soil remediation and plants || recent trends and approaches in phytoremediation
TRANSCRIPT

131Soil Remediation and Plants. http://dx.doi.org/10.1016/B978-0-12-799937-1.00006-1Copyright © 2015 Elsevier Inc. All rights reserved.
Recent Trends and Approaches in Phytoremediation
Bisma Malik, Tanveer Bilal Pirzadah, Inayatullah Tahir, Tanvir ul Hassan Dar and Reiaz Ul RehmanDepartment of Bioresources, University of Kashmir, Srinagar, India
INTRODUCTION
The tremendous progress in scientific technology has evolved human life globally and these developments have raised new issues which pose chal-lenges to the environment, particularly protection and conservation (Bennett et al., 2003). Further, in industrialized society exposure to toxic chemicals and metals becomes unavoidable. Various anthropogenic activities lead to heavy metal enrichment, especially in developing countries where it has become a serious threat to the environment (Wang et al., 2001) by affecting the health of animals as well as humans. The pollution by heavy metals degrades the qual-ity of water, soil and air, thus having deleterious effects on agricultural pro-duction. The governments around the world are duty bound to their citizens to advocate a safe environment free from pollution. However, these environ-mental issues are outweighed by other concerns for the countries’ economic, agricultural and industrial development for ever-increasing populations. Thus this prioritization in a particular direction becomes the driving force actu-ally responsible for environmental pollution (Ikhuoria and Okieimen, 2000). Plants play a pivotal role in transforming solar energy to green energy and have tremendous potential to remediate soil from heavy metals. The conventional methods available currently for the remediation of heavy-metal-contaminated soils are expensive and are not necessarily eco-friendly. However, phytoreme-diation is a novel ‘green technology’ which utilizes plants’ potential to restore the health of the environment. The conclusions drawn on the basis of funda-mental and applied research is that the plants possess tremendous potential for eliminating, degrading or neutralizing a variety of heavy metal toxicants. The wonderful nature of this technology is in its cost effectiveness, simplic-ity, sustainability, environmental compatibility and the fact that it is more aesthetically attractive than the conventional classical technologies. It can be
Chapter 6

132 Soil Remediation and Plants
implemented in situ to remediate large expanses of contaminated ground or to treat large volumes of dilute wastewater. To enhance the efficiency of phytore-mediation it is necessary to understand the biology of the plant kingdom at the molecular level. This review is an attempt to summarize the recent approaches in phytoremediation and how genetic engineering plays an important role in phytoremediation technology.
PHYTOREMEDIATION TECHNOLOGIES
Phytoremediation is a plant-based technology broadly classified into four classes, each having its own mechanism of degrading heavy metals. Figure 6.1. These include the following.
Phytoextraction
Phytoextraction or phytomining is generally an innovative technology to pro-duce bio-ore and restore the health of the soil. This technology involves plant-ing hyperaccumulator species especially in heavy-metal-contaminated soils and later harvesting the biomass leading to the permanent heavy metals removal from the soil. This process can be termed as ‘concentration technology’ which produces smaller mass for disposal in comparison to conventional engineering including excavation and landfilling. Phytoremediation evaluation by Super-fund Innovative Technology Evaluation (SITE) should demonstrate its amena-bility for recovery and recycling of contaminants. As compared to conventional soil remediation technologies, the cost involved in phytoextraction is more than
Redoxreaction &sorption in
organometallic colloids
Co-precipitation
& Dissolution &ion exchange/
Adsorption
SUNSoil inorganicfraction Soil system
Plant SystemPhytoextraction
Folliage
Energy recovery system
GasificationPyrolysis or Direct
combustion
Burner/ ICengine
Fuelgases
Ash, char &Clinkers
Energygeneration
Metalrecovered &
recycled
BIOMASS
PROCESSING
ROOT
STRUCTURE
Stem (Tuber)
Root
Root(Complex in
dead tissues)Complexeswith humus& acid-base reaction
WitheringFree metalion conc. in
soil
Soil organicfractionLitter
breakdown
Minerals
Leaching losses
Biomass µorganisms
Volatilizationlosses
FIGURE 6.1 Soil, plant and green energy recovery system depicting the key components associ-ated with the mass transfer and dynamics of phytoextraction.

133Chapter | 6 Recent Trends and Approaches in Phytoremediation
10-times less per hectare and is therefore considered to be a cost-effective tech-nology (Salt et al., 1995). This technology not only helps in producing bio-ore, but also controls leaching and soil erosion. Besides, the concentration of heavy metals can be reduced after successive cropping and harvesting (Vandenhove et al., 2001). The efficiency of this plant-based technology usually depends on following criteria: (1) site selection is of fundamental importance, i.e. it should be conducive; (2) it must possess the property to accumulate and scavenge bulk quantity of heavy metals (McGrath, 1998); (3) plants must possess fast growth rate and produce large amounts of biomass; (4) bioavailability of heavy metals so that it gets easily absorbed by the plants; (5) acceptable land topography and cheap agronomic practices; (6) capacity to resist hardy soil conditions like soil pH, salinity, soil texture, water content, etc.; root depth also determines the efficiency of effective phytoextraction. Currently, the two main approaches of phytoextraction used include: (i) chelate-assisted phytoextraction or induced phytoextraction – this approach involves the use of artificial chelates in order to enhance the mobility of heavy metal ions so that they become amenable to plants; (ii) continuous phytoextraction – this approach involves the natural capa-bility of plants to uptake and scavenge the toxicants (Salt et al., 1997). Currently research is ongoing to improve the efficiency of phytoextraction. Researchers all over the world target the novel hyperaccumulating plant species and strive to understand their biological pathways. Some plant families are reported to accu-mulate high concentrations of heavy metals which include Asteraceae, Bras-sicaceae, Euphorbiaceae, Fabaceae, Flacourtiaceae and Violaceae (Kumar et al., 1995). Among the Brasicaceae family, some species possess enormous potential to scavenge heavy metal ions and are thus considered as potential can-didates for phytoextraction of metals. Generally, this family is known to scav-enge metals like Pb, Cd, Zn and Ni (Robinson et al., 2009). Although B. juncea that belongs to the family Brassicaceae possess one-third of the concentration of zinc in its tissues, it is more efficient to remediate zinc than T. caerulescens (known zinc hyperaccumulator) and the reason behind this fact is that the bio-mass production of B. juncea is about 10-times more than T. caerulescens (Ebbs and Kochian, 1997). Experimental analysis was carried out in different Bras-sica species in order to check their ability to resist and accumulate heavy met-als. These include: Indian mustard (B. juncea L.), kale (B. oleracea L.), turnip (B. campestris L.), black mustard (B. nigra Koch) and rape (B. napus L.) (Kumar et al., 1995). Among different Brassica species, Indian mustard (B. juncea) is the leading candidate crop to remediate numerous heavy metals from the soil which include: cadmium (Cd), chromium-IV (CrIV), caesium-137 (137Cs) copper (Cu), nickel (Ni), lead (Pb), uranium (U) and zinc (Zn) (Jiang et al., 2000). In another study, scientists at the University of Florida have discovered that the Chinese brake fern, P. vittata, possesses efficient capacity to hyperaccumulate arsenic metal. This fern, when planted at a wood-preserving site containing soil loaded with arsenic (18.8–1603 ppm), exhibits the highest accumulation of arsenic heavy metal (3280–4980 ppm) in its tissues (Ma et al., 2001). Some plant species

134 Soil Remediation and Plants
also have the potential to decontaminate the soil from radionuclides and the best example is Sunflower (H. annus). Experimental analysis revealed that the level of radionuclides such as caesium-137 and strontium-90 in water declines by 90% within a 2-week time period; besides, the highest concentration of these radio-nuclides was found in roots (8000 times) rather than water when flowers were planted as a demonstration of phytoremediation in a pond contaminated with these radioactive elements (Chernobyl nuclear disaster in the Ukraine). Another study carried out by Phytotech for the US Department of Energy, shows that H. annus has tremendous potential to diminish the uranium level at the site from 350 parts per billion to 5 parts per billion, thus achieving about 95% reduc-tion within 24 hours (Schnoor, 1997). Currently very few plants are known for phytoextraction although they possess few desirable traits, but with the help of biotechnological approaches transgenic plants can be developed which possess all the traits essential for phytoextraction technology.
Phytovolatilization
Phytovolatilization generally involves biomethylation of some obnoxious and heavy metals such as selenium, arsenic and mercury and transforming them into volatile molecules that can easily be lost into the atmosphere. This technology is generally applied to remediate groundwater, soil, sediments and sludges. For-merly only microorganisms were known to play this role (Karlson and Franken-berger, 1989), but recently it has been discovered that plants (B. juncea, B. napus) also possess an excellent ability to perform the same function (Terry et al., 1992). A few aquatic plant species, viz. Azolla, rabbit foot grass, rice, cattail (Typha latifolia L.) and pickle weed, also have the ability to act as the best volatilizers of selenium (Zayed et al., 2000). Nowadays research is focussed on selenium volatilization due to its deleterious effects in many parts of the world. Gener-ally, phytovolatilization has been utilized to decontaminate soil from mercury as it converts mercuric ions into less toxic elemental mercury. The mechanism behind selenium phytoremediation involves assimilation of inorganic Se and later converting it into organic forms such as selenoaminoacids, selenocysteine (SeCys) and selenomethionine (SeMet). These compounds are further processed and can be methylated to form dimethylselenide (DMSe), which is volatile in nature (Terry et al., 2000). Microorganisms have the ability to transform mercury (II) the known toxic element into organomercurials which, in turn, changes into elemental mercury (less toxic) under optimal conditions (Bizily et al., 2000) and later enters into the global biogeochemical cycle upon volatilization. As the pro-cess of volatilization does not require plant harvesting and disposal, it becomes an attractive and cheap remediation technology (Rugh et al., 2000). Meagher et al. (2000) carried out a risk assessment study for volatile selenium and mer-cury and the results revealed that these pollutants were dispersed and diluted in such a way that they did not pose a threat to the ecosystem. Although the pace of the phytovolatilization process is very slow, it can be enhanced by using

135Chapter | 6 Recent Trends and Approaches in Phytoremediation
novel plant species with high rates of transpiration, overexpression of enzymes such as cystathionine-V-synthase that mediates S / Se volatilization (Van Huysen et al., 2003) and also by means of transgenic technology to produce transgenic plants with efficient selenium volatilization (Le Duc et al., 2004). Recently, a bacterial Hg ion reductase gene was incorporated into plants to produce a trans-genic plant which shows excellent mercury volatilization. In addition, it has also been reported that bacterial organomecurial lyase (merB) and mercuric reduc-tase (merA) genes were incorporated into model plants such as A. thaliana and N. tabacum; the resulting transgenic plants have the potential to absorb elemen-tal mercury (II) as well as methyl mercury from the soil and transform it into a volatile form (Hg0) (Heaton et al., 1998). Transgenic yellow poplar (Lirioden-dron tulipifera) plantlets were produced which exhibited resistance to and grew well in normally toxic levels of ionic mercury, and the transformed plantlets volatilized about 10-times more elemental mercury than non-transgenic plantlets (Rugh et al., 1998). Moreover it has also been revealed that the addition of Hg0 into the atmosphere would not contribute significantly to the atmospheric pool. Higher plants release the volatile selenium and it was first reported by Lewis et al. (1966). Few members of Brassicaceae family are known to release 40 g of selenium ha−1 / day as a volatile compound. Volatilization of these toxic metals is essential in various aspects such as the volatile form of selenium (dimethylsele-nide) is less toxic than inorganic forms of selenium found in soil (DeSouza et al., 2000). Furthermore, it also involves the permanent removal of these toxicants (Atkinson et al., 1990). It is also recommended that phytovolatilization is not applicable for the sites near population centres or at places with unique meteo-rological conditions as it promotes the rapid deposition of volatile compounds (Rugh et al., 2000). Although this technology is a promising tool for the reme-diation of selenium and mercury, at the same time it has some demerits like loss of control of volatile compounds over their migration to other areas. Currently by means of this technology, tritium (3H), a radioactive isotope of hydrogen is decayed to stable helium with a half-life of about 12 years as reported Dushen-kov et al. (1995).
Phytostabilization
Phytostabilization, also known as phytorestoration or in-place inactivation, is generally used for the remediation of soil, sediment and sludges (United States Protection Agency, 2000). One of the important applications of this technol-ogy is to stabilize the toxicants and prevent exposure to wind and water ero-sion. In addition, it provides hydraulic control to suppress the migration of these toxicants into the water table and thus physiochemically immobilizes contami-nants by root sorption, precipitation, complex action or metal valence reduction with different soil amendments (Berti and Cunningham, 2000). The main aim of this technique is not to restore sites from toxicants but rather to stabilize them and reduce the risks to the ecosystem. It is usually meant for those soils

136 Soil Remediation and Plants
contaminated with Zn / As / Cr / Cd / Pb / Cu. Generally the role of plants in this technique is to reduce the amount of water percolating through the soil matrix that will ultimately lead to the formation of toxicants (hazardous leachates) and avoid soil erosion and transport of toxic metals to distinct areas. The plants cho-sen for this technology should obey certain criteria: (1) efficiency of transloca-tion of heavy metals from root-to-shoot system should be low; (2) they should have a fast growth rate and resistance to heavy metals and (3) there should be cost-effective management. Practically this technique is not only applicable at sites with high organic load and porosity but is also effective for a wide range of surface contamination sites (Berti and Cunningham, 2000). One of the draw-backs of this technique is that it is not applicable to those areas which are heav-ily contaminated because such conditions become a hindrance in plant growth and development (Berti and Cunningham, 2000). Besides, the contaminant remains in soil as it is, and therefore requires regular monitoring.
Rhizofiltration
This phytoremediation technique is usually intended to restore aquatic ecosys-tems. It involves the use of both aquatic as well as terrestrial plants to treat industrial discharge, agricultural runoff or acid mine drainage. It is mainly used for the removal of Ni, Cd, Cu, Zn, Cr and Pb which are primarily retained within the roots (United States Protection Agency, 2000). Formerly some chemical- and mechanical-based techniques such as precipitation, flocculation followed by sedimentation were utilized for the removal of industrial pollutants. How-ever, there are certain limitations to this conventional technology as it is not eco-friendly and cost-effective. Thus, the rhizofiltration technique which is sus-tainable, eco-friendly and cost-effective has gained worldwide importance to restore contaminated sites. In this process, plants are raised hydroponically and later transplanted into contaminated sites in order to absorb and concentrate the toxicants in their roots and shoots (Zhu et al., 1999b). It has also been reported that some chemicals ooze out of the roots that lead to a change in rhizosphere pH which ultimately causes metals to precipitate on to the root surface. Once roots get saturated with these toxicants, either only the roots or the whole plants are harvested for further processing (Zhu et al., 1999b). This process is not feasible in those areas where metal concentration is high in water because the contaminants should be in the solution form in order to be sorbed into the plant system. Other complications associated with this technique involve dry-ing, composting or incineration. For efficient removal of contaminants from the site, plants must possess efficient and fast-growing root systems with the abil-ity to scavenge these toxicants and thus neutralize their harmful effects. Low maintenance costs, easy handling and resistance of plants to heavy metals are other important criteria for effective removal of heavy metals from a particular area. A number of aquatic plant species show a better response in the removal of toxicants from water. These include: water hyacinth (Eichhornia crassipes

137Chapter | 6 Recent Trends and Approaches in Phytoremediation
Mart.; Zhu et al., 1999c), pennywort (Hydrocotyle umbellata L; Dierberg et al., 1987) and duckweed (Lemna minor L; Mo et al., 1989). However, there are certain limitations to these plants as they have limited potential for rhizofil-tration, because of their small, slow-growing root systems (Dushenkov et al., 1995). Zhu et al. (1999c) reported that water hyacinth is a potential candidate for the removal of trace elements from waste streams. A number of plants such as tobacco, corn, rye, spinach, Indian mustard and sunflower have been studied for their ability to remove Pb from effluents and the results revealed that sun-flower has the greatest potential to detoxify Pb content. Experimental evidences show that terrestrial plants with dense and fibrous root systems are suitable for this technique as they possess greater metal-absorbing powers, and the lead-ing examples include Sunflower (Helianthus annuus L.) and Indian mustard (Brassica juncea Czern.). Indian mustard is known to remove a wide concentra-tion of Pb (4–500 mg l−1) (Raskin and Ensley, 2000). These terrestrial plants are also being used to remove various metals such as Cd, Zn, Cu, Ni and Cr (Dush-enkov et al., 1995), uranium (Dushenkov et al., 1997a) and 90Sr ( Dushenkov et al., 1997b) from hydroponic solutions. Currently, researchers are in the process of improving the efficiency of rhizofiltration technology in order to decontaminate the soil. Salt et al. (1995) carried out an experimental analysis and concluded that young seedlings exhibit better abilities to remove heavy metals from water. This technique of using young plant seedlings in order to remove toxicants (heavy metals) from water is known as blastofiltration (blasto is ‘seedling’ in Greek) and is considered to be second-generation, plant-based water treatment technology. In this technique there is a dramatic enhancement in surface to volume ratio that usually occurs after germination and some ger-minating seedlings also ab / adsorb huge quantities of toxic metal ions; this is why young seedlings are suitable for restoring water quality. Indian mustard is considered to be a potential crop for blastofiltration as it is effective in sorbing divalent cations of toxic metals (Salt et al., 1997) due to some unique features of Indian mustard such as fast growth rate and resistance to heavy metals or microbial infection. Through data analysis, it has been reported that for a few metals, blastofiltration techniques are more potent and economical than rhizo-filtration; however the main advantage of rhizofiltration is that it can be utilized both in situ as well as ex situ and species other than hyperaccumulators can also be taken into account.
GENETIC ENGINEERING TO IMPROVE PHYTOREMEDIATION
Genetic engineering is one of the important approaches that can be used as an alternative to improve the phytoremediation potential of plant species with high biomass production. It was very hard to develop general plant productiv-ity, which is regulated by several genes, by the insertion of only one gene. Biotechnological approaches (genetic engineering) are used to insert more effective accumulator genes into other plants, and these methods have been

138 Soil Remediation and Plants
recommended by various authors (Chaney et al., 2000). This approach of inserting many effective accumulator genes from taller plants into other natural plants enhanced the final biomass. Zhu et al. (1999a) studied the transgenic Brassica juncea for rate-limiting factors by inserting gshl-gene from Escherichia coli for glutathione and phytochelatin production. The γ-ECS genetically engi-neered seedlings exhibited improved resistance to Cd and contained greater amounts of phytochelatins, γ-GluCys, glutathione and totally non-proteinthi-ols when compared to wild-type seedlings. There are several factors that restrict the achievement of genetic engineering and one of the limiting factors is the anatomical restraint (Ow, 1996). Commercial utilization of phytoreme-diation can be increased by transforming metal-scavenging properties of hyperaccumulating plants, such as T. caerulescens, to high-biomass generating species, such as Indian mustard (Brassica juncea) or maize (Zea mays) (Brown et al., 1995). At present, genetic engineering is successfully utilized to manipu-late metal uptake and stress resistance properties in various species. For exam-ple, enhanced metal resistance in tobacco (Nicotiana tabacum) was achieved by expressing the mammalian metallothionein metal binding proteins (Maiti et al., 1991). Currently, genetic engineering plays a pivotal role in the produc-tion of transgenic plant species that have tremendous potential to remediate soil contaminated with methyl mercury (a neurotoxic agent). Examples are transgenic Tobacco and Arabidopsis which express bacterial genes merB and merA, and have the capacity to remove mercury from the soil. The merB gene present in these transgenic plants has the capacity to carry out protonolysis of the carbon–mercury bond and thus liberate Hg2+, a reduced mobile mercury species, and the MerA gene converts Hg2+ to Hg0 (elemental-Hg) a less toxic, volatile element which is liberated into the atmosphere (Rugh et al., 1996). Despite the fact that regulatory concerns have confined the application of genetically engineered plants transformed with merA and merB, this work demonstrates the enormous capability of genetic engineering for restoration of the environment. In order to declaim these regulatory concerns related to phy-tovolatilization of mercury, Bizily et al. (1999) illustrated genetically engi-neered plants that express MerBpe (an organomercurial lyase under the control of a plant promoter) employed to decompose methyl mercury and consequently take-off ionic mercury through extraction. In spite of current progress in bio-technology, less is known about the eugenics of metal hyperaccumulation in plants. Specifically, the genetics pertinent to plant mechanisms, such as metal transport and storage (Lasat et al., 2000) and metal resistance (Ortiz et al., 1992) must be clearly explained. Chaney et al. (1999) recently suggested the application of conventional breeding processes for enhancing metal-hyperac-cumulating species and bioengineering by inducing some important traits such as metal resistance and uptake characteristics into high-biomass-yielding plants. Complete success has not been reported in the literature. For example, Brewer et al. (1997) in an investigation attempted to correct the small size of hyperaccumulating plants by producing somatic hybrids between

139Chapter | 6 Recent Trends and Approaches in Phytoremediation
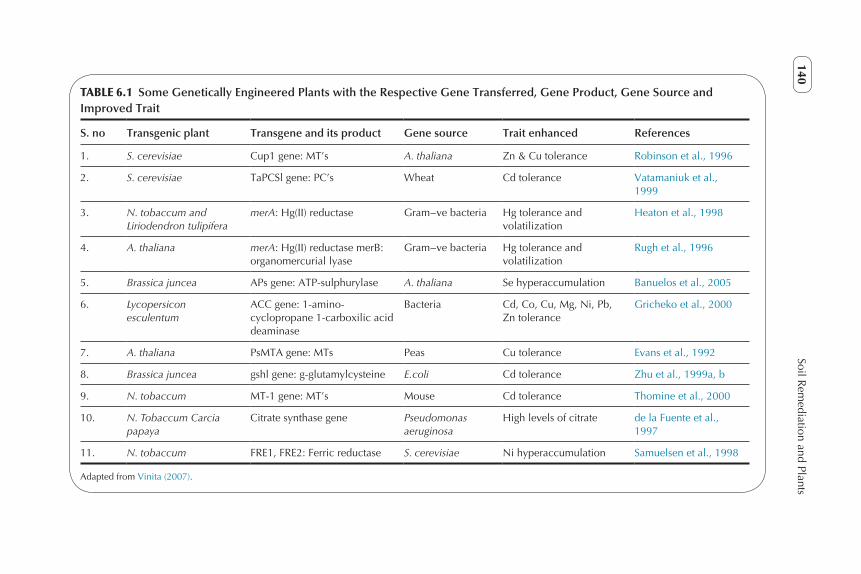
T. caerulescens (a Zn hyperaccumulator) and Brassica napus (canola) fol-lowed by hybrid selection for Zn resistance. High biomass hybrids with extreme Zn resistance were obtained. These researchers have also made an effort to collect and conserve germplasm of hyperaccumulating species. Com-prehensively, any dicotyledonous plant species can be modified by employing the Agrobacterium vector system, while as the majority of the monocotyledon-ous plants can be reconstructed employing particle gun or electroporation methods. Several feasible methods have been developed that describe greater resistance, concentration and / or degradation capabilities of several adulter-ants. These were carried out either by the overproduction of citrate (metal chelating molecule) (de la Fuente et al., 1997), phytochelatins (PCs) (Zhu et al., 1999a, b), metallothioneins (MTs) (Hasegawa et al., 1997) or ferritin (Goto et al., 1999) or by overexpression of metal transporter proteins (Curie et al., 2001). Using these bioengineering techniques in order to enhance the metal concentration in plants in turn increases the phytoremediation capability by the same factor. However, it has not been fully investigated how appropriate these genetically modified plants are for environmental restoration, because only a little field work has been done so far, including one study using geneti-cally modified Indian mustard plant that overproduces enzymes that catalyse the sulphate / selenate reduction process (Pilon-Smits et al., 1999). Also, it has been suggested by many researchers that the consequences of the genetically modified plants should be regularly observed, such as the competition between altered plants and wild ones, the influence on birds, insects, etc., that might feed on plant biomass exhibiting extreme amount of noxious metals, and also there is a probability of gene transfer to other plants through pollination. Bio-engineering of the chloroplast genome put forward a unique alternative to acquire high expression and reduce the risk of gene transfer through pollina-tion (Ruiz et al., 2003). Table 6.1 lists some of the transgenic plants, genes incorporated, gene sources and the enhanced trait of modified plants (Vinita, 2007). Recently, the transcriptomic studies on hyperaccumulators have pro-vided novel insights into molecular mechanisms underlying metal resistance and concentration, as well as reaching into the classification of a huge pattern of genes which are constitutively (in the absence of excess of metallic ions) overexpressed in the hyperaccumulation trait (Filatov et al., 2006). Classic genetic studies and recent Quantitative trait loci (QTL) analysis of hyperac-cumulators support the existence of only a few genes that are responsible for tolerance to accumulation (Willems et al., 2007). Genetic engineering of metal transporters also plays a vital role in phytoremediation, as these are extremely complex and diverse. HMA4 [heavy metal transporting adenosine triphospha-tase (ATPase)] was the first plant P1B- ATPase of the divalent transport group to be cloned and characterized in A. thaliana (Mills et al., 2003). The function of HMA4 has been described in Arabidopsis thaliana for Zn homeostasis, removal of Cd and in the transportation of these metals from the root to the shoot (Verret et al., 2005). In A. halleri and T. caerulescens, which are
140Soil R
emediation and Plants
TABLE 6.1 Some Genetically Engineered Plants with the Respective Gene Transferred, Gene Product, Gene Source and Improved Trait
S. no Transgenic plant Transgene and its product Gene source Trait enhanced References
1. S. cerevisiae Cup1 gene: MT’s A. thaliana Zn & Cu tolerance Robinson et al., 1996
2. S. cerevisiae TaPCSl gene: PC’s Wheat Cd tolerance Vatamaniuk et al., 1999
3. N. tobaccum and Liriodendron tulipifera
merA: Hg(II) reductase Gram−ve bacteria Hg tolerance andvolatilization
Heaton et al., 1998
4. A. thaliana merA: Hg(II) reductase merB: organomercurial lyase
Gram−ve bacteria Hg tolerance andvolatilization
Rugh et al., 1996
5. Brassica juncea APs gene: ATP-sulphurylase A. thaliana Se hyperaccumulation Banuelos et al., 2005
6. Lycopersicon esculentum
ACC gene: 1-amino-cyclopropane 1-carboxilic acid deaminase
Bacteria Cd, Co, Cu, Mg, Ni, Pb, Zn tolerance
Gricheko et al., 2000
7. A. thaliana PsMTA gene: MTs Peas Cu tolerance Evans et al., 1992
8. Brassica juncea gshl gene: g-glutamylcysteine E.coli Cd tolerance Zhu et al., 1999a, b
9. N. tobaccum MT-1 gene: MT’s Mouse Cd tolerance Thomine et al., 2000
10. N. Tobaccum Carcia papaya
Citrate synthase gene Pseudomonas aeruginosa
High levels of citrate de la Fuente et al., 1997
11. N. tobaccum FRE1, FRE2: Ferric reductase S. cerevisiae Ni hyperaccumulation Samuelsen et al., 1998
Adapted from Vinita (2007).

141Chapter | 6 Recent Trends and Approaches in Phytoremediation
considered to be Cd / Zn hyperaccumulators, the HMA4 shows overexpression both in roots and shoots in comparison to Cd- / Zn-sensitive close relatives (Courbot et al., 2007). Two distinct autonomously produced Zn / Cd hyperac-cumulating plant species with increased expression of HMA4 have been found, powerfully supporting the concept that HMA4 plays a vital role in resistance to both metals. In A. halleri this presumption is supported by the QTL analysis of Cd and Zn resistance indicating the co-localization of major QTLs of Cd and Zn resistance with HMA4 gene (Courbot et al., 2007). At present there is genetic evidence for only one gene, i.e. HMA4, which has a function in both Zn and Cd resistance. HMA4 is involved in the detoxification of roots by trans-porting Zn and Cd to the shoots as observed in AtHMA4-overexpressing lines (Verret et al., 2004), which translocate more Zn and Cd to the shoot and also show resistance to hyperaccumulation of Zn and Cd. The enhanced expression of HMA4 in A. halleri or T. Caerulescens may be related to the enhanced resis-tance to Zn and Cd and the hyperaccumulation phenotype of these species (Willems et al., 2007). Several ZIP (zinc regulated transporters, iron regulated transporter proteins) members are overexpressed in A. halleri and T. caerules-cens (ZIP 3, 4, 6, 9 and 10 and IRT3) and are supposed to function by increas-ing root metal uptake of those Zn hyperaccumulators (Kramer et al., 2007). IRT1, one of the first-discovered members of the large ZIP family, is involved in the uptake of Fe2+ from the soil. Plants overexpressing IRT1 engineered to improve Fe nutrition also accumulated more Cd (Cannoly et al., 2002). Also some MATE (multi-drug and toxin extrusion) member proteins, e.g. FRD3, are constitutively overexpressed in A. halleri compared to its close relative and play an important role in Zn translocation (Talke et al., 2006). Overexpression of ZAT (zinc transporter of Arabidopsis Thaliana) in A. thaliana described extreme Zn resistance and a two-fold enhanced Zn concentration in roots (Van der Zaal et al., 1999). Manganese-resistant Strylosanthes hamate (a tropical legume) includes ShMTP in the vacuolar storage of manganese and when over-expressed in A. thaliana confirmed greater resistance and concentration of manganese (Delhaize et al., 2003). Using CAX (CAtioneXchanger) under the influence of the constitutive 35S CaMV promoter it has been revealed that the expression of AtCAX2 and AtCAX4 in root vacuoles of A. thaliana resulted in greater deposition of Cd (Korenkov et al., 2007). Another study described that the overexpression of SMT (Se cysMethylTransferase) from Se-hyperaccumu-lator A. bisulcatus in A. thaliana and Indian mustard resulted in the consider-able enhancement of Se resistance, concentration and volatilization in both species (Le Duc et al., 2006). Interestingly, constitutive expression of existing genes seems to be a general process in the acclimatization of plants into the worst environmental conditions (Taji et al., 2004). Novel powerful tools to control the timing, the location and the level of the transgene expression are expected from the characterization of native promoters of hyperaccumulators. Future studies require more information from fieldwork and risk assessment monitoring along with biotechnological techniques; this will directly result in the commercializing of phytoremediation on a large scale.

142 Soil Remediation and Plants
CONCLUSIONS AND FUTURE PERSPECTIVES
Phytoremediation is currently an emerging technology and is a fast-growing field involving the approaches corresponding with green technology. Scientists as well as the general masses around the world are concerned about the chang-ing environmental conditions, and since the last decade we have witnessed the initiation of many field applications including the phytoremediation of organic, inorganic and radionuclides. Phytoremediation, being a cost-effective, sustain-able and eco-friendly technology, is one of the feasible alternatives, especially in developing countries. This technology can be applied in situ to immediate shallow soil, ground water and runoff water bodies. Currently, phytoremedia-tion technology is in its infancy and there are many technical hindrances that need to be addressed. However, with the help of modern innovative agronomic practices and genetic engineering approaches to its efficiency, this green tech-nology would be enhanced. Many hyperaccumulator plants remain to be dis-covered and there is an urgent need to explore more about their physiology and molecular biology. Optimization of the process, proper understanding of the dynamics of plant–heavy metal interactions, plant–microbe interactions and proper disposal of biomass produced is still needed.
REFERENCES
Atkinson, R., Aschmann, S.M., Hasegawa, D., Eagle-Thompson, E.T., Frankenberger Jr., W.T., 1990. Kinetics of the atmospherically important reactions of dimethyl selenide. Environ. Sci. Technol. 24, 1326–1332.
Banuelos, Terry, D.N., Leduc, D.L., Pilon Smits, E.A., Mackey, B., 2005. Field trial of transgenic Indian mustard plants shows enhanced phytoremediation of selenium contaminated sediment. Environ. Sci. Technol. 39 (6), 1771–1777.
Bennett, L.E., Burkhead, J.L., Hale, K.L., Terry, N., Pilon, M., Pilon-smits, E.A.H., 2003. Analysis of transgenic Indian Mustard plants for phytoremediation of metals-contaminated mine tail-ings. J. Environ. Qual. 32, 432–440.
Berti, W.R., Cunningham, S.D., 2000. Phytostabilization of metals. In: Raskin, I., Ensley, B.D. (Eds.), Phytoremediation of toxic metals – using plants to clean-up the environment. John Wiley & Sons, Inc, New York, pp. 71–88.
Bizily, S.P., Clayton, L.R., Summers, A.O., Meagher, R.B., 1999. Phytoremediation of methylmer-cury pollution: merB expression in Arabidopsis thaliana confers resistance to organomercuri-als. Proc. Natl. Acad. Sci. 96, 6808–6813.
Bizily, S.P., Rugh, C.L., Meagher, R.B., 2000. Phytodetoxification of hazardous organomercurials by genetically engineered plants. Nat. Biotechnol. 18, 213–217.
Brewer, E.P., Saunders, J.A., Angle, J.S., Chaney, R.L., McIntosh, M.S., 1997. Somatic hybridiza-tion between heavy metal hyperaccumulating Thlaspi caerulescens and canola. Agron Abstr. 154.
Brown, S.L., Chaney, R.L., Angle, J.S., Baker, A.J.M., 1995. Zinc and cadmium uptake by hyperac-cumulator Thlaspi caerulescens grown in nutrient solution. Soil Sci. Soc. Am. J. 59, 125–133.
Cannoly, E.L., Fett, J.P., Guerinot, M.L., 2002. Expression of the IRT1 metal transporter is con-trolled by metals at the levels of transcript and protein accumulation. Plant Cell 14, 1347– 1357.

143Chapter | 6 Recent Trends and Approaches in Phytoremediation
Chaney, R.L., Li, Y.M., Angle, J.S., Baker, A.J.M., Reeves, R.D., Brown, S.L., Homer, F.A., Malik, M., Chin, M., 1999. Improving metal hyperaccumulators wild plants to develop commercial phytoextraction systems: Approaches and progress. In: Terry, N., Bañuelos, G.S. (Eds.), Phy-toremediation of Contaminated Soil and Water. CRC Press, Boca Raton, FL.
Chaney, R.L., Li, Y.M., Angle, J.S., Baker, A.J.M., Reeves, R.D., Brown, S.L., Homer, F.A., Malik, M., Chin, M., 2000. In: Terry, N., Banelos, G. (Eds.), Phytoremediation of contaminated soil and water. Lewis Publishers, Boca Raton, FL, pp. 129–158.
Courbot, M., Willems, G., Motte, P., Arvidsson, S., Roosens, N., Saumitou-laprade, P., Verbrugger, N., 2007. A major quantitative trait locus for cadmium tolerance in Arabidopsis halleri colo-calizes with HMA4, a gene encoding a heavy metal ATPase. Plant Physiol. 144, 1052–1065.
Curie, C.Z., Panaviene, C., Loulergue, S.L., Dellaporta, J.F., Briat, Walker EL., 2001. Maize yellow stripe 1 encodes a membrane protein directly involved in Fe(III) uptake. Nature 409, 346–349.
de la Fuente, J.M., Ramírez-Rodriguez, V., Cabrera-Ponce, J.L., Herrera-Estrella, L., 1997. Alumi-num tolerance in transgenic plants by alteration of citrate synthesis. Science 276, 1566–1568.
Delhaize, E., Kataoka, T., Hebb, D.M., White, R.G., Ryan, P.R., 2003. Genes encoding proteins of the cation diffusion facilitator family that confer manganese tolerance. Plant Cell 15, 1131–1142.
DeSouza, M.P., Pilon-Smits, E.A.H., Terry, N., 2000. The physiology and biochemistry of selenium volatilization by plants. In: Raskin, I., Ensley, B.D. (Eds.), Phytoremediation of Toxic Metals – Using Plants to Clean-Up the Environment. John Wiley & Sons, Inc., New York, pp. 171–190.
Dierberg, F.E., DeBusk, T.A., Goulet Jr., N.A., 1987. In: Reddy, K.R., Smith, W.H. (Eds.), Aquatic Plants for Water Treatment and Resource Recovery. Magnolia Publishing, Orlando, FL, pp. 497–504.
Dushenkov, V., Kumar, P.B.A.N., Motto, H., Raskin, I., 1995. Rhizofiltration: The use of plants to remove heavy metals from aqueous streams. Environ. Sci. Technol. 29, 1239–1245.
Dushenkov, S., Vasudev, D., Kapulnik, Y., Gleba, D., Fleisher, D., Ting, K.C., Ensley, B., 1997a. Removal of uranium from water using terrestrial plants. Environ. Sci. Technol. 31, 3468–3474.
Dushenkov, S., Vasudev, D., Kapolnik, Y., Gleba, D., Fleisher, D., Ting, K.C., Ensley, B., 1997b. Environ. Sci. Technol. 31 (12), 3468–3476.
Ebbs, S.D., Kochian, L.V., 1997. Toxicity of zinc and copper to Brassica species: Implications for phytoremediation. J. Environ. Qual. 26, 776–781.
Evans, K.M., Gatehouse, J.A., Lindsay, W.P., Shi, J., Tommey, A.M., Robinson, N.J., 1992. Expres-sion of the pea metallothionein like gene PsMTA in Escherichia coli and Arabidopsis thaliana and analysis of trace metal ion accumulation: implications for gene PsMTA function. Plant Mol. Biol. 20, 1019–1028.
Filatov, V., Dowdle, J., Smirnoff, N., Ford-Llyod, B., Newburry, H.J., Macnain, M.M., 2006. Comparison of genes expression in segregating families identifies genes and genomic regions involved in a novel adaptation, zinc hyperaccumulation. Mol. Ecol. 15, 3045–3059.
Goto, F., Yoshihara, T., Shigemoto, N., Toki, S., Takaiwa, F., 1999. Iron fortification of rice seed by the soybean ferritin gene. Nature Biotechnol. 17, 282–286.
Gricheko, V.P., Filby, B., Glick, B.R., 2000. Increased ability of transgenic plants expressing the bacterial enzyme ACC deaminase to accumulate Cd, Co, Cu, Ni, Pb and Zn. J. Biotech. 81, 45–53.
Hasegawa, I., Terada, E., Sunairi, M., Wakita, H., Shinmachi, F., 1997. Genetic improvement of heavy metal tolerance in plants by transfer of the yeast metallothionein gene (CUP1). Plant Soil 196, 277–281.
Heaton, A.C.P., Rugh, C.L., Wang, N., Meagher, R.B., 1998. Phytoremediation of mercury and methyl mercury polluted soils using genetically engineered plants. J. Soil Contam. 7 (4), 497–509.

144 Soil Remediation and Plants
Ikhuoria, E.U., Okieimen, F.E., 2000. Scavenging Cadmium, copper, lead, nickel and zinc ions from aqueous solution by modified cellulosic sorbent. Int. J. Environ. Stud. 57 (4), 401.
Jiang, W., Liu, D., Hou, W., 2000. Hyperaccumulation of lead by roots, hypocotyls, and shoots of Brassica juncea. Biol. Plantarum 43 (4), 603–606.
Karlson, U., Frankenberger, W.T., 1989. Accelerated rates of selenium volatilization from Califor-nia soils. Soil Sci. Soc. Am. J. 53, 749–753.
Korenkov, V., Park, S.H., Chang, N.H., Sreevidya, C., Jlachmansingh, J., Hirschi, K., Wagner, G.J., 2007. Enhanced Cd2+ selective root-transport-transport in tobaccos expressing Arabidopsis cat-ion exchangers. Planta 225, 403–411.
Kramer, U., Talke, I., Hanikenne, M., 2007. Transition metal transport. FEBS Lett. 581, 2263– 2272.
Kumar, P.B.A.N., Dushenkov, V., Motto, H., Raskin, I., 1995. Phytoextraction: The use of plants to remove heavy metals from soils. Environ. Sci. Technol. 29, 1232–1238.
Lasat, M.M., Pence, N.S., Garvin, D.F., Ebbs, S.D., Kochian, L.V., 2000. Molecular physiology of zinc transport in the Zn hyperaccumulator Thlaspi caerulescens. J. Exp. Bot. 51 (342), 71–79.
Le Duc, D.L., Tarun, A.S., Montes-Bayon, M., Meija, J., Malit, M.F., 2004. Overexpression of sele-nocysteine methyltransferase in Arabidopsis and Indian mustard increases selenium tolerance and accumulation. Plant Physiol. 135, 377–383.
LeDuc, D.L., Abdelsamie, M., Montes-Bayon, M., Wu, C.P., Reisinger, S.J., Terry, N., 2006. Over-expressing both ATPsulfurylase and selenocysteine methyl transferase enhances selenium phy-toremediation traits in Indian mustard. Environ. Pollut. 144, 70–76.
Lewis, B.G., Johnson, C.M., Delwiche, C.C., 1966. Release of volatile selenium compounds by plants: Collection procedures and preliminary observations. J. Agric. Food Chem. 14, 638–640.
Ma, L.Q., Komar, K.M., Tu, C., 2001. A fern that accumulates arsenic. Nature 409, 579.Maiti, I.B., Wagner, G.J., Hunt, A.G., 1991. Light inducible and tissue specific expression of achi-
meric mouse metallothionein cDNA gene in tobacco. Plant Sci. 76, 99–107.McGrath, S.P., 1998. Phytoextraction for soil remediation. In: Brooks, R.R. (Ed.), Plants that hyper-
accumulate heavy metals: Their role in phytoremediation, microbiology, archeology, mineral exploration and phytomining. CAN International, New York, pp. 261–288.
Meagher, R.B., Rugh, C.L., Kandasamy, M.K., Gragson, G., Wang, N.J., 2000. Engineered phytore-mediation of mercury pollution in soil and water using bacterial genes. In: Terry, N., Banuelos, G. (Eds.), Phytoremediation of contaminated soil and water. Lewis, Boca Raton, pp. 201–221.
Mills, R.F., Krijger, G.C., Baccarini, P.J., Hall, J.L., Williams, L.E., 2003. Functional expression of AtHMA4, a p-13-type ATPase of the Zn / Co / Cd / Pb subclass. Plant J. 35, 164–176.
Mo, S.C., Choi, D.S., Robinson, J.W., 1989. Uptake of mercury from aqueous solution by duck-weed: The effect of pH, copper, and humic acid. J. Environ. Health Sci. A. 24, 135–146.
Ortiz, D.F., Kreppel, L., Speiser, D.M., Scheel, G., McDonald, G., Ow, D.V., 1992. Heavy metal tolerance in the fission yeast requires an ATP-binding cassette-type vacuolar membrane trans-porter. EMBO J. 11, 3491–3499.
Ow, D.W., 1996. Heavy metal tolerance genes—prospective tools for bioremediation. Res. Conserv. Recycling 18, 135–149.
Pilon-Smits, E.A.H., de Souza, M.P., Hong, G., Amini, A., Bravo, R.C., 1999. Selenium volatiliza-tion and accumulation by twenty aquatic plant species. J. Environ. Qual. 28, 1011–1017.
Raskin, I., Ensley, B.D., 2000. Phytoremediation of Toxic Metals: Using Plants to Clean Up the Environment. John Wiley & Sons, Inc., New York. 53–70.
Robinson, N.J., Wilson, J.R., Turner, J.S., 1996. Expression of the type 2 metallotionein-like gene MT2 from Arabidopsis thaliana in Zn2+- metallothionein deficient Synechococcus PCC 7942: Putative role for MT2 in Zn2+-metabolism. Plant Mol. Biol. 30, 1169–1179.

145Chapter | 6 Recent Trends and Approaches in Phytoremediation
Robinson, B.H., Banuelos, G., Conesa, H.M., Evangelon, W.H., Schulin, R., 2009. The phytoman-agement of trace elements in soil. Crit. Rev. Plant Sci. 28 (4), 240–266.
Rugh, C.L., Bizily, S.P., Meagher, R.B., 2000. Phytoreduction of environmental mercury pollution. In: Raskin, I., Ensley, B.D. (Eds.), Phytoremediation of Toxic Metals – Using Plants to Clean-Up the Environment. John Wiley & Sons, Inc., New York, NY, pp. 151–170.
Rugh, C.L., Gragson, G.M., Meagher, R.B., Merkle, S.A., 1998. Toxic mercury reduction and reme-diation using transgenic plants with a modified bacterial gene. Hort. Sci. 33 (4), 618–621.
Rugh, C.L., Wilde, H.D., Stack, N.M., Thompson, D.M., Summers, A.O., Meagher, R.B., 1996. Mercuric ion reduction and resistance in transgenic Arabidopsis thaliana plants expressing a modified bacterial merA gene. Proc. Natl. Acad. Sci. USA 93, 3182–3187.
Ruiz, O.N., Hussein, H.S., Terry, N., Daniell, H., 2003. Phytoremediation of organomercurial com-pounds via chloroplast genetic engineering. Plant Physiol. 132, 1344–1352.
Salt, D.E., Blaylock, M., Kumar, N.P.B.A., Dushenkov, V., Ensley, B.D., Chet, I., Raskin, I., 1995. Phytoremediation: A novel strategy for the removal of toxic metals from the environment using plants. Biotechnol. 13, 468–475.
Salt, D.E., Pickering, I.J., Prince, R.C., Gleba, D., Dushenkov, S., Smith, R.D., Raskin, I., 1997. Metal accumulation by aquacultured seedlings of Indian Mustard. Environ. Sci. Technol. 31 (6), 1636–1644.
Samuelsen, A.I., Martin, R.C., Mok, D.W.S., Machteld, C.M., 1998. Expression of the yeast FRE genes in transgenic tobacco. Plant Physiol. 118, 51–58.
Schnoor, J.L., 1997. Phytoremediation. University of Lowa, Department of Civil and Engineering. 1, 62.
Taji, T., Seki, M., Satou, M., Sakurai, T., Kobayashi, M., Ishiyama, K., Narusaka, M., Zhu, J.K., Shinozaki, K., 2004. Comparative genomics in salt tolerance between Arabidopsis and Arabi-dopsis- Related halophyte salt cress using Arbidopsis microarray. Plant Physiol. 135, 1697– 1709.
Talke, I., Hanikenne, M., Kramer, U., 2006. Zinc dependent global transcriptional control, tran-scriptional de-regulation and higher gene copy number for genes in metal homeostasis of the hyperaccumulator Arabidopsis halleri. Plant Physiol. 142, 148–167.
Terry, N., Carlson, C., Raab, T.K., Zayed, A., 1992. Rates of selenium volatilization among crop species. J. Environ. Qual. 21, 341–344.
Terry, N., Zayed, A.D., de Souza, M.P., Tarun, A.S., 2000. Selenium in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 51, 401–432.
Thomine, S., Wang, R., Ward, J.M., Crawford, N.M., Schroeder, J.I., 2000. Cadmium and iron transport by members of a plant metal transporter family in Arabidopsis with homology to Nramp genes. Proc. Natl. Acad. Sci. USA 97, 4991–4996.
United States Protection Agency (USPA), 2000. Introduction to Phytoremediation. EPA 600/R-99/107. U.S. Environmental Protection Agency, Office of Research and Development, Cincinnati, OH.
Van der Zaal, B.J., Neuteboom, L.W., Pinas, J.E., Chardonnens, A.N., Schat, H., 1999. Overexpres-sion of a novel Arabidopsis gene related to putative zinc-transporter genes from animals can lead to enhanced zinc resistance and accumulation. Plant Physiol. 119, 1047–1055.
Van Huysen, T., Abdel Ghany, S., Hale, K.L., Le Duc, D., Terry, N., Pilon Smits, E.A.H., 2003. Overexpression of cystathionine synthase enhances selenium volatilization in Brassica juncea. Planta 218, 71–78.
Vandenhove, H., van Hees, M., van Winkel, S., 2001. Feasibility of phytoextraction to clean up low-level uranium-contaminated soil. Int. J. Phytoremediation 3, 301–320.
Vatamaniuk, O.K., Mari, S., Lu, Y.P., Rea, Y.A., 1999. AtPCS1, a phytochelatin synthase from Arabidopsis: Isolation and in vitro reconstruction. Proc. Natl. Acad. Sci. USA 96, 7110–7115.

146 Soil Remediation and Plants
Verret, F., Gravot, A., Auroy, P., Leonhardt, N., David, P., Nussaume, L., Vavasseur, A., Richaud, P., 2004. Overexpression of AtHMA4 enhances root-to-shoot translocation of zinc and cadmium and plant heavy metal tolerance. FEBS Lett. 576, 306–312.
Verret, F., Gravot, A., Auroy, P., Preveral, S., Forestier, C., Vavasseur, A., Richaud, P., 2005. Heavy metal transport by AtMHA4 involves N-terminal degenerated metal binding domain and the C-terminal HIS (II) stretch. FEBS Lett. 579, 1515–1522.
Vinita, H., 2007. Phytoremediation of toxic metals from soil and waste water. J. Environ. Biol. 28 (2), 367–376.
Wang, H., Kimberley, M.O., Schlegelmilch, M., 2001. Biosolids derived nitrogen mineralization and transformation in forest soils. J. Environ. Qual. 32, 1851–1856.
Willems, G., Gode, C., Verbrugger, N., Saumitou-laprade, P., 2007. Quantitative trait loci mapping zinc tolerance in the metallophyte Arabidopsis halleri spp. halleri. Genetics 176, 659–674.
Zayed, A., Pilon Smits, E., DeSouza, M., Lin, Z.Q., Terry, N., 2000. Remediation of selenium pol-luted soils and waters by phytovolatilization. In: Terry, N., Bañuelos, G. (Eds.), Phytoremedia-tion of Contaminated Soil and Water. Lewis, Boca Raton, pp. 61–83.
Zhu, Y.L., Pilon-Smits, E.A.H., Jouanin, L., Terry, N., 1999a. Overexpression of gluthathione syn-thetase in Brassica juncea enhances cadmium tolerance and accumulation. Plant Physiol. 119, 73–79.
Zhu, Y., Pilon Smits, E.A.H., Tarun, A., Weber, S.U., Jouanin, L., Terry, N., 1999b. Cadmium tol-erance and accumulation in Indian mustard is enhanced by overexpressing glutamyl cysteine synthetase. Plant Physiol. 121, 1169–1177.
Zhu, Y.L., Zayed, A.M., Quian, J.H., de Souza, M., Terry, N., 1999c. Phytoaccumulation of trace elements by wetland plants: II. Water hyacinth. J. Environ. Qual. 28, 339–344.