simultaneous fermentation of glucose and xylose to butanol ... · materials and methods growth...
TRANSCRIPT

Simultaneous Fermentation of Glucose and Xylose to Butanol byClostridium sp. Strain BOH3
Fengxue Xin, Yi-Rui Wu, Jianzhong He
Department of Civil and Environmental Engineering, National University of Singapore, Singapore, Republic of Singapore
Cellulose and hemicellulose constitute the major components in sustainable feedstocks which could be used as substrates forbiofuel generation. However, following hydrolysis to monomer sugars, the solventogenic Clostridium will preferentially con-sume glucose due to transcriptional repression of xylose utilization genes. This is one of the major barriers in optimizing ligno-cellulosic hydrolysates that produce butanol. Unlike studies on existing bacteria, this study demonstrates that newly reportedClostridium sp. strain BOH3 is capable of fermenting 60 g/liter of xylose to 14.9 g/liter butanol, which is similar to the 14.5 g/literbutanol produced from 60 g/liter of glucose. More importantly, strain BOH3 consumes glucose and xylose simultaneously,which is shown by its capability for generating 11.7 g/liter butanol from a horticultural waste cellulosic hydrolysate containing39.8 g/liter glucose and 20.5 g/liter xylose, as well as producing 11.9 g/liter butanol from another horticultural waste hemicellu-losic hydrolysate containing 58.3 g/liter xylose and 5.9 g/liter glucose. The high-xylose-utilization capability of strain BOH3 isattributed to its high xylose-isomerase (0.97 U/mg protein) and xylulokinase (1.16 U/mg protein) activities compared to the low-xylose-utilizing solventogenic strains, such as Clostridium sp. strain G117. Interestingly, strain BOH3 was also found to produceriboflavin at 110.5 mg/liter from xylose and 76.8 mg/liter from glucose during the fermentation process. In summary, Clostrid-ium sp. strain BOH3 is an attractive candidate for application in efficiently converting lignocellulosic hydrolysates to biofuelsand other value-added products, such as riboflavin.
Conversion of lignocellulosic biomass to biofuels (e.g., buta-nol) is usually limited by the inefficiency of the bacteria in the
utilization of pentoses, which constitute 20 to 60% of the sugars inthe hydrolysate of lignocellulose (39). So far, the biofuel-generat-ing microbes in common use are either unable to utilize pentosesat all (e.g., Saccharomyces cerevisiae) or consume hexoses first andthen pentoses with a low yield. Good examples are the solvento-genic Clostridium species (e.g., C. acetobutylicum, C. beijerinckii,and C. pasteurianum), which are among the few microorganismsable to ferment both pentose and hexose sugars (10). However,the solvents, including butanol, are still produced mainly fromglucose, while small amounts are produced from xylose, as ob-served in cultures of C. acetobutylicum ATCC 824 and C. beijer-inckii NCIMB 8052 when fermenting a mixture of glucose andxylose (14, 17, 38). Gu et al. have shown that C. acetobutylicumATCC 824 utilized 86% of glucose within 40 h, whereas only 6% ofxylose was consumed even after an elongated incubation time(14). This sequential utilization extends fermentation time andresults in incomplete substrate consumption (21, 33). The phe-nomenon of preferring glucose over xylose is referred to as carboncatabolite repression, in which microorganisms preferentially uti-lize a rapidly metabolizable carbon source and inhibit the expres-sion of some genes and enzyme activities related to the catabolismof nonpreferred carbon resources (21, 33). Unless glucose andxylose are both utilized efficiently, converting lignocellulosic bio-mass into biologically based products is unfavorable from an eco-nomic viewpoint (19, 20, 22), as yields would be limited. Hence, inorder to fully utilize the sugars in the hydrolysates of lignocellu-losic biomass, microbes capable of fermenting both glucose andxylose with a high butanol yield are highly desirable to improvethe efficiency of biofuel production systems.
In addition to butanol production, certain biofuel-generatingmicrobes, such as Clostridium, can produce other value-addedproducts (e.g., riboflavin, also known as vitamin B2), which adds
to the value of the butanol fermentation process. Actually, in ad-dition to producing butanol, Clostridium species are also one ofthe commercial microorganisms used to produce riboflavin on anindustrial scale, although they only generate a very small amountof riboflavin (�50 mg/liter) (16, 26, 27). Riboflavin, a yellow,water-soluble vitamin, is biosynthesized solely by plants andnumerous microorganisms but not by vertebrates. Thus, thisvitamin has to be supplied by foods and dietary supplements.Currently, industrial production has shifted completely fromchemical synthesis to microbial fermentation (32), which hasbecome one of the major fermentation products in the biotech-nology industry, with an annual market demand of several mil-lion kilograms (31). Therefore, the coproduction of butanoland riboflavin would make the fermentation process more ec-onomically feasible (28).
The aim of this study is to explore wild-type Clostridium sp.strain BOH’s capability to utilize the five-carbon sugar xylose. Afurther study then was conducted on both butanol and riboflavinproduction from fermenting a mixture of glucose and xylose andhydrolysates from horticultural wastes. Lastly, the activities of xy-lose-utilizing enzymes (xylose isomerase and xylulose kinase) instrain BOH3 were evaluated to determine the underlying mecha-nism of cofermentation of glucose and xylose.
Received 31 January 2014 Accepted 17 May 2014
Published ahead of print 23 May 2014
Editor: A. M. Spormann
Address correspondence to Jianzhong He, [email protected].
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AEM.00337-14
August 2014 Volume 80 Number 15 Applied and Environmental Microbiology p. 4771– 4778 aem.asm.org 4771
on May 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from
MATERIALS AND METHODSGrowth medium and culture conditions. Clostridium sp. strain BOH3and Clostridium sp. strain G117 were used in this study (6, 9). Batchcultures were grown at 35°C in defined mineral salts medium containing(per liter of distilled water) the following: KH2PO4, 0.75 g; K2HPO4, 0.75g; CH3COONH4, 2 g; and yeast extract, 5 g. In addition, 1 ml of traceelement solution (35), 1 ml of Na2SeO3-Na2WO4 solution (7), and 10 mgof resazurin were added to 1 liter of the medium. After the medium wasboiled and cooled to room temperature under N2, reductants Na2S, L-cys-teine, and DL-dithiothreitol were added to a final concentration of 0.2, 0.2,and 0.5 mM, respectively (15). Subsequently, 20 mM 2-(N-morpholino)ethanesulfonic acid (MES) was added to the medium to adjust its initialpH to 6.0. The medium then was dispensed to serum bottles, which weresealed with butyl stoppers, autoclaved for 20 min, and cooled to roomtemperature. Glucose, xylose, or a mixture of glucose and xylose wasamended to the medium described above before inoculation.
Cultures for inoculation were grown in 50 ml mineral salts mediumamended with glucose, xylose, or a glucose-xylose mixture at 35°C for�20 h (late exponential phase) unless otherwise stated. Inocula of 5 mlwere added to 45 ml of the reduced mineral salts medium in 160-ml serumbottles, which were incubated in a shaker at a rotary rate of 150 rpm at35°C. The pH was adjusted to 5.0 using 2 M NaOH after 24 h. Experimentswere carried out in duplicate.
Preparation of horticultural waste hydrolysates. Horticulturalwastes collected from a horticultural waste treatment plant (ecoWise So-lution Pte. Ltd.) were used as raw materials and were dried and milled toa small size (200 to 500 �m). Pretreatment of the horticultural waste wascarried out by using the organosolv method as described by Zhang et al.(40) and Geng et al. (12). Initially, the organic solvent ethanol was used to
dissolve hemicellulose and lignin to the solvent phase. The ethanol thenwas removed by vacuum evaporation, and the hemicellulosic fractioncarrying mainly xylose was further detoxified using activated charcoalpowder and concentrated to obtain a final sugar mixture containing 5.9g/liter of glucose and 58.3 g/liter of xylose (40). The solid residues from thepretreatment step contained mainly cellulose, which was hydrolyzed byenzymes of Celluclast 1.5L and Novozyme 188 to glucose and designatedcellulosic hydrolysate (12). Supplemental xylose (16.9 g/liter) was addedto the cellulosic hydrolysate to attain a 2:1 ratio of glucose (39.8 g/liter) toxylose (20.8 g/liter). Prior to filter sterilization, the pHs of the detoxifiedhemicellulosic hydrolysate (containing 5.9 g/liter of glucose and 58.3 g/li-ter of xylose) and cellulosic hydrolysate (containing 39.8 g/liter of glucoseand 20.8 g/liter of xylose) were adjusted to 6.0 by titrating with 10 MNaOH solution. This was followed by supplementing KH2PO4, K2HPO4,CH3COONH4, yeast extract, trace element solution, Na2SeO3-Na2WO4
solution, and reductants as described in the above section.Expression of genes related to xylose utilization as measured by
qPCR. Total RNA from cell pellets of Clostridium sp. strain BOH3 andG117 was extracted by using a modified TRIzol method and RNeasy mini-kit (Qiagen, Hilden, Germany) according to the supplier’s instructions.The remaining trace DNA was removed using an RNase-free DNase kit(Qiagen, Hilden, Germany). cDNA was synthesized in one step using aniScript One-Step reverse transcription-PCR (RT-PCR) kit (Bio-Rad Lab-oratories, Munich, Germany). A quantitative real-time PCR (qPCR) (ABI7500 Fast real-time PCR system; ABI, Foster City, CA) assay was per-formed by using QuantiTest SYBR green (Qiagen, GmBH, Germany) toquantify the transcription levels of the xylose isomerase gene. The xyloseisomerase gene-specific primers used for qPCR were designed based onthe xylose isomerase gene sequence of cultured BOH3 and G117, with
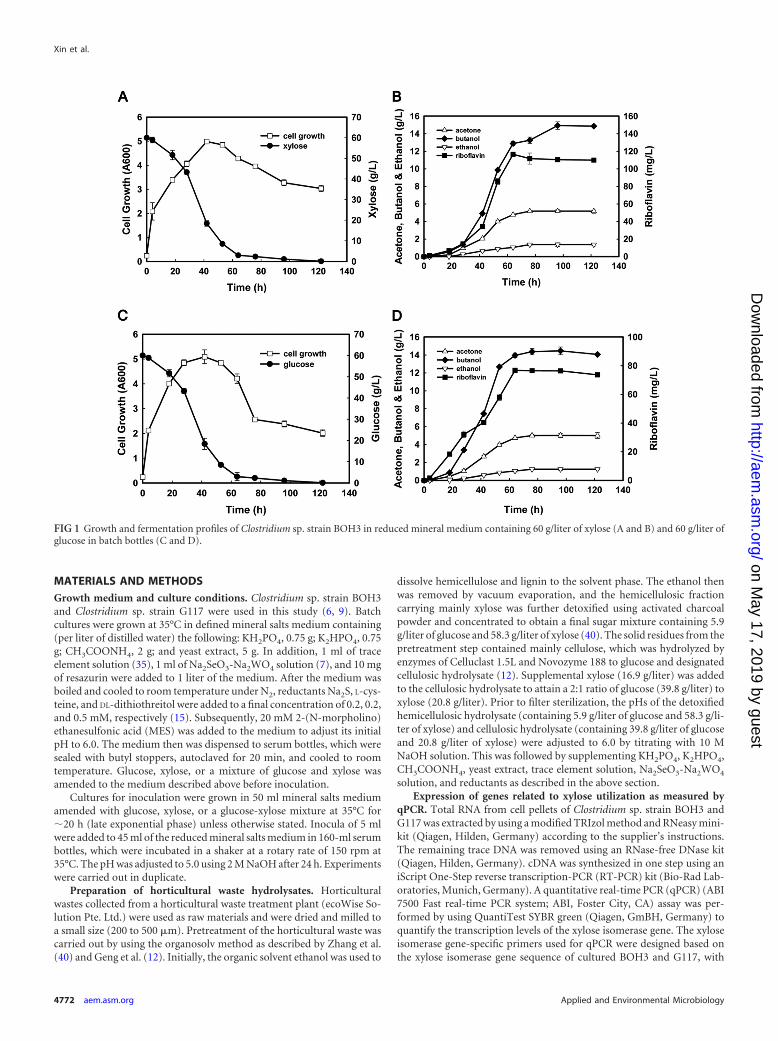
FIG 1 Growth and fermentation profiles of Clostridium sp. strain BOH3 in reduced mineral medium containing 60 g/liter of xylose (A and B) and 60 g/liter ofglucose in batch bottles (C and D).
Xin et al.
4772 aem.asm.org Applied and Environmental Microbiology
on May 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from

the forward primers 5=-ATGTTGCAGTTACAGAGGGAGA-3= (strainBOH3) and 5=-CTGTTTTACTAATCCAA GGTATGTTCA-3 (strainG117) and reverse primers 5=-TTTCATCTTGGCTTACCTT GTC-3=(strain BOH3) and 5=-ATCTAGCTCTTTTGTTATTTCA ATTGC-3=(strain G117). Primers for the housekeeping genes [(3R)-hydroxymyris-toyl-ACP dehydratase (fabZ) for strain BOH3 (23) and peptidase T (pepT)for strain G117 (36)] are the following: strain BOH3-forward, 5=-AAATAGAACCAGGGAAAAGAGCA-3=; strain BOH3-reverse, 5=-GCAACACCACCAAGTTGAGC-3=; strain G117-forward, 5=-TGATGGAG GCGAGGAAGGTG-3=; and strain G117-reverse, 5=-CATTGTATTCTTTGCAGACCCTGG-3=. The following real-time PCR program was used: 10 min at95°C, 45 cycles of 10 s at 95°C, and 30 s at 55°C, followed by a melting-point analysis (45 to 95°C) after the final PCR step to verify the specificityof amplified products. The real-time PCR data were analyzed with iQ5software, and relative quantification was done by using the ��CT method(25) and normalized to the abundance of housekeeping genes.
Xylose isomerase and xylulose kinase assays. Cell extracts obtainedfrom the batch cultures were used for enzymatic activity assays. Thechemicals used in the enzymatic activity assessment were obtained fromSigma-Aldrich (St. Louis, MO). Cultures at the exponential phase wereharvested by centrifugation at 10,000 � g at 4°C for 10 min. Cell pelletswere washed once with extraction buffer (100 mM Tris-HCl [pH 7.5], 20mM KCl, 20 mM MgCl2, 5 mM MnSO4, 0.1 mM EDTA, and 2 mM
DL-dithiothreitol). The resulting cell pellets were sonicated using an ultra-sonic homogenizer in an ice water bath for 2 min followed by 2-mincooling intervals, which were repeated three times. Cell debris was re-moved by ultracentrifugation (12,000 � g, 4°C, 30 min). The cell lysateswere used subsequently for enzyme assays.
The activity of xylose isomerase of the whole-cell lysate was deter-mined in a 1-ml reaction volume containing 70 mM xylose, 20 mMMgCl2, 5 mM MnSO4, and 2 mM DL-dithiothreitol in 100 mM Tris buffer(pH 7.5) (30). The reaction mixture was incubated for 30 min at 30°C, andthen 0.5 M HClO4 was added to stop the reaction. The xylulose producedwas quantified by using the cysteine-carbazole-sulfuric acid assay by mea-suring the absorbance at 540 nm (11). The enzyme activity of xylulosekinase was determined by the reduction of xylulose in the reaction mix-ture as described previously, with minor modifications (30). The reactionmixture contained the following: 50 mM Tris-HCl buffer (pH 7.5), 2.0mM MgCl2, 2.0 mM ATP, 0.2 mM phosphoenolpyruvate, 0.2 mMNADH, 10 U pyruvate kinase (EC 2.7.1.40), 10 U lactate dehydrogenase(EC 1.1.1.27), and 8.5 mM xylulose. Consumption of NADH was mea-sured by a spectrophotometer at 340 nm. One unit of enzyme activity wasdefined as the amount of enzyme which generated 1 �mol of product permin. The protein amount in the cell extracts was detected by using theCoomassie protein assay reagent (5).
Analytical methods. The biomass in the fermentation broth was de-termined on a UV-visible spectrophotometer set at a wavelength of 600
nm (Lambda-25; Perkin-Elmer, USA). Riboflavin initially was identifiedand quantified by using high-performance liquid chromatography(HPLC) and later was quantified at a wavelength of 444 nm on a BeckmanDU-800 spectrophotometer based on a modified protocol from Sauer etal. (29). Briefly, the fermentation broth was diluted with 0.1 M phosphatebuffer and heated at 80°C for 10 min before centrifugation. The superna-tant then was assayed. Similarly, for sugar and biosolvent analysis, culturebroths were centrifuged at 10,000 � g for 10 min at 4°C, and the super-natant fluids were stored at �20°C until further analysis. The concentra-tions of glucose and xylose were measured using the YSI Life Sciences 7100MBS multiparameter biochemistry analyzer. Glucose and xylose sensorsare configured in the same electrode chamber, allowing simultaneousmeasurement of these sugars in 1 min. Biosolvents (i.e., acetone, ethanol,and butanol) were measured by a gas chromatograph (GC; model 7890A;Agilent Technologies, USA) on a Durabond (DB)-WAXetr column (30 mby 0.25 mm by 0.25 �m; J&W, USA) equipped with a flame ionizationdetector (FID). The oven temperature was held initially at 60°C for 2 min,increased at 15°C/min to 230°C, and then held for 1.7 min. Helium wasused as the carrier gas, with a column flow rate of 1.5 ml/min. Five-pointstandard curves were established by running standard solutions contain-ing acetone, butanol, and ethanol.
RESULTSEfficient coproduction of butanol and riboflavin from xylose bystrain BOH3. In contrast to the two most well-known butanol-producing species, C. acetobutylicum ATCC 824 and C. beijerinckiiNCIMB 8052, the newly discovered Clostridium sp. strain BOH3could produce slightly larger amounts of butanol (7.4 versus 7.0g/liter) from xylose than from glucose (30 g/liter each) (6). Thisobservation inspired further investigation on strain BOH3’s capa-bility to utilize xylose at a higher concentration. When 60 g/liter ofxylose was fed as the sole carbon source to cultures of BOH3, cellsmultiplied rapidly (the optical density at a wavelength of 600 nmreached 4.9) during the first 28 h with only small amounts ofbutanol production (3.4 g/liter). After that, acetone, butanol, andethanol concentrations increased rapidly to highest values of 5.3,14.9, and 1.2 g/liter, respectively, within 62 h (Fig. 1A and B). Thiscorresponds to a yield of 0.25 g/g for butanol and 0.36 g/g for totalsolvents of acetone, butanol, and ethanol (ABE). Interestingly,riboflavin also appeared during the fermentation process andreached a concentration of 110.5 mg/liter, which is much higherthan those found in previous studies using either wild-type (50mg/liter) or gene-modified Clostridium strains (70 mg/liter) (3, 8).In comparison, when glucose (60 g/liter) was used as the solecarbon source, the substrate consumption curves almost over-
TABLE 1 Xylose consumption and total solvent (acetone, butanol, and ethanol) production by different Clostridium species fed with 60 g/literxylose
Strain Gene type
Xylose concn (g/liter) Solvent titer (g/liter)
Reference or sourceInitial Residual Butanol ABE
C. acetobutylicum ATCC 824 Wild type 60 36.7 � 0.2 4.2 � 0.1 6.7 � 0.2 14C. acetobutylicum 824-TALa Modified 60 32.5 � 0.3 5.1 � 0.2 8.1 � 0.2 14C. acetobutylicum ATCC 824 Wild type 60 35.7 � 0.5 4.3 � 0.3 6.8 � 0.1 17C. acetobutylicum EA2018b Mutant 60 23.6 � 1.2 8.5 � 0.1 11.6 � 0.1 17C. beijerinckii NCIMB 8052 Wild type 60 27.3 � 0.9 5.8 � 0.2 7.9 � 0.7 38C. beijerinckii 8052xylR-xylTptb
c Modified 60 4.2 � 0.1 11.6 � 0.1 15.9 � 0.3 38Clostridium sp. strain G117 Wild type 60 24.6 � 0.6 5.2 � 0.1 7.2 � 0.1 This studyClostridium sp. strain BOH3 Wild type 60 0 14.9 � 0.2 21.4 � 0.2 This studya C. acetobutylicum 824-TAL is a transformant from C. acetobutylicum ATCC 824 bearing the E. coli transaldolase (tal) gene (14).b C. acetobutylicum EA2018 is a mutant obtained from C. acetobutylicum ATCC 824 after several rounds of mutagenesis using NTG (N-methyl-N=-nitrosoguanidine) (17).c C. beijerinckii 8052xylR-xylTptb is an engineered strain from C. beijerinckii NCIMB 8052 (putative xylose repressor gene [xylR] inactivation plus xylose proton-symporter gene[xylT] overexpression driven by the ptb promoter) (38).
Cofermentation of Glucose and Xylose to Butanol
August 2014 Volume 80 Number 15 aem.asm.org 4773
on May 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from
lapped with that of xylose (Fig. 1A and C). Strain BOH3 exhibiteda similar sugar consumption rate of 0.83 g/liter/h and generatedslightly smaller amounts of riboflavin (76.8 mg/liter), butanol(14.5 g/liter), and ABE (20.1 g/liter) than those using xylose as asole substrate (Fig. 1C and D). Thus, strain BOH3 represents thefirst wild-type solventogenic Clostridium that could ferment xy-lose to butanol (14.9 g/liter) more efficiently than previously re-ported wild-type (5.8 g/liter) or gene-modified (11.6 g/liter) Clos-tridium strains under similar operational conditions (Table 1).
Simultaneous utilization of glucose and xylose by strainBOH3. Glucose and xylose are the two major components in the
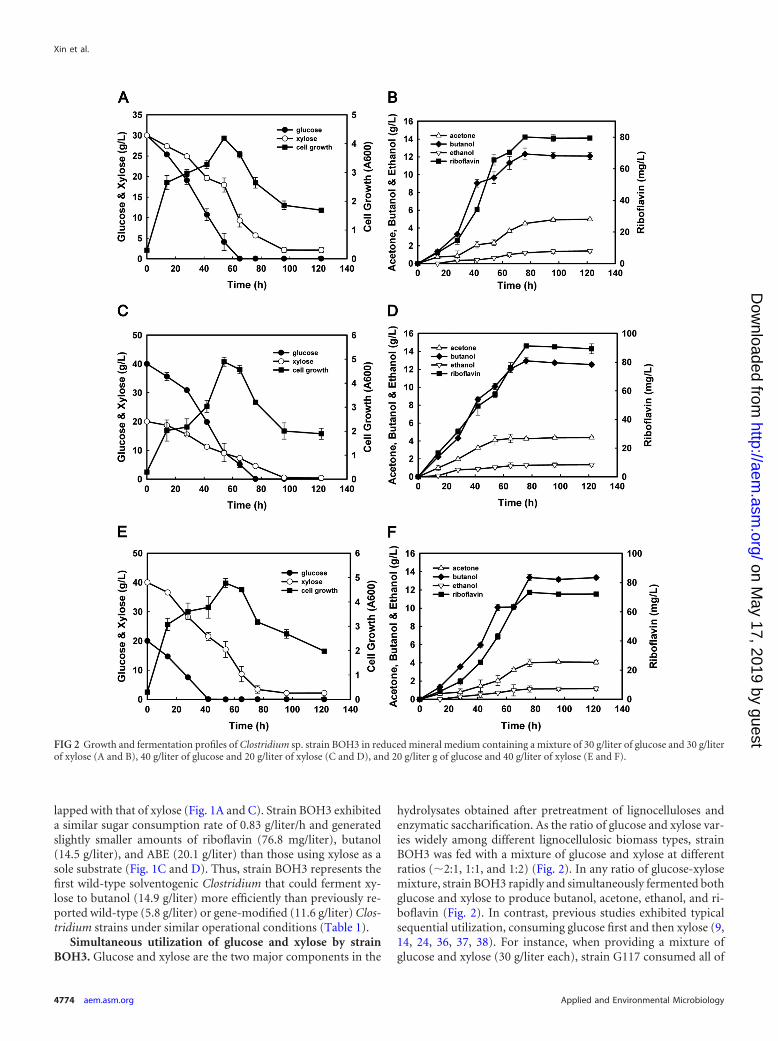
hydrolysates obtained after pretreatment of lignocelluloses andenzymatic saccharification. As the ratio of glucose and xylose var-ies widely among different lignocellulosic biomass types, strainBOH3 was fed with a mixture of glucose and xylose at differentratios (�2:1, 1:1, and 1:2) (Fig. 2). In any ratio of glucose-xylosemixture, strain BOH3 rapidly and simultaneously fermented bothglucose and xylose to produce butanol, acetone, ethanol, and ri-boflavin (Fig. 2). In contrast, previous studies exhibited typicalsequential utilization, consuming glucose first and then xylose (9,14, 24, 36, 37, 38). For instance, when providing a mixture ofglucose and xylose (30 g/liter each), strain G117 consumed all of
FIG 2 Growth and fermentation profiles of Clostridium sp. strain BOH3 in reduced mineral medium containing a mixture of 30 g/liter of glucose and 30 g/literof xylose (A and B), 40 g/liter of glucose and 20 g/liter of xylose (C and D), and 20 g/liter g of glucose and 40 g/liter of xylose (E and F).
Xin et al.
4774 aem.asm.org Applied and Environmental Microbiology
on May 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from
the glucose in the first 48 h, and xylose utilization was initiatedonly after the glucose was completely consumed (after 48 h), re-sulting in a total of 6.5 g/liter butanol after 100 h of fermentation.In comparison, strain BOH3 consumed all of the glucose andmost of the xylose in the glucose-xylose mixtures after 72 h ofincubation (Fig. 2A, C, and E). More xylose in the glucose-xylosemixture led to slightly larger amounts of butanol and riboflavinbeing produced (Fig. 2B, D, and F). For example, when fed withglucose and xylose in a ratio of 1:2, strain BOH3 produced thehighest levels of butanol (13.0 g/liter) and riboflavin (91.4 mg/liter) among the above-described three substrate ratios, which isconsistent with the results in Fig. 1 showing higher butanol andriboflavin production after consuming the same amount of xyloseas glucose.
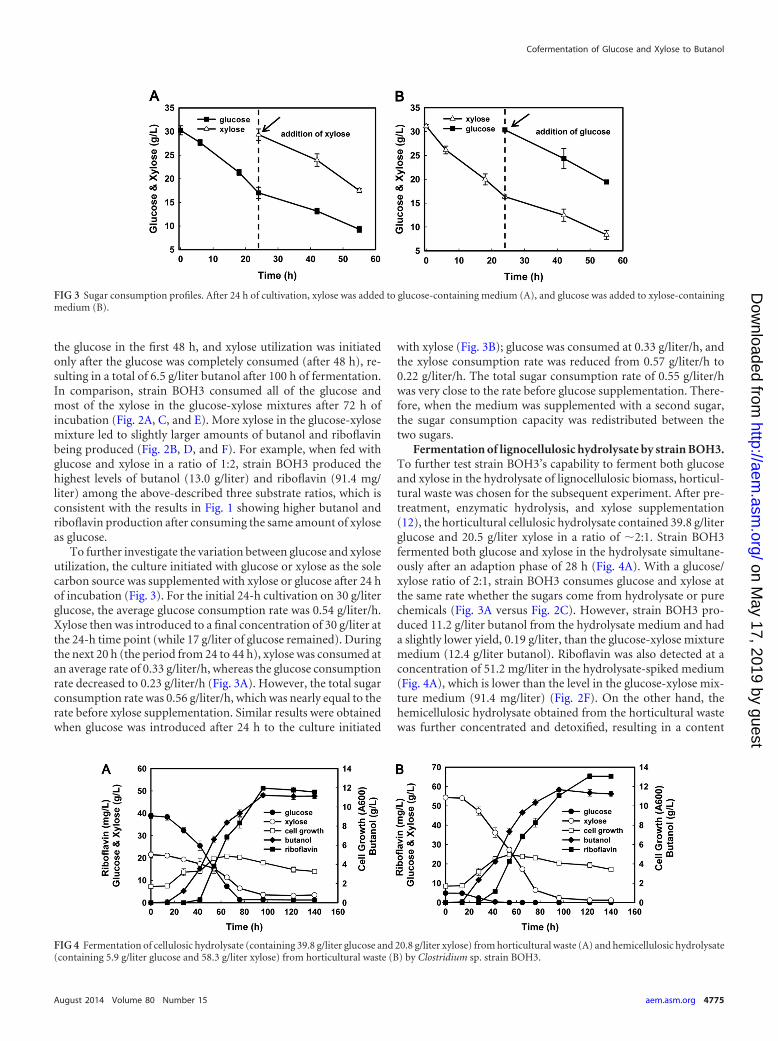
To further investigate the variation between glucose and xyloseutilization, the culture initiated with glucose or xylose as the solecarbon source was supplemented with xylose or glucose after 24 hof incubation (Fig. 3). For the initial 24-h cultivation on 30 g/literglucose, the average glucose consumption rate was 0.54 g/liter/h.Xylose then was introduced to a final concentration of 30 g/liter atthe 24-h time point (while 17 g/liter of glucose remained). Duringthe next 20 h (the period from 24 to 44 h), xylose was consumed atan average rate of 0.33 g/liter/h, whereas the glucose consumptionrate decreased to 0.23 g/liter/h (Fig. 3A). However, the total sugarconsumption rate was 0.56 g/liter/h, which was nearly equal to therate before xylose supplementation. Similar results were obtainedwhen glucose was introduced after 24 h to the culture initiated
with xylose (Fig. 3B); glucose was consumed at 0.33 g/liter/h, andthe xylose consumption rate was reduced from 0.57 g/liter/h to0.22 g/liter/h. The total sugar consumption rate of 0.55 g/liter/hwas very close to the rate before glucose supplementation. There-fore, when the medium was supplemented with a second sugar,the sugar consumption capacity was redistributed between thetwo sugars.
Fermentation of lignocellulosic hydrolysate by strain BOH3.To further test strain BOH3’s capability to ferment both glucoseand xylose in the hydrolysate of lignocellulosic biomass, horticul-tural waste was chosen for the subsequent experiment. After pre-treatment, enzymatic hydrolysis, and xylose supplementation(12), the horticultural cellulosic hydrolysate contained 39.8 g/literglucose and 20.5 g/liter xylose in a ratio of �2:1. Strain BOH3fermented both glucose and xylose in the hydrolysate simultane-ously after an adaption phase of 28 h (Fig. 4A). With a glucose/xylose ratio of 2:1, strain BOH3 consumes glucose and xylose atthe same rate whether the sugars come from hydrolysate or purechemicals (Fig. 3A versus Fig. 2C). However, strain BOH3 pro-duced 11.2 g/liter butanol from the hydrolysate medium and hada slightly lower yield, 0.19 g/liter, than the glucose-xylose mixturemedium (12.4 g/liter butanol). Riboflavin was also detected at aconcentration of 51.2 mg/liter in the hydrolysate-spiked medium(Fig. 4A), which is lower than the level in the glucose-xylose mix-ture medium (91.4 mg/liter) (Fig. 2F). On the other hand, thehemicellulosic hydrolysate obtained from the horticultural wastewas further concentrated and detoxified, resulting in a content
FIG 3 Sugar consumption profiles. After 24 h of cultivation, xylose was added to glucose-containing medium (A), and glucose was added to xylose-containingmedium (B).
FIG 4 Fermentation of cellulosic hydrolysate (containing 39.8 g/liter glucose and 20.8 g/liter xylose) from horticultural waste (A) and hemicellulosic hydrolysate(containing 5.9 g/liter glucose and 58.3 g/liter xylose) from horticultural waste (B) by Clostridium sp. strain BOH3.
Cofermentation of Glucose and Xylose to Butanol
August 2014 Volume 80 Number 15 aem.asm.org 4775
on May 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from
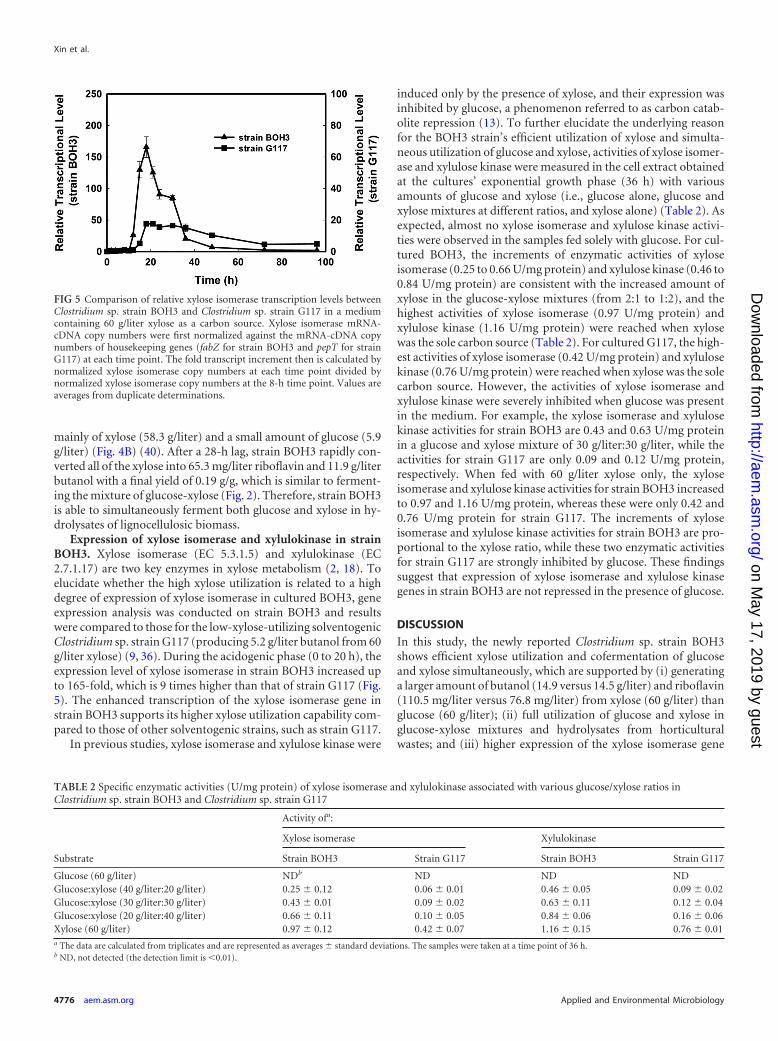
mainly of xylose (58.3 g/liter) and a small amount of glucose (5.9g/liter) (Fig. 4B) (40). After a 28-h lag, strain BOH3 rapidly con-verted all of the xylose into 65.3 mg/liter riboflavin and 11.9 g/literbutanol with a final yield of 0.19 g/g, which is similar to ferment-ing the mixture of glucose-xylose (Fig. 2). Therefore, strain BOH3is able to simultaneously ferment both glucose and xylose in hy-drolysates of lignocellulosic biomass.
Expression of xylose isomerase and xylulokinase in strainBOH3. Xylose isomerase (EC 5.3.1.5) and xylulokinase (EC2.7.1.17) are two key enzymes in xylose metabolism (2, 18). Toelucidate whether the high xylose utilization is related to a highdegree of expression of xylose isomerase in cultured BOH3, geneexpression analysis was conducted on strain BOH3 and resultswere compared to those for the low-xylose-utilizing solventogenicClostridium sp. strain G117 (producing 5.2 g/liter butanol from 60g/liter xylose) (9, 36). During the acidogenic phase (0 to 20 h), theexpression level of xylose isomerase in strain BOH3 increased upto 165-fold, which is 9 times higher than that of strain G117 (Fig.5). The enhanced transcription of the xylose isomerase gene instrain BOH3 supports its higher xylose utilization capability com-pared to those of other solventogenic strains, such as strain G117.
In previous studies, xylose isomerase and xylulose kinase were
induced only by the presence of xylose, and their expression wasinhibited by glucose, a phenomenon referred to as carbon catab-olite repression (13). To further elucidate the underlying reasonfor the BOH3 strain’s efficient utilization of xylose and simulta-neous utilization of glucose and xylose, activities of xylose isomer-ase and xylulose kinase were measured in the cell extract obtainedat the cultures’ exponential growth phase (36 h) with variousamounts of glucose and xylose (i.e., glucose alone, glucose andxylose mixtures at different ratios, and xylose alone) (Table 2). Asexpected, almost no xylose isomerase and xylulose kinase activi-ties were observed in the samples fed solely with glucose. For cul-tured BOH3, the increments of enzymatic activities of xyloseisomerase (0.25 to 0.66 U/mg protein) and xylulose kinase (0.46 to0.84 U/mg protein) are consistent with the increased amount ofxylose in the glucose-xylose mixtures (from 2:1 to 1:2), and thehighest activities of xylose isomerase (0.97 U/mg protein) andxylulose kinase (1.16 U/mg protein) were reached when xylosewas the sole carbon source (Table 2). For cultured G117, the high-est activities of xylose isomerase (0.42 U/mg protein) and xylulosekinase (0.76 U/mg protein) were reached when xylose was the solecarbon source. However, the activities of xylose isomerase andxylulose kinase were severely inhibited when glucose was presentin the medium. For example, the xylose isomerase and xylulosekinase activities for strain BOH3 are 0.43 and 0.63 U/mg proteinin a glucose and xylose mixture of 30 g/liter:30 g/liter, while theactivities for strain G117 are only 0.09 and 0.12 U/mg protein,respectively. When fed with 60 g/liter xylose only, the xyloseisomerase and xylulose kinase activities for strain BOH3 increasedto 0.97 and 1.16 U/mg protein, whereas these were only 0.42 and0.76 U/mg protein for strain G117. The increments of xyloseisomerase and xylulose kinase activities for strain BOH3 are pro-portional to the xylose ratio, while these two enzymatic activitiesfor strain G117 are strongly inhibited by glucose. These findingssuggest that expression of xylose isomerase and xylulose kinasegenes in strain BOH3 are not repressed in the presence of glucose.
DISCUSSION
In this study, the newly reported Clostridium sp. strain BOH3shows efficient xylose utilization and cofermentation of glucoseand xylose simultaneously, which are supported by (i) generatinga larger amount of butanol (14.9 versus 14.5 g/liter) and riboflavin(110.5 mg/liter versus 76.8 mg/liter) from xylose (60 g/liter) thanglucose (60 g/liter); (ii) full utilization of glucose and xylose inglucose-xylose mixtures and hydrolysates from horticulturalwastes; and (iii) higher expression of the xylose isomerase gene
FIG 5 Comparison of relative xylose isomerase transcription levels betweenClostridium sp. strain BOH3 and Clostridium sp. strain G117 in a mediumcontaining 60 g/liter xylose as a carbon source. Xylose isomerase mRNA-cDNA copy numbers were first normalized against the mRNA-cDNA copynumbers of housekeeping genes (fabZ for strain BOH3 and pepT for strainG117) at each time point. The fold transcript increment then is calculated bynormalized xylose isomerase copy numbers at each time point divided bynormalized xylose isomerase copy numbers at the 8-h time point. Values areaverages from duplicate determinations.
TABLE 2 Specific enzymatic activities (U/mg protein) of xylose isomerase and xylulokinase associated with various glucose/xylose ratios inClostridium sp. strain BOH3 and Clostridium sp. strain G117
Substrate
Activity ofa:
Xylose isomerase Xylulokinase
Strain BOH3 Strain G117 Strain BOH3 Strain G117
Glucose (60 g/liter) NDb ND ND NDGlucose:xylose (40 g/liter:20 g/liter) 0.25 � 0.12 0.06 � 0.01 0.46 � 0.05 0.09 � 0.02Glucose:xylose (30 g/liter:30 g/liter) 0.43 � 0.01 0.09 � 0.02 0.63 � 0.11 0.12 � 0.04Glucose:xylose (20 g/liter:40 g/liter) 0.66 � 0.11 0.10 � 0.05 0.84 � 0.06 0.16 � 0.06Xylose (60 g/liter) 0.97 � 0.12 0.42 � 0.07 1.16 � 0.15 0.76 � 0.01a The data are calculated from triplicates and are represented as averages � standard deviations. The samples were taken at a time point of 36 h.b ND, not detected (the detection limit is �0.01).
Xin et al.
4776 aem.asm.org Applied and Environmental Microbiology
on May 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from

and higher enzymatic activities of xylose isomerase and xylulosekinase, responsible for xylose utilization. Moreover, compared topreviously reported wild and mutant strains in batch fermenta-tions fed with 60 g/liter xylose, strain BOH3 showed the highestxylose utilization (100%) and butanol production (14.9 g/liter)(Table 1). Therefore, among all of the reported solventogenic mi-croorganisms, simultaneous utilization of glucose and xylose en-ables Clostridium sp. strain BOH3 to produce the largest amountof butanol (12.4 g/liter) from glucose and xylose mixtures undersimilar growth conditions (14, 37, 38).
During the fermentation process, strain BOH3 coproduces ri-boflavin, which adds to the economic value of the process. Usu-ally, a major concern for a coproduction process is whether thesecond product affects the yield of the major product. For strainBOH3, high butanol production (14.9 g/liter) still occurred, ac-companying 110.5 mg/liter riboflavin generation, which is close toits saturation amount (120 mg/liter) in the medium (26, 32).Therefore, results in this study support the rationale of coproduc-tion of another value-added product in the ABE fermentationprocess so as to make it more economically feasible. However,levels of butanol and riboflavin produced from the hydrolysates ofhorticultural wastes are still lower than those from a pure mono-saccharide mixture. It is known that furfural and hydroxymethyl-furfural present in lignocellulosic hydrolysates can be convertedto their alcohol or acid derivatives by microorganisms throughNAD(P)H/NAD(P)-dependent redox reactions (1, 40). Theseprocesses may break the cofactor balance in sugar metabolism ofClostridium to affect butanol formation (1). In addition, othercomponents (e.g., vanillin) in the hydrolysate may also affect thesugar transport system (1, 40).
For solventogenic Clostridium, xylose is usually transportedinto cytoplasm initially by a xylose transporter and then is furtherconverted into xylulose-5-phosphate by xylose isomerase and xy-lulose kinase (18, 37, 38). Strengthening this xylose transport sys-tem in C. acetobutylicum ATCC 824 and C. beijerinckii NCIMB8052 has been proven to partially improve xylose uptake; however,they still show lower xylose utilization than glucose utilization,and carbon catabolite repression still exists in C. acetobutylicumATCC 824 (37, 38). For example, xylose was still left at 13 g/literafter glucose was fully utilized in a 40 g/liter glucose-20 g/literxylose mixture when using gene-modified C. acetobutylicumATCC 824 (overexpression of xylT and inactivation of phosphoe-nolpyruvate [PES]-dependent phosphotransferase system [PTS])(37). By further overexpression of xylose isomerase and xylulosekinase, C. acetobutylicum ATCC 824 consumed nearly all of thexylose (20 g/liter) in the presence of glucose (40 g/liter) and im-proved butanol production from 7.9 g/liter to 9.1 g/liter, althoughthe bottleneck still existed because of its low butanol tolerance inxylose (37). In contrast, strain BOH3 shows a higher transcriptionlevel of xylose isomerase and enzymatic activities of xylose isomer-ase and xylulose kinase when using xylose alone than those of thelow-xylose-utilizing strains, such as G117 (Table 2). Most impor-tantly, the activities of xylose isomerase and xylulose kinase instrain BOH3 are not inhibited even in the presence of glucose. Incontrast, their enzymatic activities in strain G117 were stronglyinhibited by glucose (Table 2). As is known, the inhibition ofexpression of genes encoding enzymes responsible for catabolismof the carbon source over the preferred ones is the main cause ofcarbon catabolic repression. Moreover, the higher butanol toler-ance (15.2 g/liter) of strain BOH3 (6) than for previously reported
wild-type microbes (e.g., 9.0 g/liter for C. acetobutylicum ATCC824 and 7.0 g/liter Clostridium sp. strain G117) could efficientlyalleviate the butanol inhibitory effect on the energy-requiring xy-lose transport system located in the cell membrane (4, 24, 34).Therefore, the high expression of xylose degradation enzymes (xy-lose isomerase and xylulose kinase) in the presence of glucosecontributes to its efficient xylose consumption and relief fromcarbon catabolite repression.
Conclusions. Clostridium sp. strain BOH3 shows unique capa-bilities in simultaneously consuming glucose and xylose to pro-duce butanol at yields of up to 14.9 g/liter with riboflavin as acoproduct. Therefore, strain BOH3 is a promising candidate intransforming lignocellulosic biomass into biofuel due to its effi-cient use of both xylose and glucose.
ACKNOWLEDGMENTS
This research is supported by the National Research Foundation, PrimeMinister’s Office, Singapore, under the Competitive Research Programme,with project no. NRF-CRP5-2009-05.
REFERENCES1. Almeida JR, Bertilsson M, Gorwa-Grauslund MF, Gorsich S, Liden G.
2009. Metabolic effects of furaldehydes and impacts on biotechnologicalprocesses. Appl. Microbiol. Biotechnol. 82:625– 638. http://dx.doi.org/10.1007/s00253-009-1875-1.
2. Aristidou A, Penttila M. 2000. Metabolic engineering applications torenewable resource utilization. Curr. Opin. Biotechnol. 11:187–198. http://dx.doi.org/10.1016/S0958-1669(00)00085-9.
3. Bacher A, Eberhardt S, Eisenreich W, Fischer M, Herz S, Illarionov B,Kis K, Richter G. 2001. Biosynthesis of riboflavin. Vitam. Horm. 61:1– 49.http://dx.doi.org/10.1016/S0083-6729(01)61001-X.
4. Bowles LK, Ellefson WL. 1985. Effects of butanol on Clostridium aceto-butylicum. Appl. Environ. Microbiol. 50:1165–1170.
5. Bradford MM. 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding.Anal.Biochem.72:248 –254.http://dx.doi.org/10.1016/0003-2697(76)90527-3.
6. Bramono SE, Lam YS, Ong SL, He JZ. 2011. A mesophilic Clostridiumspecies that produces butanol from monosaccharides and hydrogen frompolysaccharides. Bioresour. Technol. 102:9558 –9563. http://dx.doi.org/10.1016/j.biortech.2011.07.077.
7. Brysch K, Schneider C, Fuchs G, Widdel F. 1987. Lithoautotrophicgrowth of sulfate-reducing bacteria, and description of Desulfobacteriumautotrophicum gen. nov., sp. nov. Arch. Microbiol. 148:264 –274. http://dx.doi.org/10.1007/BF00456703.
8. Cai X, Bennett GN. 2011. Improving the Clostridium acetobutylicumbutanol fermentation by engineering the strain for co-production of ribo-flavin. J. Ind. Microbiol. Biotechnol. 38:1013–1025. http://dx.doi.org/10.1007/s10295-010-0875-6.
9. Chua TK, Liang DW, Qi C, Yang KL, He J. 2013. Characterization of abutanol-acetone-producing Clostridium strain and characterization of itssolventogenic genes. Bioresour. Technol. 135:372–378. http://dx.doi.org/10.1016/j.biortech.2012.08.085.
10. Compere A, Griffith W. 1979. Evaluation of substrates for butanol pro-duction. Dev. Ind. Microbiol. 20:509 –517.
11. Dische Z, Borenfreund E. 1951. A new spectrophotometric method forthe detection and determination of keto sugars and trioses. J. Biol. Chem.192:583–587.
12. Geng AL, Xin FX, Ip JY. 2012. Ethanol production from horticulturalwaste treated by a modified organosolv method. Bioresour. Technol. 104:715–721. http://dx.doi.org/10.1016/j.biortech.2011.10.076.
13. Grimmler C, Held CC, Liebl W, Ehrenreich A. 2010. Transcriptionalanalysis of catabolite repression in Clostridium acetobutylicum growing onmixtures of D-glucose and D-xylose. J. Biotechnol. 150:315–323. http://dx.doi.org/10.1016/j.jbiotec.2010.09.938.
14. Gu Y, Li J, Zhang L, Chen J, Niu LX, Yang YL, Yang S, Jiang HW.2009. Improvement of D-xylose utilization in Clostridium acetobutyli-cum via expression of the talA gene encoding transaldolase from Esche-
Cofermentation of Glucose and Xylose to Butanol
August 2014 Volume 80 Number 15 aem.asm.org 4777
on May 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from

richia coli. J. Biotechnol. 143:284 –287. http://dx.doi.org/10.1016/j.jbiotec.2009.08.009.
15. He J, Ritalahti KM, Aiello MR, Löffler FE. 2003. Complete detoxificationof vinyl chloride by an anaerobic enrichment culture and identification ofthe reductively dechlorinating population as a Dehalococcoides popula-tion. Appl. Environ. Microbiol. 69:996 –1003. http://dx.doi.org/10.1128/AEM.69.2.996-1003.2003.
16. Hickey RJ. 1945. The inactivation of iron by 2,2=-bipyrimidine and itseffect on riboflavin synthesis by Clostridium acetobutylicum. Arch.Biochem. 8:439 – 447.
17. Hu SY, Zheng HJ, Gu Y, Zhao JB, Zhang WW, Yang YL, Wang SY,Zhao GP, Yang S, Jiang WH. 2011. Comparative genomic and transcrip-tomic analysis revealed genetic characteristics related to solvent formationand D-xylose utilization in Clostridium acetobutylicum EA 2018. BMCGenomics 12:93–111. http://dx.doi.org/10.1186/1471-2164-12-93.
18. Jeffries TW. 1983. Utilization of D-xylose by bacteria, yeasts, andfungi. Adv. Biochem. Eng. Biotechnol. 27:1–32. http://dx.doi.org/10.1007/BFb0009101.
19. Joshua CJ, Dahl R, Benke PI, Keasling JD. 2011. Absence of diauxieduring simultaneous utilization of D-glucose and D-xylose by Sulfolobusacidocaldarius. J. Bacteriol. 193:1293–1301. http://dx.doi.org/10.1128/JB.01219-10.
20. Kim JH, Block DE, Mills DA. 2010. Simultaneous consumption of pen-tose and hexose sugars: an optimal microbial phenotype for efficient fer-mentation of lignocellulosic biomass. Appl. Microbiol. Biotechnol. 88:1077–1085. http://dx.doi.org/10.1007/s00253-010-2839-1.
21. Mitchell WJ. 1998. Physiology of carbohydrate to solvent conversion byclostridia. Adv. Microb. Physiol. 39:31–130.
22. Nichols NN, Dien BS, Bothast RJ. 2001. Use of catabolite repressionmutants for fermentation of sugar mixtures to ethanol. Appl. Microbiol.Biotechnol. 56:120 –125. http://dx.doi.org/10.1007/s002530100628.
23. Nolling J, Breton G, Omelchenko MV, Makarova KS, Zeng Q, GibsonR, Lee HM, Dubois J, Qiu D, Hitti J, Wolf YI, Tatusov RL, Sabathe F,Doucette-Stamm L, Soucaille P, Daly MJ, Bennett GN, Koonin EV,Smith DR. 2001. Genome sequence and comparative analysis of the sol-vent-producing bacterium Clostridium acetobutylicum. J. Bacteriol. 183:4823– 4838. http://dx.doi.org/10.1128/JB.183.16.4823-4838.2001.
24. Ounine K, Petitdemange H, Raval G, Gay R. 1985. Regulation andbutanol inhibition of D-xylose and D-glucoseuptake in Clostridium ace-tobutylicum. Appl. Environ. Microbiol. 49:874 – 878.
25. Pfaffl MW. 2001. A new mathematical model for relative quantification inreal-time PCR. Nucleic Acids Res. 29:e45. http://dx.doi.org/10.1093/nar/29.9.e45.
26. Perlman D. 1979. Microbial process for riboflavin production, p 521–527.In Pepplev HJ, Pevlman D (ed), Microbial technology, vol I. AcademicPress, New York, NY.
27. Pridham TG. 1946. Microbial synthesis of riboflavin. Econ. Bot. 6:185–205.
28. Qureshi N, Blaschek HP. 2001. ABE production from corn: a recenteconomic evaluation. J. Ind. Microbiol. Biotechnol. 27:292–297. http://dx.doi.org/10.1038/sj.jim.7000123.
29. Sauer U, Hatzimanikatis V, Hohmann HP, Manneberg M, van LoonAP, Bailey JE. 1996. Physiology and metabolic fluxes of wild-type andriboflavin-producing Bacillus subtilis. Appl. Environ. Microbiol. 62:3687–3696.
30. Shamanna DK, Sanderson KE. 1979. Uptake and catabolism of D-xylosein Salmonella typhimurium LT2. J. Bacteriol. 139:64 –70.
31. Stahmann KP, Revuelta JL, Seulberger H. 2000. Three biotechnicalprocesses using Ashbya gossypii, Candida famata, or Bacillus subtilis com-pete with chemical riboflavin production. Appl. Microbiol. Biotechnol.53:509 –516. http://dx.doi.org/10.1007/s002530051649.
32. Vandamme EJ. 1992. Production of vitamins, coenzymes and relatedbiochemicals by biotechnological processes. J. Chem. Tech. Biotechnol.53:313–327.
33. Vinuselvi P, Kim MK, Lee SK, Ghim CM. 2012. Rewiring carbon catab-olite repression for microbial cell factory. BMB Rep. 45:59 –70. http://dx.doi.org/10.5483/BMBRep.2012.45.2.59.
34. Vollherbst-Schneck K, Sands JA, Montenecourt BS. 1984. Effect ofbutanol on lipid composition and fluidity of Clostridium acetobutylicumATCC 824. Appl. Environ. Microbiol. 47:193–194.
35. Widdel F, Hansen TA. 1992. The dissimilatory sulfate- and sulfur-reducing bacteria. In Dworkin M, Falkow S, Rosenberg E, Schleifer K-H,Stackebrandt (ed), The prokaryotes: a handbook on the biology of bacte-ria, vol 1, 2nd ed. Springer Verlag, New York, NY.
36. Wu YR, Li Y, Yang KL, He J. 2012. Draft genome sequence of butanol-acetone-producing Clostridium beijerinckii strain G117. J. Bacteriol. 194:5470 –5471. http://dx.doi.org/10.1128/JB.01139-12.
37. Xiao H, Gu Y, Ning Y, Yang Y, Mitchell WJ, Jiang W, Yang S. 2011.Confirmation and elimination of D-xylose metabolism bottlenecks in D-glucose phosphoenolpyruvate-dependent phosphotransferase system-deficient Clostridium acetobutylicum for simultaneous utilization of D-glucose, D-xylose, and arabinose. Appl. Environ. Microbiol. 77:7886 –7895. http://dx.doi.org/10.1128/AEM.00644-11.
38. Xiao H, Li Z, Jiang Y, Yang Y, Jiang W, Gu Y, Yang S. 2012. Metabolicengineering of D-xylose pathway in Clostridium beijerinckii to optimizesolvent production from D-xylose mother liquid. Metab. Eng. 14:569 –578. http://dx.doi.org/10.1016/j.ymben.2012.05.003.
39. Zaldivar J, Nielsen J, Olsson L. 2001. Fuel ethanol production fromlignocellulose: a challenge for metabolic engineering and process integra-tion. Appl. Microbiol. Biotechnol. 56:17–34. http://dx.doi.org/10.1007/s002530100624.
40. Zhang J, Geng A, Yao C, Lu Y, Li Q. 2012. Xylitol production fromD-xylose and horticultural waste hemicellulosic hydrolysate by a new iso-late of Candida athensensis SB18. Bioresour. Technol. 105:134 –141. http://dx.doi.org/10.1016/j.biortech.2011.11.119.
Xin et al.
4778 aem.asm.org Applied and Environmental Microbiology
on May 17, 2019 by guest
http://aem.asm
.org/D
ownloaded from