soybean agglutinin binding to the olfactory systems of the rat and mouse
TRANSCRIPT

Neuroscience Let ters, 69 (1986) 131-136 131
Elsevier Scientific Publishers Ireland Ltd.
NSL 04115
S O Y B E A N AGGLUTININ BINDING TO THE O L F A C T O R Y S Y S T E M S OF THE RAT A N D M O U S E
BRIAN KEY* and PIERO P. G1ORGI
Neuroemho,ology Laboratory, School o f Anatomy, University o/Queensland, St. Lucia, 4067 Brisbane, Qhl.
( A us t ralia )
(Received April 8th, 1986; Revised version received May 21st, 1986: Accepted May 26th, 1986)
Key words. olfitctory system - lectin b i nd i ng - neuronal subclass rodent
The binding of the lectin soybean agglutinin (SBA) to the olfactory system of both the rat and mouse was investigated histochemically. SBA bound to fibers in the accessory olfactory nerve and to glomeruli
in the accessory olfactory bulb. In addition, SBA binding sites were present in some, but not all, glomeruli
in the ventrolatcral and ventromedial portions of the main olfactory bulb of only the rat. Under standard experimental conditions, SBA did not bind to neurons in other regions of the olfactory system nor to any
other neurons in the brain. This selective binding of SBA to only some glomeruli in the olfactory bulb provides additional support for the presence of, at least, two subclasses of olfactory receptor cells in the nasal cavity. Whether these neuronal subclasses are the same as those previously characterized by mono-
clonal antibodies in rabbit remains to be determined.
The mammalian olfactory system consists of 3 morphologically distinct olfactory mucosae: the olfactory mucosa proper (OP), the vomeronasal organ (VNO) and the septal organ of Masera [2, 8]. Recently two subclasses of receptor cells have been identified in the OP and VNO of the rabbit on the basis of the molecular heteroge- neity observed using monoclonal antibodies [5, 9, 16]. We also have described a sub- class of olfactory neurons that innervates only the ventral portion of the main olfac- tory bulb (MOB) in Xenopus [18, 19]. These neurons were identified by cell surface glycoconjugates which were selectively labelled by the lectin soybean agglutinin (SBA). In addition, this lectin also labelled the VNO neurons. The ventral/dorsal dif- ference in the MOB was, however, particularly interesting considering a similar dis- tribution of subtypes is observed in the rabbit using monoclonal antibody technol-
ogy. The aim of the present study was to determine whether olfactory neurons, or sub-
classes of these cells carry specific SBA binding sites in the rat and mouse. We wish to address the important question of whether SBA binding could identify homolo- gous neurons in the olfactory system of vertebrates.
Four adult rats of the Wistar strain and 4 adult mice of the A 6 strain were anesthe-
*Author lk~r Correspondence.
0304 3040 ~65 03.50 © 1986 Elsevier Scientilic Publishers Ireland Ltd.

132
tized with i.p. injections of Nembutal and sacrificed by cardiac perfusion with 4% para- formaldehyde in 0.2 M potassium phosphate buffer, pH 7.2. The brain was removed and serial 40-gm coronal frozen section were cut and allowed to dry on gelatin- coated microscope slides. SBA was purchased from Sigma as a conjugate to horse- radish peroxidase (SBA-HRP). Sections were incubated in S B A - H R P (13 gg/ml in 0.1 M potassium phosphate buffer, pH 7.2, containing 1 mM CaCI2, MnC12, MgCI2) for 40 min. This incubation was lbllowed by several washes in the buffered salts so-
lution to remove unbound lectin before sections were reacted for the histochemical demonstrat ion of HRP [12]. Control sections incubated in the presence of 0.2 M N- acetyI-D-galactosamine and S B A - H R P or just HRP alone revealed that SBA--HRP labelling in the tissue was specific for this sugar.
In the mouse, S B A - H R P labelling (dark reaction product) was present in only the accessory olfactory nerve (AON) which courses along the medial surface of the MOB, and was absent from the olfactory nerve fiber layer and glomeruli of the MOB (Fig. la). In the dorso caudal MOB, SBA H R P labelling was present in the accessory olfactory bulb (AOB), the terminal field tbr accessory olfactory axons from the VNO (Fig. lb). Serial sections revealed that this labelling was present in all glomeruli m the AOB. All other neurons in the brain, at least under the present histochemical conditions, failed to bind this lectin. The specific binding of S B A - H R P to olfactory neurons in mice was observed in all animals analysed.
In the rat, in addition to the dense labelling of all glomeruli in the AOB (Fig. IC). SBA HRP lightly labelled a limited number of olfactory glomeruli in the MOB (Fig. I D). The differences in the density of reaction product between glomeruli in the AOB and MOB suggests that there are far less SBA binding molecules or sites on olfactory axons than on accessory olfactory axons. The labelled olfactory glomeruli in the MOB were located in the vemromedial and ventrolateral regions of the rostral bulb. In the caudal MOB, however, only the more ventral glomeruli were labelled. It is important to note that not all glomeruli in these regions were labelled and that unla- belled glomeruli often separated adjacently labelled glomeruli. As for mice, results observed in rats were consistent in all animals analysed.
The SBA HRP labelling in the olfactory systems of Xenopus [18, 19] as well as in mammals (rat and mouse) suggests that the specific binding molecules have impor- tant cellular functions in these neurons. This is particularly so fbr accessory olfactory neurons, since all glomeruli in the AOB of the 3 species, are labelled by SBA-HRP. In the MOB, however, there appears to be some inter-species differences. We have previously shown that all glomeruli in the ventral half of the MOB in Xenopus bind SBA [18, 19] whereas in the rat only some glomeruli in these regions were labelled. Furthermore, in the mouse no MOB glomeruli were labelled. Although the differ- ences in the labelling of the MOB between rat and mouse are most likely species- related differences, we have not excluded the possibility that higher concentrations of S B A - H R P may detect binding sites in some glomeruli of the mouse. Preliminary investigations in Xemq~us and rat [I 8] revealed that the present concentration reduced non-speeilic background labelling and gave optimal results. We are, however, cur- rently investigating the effect of different concentrations of S B A - H R P on labelling patterns in the mouse brain.
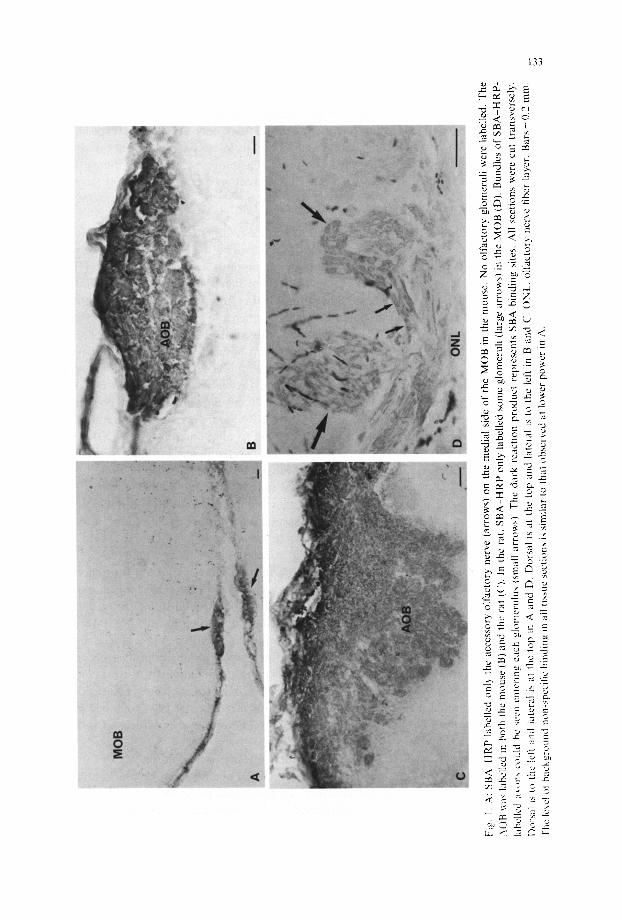
MO
B
B
Fie.
1.
A:
SB
A
HR
P l
abel
led
only
the
acc
esso
ry o
lfac
tory
ner
ve (
arro
ws)
on
the
med
ial
side
of
the
MO
B i
n th
e m
ou
se.
No
olf
acto
ry g
lom
erul
i w
ere
labe
lled
. T
he
..\(~
)B ~
as l
abel
led
in b
oth
the
mo
use
(B
) an
d th
e ra
t (C
). I
n th
e ra
t, S
BA
H
RP
onl
y la
bell
ed s
om
e gl
omer
uli
(lar
ge a
rro
ws)
in
the
MO
B (
D).
Bun
dles
ofS
BA
H
RP
-
labe
lled
axon
s co
uld
bc s
een
ente
ring
eac
h g
lom
eru
lus
(sm
all
arro
ws)
. T
he d
ark
reac
tion
pro
du
ct r
epre
sen
ts S
BA
bin
ding
site
s.
All
sect
ions
wer
e cu
t tr
ansv
erse
ly.
DoF
sal
i~ t
o th
c le
ft a
nd l
ater
al i
s at
the
to
p i
n A
and
D.
Do
rsal
is
at t
he t
op
and
lat
eral
is
to t
he l
eft
in B
and
C.
ON
L.
olfa
ctor
y ne
rve
fibe
r la
yer.
Ba
rs-0
.2 m
m.
Ihe
lexe
l o
f b
ack
gro
un
d n
on-~
pcci
fic
bind
ing
in a
ll ti
ssue
sec
tion
s is
sim
ilar
to
that
obs
erve
d at
low
er p
ow
er i
n A
. ,..
.a

134
Despite having previously shown SBA to bind to the cell surface of a subclass of receptor cells in Xenopus it is not possible from the present results to identify the cel- lular location of binding sites in the rat and mouse. Also, because SBA binds specifi- cally to the sugar N-acetyl-D-galactosamine [7] very little can be predicted about thc molecular nature of the binding glycoconjugate. Therefore, whether SBA binds to the same molecule or family of molecules in different species or even to the same molecules within the AOB and MOB of the rat can only be clarified by extracting and characterizing these molecules.
The results of the present study suggest that, in the rat, there are at least two mole- cularly distinct subclasses of olfactory neurons innervating the MOB. Similar results have previously been described in the rat using immunohistochemical procedures [1 ]. A subset of olfactory receptor cells were identified by the monoclonal antibody 2B8. Although the antigen recognized by this antibody was found on all glomeruli in the olfactory bulb some glomeruli contained much higher levels of this molecule. It is unlikely that SBA and 2B8 recognize the same antigen since this lectin, unlike other lectins, was shown to have no affinity for the antigen bound by 2B8 [1]. Two subsets of olfactory receptor cells were also identified in the rabbit [5, 16]. The monoclonal antibody R4BI2 was found to label glomeruli only in the ventro-lateral portion of the MOB. The same antibody also labelled glomeruli only in the rostrolateral portion of the AOB [9]. Although, in the present study, SBA bound to some glomeruli in the ventrolateral MOB of the rat this lectin bound to all glomeruli in the AOB of both rat and mouse. An appropriate comparison between different labelling methods should involve SBA-HRP binding and monoclonal antibody binding in adjacent tis- sue sections. However, evidence available so far suggests different binding patterns.
It would be interesting to label the rat olfactory epithelium with SBA-HRP in or- der to determine the regional location of receptor neurons which terminate in the ventral olfactory bulb. We have already shown in Xenopus that neurons in the middle nasal cavity project to glomeruli in the ventral portion of the olfactory bulb [18].
The classification of primary afferent neurons into subsets is important for under- standing the functional divisions of the sensory system. For example, dorsal root ganglion cells were classified according to their morphology and histology as well as to their biophysical and eleetrophysiological characteristerics [6, 10, 13]. More recently, there have been immunological and histochemical attempts to subclassify these cells [4, 11, 22]. In particular, SBA was found to bind almost exclusively to small dorsal root ganglion cells as well as their central terminations in the rat [22]. These small neurons were suggested to be involved with the transmission of nocicep- tive information [22]. However, the labelling of olfactory receptor cells described in the present study would suggest that SBA labelling is not necessarily associated with the sensory modality of pain.
The olfactory system has provided considerable information on the chemical com- plexity and diversity of neurons [3, 14, 15, 17, 21, 23] and, as such, appears to be a particularly suitable model for studying the functional basis for this molecular heterogeneity. By combining electrophysiology or 2-deoxyglucose methodology with the present SBA-HRP labelling technique it may be possible to define not only the

135
functional differences in odour recognition by subclasses of olfactory receptor cells but also provide an understanding of the role of SBA binding glycoconjugates in primary sensory neurons. Furthermore, SBA-HRP labelling of accessory olfactory neurons may also prove suitable as a marker for the study of cell dynamics during development and regeneration of this system.
This work was supported by an N.H. and M.R.C. Biomedical Scholarship to B.K. and by a University of Queensland Special Project Grant to P.P.G. The attthors wish to thank Dr. W. Brcipohl for critically reading this article.
1 Allen, W.K. and Akeson, R., ldentilication of a cell surface glycoprotcin family of olfactory receptor
neurons with a monoclonal antibody, J. Neurosci., 5 (1985) 284- 296.
2 Altner, It. and Kolnberger. 1., The application of transmission electron microscopy to the study of
the olfactory epithelium of vertebrates, In D.G. Moulton, A. Turk and J.W. Johnson, Jr. ([!ds.), Melll ods in ()lfactory Research, Academic Press, London, 1975, pp. 163 190.
3 Choung, C-M. and Edehnan, G.M., Alterations in neural cell adhesion molecules during development
of different regions of the nerw~us system, J. Neurosci., 4 (1984) 2354 2368.
4 Dodd, J., Solter. D. and Jcssell, T.M., Monoclonal antibodies against carbohydrate differentiation antigens identify subsets of primary sensory neurons, Nature (London), 311 (1984) 469 472.
5 I'ujita, S.('., Mori, K., lmamura, K. and ()bata, K., Subclasses of olfactory reccptor cells and their
segregated central projections demonstrated by a monoclonal antibody, Brain Res., 326 (19851 192 196.
6 Gasser, 11.S.. Effect of method leading on the recording of the nerve fiber spectrum, .1. Gen. Physiol., 43 (1960) 927 940.
7 (Toldstein, l.J. and Hayes, C.E., The lcctins: carbohydrate-binding proteins of plants and animals. In
R.S. Tipton and I). t tor ton (Eds.), Advances in Carbohydrate Chemistry and Biochemistry. Academic Press. New York, 1978, pp. 127 340.
8 Graziudei, P.P.('., The olfactory nmcosa of vertebrates. In L.M. Beidler (Ed.), l t andbook of Scns~rv Physiology, Vol. IV, Springer, New York, 1971, pp. 27 58.
9 lmamuru. K., Mora, K., Fujita, S. and Obata, K., lmmunochemical identification of subgroups of
~omeronasal nerve fibers and their terminations in the accessory olfactory bulb, Brain Res., 328 f 19851 362 366.
10 Lawson, S.N., The postnatal development of large light and small dark neurons in mouse dorsal root
ganglion: a statistical analysis of cell numbers and sizes, J. Neurocytol., 8 (1979) 275 294.
11 Lawson, S.N., Harper, A.A., t tarper, E.I., Garson, J.A. and Anderton, B.H., A monoclonal antibody against neurofilament protein specifically labels a subpopulat ion of rat sensory neurons, J. Comp. Neurol., 228 (1984) 263- 272.
12 LaVail, J.H. and LaVail, M.M., The retrograde intraaxonal transport of horseradish pcroxidasc m the chick visual system: a light and electron microscopic study, J. Comp. Neurol., 157 (1974) 3(13 358.
13 Lloyd, D.P.C., Neuron patterns controlling transmission of ipsilateral hindlimb reflexes in cat, J. Neu- roph3siol., 6 (1943) 293~ 326.
14 Margolis, V.L., A brain protein unique to the olfactory bulb, Proc. Natl. Acad, Sci. USA, 69 (1972) 1221 1224.
15 Molliconc, R., Trojan, J. and Oriol, R., Appearance of It and B antigens in primary sensory cells of the rat olfactory apparatus and inner ear, Dev. Brain Res., 17 (1985) 275-279.
16 Mori, K., Fujila, S.C., lmamura , K. and Obata, K., Immunohistochemical study of subclasses of olfactory nerve fibers and their projections to the olfactory bulb in the rabbit, J. Comp. Neurol.. 242 (1985) 214-229.
17 Muller, L., Mares, V., Sykorova, J. and Biesold, D., Regional and cellular diflizrences in fucosylation of glycomacromolecules in the mouse brain. A biochemical and autoradiographic study of early post- natal and adolescent animals, Neuroscience, 14 (1985) 875-880.

136
l~q Key, B. and Giorgi, P.P., Soybean agglutmm binding in the olfactory system, Neurosci. Lett., Suppt. 23 (1986) 60.
19 Key, B. and Giorgi, P.P., Selective binding of soybean agglutinin to the olfactory system of )(enopu~. Neuroscience, 18 (1986) 507-515.
20 Reeber, A., Vincendon, (;. and Zanetta, J.P., Isolation and immunohistochemical localization of ~ Purkinje cell-specitic glycoprotein subunit from the rat cerebellum, Brain Res., 229 (1981) 53 65+
21 Stallcup, W.B.. geasley. L.L. and Levine, J.M., Antibody against nerve growth factor-inducible lat+ge external (NILE) glycoprotein labels nerve tiber tracts in the developing rat nervous system, J. Neu- rosci., 5 (1985) 1090+ 1011.
22 Streit. W.J., Schutte, B.A.. Balentine, +I.D. and Spicer, S.S., Histochemical localization of galactose- containing glycocoujugates in sensory neurons and their processes in the central and peripheral ner- vous ~,ystcm of the rat+ ,l. Histochem. Cytochem., 33 (1985) 1042-1052.
23 Wallis, 1.+ Ellis. L., Suh, K. and Pfenninger, K.H., Immunolocation of a neuronal growth-dependent membrane glycoprotcin, J. ('ell Biol., 101 (1985) 1990-1998.