stereotypes and stereotyping: what's the brain got to do with it?
TRANSCRIPT

This article was downloaded by: [University of California Davis]On: 18 October 2014, At: 15:52Publisher: RoutledgeInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
European Review of SocialPsychologyPublication details, including instructions for authorsand subscription information:http://www.tandfonline.com/loi/pers20
Stereotypes and stereotyping:What's the brain got to do withit?Susanne Quadflieg a & C. Neil Macrae ba Division of Psychology , New York University AbuDhabi , UAE/USAb School of Psychology , University of Aberdeen , UKPublished online: 21 Dec 2011.
To cite this article: Susanne Quadflieg & C. Neil Macrae (2011) Stereotypes andstereotyping: What's the brain got to do with it?, European Review of Social Psychology,22:1, 215-273, DOI: 10.1080/10463283.2011.627998
To link to this article: http://dx.doi.org/10.1080/10463283.2011.627998
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, orsuitability for any purpose of the Content. Any opinions and views expressedin this publication are the opinions and views of the authors, and are not theviews of or endorsed by Taylor & Francis. The accuracy of the Content shouldnot be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions,claims, proceedings, demands, costs, expenses, damages, and other liabilitieswhatsoever or howsoever caused arising directly or indirectly in connectionwith, in relation to or arising out of the use of the Content.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan, sub-

licensing, systematic supply, or distribution in any form to anyone is expresslyforbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Stereotypes and stereotyping: What’s the brain got to do
with it?
Susanne Quadflieg1and C. Neil Macrae
2
1Division of Psychology, New York University Abu Dhabi, UAE/USA2School of Psychology, University of Aberdeen, UK
Influential social-psychological models suggest that perceivers frequently useinformation-processing strategies that result in the stereotype-based construaland treatment of other people. To elucidate in greater detail when, why, andhow social encounters elicit stereotyping, researchers have begun toincorporate neuroscientific tools into their repertoire of investigativemethods. The current article reviews these studies and considers theirsignificance in light of existing theories of person perception, knowledgerepresentation, and impression formation. Finally, using the concrete exampleof stereotyping, benefits and pitfalls of adopting a social neuroscienceapproach are discussed.
Keywords: Bias; Body perception; Event-related potentials; Expectancyviolations; Face perception; Functional magnetic resonance imaging; Ingroup;Outgroup; Person categorisation; Person evaluation; Person individuation;Person knowledge; Person perception; Prejudice; Social neuroscience;Stereotype activation; Stereotype application; Stereotype content; Stereotypecontrol.
As beliefs about individuals based on knowledge of the social groups towhich they belong, stereotypes offer apparent insights into the personalitiesand deeds of others without the cumbersome necessity of getting to knowthem (Allport, 1954; Brewer, 1988; Fiske & Neuberg, 1990; Hilton &
Correspondence should be addressed to Susanne Quadflieg, New York University, 19
Washington Square North, New York, NY 10011, USA. E-mail: [email protected]
SQ was supported by a University of Aberdeen Graduate Award and CNM by a Royal
Society-Wolfson Fellowship. This manuscript reflects in large parts SQ’s PhD Dissertation at
the University of Aberdeen. The section on benefits and pitfalls of social neuroscientific
approaches was additionally inspired by SQ’s participation at the interdisciplinary ENSN
Neuroschool in Bergen (March 2011), for which she would like to express her gratitude to the
organisers and other attendees.
EUROPEAN REVIEW OF SOCIAL PSYCHOLOGY
2011, 22, 215–273
� 2011 European Association of Social Psychology
http://www.psypress.com/ersp http://dx.doi.org/10.1080/10463283.2011.627998
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

von Hippel, 1996; Macrae & Bodenhausen, 2000). While clearly simplifyingthe process of person understanding, stereotypical thinking poses the dangerof promoting judgemental inaccuracy, societal inequality, and inter-groupconflict (Kunda, 1999). Recognising the impact that stereotyping exerts indaily life, researchers have tried to elucidate when, why, and how people usestereotypes for over half a century. What originally began with thesystematic examination of racial stereotypes (Katz & Braly, 1933),eventually gave rise to a field of study targeting a wide range of socialcategories (e.g., women, the elderly, Jews, Muslims, homosexuals, con-servatives, and obese individuals, to name just a few).
Summarising the resulting body of research, 15 years ago a book with thetitle Stereotypes and Stereotyping was published (Macrae, Stangor, &Hewstone, 1996). In its concluding chapter it was argued that amongscientists willing to conduct future work on the topic those ‘‘with moderntheories and powerful methods’’ would succeed (Schneider, 1996, p. 449). Intheir search for powerful methods researchers interested in stereotypinghave recently turned to neuroscientific tools of investigation. By examiningpatients with brain damage and through the use of functional magneticresonance imaging (fMRI), event-related potentials (ERP), and transcranialmagnetic stimulation (TMS), the neural underpinnings of stereotypicthinking have become a novel focus of inquiry. This focus, including itsbenefits and pitfalls, will form the main body of exploration in the presentreview. Beforehand, however, important theoretical advancements onstereotyping gained via traditional research methods will be outlined. Byconsidering behavioural and neuroscientific approaches of investigation sideby side, it will be possible to establish if these methods providecomplementary insights into the nature of stereotyping.
STEREOTYPES AND STEREOTYPING:A SOCIAL-COGNITIVE PERSPECTIVE
Among the diverse array of tactics that humans adopt to understand theirconspecifics, stereotyping has long been considered a pivotal and influentialstrategy. Almost a century ago the renowned American journalist WalterLippmann (1922) was the first to use the term stereotype in a psychologicalcontext. In his ground breaking text Public Opinion he observed that peopleoften relied on a ‘‘repertory of fixed impressions’’ (i.e., stereotypes) to makesense of an overwhelmingly complex and constantly changing world. Then32 years later the psychologist Gordon Allport narrowed the definition ofstereotypes by describing them as ‘‘exaggerated belief[s] associated with acategory’’ (1954, p. 191). Largely in line with his description, contemporaryconceptualisations define stereotypes as cognitive structures that provideknowledge, beliefs, and expectations about individuals based on their social
216 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

group membership (Hamilton & Sherman, 1994; Hamilton & Trolier, 1986;Kunda, 1999).
Stereotype content
As cognitive representations of social categories, stereotypes can target anumber of separate facets such as the personality, behaviour, and apparel ofothers (Deaux & Lewis, 1984; Jackson & Cash, 1985; Kawakami, Young, &Dovidio, 2002). According to popular stereotypes, for instance, women arecaring, enjoy shopping, and dress in pink; Jews are stingy, play the violin,and have large noses; and Germans are punctual, reserve deck chairs bythrowing their towels on them, and wear white socks with sandals. Despitetheir diversity, various types of stereotypes about social groups can often bedifferentiated along the common dimensions of warmth and competence(Cuddy et al., 2009). In this respect, Germans typically count as efficient butheartless (remember the selfish reserving of deckchairs!), while women (inparticular housewives) are seen as nurturing but ineffectual (outside thehouse, that is; Fiske, Cuddy, Glick, & Xu, 2002).
Stereotype acquisition
People seem to acquire stereotypic beliefs at a very early age. By the timethey turn 5 or 6 children display stereotyping across a wide range ofcategories such as sex, age, race, body weight, and attractiveness (for areview see Bigler & Liben, 2006). Pre-schoolers, for instance, tend to assumethat a physically aggressive story character is male rather than female (Giles& Heyman, 2005) and that overweight children possess more negativepersonality characteristics than other children (Musher-Eizenman, Holub,Miller, Goldstein, & Edwards-Leeper, 2004). Based on such observations ofearly acquisition and the prevalence of stereotype use throughoutadulthood, many theories have attempted to understand how peopleacquire stereotypic beliefs.
To date, stereotype acquisition is predominantly traced back to acompelling set of cognitive biases, motivational needs, and social learningmechanisms. Humans seem vulnerable, for example, to associate membersof minority groups with uncommon attributes and members of majoritygroups with frequent attributes even though no actual association betweenthe attributes and group membership may exist (Hamilton & Gifford, 1976;Mullen & Johnson, 1990; Sherman et al., 2009; for a detailed review seeMeiser & Hewstone, 2010). In addition we tend to see the behaviour ofothers as reflecting their inner dispositions (a phenomenon typically calledthe correspondence bias; Gilbert & Malone, 1995). Thus, when specificgroups of people are encountered in roles that call for a particular behaviour
STEREOTYPES AND STEREOTYPING 217
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

(e.g., women working as nurses who are required to show nurturingbehaviour), perceivers are likely to infer that persons enacting those roleshave corresponding dispositions—rather than that they display efforts ofrole fulfilment (e.g., Conway, Pizzamiglio, & Mount, 1996; Diekman &Eagly, 2000). Furthermore, the mere categorisation of people into groupsseems to influence person judgements. Members of the same group aregenerally considered more similar (i.e., within-group assimilation) andmembers of different groups as more different from each other (i.e.,between-group contrast) than when they are considered as mere individuals(Ford & Stangor, 1992; Wilder, 1978).
Besides cognitive biases, motivational needs can lead to members ofdifferent social groups being ascribed diverging characteristics and qualities.Based on the need to attain and maintain a positive self-image from thegroups to which they belong, humans attribute positive qualities to in-groups (i.e., social groups of which they are a member) and negativequalities to out-groups (i.e., social groups to which they do not belong;Bigler & Liben, 2006; Brewer, 1979). Similarly, perceivers are inclined toexplain positive behaviours of in-group members to advantageous disposi-tions and negative behaviours to situational factors, while positive andnegative acts of out-group members are explained in the opposite manner(Hewstone, 1990).
Finally, from a social-learning perspective, it has been argued thatstereotypes form an element of cultural heritage that is passed on from onegeneration to the next through communication, observation, and imitation(e.g., Castelli, De Dea & Nesdale, 2008; Ehrlich, 1973; Kashima, 2000; Perry& Bussey, 1979; Thompson, Judd, & Park, 2000). Not only have primarycaretakers and peers been identified as potent sources of a child’sinformation about social groups (e.g., Castelli, Carraro, Tomelleri, &Amari, 2007; Patchen, Davidson, Hofman, & Brown, 1977; Sinclar, Dunn,& Lowery, 2005), so have media portrayals. Analyses of broadcasting in theUS, for example, have revealed that women are presented predominantly indomestic or sexual roles, Latin Americans as family members engaged inconversations about crime, and Asian Americans as high-status profes-sionals (Mastro, 2008; Smith & Granados, 2008). The effects of suchportrayals can be pervasive. Extensive TV consumption appears to beassociated with enhanced stereotyping in both children and adults (Herrett-Skjellum & Allen, 1995; Oppliger, 2006).
Stereotyping others
In their attempt to better understand how, when, and why humans usestereotypes to make sense of each other, researchers have begun to delineatepivotal subcomponents of the stereotyping process (Devine, 1989; Fiske &
218 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Neuberg, 1990; Kunda & Sinclair, 1999; Sherman, Macrae, & Bodenhausen,2000; see Figure 1).
Person perception. The detection of conspecifics and the formation of acorresponding person percept often act as the starting point for stereotypingothers. Although work in social psychology tends to overlook theimportance of these perceptual operations (see Macrae & Quadflieg,2010), the ability to rapidly transform complex three-dimensional, dynamicvisual stimuli that are encountered under varying processing conditions(e.g., lighting, distance, posture, and viewpoint variations) into percepts ofpersons is nothing short of astonishing. Not only does it require theencoding of visual information relating to facial and bodily features (e.g.,mouth, nose, eyes, ears, chin, cheekbones, arms, legs etc.), in addition thespatial arrangement and configuration of these features is typicallyextracted. Thus, when seeing conspecifics, facial and bodily features arenot considered independently from each other but as one holistic entity(Weigelt, Koldewyn, & Doehrmann, 2010), as indexed by the face compositeeffect (i.e., when the top and bottom halves of a face comprise differentpeople, participants experience difficulty reporting the identity of the personportrayed in the top half of the face because the two parts of the compositestimulus are fused into a novel person; Hole, 1994; Kuefner, Jacques, Prieto,& Rossion, 2010) and by face/body inversion effects (i.e., when faces, bodies,and objects are inverted, recognition performance decreases more drama-tically for person than object stimuli due to inversion specificallycompromising the extraction of configural person information; Ramm,Cummins, & Slaughter, 2010; Reed, Stone, Bozova, & Tanaka, 2003; Reed,Stone, Grubb, & McGoldrick, 2006; Rossion & Gauthier, 2002; Yin, 1969).
Figure 1. Stereotyping at a glance.
STEREOTYPES AND STEREOTYPING 219
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Perceiving other people in a holistic manner is a skill typically acquiredand honed during early childhood (Crookes &McKone, 2009; Nelson, 2001;Pascalis & Kelly, 2009; Slaughter, Heron-Delaney, & Christie, 2011; Zieberet al., 2010). As a result, adult perceivers have an extraordinary level ofexpertise in the domain of person perception, allowing them not only toquickly spot and attend towards conspecifics (e.g., Bindemann, Scheepers,Ferguson, & Burton, 2010; Downing, Bray, Rogers, & Childs, 2004;Fletcher-Watson, Findlay, Leekam, & Benson, 2008; Hershler & Hochstein,2005; Langton, Law, Burton, & Schweinberger, 2008; Lewis & Edmonds,2005; Ro, Friggel, & Lavie, 2007; Theeuwes & Van der Stigchel, 2006), butalso to rapidly draw far-reaching social inferences about them. Typically amere glance at others is sufficient to evaluate not only their attractiveness,but also their emotional state, direction of attention, and even likelypersonality (for a review see Macrae & Quadflieg, 2010).
Given this plethora of possible social inferences on mere perception, oneof the most vexing problems in social cognition is how humans deal with theopportunity of multiple person construal. Put differently, based on which ofthe manifold available cues do perceivers spontaneously form impressions ofothers? To further illustrate this question, consider Figure 2. Both of thedepicted people can easily be construed along several dimensions of socialinterest (as spelled out by the descriptions in the white boxes framing thepicture). But is there reason to assume that different perceivers attend to andmake sense of others (such as the interacting couple) in a systematicmanner?
Figure 2. The puzzle of multiple person construal: Which cues will you use?
220 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Person categorisation. In response to this question, several influentialsocial psychological models (e.g., Allport, 1954; Brewer, 1988; Fiske &Neuberg, 1990) suggest that perceivers initially classify others according tosalient social categories (such as sex, race, and age). More specifically,according to these models, others are seen as random instances of genericsocial groups until a perceiver is willing to engage in individuated impressionformation (i.e., in making sense of others as unique entities)—either becausea specific target gains heightened relevance or because it consistently fails toconfirm category-based expectations (Brewer, 1988; Fiske & Neuberg, 1990;Quadflieg & Macrae, in press).
Naturally, a perceiver’s propensity to classify others upon mereperception depends on the ease with which categorical markers can beextracted from available visual cues. Critically, the ‘‘big three’’ categories—sex, race, and age—can instantly be gleaned from numerous facial andbodily features (e.g., body shape, facial configuration, skin colour, andtexture) and therefore dominate people’s encounters with others across awide range of social situations (Brewer, 1988, Fiske & Neuberg, 1990; VanKnippenberg & Dijksterhuis, 2000). Diverse additional group memberships,however, are often made visible by mere choice (e.g., believers dressedaccording to the requirements of their faith or sports fans decorated in thecolours of their team) or may even remain hidden until closer acquaintance(e.g., a person’s profession or political beliefs; but see Rule & Ambady,2010). Therefore person categorisation does not only result from the visualinspection of others, but can also follow from learning people’s names (thatmay reveal sex, race, or religious information) or from getting to know them(e.g., by going to lunch with Jim you may learn that he is a vegetarian).
To circumvent the various routes through which person categorisationcan be elicited, a favoured technique for exploring its consequencesexperimentally has been to present participants with triggering categorylabels (e.g., ‘‘Black’’ and ‘‘White’’, ‘‘Man’’ and ‘‘Woman’’, or ‘‘banker’’ and‘‘doctor’’). Unfortunately, such an approach often obscures naturallyoccurring complexities. Evidence indicates, for instance, that personcategorisation is a dynamic process in which for ambiguous targets (suchas men with long hair or women with short hair) contradicting groupmemberships can be activated simultaneously; (Freeman, Ambady, & Rule,2008; Freeman, Pauker, Apfelbaum, & Ambady, 2010; Macrae & Martin,2007; Martin & Macrae, 2007). Moreover, people usually belong to multiplerather than a single social category (e.g., a woman could be a youngCaucasian woman working as a nurse or an old Asian woman working as apolice officer; see also Remedios, Chasteen, Rule, & Plaks, 2011). Not toosurprisingly, perceivers find some conjunctions of categories easier tointegrate than others (e.g., Hutter, Crisp, Humphreys, Waters, & Moffitt,2009). Such observations indicate that the use of category labels rarely
STEREOTYPES AND STEREOTYPING 221
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

captures the richness of perceptual and/or cognitive operations contributingtowards person categorisation as occurring in daily life.
Stereotype activation. It is frequently assumed that once another personhas been categorised according to his or her group membership(s)stereotypic beliefs are automatically activated. Hence in the social-psychological literature the term person categorisation is sometimes usedinterchangeably with the term stereotyping (e.g., Stangor, 2000; Yzerbyt &Corneille, 2005). Strictly speaking though, the mental processes contributingtowards person categorisation and stereotyping are not the same (Shermanet al., 2000). Whereas person categorisation denotes the classification ofothers according to salient categorical markers, stereotyping goes beyondmere classification by allowing perceivers to hold expectations about thepersonality and/or behaviour of others. As Allport used to put it (1954, p.191), ‘‘a stereotype is not identical with a category; it is rather a fixed ideathat accompanies the category’’. Therefore, ‘‘categorisation is a necessarybut not sufficient condition for the occurrence of stereotyping’’ (Lepore &Brown, 1999, p. 145).
The assumption that stereotypes are inevitably activated following personcategorisation rests on the idea that stereotypic attributes are inherentlylinked with a category in a conceptual framework after frequent andconsistent co-occurrence (Fazio, Sanbonmatsu, Powell, & Kardes, 1986;Stephan & Stephan, 1993). Upon category activation, excitation is thoughtto spread automatically through the pertinent knowledge network todirectly linked characteristics. Contrary to this prediction, accumulatingdata suggest that the spreading of activation from a category representationto associated knowledge is constrained, and that processes underlying targetcategorisation and stereotype activation must be distinguished (Bartholow,2010; Mason, Cloutier, & Macrae, 2006). Put differently, although targetsare often unambiguously and rapidly categorised as members of certaingroups, whether and to which extent they elicit stereotypes based on theirgroup memberships is influenced by a variety of factors.
For instance, the extent of stereotype activation following personcategorisation critically depends on how typical for a certain social groupperceivers deem a target to be (Barden, Maddux, Petty, & Brewer, 2004;Livingston & Brewer, 2002; Locke, Macrae, & Eaton, 2005; Maddox, 2004).Faces displaying marked Afrocentric features have been shown to elicitstronger activation of the African-American stereotype than those with less-prototypical features (e.g., Blair, Judd, Saddler, & Jenkins, 2002). Capturinga similar effect, when clothing assigns stereotype-typical or atypical rolestowards a black individual (i.e., when a target is dressed as a criminal or alawyer in prison) the expected pattern of racial stereotyping can even bereversed (Barden et al., 2004). But not only does the physical typicality of
222 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

the cue provider modulate stereotype activation, so too can the circum-stances of an encounter. For example, viewing an Asian woman performinga typical female activity (e.g., applying cosmetics) or a typical Asian activity(e.g., eating with chopsticks) is sufficient to prompt either the activation ofsex or race stereotypes (Macrae, Bodenhausen, & Milne, 1995). In a similarvein, stereotype activation is amplified or reduced when a target isencountered in an expected or unexpected setting (e.g., when a black personis encountered at a street corner vs outside a church; see Wittenbrink, Judd,& Park, 2001a). In combination, these studies tell an emerging story.Whether a person looking young, black and male will elicit the activation ofstereotypic beliefs along any (or all) of those dimensions depends on hisunique appearance and the situation in question.
Does it also depend on the perceiver? It has sometimes been argued thatdue to the cultural transmission of stereotypes, all individuals within asociety are equally susceptible to learn about them. Indeed, it has beenshown that regardless of personal opinions, individuals high and low inprejudice tend to be equally knowledgeable about major stereotypes in theirculture (Devine, 1989; Devine & Elliot, 1995; Lepore & Brown, 1997;Moskowitz, Gollwitzer, Wasel, & Schaal, 1999). However, further dataindicate that egalitarian compared to prejudiced individuals are less likely toshow stereotype activation when perceiving and/or categorising others(Consalkorale, Sherman, Allen, Klauer, & Amodio, 2011; Johns, Cullum,Smith, & Freng, 2008; Kawakami, Dion, & Dovidio, 1998; Lepore &Brown, 1997; Moskowitz et al., 1999; Wittenbrink et al., 1997). Intriguingly,even when they do activate stereotypes, people with egalitarian attitudesseem less likely to apply these beliefs when engaging in person under-standing (Devine, 1989; Sherman, Stroessner, Conrey, & Azam, 2005).
Stereotype application. By examining the effects of personal attitudes onstereotyping, researchers have observed another important aspect of thestereotyping process—the activation of stereotypic beliefs does notnecessarily translate into their application. Ultimately people seem to makeuse of stereotypic beliefs when they can profit from this strategy. Throughstreamlining which information about others is sought out, attended to,interpreted, and remembered (Biernat, 2003; Biernat & Manis, 1994;Cameron & Trope, 2004; Darley & Gross, 1983; Duncan, 1976; Harnad,2005; Jussim & Fleming, 1996; Kunda & Sherman-Williams, 1993; Macrae& Bodenhausen, 2000; Sagar & Schofield, 1980; Trope & Thompson, 1997),for instance, stereotypes can save perceivers the effort of putting limitedprocessing resources into individuated impression formation (Allport, 1954;Brewer, 1988; Fiske & Neuberg, 1990).
Without doubt, stereotypes enhance the (apparent) predictability of asituation and reduce uncertainty in social interactions, especially when other
STEREOTYPES AND STEREOTYPING 223
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

sources of person knowledge are absent, ambiguous, or difficult to acquire(e.g., Allport, 1954; Kruglanski, 1989; Mendes, Blascovich, Hunter, Lickel,& Jost, 2007; Van Knippenberg & Dijksterhuis, 2000). The use ofstereotypes can also simplify communicative processes. By referring to aperson as ‘‘gay’’, ‘‘feminist’’, or ‘‘right-wing’’ a speaker implies anabundance of information and meaning likely to be easily understood bya listener of the same cultural background (Maass & Arcuri, 1996). Giventhese economising effects, stereotyping has been characterised as a cognitivetool that allows perceivers ‘‘to understand new and unique individuals interms of old and general beliefs’’ (Gilbert & Hixon, 1991, p. 509).
Besides providing an economic strategy to navigate one’s social world,stereotyping can also serve people’s motivational needs. Judging others interms of stereotypes has been found to help regulate and boost one’s self-esteem (Fein & Spencer, 1997; Sinclair & Kunda, 1999, 2000). Whenparticipants received feedback from an outgroup member, for instance, theywere more likely to activate the negative stereotype about the group theperson belonged to when the feedback was disadvantageous rather thanadvantageous (Sinclair & Kunda, 1999, 2000). In other words, stereotypeuse can be driven by a perceiver’s motivational need to disregard others andto hold up a positive self-image. Furthermore, stereotypic beliefs can beutilised to justify the distribution of resources between members of differentsocial groups (Hoffman & Hurst, 1990; Jost & Banaji, 1994; Jost & Kay,2005; Kay, Jost, & Young, 2005; Yzerbyt, Rocher, & Schadron, 1997).Because people want to believe in a just world (Hafer & Begue, 2005),stereotypes can function to legitimise inequality by ascribing differentweaknesses and strengths to members of advantaged and disadvantagedgroups. Complementary gender stereotypes describing men as agentic andwomen as communal, for example, can serve to maintain support for apatriarchal society (Glick & Fiske, 2001).
Although stereotypic beliefs are represented within the mind of theindividual person, they frequently reflect shared cultural knowledge(Haslam, Turner, Oakes, McGarty, & Reynolds, 1997). Due to thiscollective nature, people often assume social permission to express and acton stereotypes, thus reinforcing their own and other people’s biases (Fiske,2005; Sechrist & Stangor, 2001). As a result, the more stereotypes areconsensually held, the more they impact entire groups of people in the sameway (Stangor & Schaller, 1996). The societal consequences of stereotypeapplication can be intrusive and pernicious. Individuals of stereotypedgroups face differential treatment and discrimination in many public settingssuch as schools, universities, courtrooms, and at the workplace (e.g.,Agerstrom & Rooth, 2011; Blair, Judd, & Chapleau, 2004; Fiske & Lee,2008; Martin & Thompson, 2002; Zirkel, 2004). In addition, stereotypicbeliefs create a social climate in which targeted individuals feel burdened
224 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

and hindered by the predictable expectations of others (e.g., Appel,Kronberger, & Aronson, 2011; Aronson & Inzlicht, 2004; Crocker, Voelkl,Testa, & Major, 1991; Shapiro, 2011; Steele, 1997; Walton & Cohen, 2007).
OPEN QUESTIONS
The stereotyping process is often described in a domino-like fashion. Uponthe perception of a target, social categorisation is expected to occur, whichin turn activates stereotypic knowledge, that is then used to evaluate, judge,or predict a person’s personality and/or behaviour. While it is increasinglyacknowledged that this cascade of events does not necessarily unfoldautomatically, the exact factors influencing whether one stage of the processtriggers the next remains a matter of empirical investigation. In addition, theobservation that person categorisation can result from various types ofinformation (e.g., through merely perceiving others or through learninginformation about them) raises the question of whether these differentchannels lead to unique downstream consequences in the stereotypingcascade.
Along similar lines, it must be asked how different types of stereotypeacquisition (through experience, observation, instruction etc.) may shape theactivation and application of stereotypic beliefs. Furthermore, the matter ofwhether stereotyping unfolds in a diverging fashion for different types ofsocial categories, and different conjunctions of categories deserves closerscientific attention. Finally, differences in the stereotyping cascade depend-ing on whether stereotypic beliefs are evoked implicitly (i.e., without theperceiver’s introspective awareness) or explicitly (i.e., in a manner that couldbe consciously detected and reported) remain a matter of debate (Amodio &Mendoza, 2010). Although such topics of inquiry may appear of merescholarly importance at first glance, resolving them will speak towardsimportant issues of stereotype prevention and intervention. If, for instance,the mere categorisation of others were to automatically trigger stereotypeactivation and application, potential intervention programmes would bewell advised to focus particularly on aspects of social categorisation (e.g.,Bigler & Liben, 2007). If, however, multiple factors must come together forstereotyping to occur (such as a prejudiced attitude, a motivational need,and/or a situation-specific benefit), approaches considering those additionalfactors should enhance prevention and intervention success (e.g., Paluck &Green, 2009).
Aside from elucidating how subcomponents of the stereotyping processwork together in a feedforward manner, recent evidence has also emphasisedthe need to consider potential feedback connections. According to initialdata, for instance, social categorisation influences fundamental processes ofperson perception. Thus, when racially ambiguous faces are expected to
STEREOTYPES AND STEREOTYPING 225
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

belong to a group of African Americans, participants judge skin tone to bedarker than when the identical faces are understood to be Caucasian (Levin& Banaji, 2006; see also MacLin & Malpass, 2003). Along similar lines,ambiguous gender faces are encoded as more masculine or feminine in theirappearance when an associated name informs the perceiver that the personis male or female, respectively (Huart, Corneille, & Becquart, 2005). In otherwords, the expectation to see members of different groups induces anenhanced perception of targets in terms of their assigned group membership.
Further evidence indicates that social categorisation can even alterprocesses of person individuation. It is well known that healthy adults areusually very accurate at differentiating between distinct individual faces(e.g., Bahrick, Bahrick, & Wittlinger, 1975; Bruce & Young, 1998).However, this ability is typically reduced for people belonging to socialoutgroups compared to ingroups (Anastasi & Rhodes, 2005; Kuefner,Maachi Cassia, Picozzi, & Bricolo, 2008; Meissner & Brigham, 2001).Perceivers also show a categorisation advantage for outgroup faces suchthat other-race faces, for example, are categorised faster according to theirrace than own-race faces (e.g., Levin, 1996; Valentine & Endo, 1992). Suchdata support the idea of reduced individuation and enhanced categoricalprocessing of outgroup faces—a perceptual mechanism that may eventuallysupport the construal of outgroup members as ‘‘all alike’’ (an occurrencealso known as the outgroup homogeneity effect; see Boldry, Gaertner, &Quinn, 2007; Park & Rothbart, 1982).
Despite the fact that there is little disagreement about the existence ofoutgroup-related individuation deficits, their underlying mechanisms arestill poorly understood. According to perceptual-expertise accounts (e.g.,Tanaka, & Pierce, 2009; Valentine, 1991), perceivers hone their faceperception skills based on experience. Due to this tuning, the encoding ofunusual faces (i.e., faces that diverge from previous experience with fellowingroup members) poses a perceptual challenge. In contrast, according tosocial-motivational accounts, recognition differences are not elicited byperceptual difficulties alone but arise also due to assigning a person to anoutgroup. As a result of this categorisation, it is argued that the motivationto process a face in depth is reduced and the search for unique, individuatingcharacteristics is diminished (e.g., Hugenberg, Miller, & Claypool, 2007;Levin, 2000).
In summary, these data illustrate that person perception not onlydetermines person categorisation, but that person categorisation also shapesperson perception. Along similar lines, it could be expected that stereotypeactivation influences person categorisation (e.g., seeing someone reversingbadly into a parking spot should facilitate determining the driver’s sexrather than his race), and that stereotype application facilitates stereotypeactivation (e.g., holding the door for a woman may perpetuate thinking of
226 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

her in stereotypic terms). Intriguingly, collecting empirical data on feedbackloops in the stereotyping cascade may not only help us to comprehend thephenomenon in its entire complexity, but also to enhance our comprehen-sion of why stereotyping forms such a resistant and persuasive strategy ofperson understanding.
Last but not least, an ongoing issue of dispute arises from the definitionof stereotypes as mere cognitive structures. As such, stereotypic beliefs havetraditionally been differentiated from prejudice, which signifies feelings oflike or dislike, anger, fear, disgust, discomfort, or hatred towards membersof certain social groups (Allport, 1954; Eagly & Diekman, 2005). However,it has repeatedly been questioned whether representations of socialcategories can really be separated into their cognitive and evaluativecomponents. An alternative view suggests that affect forms a centralbuilding block of the stereotype itself (e.g., Czopp & Monteith, 2006;Mackie, Hamilton, Susskind, & Rosselli, 1996; Stangor & Leary, 2006;Wilson, Lindsey, & Schooler, 2000). Supporting this idea it has beenshown that among white Americans being black is not only stereo-typically associated with violence but also with feelings of threat anddislike (e.g., Wittenbrink, Judd, & Park, 2001b). As a result of suchobservations some researchers have argued that ‘‘it is difficult to conceiveof a stereotype of a group that is totally devoid of affect, and therefore tothink of affect as separate from, but merely associated with, the cognitiveelements seems implausible’’ (Mackie et al., 1996, p.52). Unfortunately atthis point the issue remains largely a question of plausibility due ascarcity of studies targeting the exact interplay of evaluative and cognitiveelements during stereotyping (for a noteworthy exception see Amodio &Devine, 2006).
Albeit this list of future issues of inquiry is far from complete, it showsthat traditional research methods (i.e., the use of response latencies, self-report measures, recall and recognition scores and so on) have left plenty ofroom for further investigation. At the same time it goes without saying thatinsights provided by behavioural methods have fundamentally advanced ourunderstanding of stereotypes and stereotyping. Whether this can be claimedfor studies using neuroscientific methods, and whether the use of ERPs,fMRI, and TMS has helped to address any previously unresolved issues, willbe discussed in the next sections of this chapter.
STEREOTYPES AND STEREOTYPING: A SOCIAL-NEUROSCIENCE PERSPECTIVE
Judging people on the basis of cultural stereotypes is a ubiquitous facet ofdaily life, yet little is known about how this fundamental inferential strategyis implemented in the brain. Consideration is now directed to this topic. For
STEREOTYPES AND STEREOTYPING 227
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

readers less familiar with neuroscientific methods, a brief description offrequently used techniques can be found in Table 1.
Stereotype content
Based on investigations targeting diverse types of concepts, it is currentlyassumed that the storage of knowledge in the brain is jointly constrained bydomain and sensory modality (Mahon & Caramazza, 2009; Martin, 2007).The idea of domain specificity is largely derived from experiments withbrain-lesioned patients who can display surprising conceptual impairments(for a review see Mahon & Caramazza, 2009). Patients with deficits foranimals, for instance, do not only fail to name animals from pictures, butare also unable to answer questions about them (e.g., ‘‘Does a whale havelegs?’’). At the same time, these patients are unimpaired at naming oranswering questions about objects or entities that do not belong to theimpaired category. According to this domain-specific approach of knowl-edge representation, it has been claimed that domain-specific constraintsexist for categories of evolutionary relevance such as animals, fruits/vegetables, tools, and also conspecifics.
TABLE 1Frequently used neuroscientific methods at a glance
Method Description Strength
Functional Magnetic
Resonance Imaging
(fMRI)
The non-invasive
measurement of brain
activity based on
recording oxygen levels
in the brain’s blood
vessels.
fMRI allows exploration of
which brain regions are
active during specific
mental operations.
Event-Related Potentials
(ERPs)
The non-invasive recording
of electrical brain activity
following the
presentation of a
stimulus by placing
electrodes on the scalp.
ERPs allow exploration of
rapid changes in brain
activity (within the
millisecond range)
related to different
mental operations.
Transcranial Magnetic
Stimulation (TMS)
The stimulation of cells in
the brain via rapidly
changing magnetic fields.
Depending on the kind
of stimulation an
increased or decreased
excitability of cells can be
evoked.
Unlike fMRI and ERPs,
which are correlational
techniques, TMS allows
exploration of the
causality and necessity of
brain regions for specific
mental operations.
228 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Although conspecifics may form a separate domain of knowledge, thisinference has so far been based on data from patients who failed to retrieveautobiographical knowledge about familiar others (e.g., Ellis, Young, &Critchley, 1989; Miceli, Capasso, Daniele, Esposito, Magarelli, & Tomaiuo-lo, 2000). Thus it remains uncertain whether generalised person knowledge,such as stereotypic beliefs (e.g., Do men wear skirts?) may recruit specialisedneural networks that can selectively be impaired by brain damage. Studieson the topic are rare. Recent fMRI data indicate that the retrieval ofstereotypic knowledge (compared to other types of conceptual knowledge)activates a set of brain regions considered key players in social cognition,such as the medial prefrontal cortex (MPFC), the posterior cingulate andthe anterior temporal lobe (Contreras, Banaji, & Mitchell, in press).
However, preliminary evidence from patients with damage to the MPFC,the anterior temporal lobe, or the amygdala, has revealed neither diminishedrapid gender- or race-based evaluative associations as measured with theimplicit association test (IAT), nor reduced deliberate gender-stereotypic orrace-biased judgements as measured with the Attitudes toward WomenScale (AWS) or the Modern Racism Scale(MRS) (Gozzi, Raymont,Solomon, Koenigs, & Grafman, 2009; Phelps, Cannistraci, & Cunningham,2003). Apart from the AWS, however, these measures tap more stronglyinto prejudice than stereotypes, and are therefore not directly informativeregarding the effects of brain damage on the representation of stereotypicknowledge. To elucidate the issue in its full scope, experiments using morediverse ways of probing stereotypic knowledge and studies covering a widerset of brain lesions are needed. Based on previous patient research on thetopic of moral reasoning, it may also be advisable to differentiate whetherbrain-damaged patients have preserved stereotypic knowledge but fail toapply this knowledge in personal matters or during social interactions(Damasio, Grabowski, Frank, Galaburda, & Damasio, 1994; Mah, Arnold,& Grafman, 2005; Saver & Damasio, 1991).
Importantly, additional data suggest that the organisation of conceptualknowledge may be constrained not only by domain but also by sensorymodality. According to this account, conceptual knowledge resides in thebrain’s model systems for perception, action, and introspection. In supportof this theory different concepts are represented by distinct neural networksaccording to their salient properties. For instance, whereas thinking aboutanimals activates brain regions in the ventral visual stream that encodevisual features such as shape and colour, thoughts about tools elicitactivation in motor systems related to object use (e.g., Chao, Haxby, &Martin, 1999; Hauk, Davis, Kherif, & Pulvermuller, 2008; Marques,Canessa, Siri, Catricala, & Cappa, 2008). Given that the content ofstereotypes is multifaceted (i.e., constituted by several different facets whichalso differ according to their salient properties), it can be argued that
STEREOTYPES AND STEREOTYPING 229
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

distributed neural networks may contribute towards different facets ofstereotyping.
As a case in point, consider the various forms that gender stereotypingcan take. Stereotyped judgements about the sexes typically tap knowledgepertaining to the apparel (e.g., women wear skirts, men wear neck ties),preferred activities (e.g., women bake cakes, men play poker), and likelypersonalities (e.g., men are aggressive; women are emotional) of women andmen. Based on evidence that conceptual knowledge is represented indistributed networks of modality-specific cortical areas, this then suggeststhat apparel, action, and personality stereotypes should also elicit activity inpartially distinct neural networks. More specifically, whereas apparelstereotypes (i.e., ‘‘visual form’’ knowledge) should be accompanied byactivity in visual association cortices located in the ventral occipital andtemporal region of the brain (e.g., Ishai, Ungerleider, Martin, & Haxby,2000; O’Craven & Kanwisher, 2000), action stereotypes (i.e., ‘‘action’’knowledge) should yield activity in regions to be found within the posteriortemporal and parietal cortices which are dedicated towards the representa-tion of motor knowledge (e.g., Assmus, Giesing, Weiss, & Fink, 2007; Chaoet al., 1999; Kable, Kan, Wilson, Thompson-Schill, & Chatterjee, 2005).Additionally, personality stereotypes (i.e., ‘‘trait’’ knowledge) should beassociated with activation in regions known to support the representation ofmore abstract social attributes such as the anterior superior temporal gyrus(aSTG; e.g., Zahn et al., 2007).
Thus the neural implementation of stereotypic person knowledge may bewidely distributed in the brain, with systematically different neural networksengaged depending on the specific content of stereotypic knowledge beingprobed. To date this important theoretical possibility awaits exploration.Initial support for the idea comes from one of our own studies. By askingpeople to report the likely actor (man or woman) of a series of everydayactivities (e.g., mowing the lawn, watching talk shows, taking photographs) weprobed the representation of action-related gender stereotypes. Comparedto actions that participants judged as equally likely to be undertaken byboth genders, those considered gender-stereotypic elicited enhanced activa-tion in left hemispheric regions known to subserve the representation ofaction knowledge (such as the posterior middle temporal gyrus and thesupramarginal gyrus, two gyri located in the posterior temporal and parietalcortex; Quadflieg et al., 2009).
Stereotype acquisition
To the best of our knowledge, neuroscientific methods have not yet beenused to study aspects of stereotype acquisition. There are, however,intriguing data indicating that the complex network of brain regions
230 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

involved in understanding and interacting with social agents continues todevelop until late into adolescence (Burnett, Sebastian, Kadosh, &Blakemore, 2011). If and how such developmental changes determineaspects of stereotype acquisition (as well as stereotype activation andapplication) poses an exciting avenue for future investigations.
Stereotyping others
While social psychologists have made noticeable progress at elucidating allaspects of the stereotyping process, neuroscientific studies have focusedalmost exclusively on aspects of person perception and person categorisa-tion. As a result of this emphasis, the neural structures that implement thedefining feature of the stereotyping process, the activation and use ofculturally shared knowledge about prominent social groups are still poorlyunderstood.
Person perception. Recordings of ERPs suggest that human faces andbodies are reliably discriminated from other object categories at an earlystage of visual processing. Relative to a variety of other objects, faces elicitan enhanced negative potential at around 170 ms after the onset of astimulus. This so-called N170 response has been detected using diverse facialstimuli such as schematic, sketched, painted, drawn, and photographedfaces, prompting researchers to conclude that the signal is associated withdetecting common structural properties of facial stimuli rather than aresponse to low-level visual properties of faces (for a review see Rossion &Jacques, 2008). This conclusion receives further support from studiesdemonstrating that the N170 is not abolished when elements of the facialstimulus are rearranged, removed, or isolated, as long as a stimulus remainsto be recognisable as a face (Bentin et al., 1996; George et al., 1996; Itier &Taylor, 2002; Sagiv & Bentin, 2001).
In contrast, the N170 response does disappear when a target is no longerconsidered a face—e.g., when eyes or the mouth are presented in isolation(Bentin et al., 1996), when noise in the frequency bands critical for faceperception is superimposed on the stimulus (Tanskanen et al., 2005), orwhen so-called mooney or arcimboldo faces are presented upside down, anarrangement that typically prevents perceiver to interpret these stimuli asfaces (George et al., 2005). In a similar vein, photographs and point-lightanimations depicting human bodies, silhouettes, and stick figures all seem toelicit a body-specific negative component peaking at 190 ms after stimulusonset compared to pictures displaying faces or objects (Hirai, Fukushima, &Hiraki, 2003; Jokisch, Daum, Suchan, & Troje, 2005; Pourtois, Peelen,Spinelli, Seeck, & Vuilleumier, 2007; Taylor, Roberts, Downing, & Thierry,2010). These findings suggest that the brain discriminates the unique
STEREOTYPES AND STEREOTYPING 231
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

properties of human faces and bodies rapidly from various kinds of visualinput.
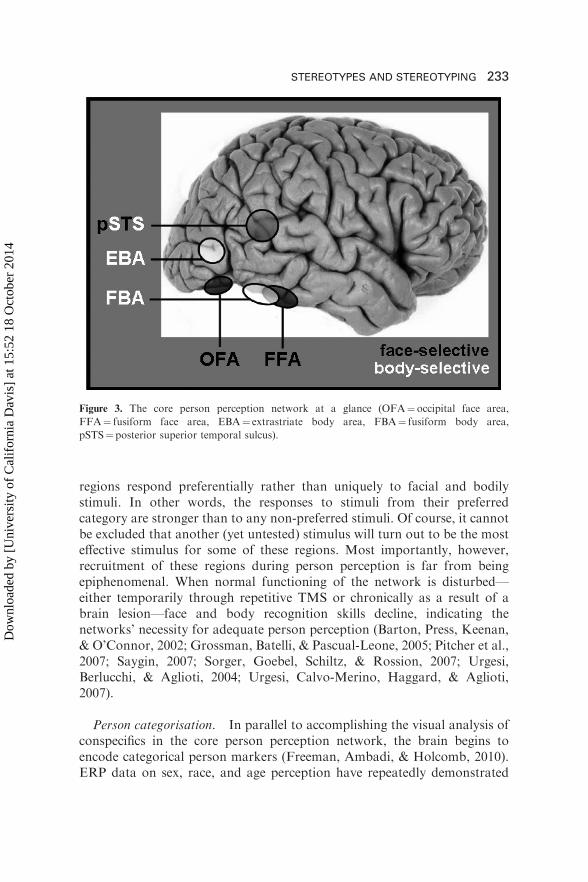
In line with this observation, several brain regions have been identifiedthat are specialised for extracting person-related visual information from theenvironment (Bracci, Ietswaart, Peelen, & Cavina-Pratesi, 2010; Kanwisher,McDermott, & Chun, 1997; Orlov, Makin, & Zohary, 2010; Weiner & Grill-Spector, 2010). Most reliably, two face-specific areas, the occipital face area(OFA) in the inferior occipital sulcus as well as the fusiform face area (FFA)in the middle fusiform gyrus are recruited when participants view humanfaces compared to other common objects and nonsense stimuli (for reviewssee Haxby, Hoffmann, & Gobbini, 2000; Yovel & Kanwisher, 2005).Similarly, two body-specific areas, the extrastriate body area (EBA) in closeproximity but slightly superior to the OFA located in the lateraloccipitotemporal cortex and the fusiform body area (FBA) situated adjacentto and overlapping with the FFA, selectively respond toward human bodies(for reviews see Downing & Peelen, 2011; Peelen & Downing, 2007).Neuroimaging data further indicate that these regions play differentfunctional roles. Whereas the OFA and the EBA encode person input byprocessing faces and bodies on the basis of their parts, FFA and the FBAanalyse the configuration and holistic appearance of these parts (Haxby,Ungerleider, Clark, Schouten, Hoffman, & Martin, 1999; Kanwisher &Yovel, 2006; Liu, Harris, & Kanwisher, 2010; Nichols, Betts, & Wilson,2010; Pitcher, Walsh, & Duchaine, 2011; Pitcher, Walsh, Yovel, &Duchaine, 2007; Taylor, Wiggett, & Downing, 2007; Yovel & Kanwisher,2005).
Additionally, perceiving movements of the eyes, the mouth, or the bodyhas consistently been found to increase activation in the posterior superiortemporal sulcus (pSTS; Campbell et al., 2001; Giese & Poggio, 2003;Grossman & Blake, 2002; Hoffman & Haxby, 2000; Lee et al., 2010;Michels, Lappe, & Vaina, 2005; Pitcher, Dilks, Saxe, Triantafyllou, &Kanwisher, 2011; Saxe, Xiao, Kovacs, Perrett, & Kanwisher, 2004). Thisregion seems to integrate form and motion information emanating from aperson’s face and body (Blake & Shiffrar, 2007; Thompson, Hardee,Panayiotou, Crewther, & Puce, 2007). Thus, by complementing the analysisobtained in the OFA, FFA, EBA, and FBA, the pSTS enables thegeneration of dynamic rather than mere static person percepts. Together thisset of brain regions (OFA, FFA, FBA, EBA, pSTS; see Figure 3) has beentermed the core person perception network and is assumed to undertake theperceptual analysis of an individual’s facial and bodily appearance (Haxbyet al., 2000; Macrae & Quadflieg, 2010).
But does this mean that these brain regions are exclusively involved in theperceptual analysis of faces and bodies (Kanwisher, 2010; Peelen &Caramazza, 2010)? Most likely not. The available data suggest that these
232 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014
regions respond preferentially rather than uniquely to facial and bodilystimuli. In other words, the responses to stimuli from their preferredcategory are stronger than to any non-preferred stimuli. Of course, it cannotbe excluded that another (yet untested) stimulus will turn out to be the mosteffective stimulus for some of these regions. Most importantly, however,recruitment of these regions during person perception is far from beingepiphenomenal. When normal functioning of the network is disturbed—either temporarily through repetitive TMS or chronically as a result of abrain lesion—face and body recognition skills decline, indicating thenetworks’ necessity for adequate person perception (Barton, Press, Keenan,& O’Connor, 2002; Grossman, Batelli, & Pascual-Leone, 2005; Pitcher et al.,2007; Saygin, 2007; Sorger, Goebel, Schiltz, & Rossion, 2007; Urgesi,Berlucchi, & Aglioti, 2004; Urgesi, Calvo-Merino, Haggard, & Aglioti,2007).
Person categorisation. In parallel to accomplishing the visual analysis ofconspecifics in the core person perception network, the brain begins toencode categorical person markers (Freeman, Ambadi, & Holcomb, 2010).ERP data on sex, race, and age perception have repeatedly demonstrated
Figure 3. The core person perception network at a glance (OFA¼ occipital face area,
FFA¼ fusiform face area, EBA¼ extrastriate body area, FBA¼ fusiform body area,
pSTS¼posterior superior temporal sulcus).
STEREOTYPES AND STEREOTYPING 233
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

that the extraction of these categories from faces can occur as quickly as150–200 ms after face onset (Brebner, Krigolson, Handy, Quadflieg, &Turk, 2011; Dickter & Bartholow, 2007; Ito, Thompson, & Cacioppo, 2004;Mouchetant-Rostaing & Giard, 2003; Walker, Silvert, Hewstone, & Nobre,2008). However, whether the face-sensitive N170 component is specificallymodulated by categorical markers remains an issue of debate. Most recently,a ‘‘release from adaptation’’ effect to face identity as typically observed inthe N170 amplitude has been found to be larger for racial ingroup thanoutgroup faces, indicating that to neural populations underlying the N170,outgroup faces largely ‘‘look alike’’ (Vizioli, Rousellet, & Caldara, 2010).
In addition some studies have reported increased negativity in N170amplitude towards racial outgroup relative to ingroup faces (Brebner et al.,2011; Herrmann et al., 2007; Stahl, Wiese, & Schweinberger, 2008; Walkeret al., 2008). Others, however, found such a difference only for inverted butnot for upright faces (Vizioli, Foreman, Rousselet, & Caldara, 2010), failedto observe any differences in this component (Caldara, Rossion, Bovet, &Hauert, 2004; Caldara, Thut, Servoir, Michel, Bovet, & Renault, 2003; He,Johnson, Dovidio, & McCarthy, 2009; James, Johnstone, & Hayward, 2001;Ofan, Rubin, & Amodio, 2011), or even detected the opposite pattern ofresults (i.e., decreased N170s for outgroup relative to ingroup faces; Ito &Urland, 2005).
These inconsistencies in the literature may reflect differences in theexperimental tasks used and corresponding differences in participants’temporary processing goals. As noted by Barthalow & Dickter (2011), forinstance, studies in which the N170 was larger to outgroup than ingroupfaces often required perceivers to focus on face identity, whereas studiesreporting other types of activity patterns asked participants to focus onother stimulus dimensions (e.g., detecting non-face stimuli). Under theassumptions that the N170 reflects holistic face encoding (e.g., Jacques &Rossion, 2009; Lewis & Edmonds, 2003) and that perceivers typicallyprocess outgroup faces in a less holistic manner (e.g., Hayward, Rhodes, &Schwaninger, 2008; Michel, Caldara, & Rossion, 2006; Michel, Rossion,Han, Chung, & Caldara, 2006; Tanaka, Kiefer, & Bukach, 2004), tasks thatrequired attention to identity may have selectively increased recruitment ofholistic face processing mechanisms to racial outgroup relative to ingroupfaces. It also needs to be considered that not all of the studies conductedcontrolled for low-level visual differences (e.g., luminance and contrast)across racial groups in the stimulus material. Furthermore, participants’racial attitudes have often been neglected during data analyses, anunfortunate circumstance given that the extent to which the N170component is amplified toward outgroup relative to ingroup facesdepends on the strength of a perceiver’s pro-ingroup bias (Ofan et al.,2011).
234 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Aside from the N170, another ERP component, the N200 has typicallyfound to be larger for (racial) ingroup compared to outgroup faces (Brebneret al., 2011; Dickter & Bartholow, 2007; Ito et al., 2004; Ito & Urland, 2005;James et al., 2001; Walker et al., 2008). This difference has been argued toreflect a bias in attention toward faces containing ingroup cues (Ito &Urland, 2003), and concomitant increases in the depth of encoding processes(Kubota & Ito, 2007). Such interpretations are supported by recentinvestigations demonstrating that a reduced amplitude in this componentduring face perception is linked to diminished outgroup face recognition at alater time (Brebner et al., 2011; Lucas, Chiao, & Paller, 2011; for alternativeexplanations see Barthalow & Dickter, 2011).
Complementing the collected ERP data, fMRI data indicate that thefusiform cortex plays a pivotal role during person categorisation (Cloutier,Turk, & Macrae, 2007; Freeman, Rule, Adams, & Ambady, 2010; Ng,Ciaramitaro, Anstis, Boynton, & Fine, 2006). In particular it has beenshown that fusiform activation (including activity in the FFA) is increasedduring the perception of ingroup compared to outgroup faces (Golby,Gabrieli, Chiao, & Eberhardt, 2001; Van Bavel, Packer, & Cunningham,2008; 2011). This observation is in line with previous behavioural researchaccording to which ingroup faces profit from enhanced individualisticprocessing (e.g., Michel et al., 2006; Tanaka et al., 2004). In support of thisidea it has also been observed that the difference in fusiform activationtowards ingroup and outgroup faces disappears when outgroup faces depictwell-known others whose unique facial characteristics are perceptuallyfamiliar (Kim et al., 2006).
Finally, perceiving others as members of generic social groups rather thanas unique individuals may rely particularly strongly on the left hemisphereof the brain (e.g., Rivera, Arms-Chavez, & Zarate, 2009; Zarate, Stoever,MacLin, & Arms-Chavez, 2008). In line with behavioural data, an earlypositron emission tomography (PET) study as well as a more recent fMRIinvestigation both revealed a predominantly left-hemispheric neuralresponse during sex categorisation compared to person individuation(Andreasen et al., 1996; Mason & Macrae, 2004). Due to a lack ofconverging localisations in these studies, however, further work is requiredto examine the lateralisation hypothesis. Alternatively, it has been claimedthat the perception of outgroup but not ingroup members may relyparticularly strongly on left hemispheric resources (Proverbio, Riva, Martin,& Zani, 2010).
Stereotype activation. Tentative attempts to study the neural correlatesof stereotype activation have often focused on the brain’s response towardsviolations of stereotypic expectancies. ERP data indicate that suchviolations can be captured in the N400 effect and the P600 effect. In one
STEREOTYPES AND STEREOTYPING 235
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

of the initial studies, for instance, participants were primed with categorylabels (‘‘women’’ or ‘‘men’’) that were followed by target words eitherconsistent or inconsistent with common gender stereotypes (e.g. ‘‘women’’ –‘‘nurturing’’ or ‘‘women’’ – ‘‘aggressive’’). The participants’ task was toindicate whether the shown word pairs matched or mismatched with regardto gender. Under these conditions it was found that the processing ofstereotype-inconsistent word pairs elicited not only slower response timesbut also a larger N400 effect (White, Crites, Taylor, & Corral, 2009).
Although this study was unable to disentangle the effect of stereotype-consistent and -inconsistent words pairs from the type of answer given (i.e.,stereotype-inconsistent pairs always required a mismatch response), asubsequent study using a similar approach and excluding this confoundreplicated the findings in the evaluative domain. In this study, Chineseparticipants were primed with category labels (urban vs migrant workers)followed by adjectives that participants had to judge according to valence(Wang et al., 2010). Response times to identify positive adjectives weresignificantly slower when they followed the label ‘‘migrant worker’’ (a low-status ostracised group in China) relative to ‘‘urban worker’’. Again, theeffect was accompanied by a larger N400 amplitude, supporting the ideathat this ERP component can reflect a category-based associative (i.e.,stereotypic or evaluative) mismatch.
The interpretation of both studies in concert requires caution given thatthe first study targeted the violation of stereotype-based associations,whereas the second one made use of prejudice-based associations. Underconsideration of previous insights by psycholinguists, however, such ageneralised effect does hardly come as a surprise. The N400 has been studiedextensively in the context of two paradigms (for a review see Lau, Phillips, &Poeppel, 2008): In the semantic priming paradigm word pairs that are eithersemantically related or unrelated are presented (e.g., ‘‘coffee-tea’’ vs ‘‘socks-tea’’), whereas in the semantic anomaly paradigm either a congruous orincongruous word is presented as a continuation of a preceding sentence(e.g., ‘‘I like my tea with milk and sugar/socks’’). In both paradigms thesemantically incongruent context elicits a N400 response of largeramplitude. Importantly, the effect is not modulated only by the degree ofanomaly but also by a reader’s expectations. Less-expected sentence endingselicit a larger N400 response than highly expected ones, even when bothendings are semantically congruent (e.g., ‘‘I like my tea with milk andhoney’’ would generate a larger N400 response than ‘‘I like my tea with milkand sugar’’; Kutas & Hillyard, 1984). Based on such findings, the N400effect has been considered an indicator of associative fit in semantic memory(Kutas & Federmeier, 2000). More specifically, a N400 reduction has beenargued to reflect facilitated access of semantic information (Lau, Almeida,Hines, & Poeppel, 2009). As such, in the realm of social cognition, the N400
236 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

could turn into an important and unobtrusive indicator of category-basedassociations.
In addition to the N400, violations of stereotypic expectations have beenfound to modulate a late positive ERP potential, the P600. Osterhout,Bersick, and McLaughlin (1997), for instance, presented participants withsentences containing gender-stereotypic occupations and pronouns that didor did not match the gender stereotypically implied by the occupation (e.g.,‘‘The nurse prepared herself for the operation.’’ vs ‘‘The nurse preparedhimself for the operation.’’). The violation of occupation-based stereotypicexpectancies in these sentences elicited a larger P600. A similar effect wasobserved when participants listened to male and female utterances that wereeither congruent or incongruent with a speaker’s gender (e.g., ‘‘I like to playsoccer’’ said by a male or a female voice). Again, incongruent informationtriggered an enhanced P600 effect (Lattner & Friederici, 2003). The P600 haspreviously been associated with the processing of grammatical incongruities(Hagoort, 2003). In domain-general terms the component has also beenlinked to the processing of anomalies that require the integration ofsurprising relations (Gouvea, Philips, Kazanina, & Poeppel, 2010). At thispoint, however, the exact process underlying the P600 during stereotypeviolations requires further investigation.
Stereotype application. Surprisingly, social neuroscientists interested inthe neural signature of stereotyping have overwhelmingly studied aspects ofstereotype control rather than stereotype application in the first place. Inlight of evidence demonstrating that brain regions such as the dorsolateralprefrontal cortex (DLPFC) and the anterior cingulate cortex (ACC) areinvolved in aspects of cognitive control (for a review see Mansouri, Tanaka,& Buckley, 2009), early on the field became interested in elucidating whetherthese regions also contribute to the inhibition or suppression of stereotypicbeliefs. Indeed, activity in both regions has been found to increase whentasks require participants to suppress rapid group-based associations (Beeret al., 2008; Knutson, Mah, Manyl, & Grafman, 2007). Using a deductivereasoning paradigm, it has also been shown that overcoming stereotypicthinking elicits increased activity in the DLPFC (De Neys, Vartanian, &Goel, 2008). Additionally, a TMS-based temporal inability to recruit theDLPFC has been linked to the enhanced expression of stereotypicassociations (Cattaneo, Mattavelli, Platania, & Papagno, 2011).
Evidence for the ACC’s role in the control of stereotyping has beenprovided by the so-called weapon identification task. This paradigm requiresparticipants to categorise pictures of either guns or tools following thepresentation of black and white faces (Payne, 2001). Utilising this task, itwas shown that the error-related negativity (ERN) wave, an ERP-component assumed to be generated by the anterior cingulate, is larger
STEREOTYPES AND STEREOTYPING 237
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

when tools are erroneously classified as guns following black relative towhite faces (Amodio et al., 2004; Amodio, Kubota, Harmon-Jones, &Devine, 2006). Put differently, responses that are race-biased (or gender-biased, see Ma, Shu, Wang, Dai, & Che, 2008) elicit a stronger ERN (i.e.,stronger activity in the anterior cingulate), most likely due to peoplemonitoring their performance and attempting to control their bias. Thus theamplitude of the ERN effect predicts better response control in the task andis most pronounced among participants who are internally motivated toavoid the expression of race bias (Amodio, Devine, & Harmon-Jones, 2008).
Intriguingly, some neuroimaging studies measuring differences in theincidental processing of unfamiliar own-race versus other-race faces havealso reported differential activity in the DLPFC and the ACC (Cunninghamet al., 2004; Richeson et al., 2003). Based on previous insights on theseregion’s functions, it has been argued that even the mere detection ofoutgroup members may elicit attempts to control automatically triggeredrace-related stereotypes or prejudice (see also Bartholow, 2010; Stanley,Phelps, & Banaji, 2008). Although this may be a valid interpretation, severalcrucial questions remain. What exactly is it that white participants try tosuppress or inhibit when they see a black face and why would they feelmotivated to do so in a mere perceptual task (see also Amodio, 2008, 2009)?These questions are particularly hard to answer given that it has not beenclarified yet whether the mere perceptual exposure to others is sufficient toelicit social categorisation (see Macrae, Quinn, Mason, & Quadflieg, 2005;Quinn & Macrae, 2005; Santos & Young, 2005)—not to mention stereotypeand/or prejudice activation. To clarify these issues, future neuroscientificstudies will need to provide evidence of spontaneous stereotype and/orprejudice control following mere person detection.
Further neuroimaging studies interested in stereotyping have focused onimpression formation for stereotype-violating individuals. People tend to beeasily stirred by others who defy prevailing stereotypes, such as femalemotor mechanics or accomplished African Americans (Hutter et al., 2009;Kernahan, Bartholow, & Bettencourt, 2000; Mendes et al., 2007).Presumably such atypical individuals receive additional attention becausestereotypic beliefs fail to explain their behaviour. Encounters withstereotype-discrepant persons have been found to trigger elaborativeprocesses that strive to resolve apparent categorical inconsistencies andenhance person memory (Crocker, Hannah, & Weber, 1983; Jackson,Sullivan, & Hodge, 1993; Macrae, Bodenhausen, Schloerscheidt, & Milne,1999). In line with these behavioural findings, fMRI data indicate that theexplicit attempt to form impressions of stereotype-violating targets enhancesactivity in the MPFC and the temporoparietal junction (Cloutier, Gabrieli,O’Young, & Ambady, 2011). Both regions are considered the brain’s keyplayers in social-cognitive functioning and typically display enhanced
238 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

activity whenever participants try to infer the mental states, intentions and/or personalities of others (see Adolphs, 2006; Amodio & Frith, 2006; Saxe,2006; van Overwalle, 2009).
In one of our own studies we were additionally interested in theperceptual consequences that can result from expectancy violations(Quadflieg et al., 2011). More specifically, we reasoned that stereotypicbeliefs may alter fundamental processes of person perception (see alsoQuadflieg & Rossion, in press), based on the assumption that perceptioncomprises an interaction between available stimulus input and the brain’spredictions regarding likely stimulus identities (Gilbert & Sigman, 2007;Gregory, 1997; Sterzer, Kleinschmidt, & Reese, 2009). To explore this issuewe asked participants to make judgements about men and women depictedin various stereotype-consistent and stereotype-inconsistent occupations.Specifically, they either reported the sex of each target (i.e., personcategorisation) or the colour of a dot (i.e., colour classification) that waslocated on the image. Goal orientation was manipulated in this way as priorwork has suggested that stereotype activation is impeded when non-socialprocessing goals guide target appraisal (Macrae, Bodenhausen, Milne,Thorn, & Castelli, 1997; Wheeler & Fiske, 2005).
Analyses of the resulting data revealed that completion of these two tasksrecruited fundamentally divergent neural substrates. While social judgementselicited enhanced activation in the MPFC, dot judgements specifically engagedlow-level visual areas dedicated towards colour processing. These findingssuggest that, depending on their temporary processing objectives, participantsconstrued targets either more strongly as meaningful social agents or as mereperceptual entities. Most importantly, only during trials that required the sex-categorisation of stereotype-inconsistent targets did we also observe increasedactivity in the DLPFC as well as in core person perception areas, such as theFFA, FBA, and EBA. Thus, in further support of previous work that foundDLPFC involvement when participants were required to refrain fromstereotypical thinking in reasoning or associative tasks (Beer et al., 2008; DeNeys et al., 2008; Knutson et al., 2007), we observed enhanced activity in thisregion when participants had to ignore stereotypic associations triggered byoccupational roles that conflicted with a target’s actual sex.
In addition, however, amplified activation was also found in the FFA,FBA, and EBA, signalling that forming a coherent person representationduring sex categorisation was more demanding when targets failed toconfirm prevailing gender. In line with this claim participants’ responsetimes during task performance showed a target typicality by taskinteraction, such that during sex categorisation response times were fasterwhen targets were displayed in stereotype-consistent than stereotype-inconsistent occupations, whereas during dot categorisation response timeswere comparable for both types of targets (see Table 2).
STEREOTYPES AND STEREOTYPING 239
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

For exploratory reasons we also asked participants to complete the Needto Evaluate Scale (NES; Jarvis & Petty, 1996) after scanning. This scale hasbeen designed to quantify how people differ in the likelihood and extent towhich they show spontaneous evaluative responding when processingdifferent types of objects, including other people (e.g., Federico, 2004;Tormala & Petty, 2001). To further examine the activity observed during sexjudgements, correlations between participants’ NES scores and the brainresponse in face- and body-selective brain regions (OFA, FFA, FBA, EBA)and the right DLPFC were examined. It was found that the strongerparticipants’ chronic need to evaluate, the greater the activity in the rightDLPFC, r(16)¼ .47, p5 .05 (see Figure 4, Panel A), the left FBA,r(16)¼ .71, p5 .05 (see Figure 4, Panel B), and the left EBA, r(16)¼ .56,p5 .05 (see Figure 4, Panel C) to stereotype-inconsistent than stereotype-consistent targets during sex categorisation trials. No such effects emergedduring colour judgements.
Why body perception areas in the right hemisphere were not impactedrequires further investigation. However, the data could support the argumentthat the left hemisphere plays a more prominent role when targets areconsidered as examples of generic social categories (e.g., men and women)rather than unique individuals (Rivera et al., 2009; Zarate et al., 2008). Moredefinitely, the study revealed that encountering targets that violate stereotypicbeliefs does not only increase neural activity related to cognitive control (i.e., inthe DLPFC) but also in areas dedicated toward person perception (see alsoSchreiber & Iacoboni, in press) and that this effect is modulated by a person’stemporary processing goal and chronic evaluative needs.
Finally, social neuroscientists have recently begun to look at the neuralimplementation of impression formation strategies shaped by stereotypicbeliefs. Many studies have previously shown that perceivers judge theidentical behaviour of others differently depending on the social groups towhich they belong (e.g., Biernat & Manis, 1994; Darley & Gross, 1983;Kunda & Sherman-Williams, 1993; Sagar & Schofield, 1980). A classic
TABLE 2Median response times in milliseconds as function of target type and processing goal
(Quadflieg et al., 2011)
Sex categorisation Dot categorisation
Stereotype-consistent target 675
(137)
681
(130)
Stereotype-inconsistent target 704
(145)
672
(137)
Standard deviations are given in parentheses. The data exclude all trials with incorrect
responses.
240 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

study, for instance, demonstrated that when white students watched aheated discussion between two men, which ended in one man shoving theother, the observed action was considered more aggressive when the shovewas delivered by a black man relative to a white man (Duncan, 1976).Neuroimaging data now indicate that perceivers may adopt differentstrategies to make sense of the behaviour of ingroup relative to outgroupmembers.
One important tactic through which people can understand and interpretthe behaviour of others seems to lie in the use of their own ‘‘motorknowledge’’ as a guiding framework. That is, incoming visual informationfrom others can be mapped onto one’s own motor representation of theaction in question (e.g., Decety & Grezes, 1999; Rizzolatti, Fogassi, &Gallese, 2001). Intriguingly, initial evidence suggests that perceivers aremore likely to show this type of motor resonance during action observationwhen targets belong to a common ingroup rather than an outgroup(Molnar-Szakacs, Wu, Robles, & Iacobini, 2007; Liew, Han, & Aziz-Zadeh,2011). This differential effect appears to be amplified by prejudiced attitudesand for particularly disliked groups (Gutsell & Inzlicht, 2010). Whether theextent of motor resonance is influenced not only by a person’s outgroupstatus per se but also by the group-specific stereotype is an empiricalquestion that requires further experimentation. It may be the case, however,that motor resonance is particularly low towards outgroups for whom thestereotype indicates a deviation in motoric abilities relative to the perceiver.A young Caucasian American banker, for instance, may show less resonanceto blacks (who are expected to be more athletic) or to the elderly (who areconsidered to be weaker and fragile) compared to Germans or taxi-driverswho do not usually elicit strong group-based expectations about divergingmotor abilities.
Figure 4. Relationship between brain activity obtained for stereotype-inconsistent4 stereotype-
consistent sex trials and participants’ chronic need to evaluate as measured with the Need to
Evaluate Scale (NES).
STEREOTYPES AND STEREOTYPING 241
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Additional research suggests that strategies used to make sense of othersmay also differ based on their category-related warmth-competence status(Fiske et al., 2002). In particular, members of social groups perceived as lowin both competence and warmth (e.g., homeless, drug addicts) tend to beassociated with a diminished neural response in the medial prefrontal cortex(MPFC; Harris & Fiske, 2006). As previously mentioned, enhanced MPFCactivation is typically observed whenever participants try to infer the mentalstates, intentions and/or personalities of others (see Adolphs, 2006; Amodio& Frith, 2006; Saxe, 2006; van Overwalle, 2009). Therefore it has beenargued that the lack of MPFC activation may signal a lack of the attempt toattribute mental states, intentions, or personalities to others—an act oftenreferred to as dehumanisation. In line with a dehumanisation account,MPFC recruitment towards low-warmth-low-competence targets appears toreturn when perceivers are experimentally forced to consider them asindividuals with unique likes and preferences (Harris & Fiske, 2007).
Along similar dehumanising lines, it has recently been observed that menviewing women (relative to other men) in revealing outfits activate theMPFC less, the stronger they hold hostile sexist attitudes (Cikara,Eberhardt, & Fiske, 2010). Such a pattern of results could be explainedby the fact that such men consider scantily dressed women more object- thenagent-like. Indeed, additional behavioural data have shown that men withhigher hostile sexism scores associated such women more quickly with first-person action verbs (e.g., ‘‘handle’’) and clothed women with third-personaction verbs (e.g., ‘‘handles’’) than the inverse, compared to their less-sexistpeers. In summary, these data suggest that reduced MPFC activity maysignal a systematic lack of mentalising about others. However, given that theMPFC comprises a large area of cortex and is involved in many diversesocial-cognitive tasks, this dehumanisation account awaits further replica-tion and refinement (see discussions in Harris & Fiske, 2007, 2009; Schreiber& Iacoboni, in press).
THE BENEFITS AND PITFALLS OF USING A SOCIALNEUROSCIENCE APPROACH TO ADDRESS ISSUES
OF STEREOTYPING
Whether and how neuroscientific findings can inform psychologicaltheorising and, in particular, social psychological reasoning has been anissue of controversy for more than 10 years (Kihlstrom, 2010; Kosslyn,1999; Quadflieg & Macrae, 2011; Sherman, 2010; Willingham & Dunn,2003). Usually, the debate centres on the question whether neuroscientificmethods really open an intriguing new window towards the workings of the(social) mind, or if they merely furnish an expensive but meaninglesscollection of brain pictures and graphs (Fodor, 1999; Uttal, 2003). Since
242 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

stereotypes and stereotyping have been topics of utmost importance in therealm of social-cognitive research for decades, their investigation withneuroscientific methods can be considered a well-suited example to judge theelucidating potential of such an approach. Therefore the followingparagraphs discuss a range of critical issues that are typically raised whenit comes to the evaluation of insights gained by the use of social-neuroscientific methods.
What is the object of investigation?
The constructs that can be localized in the brain are simply not the best level ofdescription for theories of many of the problems of interest to social psychologists’’.
Willingham & Dunn, 2003, p. 269)
It is tautological to state that social neuroscientists decide to approachtopics of interest from a neural perspective. They dare to do so based on theassumption that complex social phenomena may be better understood byexploring them from multiple levels of analysis. In particular, socialneuroscientists are fascinated by the potential of scientific insights that couldemerge from the combination of social and biological theories and methods(Cacioppo, Berntson, & Decety, 2010). What to some of us sounds like apromising idea has often been criticised based on the claim that constructs ofinterest in social psychology are typically higher-order constructs (such asstereotypes and stereotyping) that comprise numerous lower-order pro-cesses, representations, and operations (such as perception, attention,memory, inhibition). As a result, social neuroscientists are faced with theproblem of having to map complex social-psychological constructs onto acomplex biological system like the brain.
This attempt often results in the decomposition of the phenomenon inquestion into its subcomponents. However, such an approach is not uniqueto social neuroscience. It is actually quite congenial to one of the centralgoals of social cognition, namely to understand the cognitive (andincreasingly often also perceptual, emotional, and motivational) processesthat underlie social psychological phenomena (Sherman, 2010). Asportrayed in the first part of this chapter, social psychologists have oftentried to decompose complex social constructs in their attempt to understandthem in greater depth. Put differently, in order to find an answer to a generalquestion (e.g., whether a woman or a black individual would be stereotypedduring a job interview), researchers quickly noticed that it was not enoughto know the prevailing stereotypic beliefs about women or black people in acertain culture. The peculiarities of the situation and those of the peopleinvolved, their concrete appearance, attitudes, attentional resources,emotional reactions, and so forth mattered. Therefore distinguishingbetween aspects of person perception, person categorisation, stereotype
STEREOTYPES AND STEREOTYPING 243
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

activation, stereotype content, stereotype application, and prejudice helpedto illuminate the complexity of the phenomenon.
Does such a compartmentalising approach change the original researchquestion? Potentially. By focusing on particular building blocks of thephenomenon, the scope of specific studies may have narrowed—even thoughtheir scope, when taken together, most certainly broadened. But does thismean that these studies are irrelevant with regard to the original question oreven investigate the ‘‘wrong’’ constructs? We doubt that such an argumentcan be made. Stern critics have nevertheless argued that ‘‘the fact that aconstruct can be decomposed does not necessarily mean that it is useful todo so. [. . .]. The constructs that social psychologists use have a theoreticalintegrity of their own and should not be abandoned in favor of constructsthat may be localizable’’ (Willingham & Dunn, 2003, p. 268). As years ofsocial-cognitive research have shown, however, breaking a phenomenondown into its constituent elements does not lead to abandoning the higher-order construct. Chances are that this construct developed in the first placebecause it summarised a unique constellation of elements and processes.Thus, although its subcomponents may not be unique, their specificcombination might best be summarised with the higher-order construct.As in the case of stereotyping, the term is traditionally (and as far as we cansee also in the future) used to refer to the fact that a human encounter isshaped by the use of specific, culturally shared beliefs that are linked tosocial group memberships. Breaking down the construct in its subcompo-nents does not eliminate the need for referring to their interplay asstereotyping. Along similar lines, we do not see how learning to understandthe neural implementation of these subcomponents (and their interplay)would threaten to replace the construct.
Do these studies have any ecological validity?
‘‘Brain mappers turn individual persons into experimental subjects and put them intonarrow tunnels. [. . .] While staying virtually immobile, the person has to focusattention on various strange stimuli while responding in highly restricted ways thatwere specified before entering the scanner.’’
(Roepstorff, 2004, p. 1106 & p. 1108)
Critics of the field of social neuroscience often point out the artificialenvironment of the testing procedures (particularly with fMRI). Numerousethnographic observations have emphasised that by accepting to take partin neuroscientific studies, participants agree to play a relatively standar-dised role in a well-schemed endeavour (e.g., Cohn, 2008; Roepstorff,2004). However, many of these issues raised also apply to traditionalsocial-cognitive studies. It can hardly be argued, for instance, thatresponding to the computer-paced presentation of a classical word/
244 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

nonword priming task resembles natural human behaviour. Nevertheless,it is not often questioned that such tasks allow us to learn (with respect tocertain well-defined questions) about the unfolding of social-cognitiveprocesses.
Importantly, social neuroscience studies tend to adopt tasks traditionallyused in social psychology, thus using stimuli and modes of response (e.g., abutton presses) that have long characterised the field. This approach mainlydiffers from previous work in the fact that while doing so, participants aretypically asked to lie down in a confined and darkened space and to suppresshead movements. Luckily many of us have daily (or more precisely nightly)experience with lying down, and issues of head movement in the scanner canbe confined by adequate padding (to a certain extent). Far from arguing thatthis is a normal or particularly comfortable state for humans to be in, wewould nevertheless like to point out that these requirements are not so unusualor demanding either that they would interfere with any type of meaningfultask performance. Encouragingly, behavioural effects found in classical social-psychological studies (e.g., the IAT effect, the other-race effect etc.) have nowbeen replicated in the scanner (e.g., Beer et al., 2008; Golby et al., 2001;Knutson et al., 2007), suggesting that participants perform similarly regardlessof whether they are in the scanner or just a ‘‘normal’’ laboratory.
The main criticism that is usually put forward then applies to manysocial-psychological and social neuroscientific studies alike. In everyday lifesocial interactions unfold over time and people change their mental statesand behaviours based on what others do or are expected to do (De Jaegher,Di Paolo, & Gallagher, 2010). Such aspects of temporality and inter-dependency can certainly not be captured during scanning (Willingham &Dunn, 2003). Or can they? Recent studies have not only reportedparticipants playing charades together (Schippers, Roebroeck, Renken,Nanetti, & Keysers, 2010) or engaging in joint actions with the experimenter(Kokal & Keysers, 2010), but also interacting with ‘‘responsive’’ avatars(Wilms et al., 2010) or with co-participants placed in another fMRI scanner(Montague et al., 2002). Although such experimental approaches are not yetthe norm, they demonstrate that creative and inquisitive neuroscientistsconstantly develop new designs with the goal of resembling social lifeoutside the scanner.
What about distributing social neuroscience findings in themedia?
‘‘It is as if [neuroimaging] pictures are stripped of their usual interpretations oncethey enter the world of catchy stories and popular science. They are no longer read asconventional representations that result from a long series of choices andtransformations.’’
(Roepstorff, 2004, p. 1112).
STEREOTYPES AND STEREOTYPING 245
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Research dedicated towards questions of how the social mind works hasgenerally been very popular not only within the scientific community butalso with the general public. Typical social-psychological topics of inquirysuch as our sense of self, morality, altruism, issues of prejudice andintergroup behaviour have often attracted the attention of the media.Milgram’s study of obedience to authority (1963) as well as Zimbardo’sprison experiment (Haney, Banks, & Zimbardo, 1973) are well-knownhistoric examples of social-psychological studies that received coverage inmainstream movies and magazines around the world. With this in mind, ithardly comes as a surprise that social neuroscience as a field aiming toexamine these social phenomena from a new perspective also has thepotential to stir public interest.
There are many reasons to be positive about this. For instance, the merefact that popular media outlets report on science and encourage people toreflect on how scientific findings may shape their understanding of what itmeans to be human is a trend to be welcomed. Indeed, there is now plenty ofevidence that in Western societies neuroscientific ideas and concepts havebecome widespread commodities through their frequent portrayal in themedia, literature, film, and works of art (Dumit, 2004; Frazzetto & Anker,2009). What is less positive, however, is the fact that neuroscientificexplanations seem to have a particularly persuasive power, lendingcredibility to research findings irrespective of the actual scientific reasoningon which they are based. Their persuasiveness has been traced back to apeculiar combination of visually appealing pictures that often elicit theimpression to capture brain activity in photographic terms (i.e., as it unfoldsin reality; Keehner & Fischer, 2011; Keehner, Mayberry, & Fischer, 2011;McCabe & Castel, 2008), and people’s misleading intuitive beliefs about thenature of the mind’s relationship to the brain (Beck, 2010; Weisberg, 2008;Weisberg, Keil, Goodstein, Rawson, & Gray, 2008).
What makes matters worse is that popular media ultimately report onscience to sell newspapers, magazines, podcasts, and so forth. To do so,findings are sometimes oversimplified and distorted, encouraging non-expertconsumers to draw wrong conclusions, to misunderstand the scope of theclaims and/or to believe that the results of a certain experiment are moredefinite then they actually are (Weisberg, 2008). In the realm of personcategorisation, for instance, it has been suggested in Newsweek magazinethat ‘‘the amygdala gets upset when it sees a member of the other team [. . .]but not members of its own team’’ (Begley, 2008, p. 3 referring to findingsfrom Van Bavel et al., 2008). Along similarly obscure lines, results reportedby Cikara and co-workers (2011) have been linked on CNN to the idea that‘‘there’s some hard-wiring [. . .] that can interfere with the average man’sability to interact on deeper levels with really hot looking stranger women inbikinis’’ (Landau, 2009, p. 1).
246 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

It should go without saying that activity in the brain does not indicate inany way whether a mental act is hard-wired (Beck, 2010). It is equallyabsurd to argue that the amygdala is on anyone’s team or feels occasionallyupset. Alas, non-experts should not be expected to spot such fundamentalflaws in reasoning without help. Thus scientists using neuroscientificmethods to study phenomena of social relevance are not only expected tobe particularly critical towards over-interpreting their own findings, but alsoto monitor the ways in which their and other researchers’ data are reportedin the media. Courageous attempts to counter overblown neuroscience-based claims in non-scientific outlets have so far resulted in numerouscritical blogs (e.g., http://www.talkingbrains.org; http://neuroskeptic.blog-spot.com) as well as in the publication of counterstatements in popularmagazines (e.g., Aron et al., 2007). Ultimately, the credibility of the field ofsocial neuroscience will depend on how scientists and journalists manage towork together to communicate not only the fascination but also thecomplexities of social neuroscientific research to the wider public (see Beck,2010; Farah, 2008; Illes et al., 2010; Racine, Bar-Illan, & Illes, 2005).
Are social neuroscience data principally flawed?
‘‘I argue that [social-]cognitive neuroscientists should be circumspect in the use ofreverse inference, particularly when selectivity of the region in question cannot beestablished or is known to be weak.’’
(Poldrack, 2006, p. 59)
It comes as no surprise that mental processes are not directly measurable.Therefore they need to be operationalised as assessable variables in order tobe studied. Over the years (social) psychologists have been highly inventiveat finding dependent measures to quantify their constructs of interest. In ourview, brain activity (as measured via ERP and fMRI) provides just anotherdependent measure that is neither more nor less flawed than many others.On first glance, the use of this measure may seem intuitively appealing.Healthy humans can be tested in various experimental conditions while theirbrain activity (or trackable derivatives such as a variation in bloodoxygenation) is recorded. Based on the assumption that certain patternsof neural activity reflect the engagement of a particular mental process,studying brain activity can inform theories about how the human mindworks and which mental processes contribute to a specific mentalphenomenon (Kosslyn, 1999; Poldrack, 2006, 2008).
This assumption indicates, however, that brain data are only useful forscientific purposes if there is a systematic relation between ‘‘which’’ mentalprocesses are engaged at a certain time and the type of activity (‘‘Where?When?’’) that is observed in the brain during this engagement. When this
STEREOTYPES AND STEREOTYPING 247
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

assumption is met, two types of inferences can theoretically be drawn(Henson, 2005): Function-to-structure deduction describes the idea thatdifferent patterns of neural activity under two experimental conditions implythe engagement of at least one different mental process. In contrast,structure-to-function induction assumes that activity in the same neuralnetwork of brain regions under two experimental conditions implies acommon function that possibly was not previously predicted. In both casesneuroscientific data form a dependent variable that can be considered to testcompeting hypotheses. Unfortunately, warranted structure-to-functioninductions turned out to be more challenging than originally assumed. Thisis due to the fact that many brain regions are recruited by many differentmental processes (Poldrack, 2006). Therefore interpretation of fMRI dataresting on the assumption that activation in a particular brain region (ornetwork of brain regions) is a marker of engagement of a particular mentalprocess (a practice commonly referred to as reverse inference) can often bequestioned.
As a case in point, consider the amygdala’s role in the stereotypingprocess. It has repeatedly been observed that amygdala activity is enhancedwhen white participants view black relative to white faces (Cunninghamet al., 2004; Hart et al., 2000). In addition there is plenty of evidence thatactivity in the amygdala can be linked to the experience of threat and fear(see Adolphs, 2008). Does that imply that white perceivers feel threatened orexperience fear when looking at black faces? Such a conclusion wouldcertainly count as a questionable reverse inference. First of all, the amygdalais also involved in the processing of reward, novelty, and ambiguity, anddoes not act as a mere marker of threat and fear (for reviews see Amodio,2008; Whalen, 1998). Thus, since the original face perception studies lackconverging (e.g., behavioural, neurophysiological) markers indicative of afear and/or threat response, the amygdala’s specific function in these studiesremains elusive.
Second, the mechanisms through which a threat/fear response could havebeen triggered in these studies are highly speculative at this point. Based onprevious behavioural findings, it may be argued that fear or threat responsesresult from rapid stereotype or prejudice activation. Such reasoning,however, has not received support from subsequent neuroimaging studies.It is well known from classic social-psychological research, for example, thatfaces with more Afrocentric features elicit stronger activation of the African-American stereotype than faces with less prototypical features (e.g., Blairet al., 2004). Amygdala activation, however, fails to respond differentlytowards black faces differing in race typicality (Ronquillo et al., 2007),making a stereotype-based explanation a poor fit with previous insights. Theobserved amygdala activation is also unlikely to reflect processes of explicitprejudice given that none of the studies required participants to respond in a
248 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

prejudiced manner while perceiving the faces, nor were the studies able todemonstrate that a relationship exists between the magnitude of amygdalaresponse to outgroup members and participants’ explicit prejudice(Cunningham et al., 2004; Phelps et al., 2000; Wheeler & Fiske, 2005). Sowhat about the idea that it may reflect traces of implicit prejudice? So far, anfMRI study examining the amygdala’s role in rapid race-based associations(as measured with the IAT) found increased amygdala activity during rapidpositive associations with ingroup members rather than during negativeassociations with outgroup members (Beer et al., 2008).
Alternatively, it has been proposed that the amygdala may signal agenerally increased vigilance towards outgroup members. This assumptionhas been challenged by findings showing that it can also be the perception ofingroup compared to outgroup faces that elicits increased amygdalaactivation (Van Bavel et al., 2008; Wright et al., 2008). It also needs to benoted that the circumstances under which the amygdala differentiatesbetween black and white faces are poorly understood. When participantsare making socially meaningful judgements about others such as judgingtheir age, sex or race (Hart et al., 2000; Lieberman, Hariri, Jarcho,Eisenberger, & Bookheimer, 2005; Ronquillo et al., 2007; Wheeler &Fiske, 2005), activity in the amygdala seems to spontaneously track withthe target’s racial status such that increased responses for faces that do notmatch the perceiver’s own race are elicited. In contrast, though, activity inthe amygdala remains insensitive to target race when faces are processed ata superficial visual level (i.e., during a dot detection task, a localisationtask, or a n-back task; Cunningham et al., 2004; Phelps et al., 2000;Wheeler & Fiske, 2005).
What this example shows is that drawing inferences about the functionalcontribution of a certain brain area is often difficult. It becomes even moredifficult when a brain structure (even one as well defined as the amygdala)consists of important substructures that are likely to differ in functionality.Given the limited resolution of many fMRI studies at this point, the specificsubstructures contributing to observable activation remain often undeter-minable. Thus different fMRI studies that seem to report activity in the samebrain region may actually pick up activity in slightly different subregions.Therefore, rather than merely explaining observed activations in a post hocmanner, the replication of consistent and specific patterns of activation andthorough experimentation to determine their boundary conditions arenecessary prerequisites to understand the functional mapping between mindand brain.
Also, acquiring converging evidence from behavioural and psychophy-siological markers is frequently of pivotal importance to inform adequatedata interpretation. After all, these markers can help researchers tounderstand the exact processes contributing to the observed neuroscientific
STEREOTYPES AND STEREOTYPING 249
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

effects (see Amodio, Harmon-Jones, & Devine, 2003; Wilkinson & Halligan,2004). To further probe amygdala activity during outgroup face perception,for instance, Amodio and colleagues (2003) made use of an emotion-modulated startle-eyeblink procedure. When a person is startled, theytypically show a defensive eyeblink. This blink response is stronger if aperson is in an aversive rather than appetitive state just prior to beingstartled—an effect mediated by the central nucleus of the amygdala (Davis,1992; LeDoux, 2000). Amodio et al. (2003) chose to use the startle eyeblinkmeasure because it could target a specific substructure of the amygdala.Using this approach they discovered larger startle eyeblink amplitudes toblack relative to white faces, supporting the idea of an aversive response toblack people among white participants. Given that the degree of affectivebias was unrelated to self-reported racial attitudes, he interpreted thefindings as evidence for an implicit intergroup bias.
Another often-discussed problem with ERP and fMRI studies is that theygenerally cannot speak to the issue of causality. The mere observation ofbrain activity during a certain mental task does neither reflect that thisregion is necessary nor sufficient for the completion of a specific task(Poldrack, 2008). However, studies evaluating the impact of temporary (asinduced by TMS) or permanent brain lesions (as following brain damage)allow the gap in establishing causal relationships to be filled. Morespecifically, such investigations offer the opportunity to elucidate whichsocial-cognitive abilities are impaired following the inability to recruitcertain brain regions. In this respect the interpretation of fMRI and ERPdata builds inherently on converging evidence from other methodologies(Klein, Lax, & Gangi, 2010). In the realm of person perception andstereotype control, for instance, TMS studies have managed to challenge theidea of mere epiphenomenality of the identified neural networks (e.g.,Barton et al., 2002; Cattaneo et al., 2011; Grossman et al., 2005; Pitcheret al., 2007; Saygin, 2007; Sorger et al., 2007; Urgesi et al., 2004, 2007).
Finally, when judging the quality of social neuroscience studies, it shouldalso be considered that this young field is still in the process of establishing acommon and well-justified ‘‘code of practice’’. How much so was recentlyseen when puzzlingly high correlations between brain activity andpersonality measures caused substantial controversy about adequatemethods of fMRI data analyses—see Perspectives on Psychological Science,4(3), 2009, for details. Luckily, this upheaval has led to reassuringdevelopments. The wider neuroimaging community, for instance, has begunto actively exchange views on which statistical analyses are adequate andwhich are less so (see, e.g., Bennett, Baird, Miller, & Wolford, 2009;Kriegeskorte, Simmons, Bellgowan, & Baker, 2009). From this dialogue andagreement on statistical recommendations, fMRI results are likely to gain invalidity and reliability in the future. With a similar goal in mind, leading
250 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

experts in the field have specified guidelines according to whichresearchers should report their methods and results in fMRI studies(Poldrack et al., 2008). By providing standards to overcome thewidespread lack of methodological detail in peer-reviewed fMRI articles,reviewers and researchers will be able to contribute to stricter qualitycontrol by being able to question and/or to replicate conducted studies atgreater detail.
Have we learned anything (yet)?
‘‘It isn’t, after all, seriously in doubt that talking (or riding a bicycle, or building abridge) depends on things that go on in the brain somewhere or other. If the mindhappens in space at all, it happens somewhere north of the neck. What exactly turnson knowing how far north?’’
(Fodor, 1999, p. 4)
In the field of social neuroscience two types of learning are currently takingplace (Amodio, 2010; Cunningham, 2010). The first type is generally referredto as brain mapping. Brain mapping allows us to gain an understanding ofwhich neural networks implement which social processes or constructs. Withregard to stereotyping, for instance, this approach has helped us todifferentiate brain networks dedicated to person perception and stereotypecontrol. Other networks are currently speculative at best and remain to belocalised such as, for instance, networks dedicated to person categorisation,or storing and activating stereotypic knowledge. Unfortunately, the resultsof brain-mapping studies may not keep you awake all night bouncing withexcitement, unless you do actually have a certain nerdy interest in brainlocalisation (as we do).
In addition to brain mapping experiments, however, learning alsotakes place through studies aimed at hypothesis testing. This approachinherently builds on the results of brain-mapping studies by assumingthat a particular brain region or a network thereof reflects a specificprocess or construct. These studies can therefore help us to refine ourunderstanding of social phenomena. Social neuroscientists are often askedto provide specific examples of studies that have successfully distinguishedbetween competing social-psychological theories. It needs to be pointedout, though, that in psychological science single experiments providingunequivocal support for only one theory are hard to find no matter whatkind of dependent variable is considered, given that progress is usuallydriven by the accumulation of converging data over time and by iterativetheory development (Henson, 2006). Thus a fair judgement about theusefulness of social-neuroscientific data should certainly consider itscumulative contribution to topics of interest (Yarkoni, Poldrack, VanEssen, & Wager, 2010).
STEREOTYPES AND STEREOTYPING 251
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Nevertheless, we would like to give an example. Consider, for instance,our recent study that examined the effects of occupational genderstereotypes on person perception (Quadflieg et al., 2011). Response timedata can help us to understand that sex-categorisation of unexpected targetsrequires additional processing resources (e.g., as reflected in slower responsetimes). What remains less certain is whether this observed change is drivenby altered perceptual, cognitive, or motor operations. For instance, theeffect can be conceptualised as a conflict at the response output stage (i.e.,the face indicates that the person is a woman, whereas the stereotypesuggests the opposite—in order to provide a button press a selection of theadequate motor response needs to be made). In contrast, it could also reflectaltered perceptual processing due to stereotype-related prediction errors. Bymaking use of previous neuroscientific research that informs us about theneural networks underlying response selection vs person perception, we wereable to disentangle these possibilities. The example also shows that carefulwork on brain mapping has to precede hypothesis testing (see for furtherdetails Amodio, 2010). If the basic mapping of brain function is poor, thensubsequent hypothesis testing becomes problematic, clearly diminishing thepotential contribution of the work. In this respect, rushing as a fieldfrom attempts of brain mapping to hypothesis testing is likely to provecounter-productive (Cunningham, 2010). With these thoughts in mind, let usevaluate in how far neuroscientific methods have contributed to answeringany of the unresolved questions raised in the first section of this chapter.
Open questions revisited
‘‘I always suspected that social neuroscience has contributed more to helpingneuroscientists understanding the brain than it has done to help social psychologistsunderstanding psychological processes. So far, reading your manuscript has not reallysucceeded in alleviating my suspicion.’’(Stroebe, 2011, editor of European Review of Social Psychology in response to this
manuscript)
There is no denying that so far studies investigating the neural signature ofstereotyping have generally been in the brain mapping rather than in thehypothesis testing stage of events. Nevertheless we would argue that the useof neuroscientific methods has begun to advance our understanding of thenature of stereotyping. As a case in point, think of the idea that stereotypingunfolds inevitably upon the perception of others (Brewer, 1988; Fiske &Neuberg, 1990)? Although challenged before, many social psychologists stillargue in favour of a strong notion of automaticity and inevitability in sociallife (e.g., Bargh & Williams, 2006). Neuroimaging studies have repeatedlyshown, however, that diverging processing goals can lead to very diverseneural networks (and strategies of impression formation) being engaged
252 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

during person construal (e.g., Quadflieg et al., 2011; Wheeler & Fiske, 2005).Also, conflicting findings, as in the case of outgroup face processing and theN170, give emphasis to the idea that processing goals play an importantmodulatory role during all stages of the stereotyping cascade—even forthose typically considered very basic (such as person perception). In thisrespect, highly sensitive neuroscientific markers have emphasised the need todefine all the building blocks of the phenomenon in question and to specifyprecisely what participants exactly do when asked to complete stereotype-related tasks.
In addition, researchers are beginning to disentangle the neural correlatesof different types of social learning (e.g., based on observation vsinstruction; Li, Delgado, & Phelps, 2010; Olsson & Phelps, 2007), and toshow that social categorisation based on photos of people relative to merelabels have fundamentally different neural consequences (Lieberman et al.,2005). Although both lines of research are relatively recent, their replication,advancement, and extension to the stereotyping literature will most likelyallow us to address previously raised issues of how factor such as the mannerof stereotype acquisition and the type of category/triggering stimulusmaterial shape the stereotyping process. Along similar lines, understandingthe neural networks involved in evaluative and cognitive functioning opensthe exciting opportunity to study the interplay of stereotypic and prejudicedaspects of category-based responses towards others (see Amodio & Devine,2006, for a discussion).
Most prominently to date, neuroscientific studies have shown thatcategorical and stereotypic beliefs can modulate person perception (Quad-flieg et al., 2011; van Bavel et al., 2008; 2011). As such, these data provideconverging evidence that outgroup members are encoded in a lessindividualistic manner than ingroup members, and that stereotypic beliefsare typically used to make sense of visual person input.
Apart from tackling previous questions of interest, in our opinion,neuroscientific methods have also raised intriguing new issues and possibilities.The use of neuroscientific methods was originally fuelled by the idea thatidentifying neural ‘‘footprints’’ of various mental processes might provide acommon language for psychology (Kosslyn, 1999). Indeed, insights on how thebrain stores conceptual knowledge more generally (i.e., according to domainsand sensory modalities), have inspired us to think in new ways about thestorage of stereotypic beliefs. Thus, by learning about general rules of brainfunction, highly specific psychological constructs can be redefined andspecified. On another note, since people rarely know how to voluntarilymanipulate their level of neural activity, neuroscientific markers of stereotyp-ing are likely to be less biased by effects of strategic responding—a concernparticularly relevant when assessing stereotypes of social groups that conflictwith widespread norms of social desirability. In this respect, indicators like the
STEREOTYPES AND STEREOTYPING 253
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

N400 may open up the development of exciting new implicit measures ofcategory-based processing. As such, of course, neuroscientific markers mayultimately become a powerful mean to evaluate the effectiveness of stereotypeprevention and intervention programs.
CONCLUSION
To describe fixed impressions through which people make sense of theworld, the American journalist Walter Lippmann (1922) originallyintroduced the term stereotype. Almost a century later, social psychologistsstill struggle to understand the antecedents and consequences of stereotypesand stereotyping in their entire complexity. Through the use of modernneuroscientific methods, some researchers hope to advance the field. So far,this work indicates that a network of neural areas is specifically dedicatedtowards the perception of conspecifics. However, how the encoded visualperson input is subsequently transformed and used by the brain torapidly categorise and stereotype others is less well understood.Admittedly though, questions of how person perception triggers personcategorisation, stereotype activation, and stereotype application havepuzzled social psychologists working with traditional research methodsfor quite some time. On a very positive note this means that socialpsychologists and social neuroscientists are presently united in their topicsof interest. Nevertheless, progress on these issues will not easily beobtained. As Willingham and Dunn (2003) once noted ‘‘If [brainmapping] is difficult with [lower-order] processes [. . .], how much moredifficult will it be when studying processes (e.g., stereotyping) that haveother processes (attention) embedded in them?’’ (p. 268). Although attimes we are certainly inclined to mumble a grumpy ‘‘Indeed!’’, the factthat something is difficult to do, does luckily not mean that it cannotprincipally be done. It ‘‘just’’ requires the careful coordination of theorieson how the social mind works with those on how the brain works. Attimes the former, at times the latter, will profit most from this intellectualmarriage. Due to their cultural origin, however, it is worth keeping inmind that stereotypes cannot be captured by studying single brains alone.Only a multidisciplinary approach comprising the work of psychologists,sociologists, anthropologists, and neuroscientists can ultimately succeed inexplaining when, why, and how the human brain and mind are drawn tostereotypes and stereotyping.
REFERENCES
Adolphs, R. (2006). How do we know the minds of others? Domain-specificity, simulation, and
enactive social cognition. Brain Research, 1079, 25–35.
254 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Adolphs, R. (2008). Fear, faces, and the human amygdala. Current Opinion in Neurobiology, 18,
166–172.
Adolphs, R. (2009). The social brain: Neural basis of social knowledge. Annual Review of
Psychology, 60, 693–716.
Agerstrom, J., & Rooth, D-O. (2011). The role of automatic obesity stereotypes in real hiring
discrimination. Journal of Applied Psychology, 96, 790–805.
Allport, G. W. (1954). The nature of prejudice. Reading, MA: Addison Wesley.
Amodio, D. M. (2008). The social neuroscience of intergroup relations. European Review of
Social Psychology, 19, 1–54.
Amodio, D. M. (2009). Coordinated roles of motivation and perception in the regulation of
intergroup responses: Frontal cortical asymmetry effects on the P2 event-related potential
and behavior. Journal of Cognitive Neuroscience, 22, 2609–2617.
Amodio, D. M. (2010). Can neuroscience advance social psychological theory? Social
neuroscience for the behavioral social psychologist. Social Cognition, 28, 695–716.
Amodio, D. M., & Devine, P. G. (2006). Stereotyping and evaluation in implicit race bias:
Evidence for independent constructs and unique effects on behavior. Journal of Personality
and Social Psychology, 91, 652–661.
Amodio, D. M., Devine, P. G., & Harmon-Jones, E. (2008). Individual differences in the
regulation of intergroup bias: The role of conflict monitoring and neural signals for control.
Journal of Personality and Social Psychology, 94, 60–74.
Amodio, D. M., & Frith, C. D. (2006). Meeting of minds: The medial prefrontal cortex and
social cognition. Nature Reviews Neuroscience, 7, 268–277.
Amodio, D. M., Harmon-Jones, E., & Devine, P. G. (2003). Individual differences in the
activation and control of affective race bias as assessed by startle eyeblink response and self-
report. Journal of Personality and Social Psychology, 84, 738–753.
Amodio, D. M., Harmon-Jones, E., Devine, P. G., Curtin, J. J., Hartley, S. L., & Covert, A. E.
(2004). Neural signals for the detection of unintentional race bias. Psychological Science, 15,
88–93.
Amodio, D. M., Kubota, J. T., Harmon-Jones, E., & Devine, P. G. (2006). Alternative
mechanisms for regulating racial responses according to internal vs. external cues. Social
Cognitive and Affective Neuroscience, 1, 26–36.
Amodio, D. M. & Mendoza, S. (2010). Implicit intergroup bias: Cognitive, affective, and
motivational underpinnings. In B. Gawronski & B. K. Payne (Eds.), Handbook of implicit
social cognition (pp. 353–374). New York: Guilford Press.
Anastasi, J. S., & Rhodes, M. G. (2005). An own-age bias in face recognition for children and
older adults. Psychonomic Bulletin & Review, 12, 1043–1047.
Andreasen, N. C., O’Leary, D. S., Arndt, S., Cizadlo, T., Hurtig, R., Rezai, K., et al. (1996).
Neural substrates of facial recognition. Journal of Neuropsychiatry and Clinical
Neuroscience, 8, 139–146.
Appel, M., Kronberger, N., & Aronson, J. (2011). Stereotype threat impairs ability building:
Effects on test preparation among women in science and technology. European Journal of
Social Psychology, doi: 10.1002/ejsp.835.
Aron, A., Badre, D., Brett, M., Cacioppo, J., Chambers, C., Cools, R., et al. (2007). Politics
and the brain. [Letter to the editor.] The New York Times, http://www.nytimes.com/2007/11/
14/opinion/lweb14brain.html
Aronson, J., & Inzlicht, M. (2004). The ups and downs of attributional ambiguity. Stereotype
vulnerability and the academic self-knowledge of African American College Students.
Psychological Science, 15, 829–836.
Assmus, A., Giessing, C., Weiss, P. H., & Fink, G. R. (2007). Functional interactions during the
retrieval of conceptual action knowledge: An fMRI study. Journal of Cognitive
Neuroscience, 19, 1004–1012.
STEREOTYPES AND STEREOTYPING 255
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Bahrick, H. P., Bahrick, P. O., & Wittlinger, R. (1975). Fifty years on memory for names and
faces: A cross-sectional approach. Journal of Experimental Psychology: General, 104, 54–75.
Barden, J., Maddux, W. W., Petty, R. E., & Brewer, M. B. (2004). Contextual moderation of
racial bias: The impact of social roles on controlled and automatically activated attitudes.
Journal of Personality and Social Psychology, 87, 5–22.
Bargh, J. A., & Williams, L. E. (2006). The automaticity of social life. Current Directions in
Psychological Science, 15, 1–4.
Bartholow, B. D. (2010). On the role of conflict and control in social cognition: Event-related
brain potential investigations. Psychophysiology, 47, 201–212.
Bartholow, B. D., & Dickter, C. L. (2011). Person perception. In J. Decety & J. T. Cacioppo
(Eds.), Handbook of social neuroscience (pp. 404–418). New York: Oxford University Press.
Barton, J. J., Press, D. Z., Keenan, J. P., & O’Connor, M. (2002). Lesions of the fusiform face
area impair perception of facial configuration in prosopagnosia. Neurology, 58, 71–78.
Beck, D. M. (2010). The appeal of the brain in the popular press. Perspectives on Psychological
Science, 5, 762–766.
Beer, J. S., Stallen, M., Lombardo, M. V., Gonsalkorale, K., Cunningham, W. A., & Sherman,
J. W. (2008). The quadruple process model approach to examining the neural underpinnings
of prejudice. NeuroImage, 43, 775–783.
Begley, S. (2008). How your brain looks at race. Newsweek, http://www.newsweek.com/2008/
02/23/how-your-brain-looks-at-race.html
Bennett, C. M., Baird, A. A., Miller, M. B., & Wolford, G. L. (2009). Neural correlates of
interspecies perspective taking in the post-mortem Atlantic salmon: An argument for multiple
comparisons correction. Poster presented at the 15th Annual Meeting of the Organisation for
Human Brain Mapping. San Francisco, CA.
Bentin, S., Allison, T., Puce, A., Perez, E., & McCarthy, G. (1996). Electrophysiological studies
of face perception in humans. Journal of Cognitive Neuroscience, 8, 551–565.
Biernat, M. (2003). Toward a broader view of social stereotyping. American Psychologist, 58,
1019–1027.
Biernat, M., & Manis, M. (1994). Shifting standards and stereotype-based judgements. Journal
of Personality and Social Psychology, 66, 5–20.
Bigler, R. S., & Liben, L. S. (2006). A developmental intergroup theory of social stereotypes and
prejudice. In R. V. Kail (Ed.), Advances in child development and behavior (Vol. 34, pp. 39–
89). San Diego: Elsevier.
Bigler, R. S., & Liben, L. S. (2007). Developmental intergroup theory: Explaining and reducing
children’s social stereotyping and prejudice. Current Directions in Psychological Science, 16,
162–166.
Bindemann, M., Scheepers, C., Ferguson, H. J., & Burton, A. M. (2010). Face, body, and center
of gravity mediate person detection in natural scenes. Journal of Experimental Psychology:
Human Perception and Performance, 36, 1477–1485.
Blair, I. V., Judd, C. M., & Chapleau, K. M. (2004). The influence of Afrocentric facial features
in criminal sentencing. Psychological Science, 15, 674–679.
Blair, I. V., Judd, M., Sadler, M. S., & Jenkins, C. (2002). The role of Afrocentric features in
person perception: Judging by features and categories. Journal of Personality and Social
Psychology, 83, 5–25.
Blake, R., & Shiffrar, M. (2007). Perception of human motion. Annual Review of Psychology, 58,
47–73.
Bodenhausen, G. V., & Lichtenstein, M. (1987). Social stereotypes and information-processing
strategies: The impact of task complexity. Journal of Personality and Social Psychology, 52,
871–880.
Bodenhausen, G. V., & Wyer, R. S. Jr. (1985). Effects of stereotypes on decision making and
information-processing strategies. Journal of Personality and Social Psychology, 48, 267–282.
256 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Boldry, J. G., Gaertner, L., & Quinn, J. (2007). Measuring the measures: A meta-analytic
investigation of the measures of outgroup homogeneity. Group Processes and Intergroup
Relations, 10, 157–178.
Bracci, S., Ietswaart, M., Peelen, M. V., & Cavina-Pratesi, C. (2010). Dissociable neural
responses to hands and non-hand body parts in human left extrastriate visual cortex. Journal
of Neurophysiology, 103, 3389–3397.
Brebner, J. L., Krigolson, O., Handy, T. C., Quadflieg, S., & Turk, D. J. (2011). The importance
of skin color and facial structure in perceiving and remembering others: An electro-
physiological study. Brain Research, 1388, 123–133.
Brewer, M. B. (1979). In-group bias in the minimal intergroup situation: A cognitive-
motivational analysis. Psychological Bulletin, 86, 307–324.
Brewer, M. B. (1988). A dual-process model of impression formation. In R. S. Wyer Jr. & T. K.
Srull (Eds.), Advances in social cognition (Vol. 1, pp. 1–36). Hillsdale, NJ: Lawrence Erlbaum
Associates Inc.
Bruce, V., & Young, A. W. (1998). In the eye of the beholder: The science of face perception.
Oxford, UK: Oxford University Press.
Burnett, S., Sebastian, C., Cohen Kadosh, K., & Blakemore, S-J. (2011). The social brain in
adolescence: Evidence from functional magnetic resonance imaging and behavioural studies.
Neuroscience and Biobehavioral Reviews, 35, 1654–1664.
Cacioppo, J. T., Berntson, G. G., & Decety, J. (2010). Social neuroscience and its relationship to
social psychology. Social Cognition, 28, 675–685.
Caldara, R., Rossion, B., Bovet, P., & Hauert, C. (2004). Event-related potentials and time
course of the ‘other-race’ face classification advantage. NeuroReport, 15, 905–910.
Caldara, R., Thut, G., Servoir, P., Michel, C., Bovet, P., & Renault, B. (2003). Face versus
nonface object perception and the ‘other-race’ effect: A spatio-temporal event-related
potential study. Clinical Neurophysiology, 114, 515–528.
Cameron, J. A., & Trope, Y. (2004). Stereotype-biased search and processing of information
about group members. Social Cognition, 22, 650-672.
Campbell, R., MacSweeney, M., Surguladze, S., Calvert, G., McGuire, P., Suckling, J., et al.
(2001). Cortical substrates for the perception of face actions: An fMRI study of the
specificity of activation for seen speech and for meaningless lower-face acts. Cognitive Brain
Research, 12, 233–243.
Castelli, L., Carraro, L., Tomelleri, S., & Amari, A. (2007). White children’s alignment to the
perceived racial attitudes of the parents: Closer to the mother than the father. British Journal
of Developmental Psychology, 25, 353–357.
Castelli, L., De Dea, C., & Nesdale, D. (2008). Learning social attitudes: Children’s sensitivity to the
nonverbal behavior of adult models. Personality and Social Psychology Bulletin, 34, 1504–1513.
Cattaneo, Z., Mattavelli, G., Platania, E., & Papagno, C. (2011). The role of the prefrontal
cortex in controlling gender-stereotypical associations: A TMS investigation. NeuroImage,
56, 1839–1846.
Chao, L. L., Haxby, J. V., & Martin, A. (1999). Attribute-based neural substrates in
temporal cortex for perceiving and knowing about objects. Nature Neuroscience, 2, 913–
919.
Cikara, M., Eberhardt, J. L., & Fiske, S. T. (2010). From agents to objects: Sexist attitudes and
neural responses to sexualized targets. Journal of Cognitive Neuroscience, 23, 1–12.
Cloutier, J., Gabrieli, J. D. E., O’Young, D., & Ambady, N. (2011). An fMRI study of
violations of social expectations: When people are not who we expect them to be.
NeuroImage, 57, 583–588.
Cloutier, J., Turk, D. J., & Macrae, C. N. (2007). Extracting variant and invariant information
from faces: The neural substrates of gaze detection and sex categorisation. Social
Neuroscience, 3, 69–78.
STEREOTYPES AND STEREOTYPING 257
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Cohn, S. (2008). Petty cash and the neuroscientific mapping of pleasure. Biosocieties, 3, 151–163.
Consalkorale, K., Sherman, J. W., Allen, T. J., Klauer, K. C., & Amodio, D. M. (2011).
Accounting for successful control of implicit racial bias: The roles of association activation,
response monitoring, and overcoming bias. Personality and Social Psychology Bulletin, 37,
1534–1545.
Contreras, J. M., Banaji, M. R., & Mitchell, J. P. (2010). Dissociable neural correlates of
stereotypes and other forms of semantic knowledge. Social Cognitive and Affective
Neuroscience, doi: 10.1093/scan/nsr053.
Conway, M., Pizzamiglio, M. T., & Mount, L. (1996). Status, communality, and agency:
Implications for stereotypes of gender and other groups. Journal of Personality and Social
Psychology, 71, 25–38.
Crocker, J., Hannah, D., & Weber, R. (1983). Person memory and causal attribution. Journal of
Personality and Social Psychology, 44, 55–66.
Crocker, J., Voelkl, K., Testa, M., & Major, B. (1991). Social stigma: The affective
consequences of attributional ambiguity. Journal of Personality and Social Psychology, 60,
218–228.
Crookes, K., &McKone, E. (2009). Early maturity of face recognition: No childhood development
of holistic processing, novel face encoding, or face-space. Cognition, 111, 219–247.
Cuddy, A. J. C., Fiske, S. T., Kwan, V. S. Y., Glick, P., Demoulin, S., Leyens, J. P., et al.
(2009). Stereotype content model across cultures: Towards universal similarities and some
differences. British Journal of Social Psychology, 48, 1–33.
Cunningham, W. A. (2010). In defense of brain mapping in social and affective neuroscience.
Social Cognition, 28, 716–721.
Cunningham, W. A., Johnson, M. K., Raye, C. L., Gatenby, J. C., Gore, J. C., & Banaji, M. R.
(2004). Separable neural components in the processing of black and white faces.
Psychological Science, 15, 806–813.
Czopp, A. M., & Monteith, M. J. (2006). Thinking well of African Americans: Measuring
complimentary stereotypes and negative prejudice. Basic and Applied Social Psychology, 28,
233–250.
Damasio, H., Grabowski, T. J., Frank, R., Galaburda, A. M., & Damasio, A. R. (1994). The
return of Phineas Gage: Clues about the brain from the skull of a famous patient. Science,
264, 1102–1105.
Darley, J. M., & Gross, P. H. (1983). A hypothesis-confirming bias in labelling effects. Journal
of Personality and Social Psychology, 44, 20–33.
Davis, M. (1992). The role of the amygdala in fear and anxiety. Annual Review of Neuroscience,
15, 353–375.
Deaux, K., & Lewis, L. L. (1984). Structure of gender stereotypes: interrelationships among
components and gender label. Journal of Personality and Social Psychology, 46, 991–1004.
Decety, J., & Grezes, J. (1999). Neural mechanisms subserving the perception of human actions.
Trends in Cognitive Sciences, 3, 172–178.
De Jaegher, H., Di Paolo, E., & Gallagher, S. (2010). Does social interaction constitute social
cognition? Trends in Cognitive Sciences, 14, 441–447.
De Neys, W., Vartanian, O., & Goel, V. (2008). Smarter than we think. When our brains detect
that we are biased. Psychological Science, 19, 483–489.
Devine, P. G. (1989). Stereotypes and prejudice: Their automatic and controlled components.
Journal of Personality and Social Psychology, 56, 5–18.
Devine, P. G., & Elliot, A. J. (1995). Are racial stereotypes really fading? The Princeton trilogy
revisited. Personality and Social Psychology Bulletin, 22, 22–37.
Diekman, A. B., & Eagly, A. H. (2000). Stereotypes as dynamic constructs: Women and
men of the past, present, and future. Personality and Social Psychology Bulletin, 26,
1171–1188.
258 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Dickter, C. L., & Bartholow, B. D. (2007). Racial ingroup and outgroup attention biases revealed
by event-related potentials. Social, Cognitive, and Affective Neuroscience, 2, 189–198.
Downing, P. E., Bray, D., Rogers, J., & Childs, C. (2004). Bodies capture attention when
nothing is expected. Cognition, 93, B27–B38.
Downing, P. E., & Peelen, M. V. (2011). The role of occipitotemporal body-selective regions in
person perception. Cognitive Neuroscience, doi: 10.1080/17588928.2011.582945.
Dumit, J. (2004). Picturing personhood: Brain scans and biomedical identity. Princeton, NJ:
Princeton UP.
Duncan, B. L. (1976). Differential social perception and attribution of intergroup violence:
Testing the lower limits of stereotyping of blacks. Journal of Personality and Social
Psychology, 34, 590–598.
Eagly, A. H., & Diekman, A. B. (2005). What is the problem? Prejudice as an attitude-in-
context. In J. F. Dovidio, P. Glick, & L. A. Rudman (Eds.), On the nature of prejudice (pp.
175–191). Malden, MA: Blackwell Publishing.
Ehrlich, H. J. (1973). The social psychology of prejudice. New York: Wiley.
Ellis, A. W., Young, A. W., Critchley, A. M. R. (1989). Loss of memory for people following
temporal lobe damage. Brain, 112, 1469–1483.
Farah, M. J. (2008). A picture is worth a thousand dollars. Journal of Cognitive Neuroscience,
21, 623–624.
Fazio, R. H., Sanbonmatsu, D. M., Powell, M. C., & Kardes, F. R. (1986). On the automatic
activation of attitudes. Journal of Personality and Social Psychology, 50, 229–238.
Federico, C. M. (2004). Predicting attitude extremity: The interactive effects of schema
development and the need to evaluate and their mediation by evaluative integration.
Personality and Social Psychology Bulletin, 30, 1281–1294.
Fein, S., & Spencer, S. J. (1997). Prejudice as self-image maintenance: Affirming the self through
derogating others. Journal of Personality and Social Psychology, 73, 31–44.
Fiedler, K., Freytag, P., & Meiser, T. (2009). Pseudocontingencies: An integrative account of an
intriguing cognitive illusion. Psychological Review, 116, 187–206.
Fiske, S. T. (2005). Social cognition and the normality of prejudgement. In J. F. Dovidio, P.
Glick, & L. A. Rudman (Eds.), On the nature of prejudice (pp. 36–53). Malden, MA:
Blackwell Publishing.
Fiske, S. T., Cuddy, A. J. C., Glick, P., & Xu, J. (2002). Model of (often mixed) stereotype
content: Competence and warmth respectively follow from perceived status and competi-
tion. Journal of Personality and Social Psychology, 82, 878–902.
Fiske, S. T., & Lee, T. L. (2008). Stereotypes and prejudice create workplace discrimination. In A.
P. Brief (Ed.), Diversity at work (pp. 13–52). Cambridge, UK: Cambridge University Press.
Fiske, S. T., & Neuberg, S. L. (1990). A continuum of impression formation, from category-
based to individuating processes: Influences of information and motivation on attention and
interpretation. Advances in Experimental Social Psychology, 23, 1–74.
Fletcher-Watson, S., Findlay, J. M., Leekam, S. R., & Benson, V. (2008). Rapid detection of
person information in a naturalistic scene. Perception, 37, 571–583.
Fodor, J. (1999). Let your brain alone. London Book Review, 21, 1–5.
Ford, T. E., & Stangor, C. (1992). The role of diagnosticity in stereotype formation:
Perceiving group means and variances. Journal of Personality and Social Psychology, 63,
356–367.
Frazzetto, G., & Anker, S. (2009). Neuroculture. Nature Reviews Neuroscience, 10, 815–821.
Freeman, J. B., Ambadi, N., & Holcomb, P. J. (2010). The face-sensitive N170 encodes social
category information. NeuroReport, 21, 24–28.
Freeman, J. B., Ambady, N., & Rule, N. (2008). Will a category cue attract you? Motor output
reveals dynamic competition across person construal. Journal of Experimental Psychology:
General, 137, 673–690.
STEREOTYPES AND STEREOTYPING 259
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Freeman, J. B., Pauker, K., Apfelbaum, E. P., & Ambady, N. (2010). Continuous dynamics
in the real-time perception of race. Journal of Experimental Social Psychology, 46, 179–
185.
Freeman, J. B., Rule, N., Adams, R. B., & Ambady, N. (2010). The neural basis of categorical
face perception: Graded representations of face gender in fusiform and orbitofrontal
cortices. Cerebral Cortex, 20, 1314–1322.
George, N., Evans, J., Fiori, N., Davidoff, J., & Renault, B. (1996). Brain events related to
normal and moderately scrambled faces. Cognitive Brain Research, 4, 65–78.
George, N., Jemel, B., Fiori, N., Chaby, L., & Renault, B. (2005). Electrophysiological
correlates of facial decision: Insights from upright and upside-down mooney-face
perception. Cognitive Brain Research, 24, 663–673.
Giese, M. A., & Poggio, T. (2003). Neural mechanisms for the recognition of biological
movements. Nature Reviews Neuroscience, 4, 179–192.
Gilbert, C. D., & Sigman, M. (2007). Brain states: Top-down influences in sensory processing.
Neuron, 54, 677–696.
Gilbert, D. T., & Hixon, J. G. (1991). The trouble of thinking. Activation and application of
stereotypic beliefs. Journal of Personality and Social Psychology, 60, 509–517.
Gilbert, D. T., & Malone, P. S. (1995). The correspondence bias. Psychological Bulletin, 117,
21–38.
Giles, J. W., & Heyman, G. D. (2005). Young children’s beliefs about the relationship between
gender and aggressive behavior. Child Development, 76, 107–121.
Glick, P., & Fiske, S. T. (2001). An ambivalent alliance: Hostile and benevolent sexism as
complementary justifications for gender inequality. American Psychologist, 56, 109–118.
Golby, A. J., Gabrieli, J. D. E., Chiao, J. Y., & Eberhardt, J. L. (2001). Differential responses in
the fusiform region to same-race and other-race faces. Nature Neuroscience, 4, 845–850.
Gouvea, A.C., Phillips, C., Kazanina, N., & Poeppel, D. (2010). The linguistic processes
underlying the P600. Language & Cognitive Processes, 25, 149–188.
Gozzi, M., Raymont, V., Solomon, J., Koenigs, M., & Grafman, J. (2009). Dissociable effects of
prefrontal and anterior temporal cortical lesions on stereotypical gender attitudes.
Neuropsychologia, 47, 2125–2132.
Gregory, R. L. (1997). Knowledge in perception and illusion. Philosophical Transactions of the
Royal Society B, 352, 1121–1128.
Grossman, E. D., Battelli, L., & Pascual-Leone, A. (2005). Repetitive TMS over STSp disrupts
perception of biological motion. Vision Research, 45, 2847–2853.
Grossman, E. D., & Blake, R. (2002). Brain areas active during visual perception of biological
motion. Neuron, 35, 1167–1175.
Gutsell, J. N., & Inzlicht, M. (2010). Empathy constrained: Prejudice predicts reduced mental
simulation of actions during observation of outgroups. Journal of Experimental Social
Psychology, 46, 841–845.
Hafer, C. L., & Begue, L. (2005). Experimental research on Just-World Theory: Problems,
developments, and future challenges. Psychological Bulletin, 131, 128–167.
Hagoort, P. (2003). How the brain solves the binding problem for language: A neurocomputa-
tional model of syntactic processing. Neuroimage, 20, S18–S29.
Hamilton, D. L., & Gifford, R. K. (1976). Illusory correlation in interpersonal perception: A cogni-
tive basis of stereotypic judgements. Journal of Experimental Social Psychology, 12, 392–407.
Hamilton, D. L., & Sherman, J. W. (1994). Stereotypes. In R. S. Wyer Jr. & T. K. Srull (Eds.),
Handbook of social cognition (Vol. 2, pp. 1–68). Hillsdale, NJ: Lawrence Erlbaum Associates
Inc.
Hamilton, D. L., & Trolier, T. K. (1986). Stereotypes and stereotyping: An overview of the
cognitive approach. In J. F. Dovidio & S. L. Gaertner (Eds.), Prejudice, discrimination, and
racism (pp. 127–163). Orlando, FL: Academic Press.
260 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Haney, C., Banks, W. C., & Zimbardo, P. G. (1973). Interpersonal dynamics in a simulated
prison. International Journal of Criminology and Penology, 1, 69–97.
Harnad, S. (2005). To cognize is to categorise: Cognition is categorisation. In H. Cohen &
C. Lefebvre (Eds.), Handbook of categorisation in cognitive science (pp. 20–45). Amsterdam:
Elsevier.
Harris, L. T., & Fiske, S. T. (2006). Dehumanizing the lowest of the low. Neuroimaging
responses to extreme out-groups. Psychological Science, 17, 847–853.
Harris, L. T., & Fiske, S. T. (2007). Social groups that elicit disgust are differentially processed
in mPFC. Social Cognitive and Affective Neuroscience, 2, 45–51.
Harris, L. T., & Fiske, S. T. (2009). Social neuroscience evidence for dehumanised perception.
European Review of Social Psychology, 20, 192–231.
Hart, A. J., Whalen, P. J., Shin, L. M., McInerney, S. C., Fischer, H., & Rauch, S. L. (2000).
Differential response in the human amygdala to racial outgroup vs. ingroup face stimuli.
NeuroReport, 11, 2351–2355.
Haslam, S. A., Turner, J. C., Oakes, P. J., McGarty, C., & Reynolds, K. J. (1997). The group as
a basis for emergent stereotype consensus. European Review of Social Psychology, 8, 203–
239.
Hauk, O., Davis, M. H., Kherif, F., & Pulvermuller, F. (2008). Imagery or meaning? Evidence
for a semantic origin of category-specific brain activity in metabolic imaging. European
Journal of Neuroscience, 27, 1856–1866.
Haxby, J. V., Hoffman, E. A., & Gobbini, M. A. (2000). The distributed human neural system
for face perception. Trends in Cognitive Sciences, 4, 223–233.
Haxby, J., Ungerleider, L. G., Clark, V. P., Schouten, J. L., Hoffman, E. A., & Martin, A.
(1999). The effect of face inversion on activity in human neural systems for face and object
perception. Neuron, 22, 189–199.
Hayward, W. G., Rhodes, G., & Schwaninger, A. (2008). An own-race advantage for
components as well as configurations in face recognition. Cognition, 106, 1017–1027.
He, Y., Johnson, M., Dovidio, J., & McCarthy, G. (2009). The relation between race-related
implicit associations and scalp-recorded neural activity evoked by faces from different races.
Social Neuroscience, 4, 426–442.
Henson, R. N. (2005). What can functional imaging tell the experimental psychologist?
Quarterly Journal of Experimental Psychology, 58, 193–233.
Henson, R. N. (2006). What has (neuro)psychology told us about the mind (so far)? A reply to
Coltheart (2006). Cortex, 42, 387–392.
Herrmann, M. J., Schreppel, T., Jager, D., Koehler, S., Ehlis, A. C. & Fallgatter, A. J. (2007).
The other-race effect for face perception: an event-related potential study. Journal of Neural
Transmision, 114, 951–957.
Herrett-Skjellum, J., & Allen, A. (1995). Television programming and sex stereotyping: A meta-
analysis. In B. R. Burleson (Ed.), Communication yearbook 19 (pp. 157–185). Thousand
Oaks, CA: Sage.
Hershler, O., & Hochstein, S. (2005). At first sight: A high-level pop out effect for faces. Vision
Research, 45, 1707–1724.
Hewstone, M. (1990). The ‘ultimate attribution error’? A review of the literature on intergroup
causal attribution. European Journal of Social Psychology, 20, 311–335.
Hilton, J. L., & von Hippel, W. (1996). Stereotypes. Annual Review of Psychology, 47, 237–271.
Hirai, M., Fukushima, H., & Hiraki, K. (2003). An event-related potentials study of biological
motion perception in humans. Neuroscience Letters, 344, 41–44.
Hoffman, E., & Haxby, J. (2000). Distinct representations of eye gaze and identity in the
distributed human neural system for face perception. Nature Neuroscience, 3, 80–84.
Hoffman, C., & Hurst, N. (1990). Gender stereotypes: Perception or rationalization? Journal of
Personality and Social Psychology, 58, 197–208.
STEREOTYPES AND STEREOTYPING 261
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Hole, G. (1994). Configurational factors in the perception of unfamiliar faces. Perception, 23,
65–74.
Huart, J., Corneille, O., & Becquart, E. (2005). Face-based categorisation, context-based
categorisation, and distortions in the recollection of gender ambiguous faces. Journal of
Experimental Social Psychology, 41, 598–608.
Hugenberg, K., Miller, J., & Claypool, H. M. (2007). Categorisation and individuation in the
cross-race recognition deficit: Toward a solution to an insidious problem. Journal of
Experimental Social Psychology, 43, 334–340.
Hutter, R. R. C., Crisp, R. J., Humphreys, G. W., Waters, G. M., & Moffitt, G. (2009).
The dynamics of category conjunctions. Group Processes & Intergroup Relations, 12, 673–
686.
Illes, J., Moser M. A., McCormick, J. B., Racine, E., Blakeslee, S., Caplan, A., et al. (2010).
Neurotalk: Improving the communication of neuroscience research. Nature Reviews
Neuroscience, 11, 61–69.
Ishai, A., Ungerleider, L. G., Martin, A., & Haxby, J. V. (2000). The representation of
objects in the human occipital and temporal cortex. Journal of Cognitive Neuroscience,
12, 35–51.
Itier, R. J., & Taylor, M. J. (2002). Inversion and contrast polarity reversal affect both encoding
and recognition processes of unfamiliar faces: a repetition study using ERPs. NeuroImage,
15, 353–372.
Ito, T. A., Thompson, E., & Cacioppo, J. T. (2004). Tracking the timecourse of social
perception: The effects of racial cues on event-related brain potentials. Personality and Social
Psychology Bulletin, 30, 1267–1280.
Ito, T. A., & Urland, G. R. (2003). Race and gender on the brain: Electrocortical measures of
attention to race and gender of multiply categorisable individuals. Journal of Personality and
Social Psychology, 85, 616–626.
Ito, T. A., & Urland, G. R. (2005). The influence of processing objectives on the perception of
faces: An ERP study of race and gender perception. Cognitive, Affective, & Behavioral
Neuroscience, 5, 21–36.
Jackson, L. A., & Cash, T. F. (1985). Components of gender stereotypes: Their implications for
inferences on stereotypic and nonstereotypic dimensions. Personality and Social Psychology
Bulletin, 11, 326–344.
Jackson, L. A., Sullivan, L. A., & Hodge, C. N. (1993). Stereotype effects on attributions,
predictions, and evaluations: No two social judgements are quite alike. Journal of
Personality and Social Psychology, 65, 69–84.
Jacques, C., & Rossion, B. (2009). The initial representation of individual faces in the right
occipito-temporal cortex is holistic: Electrophysiological evidence from the composite face
illusion. Journal of Vision, 9, 8.1–16.
James, M. S., Johnstone, S. J., & Hayward, W. G. (2001). Event-related potentials, configural
encoding, and feature-based encoding in face recognition. Journal of Psychophysiology, 15,
275–285.
Jarvis, W. B. G., & Petty, R. E. (1996). The need to evaluate. Journal of Personality and Social
Psychology, 70, 172–194.
Johns, M., Cullum, J., Smith, T., & Freng, S. (2008). Internal motivation to respond without
prejudice and automatic egalitarian goal activation. Journal of Experimental Social
Psychology, 44, 1514–1519.
Jokisch, D., Daum, I., Suchan, B., & Troje, N. F. (2005). Structural encoding and recognition
of biological motion: Evidence from event-related potentials and source analysis.
Behavioural Brain Research, 157, 195–204.
Jost, J. T., & Banaji, M. R. (1994). The role of stereotyping in system justification and the
production of false consciousness. British Journal of Social Psychology, 33, 1–27.
262 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Jost, J. T., & Kay, A. C. (2005). Exposure to benevolent sexism and complementary gender
stereotypes: Consequences for specific and diffuse forms of system justification. Journal of
Personality and Social Psychology, 88, 498–509.
Jussim, L., &Fleming, C. (1996). Self-fulfilling prophecies and themaintenance of social stereotypes:
The role of dyadic interactions and social forces. In C. N. Macrae, C. Stangor, & M. Hewstone
(Eds.), Stereotypes and stereotyping (pp. 161–192). New York: Guilford Press.
Kable, J. W., Kan, I. P., Wilson, A., Thompson-Schill, S. L., & Chatterjee, A. (2005).
Conceptual representations of action in the lateral temporal cortex. Journal of Cognitive
Neuroscience, 17, 1855–1870.
Kanwisher, N. (2010). Functional specificity in the human brain: A window into the functional
architecture of the mind. Proceedings of the National Academy of Sciences, 107, 17927–
17932.
Kanwisher, N., McDermott, J., & Chun, M.M. (1997). The fusiform face area: A module in human
extrastriate cortex specialized for face perception. Journal of Neuroscience, 17, 4302–4311.
Kanwisher, N., & Yovel, G. (2006). The fusiform face area: A cortical region specialized for
the perception of faces. Philosophical Transactions of the Royal Society B, 361, 2109–
2128.
Kashima, Y. (2000). Maintaining cultural stereotypes in the serial reproduction of narratives.
Personality and Social Psychology Bulletin, 26, 594-604.
Katz, D., & Braly, K. W. (1933). Racial stereotypes of one hundred college students. Journal of
Abnormal and Social Psychology, 28, 280–290.
Kawakami, K., Dion, K. L., & Dovidio, J. F. (1998). Racial prejudice and stereotype activation.
Personality and Social Psychology Bulletin, 24, 407–416.
Kawakami, K., Young, H., & Dovidio, J .F. (2002). Automatic stereotyping: Category, trait,
and behavioral activations. Personality and Social Psychology Bulletin, 28, 3–15.
Kay, A. C., Jost, J. T., & Young, S. (2005). Victim derogation and victim enhancement as
alternate routes to system justification. Psychological Science, 16, 240–246.
Keehner, M., & Fischer, M. H. (2011). Naive realism in public perceptions of neuroimages.
Nature Reviews Neuroscience, 12, 118–165.
Keehner, M., Mayberry, L., & Fischer, M. H. (2011). Different clues from different views: The
role of image format in public perceptions of neuroimaging results. Psychonomic Bulletin &
Review, 18, 422–428.
Kernahan, C., Bartholow, B. D., & Bettencourt, B. A. (2000). Effects of category-based
expectancy violation on affect-related evaluations: Toward a comprehensive model. Basic
and Applied Social Psychology, 22, 85–100.
Kihlstrom, J. F. (2010). Social neuroscience: The footprints of Phineas Gage. Social Cognition,
28, 757–783.
Kim, J. S., Yoon, H. W., Kim, B. S., Jeun, S. S., Jung, S. L., & Choe, B. Y. (2006). Racial
distinction of the unknown facial identity recognition mechanism by event-related fMRI.
Neuroscience Letters, 397, 279–284.
Klein, S. B., Lax, M. L., Gangi, C. E. (2010). A call for an inclusive approach to the social
cognitive neurosciences. Social Cognition, 28, 748–756.
Knutson, K. M., Mah, L., Manly, C. F., & Grafman, J. (2007). Neural correlates of automatic
beliefs about gender and race. Human Brain Mapping, 28, 915–930.
Kokal, I., & Keysers, C. (2010). Granger causality mapping during joint actions reveals
evidence for forward models that could overcome sensory-motor delays. PLoS ONE, 5,
e13507.
Kosslyn, S. M. (1999). If neuroimaging is the answer, what is the question? Philosophical
Transactions of the Royal Society B, 354, 1283–1294.
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S. F., & Baker, C. I. (2009). Circular analysis
in systems neuroscience—the danger of double dipping. Nature Neuroscience, 12, 535–540.
STEREOTYPES AND STEREOTYPING 263
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Kruglanski, A. W. (1989). Lay epistemics and human knowledge. New York: Plenum.
Kubota, J. T., & Ito, T. A. (2007). Multiple cues in social perception: The time course of processing
race and facial expression. Journal of Experimental Social Psychology, 43, 738–752.
Kuefner, D., Jacques, C., Prieto, E. A., & Rossion, B. (2010). Electrophysiological correlates of
the composite face illusion: Disentangling perceptual and decisional components of holistic
face processing in the human brain. Brain and Cognition, 74, 225–238.
Kuefner, D., Macchi Cassia, V., Picozzi, M., & Bricolo, E. (2008). Do all kids look alike?
Evidence for an other-age effect in adults. Journal of Experimental Psychology: Human
Perception and Performance, 34, 811–817.
Kunda, Z. (1999). Stereotypes. In Z. Kunda (Ed.), Social cognition: Making sense of people (pp.
313–393). Cambridge, MA: MIT Press.
Kunda, Z., & Sherman-Williams, B. (1993). Stereotypes and the construal of individuating
information. Personality and Social Psychology Bulletin, 19, 90–99.
Kunda, Z., & Sinclair, L. (1999). Motivated reasoning with stereotypes: Activation, application,
and inhibition. Psychological Inquiry, 10, 12–22.
Kutas, M., & Federmeier, K. D. (2000). Electrophysiology reveals semantic memory use in
language comprehension. Trends in Cognitive Sciences, 4, 463–470.
Kutas, M., & Hillyard, S.A. (1984). Brain potentials during reading reflect word expectancy and
semantic association. Nature, 307, 161–163.
Landau, E. (2009). Men see bikini-clad women as objects, psychologists say. CNN.com, http://
edition.cnn.com/2009/HEALTH/02/19/women.bikinis.objects/index.html?iref¼allsearchLangton, S. R. H., Law, A. S., Burton, A. M., & Schweinberger, S. R. (2008). Attention capture
by faces. Cognition, 107, 330–342.
Lattner, S., & Friederici, A. D. (2003). Talker’s voice and gender stereotype in human auditory
sentence processing – evidence from event-related brain potentials. Neuroscience Letters,
191–194.
Lau, E., Almeida, D., Hines, P., & Poeppel, D. (2009). A lexical basis for N400 context effects:
Evidence from MEG. Brain & Language, 111, 161–172.
Lau, E., Phillips, C., & Poeppel, D. (2008). A cortical network for semantics: (de)constructing
the N400. Nature Reviews Neuroscience, 9, 920–933.
LeDoux, J. E. (2000). Emotion circuits in the brain. Annual Review of Neuroscience, 23, 155–
184.
Lee, L. C., Andrews, T., Johnson, S. J., Woods, W., Gouws, A., Green, G. G. R., et al. (2010).
Neural responses to rigidly moving faces displaying shifts in social attention investigated
with fMRI and MEG. Neuropsychologia, 48, 477–490.
Lepore, L., & Brown, R. (1997). Category and stereotype activation: Is prejudice inevitable?
Journal of Personality and Social Psychology, 72, 275–287.
Lepore, L., & Brown, R. (1999). Exploring automatic stereotype activation: A challenge to the
inevitability of prejudice. In D. Abrams & M. A. Hogg (Eds.), Social identity and social
cognition (pp. 141–163), London: Blackwell Publishing.
Levin, D. T. (1996). Classifying faces by race: The structure of face categories. Journal of
Experimental Psychology: Learning, Memory, and Cognition, 22, 1364–1382.
Levin, D. T. (2000). Race as a visual feature: Using visual search and perceptual discrimination
tasks to understand face categories and the cross-race recognition deficit. Journal of
Experimental Psychology: General, 129, 559–574.
Levin, D. T., & Banaji, R. (2006). Distortions in the perceived lightness of faces: The role of race
categories. Journal of Experimental Psychology: General, 135, 501–512.
Lewis, M. B., & Edmonds, A. J. (2003). Face detection: Mapping human performance.
Perception, 32, 903–920.
Lewis, M. B., & Edmonds, A. J. (2005). Searching for faces in scrambled scenes. Visual
Cognition, 12, 1309–1336.
264 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Li, J., Delgado, M. R., & Phelps, E. A. (2010). How instructed knowledge modulates the neural
systems of reward learning. Proceedings of the National Academy of Sciences of the United
States of America, 108, 55–60.
Lieberman, M. D., Hariri, A., Jarcho, J. M., Eisenberger, N. I., & Bookheimer, S. Y. (2005). An
fMRI investigation of race-related amygdala activity in African-American and Caucasian-
American individuals. Nature Neuroscience, 8, 720–722.
Liew, S-L., Han, S., & Aziz-Zadeh, L. (2011). Familiarity modulates mirror neuron and
mentalizing regions during intention understanding. Human Brain Mapping, 32, 1986–
1997.
Lippmann, W. (1922). Public opinion. New York: Harcourt, Brace Jovanovitch.
Liu, J., Harris, A., & Kanwisher, N. (2010). Perception of face parts and face configurations: An
fMRI study. Journal of Cognitive Neuroscience, 22, 203–211.
Livingston, R. W., & Brewer, M. B. (2002). What are we really priming? Cue-based versus
category-based processing of facial stimuli. Journal of Personality and Social Psychology, 82,
5–18.
Locke, V., Macrae, C. N., & Eaton, J. L. (2005). Is person categorisation modulated by
exemplar typicality. Social Cognition, 23, 417–428.
Lucas, H. D., Chiao, J. Y., & Paller, K. A. (2011). Why some faces won’t be remembered: Brain
potentials illuminate successful versus unsuccessful encoding for same-race and other-race
faces. Frontiers in Human Neuroscience, 5, doi: 10.3389/fnhum.2011.00020
Ma, Q., Shu, L., Wang, X., Dai, S., & Che, H. (2008). Error-related negativity varies with the
activation of gender stereotypes. Neuroscience Letters, 442, 186–189.
Maass, A., & Arcuri, L. (1996). Language and stereotyping. In C. N. Macrae, C. Stangor, &
M. Hewstone (Eds.), Stereotypes and stereotyping (pp. 193–226). New York: Guilford
Press.
Mackie, D. M., Hamilton, D. L., Susskind, J., & Rosselli, F. (1996). Social psychological
foundations of stereotype formation. In C. N. Macrae, C. Stangor, & M. Hewstone (Eds.),
Stereotypes and stereotyping (pp. 41–78). New York: Guilford Press.
MacLin, O. H., & Malpass, S. (2003). The ambiguous-race face illusion. Perception, 32, 249–
252.
Macrae, C. N., & Bodenhausen, G. V. (2000). Social cognition: Thinking categorically about
others. Annual Review of Psychology, 51, 93–120.
Macrae, C. N., Bodenhausen, G. V., & Milne, A. B. (1995). The dissection of selection in person
perception: Inhibitory processes in social stereotyping. Journal of Personality and Social
Psychology, 69, 397–407.
Macrae, C. N., Bodenhausen, G. V., Milne, A. B., Thorn, T. M. J., & Castelli, L. (1997). On the
activation of social stereotypes: The moderating role of processing objectives. Journal of
Experimental Social Psychology, 33, 471–489.
Macrae, C. N., Bodenhausen, G. V., Schloerscheidt, A. M., & Milne, A. B. (1999). Tales of the
unexpected: Executive processes in person perception. Journal of Personality and Social
Psychology, 76, 200–213.
Macrae, C. N., & Martin, D. (2007). A boy primed Sue: Feature-based processing and person
construal. European Journal of Social Psychology, 37, 793–805.
Macrae, C. N., Milne, A. B., & Bodenhausen, G. V. (1994). Stereotypes as energy-saving
devices: A peek inside the cognitive toolbox. Journal of Personality and Social Psychology,
66, 37–47.
Macrae, C. N., & Quadflieg, S. (2010). Perceiving people. In Fiske, S., Gilbert, D. T., &
Lindzey, G. (Eds.), The handbook of social psychology (5th ed., pp. 428–463). New York:
McGraw-Hill.
Macrae, C. N., Quinn, K. A., Mason, M. F., & Quadflieg, S. (2005). Understanding others: The
face and person construal. Journal of Personality and Social Psychology, 89, 686–695.
STEREOTYPES AND STEREOTYPING 265
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Macrae, C. N., Stangor, C., & Hewstone, M. (1996). Stereotypes and stereotyping. New York:
Guilford Press.
Maddox, K. B. (2004). Perspectives on racial phenotypicality bias. Personality and Social
Psychology Review, 8, 383–401.
Mah, L. W. Y., Arnold, C. M., & Grafman, J. (2005). Deficits in social knowledge following
damage to ventromedial prefrontal cortex. Journal of Neuropsychiatry and Clinical
Neuroscience, 17, 66–74.
Mahon, B. Z., & Caramazza, A. (2009). Concepts and categories: A cognitive neuropsycho-
logical perspective. Annual Review of Psychology, 60, 27–51.
Mansouri, F. A., Tanaka, K., & Buckley, M. J. (2009). Conflict-induced behavioural
adjustment: A clue to the executive functions of the prefrontal cortex. Nature Reviews
Neuroscience, 10, 141–152.
Martin, A. (2007). The representation of object concepts in the brain. Annual Review of
Psychology, 58, 25–45.
Martin, D., & Macrae, C. N. (2007). A face with a cue: Exploring the inevitability of person
categorisation. European Journal of Social Psychology, 37, 806–816.
Martin, W. E., & Thompson, P. N. (2002). Judicial toleration of racial bias in the Minnesota
Justice System. Hamline Law Review, 25, 235–270.
Marques, J. F., Canessa, N., Siri, S., Catricala, E., & Cappa, S. (2008). Conceptual knowledge
in the brain: fMRI evidence for featural organisation. Brain Research, 1194, 90–99.
Mason, M. F., Cloutier, J., & Macrae, C. N. (2006). On construing others: Category and
stereotype activation from facial cues. Social Cognition, 24, 540–562.
Mason, M. F., & Macrae, C. N. (2004). Categorising and individuating others: The neural
substrates of person perception. Journal of Cognitive Neuroscience, 16, 1785–1795.
Mastro, D. (2008). Effects of racial and ethnic stereotyping. In J. Bryant & M. B. Oliver (Eds.),
Media effects. Advances in theory and research (3rd ed., pp. 325–341). London: Routledge,
Taylor & Francis Group.
McCabe, D. P., & Castel, A. D. (2008). Seeing is believing: The effect of brain images on
judgements of scientific reasoning. Cognition, 107, 343–352.
Meiser, T., & Hewstone, M. (2010). Contingency learning and stereotype formation: Illusory
and spurious correlations revisited. European Review of Social Psychology, 21, 285–331.
Meissner, C. A., & Brigham, J. C. (2001). Thirty years of investigating the own-race bias in
memory for faces: A meta-analytic review. Psychology, Public Policy, and Law, 7, 3–35.
Mendes, W. B., Blascovich, J., Hunter, S. B., Lickel, B., & Jost, J. T. (2007). Threatened by the
unexpected: Physiological responses during social interactions with expectancy-violating
partners. Journal of Personality and Social Psychology, 92, 698–716.
Miceli, G., Capasso, R., Daniele, A., Esposito, T., Magarelli, M., & Tomaiuolo, F. (2000).
Selective deficit for people’s names following left temporal damage: An impairment of
domain-specific conceptual knowledge. Cognitive Neuropsychology, 17, 489–516.
Michel, C., Caldara, R., & Rossion, B. (2006). Same-race faces are perceived more holistically
than other race faces. Visual Cognition, 14, 55–73.
Michel, C., Rossion, B., Han, J., Chung, C-S., & Caldara, R. (2006). Holistic processing is finely
tuned for faces of one’s own race. Psychological Science, 17, 608–615.
Michels, L., Lappe, M., & Vaina, L. M. (2005). Visual areas involved in the perception of
human movement from dynamic form analysis. NeuroReport, 16, 1037–1041.
Milgram, S. (1963). Behavioral study of obedience. Journal of Abnormal and Social Psychology,
67, 371–378.
Mitchell, J. P. (2009). Social psychology as a natural kind. Trends in Cognitive Sciences, 13, 246–
251.
Molnar-Szakacs, I., Wu, A. D., Robles, F. J., & Iacobini, M. (2007). Do you see what I mean?
Corticospinal excitability during observation of culture-specific gestures. Plos One, 7, e626.
266 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Montague, P. R., Berns, G. S., Cohen, J. D., McClure, S. M., Pagnoni, G., Dhamala, M., et al.
(2002). Hyperscanning: Simultaneous fMRI during linked social interactions. NeuroImage,
16, 1159–1164.
Moskowitz, G. B., Gollwitzer, P. M., Wasel, W., & Schaal, B. (1999). Preconscious control of
stereotype activation through chronic egalitarian goals. Journal of Personality and Social
Psychology, 77, 167–184.
Mouchetant-Rostaing, Y., & Giard, M. H. (2003). Electrophysiological correlates of age and
gender perception on human faces. Journal of Cognitive Neuroscience, 15, 900–910.
Mullen, B., & Johnson, C. (1990). Distinctiveness-based illusory correlations and stereotyping:
A meta-analytic integration. British Journal of Social Psychology, 29, 11–27.
Musher-Eizenman, D. R., Holub, S. C., Miller, A. B., Goldstein, S. E., & Edwards-Leeper, L.
(2004). Body size stigmatization in preschool children: The role of control attributions.
Journal of Pediatric Psychology, 29, 613–620.
Nelson, C. A. (2001). The development and neural bases of face recognition. Infant and Child
Development, 10, 3–18.
Ng, M., Ciaramitaro, V. M., Anstis, S., Boynton, G. M., & Fine, I. (2006). Selectivity for the
configural cues that identify the gender, ethnicity, and identity of faces in human cortex.
Proceedings of the National Academy of Sciences, 103, 19552–19557.
Nichols, D. F., Betts, L. R., & Wilson, H. R. (2010). Decoding of faces and face
components in face-sensitive human visual cortex. Frontiers in Psychology, doi: 10.3389/
fpsyg.2010.00028.
O’Craven, K., & Kanwisher, N. (2000). Mental imagery of faces and places activates corresponding
stimulus-specific brain regions. Journal of Cognitive Neuroscience, 12, 1013–1023.
Ofan, R. H., Rubin, N., & Amodio, D. M. (2011). Seeing race: N170 responses to race and their
relation to automatic racial attitudes and controlled processing. Journal of Cognitive
Neuroscience, 23, 3152–3161.
Olsson, A., & Phelps, E. A. (2007). Social learning of fear. Nature Neuroscience, 10, 1095–1102.
Oppliger, P. A. (2006). Effects of gender stereotyping on socialization. In R. W. Preiss, B. M.
Gayle, N. Burrell, M. Allen, & J. Bryant (Eds.),Mass media effects research. Advances through
meta-analysis (1st ed., pp. 199–214). Mahwah, NJ: Lawrence Erlbaum Associates Inc.
Orlov, T., Makin, T. R., & Zohary, E. (2010). Topographic representation of the human body
in the occipitotemporal cortex. Neuron, 68, 586–600.
Osterhout, L., Bersick, M., & McLaughlin, J. (1997). Brain potentials reflect violations of
gender stereotypes. Memory & Cognition, 25, 273–285.
Paluck, E. L., & Green, D. P. (2009). Prejudice reduction: What works? A review and
assessment of research and practise. Annual Review of Psychology, 60, 339–367.
Park, B., & Rothbart, M. (1982). Perception of outgroup homogeneity and levels of social
categorisation: Memory for the subordinate attributes of ingroup and outgroup members.
Journal of Personality and Social Psychology, 42, 1051–1068.
Pascalis, O., & Kelly, D. J. (2009). The origins of face processing in humans: Phylogeny and
ontogeny. Perspectives on Psychological Science, 4, 200–209.
Patchen, M., Davidson, J. D., Hofman, G., & Brown, W. R. (1977). Determinants of students’
interracial behavior and opinion change. Sociology of Education, 50, 55–75.
Payne, B. K. (2001). Prejudice and perception: The role of automatic and controlled processes in
misperceiving a weapon. Journal of Personality and Social Psychology, 81, 181–192.
Peelen, M. V., & Caramazza, A. (2010). What body parts reveal about the organisation of the
brain. Neuron, 68, 331–333.
Peelen, M. V., & Downing, P. E. (2007). The neural basis of visual body perception. Nature
Reviews Neuroscience, 8, 636–648.
Perry, D. B., & Bussey, K. (1979). The social learning theory of sex differences: Imitation is alive
and well. Journal of Personality and Social Psychology, 37, 1699–1712.
STEREOTYPES AND STEREOTYPING 267
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Phelps, E., Cannistraci, C., & Cunningham, W. (2003). Intact performance on an
indirect measure of race bias following amygdala damage. Neuropsychologia, 41, 203–
208.
Phelps, E. A., O’Connor, K. J., Cunningham, W. A., Funayama, E. S., Gatenby, J C., Gore, J.
C., et al. (2000). Performance on indirect measures of race evaluation predicts amygdala
activation. Journal of Cognitive Neuroscience, 12, 729–738.
Pitcher, D., Dilks, D. D., Saxe, R. R., Triantafyllou, C., & Kanwisher, N. (2011). Differential
selectivity for dynamic versus static information in face-selective cortical regions. Neuro-
Image, 56, 2356–2363.
Pitcher, D., Walsh, V., & Duchaine, B. (2011). The role of the occiptial face area in the cortical
face perception network. Experimental Brain Research, 209, 481–493.
Pitcher, D., Walsh, V., Yovel, G., & Duchaine, B. (2007). TMS evidence for the
involvement of the right occipital face area in early face processing. Current Biology, 17,
1568–1573.
Poldrack, R. A. (2006). Can cognitive processes be inferred from neuroimaging data? Trends in
Cognitive Sciences, 10, 59–63.
Poldrack, R. A. (2008). The role of fMRI in cognitive neuroscience: Where do we stand?
Current Opinion in Neurobiology, 18, 223–227.
Poldrack, R. A., Fletcher, P. C., Henson, R. N., Worsley, K. J., Brett, M., & Nichols, T. E.
(2008). Guidelines for reporting an fMRI study. NeuroImage, 40, 409–414.
Pourtois, G., Peelen, M., Spinelli, L., Seeck, M., & Vuilleumier, P. (2007). Direct intracranial
recording of body-selective responses in human extrastriate visual cortex. Neuropsychologia,
45, 2621–2625.
Proverbio, A. M., Riva, F., Martin, E., & Zani, A. (2010). Neural markers of opposite-sex bias
in face processing. Frontiers in Psychology, 1, e169.
Quadflieg, S., Flannigan, N., Waiter, G. D., Rossion, B., Wig, S. G., Turk, D. J., et al. (2011).
Stereotype-based modulation of person perception. NeuroImage, 57, 549–557.
Quadflieg, S., & Macrae, C. N. (2011). Neuroimaging methods in social cognition. In K. C.
Klauer, C. Stahl, & A. Voss (Eds.), Cognitive methods in social psychology (pp. 340–366).
New York: Guilford Press.
Quadflieg, S., & Macrae, C. N. (in press). Impression formation. In V. S. Ramachandran (Ed.),
Encyclopedia of human behavior (2nd ed.). Oxford, UK: Elsevier.
Quadflieg, S., & Rossion, B. (in press). When perception and attention collide: Neural
processing in EBA and FBA. Commentary in Cognitive Neuroscience.
Quadflieg, S., Turk, D. J., Waiter, G. D., Mitchell, J. P., Jenkins, A. C., & Macrae, C. N. (2009).
Exploring the neural correlates of social stereotyping. Journal of Cognitive Neuroscience, 21,
1560–1570.
Quinn, K. A., & Macrae, C. N. (2005). Categorising others: The dynamics of person construal.
Journal of Personality and Social Psychology, 88, 467–479.
Racine, E., Bar-Illan, O., & Illes, J. (2005). fMRI in the public eye. Nature Reviews
Neuroscience, 6, 159–164.
Ramm, B., Cummins, T., & Slaughter, V. (2010). Specifying the human body configuration.
Visual Cognition, 18, 898–919.
Reed, C. L., Stone, V., Bozova, S., & Tanaka, J. (2003). The body inversion effect. Psychological
Science, 14, 302–308.
Reed, C. L., Stone, V. E., Grubb, J. D., & McGoldrick, J. E. (2006). Turning configural
processing upside down: Part and whole body postures. Journal of Experimental Psychology:
Human Perception and Performance, 32, 73–87.
Remedios, J. D., Chasteen, A. L., Rule, N. O., & Plaks, J. E. (2011). Impressions at the
intersection of ambiguous and obvious social categories: Does gayþBlack¼ likable?
Journal of Experimental Social Psychology, 47, 1312–1315.
268 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Richeson, J. A., Baird, A. A., Gordon, H. L., Heatherton, T. F., Wyland, C. L., Trawalter, S.,
et al. (2003). An fMRI investigation of the impact of interracial contact on executive
function. Nature Neuroscience, 6, 1323–1328.
Rivera, L. O., Arms-Chavez, C. J., & Zarate, M. A. (2009). Resource dependent effects during
sex categorisation. Journal of Experimental Social Psychology, 45, 908–912.
Rizzolatti, G., Fogassi, L., & Gallese, V. (2001). Neurophysiological mechanisms
underlying the understanding and imitation of action. Nature Reviews Neuroscience, 2,
661–670.
Ro, T., Friggel, A., & Lavie, N. (2007). Attentional biases for faces and body parts. Visual
Cognition, 15, 322–348.
Roepstorff, A. (2004). Mapping brain mappers: An ethnographic coda. In R. S. J. Frackowiak
et al. (Eds.), Human brain function (2nd ed., pp. 1105–1117). London: Elsevier.
Ronquillo, J., Denson, D. F., Lickel, B., Lu, Z-L., Nandy, A., & Maddox, K. B. (2007). The
effects of skin tone on race-related amygdala activity: An fMRI investigation. Social
Cognitive and Affective Neuroscience, 2, 39–44.
Rossion, B., & Gauthier, I. (2002). How does the brain process upright and inverted faces?
Behavioral and Cognitive Neuroscience Reviews, 1, 62–74.
Rossion, B., & Jacques, C. (2008). Does physical interstimulus variance account for early
electrophysiological face sensitive responses in the human brain? Ten lessons on the N170.
NeuroImage, 39, 1959–1979.
Rule, N. O., & Ambady, N. (2010). Democrats and Republicans can be differentiated from their
faces. PLoS ONE, 5, e8733.
Sagar, H. A., & Schofield, J. W. (1980). Racial and behavioral cues in Black and White
children’s perceptions of ambiguously aggressive acts. Journal of Personality and Social
Psychology, 39, 590–598.
Sagiv, N., & Bentin, S. (2001). Structural encoding of human and schematic faces: Holistic and
part-based processes. Journal of Cognitive Neuroscience, 13, 937–951.
Santos, I. M., & Young, A. W. (2005). Exploring the perception of social characteristics in faces
using the isolation effect. Visual Cognition, 12, 213–247.
Saver, J. L., & Damasio, A. R. (1991). Preserved access and processing of social knowledge in a
patient with acquired sociopathy due to ventromedial frontal damage. Neuropsychologia, 29,
1241–1249.
Saxe, R. (2006). Uniquely human social cognition. Current Opinion in Neurobiology, 16, 235–239.
Saxe, R., Xiao, D. K., Kovacs, G., Perrett, D. I., & Kanwisher, N. (2004). A region of right
posterior superior temporal sulcus responds to observed intentional actions. Neuropsycho-
logia, 42, 1435–1446.
Saygin, A. P. (2007). Superior temporal and premotor brain areas necessary for biological
motion perception. Brain, 130, 2452–2461.
Schippers, M. B., Roebroeck, A., Renken, R., Nanetti, L., & Keysers, C. (2010). Mapping the
information flow from one brain to another during gestural communication. Proceedings of
the National Academy of Sciences, 107, 9388–9393.
Schneider, D. J. (1996). Modern stereotype research: unfinished business. In C. N. Macrae, C.
Stangor, & M. Hewstone (Eds.), Stereotypes and stereotyping (pp. 419–453). New York:
Guilford Press.
Schreiber, D., & Iacoboni, M. (in press). Huxtables on the brain: An fMRI study of race and
norm violation. Political Psychology.
Schwarzlose, R. F., Swisher, J. D., Dang, S., & Kanwisher, N. (2008). The distribution of
category and location information across object-selective regions of visual cortex.
Proceedings of the National Academy of Scienes, 105, 4447–4452.
Sechrist, G. B., & Stangor, C. (2001). Perceived consensus influences intergroup behavior and
stereotype accessibility. Journal of Personality and Social Psychology, 80, 645–654.
STEREOTYPES AND STEREOTYPING 269
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Shapiro, J. R. (2011). Different groups, different threats: A multi-threat approach to
the experience of stereotype threats. Personality and Social Psychology Bulletin, 37, 464–
480.
Sherman, J. W. (2010). Social neuroscience and its contribution to social psychological theory:
Introduction to the special issue. Social Cognition, 28, 663–666.
Sherman, J.W.,Kruschke, J.K., Sherman, S. J., Percy, E. J., Petrocelli, J.V.,&Conrey, F.R. (2009).
Attentional processes in stereotype formation: A common model for category accentuation and
illusory correlation. Journal of Personality and Social Psychology, 96, 305–323.
Sherman, J. W., Macrae, C. N., & Bodenhausen, G. V. (2000). Attention and stereotyping:
Cognitive constraints on the construction of meaningful social impressions. European
Review of Social Psychology, 11, 145–175.
Sherman, J. W., Stroessner, S. J., Conrey, F. R., & Azam, O. A. (2005). Prejudice and
stereotype maintenance processes: Attention, attribution, and individuation. Journal of
Personality and Social Psychology, 89, 607–622.
Sinclair, L., & Kunda, Z. (1999). Reactions to a black professional: motivated inhibition and
activation of conflicting stereotypes. Journal of Personality and Social Psychology, 77, 885–904.
Sinclair, L., & Kunda, Z. (2000). Motivated stereotyping of women: She’s fine if she praised me but
incompetent if she criticized me. Personality and Social Psychology Bulletin, 26, 1329–1342.
Sinclair, S., Dunn, E., & Lowery, B. S. (2005). The relationship between parental racial
attitudes and children’s implicit prejudice. Journal of Experimental Social Psychology, 41,
283–289.
Slaughter, V., Heron-Delaney, M., & Christie, T. (2011). Developing expertise in human body
perception. In V. Slaughter & C. Brownell (Eds.), Early development of body representations
(pp. 81–100). New York: Cambridge University Press.
Smith, S. L., & Granados, A. D. (2008). Content patterns and effects surrounding sex-role
stereotyping on television and film. In J. Bryant &M. B.Oliver (Eds.),Media effects. Advances in
theory and research (3rd ed., pp. 342–361). London: Routledge, Taylor & Francis Group.
Sorger, B., Goebel, R., Schiltz, C., & Rossion, B. (2007). Understanding the functional
neuroanatomy of prosopagnosia. NeuroImage, 35, 836–852.
Stahl, J., Wiese, H., & Schweinberger, S. R. (2010). Learning task affects ERP correlates of
the own-race bias, but not recognition memory performance. Neuropsychologia, 48, 2027–
2040.
Stangor, C. (Ed.). (2000). Volume overview. In Stereotypes and prejudice: Essential readings
(pp. 1–16). New York: Psychology Press.
Stangor, C., & Leary, S. P. (2006). Intergroup beliefs: Investigations from the social side.
Advances in Experimental Social Psychology, 38, 243–281.
Stangor, C., & Schaller, M. (1996). Stereotypes as individual and collective representations. In
C. N. Macrae, C. Stangor, & M. Hewstone (Eds.), Stereotypes and stereotyping (pp. 3–40).
New York: Guilford Press.
Stanley, D., Phelps, E., & Banaji, M. (2008). The neural basis of implicit attitudes. Current
Directions in Psychological Science, 17, 164–170.
Steele, C. M. (1997). A threat in the air: How stereotypes shape intellectual identity and
performance. American Psychologist, 52, 613–629.
Stephan, W. G., & Stephan, C. W. (1993). Cognition and affect in stereotyping: Parallel
interactive networks. In D. M. Mackie & D. H. Hamilton (Eds.), Affect, cognition, and
stereotyping: Interactive processes in group perception (pp. 111–136). San Diego: Academic
Press.
Sterzer, P., Kleinschmidt, A., & Rees, G. (2009). The neural bases of multistable perception.
Trends in Cognitive Sciences, 13, 310–318.
Tanaka, J. W., Kiefer, M., & Bukach, C. M. (2004). A holistic account of the own-race effect in
face recognition: Evidence from a crosscultural study. Cognition, 93, B1–B9.
270 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Tanaka, J. W., & Pierce, L. J. (2009). The neural plasticity of other-race face recognition.
Journal of Cognitive and Behavioral Neuroscience, 9, 122–131.
Tanskanen, T., Naesaenen, R., Montez, T., Paeaellysaho, J., & Hari, R. (2005). Face
recognition and cortical responses show similar sensitivity to noise spatial frequency.
Cerebral Cortex, 15, 526–524.
Taylor, J. C., Roberts, M. V., Downing, P. E., & Thierry, G. (2010). Functional
characterisation of the extrastriate body area based on the N1 ERP component. Brain &
Cognition, 73, 153–159.
Taylor, J. C., Wiggett, A. J., & Downing, P. E. (2007). FMRI analysis of body and body part
representations in the extrastriate and fusiform body areas. Journal of Neurophysiology, 98,
1626–1633.
Theeuwes, J., & Van der Stigchel, S. (2006). Faces capture attention: Evidence from inhibition
of return. Visual Cognition, 13, 657–665.
Thompson, J. C., Hardee, J. E., Panayiotou, A., Crewther, D., & Puce, A. (2007). Common and
distinct brain activation to viewing dynamic sequences of face and hand movements.
NeuroImage, 37, 966–973.
Thompson, M. S., Judd, C. M., & Park, B. (2000). The consequences of communicating social
stereotypes. Journal of Experimental Social Psychology, 36, 567–599.
Tormala, Z. L., & Petty, R. E. (2001). On-line versus memory-based processing: The role of
‘‘need to evaluate’’ in person perception. Personality and Social Psychology Bulletin, 27,
1599–1612.
Trope, Y., & Thompson, E. (1997). Looking for the truth in all the wrong places? Asymmetric
search of individuating information about stereotyped group members. Journal of
Personality and Social Psychology, 73, 229–241.
Ungerleider, L.G. & Mishkin, M. (1982). Two cortical visual systems. In D. J. Ingle, M. A.
Goodale, & R. J. W. Mansfield (Eds.), Analysis of visual behavior (pp. 549–586). Cambridge,
MA: MIT Press.
Urgesi, C., Berlucchi, G., & Aglioti, S. M. (2004). Magnetic stimulation of extrastriate
body area impairs visual processing of non-facial body parts. Current Biology, 14, 2130–
2134.
Urgesi, C., Calvo-Merino, B., Haggard, P., & Aglioti, S. M. (2007). Transcranial magnetic
stimulation reveals two cortical pathways for visual body processing. The Journal of
Neuroscience, 27, 8023–8030.
Urgesi, C., Candidi, M., Ionta, S., & Aglioti, S. M. (2007). Representation of body identity and
body actions in extrastriate body area and ventral premotor cortex. Nature Neuroscience, 10,
30–31.
Uttal, W. R. (2003). The new phrenology. Cambridge, MA: MIT Press.
Valentine, T. (1991). A unified account of the effects of distinctiveness, inversion, and race in
face recognition. Quarterly Journal of Experimental Psychology, 43, 161–204.
Valentine, T., & Endo, M. (1992). Towards an exemplar model of face processing: The effects of
race and distinctiveness. Quarterly Journal of Experimental Psychology: Human Experi-
mental Psychology, 44, 671–703.
Van Bavel, J. J., Packer, D. J., & Cunningham, W. A. (2008). The neural substrates of in-group
bias: A functional magnetic resonance imaging investigation. Psychological Science, 19,
1131–1139.
Van Bavel, J. J., Packer, D. J., & Cunningham, W. A. (2011). Modulation of fusiform face area
following minimal exposure to motivationally relevant faces. Journal of Cognitive
Neuroscience, 23, 3343–3354.
Van Knippenberg, A., & Dijksterhuis, A. (2000). Social Categorisation and stereotyping: A
functional perspective. In W. Stroebe & M. Hewstone (Eds.), European review of social
psychology, 11 (pp. 105–144). Chichester, UK: Wiley.
STEREOTYPES AND STEREOTYPING 271
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Van Overwalle, F. (2009). Social cognition and the brain: a meta-analysis. Human Brain
Mapping, 30, 829–858.
Vizioli, L., Foreman, K., Rousselet, G. A., & Caldara, R. (2010). Inverting faces elicits sensitivity
to race on the N170 component: A cross-cultural study. Journal Of Vision, 10, 1–23.
Vizioli, L., Rousellet, G., & Caldara, R. (2010). Neural repetition suppression to identity is
abolished by other-race faces. Proceedings of the National Academy of Sciences, 107, 20081–
20086.
Walker, P. M., Silvert, L., Hewstone, M., & Nobre, A. C. (2008). Social contact and other-race
face processing in the human brain. Social, Cognitive and Affective Neuroscience, 3, 16–25.
Walton, G., & Cohen, L. (2007). A question of belonging: Race, social fit, and achievement.
Journal of Personality and Social Psychology, 92, 82–96.
Wang, L., Ma, Q., Song, Z., Shi, Y., Wang, Y., & Pfotenhauer, L. (2010). N400 and the
activation of prejudice against rural migrant workers in China. Brain Research, 1375, 103–
110.
Weigelt, S., Koldewyn, K., & Doehrmann, O. (2010). Cross-category adaptation reveals tight
coupling of face and body perception. Journal of Neurophysiology, 104, 581–583.
Weiner, K. S., & Grill-Spector, K. (2010). Sparsely-distributed organisation of face and limb
activations in human ventral temporal cortex. NeuroImage, 52, 1559–1573.
Weisberg, D. S. (2008). Caveat lector: The presentation of neuroscience information in the
popular media. The Scientific Review of Mental Health Practice, 6, 51–56.
Weisberg, D. S., Keil, F. C., Goodstein, J., Rawson, E., & Gray, J. R. (2008). The seductive
allure of neuroscience explanations. Journal of Cognitive Neuroscience, 20, 470–477.
Whalen, P. J. (1998). Fear, vigilance, and ambiguity: Initial neuroimaging studies of the human
amygdala. Current Directions in Psychological Science, 7, 177–188.
Wheeler, M. E., & Fiske, S. T. (2005). Controlling racial prejudice. Psychological Science, 16, 56–62.
White, K. R., Crites, S. L. Jr., Taylor, J. H., & Corral, G. (2009). Wait, what? Assessing
stereotype incongruencies using the N400 ERP component. Social and Affective Cognitive
Neuroscience, 4, 191–198.
Wilder, D. A. (1978). Perceiving persons as a group: Effects on attributions of causality and
beliefs. Social Psychology, 1, 13–23.
Wilkinson, D., & Halligan, P. (2004). The relevance of behavioral measures for functional-
imaging studies of cognition. Nature Reviews Neuroscience, 5, 67–73.
Willingham, D. T., & Dunn, E. W. (2003). What neuroimaging and brain localization can do,
cannot do, and should not do for social psychology. Journal of Personality and Social
Psychology, 85, 662–671.
Wilms, M., Schilbach, L., Pfeifer, U., Bente, G., Fink, G. R., & Vogeley, K. (2010). It’s in your
eyes. Using gaze feedback to create truly interactive paradigms for social cognitive and
affective neuroscience. Social Cognitive and Affective Neuroscience, 5, 98–107.
Wilson, T. D., Lindsey, S., & Schooler, T. Y. (2000). A model of dual attitudes. Psychological
Review, 107, 101–126.
Wittenbrink, B., Judd, C. M., & Park, B. (1997). Evidence for racial prejudice at the implicit
level and its relationship with questionnaire measures. Journal of Personality and Social
Psychology, 72, 262–274.
Wittenbrink, B., Judd, C. M., & Park, B. (2001a). Spontaneous prejudice in context: Variability in
automatically activated attitudes. Journal of Personality and Social Psychology, 81, 815–827.
Wittenbrink, B., Judd, C. M., & Park, B. (2001b). Evaluative versus conceptual judgements in
automatic stereotyping and prejudice. Journal of Experimental Social Psychology, 37, 244–252.
Wright, C. I., Negreira, A., Gold, A. L., Britton, J. C., Williams, D., & Feldman Barrett, L.
(2008). Neural correlates of novelty and face–age effects in young and elderly adults.
NeuroImage, 42, 956–968.
272 QUADFLIEG AND MACRAE
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014

Yarkoni, T., Poldrack, R. A., Van Essen, D. C., & Wager, T. D. (2010). Cognitive neuroscience
2.0: Building a cumulative science of human brain function. Trends in Cognitive Sciences, 14,
489–496.
Yin, R. K. (1969). Looking at upside-down faces. Journal of Experimental Psychology, 81, 141–145.
Yovel, G., & Kanwisher, N. (2005). The neural basis of the behavioral face-inversion effect.
Current Biology, 15, 2256–2262.
Yzerbyt, V., & Corneille, O. (2005). Cognitive process: Reality constraints and integrity
concerns in social perception. In J. F. Dovidio, P. Glick, & L. A. Rudman (Eds.), On the
nature of prejudice (pp. 175–191). Malden, MA: Blackwell Publishing.
Yzerbyt, V., Rocher, S., & Schadron, G. (1997). Stereotypes as explanations: A subjective
essentialistic view of group perception. In R. Spears, P. J. Oakes, N. Ellemers, & S. A.
Haslam (Eds.), The social psychology of stereotyping and group life (pp. 20–50). Oxford, UK/
Cambridge, MA: Blackwell.
Zahn, R., Moll, J., Krueger, F., Huey, E. D., Garrido, G., & Grafman, J. (2007). Social
concepts are represented in the superior anterior temporal cortex. Proceedings of the
National Academy of Sciences, 104, 6430–6435.
Zarate, M. A., Stoever, C. J., MacLin, M. K., & Chavez, C. J. (2008). Neurocognitive
underpinnings of face perception: Further evidence of distinct person and group perception
processes. Journal of Personality and Social Psychology, 94, 108–115.
Zieber, N., Bhatt, R. S., Hayden, A., Kangas, A., Collins, R., & Bada, H. (2010). Body
representation in the first year of life. Infancy, 15, 534–544.
Zirkel, S. (2004). What will you think of me? Racial integration, peer relationships and
achievement among White students and students of color. Journal of Social Issues, 60, 57–74.
STEREOTYPES AND STEREOTYPING 273
Dow
nloa
ded
by [
Uni
vers
ity o
f C
alif
orni
a D
avis
] at
15:
52 1
8 O
ctob
er 2
014