skeletal muscle - chnbaph.files.wordpress.com · skeletal muscle muscle tissue is one ... similar...
TRANSCRIPT

Skeletal Muscle
Muscle tissue is one of the four primary tissue types. Muscle tissue consists chiefly of muscle cells that are
highly specialized for contraction. Our bodies contain three types of muscle tissue:
1. Skeletal muscle,
2. Cardiac muscle, and
3. Smooth muscle.
Without these muscle tissues, nothing in the body would move, and the body itself could not move.
Skeletal muscle tissue moves the body by pulling on our bones, making it possible for us to walk,
dance, bite an apple, or play.
Cardiac muscle tissue pushes blood through the cardiovascular system.
Smooth muscle tissue pushes fluids and solids along the digestive tract and regulates the diameters
of small arteries, among other functions.
Anatomy of skeletal muscle:
Each and every individual skeletal muscle primarily is constituted by numerous muscle fibers or cells which
are the structural units. Similar to all the other muscles, skeletal muscle also contains connective tissues,
blood vessels and nerve supply.
1. Connective tissues:
Connective tissues functions to wrap, protect, support and strengthen the skeletal muscle tissue.
Subcutaneous layer separates the muscle tissue from skin. It is composed of adipose tissue and areolar
connective connective tissue and provides entry as well as exit for nerves, lymphatic vessels and blood
vessels. Based on location, connective tissue is devided as follows:
i. Epimysium,
ii. Perimysium, and
iii. Endomysium.
The epimysium is a dense layer of collagen fibers that surrounds the entire muscle. It separates the muscle
from nearby tissues and organs. It is connected to the deep fascia, a dense connective tissue layer.
The perimysium divides the skeletal muscle into a series of compartments. Each compartment contains a
bundle of muscle fibers called a fascicle (fasciculus, a bundle). Each fascicle inturn comprising 10-100
muscle fibers. Perimysium is a dense, irregular connective tissue that surrounds or bounds each and every
fascicle.
Tendon:
Within a fascicle, the delicate connective tissue of the endomysium surrounds the individual skeletal
muscle cells, called muscle fibers, and loosely interconnects adjacent muscle fibers.

The collagen fibers of the perimysium and endomysium are interwoven and blend into one another. At each
end of the muscle, the collagen fibers of the epimysium, perimysium, and endomysium come together to form
either a bundle known as a tendon, or a broad sheet called an aponeurosis. Tendons and aponeuroses usually
attach skeletal muscles to bones. Where they contact the bone, the collagen fibers extend into the bone matrix,
providing a firm attachment. As a result, any contraction of the muscle pulls on the attached bone.
Blood Vessels and Nerves
Muscle contraction requires
tremendous quantities of energy. An
extensive vascular network delivers
the necessary oxygen and nutrients
and carries away the metabolic
wastes generated by active skeletal
muscles.
Skeletal muscles contract only when
the central nervous system stimulates
them. Axons, nerve fibers extending
from the cell, penetrate the
epimysium, branch through the
perimysium, and enter the
endomysium to innervate individual
muscle fibers. Skeletal muscles are
often called voluntary muscles,
because we have voluntary control
over their contractions. We may also
control many skeletal muscles at a
subconscious level. For example,
skeletal muscles involved with
breathing, such as the diaphragm,
usually work outside conscious
awareness.
Muscle fibre
Muscle fibre is the structural and
functional unit of the skeletal muscle. These muscle fibers are long, elongated cylindrical structures with

many nuclei. Multiple nuclei are present because the muscle fiber is formed by the fusion of hundred or
more myoblasts (mesodermal cells) during the stages of embryonic development. This is reffered to as
syncytium which means fused cells. After the fusion of myoblasts, the capacity of the muscle fibers to
undergo mitotic cell division is lost. Therefore, the number of muscle fibers present at the time of birth
remains constant throughout the lifetime.
Some myoblasts do not fuse with developing muscle fibers. These unfused cells remain in adult skeletal
muscle tissue as the myosatellite cells. After an injury, myosatellite cells may enlarge, divide, and fuse with
damaged muscle fibers, thereby assisting in the repair of the tissue.
The muscle fibers lie parallel to one another with distinctive band-like appearance having alternate dark and
light striations.they are very long cells with length ranging from 10-30 cm and width ranging from 10-
100µm.
The Sarcolemma and Transverse Tubules
The sarcolemma or plasma membrane of a muscle fiber, surrounds the sarcoplasm, or cytoplasm of the
muscle fiber. Like other plasma membranes, the sarcolemma has a characteristic transmembrane potential
due to the unequal distribution of positive and negative charges across the membrane. In a skeletal muscle
fiber, a sudden change in the transmembrane potential is the first step that leads to a contraction.
Even though a skeletal muscle fiber is very large, all regions of the cell must contract at the same time. For
this reason, the signal to contract must be distributed quickly throughout the interior of the cell. This signal
is conducted through the transverse tubules.
Transverse tubules, or T tubules, are narrow tubes that are continuous with the sarcolemma and extend
deep into the sarcoplasm. They are filled with extracellular fluid and have the same general properties as the
sarcolemma, so electrical impulses conducted by the sarcolemma travel along the T tubules into the cell
interior. These impulses, called action potentials, trigger muscle fiber contraction.
Myofibrils
Inside each muscle fiber are hundreds to thousands of cylindrical structures called myofibrils, which can
actively shorten and are responsible for skeletal muscle fiber contraction. A myofibril is 1–2 µm in diameter
and as long as the entire cell. Branches of the T tubules encircle each myofibril.
Myofibrils consist of bundles of protein filaments called myofilaments. Myofibrils contain two types of
myofilaments: thin filaments composed primarily of actin, and thick filaments composed primarily of
myosin. At each end of the skeletal muscle fiber, the myofibrils are anchored to the inner surface of the
sarcolemma. In turn, the outer surface of the sarcolemma is attached to collagen fibers of the tendon or
aponeurosis of the skeletal muscle. As a result, when the myofibrils contract, the entire cell shortens and
pulls on the tendon.
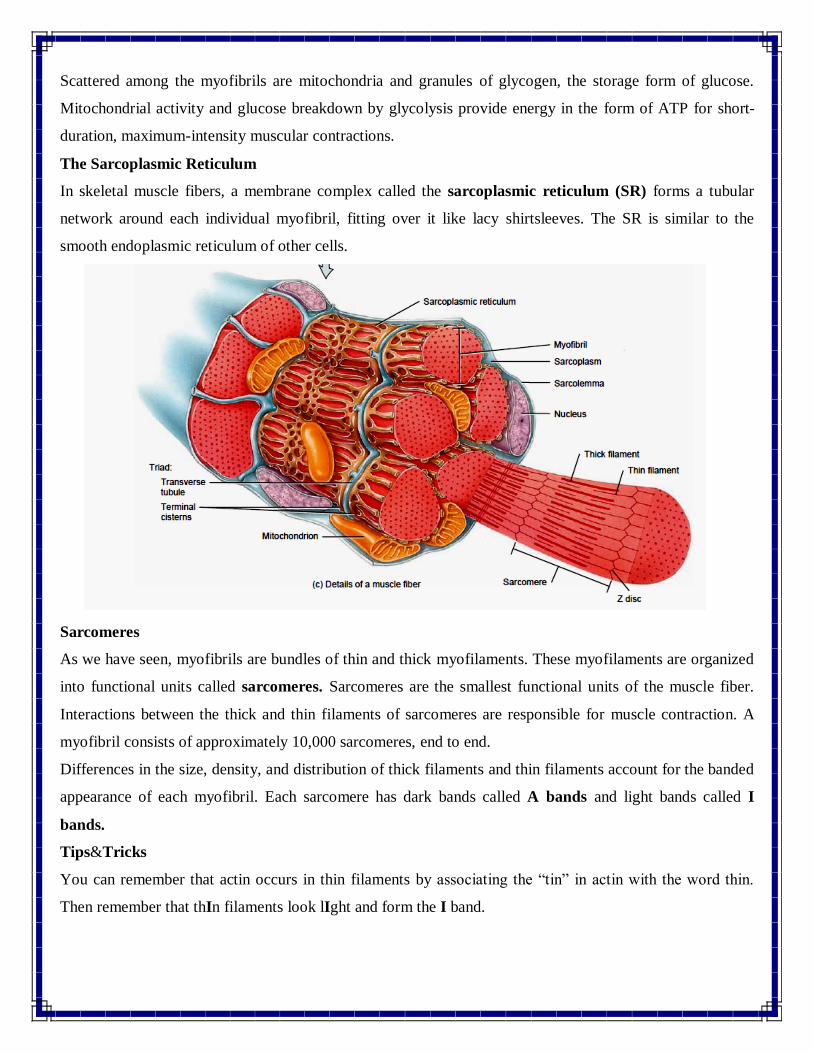
Scattered among the myofibrils are mitochondria and granules of glycogen, the storage form of glucose.
Mitochondrial activity and glucose breakdown by glycolysis provide energy in the form of ATP for short-
duration, maximum-intensity muscular contractions.
The Sarcoplasmic Reticulum
In skeletal muscle fibers, a membrane complex called the sarcoplasmic reticulum (SR) forms a tubular
network around each individual myofibril, fitting over it like lacy shirtsleeves. The SR is similar to the
smooth endoplasmic reticulum of other cells.
Sarcomeres
As we have seen, myofibrils are bundles of thin and thick myofilaments. These myofilaments are organized
into functional units called sarcomeres. Sarcomeres are the smallest functional units of the muscle fiber.
Interactions between the thick and thin filaments of sarcomeres are responsible for muscle contraction. A
myofibril consists of approximately 10,000 sarcomeres, end to end.
Differences in the size, density, and distribution of thick filaments and thin filaments account for the banded
appearance of each myofibril. Each sarcomere has dark bands called A bands and light bands called I
bands.
Tips&Tricks
You can remember that actin occurs in thin filaments by associating the “tin” in actin with the word thin.
Then remember that thIn filaments look lIght and form the I band.

Similarly, the A bands are dArk. The A Band The thick filaments are at the center of each sarcomere, in the
A band. The A band is about as long as a typical thick filament. The A band also includes portions of thin
filaments and contains these three subdivisions:
1. The M Line. Proteins of the M line connect the central portion of each thick filament to neighboring thick
filaments. (The M is for middle.) These dark-staining proteins help stabilize the positions of the thick
filaments.
2. The H band. In a resting sarcomere, the H band, or H zone, is a lighter region on either side of the M line.
The H band contains thick filaments, but no thin filaments.
3. The Zone of Overlap. The zone of overlap is a dark region where thin filaments are located between the
thick filaments. Here three thick filaments surround each thin filament, and six thin filaments surround each
thick filament.
Two T tubules encircle each sarcomere, and the triads containing them are located in the zones of overlap, at
the edges of the A band. As a result, calcium ions released by the SR enter the regions where thick and thin
filaments can interact.
Composition of myofibrils:
Myofibrils are made up of 3 types of proteins. They as follows:
1. Contractile proteins
2. Regulatory proteins
3. Structural proteins
1. Contractile proteins: These proteins are responsible for the generation of force of contraction. They are
of two types:
a. Myosin and
b. Actin
Myosin molecule is a motor protein i.e., it helps in push and pull function of different cellular components
so as to attain movement by converting chemical energy (ATP) to mechanical energy. There are about 200-
300 myosin molecules in every thick filament. Each myosin molecule consists of a rod like tail and two
globular heads or myosin heads. The myosin heads contain actin binding sites, ATP binding sites and
ATPase enzyme.
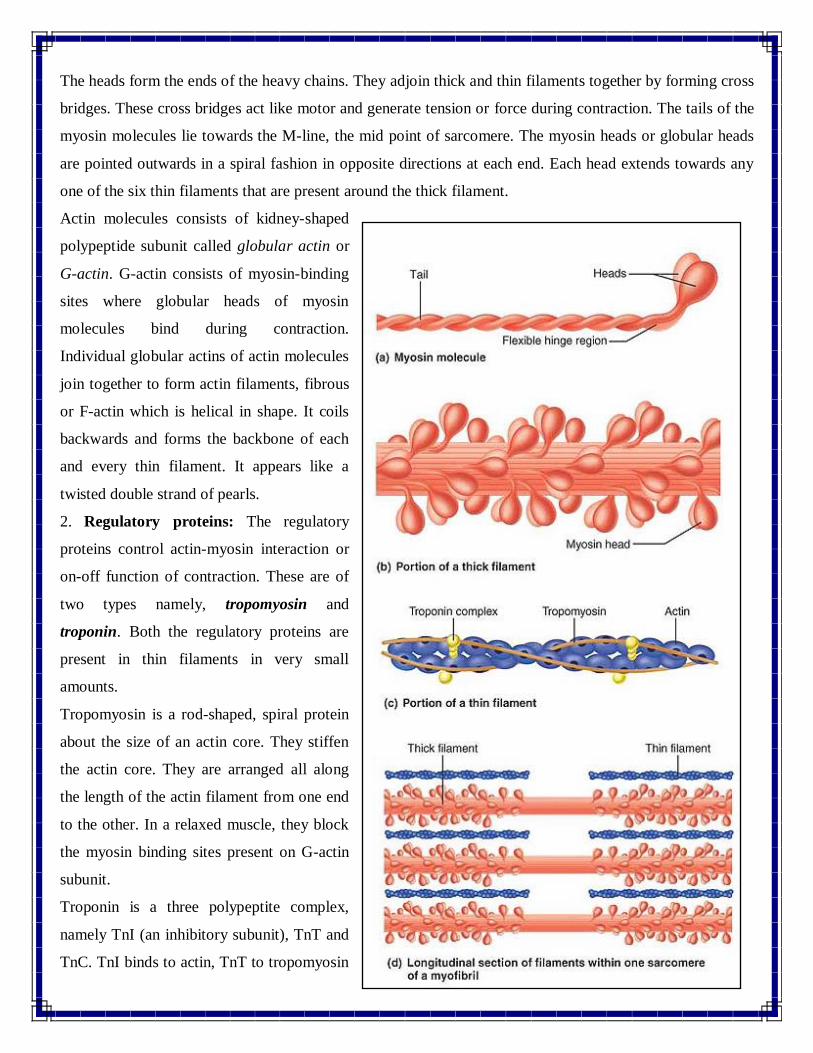
The heads form the ends of the heavy chains. They adjoin thick and thin filaments together by forming cross
bridges. These cross bridges act like motor and generate tension or force during contraction. The tails of the
myosin molecules lie towards the M-line, the mid point of sarcomere. The myosin heads or globular heads
are pointed outwards in a spiral fashion in opposite directions at each end. Each head extends towards any
one of the six thin filaments that are present around the thick filament.
Actin molecules consists of kidney-shaped
polypeptide subunit called globular actin or
G-actin. G-actin consists of myosin-binding
sites where globular heads of myosin
molecules bind during contraction.
Individual globular actins of actin molecules
join together to form actin filaments, fibrous
or F-actin which is helical in shape. It coils
backwards and forms the backbone of each
and every thin filament. It appears like a
twisted double strand of pearls.
2. Regulatory proteins: The regulatory
proteins control actin-myosin interaction or
on-off function of contraction. These are of
two types namely, tropomyosin and
troponin. Both the regulatory proteins are
present in thin filaments in very small
amounts.
Tropomyosin is a rod-shaped, spiral protein
about the size of an actin core. They stiffen
the actin core. They are arranged all along
the length of the actin filament from one end
to the other. In a relaxed muscle, they block
the myosin binding sites present on G-actin
subunit.
Troponin is a three polypeptite complex,
namely TnI (an inhibitory subunit), TnT and
TnC. TnI binds to actin, TnT to tropomyosin

and TnC to calcium ions. The major function of troponin is to place tropomyosin strands properly.
3. Structural proteins: There are about 12 structural proteins in the muscle. They help in maintaining
accurate alignment, stability, extensibility and elasticity of myofibrils.
After actin and myosin, the most abundantly present protein in the skeletal muscle is the gaint protein, titin
which was previously known as elastic protein. It has a very huge structure, is about 50 times bigger than a
protein of average size. It has a molecular weight of about 3million Daltons. It extends from Z-disc to M-
line running through the thick filament, about 1-1.2µm in distance spanning half of the sarcomere in a
relaxed muscle. It holds and stabilizes the thick filament in its place. Therefore, it maintains the organization
of the A-band. A part of the protein titin that extends from Z-line to the starting portion of thick filament
which spans I-band is elastic in nature. This helps the muscle to resume its actual shape after being stretched
or contracted excessively, as it unfolds when extended and coils when relaxed. Thus, it prevents sarcomeres
from being pulled apart.
Myomesin is a structural protein present in the M-line. It binds to titin and help in adjoining successive
thick filaments together.
Nebulin is present in thin filament all along its length. It is a very long nonelastic protein that assists in the
anchorage of thin filaments with the Z-lines. It also helps in controlling the length of thin filaments during
development.
Dystrophin is a cytoskeleton protein that binds thin filaments of sarcomere to sarcolemma via integral
membrane proteins. They inturn bind with the proteins present in the extracellular matrix of connective
tissue surrounding the muscle fibers. They help in the transfer of tension produced by sarcolemma to the
tendons.
Physiology of contraction
The physiology of contraction of skeletal muscle is well explained by sliding filament theory proposed by
Hugh Huxley in the year 1954. According to this theory, contraction is due to the sliding of thin filaments
over the thick ones such that actin and myosin filaments overlap to a greater extent.
When a muscle is relaxed, the overlapping of actin and myosin filaments in very slight. However, when the
stimulation occurs with the generation of nerve impulse, the myosin crossbridges which are the force-
producing areas attach to the actin filament at the myosin binding sites and thus initiates the process of
sliding. Many times the myosin crossbridges attach and detach from the thin filaments producing tension
and pushing them inwards such that they meet the centre of the sarcomere. They may even move very closer
and their ends may overlap. As this process occurs continuously, it progresses to the complete muscle fibre
leading to shortening of the muscle. The distance between the adjacent Z-discs is decreased as they are

pulled inwards towards the thick filaments. The length of the I-band shortens, H-line disappears and the
distance between A-bands is shortened, but the lengths of the thick and thin filaments remain unaltered.
Series of events taking place during contraction:
For a contraction to begin, it mainly needs to be stimulated by a nerve ending or axon that transmits action
potential or electrical impulse. The impulse results in an increased intracellular level of Ca2+ ion levels
which finally augments contraction. The events taking place during contraction are as follows:
1. The contraction cycle: the contraction cycle begins with the release of Ca2+ ions from the sarcoplasmic
reticulum into the cytosol. Ca2+ions binds to troponin and results in the removal of troponin- tropomyosin
complexes from the myosin binding sites of actin. Thus the contraction cycle begins which consists of the
following four steps:
a. Myosin or globular head consists of ATP binding site and ATPase. The enzymes ATPase bring about
hydrolysis of ATP to ADP and a phosphate group, thereby activating the myosin head.
b. Activated myosin heads bind to the myosin-binding site of actin by releasing the phosphate group, thus
forming crossbridges.
c. Once the formation of crossbridges is completed, power stroke takes place. The cross bridge rotates and
releases ADP and opens the site. The crossbridge while rotating inwards towards the sarcomere
produces a force which results in the sliding of thin filament over thick filament towards the M-line.
d. At the completion of power stroke, ATP binds to the ATP binding site on myosin head, due to which
myosin gets detached from the actin filament.
The contraction cycle continues as long as ATP and Ca2+ions are highly available and the newly bound ATP
is hydrolysed. One single crossbridge out of 600 present in one thick filament attach and detach for 5
times/second. At a time, few myosin heads attach to actin while few detach and get ready to bind again.
As the cells of the muscle fibre shorten, they pull the connective tissue sheaths i.e., epimysium, perimysium
and endomysium as well as tendons. The tendons inturn pull the bones to which they are attached. This
results in the movement of stimulated bone and associated structures.
2. 2. Excitation-contraction coupling: Excitation-contraction coupling is defined is defined as the series of
events taking place that result in the propagation of action potential along the length of sarcolemma which
results in the sliding of thick and thin myofilaments.
The action potential terminates well before the signs of contraction appear. Excitation-contraction coupling
occurs in the latent period present between the initiation of action potential and shortening of muscle fibers
(muscle activity). It takes place in the following order.
a) The muscle action potential propagates along the sarcolemma to the transverse T-tubules.
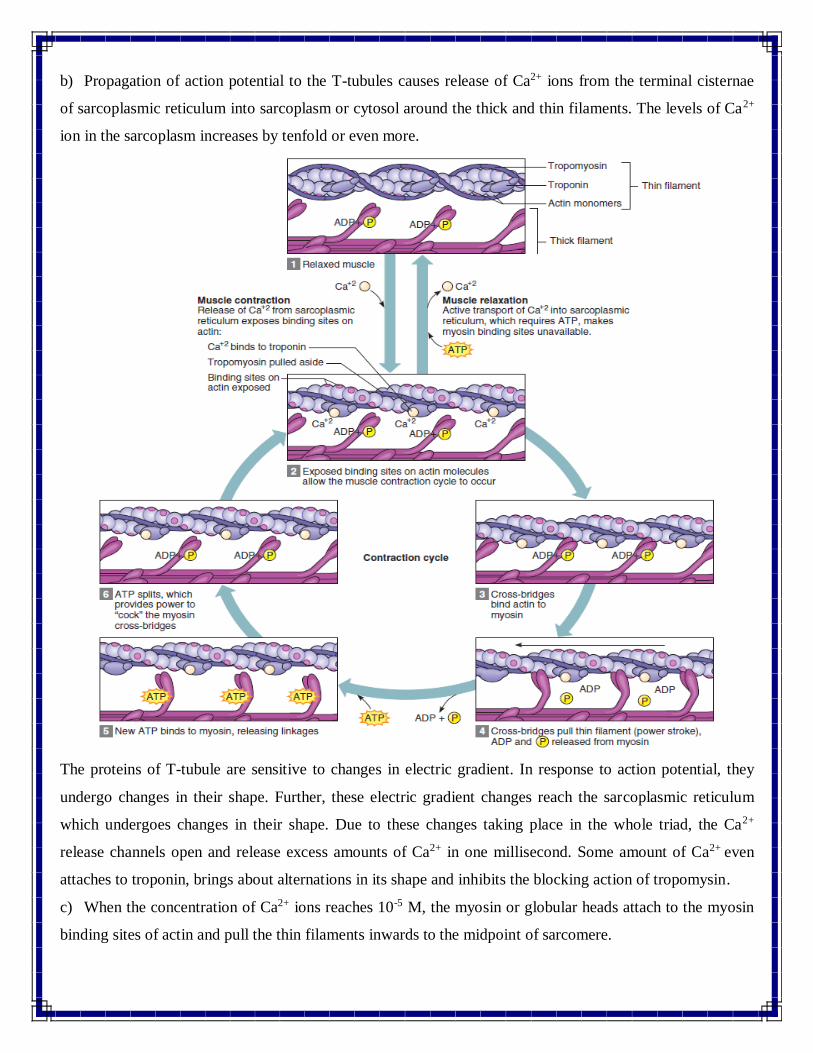
b) Propagation of action potential to the T-tubules causes release of Ca2+ ions from the terminal cisternae
of sarcoplasmic reticulum into sarcoplasm or cytosol around the thick and thin filaments. The levels of Ca2+
ion in the sarcoplasm increases by tenfold or even more.
The proteins of T-tubule are sensitive to changes in electric gradient. In response to action potential, they
undergo changes in their shape. Further, these electric gradient changes reach the sarcoplasmic reticulum
which undergoes changes in their shape. Due to these changes taking place in the whole triad, the Ca2+
release channels open and release excess amounts of Ca2+ in one millisecond. Some amount of Ca2+ even
attaches to troponin, brings about alternations in its shape and inhibits the blocking action of tropomysin.
c) When the concentration of Ca2+ ions reaches 10-5 M, the myosin or globular heads attach to the myosin
binding sites of actin and pull the thin filaments inwards to the midpoint of sarcomere.

As the concentration of Ca2+ increases in the sarcoplasm, the muscle contraction begins and immediately
ceases when the Ca2+ concentration falls down. In a relaxed muscle, the concentration of Ca2+ in sarcoplasm
is very low i.e., about 0.1µm/L. Ca2+ signals generally last for a very brief period of about 30 milliseconds
after the cessation of action potential. The events reoccur when another nerve impulse reaches the
neuromuscular junction.
The sarcoplasmic reticulum contains Ca2+ active
transport pumps. These pumps use ATP to bring
back Ca2+ from the cytosol to the sarcoplasmic
reticulum. The flow of Ca2+ ions from
sarcoplasmic reticulum to sarcoplasm is very
rapid, while vice-versa is slower. After the
cessation of last action potential, the closure of
Ca2+ release channels occurs. This rapidly
decreases calcium ion levels in the cytosol. The
calcium binding protein calsequestin present in
sarcoplasmic reticulum binds to Ca2+ resulting in
the storage of Ca2+ in the sarcoplasmic reticulum.
Therefore, in a relaxed muscle, Ca2+ levels are
10,000times higher in sarcoplasmic reticulum
than in sarcoplasm. Due to the drop in Ca2+ levels
in cytosol, the troponin-tropomysin complexes
cover the myosin binding sites on actin and
release muscle fibers thereby ceasing their
contraction.
3. Neuromuscular junction and generation of muscle action potential: the nerve cells associated with
stimulating the skeletal muscle belong to somatic nervous system. Every single somatic motor nerve cell
consists of an axon that runs from brain or spinal cord to a bundle of muscle fibers of skeletal muscle tissue.
The axon of every motor nerve cell divides profusely into a cluster of synaptic end bulbs at the
neuromuscular junction (NMJ) or synapse. NMJ is defined as an area present between the nerve cell and
target cell i.e., muscle fibre. At almost all the synapses there lies a small gap called synaptic cleft that
separates two adjacent cells from each other. Synaptic cleft consists of a gel-like extracellular substance
which contains high amount of glycoprotein. Within each ending of the axonal terminal numerous small
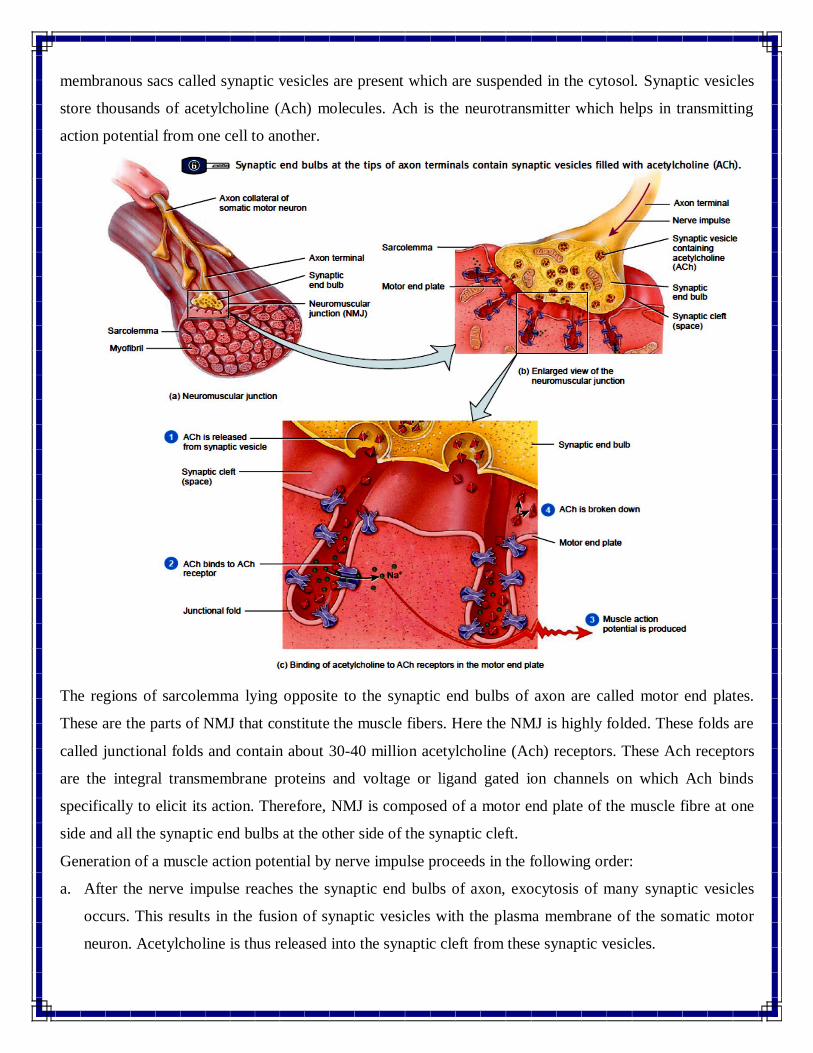
membranous sacs called synaptic vesicles are present which are suspended in the cytosol. Synaptic vesicles
store thousands of acetylcholine (Ach) molecules. Ach is the neurotransmitter which helps in transmitting
action potential from one cell to another.
The regions of sarcolemma lying opposite to the synaptic end bulbs of axon are called motor end plates.
These are the parts of NMJ that constitute the muscle fibers. Here the NMJ is highly folded. These folds are
called junctional folds and contain about 30-40 million acetylcholine (Ach) receptors. These Ach receptors
are the integral transmembrane proteins and voltage or ligand gated ion channels on which Ach binds
specifically to elicit its action. Therefore, NMJ is composed of a motor end plate of the muscle fibre at one
side and all the synaptic end bulbs at the other side of the synaptic cleft.
Generation of a muscle action potential by nerve impulse proceeds in the following order:
a. After the nerve impulse reaches the synaptic end bulbs of axon, exocytosis of many synaptic vesicles
occurs. This results in the fusion of synaptic vesicles with the plasma membrane of the somatic motor
neuron. Acetylcholine is thus released into the synaptic cleft from these synaptic vesicles.

b. Two molecules of Ach bind to an Ach receptor present at the motor end plate. This results in the
opening of ligand gated ion channel present on the Ach receptor. The opening of ion channel allows
passage or flow of small cations like Na+ and K+. Basically, the influx of Na+ ions and efflux K+ ions
takes place.
c. Influx of Na+ ion causes slight alteration in the membrane potential or electrochemical gradient. The
inner side of the membrane becomes more positively charged. This is called as depolarization. It
augments the action potential which spreads from the NMJ along with sarcolemma to T-tubules. This
results in the release of Ca2+ from the sarcoplasmic reticulum into the sarcoplasm and eventually
contraction of the muscle fibre occurs.
The action potential is due to the following series of electrical changes taking place along the length of
sarcolemma.
i. Spread of depolarization to the membrane areas adjacent to depolarized end plate via local
currents. This results in the opening of voltage-gated Na+ ions occurs causing potential difference
and initiation of action potential.
ii. The action potential propagates to the regions near sarcolemma and therefore opening of ligand
or voltage gated ion-channels takes place in those areas. This further enhances the Na+ influx,
causing the development of an electrochemical gradient.
iii. Finally, repolarization occurs subsequently after depolarization due to the closure of voltage-
gated Na+ channels and opening of ligand-gated K+ channels. This leads to the efflux (outflow) of
K+ ions from the muscle fibre to extracellular fluid. The muscle fibre stays in refractory phase
during repolarization as the cell cannot be stimulated again till the repolarization is completed.
d. The activity of Ach terminates within a very brief period. This is because the enzyme
acetylcholinesterase bound to the collagen fibers of extracellular membrane of synaptic cleft rapidly
breaks Ach to acetyl and choline molecules.
If another nerve impulse is generated and it releases more amount of Ach, then steps (b) and (c) occur
again. Inhibition of generation of action potential from the motor nerve cell inhibits Ach release. Ach is
broken down by acetylcholinesterase, thus resulting in the inhibition of action potential and closure of
sarcoplasmic reticulum leading to cessation of Ca2+ release.
The NMJ lies in the centre of the skeletal muscle fibre. Thus, the generated action potential of the
muscle is transmitted towards both the ends of muscle fibre resulting in simultaneous contraction of all
its areas.

Physiological properties of skeletal muscles
The following five properties exhibited by the skeletal muscles are essential for their effective
functioning:
1. Excitability and contractility
2. Refractory period
3. Tonicity
4. Conductivity
5. Extensibility.
1. Excitability and contractility: This property is the characteristic feature of all the living tissues. It is very
well developed in muscle and nerve tissue. The capacity of a tissue to respond to stimuli is known as
excitability and contractility. The stimuli may be of electrical, chemical (acids, bases), mechanical
(pressure, pinching) or thermal (temparature fluctuations) origin.
Generally, the nerve impulses produced in the human body act as stimulus and result in the contraction
of muscles. These nerve impulses are of electrical type. Therefore, in the laboratory electrical current,
either galvanic or faradic is used to generate stimulus.
Upon the influence of stimulus, the muscle gets excited and contracts. The contraction is immediately
followed by relaxation. Contraction of a muscle
occurs either during the make or break of
current. On application of a single induction
shock, a single twitch or contraction is produced.
This when recorded on a moving drum in three
phases or periods. They are as follows:
a. Latent period or phase: It is defined as
the interval between the application of
stimulus and initiation point of
contraction. It is generally the time taken
by the nerve impulse to propagate from the
point of stimulation to neuromuscular junction,
finally to sarcolemma of the muscle and
beginning of contraction. The length of the
muscle gets shortened in this period.
b. Contraction period or phase: The time
interval between the beginning of contraction
to the maximum contraction (M) is considered
as the contraction period.
Figure: Single Muscle Curve
A1= Point of stimulation
A2= Initiation or beginning of contraction
A1- A2= Latent period or phase
X= Contraction period or phase
Y= Relaxation period or phase
M= Maximum contraction

c. Relaxation period or phase: The time interval from the maximum contraction or summit to the
original level is the relaxation period. The muscle resumes its original length during this phase.
Factors affecting excitability and contractility
a) Strength or intensity of stimuli: a minimum strength is required by the stimulus to produce
response. This is referred to as minimal or threshold stimulus. Stimulation lower than the
threshold stimuli i.e., subminimal or subliminal stimuli does not produce any response.
Upon application of minimal or threshold stimuli, only a few motor units with low threshold are
activated and contraction occurs. With an increase in the intensity of stimulus, the degree or
force of contraction also increases progressively due to the activation of number of motor units.
This process continues until the maximum contraction occurs by the application of maximum
stimulus from the activation of all the muscle fibers or motor units. This is referred to as quantal
or multifibre stimulation. The strength of the stimuli lying in between minimal and maximal
stimuli is submaximal stimuli and the intensity of the stimuli which is more than the maximal
stimuli is supramaximal stimuli.
b) All or none law: stimulation of one muscle fibre or motor unit by the application of minimal or
threshold stimuli results in maximum contraction. Any increase in the intensity of stimulus does
not raise the force of contraction any further. This is referred to as all or none law.
However, when the whole muscle comprising numerous muscle fibers or motor units is
stimulated, all or none law slightly varies. As the intensity of the stimulus is increased, the
degree or force of contraction increases until a state where no further increase is achieved. In
contrast, according to the modern theory, all or none law is not applicable to contractile fibers. It
is true only in the case of action potential.
c) Effect of application of two successive stimuli on muscle contraction
i. Application of two successive subminimal stimuli results in the contraction of muscle.
This is referred to as “latent addition of excitation” or “summation of subminimal
stimuli”.
ii. When the second stimulus is applied after
sufficient time of application of first
stimuli i.e., after relaxation of first
contraction, then two separate simple
muscle curves are produced. Due to the
beneficial effect of the first stimuli, the
second muscle curve produced is slightly
higher than the first muscle curve. The
increase in the height of the second muscle
curve may be attributed to increase in the
availability of Ca2+ ions or due to increase
in the temperature which causes a decrease
in the viscosity of sarcoplasm. This inturn
enables the muscle fibers to contract better.

iii. When the second stimulus is applied during the relaxation period of the first stimuli, then
the second contraction occurs well before the completion of relaxation phase. This results
in superimposition of second curve on the first curve. This phenomenon is termed as
superposition.
iv. When the second stimulus is applied
during the contraction phase of the
first curve, then a curve similar to
simple muscle curve is formed. This
is called as summation of effects or
contraction.
v. When the second stimulus falls with
the latent phase but after the
completion of refractory phase of the
first curve, it gives a simple muscle
curve of greater height and longer
duration than the curve produced when
two stimuli are applied separately. This
effect is termed summation of stimuli
or wave summation.
Both the maximal and submaximal
stimuli show summation effects.
Greater heights of the muscle curves
are obtained by the application of maximal stimuli. This is due to variations in the
mechanism of contraction but not due to the increase in the activation of numerous
muscle fibers or motor units. The second contraction or twitch is initiated well before the
completion of the active phase of the first contraction. Thus, the duration of the active
state of first curve augments the response of the second stimulus. This effect does not
follow all or none law.
d. Effect of repletion of stimuli on muscle contraction
i. When a series of adequate stimuli of equal strength are applied successively by
maintaining a time interval such that the contraction resulting due to the application of
first stimulus is completed (1 second), then the muscle curves showing gradual increase
in the amplitude or force of contraction are obtained. They show a stair-like increase and
thus this effect is termed staircase phenomenon or treppe. This effect is mainly because
of slight increase in the temperature, rise in the concentration of H+ ions obtained from
previous contraction and increase in the Ca2+ ion availability which inturn results in an
increased excitability. This phenomenon can be described as beneficial effect.
ii. When the successive stimuli applied lie within the relaxation phase of the previous curve,
the muscle curve shows a series of wavy oscillations or vibrations. This phenomenon is
termed clonus or incomplete tetanus. It may be considered as summation of successive
stimuli.
iii. When the successive stimuli applied lie within the latent period of the previous curve, it
gives a clear steady line. This line shows an abrupt initial increase and then gradually

increase until maximum contraction occurs. This is termed tetanus. This is considered as
summation of stimuli. Complete fusion of the contractions occurs in tetanus and the
height of the titanic contraction curve is more than the height of the curve obtained by
single twitch or contraction.
The greater amplitude of tetanus contraction is due to increase in the amount of Ca2+
ions in sarcoplasm. The frequency of the stimulus per second necessary to produce
tetanus contraction depends upon the nature of the muscle. The muscle of a human being
requires 60/sec, frog’s muscle needs 40/sec, external eye muscle needs 350/sec and
gastrocnemius requires 100/sec.
iv. Loss of irritability, gradual decrease in the excitability of muscle and finally cessation in
response occurs when a muscle is stimulated repeatedly. This phenomenon is termed
fatigue. It is studied by using an instrument called Mosso’s ergograph.
Fatigue is defined as the inability of the muscle to work any further. In the muscle curve
obtained during fatigue. The first few contractions show rise in the amplitude in the
beginning because of the beneficial effect. Gradually the height of the fatigue curve
decreases until the contractions completely stop. This is because, the muscle completely
loose its excitability.
Prolongation of all the phases takes place in fatigue. The latent phase is lengthened and
the time interval for the completion of relaxation phase is very much prolonged. Due to
this, the muscle does not resume its original length. This shortening of the muscle
because of the time lag in the relaxation phase is termed contraction remainder. This is a
form of contracture (persistent contraction). It mainly occurs due to consumption and
depletion of sources that supply energy to the muscle, deposition of acid metabolites like
H+, lactic acid, ketone bodies etc., the end products of chemical reactions and reduced
synthesis of acetylcholine (Ach) like substances during prolonged exercise. The acidic
metabolites inactivate the enzymes that cause glycolysis. Therefore, fatigued muscle is
acidic in nature which is otherwise slightly alkaline in normal conditions. Oxygen helps
in the recovery of fatigued muscle by removing all these substances. This can be proved
while studying fatigue in muscle with and without circulation. In case of a muscle with
blood circulation, fatigue occurs very slowly and recovery occurs during rest. But in case
of a muscle without blood circulation, fatigue occurs at a faster rate and recovery is not
possible during rest.
When a muscle is stimulated directly, the seat or site of fatigue is present within the
muscle. But, when stimulation is produced via motor nerve, the site or seat of fatigue lies
in the neuromuscular junction while during physiological exercise it is present in the
CNS. This is termed central fatigue. The order of fatigability after a voluntary work is
generally in the synapse first, then in the neuromuscular junction and finally in the
muscle. However, the cause and the seat of fatigue is not clearly understood.
It may be due to physical or mental stress, a manifestation of a clinical illness, due to
local acidosis, accumulation of interstitial fluid or due to decreased amount of glycogen.
Sometimes, increase in the number of afferent impulses from proprioreceptors present in
the muscle may get transmitted to brain and cause a sensation of weakness i.e., fatigue.

e. Effect of temperature on muscle contraction: Room temperature (25oC) or moderately warm
temperature (38oC) increases both excitability as well as contractility. All the chemical processes
occur at a faster rate. The time interval of all the phases or periods of muscle curve is reduced.
In contrast, very low or cold temperature ranging from 5-10oC decreases excitability and
contractility. It prolongs the duration of all the phases of the muscle curves.
At a temperature of 45oC and above, coagulation of muscle proteins occurs leading to heat rigor
or rigidity. This process is irreversible i.e., the muscle do not recover on subsequent cooling.
Cooling to 0oC temperature leads to complete loss of excitability, but it can be reversed on
subsequent warming of the muscle.
f. Effect of load on muscle contraction: The weight applied after the muscle has just started to
contrast is referred to as after-load and the weight applied before the muscle begins to contract is
termed as load, pre-load, fore-load or free-load.
Load prolongs the latent phase but decreases the time interval of contraction as well as relaxation
period. Therefore, a loaded muscle works more efficiently because the load applied stretches the
muscles, increases the initial length and thereby increases the degree or force of contraction
(starling’s law).
g. Effect of ions and salts on muscle contraction: Calcium plays an important role in the
initiation of muscle contraction. Ca+ ions help in the development of tension in the muscle. It
also stimulates the activity of the enzymes ATPase. This helps in prompt association of actin and
myosin in the presence of ATP. Magnesium ion is necessary for the activation of phosphorylase
enzyme which is required for the transfer of phosphate during the contraction of muscle. Ca+ as
well as Mg+ ions to some extent contribute to the enzymatic action of myosin.
Sodium salts exert excitatory effect on the muscle cells, whereas potassium salts decrease
excitability and increase the rate of attainment of fatigue. The electrical phenomenon, excitability
and contractility, all depend upon the migration of potassium ions. Hydrogen ion shows similar
effect on the contraction of muscle as that shown by excess of potassium ions.
h. Effect of duration of stimulus: Duration of stimulus (current) also affects the muscle
contraction. A very strong stimulus existing for a very brief or short period of time may not be
effective and very weak stimuli though long standing does not show any effect. The more
stronger the current, the lesser is the duration required to produce contraction. However, certain
limits exist which can be measured with the help of two standards, namely Rheobase and
Chronaxie.
Rheobase is defined as the strength of minimum galvanic current required to excite the muscle
when it is allowed to flow indefinitely. The duration for which the current flow is called
utilization time. Chronaxie is defined as the shortest duration of time required by the stimuli,
which has twice the strength of rheobase current, to excite the muscle.
A tissue with less excitability has longer chronaxie, while a tissue with greater excitability has
shorter chronaxie. A neonate has chronaxie ten times longer than the adults. The chronaxie of
cold blooded animals is longer than warm blooded animals. A degenerating muscle exhibits
longer chronaxie. The skeletal muscle exhibits shorter chronaxie than any other muscle in the
body.
Stretching a muscle depletes chronaxie, thus increases excitability. Fatigue and cold temperature
prolong chronaxie, whereas adrenaline and warm temperature shorten it.

i. Rate of alteration of strength of stimuli: the stimulus which is built up very fast reaches its
peak strength and is more effective than the stimulus that builds up slowly. If the rate of
alteration of the strength or intensity of the stimulus is extremely prolonged, then it may be
ineffective.
2. Refractory period:
A brief time period for which the muscle remains stable or inexcitable to the second stimulus after the
application of the first stimulus is termed refractory period. It is very short, about 2 milliseconds
(0.002) in mammals and 5 milliseconds (0.005 sec) in frog.
Though a very strong stimulus is applied, the muscle cannot be excited during the first part of the
refractory period. This period is referred to as absolute refractory period. However, in the second part
of the refractory period, the muscle can be excited by applying very strong stimuli. This period is
termed relative refractory period. The absolute refractory period for skeletal muscles is very short,
hence they can be fatigued and tetanized. Hot temperature shortens, whereas cold temperature prolongs
the refractory period.
3. Tonicity:
The stage in which the skeletal muscles are present in light tension is termed tone. It is defined as a
reflex sustained and partial contraction. Tonicity is a reflex process whose process whose centers are
present in the spinal cord. A muscle with a tone does not undergo fatigue. This is because during tone
production, muscle fibers exist as batches, when one batch contracts the other relaxes. Thus, complete
muscle tissue is free from fatigue.
4. Conductivity:
After the stimulation of muscle, a wave of contraction begins to flow from the point of application of
stimulus in both the directions along the complete muscle. In warm blooded animals it is about 6-12
meters/sec and in frog it is 3-4 meters/sec.
5. Excitability:
The capacity of a muscle tissue to stretch or extend without being damaged is called extensibility. This
property enables the muscle to contract forcefully even if it is already stretched.
6. Elasticity:
The capacity of the muscle to resume its actual shape and length after contraction or extension is called
elasticity.
Skeletal muscle disorders
The disorders associated with the skeletal muscle tissue are termed myopathies where myo means
muscle and pathy means disease or illness.
Myasthenia Gravis: (Mys=muscle, aesthesis=sensation, gravis=serious)
It is a neurogenic autoimmune disorder occurring due to chronic and progressive destruction of
neuromuscular junction. The immune system produces antibodies which bind to acetylcholine receptors
at motor end plates and block or inhibit them. Therefore, the number of active Ach receptors gets
reduced, which results in the blockade of nerve impulse transmission to muscle fibers. This further result

in failure to trigger action potential of muscle and subsequently the contraction of muscles get
weakened. As the disease progresses, the inactivation of number of Ach receptors increases resulting in
greater muscle weakness, fatigue and finally loss of muscle function.
Initially, the disease affects the muscles of the eye leading to ptosis (drooping of the eyelids) or double
vision (diplopia), followed by weakness in the muscles of neck or throat leading to difficulty in chewing,
swallowing and speech. Finally, it affects the muscles of the limbs. Very rarely it may cause paralysis of
respiratory muscles which may lead to death of the individual. Myasthenia gravis is more commonly
seen in women than in men. Neostigmine and pyridostigmine (Acetylcholinesterase inhibitors) are used
as first line agents to treat myasthenia gravis. Thymectomy (removal of thymus by surgery) may also
serve to be helpful.
Fibromyalgia:
Fibromyalgia is mainly associated with the damage to fibrous connective tissue present in muscles,
tendons as well as ligaments. It is a painful, non-articular rheumatic disease. Generally, the age of onset
of this disease lies in between 25-50 years. Treatment procedure includes physical therapy, regular
exercise, gentle massage, and application of heat, medication for pain and a low-dose antidepressant so
as to improve sleep.
Denervation atrophy:
Denervation atrophy is associated with axonal degeneration followed by muscle atrophy. This mainly
occurs due to the loss of motor nerve supply to complete or a part of the muscle.
Muscular dystrophy:
Muscular dystrophy belongs to the class of myopathies. It is a group of genetically acquired disorders
wherein degeneration of muscle fibers takes place.
Crush syndrome:
It is also known as traumatic rhabdomyolysis or Bywater’s syndrome. It is a crushing injury of skeletal
muscle caused by sustained pressure on the trunk or limb muscles. It is believed that the breakdown
products of skeletal muscles i.e., myoglobin, potassium and phosphorous causes crush syndrome. When
the pressure is released, these breakdown products enter the circulation and may lead to renal failure.
Other symptoms include hypovolaemic shock and hyperkalemia (increased blood potassium levels). A
general complication associated with crush syndrome is infection by anaerobic microorganisms.
Examples: Gas gangrene occurs due to infection by Clostridium perfringens.