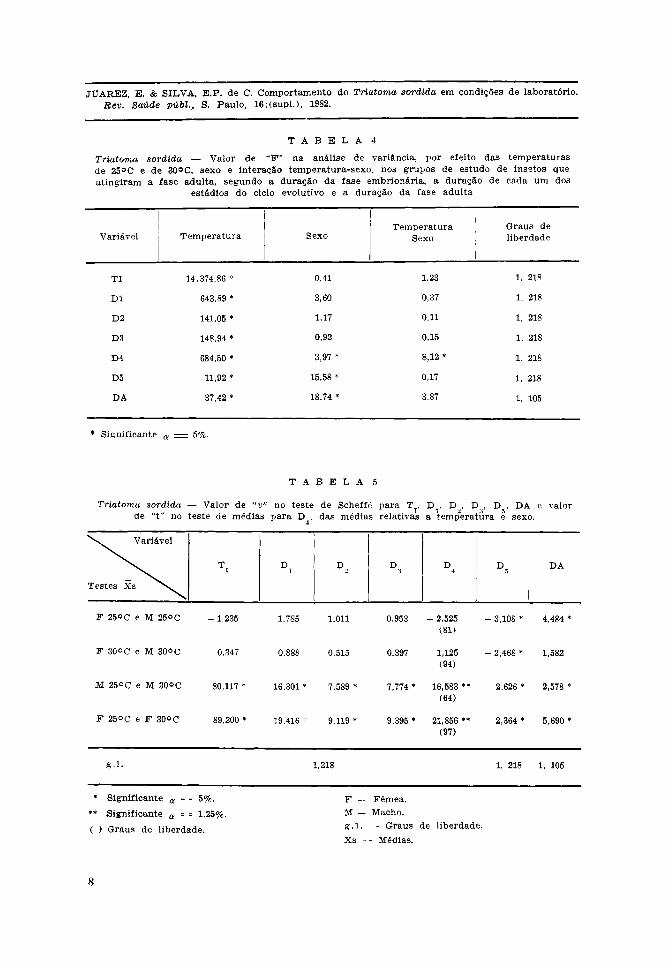
O resultado revelou que a temperaturatem um efeito marcante, pelas diferençassignificantes ao nível de 5%, para todasas variáveis, exceto para sangue ingeridono 5.° estádio, número de repastos no 5.°estádio e peso inic ia l no 1.° estádio.
A comparabilidade destes grupos deinsetos foi constatada em todas as variáveis,exceto para os grupos de estudo referentesa sangue ingerido no 4.° estádio e númerode repastos no 1.° estádio, em que as dife-renças foram significantes ao nível de 5%(F = 3,21 e F = 1,91 respectivamente).
Pelas características do trabalho desen-volvido não seria possível a tomada de todasas possíveis amostras que levassem em contatodos os estratos e, assim, optou-se pelaamostra casual simples. Este fato, associadoa possíveis d i f icu ldades operacionais, poderiaser a causa da não obtenção de gruposcomparáveis para essas variáveis.
Mortalidade e perdas observadas
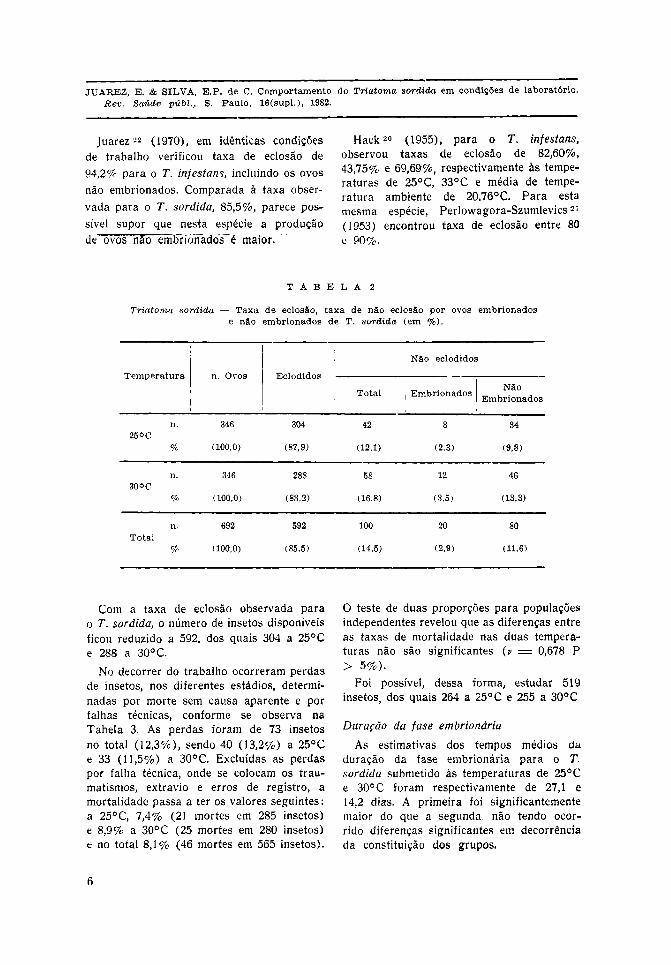
A Tabela 2 mostra que dos 692 ovosobservados, eclodiram 592 (85,5%) e nãoeclodiram 100 (14,5%). Destes, 20 (2,9%)eram embrionados e 80 (11,6%) eram nãoembrionados.
À temperatura de 25°C os ovos embrio-nados não eclodiram numa proporção maiorda de 30°C. Se, porém, forem excluídosos ovos não embrionados, o total passaa ser 612 ovos e a taxa de não eclosãose eleva para 3,3% sendo 2,6% a 25°Ce 4,0% a 30°C.
Assim, a taxa de eclosão, excluídos osovos não embrionados, assume os valoresde 97,4% e 96,0%, respectivamente àtemperatura de 25°C e 30°C.
Como não existem referências na biblio-grafia sobre a influência da temperatura,foi aplicado o teste de duas proporções parapopulações independentes, aceitando-se hipó-tese de nulidade (v = 0,99876 P > 5%),isto é, a diferença observada entre as taxasde eclosão de ovos a 25°C e 30°C nãosão significantes.
Juarez2 2 (1970), em idênticas condiçõesde trabalho verificou taxa de eclosão de94,2% para o T. in fes tans , incluindo os ovosnão embrionados. Comparada à taxa obser-vada para o T. sordida, 85,5%, parece pos-sível supor que nesta espécie a produçãode ovos não embrionados é maior.
Hack2 0 (1955), para o T. infestans,observou taxas de eclosão de 82,60%,43,75% e 69,69%, respectivamente às tempe-raturas de 25°C, 33°C e média de tempe-ratura ambiente de 20,76°C. Para estamesma espécie, Perlowagora-Szumlevics27
(1953) encontrou taxa de eclosão entre 80e 90%.
Com a taxa de eclosão observada parao T. sordida, o número de insetos disponíveisficou reduzido a 592, dos quais 304 a 25°Ce 288 a 30° C.
No decorrer do trabalho ocorreram perdasde insetos, nos diferentes estádios, determi-nadas por morte sem causa aparente e porfalhas técnicas, conforme se observa naTabela 3. As perdas foram de 73 insetosno total (12,3%), sendo 40 (13,2%) a 25°Ce 33 (11,5%) a 30°C. Excluídas as perdaspor falha técnica, onde se colocam os trau-matismos, extravio e erros de registro, amortalidade passa a ter os valores seguintes:a 25°C, 7,4% (21 mortes em 285 insetos)e 8,9% a 30°C (25 mortes em 280 insetos)e no total 8,1% (46 mortes em 565 insetos).
O teste de duas proporções para populaçõesindependentes revelou que as diferenças entreas taxas de mortalidade nas duas tempera-turas não são significantes (v = 0,678 P> 5%).
Foi possível, dessa forma, estudar 519insetos, dos quais 264 a 25°C e 255 a 30°C
Duração da fase embrionária
As estimativas dos tempos médios daduração da fase embrionária para o Tsordida submetido às temperaturas de 25°Ce 30°C foram respectivamente de 27,1 e14,2 dias. A primeira foi significantementemaior do que a segunda, não tendo ocor-rido diferenças significantes em decorrênciada constituição dos grupos.

Também não foram detectadas diferençasdeterminadas pelo sexo, nem pela interaçãotemperatura-sexo (Tabela 4).
O efeito da temperatura, observado parao total de ovos, também foi evidente naobservação dos ovos correspondentes a cadasexo. As médias observadas para a duraçãoda fase embrionária em cada sexo à tempe-ratura de 25°C foram maiores do que ascorrespondentes a 30°C (Tabela 5). Parafêmeas e machos a 25°C foram respectiva-mente de 27,1 e 27,3 dias e a 30°C asmédias correspondentes foram de 14,2 e 14,1dias (Tabelas 4 e 5).
O efeito da temperatura de 30°C, acele-rando a fase embrionária do T. sordida,foi muito semelhante ao observado porJuarez2 2 (1970) estudando o T. infestansem idênticas condições de temperatura eumidade relativa do ar. Essa semelhançafoi tão marcante a ponto de os valoresmáximos e mínimos terem sido exatamenteiguais. Apenas as estimativas das médiasdiferiram muito pouco, com valores de 27.3e 14,4, respectivamente a 25°C e 30°C noT. infestans.
Dias7 (1955) observou cinco exemplaresde ovos de T. sordida, permitindo estimarem 17,0 dias o tempo médio de duração dafase embrionária, em condições de labora-tório sem controle de temperatura.
O efeito favorável de temperaturas maisaltas na aceleração da fase embrionária foiverificado por Neiva2 5 (1913) observandoo T. infestans e afirmando que "todas asfunções são ativadas pelo calor". Esse autorencontrou um mínimo de 16 dias e ummáximo de 33 dias para a duração da faseembrionária, mas que "geralmente desa-lagam depois de 20-25 dias".
Desde então, diversos autores, estudandodiferentes espécies de triatomíneos, têmdemonstrado o mesmo efeito, indicando ovalor observado para a duração da faseembrionária em temperaturas pré-fixadas ouambientais.
Para o T. infeslans, Abalos e Wygodzins-k y 1 (1951) afirmaram que a evolução de-pende da temperatura, encontrando valoresentre 10 e 30 dias para a duração da faseembrionária. Para essa mesma espécie,Perlowagora-Szumlevics27 (1953) mostrouque a 24-28°C os ovos começam a eclodir

no 11.° dia, mas que em geral o fazem do17.° ao 20.° dias. Hack2 0 (1955) obteve, àtemperatura de 18,5-28,7°C, a eclosão dosovos em 27-46 dias, lembrando, porém, apossibilidade de haver comportamento dife-rente para essa espécie, quando submetidosàs temperaturas de seus ambientes naturais.
Lucena2 3 (1960) para o T. brasiliensisobservou a eclosão dos ovos com 17-19dias, à temperatura de 25,4-28,9°C.
Zeledon e col.35 (1970) observaram queos ovos de T. dimidiata evoluíram maisrapidamente para a eclosão quando subme-tidos à temperatura de 26,5°C (21,2 —22,9 dias) do que à temperatura variandode 21,8-24,4°C (29,0-35,9 dias).
As estimativas para a variável TI parao total de insetos, machos e fêmeas, àstemperaturas de 25°C e 30°C acham-se nasTabelas 4 e 5. Observa-se uma regula-ridade apreciável em todas as estimativas,dentro de cada temperatura, tanto entre asninfas em geral, como também em ambosos sexos.
Informações referentes à ação da tempe-ratura sobre a duração da fase embrionáriapodem ser úteis em laboratórios onde sãomantidas colônias de triatomíneos para finsdiversos e especialmente para xenodiagnós-tico. Como se pode observar, o aumentoda temperatura em 5°C reduziu o tempo deduração da fase evolutiva em cerca de 50%,tanto para o T. sordida como para o T.infestans. O fato se repete em outros triato-míneos em graus variáveis, mas sempre como resultado no mesmo sentido.
Convém ainda apontar outro aspecto deinteresse, qual seja o da aplicação dessesachados de laboratórios nos programas decombate aos vetores da doença de Chagas.Parece pertinente a proposição de que adesinsetização de casas e anexos deva serfei ta , preferentemente nos meses maisquentes do ano, propiciando, dessa forma,ação mais favorável dos inseticidas queatingiram maior número de ninfas nosprimeiros estádios, recém-eclodidos.
Duração do ciclo evolutivo
Da mesma forma que ocorreu em relaçãoà fase embrionária, a temperatura teve efeitomarcante, determinando diferenças signifi-cantes entre as durações médias de todosos estádios de evolução. Não ocorreramdiferenças significantes entre os grupos deestudo. O efeito do sexo determinou dife-renças significantes nos 4.° e 5.° estádios.Ocorreu interação entre temperatura e sexono 4.° estádio (Tabela 4).
Nos cinco estádios de evolução observou--se o mesmo efeito da temperatura, em cadasexo separadamente. O teste de Scheffé(Tabela 5) permitiu detectar diferençassignificantes das médias observadas entremachos e fêmeas, nas duas temperaturasestudadas, no 5.° estádio. Dado que esteteste permite a utilização de seu resultadomesmo para constrastes sugeridos após acoleta de dados, depreende-se que os machostêm duração média significantemente maiordo que as fêmeas no 5.° estádio, em ambasas temperaturas.
No 4.° estádio, tendo ocorrido interaçãotemperatura-sexo, os contrastes foram jul-gados por testes de médias. No tocante àtemperatura, confirmou-se o observado naanálise de variância, que à temperatura de25°C as médias são maiores do que a 30°C,tanto para os machos como para as fêmeas.No entanto, não se confirmou a hipótesede que as fêmeas teriam duração médiamaior que a dos machos. A 30°C não ocor-reram diferenças significantes, porém, a25°C o resultado do teste foi bastante indi-cativo de que os machos têm duração maiorque as fêmeas.
Nas Tabelas 6 e 7 observa-se que a 30°C,tanto para as ninfas em geral como paraas ninfas machos e fêmeas, a duração médiade evolução cresce lenta e progressivamentedo 1.° ao 4.° estádio, para atingir valoresbem mais elevados no 5.°. O acréscimo per-centual da duração média em cada estádio,com relação ao estádio imediatamente ante-rior, foi de 14,3%, 1,0%, 7,1%, 293,4% paraas ninfas em geral; de 17,8%, 1,5%, 6,3%,
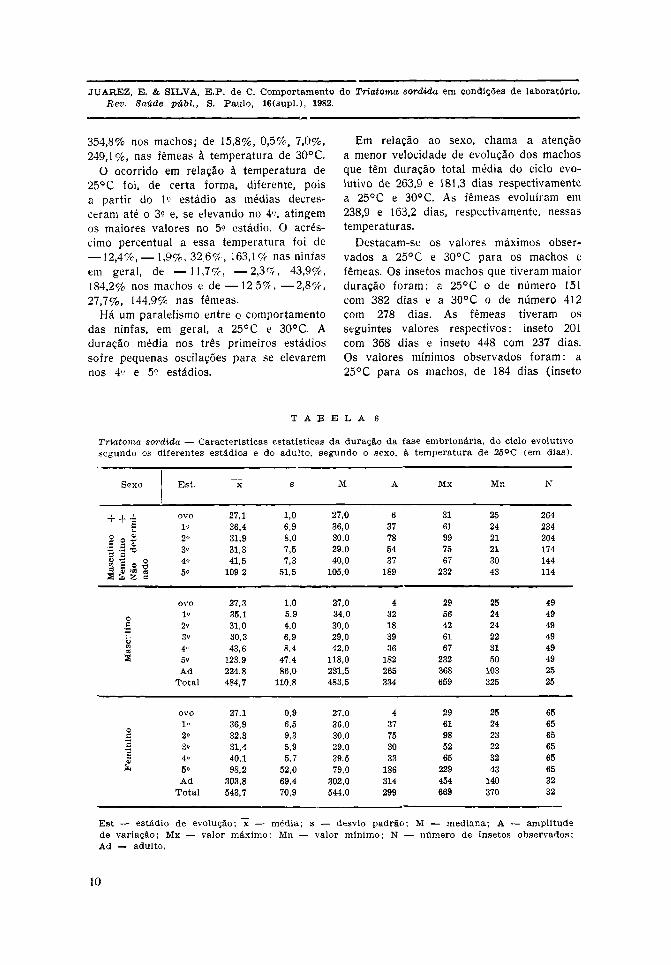
354,8% nos machos; de 15,8%, 0,5%, 7,0%,249,1%, nas fêmeas à temperatura de 30°C.
O ocorrido em relação à temperatura de25°C foi, de certa forma, diferente, poisa partir do 1.° estádio as médias decres-ceram até o 3.° e, se elevando no 4.°, atingemos maiores valores no 5.° estádio. O acrés-cimo percentual a essa temperatura foi de
- 12,4%, — 1,9%, 32,6%, 163,1% nas ninfasem geral, de — 1 1 , 7 % , —2,3%, 43,9%,184,2% nos machos e de — 125%, —2,8%,27,7%, 144,9% nas fêmeas.
Há um paralelismo entre o comportamentodas ninfas, em geral, a 25°C e 30°C. Aduração média nos três primeiros estádiossofre pequenas oscilações para se elevaremnos 4.° e 5.° estádios.
Em relação ao sexo, chama a atençãoa menor velocidade de evolução dos machosque têm duração total média do ciclo evo-lutivo de 263,9 e 181,3 dias respectivamentea 25°C e 30°C. As fêmeas evoluíram em238,9 e 163,2 dias, respectivamente, nessastemperaturas.
Destacam-se os valores máximos obser-vados a 25°C e 30°C para os machos efêmeas. Os insetos machos que tiveram maiorduração foram: a 25°C o de número 151com 382 dias e a 30°C o de número 412com 278 dias. As fêmeas tiveram osseguintes valores respectivos: inseto 201com 368 dias e inseto 448 com 237 dias.Os valores mínimos observados foram: a25°C para os machos, de 184 dias (inseto
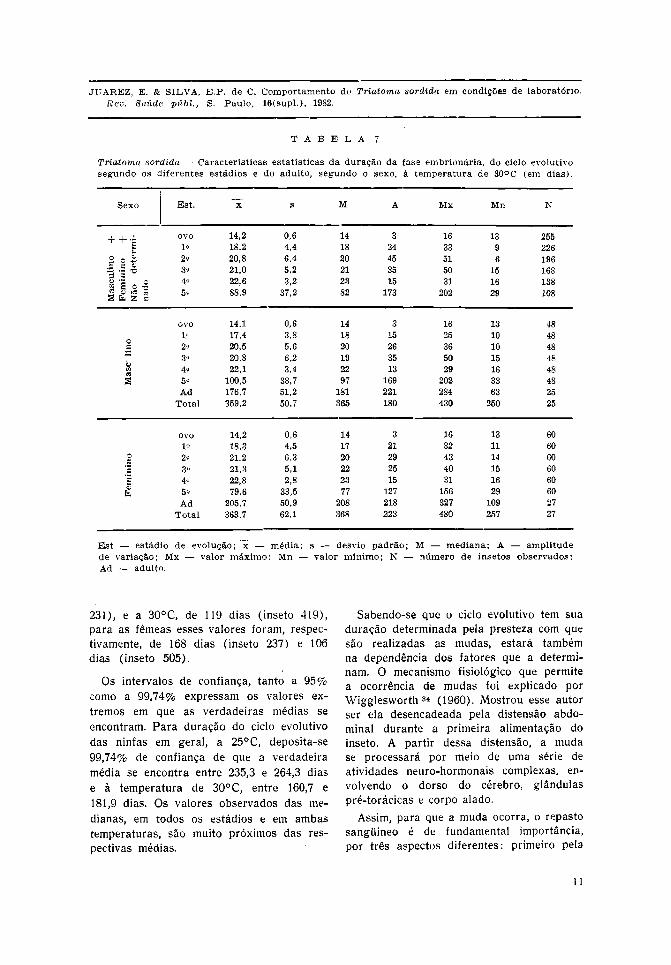
231), e a 30°C, de 119 dias (inseto 419),para as fêmeas esses valores foram, respec-tivamente, de 168 dias (inseto 237) e 106dias (inseto 505).
Os intervalos de confiança, tanto a 95%como a 99,74% expressam os valores ex-tremos em que as verdadeiras médias seencontram. Para duração do ciclo evolutivodas ninfas em geral, a 25°C, deposita-se99,74% de confiança de que a verdadeiramédia se encontra entre 235,3 e 264,3 diase à temperatura de 30°C, entre 160,7 e181,9 dias. Os valores observados das me-dianas, em todos os estádios e em ambastemperaturas, são muito próximos das res-pectivas médias.
Sabendo-se que o ciclo evolutivo tem suaduração determinada pela presteza com quesão realizadas as mudas, estará tambémna dependência dos fatores que a determi-nam. O mecanismo fisiológico que permitea ocorrência de mudas foi explicado porWigglesworth 34 (1960). Mostrou esse autorser ela desencadeada pela distensão abdo-minal durante a primeira alimentação doinseto. A partir dessa distensão, a mudase processará por meio de uma série deatividades neuro-hormonais complexas, en-volvendo o dorso do cérebro, glândulaspré-torácicas e corpo alado.
Assim, para que a muda ocorra, o repastosangüíneo é de fundamental importância,por três aspectos diferentes: primeiro pela

distensão abdominal; segundo, pela neces-sidade absoluta que as ninfas têm da hema-tina presente no sangue, sem o que a mudanão se processará, conforme o demonstradopor Lwoff e Nicolle 24 (1945); terceiro pelanecessidade do sangue como nutriente quedará as condições para o desenvolvimentofísico do inseto.
Nesta pesquisa, os insetos tiveram opor-tunidade de se alimentar semanalmente,sempre nas mesmas condições e por umperíodo de tempo julgado satisfatório paraque ocorresse a repleção abdominal. Épossível que, após a eclosão dos ovos oua realização das mudas, as ninfas aindanão estivessem em condições de se alimentarno momento em que lhes era oferecido orepasto sangüíneo.
Perlowagora-Szumlevics27 (1953) enfa-tizou a necessidade da realização do repastono 4.° dia, para as ninfas de primeiro estádiodo T. infestans poderem efetivar a muda eterem melhores perspectivas de chegar àfase adulta. Buxton 3 (1930), afirmou quea duração da evolução seria encurtada sefosse oferecido o repasto tão logo as ninfasde R. prolixus pudessem se alimentar. Cas-tanho 4 (1972) observou que as ninfas deP. megistus conseguiram se alimentar no4.° dia após a muda, porém a alimentaçãofoi maior depois desse período, não tendoverificado mudas no 5.° estádio, com apenasum repasto. Goodchild19 (1955) verificouque o T. infestans tem limitações anatô-micas que lhe conferem pequena capacidadede alimentação, necessitando de mais deum repasto para efetuar a muda, enquantoo R. prolixus o faz freqüentemente comapenas um. Este fato também foi observadopor Danilov 6 (1968) em T. infestans, no-tando que somente 1,1% das ninfas de 5.°estádio conseguem mudar com um repasto,à temperatura de 26-27°C. Ainda para estaespécie, Juarez2 2 (1970) observou a ocor-rência de mudas em 31,4% das ninfas de 5.°estádio alimentadas em camundongos emantidas a 30°C com apenas 1 repasto;as ninfas alimentadas em galinhas, a essatemperatura, e as alimentadas nas duasfontes de sangue a 25°C, não efetivaram a
muda com apenas um repasto, como o fizeramaquelas. Ferreira1 0 (1972) verificou que a30°C, 30,43% das ninfas de 5.° estádio,fêmeas, de T. brasiliensis, mudaram com umso repasto, enquanto que as ninfas machossó o fizeram em 8,1%.
Com essas considerações, os resultadosobtidos para duração do ciclo evolutivo nãorepresentará necessariamente o menor valorpossível, em condições naturais.
Souza e col.31 (1978) observaram a evo-lução do T. sordida na fase de ninfa , emlaboratório a 25-28°C de temperatura e60-80% de umidade relativa do ar. Nãofizeram a medida da duração de cada umdos estádios ninfais isoladamente, porém ototal do ciclo evolutivo teve médias de 174,5e 170,6 dias para os machos e fêmeas,respectivamente.
No presente trabalho, os resultados forammuito próximos do desses autores, a 30°C,com valores respectivos para a média dosmachos e fêmeas de 181,3 e 163,2 dias.Porém à temperatura de 25°C, os resul-tados foram apreciavelmente diferentes, comvalores respectivos de 263,9 e 238,9 dias.Essa diferença não seria esperada, uma vezque a temperatura em que os autores fizeramsuas observações está mais próxima de25°C do que de 30°C. Poder-se-ia fazer ahipótese de que, pela metodologia empre-gada, criação dos insetos em conjunto atéo 4.° estádio, tivesse ocorrido elevação detemperatura e conseqüente aceleração dociclo evolutivo. Essa possibilidade foi com-provada por Bodenstein2 (1953) e tambémverificada por Juarez 2 1 (1970) em T.arthurneivai. Este autor observou que osinsetos criados em conjunto evoluem maisrapidamente do que os isolados, notandotambém que os machos evoluem mais rapida-mente do que as fêmeas.
Castanho4 ,(1972), trabalhando com P.megistus a 30°C, não encontrou diferençaentre o tempo de evolução dos machos edas fêmeas, que tiveram respectivamente139,0 e 139,9 dias de duração do ciclo evolu-tivo. Zeledon e col.35 (1970) verificaramque a evolução das ninfas de 5.° estádio do

T. dimidiata se faz mais rapidamente nasfêmeas do que nos machos, se bem queas diferenças sejam muito pequenas. Perlo-wagora-Szumlevics28 (1969) verificou pre-dominância de adultos machos em tempe-raturas ambiente altas, e de fêmeas nastemperaturas mais baixas, sugerindo umadiferença de comportamento quanto ao sexopor efeito da temperatura.
Em que pese não haver análise estatísticapara grande parte dos trabalhos, pode-seperceber que deve haver uma variabilidademuito grande no comportamento dos tria-tomíneos.
Esses dados levam à suposição de quedevam existir muitos fatores atuando inte-rativamente e que não podem ser conhecidose controlados em experimentos de labora-tório, como estes. O fato de se procurarcontrolar algumas variáveis tais como tem-peratura, umidade relativa do ar, fonte dealimento, freqüência de oferecimento derepastos, tempo de sucção, não parecesuficiente para que seja feita a inferênciados resultados para a população de triato-míneos no seu ambiente natural. Há outrosfatores, alguns dos quais já conhecidos,referentes à fisiologia dos insetos, que podemalterar total ou parcialmente resultados detrabalhos, dif icultando sobremaneira a suainterpretação.
Decorre desse fato a necessidade de sealiar os resultados obtidos em laboratóriocom os de observação de campo para omelhor aproveitamento das informações emtermos da busca de interpretações.
Duração da fase adulta
Em relação às formas adultas, os efeitosda temperatura e do sexo foram bastanteexpressivos, o que pode ser observado naTabela 4.
Os valores médios da duração da faseadulta foram maiores a 25°C em ambosos sexos. Conquanto na análise de variânciao efeito do sexo tenha sido muito expres-sivo, os contrastes obtidos com o teste deScheffé revelaram que não houve compor-
tamento análogo em ambas as tempera-turas. A 25°C houve diferença significanteentre as médias dos sexos, as fêmeas comduração maior do que os machos na faseadulta. A 30°C não ocorreu diferença signi-ficante entre as médias dos sexos, as fêmeasnão tendo média superior à dos machos.Houve pois, um efeito desigual da tempera-tura em relação ao sexo, lembrando a pos-sibilidade de interação dos efeitos da tem-peratura e sexo. Entretanto, a análise devariância não detectou a existência de inte-ração, porém o valor de "F" foi muitopróximo de ser significante ao nível de 5%(F = 3,87 P = 0,0517).
É interessante ressaltar que a velocidadeda fase de ninfas não se mostrou diferenteem relação aos sexos nos três primeirosestádios. No 4.° estádio, em que ocorreuinteração, a indicação de que os machosevoluem mais lentamente do que as fêmeasficou evidente. No 5.° estádio, houve com-provação estatística de que a duração dosmachos é maior. Na fase adulta, a duraçãodas fêmeas foi maior, isto é, sua longe-vidade é maior do que a dos machos.
Este fato também foi observado porSouza e col.31 (1978) que verificaramperíodos praticamente iguais para a duraçãodo ciclo evolutivo dos machos e fêmeas doT. sordida. Estes autores observaram quena fase adulta as fêmeas têm longevidademédia bastante maior que a dos machos,com valores respectivos de 503,0 e 284,0dias.
Nas Tabelas 6 e 7 verifica-se que aduração média dos adultos machos e fêmeasforam, respectivamente, às temperaturas de25°C e 30°C de 224,8 — 176,7 dias e 303,8— 205,7 dias. Como se vê, estes resultadosficaram muito aquém dos obtidos poraqueles autores 31. Também com relação aosvalores máximo e mínimo observados, osresultados foram muito diferentes. Enquantoaqueles autores31 obtiveram para o má-ximo de longevidade dos machos e fêmeas,respectivamente 520 e 782 dias, neste tra-balho os valores respectivos a 25°C foramde apenas 368 e 454 dias. Haveria a pos-

sibilidade de explicar essas diferenças pelamaior adaptabilidade de uma das colôniasàs condições de laboratório com menoragressão aos adultos, o que poderia ocorrerpela menor resistência que apresentam.
Ambos os trabalhos, no entanto, mostramque as fêmeas têm maior longevidade do queos machos, mesmo considerados os resultadosnuméricos bastante diversos. Haveria poispor um determinismo biológico de perpetua-ção da espécie, um comportamento favorávelà fecundação das fêmeas. Os machos, depequena importância na perpetuação daespécie, uma vez que teriam somente afunção de fecundação, poderiam ter vidamenos longa. Por sua vez, as fêmeas quetêm o encargo de realizar a postura deovos férteis e, ainda mais, de promovera dispersão da espécie deslocando-se paraoutros ecótopos, deveria ter vida mais longa.
Quanto à velocidade de evolução da fasede ninfas, na mesma linha de pensamento,as fêmeas deveriam atingir a fase adultamais precocemente do que os machos para,tão logo estivessem em condições, dar pros-seguimento ao ciclo vital com o objetivo demanter a espécie. Assim pensando, osmachos poderiam ser mais lentos na evo-lução. Dessa forma, torna-se dif íc i l encontrarexplicação que satisfaça inteiramente, umavez que o fenômeno terá a interpretaçãoque os escassos conhecimentos da biologiadesses insetos pode oferecer.
Outro aspecto de difícil explicação, e quetalvez ajudasse no entendimento do fatoacima exposto, é o fenômeno da interaçãoentre temperatura e sexo. Nos dois tra-balhos em que se observou o comporta-mento do T. sordida, há diferenças razoáveisentre os resultados, no que diz respeito aduração do ciclo evolutivo. Em um, pratica-mente não se observou diferença entre asmédias do ciclo evolutivo dos machos edas fêmeas. No outro, nas duas tempera-turas em estudo, ficou patente a diferençadas médias de duração do ciclo evolutivopara machos e fêmeas, lembrando-se queocorreu interação entre os efeitos do sexoe temperatura no 4.° estádio.
Na fase adulta não ocorreu diferençasignificante entre as médias de longevidadedos machos e fêmeas mantidos a 30°C.Já a 25°C, ocorreu diferença entre asmédias respectivas, a das fêmeas maior doque a dos machos. É evidente, então, ofenômeno da interação, não detectado pelaestatística, porém facilmente verificado pelaobservação dos números.
Os fatores que regulam a biologia dosinsetos no ambiente natural certamenteinteragem, determinando as melhores pos-sibilidades para a permanência da espécie.Devem pois, ocorrer interações entre essesfatores, no sentido de que a presença deinsetos em todas as fases de evolução sejauma constante, mesmo que, em certasépocas, provavelmente as mais favoráveis,determinadas formas predominem sobre asoutras.
Se as condições de trabalho em labora-tório pudessem ao menos permitir a infe-rência para o ambiente natural, o T. sordidadeveria ter, em algumas épocas do ano,maior quantidade de exemplares adultos doque em outras em que esse número seriareduzido. Essa hipótese se baseia nos dadosobtidos neste trabalho em que a duraçãodas fêmeas é de 7 a 10 meses e a dosmachos de 6 a 7 meses. Considerando,também, os valores extremos observados,seria possível a presença de adultos du-rante todo o período de um ano, maisnumerosos em apenas uma época.
Sangue ingerido
A constituição dos grupos não teve osucesso esperado, pois as médias dos gruposna variável S4 não foram homogêneas entresi.
O efeito da temperatura em relação aosangue ingerido pelos insetos foi bastanteintenso do 1.° ao 4.° estádios. Nesses está-dios, as quantidades médias de sangue inge-rido a 25°C foram significantemente maioresdo que a 30°C. O efeito do sexo nos doisprimeiros estádios não foi evidente. Nosdemais, o efeito do sexo se fez presente.(Tabela 8).

O teste de Scheffé, na Tabela 8, confirmouo efeito da temperatura em todos os está-dios, isto é, ocorreram diferenças signifi-cantes em todos os estádios exceto no 5.°.O mesmo teste confirmou as diferençasobservadas entre as médias de sangueingerido nos 4.° e 5.° estádios, mostrandoque as fêmeas ingeriram maior quantidadede sangue do que os machos em ambasas temperaturas. No terceiro estádio, entre-tanto, a diferença observada na análise devariância não se confirmou.
Houve, pois, na análise de variância(Tabela 8) uma somação numérica dasdiferenças entre as médias dos sexos emambas as temperaturas, que resultou nasignificância observada.
Nas Tabelas 9 e 10, nota-se que amédia de sangue ingerido cresce do 1.° ao5.° estádios, nas duas temperaturas, e emambos os sexos.
Em relação às ninfas em geral, em todosos estádios o consumo de sangue foi maiora 25°C, exceto no 5.° estádio.
À temperatura de 30°C, as fêmeas, emtodos os estádios, consumiram maior quan-tidade de sangue do que os machos. A 25°Chouve o mesmo comportamento, à exceção
do 1.° estádio, em que os machos ingeriramigual quantidade.
Como se pode constatar, há uma regula-ridade apreciável no que concerne ao sangueingerido pelo T. sordida nas duas tempe-raturas e em ambos os sexos.
Os valores médios, mínimos e máximosde sangue ingerido que propiciou a ocor-rência da muda no 5.° estádio, foram paraos machos a 25°C e 30°C de, respectiva-mente, 484,8, 215,1, 781,6 e 503,9, 290,4,730,6 mg; para as fêmeas os valores cor-respondentes foram de 565,8, 353,1, 956,8e 559,0, 311,6, 833,4 mg.
A razão entre sangue ingerido eduração num determinado estádio, dá amedida da necessidade média diária, paraque o inseto evolua entre duas mudas con-secutivas. A necessidade média diária dasninfas em geral, a 25°C foi, do 1.° ao 5.°estádio, de 0,1, 0,4, 1,2, 3,1, 4,9, e a 30°Cde 0,2, 0,5, 1,5, 4,4, 4,9. Nota-se uma maiornecessidade de sangue à temperatura de30° C, nos diferentes estádios, exceto no 5.°.Neste, não houve diferença significante dasmédias por efeito da temperatura, nem foiobservada maior duração das fêmeas, con-forme já apresentado. No entanto, o efeitodo sexo sobre as médias de sangue ingerido
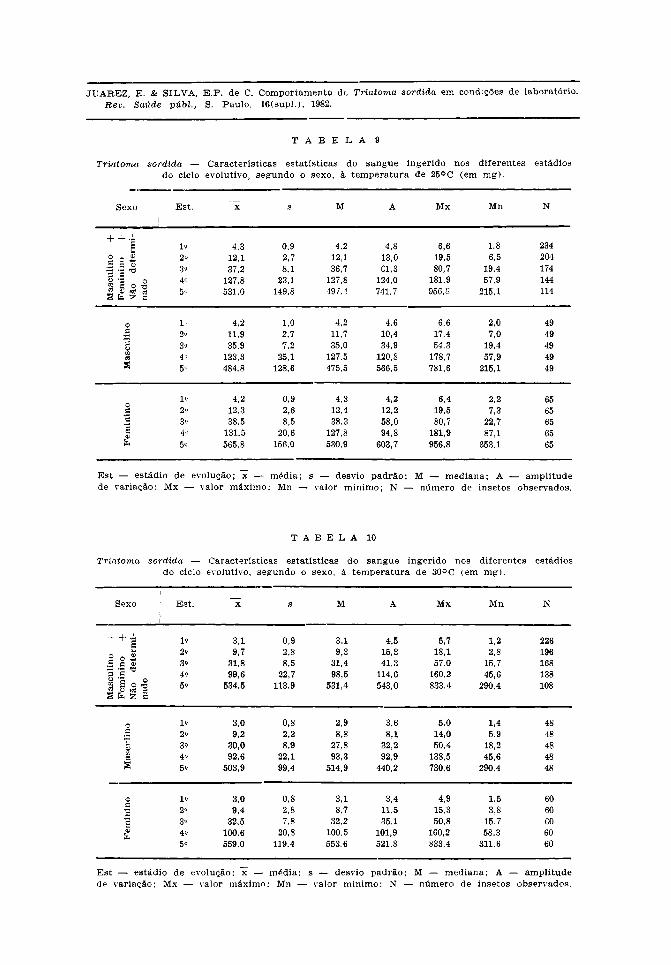

e médias de duração se fez sentir a partirdo 4.° estádio o que refletiu no consumomédio diário. Para esses estádios, o con-sumo médio diário a 25°C, foi de 2,8 e 3,9para machos e de 3,3 e 5,8 para fêmeas;a 30°C os valores correspondentes foramde 4,2 e 5,0 para os machos e de 4,4 e7,0 para as fêmeas. Há uma predominânciadas necessidades médias diárias das fêmeascomo também para ambos os sexos a 30° C.
Juarez2 2 (1970), observou no T. infestans,uma quantidade média de sangue ingeridomenor à temperatura de 30°C para as ninfasde 1.°, 4.° e 5.° estádios, ocorrendo o inversonos demais estádios. É interessante notarque os resultados observados nesta espéciee no T. sordida foram diferentes, apesar deter sido utilizada a mesma técnica o quemostra as diferenças entre as espécies detriatomíneos do mesmo gênero.
Diferentes autores têm estudado diversasespécies de triatomíneos, observando aquantidade de sangue ingerido, segundo asvárias fontes de alimento e em temperaturasfixas ou ambientais. Com relação ao T.sordida não foi encontrada bibliografiacorrespondente.
Zeledon e col.35 (1970), em T. dimidiata,observou que nos vários estádios ninfais,a quantidade de sangue ingerido foi maiornos estádios 1.°, 2.° e 4.° a 26,5°C enquantonos 3.° e 5.° foi maior à temperatura de21,8-23,9°C.
Castanho4 (1972), em P. megistus man-tidos a 30°C e alimentados em galinha,observou que as ninfas de 4.° e 5.° estádiosingerem maior quantidade de sangue doque os machos, fato também observado como T. sordida.
Os fatos observados com essas váriasespécies de triatomíneos, em relação aosangue ingerido, em especial o T. sordida,sugerem que ainda existem muitas inda-gações a serem respondidas no que dizrespeito ao seu metabolismo. Apenas existemconjecturas sobre o comportamento alimentaraos triatomíneos, salvo o fato de seu

ecletismo alimentar bem acentuado. Sobresua capacidade de ingerir em um só repasto,a quantidade de sangue necessária paraefetuar a muda, no ambiente natural ouse existem alimentos que facilitam aevolução, existem conjecturas baseadasapenas em trabalhos de laboratório, sujeitosa sérias restrições pela sua condição total-mente artificial.
Número de repastos
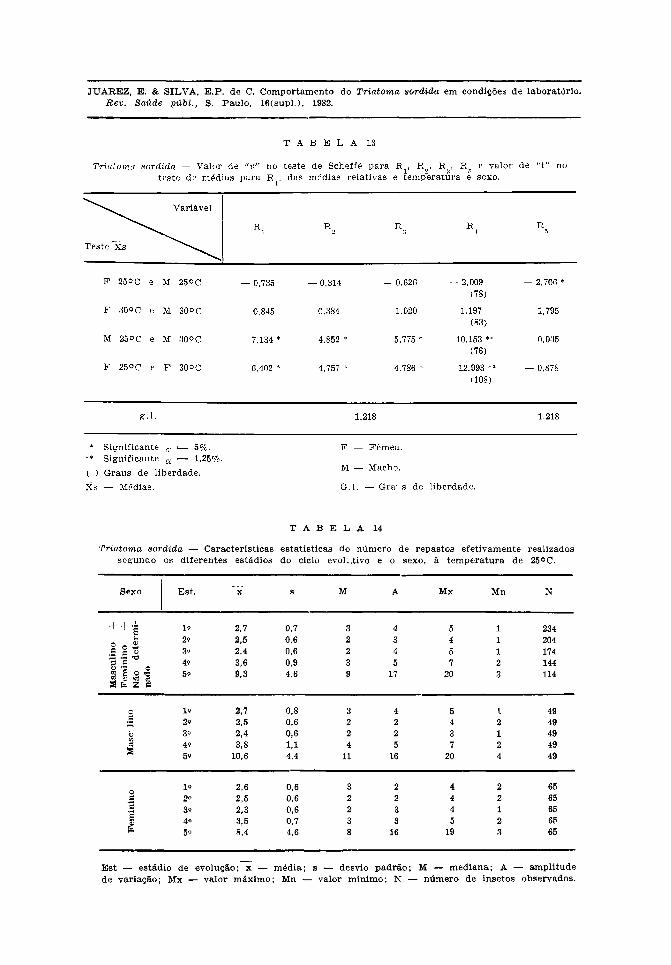
Observou-se que ocorreram diferençassignificantes na constituição dos gruposde estudo para a variável R1, e que o efeitoda temperatura determinou diferenças signi-cantes entre as médias do número derepastos a 30°C e 25°C sendo esta a maiornos estádios 1.° ao 4.°. No 5.° estádio nãoocorreu diferença por efeito da tempera-tura.
Na Tabela 12, verifica-se que o efeitoda temperatura teve o mesmo comporta-mento descrito acima. Em relação ao sexo,ocorreu diferença significante apenas no 5.°estádio. Houve interação temperatura-sexono 4.° estádio.
Na Tabela 13, nota-se que o teste deScheffé confirmou as diferenças signi-ficantes determinadas por efeito da tempe-ratura nos três primeiros estádios, bem como
a não ocorrência desse efeito no 5.° estádio.O teste de médias também mostrou as dife-renças significantes ocorridas no 4.° estádio,por efeito da temperatura. Do 1.° ao 4.°estádio, a ação da temperatura determinounúmero médio de repastos maiores para osmachos e fêmeas mantidos a 25°C.
Em relação ao efeito determinado pelosexo, verifica-se neste mesmo quadro queno 5.° estádio os machos tiveram médiamaior que as fêmeas a 25°C, e a 30°C nãohouve diferença entre eles. No quarto está-dio, as fêmeas não tiveram número médiode repastos maior do que os machos.
Nas Tabelas 14 e 15 observa-se que oT. sordida tem os menores valores denúmero médio de repastos no 3.° estádiopara as ninfas em geral, macho e fêmea,em ambas as temperaturas. No 5.° estádioencontra-se o maior número médio de re-pastos nas duas temperaturas e tambémmaior nos machos. Os demais estádios com-portam-se de forma muito semelhante.
A razão entre sangue ingerido e númerode repastos representa a média de sangueingerido para cada repasto efetivamenterealizado. Essa razão aumenta do 1.° ao5.° estádios para ambos os sexos e tempera-turas.
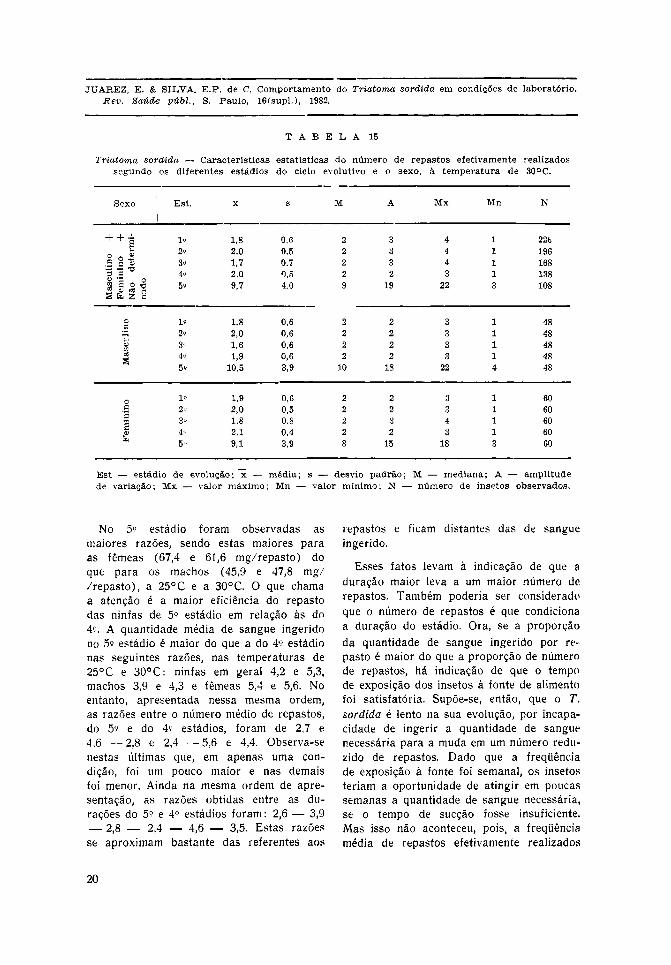
No 5.° estádio foram observadas asmaiores razões, sendo estas maiores paraas fêmeas (67,4 e 61,6 mg/repasto) doque para os machos (45,9 e 47,8 mg//repasto), a 25°C e a 30°C. O que chamaa atenção é a maior eficiência do repastodas ninfas de 5.° estádio em relação às do4.°. A quantidade média de sangue ingeridono 5.° estádio é maior do que a do 4.° estádionas seguintes razões, nas temperaturas de25°C e 30°C: ninfas em geral 4,2 e 5,3,machos 3,9 e 4,3 e fêmeas 5,4 e 5,6. Noentanto, apresentada nessa mesma ordem,as razões entre o número médio de repastos,do 5.° e do 4.° estádios, foram de 2,7 e4,6 —2,8 e 2,4 —5,6 e 4,4. Observa-senestas últimas que, em apenas uma con-dição, foi um pouco maior e nas demaisfoi menor. Ainda na mesma ordem de apre-sentação, as razões obtidas entre as du-rações do 5.° e 4.° estádios foram: 2,6 — 3,9
— 2,8 — 2,4 — 4,6 — 3,5. Estas razõesse aproximam bastante das referentes aos
repastos e ficam distantes das de sangueingerido.
Esses fatos levam à indicação de que aduração maior leva a um maior número derepastos. Também poderia ser consideradoque o número de repastos é que condicionaa duração do estádio. Ora, se a proporçãoda quantidade de sangue ingerido por re-pasto é maior do que a proporção de númerode repastos, há indicação de que o tempode exposição dos insetos à fonte de alimentofoi satisfatória. Supõe-se, então, que o T.sordida é lento na sua evolução, por incapa-cidade de ingerir a quantidade de sanguenecessária para a muda em um número redu-zido de repastos. Dado que a freqüênciade exposição à fonte foi semanal, os insetosteriam a oportunidade de atingir em poucassemanas a quantidade de sangue necessária,se o tempo de sucção fosse insuficiente.Mas isso não aconteceu, pois, a freqüênciamédia de repastos efetivamente realizados

no 5.° estádio foi, na mesma ordem, de11,2 e 9,5 — 11,7 e 11,7 — 9,5 e 8,8 dias.Como se nota, esses insetos não aproveitamtodas as oportunidades que lhe são ofere-cidas para se alimentar, mesmo se fôr levadaem conta a incapacidade de sugar logo apósa muda. Por outro lado, a observação indi-vidual mostrou que os insetos permanecemsem realizar o repasto muitas semanas con-secutivas.
Parece, pois, que a hipótese de que afreqüência e a duração do repasto inter-ferem na duração não tem muito suporte.Prefere-se, pois, a hipótese de que a duraçãodo inseto é que determina o número derepastos, pelas sucessivas necessidades doseu metabolismo.
Outro ponto a considerar é o valormáximo, mínimo e amplitude de variaçãodo número de repastos observado. A 25°Cas fêmeas tiveram, respectivamente; 19 —3 — 16 e os machos 20 — 4 — 16; e a30°C as fêmeas tiveram 18 — 3 — 15 eos machos 22 — 4 — 18, para o númerode repastos no 5.° estádio.
Comparando esses resultados com osobtidos no 5.° estádio em T. infestans, emidênticas condições de trabalho (Juarez22,1970), observa-se que existe uma grandediferença devido ao comportamento dasduas espécies de triatomíneos. Nas ninfasdessa espécie, os respectivos valores, parafins de comparação, foram: a 25°C, 7 — 2
— 5 e a 30°C, 3 — 1 — 2.
Outro aspecto interessante a ser apre-sentado é a quase total incapacidade mos-trada pelo T. soridda de realizar a muda comapenas um repasto, a 25°C, mesmo entreas ninfas mais jovens, como ocorre emoutras espécies. Enquanto a 30°C esse fatonão foi incomum, nos quatro estádiosiniciais, a 25°C, somente ocorreu com umaninfa no 1.° estádio, 2 ninfas no 2.°, 4 ninfasno 3.° e 17 no 4.°. Essa é outra diferençaencontrada com o T. infestans, no trabalhocitado 22, em que a freqüência com que osinsetos realizaram a muda com apenas umrepasto foi muito alta, inclusive no 5.°estádio.
Não se encontrou, porém, qualquer tra-balho com T. sordida que fizesse mençãode resultados acerca do número de repastosna fase de ninfas.
O número médio total de repastos efetiva-mente realizados no decorrer do ciclo evo-lutivo para as ninfas em geral, a 25°C, foide 20,5, a mediana do número total derepastos, 20 com máximo de 31 e minimode 11. A 30° C, para o total de repastos,a média foi 17,3, a mediana 17, o máximo29 e o mínimo 10.
Essas estimativas são muito importantesem epidemiologia, pois indicam a possibi-lidade de haver transmissão do T. cruzi.A simples comparação desses resultados, comos correspondentes do T. infestans, obtidosem idênticas condições de laboratório, porJua rez 2 2 (1970), indicam a possível capa-cidade de transmissão do T. sordida. Nestetrabalho, a 25°C, a média total de repastosfoi 9,8 com máximo de 15 e mínimo de7, e a 30°C a média foi 6,2, o máximo 9e o mínimo 5.
Outro aspecto interessante se refere aonúmero de repastos com relação ao sexo.As ninfas fêmeas de 5.° estádio, que emambas as temperaturas ingerem maiorquantidade de sangue do que os machos,também realizam menor número de repastos.O número total médio de repastos decresceda temperatura de 25°C, para a de 30°Ce, em ambas, do macho para a fêmea comos seguintes valores respectivos: 22,0, 19,3,17,8 e 16,9.
As possibilidades de transmissão decres-cem, pois, a medida que a temperatura seeleva e são menores para as fêmeas. Entreas ninfas, as que mostram possuir asmaiores possibilidades de transmissão sãoas do 5.° estádio, qne realizam grandespercentuais dos repastos no ciclo evolutivo.Essas proporções, na ordem apresentadaacima, são 48,0%, 43,6%, 54,7% e 54,0%.
Considerada a longevidade dos adultos,e a impressão de que raramente deixam dese alimentar quando se lhes oferece a fontede sangue semanalmente, são esses, pos-
sivelmente, os que têm maiores probabi-lidades de agir como transmissores.
Peso inicial dos insetos
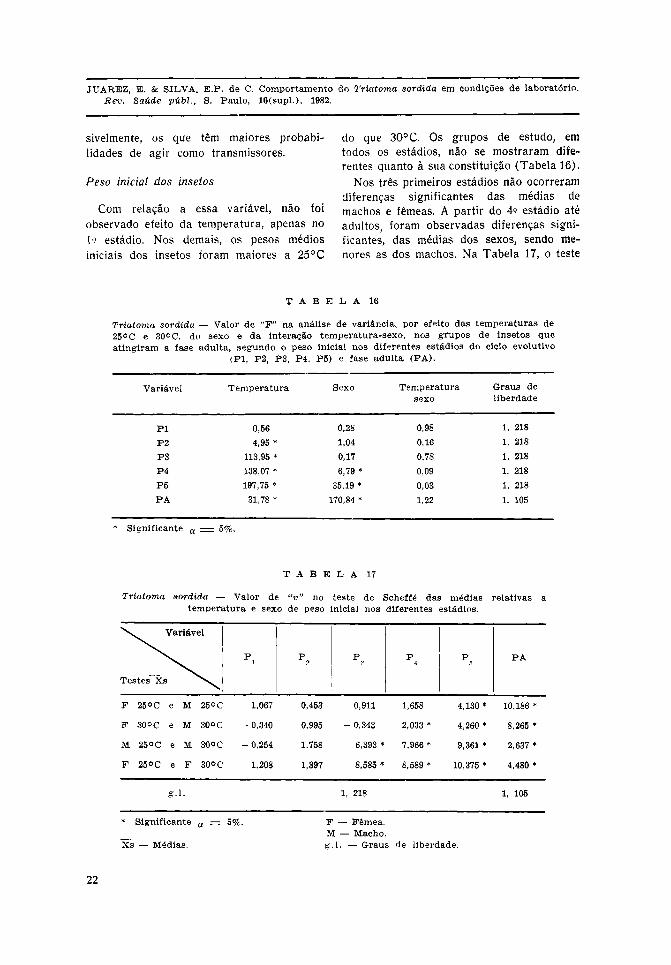
Com relação a essa variável, não foiobservado efeito da temperatura, apenas no1.° estádio. Nos demais, os pesos médiosiniciais dos insetos foram maiores a 25°C
do que 30°C. Os grupos de estudo, emtodos os estádios, não se mostraram dife-rentes quanto à sua constituição (Tabela 16).
Nos três primeiros estádios não ocorreramdiferenças significantes das médias demachos e fêmeas. A partir do 4.° estádio atéadultos, foram observadas diferenças signi-ficantes, das médias dos sexos, sendo me-nores as dos machos. Na Tabela 17, o teste

de Scheffé mostrou diferenças significantesdevidas à temperatura, somente a partir do3.° estádio.
O efeito do sexo foi evidente no 5.° estádioe nas formas adultas, as fêmeas tiverammédias de peso inicial maiores do que asdos machos. No 4.° estádio, somente foidetectada diferença entre as médias dossexos à temperatura de 30°C; a 25°C nãoocorreu diferença.
Nas Tabelas 18 e 19 nota-se que ospesos médios iniciais das ninfas crescemdo 1.° estádio até a fase adulta, de formasemelhante ao que ocorreu com as médiasde sangue ingerido.
O aproveitamento do sangue pelos insetospode ser medido pela razão entre o ganhode peso no intervalo de duas mudas conse-cutivas e o sangue ingerido nesse período.Dada a metodologia proposta para a exe-
cução deste trabalho, alguns erros poderiamser apontados como possíveis de falsearemos resultados, tais como erros de pesagem,que devem ser muito pequenos pela pre-cisão oferecida pela balança. O outro pos-sível defeito na apreciação do peso inicialdos insetos se refere ao período após amuda em que foi tomado. Como os insetoseram pesados somente no dia predetermi-nado, o registro dessa variável pode sereferir ao peso inicial no dia da eclosãoou da muda, ou até sete dias após.
No seu trabalho, Castanho4 (1972) esti-mou a perda de peso ocorrida após as mudasdo P. megistus, com o intuito de corrigiro peso inicial dos insetos medidos apósdiferentes períodos. Ferreira10 (1972) eCosta5 (1972) estimaram para as ninfas deT. brasiliensis no 5.° estádio a perda de

peso em cerca de 17% de seu peso inicial,após oito dias da muda.
No presente trabalho não se utilizou amesma técnica, nem outro tipo de tentativade correção por estimativa prévia, na espec-tativa de que as possíveis diferenças seriamdeterminadas por erros sistemáticos emtodos os estádios e, em decorrência, aanálise dos resultados não deveria ser sen-sivelmente prejudicada.
Outro fato importante é o já demonstra-do por Goodchild19 (1955) e Friend18
(1965), de que o inseto ao fazer a mudaainda é portador de apreciável quantidadede sangue no tubo digestivo. Assim, pres-supõe-se que os insetos ao realizar a mudaconservam um percentual constante emrelação ao sangue ingerido.
Feitas essas considerações, os resulta-dos, a seguir apresentados, sobre o apro-
veitamento alimentar do T. sordida, poderãoser melhor apreciados, acreditando-se quenão devam se colocar muito longe de valorescorrigidos.
De um modo geral, as razões decrescemdo 1.° estádio até o 5.°, quando observadosos machos e fêmeas a 25°C e os machosa 30°C. As fêmeas a 30°C, apresentaramrazões sensivelmente iguais nos 2.°, 3.° e 4.°estádios. Para melhor apreciação, os resul-tados obtidos estão apresentados em se-guida, na ordem de sucessão natural dosestádios: a 25°C, machos 0,35, 0,32, 0,29,0,24, 0,13 e fêmeas 0,36, 0,32, 0,29, 0,26,0,17; a 30°C machos 0,44, 0,29, 0,26, 0,25,0,13 e fêmeas 0,47, 0,27, 0,27, 0,27, 0,17Há pois, um aproveitamento maior dosangue ingerido nos estádios iniciais. O de-créscimo observado nos diferentes estádiosa 25°C é mais lento e gradual do que a30°C. Nesta temperatura, observam-se duas

quedas dignas de nota, do 1.° para o 2.°estádios e do 4.° para o 5.° estádios.
Os menores valores observados para essasrazões referem-se ao aproveitamento no 5.°estádio. Neste, em ordem decrescente, asrazões médias f o r a m : 0,174 (fêmeas a25°C), 0,166 (fêmeas a 30°C), 0,133 (machosa 25°C) e 0,129 (machos a 30°C). Há umaproveitamento maior das fêmeas e tambémà temperatura de 25°C.
A ninfa de 5.° estádio tem, pois, um menoraproveitamento de sangue em termos deaumento ponderal do que o observado nosoutros estádios. As profundas modificaçõesa que está sujeita essa ninfa , quais sejamas de originar as formas adultas, morfolo-gicamente diferente de todas as ninfas , éuma jus t i f ica t iva bastante ampla para ofato observado, uma vez que o metabolismodeve ser bem mais intenso. Os mecanismosbioquímicos e fisiológicos devem ser de talcomplexidade e demandam tal quantidadede energia que ocorre um desgaste maisacentuado do inseto no que concerne aosangue ingerido.
Ainda em relação ao peso do inseto, deve--se considerar também a razão de sangueingerido pelo peso inicial em cada estádio,indicando a necessidade de sangue relativaao peso.
Essas razões médias foram as seguintes,do 1.° ao 5.° estádios: machos a 25°C 6,6,5,6, 6,1, 7,5, 10,4; fêmeas a 25°C 6,4, 5,7,6,3, 7,7, 11,1; machos a 30°C 4,7, 4,7, 6,5,7,5, 14,4; fêmeas a 30°C 4,8, 4,6, 7,1, 7,6,14,0. As diferenças observadas nas razões,entre os sexos, são de pequena monta.Porém, o que chama a atenção são asdiferenças notadas no 5° estádio em que,a 30°C, as razões são sensivelmente maiores.Por outro lado, as razões nos dois primeirosestádios foram maiores a 25°C.
Os resultados obtidos para essas duasrazões, aproveitamento alimentar e neces-sidade de sangue em relação ao peso, decerta forma concordam com o verif icado poroutros autores. Castanho4 (1972) em P.megistus observou uma tendência ao decrés-cimo da razão média de aproveitamentoalimentar, do 1.° ao 5.° estádio, o inverso
acontecendo com a necessidade de sangueem relação ao peso. Viei ra 3 3 (1971), paraesta razão encontrou tendência semelhantepara o T. infestans. Friend1 8 (1965) eP ipp in 2 9 (1970) observaram que o R. pro-lixus tem tendência um pouco diversa, in-clusive caindo no 4.° e 4.°-5.° estádios, res-pectivamente.
É ainda digno de nota o observado comas razões médias de sangue ingerido emrelação ao peso do inseto no T. sordida,no 5.° estádio, com valores de 10,4 e 14,4que, comparadas às observadas no P.megistus de 7,62 e 8,43 para machos efêmeas a 30°C, mostram a maior voracidadeproporcional daquele triatomíneo.
Resistência ao Jejum
O efeito da temperatura sobre a resis-tência ao je jum, medida em dias após amuda, foi marcante em todos os estádiosda evolução do T. sordida. Os valores de"t", obtido nos testes de médias, mostraram-se significantes ao nível de 5%, mostrandoque a resistência média ao je jum em todosos estádios n infa is foi maior a 25°C do quea 30°C (Tabela 20).
Em relação ao comportamento dos adultosfrente ao jejum, a análise de variânciamostrou que os efeitos da temperatura esexo foram significantes ao nível de 5%,indicando que a resistência média ao je juma 25°C foi maior do que a 30°C (F = 158,6)e, também, maior nas fêmeas do que nosmachos (F = 12,1). Dado que a análisede variância detectou também a ocorrênciade interação entre temperatura e sexo(F = 4,1), significante ao nível de 5%, oque impede de se afirmar o efeito isoladode cada uma das condições, temperaturae sexo, foi aplicado o teste de médias(Tabela 20). Como resultado desse testeobtiveram-se valores de "t" significantesao nível de 1,25% indicativos de que oefeito da temperatura foi eficiente tanto paramachos como para as fêmeas, isto é, ambos
resistem ao j e jum em período médio maiora 25°C do que a 30°C. No entanto, osvalores de "t" indicaram que a 25C° as


fêmeas resistiram mais ao jejum do queos machos, enquanto a 30°C não ocorreudiferença signif icante entre os sexos.
Observadas as Tabelas 21 e 22, verifica--se que a redução da sobrevida, em jejum,das fêmeas , quando comparadas às médiasobtidas a 25°C e 30°C, foi de 36,3% (63,9e 40,7 dias) enquanto entre os machos foide 30,2% (54,9 e 38,3 dias).
Houve, pois, uma interação entre tempe-ratura e sexo resultando em aumento maiorda resistência ao j e jum do adulto fêmeaou aumento menor da resistência ao je jumdo macho quando, submetidos a 30° C, pas-sam à condição de 25°C. O inverso podeser considerado quando os insetos passamda temperatura de 25°C para a de 30C°.
A observação das Tabelas 1, 2 e 20mostra que o último peso (UP i ) no estádioimediatamente anterior àquele em que sevai observar o comportamento frente aoje jum tem resultados estatísticos semelhantesao da resistência ao j e jum (RJ i ) , com duasdiferenças importantes. No primeiro estádio,a 25°C o teste de médias revela que oúltimo peso médio do primeiro estádio a25°C não é maior do que a 30°C, enquantoque no 2.°, 3.° e 4.° estádios os últimos pesosmédios a 25°C são maiores.
O último peso médio no 5° estádio, me-dida feita para os insetos que foram subme-tidos ao je jum no estádio adulto, permitetambém a análise da inf luência do sexo.A análise de variância mostra efeitos datemperatura e do sexo significantes, nãorevelando a ocorrência de interação ao nívelde 5%. Aplicando o teste de Scheffé, osvalores de "v" indicam que o efeito do sexoé significante tanto à temperatura de 25°Ccomo à de 30° C, isto é, nas fêmeas, oúltimo peso médio nas duas temperaturasé maior do que a dos machos. O mesmoteste revela que entre as fêmeas o pesomédio é maior a 25°C do que a 30°C eque entre os machos não foi detectada dife-rença significante.
A análise da Tabela 20 permite fazerconsiderações a respeito da ação da tempe-ratura e sexo interagindo. A temperatura
de 25°C deve determinar menor velocidadeno consumo de reservas que o triatomíneopossui ao mudar do 5.° estádio para o estádioadulto. É o que sugere o observado comas médias de último peso no 5.° estádio ea resistência ao j e jum à temperatura de25°C, em que se nota uma concordâncianos resultados dos testes estatísticos apli-cados. Ambos mostram que os valores médiosdas fêmeas, nas duas variáveis, são signi-ficantemente maiores que as dos machos.Essa mesma comparação à temperatura de30°C mostra uma discordância nos resulta-dos dos testes, mostrando que o último pesomédio é maior para as fêmeas, enquantonão foi detectada diferença significanteentre as médias de resistência ao je jum dosdois sexos. Entre as fêmeas existe concor-dância nos resultados dos testes aplicados,isto é, o último peso médio e a resistênciamédia ao je jum são significantemente maioresa 25°C. O mesmo não ocorreu em relaçãoaos machos, em que houve discordância entreos resultados dos testes, isto é, a resistênciamédia ao j e jum a 25°C foi maior, a des-peito de não haver ocorrido diferença sig-nificante entre os últimos pesos médios a25°C e 30°C.
Os fatos apresentados permitem a hipó-tese de que a temperatura, ao se elevar, de-termina um efeito acelerador no consumo dereservas nutritivas do Triatoma sordida, maisacentuado nas fêmeas do que nos machos.
Ainda nas Tabelas 21 e 22, verifica-seque os valores obtidos para a mediana sãopraticamente iguais aos da média para aresistência ao je jum em ambas as tempera-turas. Ocorreram discordâncias entre essesvalores, superiores a dois dias, à tempera-tura de 25°C, nos estádios 2.° (2,8 dias),3.° (8,0 dias) e 5.° (2,2 dias), em todos, afavor da mediana; à temperatura de 30°Cnão ocorreram discordância maiores de doisdias. Esse fato permite supor que o efeitodo j e j u m está na dependência do consumodas reservas existentes no momento da muda,consumo esse que se faz segundo uma dis-tribuição normal.
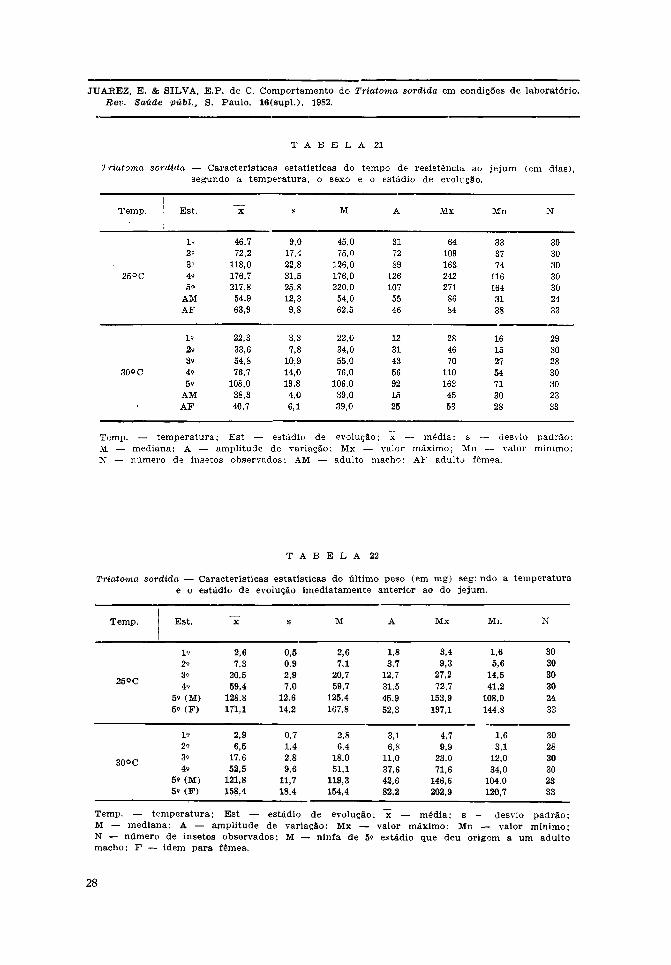

Ainda no que diz respeito às Tabelas 21e 22, a comparação dos valores médios deresistência ao jejum oferece uma visão prá-tica da influência da temperatura. Os va-lores médios à temperatura de 30°C emrelação aos de 25°C representaram, percen-tualmente, uma redução de 52,2%, 53,5%,53,6%, 56,6%, 50,4%, 30,2% e 36,3%, do1.° estádio até o estádio adulto (macho efêmea), respectivamente. O aumento de 5°C,nas condições deste trabalho, determinouum acentuado decréscimo na capacidade desobreviver ao je jum absoluto do T. sordida,mais evidente nos estádios ninfais. Entre osadultos, ocorreu decréscimo menos acen-tuado, denotando que, nesse estádio, tanto osmachos como as fêmeas têm possibilidadesmais limitadas de resistência ao jejum.
A resistência ao jejum cresce do 1.° até5.° estádio e em seguida decresce para osadultos fêmeas e machos, em ambas astemperaturas.
Em que pese as médias de resistênciaao jejum nas temperaturas estudadas seremapreciavelmente diferentes, os valores má-ximos a 30°C e os valores mínimos a 25°Caproximam-se o bastante para se super-porem no 2.° estádio e no estádio adultomacho e fêmea. Novamente, pois, sugere--se a hipótese de que o fenômeno de des-gaste no jejum se processa segundo umadistribuição normal.
As médias de resistência ao jejum crescemdo 1.° ao 5.° estádio de forma bastanteregular, com inclinação maior a 25°C doque a 30°C. Os adultos a 25°C tiverammédias intermediárias entre as dos 1.° e 2.°estádios. A 30° C as médias dos adultostiveram valores intermediários às do 2.° e3.° estádios, seja por esta temperatura sermais atuante nas ninfas de 1.° estádio, sejapor uma resistência maior dos adultos.
Convém ressaltar os valores máximos emínimos observados, especialmente no 5.°estádio. À temperatura de 25°C esses valoresforam 271 e 164 dias e a 30°C foram 163e 71 dias, lembrando que essa sobrevida sedeu em jejum absoluto. Ainda nesse estádioo valor da mediana mostra que 50% dos
insetos nessas condições de jejum, podemsobreviver 220 e 106 dias, respectivamentea 25°C e 30°C.
Ressalta-se ainda o comportamento dasninfas de 4.° estádio, em que os valoresda média e mediana praticamente seconfundem, a 25°C e 30°C, mostrando aapreciável capacidade de viverem sem qual-quer tipo de alimento.
Os resultados apresentados neste trabalho,em relação ao T. sordida, permitem fazeralgumas considerações, tendo em vista abibliografia consultada. É de se supor que,o observado para a resistência ao jejum,nesta espécie, deva ser o menor possível,tendo em vista que os insetos foram subme-tidos ao jejum absoluto. É lícito supor quea ingestão de qualquer quantidade de água,sangue ou o resultado da predação devaprolongar a sobrevida em jejum. Ryckman 30
(1962) afirmou que, teoricamente, se ostriatomíneos fossem postos em jejum, emcada estádio sucessivo, até próximo à mortee então alimentados, poderiam passar maisde três anos como ninfas.
Desde que o objetivo deste trabalho seprende à observação do T. sordida em jejumabsoluto, torna-se difícil a comparação dosresultados obtidos com o de outros autoresque observaram o jejum de triatomíneosjá alimentados ou sem conhecer a sua con-dição prévia em relação à alimentação.
Na Tabela 23 verifica-se que o T. sor-dida a 25°C, para os cinco estádios deevolução, resiste mais ao jejum do que oT. dimidiata e T. infestans à temperaturaambiente, exceto no primeiro estádio emque a última espécie resistiu mais. A 30°Co T. sordida resistiu menos ao jejum doque o P. megistus em todos os estádios deevolução, inclusive nos adultos machos efêmeas. A esta temperatura, o T. sordidaresistiu mais ao jejum do que o T. brasi-liensis nos 3.°, 4.° e 5.° estádios, tendo mos-trado sobrevida menor nos demais estádios.
Em relação aos adultos, as fêmeas deT. sordida resistiram mais do que os machosnas duas temperaturas, da mesma formaque o P. megistus. No T. dinudiata, prati-
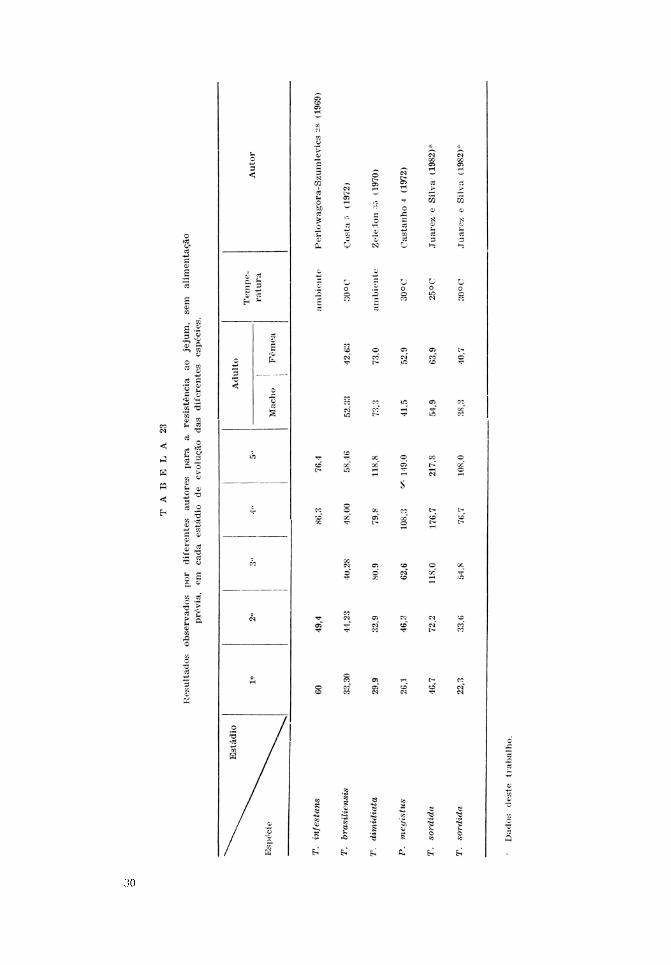

camente não ocorreu diferença, enquanto noT. brasiliensis a fêmea resistiu menos doque o macho.
Na tentativa de se conhecer quais asvariáveis que estariam interferindo naduração do jejum, foram aplicados testesde correlação entre a resistência aojejum em determinado estádio (RJ i)e as variáveis em estudo no estádioimediatamente anterior. Para o 1.° estádio,a única variável possível foi a duraçãoda fase embrionária TI. Nos demaisestádios, as correlações foram feitascom Di-1, Si-1, Ri-1, Pi-1 e UP i - 1 .
A Tabela 24 contém o resultado dascorrelações efetuadas e sua significânciaestatística. No primeiro estádio, a únicacorrelação possível mostrou que a 25°C aresistência ao je jum aumenta na medidaem que decresce o período de duração dafase embrionária. É possível que a maiorduração dessa fase de evolução dos triato-míneos acarrete um desgaste maior dasreservas nutritivas do ovo, restando, propor-cionalmente, menor quantidade para a ninfade 1.° estádio, após a eclosão. A 30°C, oresultado dessa correlação sugere que, apesardo metabolismo ser mais acelerado, provo-cando mais rapidamente a eclosão do ovo,não chega a haver maior consumo propor-cional de reservas do ovo, como decor-rência da menor amplitude de variação deTI.
A resistência ao jejum no 2.° estádio mos-trou, em ambas as temperaturas, correla-ções significantes e positivas com a quan-tidade de sangue ingerido e com o últimopeso no 1.° estádio. Considerado o pequenopeso da ninfa de 1.° estádio e a grandequantidade proporcional de sangue que écapaz de ingerir, pode-se supor que, nomomento da muda para o 2.° estádio, existauma reserva de sangue no tubo digestivodo inseto que determinará a maior ou menorresistência ao jejum, uma vez que suaestrutura física não permite prever reservasapreciáveis.
No 3.° estádio, observa-se o mesmo com-portamento em relação ao UP2, nas duas
temperaturas. Houve correlação positiva esignificante entre RJ3 e UP2. No entanto,não se observou o mesmo com a variávelS2, que apresentou correlação positiva enão significante. A resistência ao jejum foitanto maior quanto maior a soma de energiaexistente na ninfa por ocasião da muda, emsangue no tubo disgestivo ou em estruturafísica do inseto.
O 4.° estádio teve comportamento seme-lhante ao do 2.° em relação às correlaçõescom sangue ingerido e último peso. Essasduas variáveis forneceram correlações posi-tivas e significantes com a resistência aojejum. Ocorreu também correlação positivae significante com o número de repastosefetivamente realizados no 3.° estádio, a30°C. Este resultado leva à suposição deque os insetos que ingerem mais sangueo fazem em um número maior de vezes.
O 5.° estádio ofereceu resultados contra-ditórios em relação às temperaturas. A 30°C,como nos estádios anteriores, ocorreraminterações entre a resistência ao je jum e oúltimo peso no 4.° estádio, bem como, coma quantidade de sangue ingerido nesseestádio. A 25°C não ocorreram correlaçõessignificantes, porém, o último peso no 4.°estádio e a resistênciaa ao je jum no 5.°mostraram uma correlação positiva e muitopróxima de ser significante (P = 0,05145).O que chama a atenção é o fato da cor-relação com o sangue ingerido no 4.° estádioter resultado negativo e não significante,nessa temperatura, indicando que há umarelação inversa entre essas variáveis, o quenão parece ter nexo.
Em relação aos adultos machos, ocorreramcorrelações significantes e negativas, nasduas temperaturas, com o número de re-pastos do 5.° estádio. Quanto às fêmeas,a 25°C ocorreram correlações significantese negativas com a duração, sangue ingeridoe número de repastos no 5.° estádio. A 30°Cas fêmeas apresentaram apenas uma cor-relação significante e positiva, com o últimopeso no 5.° estádio.


A única correlação sistematicamente posi-tiva foi a que ocoreru entre resistência aojejum e o último peso do estádio imediata-mente anterior, porém, apenas oito foramsignificantes, das doze possíveis. As demaiscorrelações tiveram resultados ora positivos,ora negativos, não permitindo qualquer tipode suposição para sua explicação.
A série de trabalhos de Forattini ecol.11-17 (1971, 1973, 1974, 1975, 1977,1979) enfatiza a importância que estaespécie tem no momento atual, bem como aexpectativa que se poderá ter no futuro.Utilizando-se de ecótopos artificiais, gali-nheiros experimentais localizados nas áreasonde o T. sordida habita naturalmente, essesautores mostraram a valência ecológicaapreciável que tem esta espécie, o que lhepermite a colonização em diferentes tiposde ecótopos, sejam naturais, sejam artificiais,mesmo quando as condições para a alimen-tação não sejam as mais adequadas. Exem-plares dessa espécie foram encontrados emárvores secas, em ambientes muito alteradospor desmatamentos e queimadas em que aspossibilidades de obter alimento são muitoremotas. Ficou patente a atração que osecótopos artificiais podem exercer sobreesses triatomíneos. Outro aspecto mostradoé o desenvolvimento numérico da populaçãoquando desaparecem os inimigos naturais.
Esses fatos permitem supor que, comoresultado de sua valência ecológica alta oucomo determinante desta, a capacidadeampla de resistir ao jejum seja um fatomuito importante e que poderá servir paraespeculações em outros trabalhos de campo.Essa resistência ao jejum lhe confere umacapacidade de "espera" de alimento, o quepermite a suposição de que têm pequenamobilidade, especialmente as ninfas. Pos-sivelmente são insetos muito pouco agres-sivos, quando em busca de alimento, sendode certa forma cerceados pelos inimigosnaturais. A associação dos resultadosobtidos em laboratório aos observados nostrabalhos de campo permitem fazer algumasconsiderações como as que foram apresen-tadas, com base em alguns fatos. A tempe-
ratura não teve efeito significantementediferente quando se observou a quantidadede sangue ingerido no 5.° estádio. Pode-sesupor uma capacidade de resistência àsvariações de temperatura, manifestada pelanão aceleração ou não aumento de neces-sidades do seu metabolismo, o que lhe con-feriria a "espera" da época oportuna paracontinuar o desenvolvimento, uma vez queos adultos não apresentaram a elevadaresistência ao je jum observada nesse estádio.
Restou apenas a explicação, de certaforma óbvia, de que a duração do inseto emjejum depende do último peso que apresen-tava antes da muda no estádio anterior.
Novamente se enfatiza a necessidade demaiores conhecimentos sobre fisiologia, bio-química, genética e ecologia para a formu-lação de hipóteses convincentes. Quaisquerque sejam as explicações possíveis, éinegável o valor da informação referenteà capacidade dos triatomíneos resistirem aojejum por períodos tão prolongados, paraestudos ecológicos.
A perspectiva de ser mantida a infestaçãode casas e anexos por T. sordida, mesmoquando abandonados pelo homem e animaisé digna de ser considerada em termos epi-demiológicos.
CONCLUSÕES
1. A duração da fase embrionária foimaior à temperatura de 25°C que àde 30°C, não tendo ocorrido interaçãotemperatura-sexo.
2. A duração do ciclo evolutivo foi maiorà temperatura de 25°C que à de 30°Cnos estádios 1.°, 2.°, 3.° e 5./. No 4.°estádio ocorreu interação temperatura--sexo.
3. A duração do 5.° estádio do cicloevolutivo foi maior nos machos do quenas fêmeas.
4. A duração do adulto macho foi menordo que a das fêmeas, à temperaturade 25°C. À temperatura de 30°C nãoocorreu diferença significante entre asmédias dos machos e das fêmeas.

5. A quantidade média de sangue ingeridanos 1.° a 4.° estádios do ciclo evolutivo,foi maior à temperatura de 25°C doque à de 30°C. No 5.° estádio nãoocorreu diferença significante porefeito da temperatura.
6. As quantidades médias de sangue inge-rido foram significantemente diferentesnos 3.°, 4.° e 5.° estádios do ciclo evo-lutivo. Nos 4.° e 5.° estádios as médiasdas fêmeas foram maiores.
7. O número médio de repastos, nos 1.°,2.° e 3.° estádios do ciclo evolutivo,foi maior à temperatura de 25°C queà de 30°C. No 4.° estádio ocorreuinteração temperatura-sexo. No 5.°estádio não ocorreu diferença signi-ficante por efeito da temperatura.
8. O número médio de repastos dosmachos no 5.° estádio do ciclo evolu-tivo foi maior que o das fêmeas, àtemperatura de 25°C. Nos demaisestádios não ocorreram diferençassignificantes por efeito do sexo.
9. O peso médio inicial, exceto no pri-meiro estádio, foi maior, em todos osestádios do ciclo evolutivo e no estádioadulto, à temperatura de 25°C do queà de 30°C.
10. O peso médio inicial foi maior entreas fêmeas nos 4.° e 5.° estádios do cicloevolutivo e fase adulta, à temperaturade 30°C. À temperatura de 25°C, opeso médio inicial das fêmeas foi maiorno 5.° estádio e na fase adulta.
1 1 . A resistência média ao jejum foi maiorà temperatura de 25°C do que à de30°C, em todos os estádios do cicloevolutivo. Na fase adulta ocorreu inte-ração temperatura-sexo.
12. As correlações entre resistência aoje jum e último peso no estádio imedia-tamente anterior foi a única que semostrou sistematicamente positiva,sendo significante em oito das dozepossíveis, considerados os estádios deevolução e temperaturas.
JUAREZ, E. & SILVA, E. P. de C. [The behavior of Triatoma sordida under laboratoryconditions]. Rev. Saúde públ., S. Paulo, 16(suppl.), 1982.
ABSTRACT: The behavior of Triatoma sordida maintained in stoves withrelative atmospheric humidity of 60-70% and temperatures, respectively, of25°C and 30°C was observed. The higher temperature caused a shorteningof the growth phases. The growth of the fifth instar took longer in malesthan in females. The longevity of adult females was greater than that ofmales. With regard to ingested blood higher averages were observed at25°C, except in the 5th instar, and the females had the highest averagesin the 4th and 5th instars. With regard to the number of repasts, at 25°Cthe average was higher in the 1st, 2nd and 3rd instars; there was interactionof temperature and sex in the 4th instar and there was no significantdifference in the 5th instar; the number of repasts of males was largerthan that of females only in the 5th instar. The initial weight of triatomidbugs was greatest at 25°C in all the instars except the 1st; females at 25°Chad greater weight than males in the 4th and 5th instars and at 30°C inthe 5th instar only. Effects of temperature and sex in the starvation resis-tance of T. sordida were observed. The triatomid bugs were not manipulatedor fed after the move to instar study. The average of starvation resistancemeasured in days was longer in all the instars of the growth cycle, but, inthe adult phase, there was interaction of temperature and sex. The corre-lation between starvation resistance and the last weight of the immediatelyprevious instar was the only one that was consistently positive, being signi-ficant in eight only of the twelve possible correlations.
UNITERMS: Triatoma sordida. Triatominae, laboratory conditions.

REFERÊNCIAS BIBLIOGRÁFICAS
1. ABALOS, J.W. & WYGODZINSKY, P. Lastriatominae argentinas. Tucuman, Uni-versidad Nacional de Tucuman. Insti-tuto Medicina Regional, 1951. (Publi-cacion, 601).
2. BODENSTEIN, D. Postembryonic develop-ment. In: Roeder, K.D., ed. Insectphysiology. New York, Wiley & Sons,1953. p. 822-65.
3. BUXTON, P.A., 1930 Apud CORRÊA, F.M.A.Estudo comparativo do ciclo evolutivodo Triatoma infestans alimentado emdiferentes animais, (Hemiptera, Redu-viidae). Pap. avulso Zool., S. Paulo,15:177-200, 1962.
4. CASTANHO, M. L. S. Observações sobre aevolução e o jejum do Panstrongylusmegistus em laboratório. São Paulo,1972. [Tese de Doutoramento — Institutode Ciências Médicas USP]
5. COSTA, M.J. Estudo do jejum em Triatomabrasiliensis. São Paulo, 1972. [Disser-tação de Mestrado — Instituto deBiociências USP]
6. DANILOV, V. N. The effect of blood-meatsize taken in the nymphal stage onmoulting into imago in triatomid bugs.Medskaya Parazit., 46:218-23, 1969. ApudTrop. Dis. Bull, 65:982, 1968.
7. DIAS, E. Notas sobre o tempo de evoluçãode algumas espécies de triatomíneos emlaboratório. Rev. bras. Biol., 15:157-8,1955.
8. DIAS, E. Observações sobre eliminação dedejeções e tempo de sucção em algunstriatomíneos sul-americanos. Mem. Inst.Oswaldo Cruz, 54:115-24, 1956.
9. DIAS, J.C.P. Observações sobre o compor-tamento de triatomíneos brasileirosfrente ao jejum em laboratório. Rev.bras. Malar., 17:55-64, 1965.
10. FERREIRA, V.L. Relações entre a quan-tidade de sangue ingerido no 5.° estádioe a muda imaginal em Triatoma brasi-liensis. São Paulo, 1972. [Dissertação deMestrado — Instituto de BiociênciasUSP]
11. FORATTINI, O.P. et al. Aspectos eco-lógicos da tripanossomose americana. II— Distribuição e dispersão local de
triatomíneos em ecótopos naturais eartificiais. Rev. Saúde públ., S. Paulo,5:163-91, 1971.
12. FORATTINI, O.P. et al. Aspectos eco-lógicos da tripanossomose americana. III— Dispersão local de triatomíneos comespecial referência ao Triatoma sordida.Rev. Saúde públ., S. Paulo, 5:193-205,1971.
13. FORATTINI, O.P. et al. Aspectos eco-lógicos da tripanossomose americana. V— Observação sobre a colonização espon-tânea de triatomíneos silvestres em ecó-topos artificiais, com especial referênciaao Triatoma sordida. Rev. Saúde públ.,S. Paulo, 7:219-39, 1973.
14. FORATTINI, O.P. et al. Aspectos eco-lógicos da tripanossomíase americana. VI— Persistência do Triatoma sordida apósalteração ambiental e suas possíveisrelações com a dispersão da espécie.Rev. Saúde públ., S. Paulo, 8:265-82,1974.
15. FORATTINI, O.P. et al. Aspectos eco-lógicos da tripanossomíase americana. VII— Permanência e mobilidade do Tria-toma sordida em relação aos ecótoposartificiais. Rev. Saúde públ., S. Paulo,9:467-76, 1975.
16. FORATTINI, O.P. et al. Aspectos eco-lógicos da tripanossomíase americana. X— Dados populacionais das colônias dePanstrongylus megistus e de Triatomasordida espontaneamente desenvolvidasem ecótopos artificiais. Rev. Saúde públ.,S. Paulo, 11:362-74, 1977.
17. FORATTINI, O.P. et al. Aspectos eco-lógicos da tripanossomíase americana. XVI— Dispersão e ciclos anuais de colôniasde Triatoma sordida e de Panstrongylusmegistus espontaneamente desenvolvidasem ecótopos artificiais. Rev. Saúde públ.,S. Paulo, 13:299-313, 1979.
18. FRIEND, W.G. et al. The effect of nutrientintake on the development and the eggproduction of Rhodnius prolixus Stal(Hemiptera: Reduviidae). Canad. J. Zool,.43:891-904, 1965.
19. GOODCHILD, A.J.P. Some observation ongrowth an egg production of the blood--sucking Rectuviids Rhodnius prolixusand Triatoma infestans. Proc. roy. ent.Soc. London A, 30:137-44, 1955.

20. HACK, W. H. Estudios sobre la biologiadel Triatoma infestans (Klug, 1834)(Hemiptera, Reduviidae). Ann. Inst.Med. region., Comentes, 4:125-47, 1955.
21. JUAREZ, E. Observações sobre o cicloevolutivo do Triatoma arthurneivai emcondições de laboratório. (Hemiptera,Reduviidae). Rev. Saúde públ., S. Paulo,4:13-8, 1970.
22. JUAREZ, E. Comportamento do Triatomainfestans sob várias condições de labora-tório. Rev. Saúde públ., S. Paulo, 4:147--66, 1970.
23. LUCENA, D. T. Evolução do Triatoma brasi-liensis Neiva, 1911, no laboratório. Rev.bras. Biol., 20:295-302, 1960.
24. LWOFF, M. & NICOLLE, P. Necessité de1'hématyne pour la nutrition de Triatomainfestans Klug. (Reduvidé-Hémophage).C.R. Soc. Biol. Paris, 139:879-81, 1945.
25. NEIVA, A. Informações sobre a biologiada Vinchuca, Triatoma infestans KLUG.Mem. Inst. Oswaldo Cruz, 5:24-31, 1913.
26. PELEGRINO, J. Observações sobre a resis-tência do Triatoma infestans ao jejum.Rev. bras. Biol., 12:317-20, 1952.
27. PERLOWAGORA SZUMLEVICS, A. Cicloevolutivo do Triatoma infestans emcondições de laboratório. Rev. bras.Malar., 5:35-48, 1953.
28. PERLOWAGORA SZUMLEVICS, A. Estudossobre a biologia do Triatoma infestans,o principal vetor da doença de Chagasno Brasil. Rev. bras. Malar., 21:117-59,1969.
29. PIPPIN, W. E. The biology and vectorcapability of Triatoma sanguisuga texanaUsinger and Triatoma gerstaecker (Stal)compared with Rhodnius prolixus (Stal).Hemiptera: Reduviidae). J. med. Ent.,7:30-45, 1970.
30. RYCKMANN, R.E. Biosystematics andhosts of Triatoma protracta complex inNorth America. (Hemiptera: Reduviidae)(Rodentia, Cricetidae). Univ. Calif. Publ.Ent., 27:93-240, 1962.
31. SOUZA, J.M.P. de et al. Triatoma sordida:considerações sobre o tempo de vida dasformas adultas e sobre a oviposição dasfêmeas. Rev. Saúde públ., S. Paulo,12:291-6, 1978.
32. TOBAR, R.G. Capacidad de ayuno de lostriatomineos chilenos. Bol. Inf. Par. chil.,7:56-9, 1952.
33. VIEIRA, C.A.B. Necessidade e aproveitamentoda alimentação sanguínea pelo Triatomainfestans sob diferentes condições expe-rimentais. São Paulo, 1971. [Dissertaçãode Mestrado — Faculdade de SaúdePública USP]
34. WIGGLESWORTH, V.B. Nutrition andreproduction in insects. Proc. Nutr. Soc.,19:18-23, 1960.
35. ZELEDON, R. et al. Biology and ethologyof Triatoma dimidiata (Latreille, 1811)I. Life cycle, amount of blood ingested,resistance to starvation and size ofadults. J. med. Ent., 7:313-9, 1970.
Recebido para publicação em 03/11/1981
Aprovado para publicação em 03/05/1982