stress induced g-protein signalling cascade
TRANSCRIPT

1
INTRODUCTION
1.1 Stressors
Substantial loss of yield worldwide is due to abiotic stress (Shelden and
Roesnner 2013). In fact, more than 50% of the yield is lost to abiotic stress
(Boyer, 1982, Bray et al. 2000 as stated in Wang et al. 2003, Shelden and Roesnner
2013). For instance, in wheat, the record yield (average yield + yield without
loss to diseases, pests, and unfavouable environments) in 1975 was 14500
kg/hectares while the average yield was 1880 kg/hectares; further, losses to
unfavourable physicochemical environments was 12156 kg/hectares (Boyer, 1982).
Translated, this would mean a loss of 87% to diseases, pests, and unfavourable
physicochemical environments; in particular, a loss of 71.7% to unfavourable
environmental conditions (Boyer, 1982).
Drought, salinity, cold, heat, oxidative stress, and chemical toxicity are the
primary stressors of a plant (Carson 2012, Wang et al 2003). The damage and
cellular response of plant to any of the above stressors tend to be similar;
they are interconnected in terms of the response induced by the plant (Wang et
al. 2003). However, cell identity and the number of simultaneous stresses

2
determine stress-induced gene expression patterns (Dinneny et al. 2008 as
stated in Voesenek and Pierik, 2008). When both heat and drought stresses were
simultaneously subjected onto a plant – Arabidopsis thaliana – only 10% of the
genes imbricated between the double-stressed and a single-stressed treatment
were expressed (Voesenek and Pierik, 2008). The latter would imply that the
gene expression profile, and control networks of multiple stresses cannot be
adumbrated by expression profiles generated for single stress treatments
(Voesenek and Pierik, 2008).
Stressors usually cause disruption of osmotic and ionic homeostasis, or damage
to functional, and structural proteins, and membranes (Wang et al. 2003, Figure
1). The genetic makeup of any given plant is responsible for the stress-induced
response. However, acclimation can also influence the response of a plant
(Carson 2012). Genes regulated by the stressors usually are involved in
signaling cascades, or transcriptional control, function to directly protect
membranes and proteins, or are involved in water and ion uptake and
transportation (Wang et al. 2003). Tolerance to stressors is usually conferred
by modifying the internal environment to permit internal structural
stability, and function of internal components (Wang et al. 2003).
In other words, if one can manage to retain the structural integrity of the
cell, the effect of the stressor would be minimized (Wang et al. 2003). Response
to salt stress usually involves tolerance (plant maintains water uptake, and

3
growth), Na+ exclusion (removal of toxic ions from shoot tissue), and tissue
tolerance (selectively isolate toxic ions into vacuoles (Munns and Tester, 2008
as stated in Shelden and Roesnner 2013). Munns and Tester (2000, 2008) (as stated
in Shelden and Roesnner 2013) noted that a sub-population of the duram wheat
genotypes possessed the ability to tolerate salt stress via Na+ exclusion;
later, Nax1 and Nax2 were identified as the genes conferring this tolerance,
and incorporated into alternate lines to decree salt-stress tolerance to scions
(Shelden and Roesnner 2013).
In Triticum aestivum, for example, a number of genes were found to be up-
regulated by the said stressors (Khalil 2013, Tardif et al. 2007). A scad of these
genes are regulatory factors (ex: protein kinases, transcription factors,
ubiquitin ligases, GTP and calcium binding proteins; therewith, up-regulated
genes may act to modify chromatin or act as post-transcriptional regulation
(Tardif et al. 2007). However, the external architecture of the plant is also
retailored to specific stresses. For instance, in response to water stress,
plants have been noted to suppress lateral root formation, and initiate
primary root formation in an attempt to reach the deeper layers of the soil
where the probability of water discovery is superlative (Xiong et al 2006 as
stated in Wright 2014, thesis).
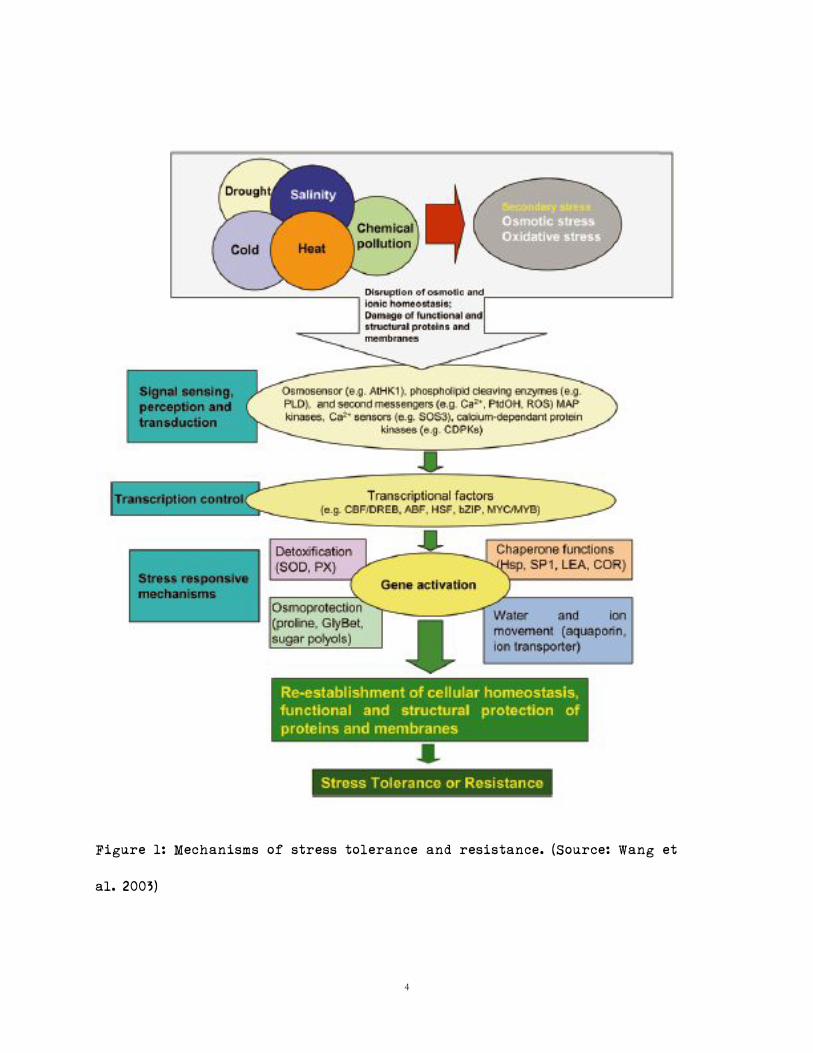
4
Figure 1: Mechanisms of stress tolerance and resistance. (Source: Wang et
al. 2003)

5
1.2 Heterotrimeric G Proteins
Animals
G proteins can be classified into two distinct categories – the heterotrimeric
G proteins, and the monomeric G proteins (the small G proteins); the monomeric G
proteins do not include the Gsβγ subunits. However, both the receptor-binding
region and GTPase activity are found on the Gsα subunit; in other words, absence
of either function on the Gsβγ subunit permits exclusion of said subunit by the
monomeric G protein complex.
The heterotrimeric G proteins are involved in signal transduction pathways.
Coupled with cell-surface receptors (henceforth referred to as the G-protein
coupled receptors (GPCR)), the Gs proteins are a network of three discrete
subunits – the α, β, and γ subunits (Lodish et al., 2000, Figure 2). Ligand
binding induces the dislocation and dissociation of the Gsα subunit. The entire
complex vacillates between two alternate states – the “on”, and the “off” state.
This vascillation is dependent upon the bound state of the Gsα subunit. In other
words, in the “off” state, the Gsα subunit is bound to a GDP molecule. However,
ligand binding to GPCR dislodges the GDP and permits a GTP molecule to bind the
α subunit – henceforth referred to as the Gsα GTP complex. The dislocated α
subunit then activates an effector protein which activates an intercellular
second messenger (Lodish et al., 2000). Simultaneously, the bound GTP is

6
hydrolyzed, and the Gsα subunit returns to the initial heterotrimeric state,
bound to the β, and γ subunits.
Plants
In plants, only five GPCRs have been found to interact with the Gsα subunit –
AtGCR1, AtRGS1, GPCR, and GTG1, and GTG2 (REF---paper, Khalil et al. 2011).
Effector proteins, on the other hand, can be any one of the following: Ca2+,
calmodulin (CaM), cAMP, cGMP, cADP-ribose, inositol 1,4,5-triphosphate (IP3) and
reactive oxygen species (ROS) (Tardif et al. 2007). The signaling cascade can
result positively or negatively regulate expression of target genes (Tardif et
al. 2007).
Response to stress – pathogens, light, high salinity, hypoxia, drought, ethylene
signaling, signaling pathways regulated by jasmonic acid, gibberellin and
abscisic acid (ABA) - in plants has been noted to involve G-protein signaling.
Plant response to pathogens, for instance, is elicited by modulating
intracellular Ca2+ levels which is itself triggered by G proteins.
Animals possess 23 Gα, 6 Gβ, and 12 Gγ encoding genes while plants possess one
Gα (GPA1), one Gβ (AGB1), and 2 Gγ (AGG1, AGG2) encoding genes (Jones 2002, Figure
2). In plants, the subunits are designated as follows: GPA1 (alpha subunit), AGB1
(beta subunit), AGG1/2 (gamma-A subunit), and AGG3 (gamma-C subunit). G proteins
are involved in regulation of ion-channels, seed germination, light responses,

7
cell division, cell elongation, and phytohormones (ABA, GA, auxin) (Jones and
Assmann 2004). In particular, heterotrimeric G proteins in plants have been
noted to induce phytohormones, and control mitosis (Jones 2002). Mutations in
Arabidopsis GPA1 leads to fewer mitosis during formation of hypocotyl, and
leaf; overexpression leads to ectopic mitosis, and overproliferation of
meristem suggesting that GPA1 is involved in the control of cellular division
(Jones 2002).
G proteins in plants are believed to function in ion channel, ABA signaling,
and cellular proliferation (Tuteja and Sopory, 2008). Mutations in members of
G-protein signaling have been shown to affect seed germination, shoot/root
growth, and control of stomata (Tuteja and Sopory, 2008). The G-protein system
in plants is still under study as not much is known about the system (Tuteja
and Sopory, 2008).
Animals have about 1000 GPCRs while plants have been noted to have a single
GPCR (Tuteja and Sopory, 2008). In Arabidopsis thaliana, a GPCR called GCR2 was
found for ABA, however, ABA is recognised by a number of plant receptors
(Tuteja and Sopory, 2008). In plants, there are three known effectors -
Phospholipase D (PLD), K+ and Ca2+ channels (Jones 2002). PLD converts
phospholipids to phosphatidic acid (PA) – where PA acts as a second messenger -
which activates genes involved in stress tolerance, defense, meiosis,
phytochrome action, membrane metabolism, and trafficking of vesicles (Tuteja

8
and Sopory, 2008).
Figure 2: Proposed model of stress-induced G-protein signaling cascade in
animals versus plants. (Source: Tuteja and Sopory 2008)
1.3 RD20/Clo3
One stress-induced gene is CLO3/RD20 – which was found to increase during
stresses (Wright 2014). Ta-CLO3 (CLO3 in Triticum aestivum) was noted to
interact with Ta-GA1 – the Galpha subunit in Triticum aestivum (Wright 2014).
In Arabidopsis thaliana, under abiotic stresses, a homolog of CLO3 was found to
be activated; a member of the caleosin family, CLO3 encodes a protein that is
highly abundant when subjected to abiotic stresses. ABA as well as other
stresses (water, salt, osmotic) were found to increase CLO3/RD20 in Arabidopsis
thaliana – a 150 fold increase in CLO3 after 2 hours of dehydration, and a 300

9
fold increase 3 hours after ABA treatment (Aubert et al. 2010). In particular,
CLO3 was found to modulate stomatal opening/closing in guard cells (Aubert et
al. 2010). RD20 has been noted to be induced by water stress, ABA, and NaCl
treatments; RD20 was not found to be upregulated by cold treatments (Takahashi
et al., 2000). RD20 mRNA can be observed in leaves, flowers, stems, and siliques,
but not in roots or mature seeds (Takahashi et al., 2000).
1.4 Bimolecular Fluorescence Complementation (BiFC)
Bimolecular fluorescence complementation (BiFC) is a method utilized to detect
protein-protein interactions in vivo. Two potential interacting partners –
the bait, and the prey – are non-fluorescing. Fusion proteins of the bait, and
prey are created in such a manner that the N-terminal of a fluorescent
protein (ex: YFP) is fused to one partner while the C-terminal is fused to the
other (Kerppola, 2008). Interaction between bait, and prey brings into close
proximity the two halves of the fluorescent proteins – a bimolecular
fluorescence complex is formed - which if excited, emits fluorescence
(Kerppola, 2008). BiFC can be used to identify the subcellular location of a
protein complex (Kerppola, 2008).
The advantage of using BiFC is that its intrinsic signal is strong enough to
enable detection; alternative methods require exogenous chromogenic or

10
fluorogenic agents (Kerppola, 2008). Further, a number of interactions can be
detected simultaneously by using distinct fluorescence proteins (ex: YFP, CYF,
etc…). However, the signal is usually detected after the formation of the
complex – there is a time lapse between actual interaction, and visibility of
observation, and is therefore not considered a real-time observation (Kerppola,
2008).
Given that not all fluorescent proteins are candidates for BiFC, two in
particular have been noted to function well – YFP, and CFP. YFP is usually
truncated in the following manner: 1-154 consists of the N-terminal portion,
and 155-238 consists of the C-terminal portion; alternatively, one can generate
an N-terminal from 1-172, and a C-terminal from 173-238 (Kerppola, 2008,
Figure 3). Further if the concentration of the fluorescent proteins is high
within a cell, random association between fragments can generate false
positives; it is therefore recommended that fusion proteins be expressed at low
levels or levels typical to the cell (Kerppola, 2008). BiFC usually requires
multiple attempts or generation of multiple combinations (FIG) to determine the
combinations that [work], and that generate the strongest signal (Kerppola,
2008).
BiFC also requires negative controls to validate the protein-protein
interaction detected. Efficiency is calculated as the ratio of the fluorescence
emitted by the YN-YC complex to an internal control (ex: CFP) (Kerppola, 2008).
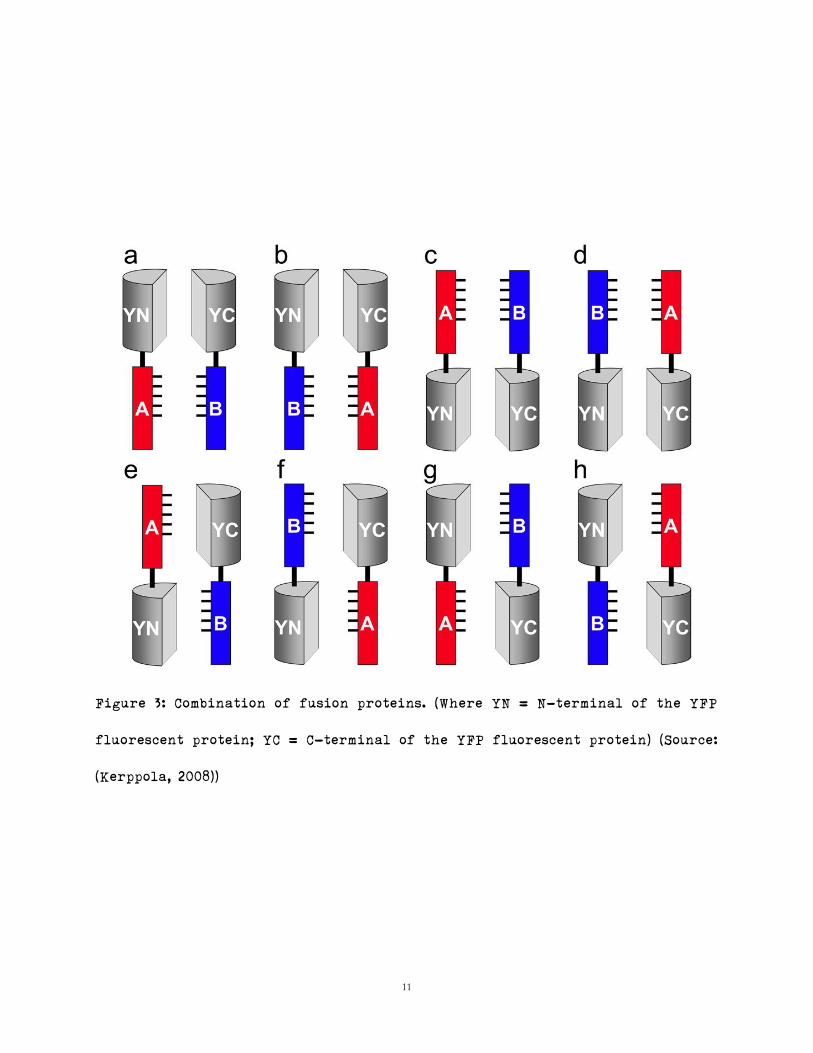
11
Figure 3: Combination of fusion proteins. (Where YN = N-terminal of the YFP
fluorescent protein; YC = C-terminal of the YFP fluorescent protein) (Source:
(Kerppola, 2008))

12
References
Aubert, Y., Vile, D., Pervent, M., Aldon, D., Ranty, B., Simonneau, T., Vavasseur,
A., and Galaud, J. (2010). RD20, a Stress-Inducible Caleosin, Participates in
Stomatal Control, Transpiration and Drought Tolerance in Arabidopsis
thaliana. Plant And Cell Physiology 51, 1975-1987.
Boyer, J. (1982). Plant Productivity and Environment. Science 218, 443-448.
Carson, R. (2012). The Interaction of the Stress Regulated Wheat (Triticum
aestivum) Proteins, ESI2 and RAN1, and the Localization of Ipp-1 and Pr-1.
Master of Science. Concordia University.
Nallamsetty, S., and Waugh, D. (2007). A generic protocol for the expression
and purification of recombinant proteins in Escherichia coli using a
combinatorial His6-maltose binding protein fusion tag. Nature Protocols 2,
383-391.
Lodish, H., Berk, A., Zipursky, L., Matsudaira, P., Baltimore, D., and Darnell, J.
(2000). Molecular Cell Biology, New York.
Jones, A. (2002). G-protein-coupled signaling in Arabidopsis. Current Opinion
In Plant Biology 5, 402-407.

13
Jones, A., and Assmann, S. (2004). Plants: the latest model system for G-protein
research. EMBO Rep 5, 572-578.
Kerppola, T. (2008). Bimolecular Fluorescence Complementation (BiFC)
Analysis as a Probe of Protein Interactions in Living Cells. Annual
Review of Biophysics, 37(1), pp.465-487.
Khalil, H. (2013). The characterization of the caleosin gene family in
Triticeae and their role in G-protein signalling and Identification
and characterization of rye genes silenced in allohexaploid triticale:
A bioinformatic study. Doctor of Philosophy. Concordia University.
Shelden MC, Roessner U. Advances in functional genomics for
investigating salinity stress tolerance mechanisms in cereals.
Frontiers in Plant Science. 2013;4, article 123 [PMC free article] [PubMed]
Takahashi, S., Katagiri, T., Yamaguchi-Shinozaki, K. and Shinozaki, K.
(2000). An Arabidopsis Gene Encoding a Ca2+-Binding Protein is Induced
by Abscisic Acid during Dehydration. Plant Cell Physiol, 41(7), pp.898-
903.

14
Tardif, G., Kane, N., Adam, H., Labrie, L., Major, G., Gulick, P., Sarhan, F., and
Laliberté, J. (2007). Interaction network of proteins associated with abiotic
stress response and development in wheat. Plant Mol Biol 63, 703-718.
Tuteja, N. and Sopory, S. (2008). Plant signaling in stress. Plant
Signaling & Behavior, 3(2), pp.79-86.
Urano, D., Chen, J., Botella, J. and Jones, A. (2013). Heterotrimeric G protein
signalling in the plant kingdom. Open Biology, 3(3), pp.120186-120186.
Voesenek, L., and Pierik, R. (2008). PLANT SCIENCE: Plant Stress Profiles.
Science 320, 880-881.
Wang W, Vinocur B & Altman A 2003 Plant responses to drought, salinity
and extreme temperatures: towards genetic engineering for stress
tolerance. Planta 218 1-14. (doi:10.1007/s00425-003-1105-5).
Wang Z 2009 The interaction between the Ga subunit of the
heterotrimeric G protein complex with calcium binding proteins,
AtClo3/RD20, and AtClo7 from Arabidopsis and TaClo3 from wheat,

15
Triticum aestivum. Concordia University.
Wright J 2014 RD20/CLO3, a stress-induced calcium-binding protein, acts
as a negative regulator of GPA1 in Arabidopsis through GAP activity.
Concordia University.