stable carbon isotope biogeochemistry of propionate and acetate in methanogenic soils and lake...
TRANSCRIPT

Accepted Manuscript
Stable carbon isotope biogeochemistry of propionate and acetate in methano-
genic soils and lake sediments
R. Conrad, P. Claus, A. Chidthaisong, Y. Lu, A. Fernandez Scavino, Y. Liu, R.
Angel, P.E. Galand, P. Casper, F. Guerin, A. Enrich-Prast
PII: S0146-6380(14)00104-1
DOI: http://dx.doi.org/10.1016/j.orggeochem.2014.03.010
Reference: OG 3116
To appear in: Organic Geochemistry
Received Date: 22 November 2013
Revised Date: 11 March 2014
Accepted Date: 12 March 2014
Please cite this article as: Conrad, R., Claus, P., Chidthaisong, A., Lu, Y., Fernandez Scavino, A., Liu, Y., Angel,
R., Galand, P.E., Casper, P., Guerin, F., Enrich-Prast, A., Stable carbon isotope biogeochemistry of propionate and
acetate in methanogenic soils and lake sediments, Organic Geochemistry (2014), doi: http://dx.doi.org/10.1016/
j.orggeochem.2014.03.010
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers
we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and
review of the resulting proof before it is published in its final form. Please note that during the production process
errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.

1
Stable carbon isotope biogeochemistry of propionate and acetate in
methanogenic soils and lake sediments
R. Conrada*, P. Clausa, A. Chidthaisongb, Y. Luc, A. Fernandez Scavinod, Y. Liue, R. Angela, 1, P.E. Galandf, P. Casperg, F. Guerinh, A. Enrich-Prasti
a Max-Planck-Institute for Terrestrial Microbiology, Karl-von-Frisch-Str.10, 35043 Marburg,
Germany b Joint Graduate School of Energy and Environment, King Mongkut´s University of
Technology Thonburi, and Center for Energy Technology and Environment, Ministry of Education, Bangkok, Thailand
c College of Resources and Environmental Sciences, China Agricultural University, Beijing, China
d Departamento de Biosciencias, Facultad de Quimica, Universidad de Republica, General Flores 2124, Montevideo, Uruguay
e Key Laboratory of Tibetan Environment Changes and Land Surface Processes, Institute of Tibetan Plateau Research, Chinese Academy of Science, 100085 Beijing, China
f UPMC, Université Paris 06, and CNRS, UMR 8222, Laboratoire d’Ecogéochimie des Environnements Benthiques-LECOB, Observatoire Océanologique de Banyuls, Banyuls/mer 66650, France
g Leibniz-Institute of Freshwater Ecology and Inland Fisheries, Department of Experimental Limnology, alte Fischerhuette 2, 16775 Stechlin, Germany
h Géosciences Environnement Toulouse (GET), Observatoire Midi Pyréneés, Université de Toulouse, CNRS, IRD, 14 avenue E. Belin, 31400 Toulouse, France
i Biogeochemistry Laboratory, Institute of Biology, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
1 Present address: University of Vienna, Division of Microbial Ecology, Althanstr.14, 1090
Vienna, Austria ([email protected]). *Corresponding author: Tel.: +49-6421-178801; fax : +49-6421-178809. E mail address: [email protected] (Ralf Conrad)

2
ABSTRACT
In anoxic environments, degradation of organic matter (OM) results in strong fractionation of
carbon isotopes, with formation of 13C-depleted CH4. Propionate and acetate are important
products of OM fermentation. Propionate is further fermented to acetate. Acetate is a direct
precursor of CH4, the remainder usually being produced from H2-mediated CO2 reduction.
There is a paucity of data for the turnover of acetate and, even more so, propionate. We
therefore analyzed the δ13C values of organic carbon, propionate, acetate and the methyl (Me)
group of acetate (acetate-Me) during the production of CH4 in anoxic incubations of various
flooded and non-flooded soils and various lake sediments. Incubation in the presence of
methylfluoride (CH3F), which inhibits CH4 production from acetate, allowed exclusion of
isotope effects during aceticlastic methanogenesis. Despite the variation inherent in the wide
diversity of sample type and origin, the data collectively showed that the δ13C value of acetate
was only marginally different (-2 ± 5‰) from that of OM, while propionate was depleted in
13C relative to total acetate (-6 ± 5‰). Acetate-Me was generally depleted in 13C relative to
total acetate (-8 ± 5‰). Thus, isotopic enrichment factors during the degradation of OM to
total propionate and acetate were much smaller than those during hydrogenotrophic and
aceticlastic methanogenesis or the intramolecular difference in δ13C between the carboxyl
(CO2H) and Me of acetate, so that the δ13C value of OM may be used as a proxy when data for
acetate are not available.
Key words: acetate; propionate; methane; organic matter; fermentation; isotope fractionation;
anoxic soil; lake sediment

3
1. Introduction
The cycling of carbon is a crucial process in ecosystems. The essential processes are
primary production and degradation of organic matter (OM), the latter being catalyzed mainly
by microorganisms. Degradation also occurs in anoxic environments. There, OM is degraded
by a complex microbial community (Conrad and Frenzel, 2002; Megonigal et al., 2003; Zinder,
1993). If there is sufficient supply of oxidants such as SO42- (in marine environments) or Fe3+
(in a few soils and freshwater environments), OM is eventually degraded to CO2 as sole
product. However, when the environment is depleted in SO42- and Fe3+, OM is degraded to
CH4 plus CO2 in most anoxic environments (and to some extent marine ones). The formation
of CH4 is the last step in the degradation, while the initial steps, hydrolysis and fermentation of
the polymeric OM, are mainly identical to those operating when SO42- or Fe3+ are available. In
these initial fermentation steps, acetate is always formed and frequently accumulates, at least
temporarily, in the environment. Besides acetate, propionate is also an important intermediate
in OM breakdown (Boschker et al., 2001; Glissmann and Conrad, 2000; Heuer et al., 2010).
Therefore, the turnover of acetate and the turnover of propionate are important for the
anaerobic degradation process and the formation of CH4 and CO2.
Methanogenic degradation of OM involves several steps that are performed by different
groups of microorganisms (McInerney and Bryant, 1981; Zinder, 1993). Degradation is
initiated by hydrolysis of polymers to oligomers and then to monomers, which become
fermentatively degraded to CO2, H2 and acetate, as well as other simple carbon compounds
such as fatty acids (e.g. propionate) and alcohols. The latter are further fermented (secondary
fermentation) to acetate, CO2 and H2, which are then finally converted to CH4. Furthermore,
acetate is not only a product of these fermentation reactions, but can also be the sole product
of fermentation and can also be formed by chemolithotrophic acetogenesis converting H2 and
CO2 to acetate (Dolfing, 1988; Drake and Küsel, 2003). Overall, acetate often contributes

4
more than two thirds of total CH4 production, the remainder being produced from the
reduction of CO2 (Conrad, 1999).
Each of these steps theoretically exhibits a characteristic stable isotope fractionation.
Production of CH4 is characterized by strong fractionation, such that CH4 is always depleted in
13C relative to the substrate from which it is formed (Whiticar et al., 1986). Methanogenesis
from acetate (aceticlastic) or from CO2 plus H2 (hydrogenotrophic) are the two most common
methanogenic pathways in freshwater sediments and anoxic soils over a wide temperature
range (Conrad et al., 2009c; Nozhevnikova et al., 2007). Since they exhibit quite distinct
carbon isotope fractionation factors, the isotopic signature (δ13C) of CH4 as compared with
those of the substrates (CO2, acetate) allows determination of the relative contribution of
aceticlastic vs. hydrogenotrophic methanogenesis to total CH4 production using mass balance
calculations (Conrad, 2005). Isotopic signatures of CH4 and CO2 in environmental samples and
microbial enrichment cultures have frequently been reported in the literature, with many
different types of OM as primary substrates, including hydrocarbons and long-chain fatty
acids (Feisthauer et al., 2010; Morris et al., 2012). However, while the precise analysis of the
carbon isotopic composition of gaseous CO2 and CH4 and of bulk OM has become routine
through the use of isotope ratio mass spectrometry (IRMS) and/or laser spectrometry, the
corresponding analysis of acetate (or propionate) has been carried out less frequently (Botsch
and Conrad, 2011; Heuer et al., 2009). Moreover, since CH4 is formed from the acetate-Me
during aceticlastic methanogenesis (Ferry, 1992), the δ13C value of this Me is the crucial one
for mass balance calculations. Obtaining such data is cumbersome and therefore relatively rare.
The δ13C values of acetate-Me and acetate-CO2H are often quite different (Conrad et al., 2011).
This intramolecular difference varies between environments and is likely dependent on
whether the acetate was formed mainly fermentatively or chemolithotrophically (Blair et al.,
1985; Gelwicks et al., 1989).
Fractionation factors have been studied in some detail for the final methanogenic steps
(Gelwicks et al., 1994; Goevert and Conrad, 2009; Penning et al., 2005), and to some extent

5
for chemolithotrophic acetogenesis (Blaser et al., 2013; Gelwicks et al., 1989). The factors are
quite substantial (ca. -10 to -90‰). However, data on carbon isotope fractionation during
primary or secondary fermentation are scarce (Blair et al., 1985; Botsch and Conrad, 2011;
Penning and Conrad, 2006). Nevertheless, the few data on fermentation reactions suggest that
fractionation factors are small (< 5‰).
The objectives of the present study were to evaluate the extent to which (i) the δ13C value
of acetate-Me and (ii) of propionate differ from those of total acetate and bulk OM in various
soils and sediments. We collected δ13C data for propionate, acetate and acetate-Me obtained
during anaerobic incubation of soil and sediment samples with different OM δ13C values. The
incubations had primarily been done for studying microbial CH4 production, in particular for
quantifying the methanogenic pathway. However, propionate and acetate had also been
analyzed routinely at the end of incubation. The δ13C values of propionate and acetate were
also analyzed in incubations in which aceticlastic methanogenesis was inhibited by CH3F
(Janssen and Frenzel, 1997), thereby avoiding effects from isotope fractionation during acetate
consumption.
2. Material and methods
Samples of freshwater sediments and soils were from the sites listed in Table 1, as
described in the quoted references. The sites included sediments and soils from different
places around the world, including tropical (Brazil, Thailand, Cameroon), temperate (Germany,
China, Uruguay), arid and semi-arid (USA, Israel, Australia), high altitude (Tibet) and boreal
(Finland) sites. Table 1 lists the δ13C analyses performed on the samples. The sediment and
soil samples had been used for determination of rates and pathways of CH4 production. For
this purpose, δ13C values of CH4, CO2 and acetate were measured, and sometimes also of
acetate-Me. The published δ13C data were collected from the references (Table 1). δ13C values
of propionate having not been published. Measurements from three sample series (Amazon

6
2010 and Lagunes, Brazil) and Cameroon have not been published. The Amazon 2010
sediment samples were from lakes with white water or clear water, as described for Amazon
2009. The lagoon samples were from coastal lagoons in the “Parque Nacional da Restinga de
Jurubatida e do Municipio de Macae” (Furtado et al., 2002). Three soils with a different
degree of moisture content were sampled from sites at Nsimi, a small experimental watershed
in the south of Cameroon (Braun et al., 2012).
The incubations and analysis were carried out as described in the references in Table 1 (e.g.
Conrad et al., 2007). Briefly, samples were placed in glass tubes (28 ml) or glass bottles (60-
120 ml) closed with butyl rubber stoppers and flushed with N2 (usually flushing the headspace
of the vessels or using several rounds of evacuation and gassing). In parallel, vessels were
treated with CH3F at initial concentrations of 0.5, 1, 2 or 3% (v/v) to inhibit aceticlastic
methanogenesis. In many experiments, CH3F was applied at more than one concentration. In
some samples (Lake Dagow sediment) CH4 production was completely inhibited by addition
of bromoethanesulfonate (5, 10, 15, 20 and 30 mM) or CHCl3 (50. 100, 150, 200 and 250 µM).
All treatments were carried out in usually 3, or sometimes 4, replicates. The samples were
incubated until CH4 was produced with a constant rate. The vessels were opened and the
contents centrifuged. The supernatant was filtered through a membrane filter and used for
analysis of acetate and propionate with high-performance liquid chromatography (HPLC) with
refraction index detection. The δ13C values of acetate and propionate was determined after
combustion (C) to CO2 using isotope ratio mass spectrometry (IRMS). The HPLC-C-IRMS
system had a detection limit of ca. 5 µM and a precision of ± 0.3‰. The δ13C values of the
methyl group of acetate were determined after off-line pyrolysis. Details have been given
before (Botsch and Conrad, 2011; Conrad et al., 2007). For δ13C analysis of OM, the samples
were acidified with HCl to remove carbonate, and dried. δ13C values were measured using an
elemental analyzer coupled to an IRMS instrument (Institute for Soil Science and Forest
Nutrition (IBW) at the University of Göttingen, Germany).

7
3. Results
Rice field soils from Italy, China and Thailand were incubated in the presence of either rice
straw or maize straw and exhibited δ13C values characteristic of C3 (-31.0 ± 0.1‰) and C4 (-
13.0 ± 0.1‰) pathways, respectively (Fig. 1). The median values for total acetate were almost
identical to those of the respective straw OM, the mean values being slightly higher (-29‰
and -11‰, respectively), while the upper and lower quartiles were of the order of ± 3-5‰.
In addition to the rice field soils, a wide variety of different soils and sediments (Table 1)
were incubated under methanogenic conditions. The δ13C values of acetate plotted against
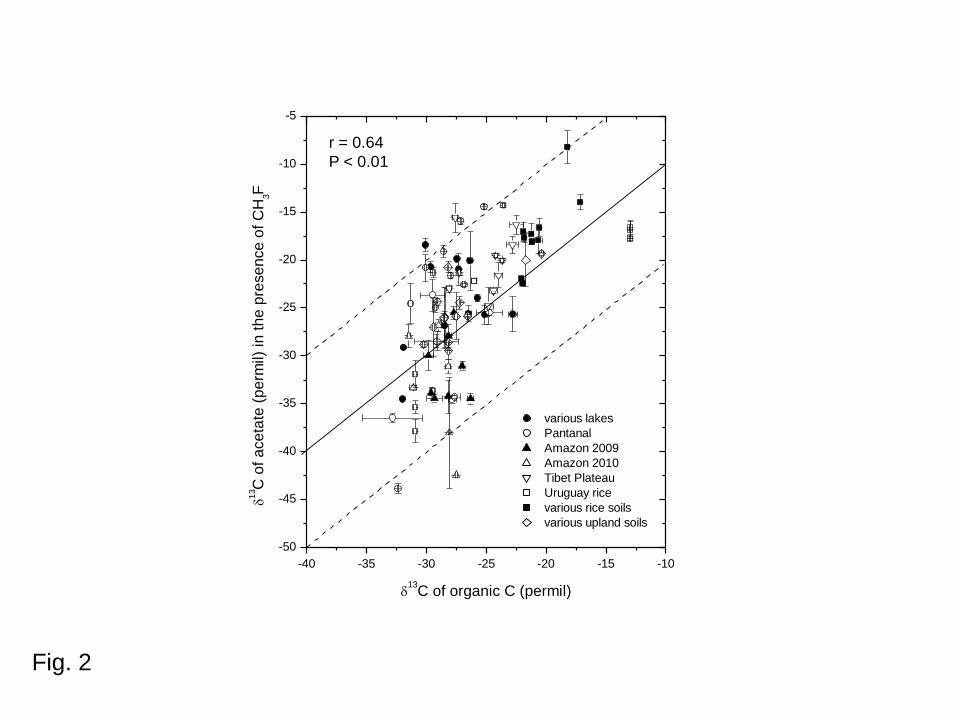
those of the OM in the respective soil and sediment samples are summarized in Fig. 2. The
data show a positive correlation between acetate and OM, the average difference (± SD) being
just -1.6 ± 5.3‰ (Fig. 2). However, the rather large variance should be noted.
The acetate was also analyzed with respect to the intramolecular difference of δ13C by
plotting the δ13C value of the Me against that of the total acetate (Fig. 3). The data included
incubations with and without CH3F, which, however, were not different. The data are
separately shown for soil samples (Fig. 3A) and lake sediments (Fig. 3B). The plots revealed
in both sets a linear relationship with a systematic offset showing that Me in soils and
sediments was on average (± SD) depleted in 13C by 6.4 ± 4.2‰ (soils) and by 11.0 ± 4.3‰
(sediments) relative to total acetate, respectively. The offset for all data together was 8.4 ±
4.8‰.
Propionate was analyzed in the incubations of rice field soils from Italy, China and
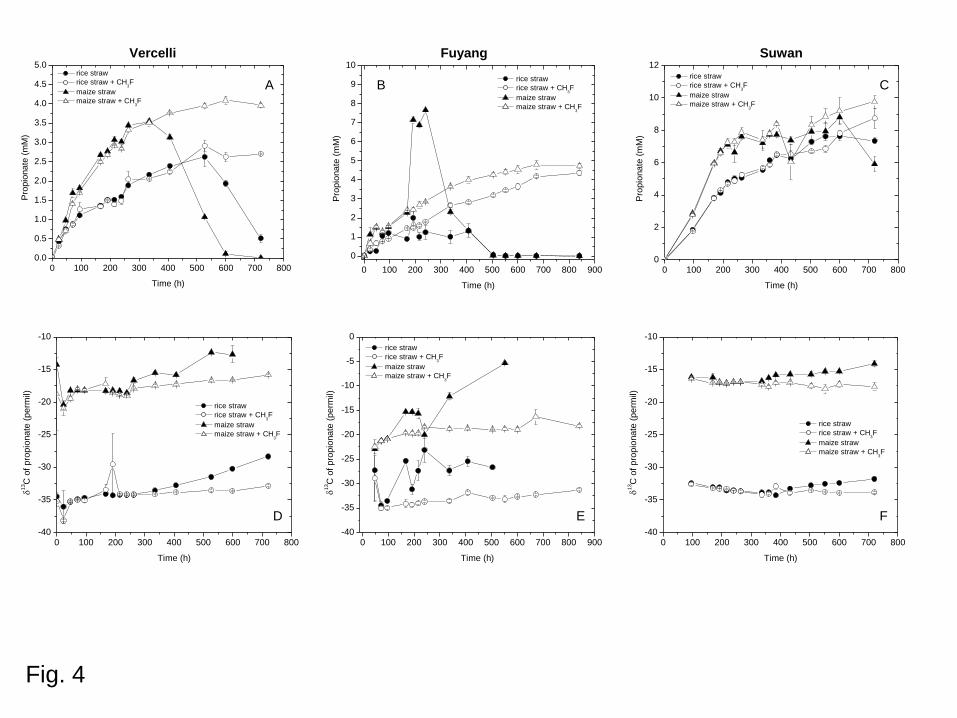
Thailand in the presence of either rice straw or maize straw (Fig. 4). It accumulated as an
intermediate reaching a maximum concentration of 2.5-9 mM, and then decreased, unless
aceticlastic methanogenesis was inhibited by CH3F. While the δ13C values of propionate
stayed relatively constant in the presence of CH3F, they slowly increased in its absence as
soon as the propionate concentration started to decrease. The δ13C values of propionate in the
non-inhibited samples increased to values that were at maximum of ca. 5-10‰ higher than for
the inhibited samples (Fig. 4). The values for propionate in the inhibited samples were similar

8
to those of the rice straw or maize straw OM. On average (median and mean nearly identical),
propionate was by 2.7 ± 0.2‰ and 5.1 ± 0.3‰ lighter than organic carbon (OC), respectively
(Fig. 1). Together with the data from all the other sediment and soil incubations, the δ13C
values of propionate increased linearly with those of total acetate (Fig. 5). We assume that the
values for propionate also increased linearly with those of OM, but many fewer data pairs
were available than for propionate and acetate. On average, propionate was depleted in 13C by
6 ± 5‰ relative to acetate.
4. Discussion
Analysis of the carbon isotopic composition in numerous samples of methanogenic
freshwater sediments and soils showed that fractionation was small during anaerobic
conversion of complex OM to acetate, propionate and CO2, while the further conversion of
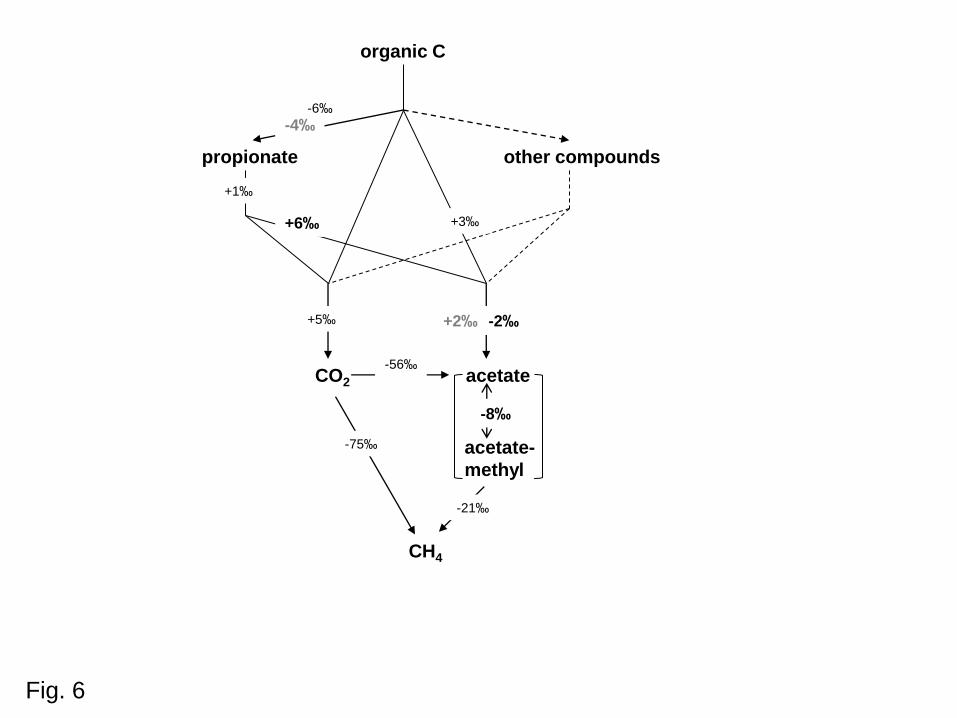
these compounds to CH4 exhibited large fractionation factors (Fig. 6). The samples were
incubated under methanogenic conditions, so that propionate and acetate could only be
degraded by processes coupled to CH4 production. However, aceticlastic methanogenesis was
inhibited by the addition of CH3F (Janssen and Frenzel, 1997). This inhibitor was found to be
quite specific and does normally not inhibit sulfate-reducing, acetogenic or fermenting
bacteria (Janssen and Frenzel, 1997). Hydrogenotrophic methanogens, however, may be
partially inhibited if CH3F concentrations are too high (Conrad and Klose, 1999). This is not
critical for interpretation of the present data as long as aceticlastic methanogenesis is
completely inhibited. Therefore, inhibition experiments were usually conducted at more than
one CH3F concentration to guarantee complete inhibition of aceticlastic methanogenesis. In
methanogenic samples, aceticlastic methanogenesis is normally the sole path for acetate
degradation, unless syntrophic acetate oxidizers exist that are able to couple acetate oxidation
to hydrogenotrophic methanogenesis (Zinder and Koch, 1984). For simplicity, we assumed
that such syntrophs did not play a major role and that acetate consumption was largely

9
impeded by CH3F. Indeed, we always observed net accumulation of acetate under these
conditions (references in Table 1). Therefore, the δ13C value of acetate was probably
unaffected by acetate consumption and should represent the δ13C value of the acetate after
production from precursors derived from OM. However, it is noteworthy that isotopic
fractionation during syntrophic acetate oxidation seems to be smaller than during aceticlastic
methanogenesis and so would have only a small effect on the δ13C value of the acetate pool
(Conrad and Klose, 2011).
The δ13C value of total acetate increased linearly with that of OM, exhibiting a difference
of ca. -6‰ to +4‰ (avg. ca. -2‰; Fig. 6). Hence, total acetate was similar to OC. Acetate
produced from the OC of C4 plants was heavier than that of C3 plants. Such similarity in δ13C
values would be expected if acetate is primarily produced by fermentative production, i.e.
anaerobic heterotrophic metabolism of OM (Blair et al., 1985). In fact, there is a paucity of
isotope fractionation data from pure cultures of fermentative microorganisms (Blair et al.,
1985; Botsch and Conrad, 2011; Penning and Conrad, 2006). The data indicate that acetate
was only slightly enriched in 13C (ca. 3‰; Fig. 6). However, it cannot be excluded that other
fermenting microorganisms would exhibit greater fractionation. Nevertheless, the similarity in
δ13C values of OM and acetate in our data suggest that organisms exhibiting a large
fractionation during fermentation (if they exist) may not be of importance for OM degradation
in anoxic freshwater sediments and soils. In addition, acetate can also be produced from
chemolitotrophic acetogenesis, i.e. the reduction of CO2 to acetate. This process exhibits rather
large fractionation factors, of the order of -56‰ (Blaser et al., 2013; Gelwicks et al., 1989).
The process is dominant in the anoxic hindgut of some termites (Breznak and Switzer, 1986)
and has been suggested to occur in some anoxic soils and sediments (Conrad et al., 1989;
Hoehler et al., 1999; Nozhevnikova et al., 2007). Indeed, we do not rule out the idea that
chemolithotrophic acetogenesis plays a certain role in some of the anoxic environments
analyzed, as suggested by the large variation in the δ13C difference between OC and acetate.

10
On the other hand, our data are such that acetate formation via heterotrophic fermentation of
OM seems to be the more common path in freshwater sediments and soils.
Acetate formation via mainly heterotrophic fermentation is consistent with the observed
intramolecular δ13C difference. Heterotrophic fermentation was reported to result in a rather
large intramolecular difference [δ13Cac-methyl - δ13Cac-carboxyl = 2 (δ13Cac-methyl - δ
13Cac)] of ca.
24‰ (Blair et al., 1985). Chemolithotrophic acetogenesis, on the other hand, exhibits only a
small intramolecular difference, i.e. < 1‰ (Gelwicks et al., 1989). The difference between
acetate-Me and total acetate in the soils (6‰) was on average a little smaller than in the
sediments (11‰). Together the difference was on average -8‰ (Fig. 6), equivalent to an
intramolecular difference of ca. 16‰. These data indicate that acetate was produced mainly by
fermentation, with a minor contribution from chemolithotrophic acetogenesis which, however,
may be more significant in soils than in sediments.
Propionate was produced as an intermediate when freshwater sediments and soils were
incubated under methanogenic conditions. Inhibition of aceticlastic methanogenesis also
resulted in inhibition of propionate degradation, at least during the time of observation, as
explicitly shown for the soils from Italy, China and Thailand (Fig. 4). This observation is
plausible, since propionate is most likely degraded by secondary fermentation, which easily
becomes endergonic when acetate or H2 accumulate (Chidthaisong and Conrad, 2000; Krylova
and Conrad, 1998). When propionate was not degraded, the δ13C values of propionate stayed
constant with time, whereas propionate became slightly heavier when it was consumed. Such
an increase in δ13C indicates isotope fractionation during propionate consumption. The
fractionation was in the range -10 to -5‰, which is relatively large compared with that in a
defined culture of propionate-utilizing Syntrophobacter fumaroxidans, where it was negligible
(Botsch and Conrad, 2011). The reason for this difference is unclear and needs further study.
One reason could be the pathway of propionate utilization. Syntrophobacter utilizes
propionate via the randomizing succinate or methyl malonyl-CoA pathway (CH3CH2COO- +
3H2O � CH3COO- + HCO3- + 3H2 + H+) (Houwen et al., 1990). The same pathway was found

11
to be operative in Italian rice field soil (Krylova et al., 1997). However, in other environments
propionate may be degraded via other pathways, e.g. the non-randomizing pathway
(2CH3CH2COO- + 2H2O � 3CH3COO- + 2H2 + H+) found in Smithella spp., which proceeds
via intermediate butyrate formation (DeBok et al., 2001). Both degradation pathways are
rather complex, so that isotope fractionation during propionate depletion would barely enable
prediction of the isotopic signature of the acetate, which is only one of the possible products of
propionate degradation.
When propionate was not degraded, it was slightly depleted in 13C (ca. 4‰; Fig. 6) relative
to the straw serving as the major substrate. This relatively small fractionation was consistent
with microbial culture studies fermenting pectin, xylan or starch, which showed similar
isotope fractionation (ca. -6‰; Fig. 6) during propionate formation by Opitutus terrae (Botsch
and Conrad, 2011). Overall, fractionation during propionate formation from OM was larger
than during acetate formation, resulting in acetate that was on average ca. 6‰ heavier than
propionate (Figs. 5, 6). However, the range of difference between propionate and acetate was
rather large, indicating that the processes involved in turnover of these compounds are not
well understood.
This notion is further illustrated by the following consideration. In Italian rice field soil,
propionate turnover can account for all of the H2 driving hydrogenotrophic methanogenesis
(Krylova et al., 1997). Hydrogenotrophic methanogenesis accounts for ca. 30% of total CH4
production, the remaining 70% is due to aceticlastic methanogenesis. On a molar basis
propionate turnover produces one acetate and three H2 molecules (succinate pathway).
Therefore, if 30% of the total CH4 is from propionate-derived H2, then 40% of the CH4 must
be from propionate-derived acetate. Thus almost 60% of the total acetate production would be
caused by propionate oxidation. Such a large contribution of propionate turnover to acetate
turnover cannot be generally true, since otherwise the difference between propionate and
acetate should be < 6‰.

12
In conclusion, our study has revealed gaps of knowledge in the detail, but showed a
relatively uniform picture for isotope fractionation during production of acetate and propionate
from OM in soils and sediments sampled from around the world. In general, fractionation
during fermentation was relatively small, of the order of only a few permil (Fig. 6). Even
when taking the large variance into account, the δ13C values of acetate differed from those of
OC on average by only -2‰ (Fig. 6). Likewise, the values of the CO2 produced are not much
different from those of the OM (Conrad et al., 2012b), similar to the situation found for
aerobic CO2 production from soil OM (Werth and Kuzyakov, 2010). The fractionation factors
for the further conversion of acetate and CO2 to CH4 are much larger (Fig. 6). Hence, if δ13C
values for acetate are unknown, those of OC could be used as a proxy for mass balance
calculations. Angel et al. ( 2012) provide an example for determining the fraction of CH4
produced by hydrogenotrophic and aceticlastic methanogenesis. In such a case, the δ13C value
of acetate-Me rather that of total acetate is required. Analysis of our data showed that acetate-
Me was depleted in 13C relative to total acetate, on average by 8 ± 5‰. Again, one might use
this value as a first approximation to correct the δ13C value of total acetate, if a value for
acetate-Me were not available (Conrad et al., 2012a). If only the δ13C value of OM is available,
one might estimate the value for acetate-Me by subtracting ca. 10‰ (Angel et al., 2012). Of
course, all these corrections bear more uncertainty than direct analysis, but may nevertheless
be useful for an initial estimate.
Acknowledgements
The study is part of the ICON project financed by the German Research Foundation. We
thank the Brazilian agencies CNPq, CAPES and FAPERJ for the financial support used to
sample sediments in Brazil. The sampling in Tibet was supported by the National Natural
Science Foundation of China, the Chinese Academy of Science and the German Research
Foundation. The soils from Cameroon were sampled in the Nsimi small experimental

13
watershed (SOERE BVET: Service Observation Recherche en Environnement – Basins
Versants Expérimentaux Tropicaux funded by IRD and INSU/CNRS; http://bvet.omp.obs-
mip.fr) in the framework of the project LIMON funded by EC2CO (INSU/CNRS). P.E.G. is
supported by the Agence Nationale de la Recherche (ANR) project MICADO (ANR-11JSV7-
003-01). We thank two anonymous reviewers for helpful comments.
References Angel, R., Claus, P., Conrad, R., 2012. Methanogenic archaea are globally ubiquitous in
aerated soils and become active under wet anoxic conditions. ISME Journal 6, 847-862.
Blair, N., Leu, A., Munoz, E., Olsen, J., Kwong, E., DesMarais, D., 1985. Carbon isotopic
fractionation in heterotrophic microbial metabolism. Applied and Environmental
Microbiology 50, 996-1001.
Blaser, M. B., Dreisbach, L. K., Conrad, R., 2013. Carbon isotope fractionation of 11
acetogenic strains grown on H2 and CO2. Applied and Environmental Microbiology 79,
1787-1794.
Boschker, H. T. S., DeGraaf, W., Koster, M., Meyer-Reil, L. A., Cappenberg, T. E., 2001.
Bacterial populations and processes involved in acetate and propionate consumption in
anoxic brackish sediment. FEMS Microbiology Ecology 35, 97-103.
Botsch, K. C. and Conrad, R., 2011. Fractionation of stable carbon isotopes during anaerobic
production and degradation of propionate in defined microbial cultures. Organic
Geochemistry 42, 289-295.
Braun, J. J., Marechal, J. C., Riotte, J., Boeglin, J. L., Bedimo Bedimo, J. P., Ndam
Ngoupayou, J. R., Nyeck, B., Robain, H., Sekhar, M., Audry, S., Viers, J., 2012. Elemental
weathering fluxes and saprolite production rate in a Central African lateritic terrain (Nsimi,
South Cameroon). Geochimica et Cosmochimica Acta 99, 243-270.
Breznak, J. A. and Switzer, J. M., 1986. Acetate synthesis from H2 plus CO2 by termite gut
microbes. Applied and Environmental Microbiology 52, 623-630.

14
Chidthaisong, A. and Conrad, R., 2000. Specificity of chloroform, 2-bromoethanesulfonate
and fluoroacetate to inhibit methanogenesis and other anaerobic processes in anoxic rice
field soil. Soil Biology and Biochemistry 32, 977-988.
Conrad, R., 1999. Contribution of hydrogen to methane production and control of hydrogen
concentrations in methanogenic soils and sediments. FEMS Microbiology Ecology 28, 193-
202.
Conrad, R., 2005. Quantification of methanogenic pathways using stable carbon isotopic
signatures: a review and a proposal. Organic Geochemistry 36, 739-752.
Conrad, R., Bak, F., Seitz, H. J., Thebrath, B., Mayer, H. P., Schütz, H., 1989. Hydrogen
turnover by psychrotrophic homoacetogenic and mesophilic methanogenic bacteria in
anoxic paddy soil and lake sediment. FEMS Microbiology Ecology 62, 285-294.
Conrad, R., Chan, O. C., Claus, P., Casper, P., 2007. Characterization of methanogenic
Archaea and stable isotope fractionation during methane production in the profundal
sediment of an oligotrophic lake (Lake Stechlin, Germany). Limnology and Oceanography
52, 1393-1406.
Conrad, R., Claus, P., Casper, P., 2009a. Characterization of stable isotope fractionation
during methane production in the sediment of a eutrophic lake, Lake Dagow, Germany.
Limnology and Oceanography 54, 457-471.
Conrad, R., Claus, P., Casper, P., 2010a. Stable isotope fractionation during the methanogenic
degradation of organic matter in the sediment of an acidic bog lake, Lake Grosse
Fuchskuhle. Limnology and Oceanography 55, 1932-1942.
Conrad, R. and Frenzel, P., 2002, Flooded soils. In: Bitton, G. (Ed.), Encyclopedia of
Environmental Microbiology, John Wiley & Sons, Inc., New York, pp. 1316-1333.
Conrad, R., Ji, Y., Noll, M., Klose, M., Claus, P., Enrich-Prast, A., 2013. Response of the
methanogenic microbial communityies in Amazonian oxbow lake sediments to desiccation
stress. Environmental Microbiology, in press, doi:10.1111/1462-2920.12267.
Conrad, R. and Klose, M., 1999. How specific is the inhibition by methyl fluoride of
acetoclastic methanogenesis in anoxic rice field soil? FEMS Microbiology Ecology 30, 47-
56.

15
Conrad, R. and Klose, M., 2011. Stable carbon isotope discrimination in rice field soil during
acetate turnover by syntrophic acetate oxidation or acetoclastic methanogenesis.
Geochimica et Cosmochimica Acta 75, 1531-1539.
Conrad, R., Klose, M., Claus, P., Dan, J. G., 2009b. Activity and composition of the
methanogenic archaeal community in soil vegetated with wild versus cultivated rice. Soil
Biology and Biochemistry 41, 1390-1395.
Conrad, R., Klose, M., Claus, P., Enrich-Prast, A., 2010b. Methanogenic pathway, 13C isotope
fractionation, and archaeal community composition in the sediment of two clearwater lakes
of Amazonia. Limnology and Oceanography 55, 689-702.
Conrad, R., Klose, M., Lu, Y., Chidthaisong, A., 2012a. Methanogenic pathway and archaeal
communities in three different anoxic soils amended with rice straw and maize straw.
Frontiers in Microbiology 3, 4, doi: 10.3389/fmicb.2012.00004.
Conrad, R., Klose, M., Noll, M., 2009c. Functional and structural response of the
methanogenic microbial community in rice field soil to temperature change. Environmental
Microbiology 11, 1844-1853.
Conrad, R., Klose, M., Yuan, Q., Lu, Y., Chidthaisong, A., 2012b. Stable carbon isotope
fractionation, carbon flux partitioning and priming effects in anoxic soils during
methanogenic degradation of straw and soil organic matter. Soil Biology and Biochemistry
49, 193-199.
Conrad, R., Noll, M., Claus, P., Klose, M., Bastos, W. R., Enrich-Prast, A., 2011. Stable
carbon isotope discrimination and microbiology of methane formation in tropical anoxic
lake sediments. Biogeosciences 8, 795-814.
DeBok, F. A. M., Stams, A. J. M., Dijkema, C., Boone, D. R., 2001. Pathway of propionate
oxidation by a syntrophic culture of Smithella propionica and Methanospirillum hungatei.
Applied and Environmental Microbiology 67, 1800-1804.
Dolfing, J., 1988, Acetogenesis. In: Zehnder, A. J. B. (Ed.), Biology of Anaerobic
Microorganisms, Wiley, New York, pp. 417-468.
Drake, H. L. and Küsel, K., 2003, How the diverse physiologic potentials of acetogens
determine their in situ realities. In: Ljungdahl, L. G., Adams, M. W., Barton, L. L., Ferry, J.
G., Johnson, M. K. (Eds.), Biochemistry and Physiology of Anaerobic Bacteria, Springer,
New York, pp. 171-190.

16
Feisthauer, S., Siegert, M., Seidel, M., Richnow, H. H., Zengler, K., Gründger, F., Krüger, M..
2010. Isotopic fingerprinting of methane and CO2 formation from aliphatic and aromatic
hydrocarbons. Organic Geochemistry 41, 482-490.Fernandez Scavino, A., Ji, Y., Pump, J.,
Klose, M., Claus, P., Conrad, R., 2013. Structure and function of the methanogenic
microbial communities in Uruguayan soils shifted between pasture and irrigated rice fields.
Environmental Microbiology 15, 2588-2602.
Ferry, J. G., 1992. Methane from acetate [Minireview]. Journal of Bacteriology 174, 5489-
5495.
Furtado, A. L. S., Casper, P., Esteves, F. A., 2002. Methanogenesis in an impacted and two
dystrophic coastal lagoons (Macae, Brazil). Brazilian Archives of Biology and Technology
45, 195-202.
Galand, P. E., Yrjälä, K., Conrad, R., 2010. Stable carbon isotope fractionation during
methanogenesis in three boreal peatland ecosystems. Biogeosciences 7, 3893-3900.
Gelwicks, J. T., Risatti, J. B., Hayes, J. M., 1989. Carbon isotope effects associated with
autotrophic acetogenesis. Organic Geochemistry 14, 441-446.
Gelwicks, J. T., Risatti, J. B., Hayes, J. M., 1994. Carbon isotope effects associated with
aceticlastic methanogenesis. Applied and Environmental Microbiology 60, 467-472.
Glissmann, K. and Conrad, R., 2000. Fermentation pattern of methanogenic degradation of
rice straw in anoxic paddy soil. FEMS Microbiology Ecology 31, 117-126.
Goevert, D. and Conrad, R., 2009. Effect of substrate concentration on carbon isotope
fractionation during acetoclastic methanogenesis by Methanosarcina barkeri and M.
acetivorans and in rice field soil. Applied and Environmental Microbiology 75, 2605-2612.
Heuer, V. B., Krüger, M., Elvert, M., Hinrichs, K. U., 2010. Experimental studies on the stable
carbon isotope biogeochemistry of acetate in lake sediments. Organic Geochemistry 41, 22-
30.
Heuer, V. B., Pohlman, J. W., Torres, M. E., Elvert, M., Hinrichs, K. U., 2009. The stable
carbon isotope biogeochemistry of acetate and other dissolved carbon species in deep
subseafloor sediments at the northern Cascadia Margin. Geochimica et Cosmochimica Acta
73, 3323-3336.
Hoehler, T. M., Albert, D. B., Alperin, M. J., Martens, C. S., 1999. Acetogenesis from CO2 in
an anoxic marine sediment. Limnology and Oceanography 44, 662-667.

17
Houwen, F. P., Plokker, J., Stams, A. J. M., Zehnder, A. J. B., 1990. Enzymatic evidence for
involvement of the methylmalonyl-CoA pathway in propionate oxidation by
Syntrophobacter wolinii. Archives of Microbiology 155, 52-55.
Janssen, P. H. and Frenzel, P., 1997. Inhibition of methanogenesis by methyl fluoride - studies
of pure and defined mixed cultures of anaerobic bacteria and archaea. Applied and
Environmental Microbiology 63, 4552-4557.
Krylova, N. I. and Conrad, R., 1998. Thermodynamics of propionate degradation in
methanogenic paddy soil. FEMS Microbiology Ecology 26, 281-288.
Krylova, N. I., Janssen, P. H., Conrad, R., 1997. Turnover of propionate in methanogenic
paddy soil. FEMS Microbiology Ecology 23, 107-117.
Liu, Y., Yao, T., Gleixner, G., Claus, P., Conrad, R., 2013. Methanogenic pathways, 13C
isotope fractionation, and archaeal community composition in lake sediments and wetland
soils on the Tibetan Plateau. Journal of Geophysical Research - Biogeosciences 118, in
press, doi:10.1002/jgrg.20055.
McInerney, M. J. and Bryant, M. P., 1981, Basic principles of bioconversions in anaerobic
digestion and methanogenesis. In: Sofer, S. S., Zaborsky, O. R. (Eds.), Biomass Conversion
Processes for Energy and Fuels, Plenum, New York, pp. 277-296.
Megonigal, J. P., Hines, M. E. and Visscher, P. T., 2003, Anaerobic metabolism: linkages to
trace gases and aerobic processes. In: Schlesinger, W. H., Holland, H. D., Turekian, K. K.
(Eds.), Treatise on Geochemistry, vol. 8: Biogeochemistry, Elsevier-Pergamon, Oxford,
UK, pp. 317-424.
Morris, B. E. L., Herbst, F. A., Bastida, F., Seifert, J., VonBergen, M., Richnow, H. H.,
Suflita, J. M., 2012. Microbial interactions during residual oil and n-fatty acid metabolism
by a methanogenic consortium. Environmental Microbiololgy Reports 4, 297-306.
Nozhevnikova, A. N., Nekrasova, V., Ammann, A., Zehnder, A. J. B., Wehrli, B., Holliger, C.,
2007. Influence of temperature and high acetate concentrations on methanogenensis in lake
sediment slurries. FEMS Microbiology Ecology 62, 336-344.
Penning, H. and Conrad, R., 2006. Carbon isotope effects associated with mixed-acid
fermentation of saccharides by Clostridium papyrosolvens. Geochimica et Cosmochimica
Acta 70, 2283-2297.

18
Penning, H. and Conrad, R., 2007. Quantification of carbon flow from stable isotope
fractionation in rice field soils with different organic matter content. Organic Geochemistry
38, 2058-2069.
Penning, H., Plugge, C. M., Galand, P. E., Conrad, R., 2005. Variation of carbon isotope
fractionation in hydrogenotrophic methanogenic microbial cultures and environmental
samples at different energy status. Global Change Biology 11, 2103-2113.
Werth, M. and Kuzyakov, Y., 2010. 13C fractionation at the root-microorganisms-soil
interface: A review and outlook for partitioning studies. Soil Biology and Biochemistry 42,
1372-1384.
Whiticar, M. J., Faber, E., Schoell, M., 1986. Biogenic methane formation in marine and
freshwater environments: CO2 reduction vs. acetate fermentation - isotopic evidence.
Geochimica et Cosmochimica Acta 50, 693-709.
Zinder, S. H., 1993, Physiological ecology of methanogens. In: Ferry, J. G. (Ed.),
Methanogenesis. Ecology, Physiology, Biochemistry and Genetics, Chapman & Hall, New
York, pp. 128-206.
Zinder, S. H. and Koch, M., 1984. Non-aceticlastic methanogenesis from acetate: acetate
oxidation by a thermophilic syntrophic coculture. Archives of Microbiology 138, 263-272.

19
Table 1: Origin of sediment and soil samples used for determination of δ13C (+, analyzed in all samples; +/-, analyzed in most samples; -, not analyzed; np, unpublished results
a 1, Conrad et al., 2011; 2, Conrad et al., 2013; 3, Galand et al., 2010; 4, Conrad et al., 2010b; 5, Liu et al., 2013; 6, Conrad et al., 2007; 7, Conrad et al., 2009a; 8, Conrad et al., 2010a; 9, Penning and Conrad, 2007; 10, Conrad et al., 2012a; 11, Conrad et al., 2009b; 12, Fernandez Scavino et al., 2013; 13, Angel et al., 2012.
Samples description δ13Corg δ
13Cprop δ13Cac δ
13Cac-methyl Referencea
Lake sediments Pantanal, Brazil Different lakes (16) + + + + 1 Amazon 2009 White water (3), black water (3), clear water lakes (3) + +/- + + 2 Amazon 2010 White water (4), clear water lakes (3) + + + + np Mires, Finland Fens (2), bog (1) + - + - 3 Amazon 2006 Clear water lakes (2; Batata, Mussura) + - + + 4 Tibetan plateau Lake sediments (7), wetlands (2) + - + - 5 Lagoons, Brazil Coastal lagoons (Cabiunas, Imboassica) at Macae, RJ + - + - np Lakes, Germany Lakes Stechlin, Dagow, Grosse Fuchskuhle + +/- + + 6, 7, 8 Soils Vercelli, Italy Rice soil, addition of rice or maize straw, no addition + + + + 9, 10 Fuyang, China Rice soil, addition of rice or maize straw + + + + 10 Suwan, Thailand Rice soil, addition of rice or maize straw + + + - 10 Hainan, China Cultivated rice, wild rice + - + + 11 Uruguay 2009 Drained rice field + + + + 12 Uruguay 2010 Fields within a pasture rice crop rotation (8), pasture (1) + + + - 12 Upland soils Germany, Israel, Cameroon, Australia, Utah + + + +/- 13, np

20
Figure legends
Fig. 1. Box plots of δ13C of total acetate (ac) and propionate (prop) measured at the end
of anaerobic incubations (with 1% CH3F) of rice field soils from Italy, China and
Thailand amended with either rice straw (RS) or maize straw (MS). Horizontal lines
indicate the δ13C of OM in MS (upper line) and RS (lower line); n = 20.
Fig. 2. Values of δ13C of total acetate vs. those of OC at the end of anaerobic incubation
of soil or sediment in the presence of CH3F. Solid and dashed lines indicate no
fractionation and ± 10‰ fractionation, respectively; mean ± SE, n = 3. Also see
supplemental data table.
Fig. 3. Values of δ13C of acetate-Me vs. those of total acetate at the end of anaerobic
incubation of (A) various soils and (B) various freshwater sediments. Incubation was with
and without CH3F. Solid line indicates no fractionation, dashed lines -10‰ and -20‰
fractionation between acetate-Me and total acetate; mean ± SE, n = 3. Also see
supplemental data table.
Fig. 4. Transient accumulation of propionate (upper panel) and δ13C values (lower panel)
of soil from (A,D) Vercelli (Italy), (B,E) Fuyang (China) and (C,F) Suwan (Thailand),
amended with either rice straw or maize straw and incubated under anoxic conditions in
the presence or absence of 1% CH3F.
Fig. 5. δ13C values of propionate vs. those of acetate at the end of anaerobic incubation of
soil or sediment with CH3F. Solid line indicates no fractionation, dashed lines -10‰ and -
20‰ fractionation between acetate-Me and total acetate; mean ± SE, n = 3. Also see
supplemental data table.

21
Fig. 6. Scheme of carbon flow and stable carbon-isotopic enrichment factors (ε) in
methanogenic environments. Bold numbers are from this study, bold black numbers refer
to data in Figs 2, 3 and 5; bold grey numbers refer to data in Fig. 1. Small black numbers
refer to representative literature data, i.e. (from top to bottom): Botsch and Conrad (2011),
Penning and Conrad (2006), Conrad et al. (2012b), Blaser et al. (2013), Conrad et al.
(2011), Goevert and Conrad (2009).

RS.pr.i RS.ac.t.i MS.pr.i MS.ac.t.i
-40
-35
-30
-25
-20
-15
-10
-5
0
5
1
3C
of
pro
pio
na
te o
r a
ce
tate
(p
erm
il)
Straw type, compound
RS, prop RS, ac MS, prop MS, ac
Fig. 1
Figure(s)
-40 -35 -30 -25 -20 -15 -10
-50
-45
-40
-35
-30
-25
-20
-15
-10
-5
13C
of
ace
tate
(p
erm
il)
in t
he
pre
se
nce
of
CH
3F
13
C of organic C (permil)
various lakes
Pantanal
Amazon 2009
Amazon 2010
Tibet Plateau
Uruguay rice
various rice soils
various upland soils
r = 0.64
P < 0.01
Fig. 2

A B
Fig. 3
-55 -50 -45 -40 -35 -30 -25 -20 -15 -10
-55
-50
-45
-40
-35
-30
-25
-20
-15
-10
13C
of
ace
tate
-me
thyl (p
erm
il)
13
C of total acetate (permil)
Pantanal Lakes
with CH3F
Amazon Lakes 2009
with CH3F
Amazon Lakes 2010
with CH3F
other lakes
with CH3F
with BES, CHCl3
-55 -50 -45 -40 -35 -30 -25 -20 -15 -10 -5 0 5 10 15 20 25 30
-55
-50
-45
-40
-35
-30
-25
-20
-15
-10
-5
0
5
10
15
20
25
30
13C
of
ace
tate
-me
thyl (p
erm
il)
13
C of total acetate (permil)
rice soils, rice straw
rice soils, rice straw + CH3F
rice soils, maize straw
rice soils, maize straw + CH3F
other rice soils
other rice soils + CH3F
upland soils + CH3F
0 100 200 300 400 500 600 700 8000.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0 rice straw
rice straw + CH3F
maize straw
maize straw + CH3F
Pro
pio
nate
(m
M)
Time (h)
0 100 200 300 400 500 600 700 800-40
-35
-30
-25
-20
-15
-10
rice straw
rice straw + CH3F
maize straw
maize straw + CH3F
1
3C
of pro
pio
nate
(perm
il)
Time (h)
0 100 200 300 400 500 600 700 800 900
0
1
2
3
4
5
6
7
8
9
10
rice straw
rice straw + CH3F
maize straw
maize straw + CH3F
Pro
pio
na
te (
mM
)
Time (h)
0 100 200 300 400 500 600 700 800 900-40
-35
-30
-25
-20
-15
-10
-5
0
rice straw
rice straw + CH3F
maize straw
maize straw + CH3F
13C
of pro
pio
nate
(p
erm
il)
Time (h)
0 100 200 300 400 500 600 700 8000
2
4
6
8
10
12
rice straw
rice straw + CH3F
maize straw
maize straw + CH3F
Pro
pio
na
te (
mM
)
Time (h)
0 100 200 300 400 500 600 700 800-40
-35
-30
-25
-20
-15
-10
rice straw
rice straw + CH3F
maize straw
maize straw + CH3F
1
3C
of pro
pio
nate
(perm
il)
Time (h)
Vercelli Fuyang Suwan
A B C
D E F
Fig. 4

-45 -40 -35 -30 -25 -20 -15 -10 -5 0 5
-45
-40
-35
-30
-25
-20
-15
-10
13C
of
pro
pio
na
te (
pe
rmil)
13
C of total acetate (permil)
Pantanal
Amazon 2009
Amazon 2010
German Lakes
rice soils, rice straw
rice soils, maize straw
Uruguay rice soil
upland soils
Fig. 5
r = 0.81
P < 0.01
organic C
propionate other compounds
-2‰
-4‰
+6‰
+2‰
-6‰
+1‰
acetate CO2
CH4
acetate-
methyl
-8‰
-56‰
-75‰
-21‰
+3‰
+5‰
Fig. 6

22
Highlights Methanogenic degradation of organic matter in anoxic environments examined. δ
13C of organic C, propionate, acetate and acetate-methyl relate to each other. Smaller fractionation during production of propionate and acetate than of CH4. We suggest δ13C value of in organic C can be used as proxy for that of acetate.