simultaneous exposure of mammalian cells to heavy ions and x-rays
TRANSCRIPT

~ Pergamon
www.elsevier.eorn/locate/asr
Adv. Space Res. Vol. 30, No. 4, pp. 877-884, 2002 © 2002 COSPAR. Published by Elsevier Science Ltd. All rights reserved
Printed in Great Britain 0273-1177/02 $22.00 + 0.00
PII" S0273-1177(02)00409-X
SIMULTANEOUS EXPOSURE OF MAMMALIAN CELLS TO HEAVY IONS AND X-RAYS
Y. Furusawa l, M. Aoki 1, and M. Durante 2
1National Institute of Radiological Sciences, 4-9-1, Anagawa, Inage, Chiba 263-8555, Japan 2Dipartimento di Scienze Fisiche, Universit~ Federico II, Monte S. Angelo, 80126 Napoli, Italy
ABSTRACT
Crews of space missions are exposed to a mixed radiation field, including sparsely and densely ionizing radiation. To determine the biological effectiveness of mixed high-/low-LET radiation fields, mammalian cells were exposed in vitro simultaneously to X-rays and heavy ions, accelerated at the HIMAC accelerator. X-ray doses ranged from 1 to 11 Gy. At the same time, cells were exposed to either 4°Ar (550 MeV/n, 86 keV/ktm), 28Si (100 MeV/n, 150 keV/gm), or 56Fe (115 MeV/n, 442 keV/gm) ions. Survival was measured in hamster V79 fibroblasts. Structural aberrations in chromosome 2 were measured by chemical-induced premature chromosome condensation combined with fluorescence in situ hybridization in isolated human lymphocytes. For argon and silicon experiments, measured damage in the mixed radiation field was consistent with the value expected using an additive function for low- and high-LET separated data. A small deviation from a simple additive function is observed with very high-LET iron ions combined to X-rays. © 2002 COSPAR. Publ ished by Elsevier Science Ltd. All rights reserved.
INTRODUCTION
Astronauts are exposed in space to mixed radiation fields, including photons, neutrons, light and heavy charged particles of different energies. The yield Y of any specified biological endpoint (e.g. cell killing, chromosomal aberrations, etc.) can be calculated as (Kraft and Scholz, 1994):
Y= Sf(Z,E).LET(Z,E).RBE(Z,E)dV = ff(Z,E).~ (Z,E)dV (1)
where RBE(Z,E) is the relative biological effectiveness for each particle of charge Z and energy E, f(Z, E) is the fluence spectrum, LET (Z,E) is the LET spectrum, and a=LETxRBE is the action cross section for the specific endpoint. The implicit assumption in Eq. 1 is that there is no interaction among different components in the spectrum. Based on this hypothesis, Y is evaluated as the weighed sum of the action cross sections of the different radiation components.
However, few data exist concerning the interaction of radiation of different qualities. Early interpretation of radiation effects were based on the assumption that densely ionizing radiation acts via a single-hit mechanism, i.e. no sublethal damage is present. An interaction with other types of radiation damage would appear unlikely. In early experiments at the Bevalac (Berkeley), it was shown that cells surviving heavy ion exposure were subsequently sensitized to X-irradiation (Ngo et al., 1981). Evidence
877

878 Y. Furusawa et al.
of a synergism for cell killing between high- and low-LET radiation was also found in later experiments (Bird et al., 1983, McNally et al., 1984, 1988, Ngo et al., 1988), using sequential exposure of mammalian cells to low- and high-LET radiation. Higgins et al. (1983, 1984) found different results in case of sequential exposure to T-rays and neutrons or simultaneous exposure of V79 cells to the same radiation sources, with an increased interaction for simultaneous exposure. Brooks et al. (1990) exposed rat lung epithelial cells simultaneously to X-rays and co-particles. These authors demonstrated that both cell killing and the induction of micronuclei were increased by combined exposures compared with that predicted for separate exposures.
Overall, deta available in the literature suggest a synergistic interaction between high- and low- LET radiation fields. To simulate the space radiation field, it is important to obtain data on simultaneous exposure to photons and HZE particles. To this purpose, a facility for simultaneous exposure to X-rays and heavy ions has been installed at the HIMAC accelerator. In this paper, we will describe the results of the first experiments concerning cell killing in hamster fibroblasts and chromosomal aberrations in human lymphocytes after mixed field exposure at this facility. V79 cells have been widely used to measure cell inactivation following exposure to heavy ions (Furusawa et al., 2000, Aoki et al., 2000). Chromosomal damage has been measured using the novel technique of chemical-induced premature chromosome condensation (PCC), combined with fluorescence in situ hybridization (FISH). Compared to conventional metaphase analysis, PCC allows the scoring of aberrations independently from cell-cycle perturbations (Durante et al., 1996, 1998). This is important in mixed-field experiments, because heavy ions induce mitotic delay and interphase death more efficiently than X-rays.
In the present paper we will present results obtained with 3 different particles: 4°Ar (550 MeV/n, 86 keV/~trn), 2SSi (100 MeV/n, 150 keV/lxrn, 0.6-1.5 Gy; chromosomal aberration data only), or 56Fe (115 MeV/n, 442 keV/~n).
MATERIALS AND METHODS
Cells
V79 hamster fibroblasts were grown in NUNC T25 flasks in MEM medium (Nissui Pharmaceutical Co., Tokyo) supplemented with 10% serum and antibiotics (Penicilin and Streptmycin). Cells were harvested by trypsinization (Trypsin 1:250, Gibco-BRL, Grand Island, NY) and seeded one day before irradiation at a concentration of about 2.5 x 105 cells/flask, then exposed in log phase.
Whole blood was collected in a Vacutainer CPT (Beckton-Dickinson, Lincoln Park, USA), from a healthy male donor. White blood cells were isolated by centrifugation and resuspended in RPMI 1640 medium (Gibco-BRL, Grand Island, NY), supplemented with 20% fetal calf serum, 1% L-glutamine, 1% penicillin/streptomycin, 0.1% sodium heparin (stock 176.2 units/mg). About 1 ml of the medium containing the cells was then loaded by a syringe into blood holders (1 mm thick), specifically designed for HIMAC experiments
Irradiation
Details of the experimental apparatus is shown in Figure 1. Flasks or blood holders are kept at 45 ° and exposed simultaneously to a vertical beam of X-rays (150 kVp, 20 mA, 0.5 mm A1 and Cu filters), and to heavy ions, from the horizontal accelerator pipe. The distance from the sample to the X-ray tube is set to achieve an exposure rate E around 100 R/min. For each dose D of X-rays, the exposure time t is calculated as:
t (min)= D(cGy) • [0.96.E(R/min)] ~ (2)

Interaction Between Heavy Ions and X-Rays 879
The fluence rate of the heavy ion beam accelerated at HIMAC is modulated to deliver the dose required in the same time t (Eq. 2). Samples exposed to X-rays only or heavy ions only were exposed in the same experimental conditions.
All exposures were performed at room temperature, and samples were processed immediately after exposure as described below.
Survival assay
V79 cells were trypsinized and counted immediately after exposure. Cell singlets were plated in 60-mm dishes at low density. Dishes were fixed and stained after 6 days incubation at 37 °C. Colonies containing more than 50 cells were scored as survivors.
X
Mo RF ML BC
RS
SC
Fig. 1. Experimental apparatus for simultaneous X-rays/heavy ion exposure. W; wall between accelerator and experimental hall. Mo; beam intensity monitors. RF; revolving unit of ridge filter. RS; inserting unit of range shifters. ML; beam profile monitor and laser pointer. BC; block collimator. Co; sample collimator. X; X-ray tube. SC; sample-changer/transporter. S; samples. SC; sample driver.
Chromosome condensation
Immediately after exposure, lymphocytes were collected from the blood holder with a syringe, and moved to a 25 c m 2 tissue culture flask containing 9 ml of RPMI medium (see above), supplemented with 1% phytohaemagglutinin (Gibco). Flasks were incubated at 37 °C in a vertical position and with a loosened cap. After 24 h, 40 ~tl of colcemid (stock 10 I.tg/ml, Wako Chemicals, Japan; final concentration: 40 ng/ml) was added to the medium, and the tube was incubated again for 22 h. Five microliters of calyculin A (Wako Chemicals, Japan. Stock 0.1 mM in ethanol, stored at -20 °C. Final concentration: 50 nM) was finally added and the tube incubated 1 h at 37 °C. Following incubation, cells were centrifuged

880 Y. Furusawa et al.
and resuspended in hypotonic solution, fixed in Carnoy's solution (methanol:acetic acid = 3:1), and dropped on wet slides. High condensation indexes (30-40%) were obtained with this procedure.
Chromosome painting
Slides were hybridized with whole-chromosome-paint probes specific for human chromosome 2 (spectrum green). The protocol suggested by the manufacturer (Vysis Inc., Downers Grove, IL) was basically followed, with few modifications (Durante et al., 1996). Slides were counterstalned in propidium iodide and viewed on an Olympus fluorescence microscope. Only spreads in GI, (32, and M- phase were scored for aberrations. All aberrations in chromosome 2 were scored. In our data analysis, we concentrate on the fraction F of cells displaying aberrations in chromosome 2, to achieve a better statistics.
Statistical analysis
The theory of dual radiation action provides a simple method to estimate the effects of mixed fields under the assumption of total independence among the different radiation components (Zaider and Rossi, 1980). If fraction of cells surviving the irradiation (S) or carrying aberrations in chromosome 2 (F) follows a linear-quadratic relationship:
S= exp(-od:)-~D 2) ; F = otD+~D 2 (3)
then after a dose dx of X-rays combined to a dose dp of charged particles, the expected fractions Sexp. and Fcxp. of surviving and aberrant ceils following the dose D=dx+dp will be:
where Sexp. =exp(-ff,~xD-~mixD 2) ; fexp.,-- ff~xD+[3mixD 2
ff, mix=fXO~X+fpOtp ; ~mix= [ fx( ~X)~i+fp([~p)~] 2
(4)
(5)
and fx=dx/D, fr~p/D are the fractions of the total dose D delivered by X-rays and charged particles, respectively. We have fitted S and F data for charged particles and X-rays (Figures 2-4) to evaluate cx and
parameters. For survival data, O~mix and 13mix were measured by fitting survival data in mixed fields and
compared with parameters calculated by equation 5 using t-test. For chromosome data, the calculated yield Fexp. (equation 4) was compared with the measured yield Fobs. at the same dose in the mixed field experiment. Standard error on Fobs. was estimated according to binomial statistics as [F(1-F)/N] ~, where N is the total number of cells scored. A ratio e=Fcxp./Fobs. <1 suggests a synergistic effect, while E=I corresponds to additive effect, and e >1 to antagonistic effect (Suzuki, 1994, 1998, Lam, 1995). The 95% confidence interval was evaluated to assess significant deviations from the expected e= 1 value.
RESULTS
Both X-rays and heavy ions induce chromosomal aberrations in lymphocytes. The fraction of cells displaying aberrations in chromosome 2 as a function of photon or charged particle dose is plotted in Figure 2. Fitting parameters for these curves were used to evaluate the expected frequencies Fcxp. as described in methods. Mixed-field data for the 3 different ions are described below. The 3 different experiments were designed to cover a wide LET range (86-442 keV/~tm), different total doses and
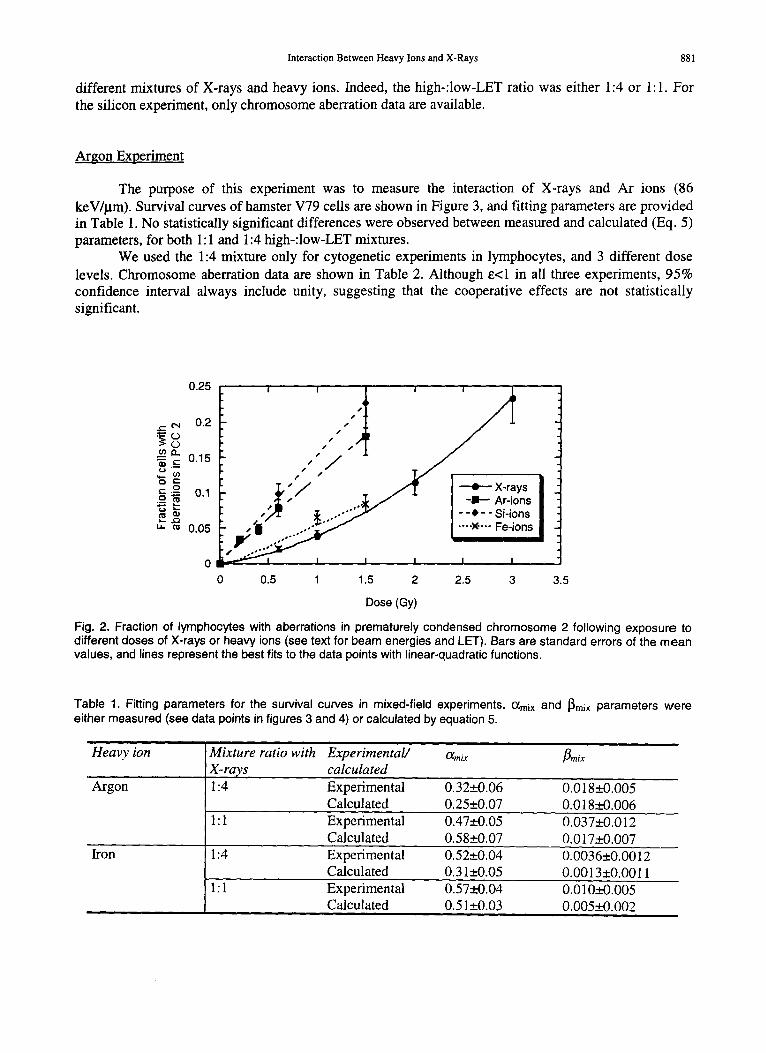
Interaction Between Heavy Ions and X-Rays 881
different mixtures of X-rays and heavy ions. Indeed, the high-:low-LET ratio was either 1:4 or 1:1. For the silicon experiment, only chromosome aberration data are available.
Argon Experiment
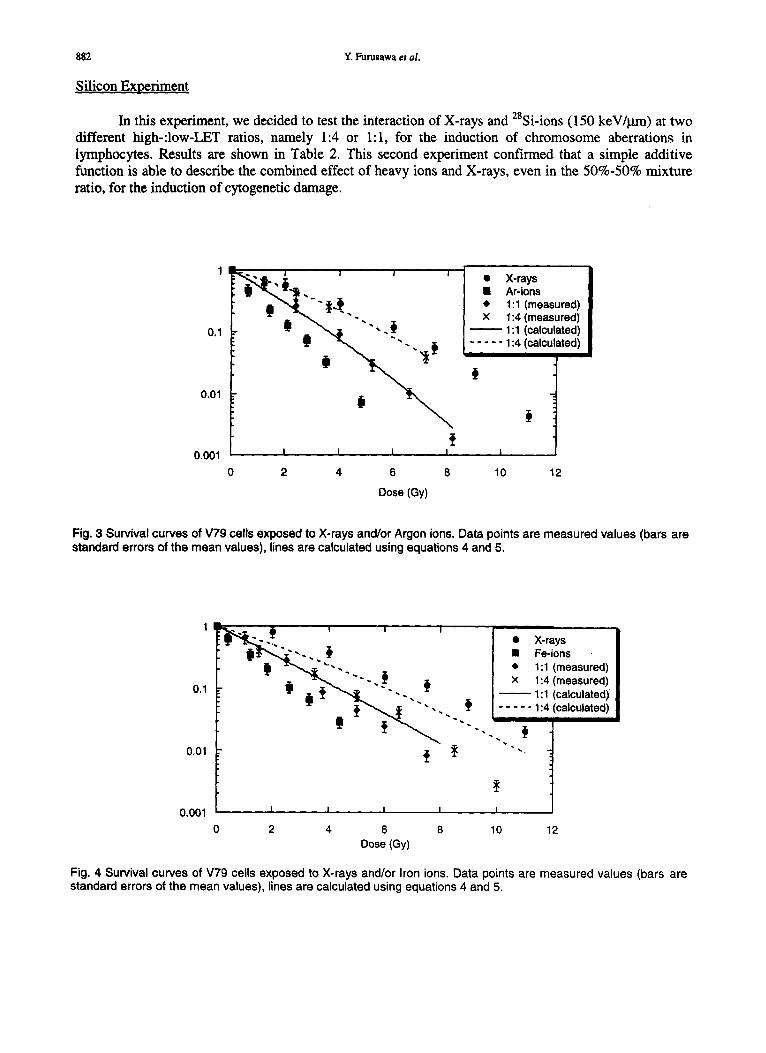
The purpose of this experiment was to measure the interaction of X-rays and Ar ions (86 keV/~m). Survival curves of hamster V79 cells are shown in Figure 3, and fitting parameters are provided in Table 1. No statistically significant differences were observed between measured and calculated (Eq. 5) parameters, for both 1:1 and 1:4 high-:low-LET mixtures.
We used the 1:4 mixture only for cytogenetic experiments in lymphocytes, and 3 different dose levels. Chromosome aberration data are shown in Table 2. Although e<l in all three experiments, 95% confidence interval always include unity, suggesting that the cooperative effects are not statistically significant.
0.25
= ~ 0.2 ~ ~/~ I
.~ 0.15 ,,,, o c-
.9 I , / .,,,11' ~ X-rays .~, ~ 0.1 , , ~ - - - ~, / - - I I - - Ar-ions
- - . - - si- io,s " 0 0 s , 'O" - - - * - - Fe-ions
0 ~ I I I I I
0 0.5 1 1.5 2 2.5 3 3.5
Dose (Gy)
Fig. 2. Fraction of lymphocytes with aberrations in prematurely condensed chromosome 2 following exposure to different doses of X-rays or heavy ions (see text for beam energies and LET). Bars are standard errors of the mean values, and lines represent the best fits to the data points with linear-quadratic functions.
Table 1. Fitting parameters for the survival curves in mixed-field experiments. ~mix and ~mix parameters were either measured (see data points in figures 3 and 4) or calculated by equation 5.
Heavy ion
Argon
Mixture ratio with Experimental/ O~mi x ~n,ix X-rays calculated 1:4 Experimental 0 . 3 2 _ + 9 . 0 6 0.018-+0.005
Calculated 0 . 2 5 _ + 0 . 0 7 0.018_+0.006 1:1 Experimental 0 . 4 7 _ + 0 . 0 5 0.037_+0.012
Calculated 0 . 5 8 _ + 0 . 0 7 0.017_+0.007 Iron 1:4 Experimental 0 .52_ .+0 .04 0.0036_+0.0012
Calculated 0.31_+0.05 0.0013 _+0.0011 1:1 Experimental 0 . 5 7 _ + 0 . 0 4 0.010_+0.005
Calculated 0.51 _+0.03 0.005_+0.002
882 Y. Furusawa et al.
Silicon Experiment
In this experiment, we decided to test the interaction of X-rays and 28Si-ions (150 keW~tm) at two different high-:low-LET ratios, namely 1:4 or 1:1, for the induction of chromosome aberrations in lymphocytes. Results are shown in Table 2. This second experiment confirmed that a simple additive function is able to describe the combined effect of heavy ions and X-rays, even in the 50%-50% mixture ratio, for the induction of cytogenetic damage.
1
0.1
0.01
0.001
".. i , i i • X-rays ~ i . ~ . , . I At-ions
~ . ,,, ~.. • 1:1 (measured) I~1 X 1:4 (measured)
1:1 (calculated) = {11 ~ " . . . . . ~ 1:4 !calculated)
I I I I I
2 4 6 8 10 12
Dose (Gy)
Fig. 3 Survival curves of V79 cells exposed to X-rays and/or Argon ions. Data points are measured values (bars are standard errors of the mean values), lines are calculated using equations 4 and 5.
0,01
0,001
1 ~ . . , ~ I I I
o.1
• X-rays • Fe-ions • 1:1 (measured) x 1:4 (measured)
1:1 (calculated) . . . . . 1:4 (calculated)
I I I
0 2 4 6 8 Dose (Gy)
I I
10 12
Fig. 4 Survival curves of V79 cells exposed to X-rays and/or Iron ions. Data points are measured values (bars are standard errors of the mean values), lines are calculated using equations 4 and 5.

Interaction Between Heavy Ions and X-Rays 883
Iron Experiment
The purpose of this experiment was to repeat the previous experiments with two mixture ratios (1:4 and 1:1), using a very high LET particle: 56Fe ions, energy in vacuum 200 MeV/n, LET in the sample 442 keV//am. Results are displayed in Figure 4 and Table 1 (cell killing) and Table 2 (chromosomal aberrations).
Table 2. Results of chromosomal aberration experiments. The fraction Fobs, of lymphocytes with aberrations in chromosome 2 is reported in this table. Corresponding theoretical value Fexp. calculated by equations 4 and 5 is also reported with its uncertainty. The ratio E=Fexp./Fobs. is provided in the last column, with the 95% confidence limit.
Radiation
X+Ar X+Ar X+Ar X+Ar X+Ar X+Ar X+Si X+Si X+Si X+Si X+Fe X+Fe X+Fe X+Fe
Dose (Gy) Cells scored Aberrant cells e (% ±SE) (95%
confidence interval)
1.0+0.2 642 48 (7.5±1.0) 1.0+0.2 Calculated 6.6±1.7 0.88 (0.4-1.4) 2.0+0.4 223 40 (17.9±2.6) 2.0+0.4 Calculated 17.0±3.3 0.95 (0.5-1.4) 3.0+0.6 172 56 (32.6_+.3.6) 3.0+0.6 Calculated 31.4±5.7 0.96 (0.6-1.4) 3.0+0.6 147 28 (19.0±3.2) 3.0+0.6 Calculated 33.5±8.4 1.76 (0.7-2.8) 1.5+1.5 88 23 (26.1_+4.7) 1.5+1.5 Calculated 32.9±9.1 1.26 (0.4-1.9) 3.0+0.6 327 79 (24.2+_2.4) 3.0+0.6 Calculated 26.8__.4.4 1.10 (0.6-1.6) 1.5+1.5 309 76 (24.6+_2.4) 1.5+1.5 Calculated 16.0±4.1 0.66 (0.3-1.0)
Calculated survival curves using equations 4 and 5 provide a poor fit of the experimental data. Differences between measured and calculated ~mJx parameter from survival curves is significant for the 1:5 mixture ratio (t-test, p<0.05) only. Some difference is also observed for chromosomal aberration data, but this time a significant deviation from predictions (equations 4 and 5) is observed for the 1:1 mixture (0.3<e<1.0 at 95% confidence level).
DISCUSSION
We have presented results of the interaction between heavy ions and X-rays in the induction of cell killing in hamster fibroblasts and chromosomal aberrations in human lymphocytes. Within the limits of the statistics achieved, Ar-ions (86 keV//.tm) and Si-ions (150 keV/~tm) data are well described by an additive function. No statistically significant interaction is observed for these ions, nor for cell killing

884 / Y. Furusawa et al.
neither for chromosomal damage. For Fe-ions (442 keV/~'a), a small deviation from a simple additive function is observed in survival and chromosomal aberration data, although the effect is barely statistically significant, and appears to be strongly dependent on the fitting function used. Overall, these data indicate that the additive function describes adequately a simultaneous exposure to heavy ions and photons, although more experiments seems to be necessary for very high-LET particles.
REFERENCES
Aoki, M., Y. Furusawa, and T. Yamada, LET Dependency of Heavy-Ion Induced Apoptosis in V79 Ceils, J. Radiat. Res., 41, 163-175, 2000.
Bird, R.P., M. Zaider, H.H. Rossi, E.J. Hall, S.A. Marino, and N. Rohrig, The Sequential Irradiation of Mammalian Cells with X Rays and Charged Particles of High LET, Radiat. Res., 93, 444-452, 1983.
Brooks, A.L., G.J. Newton, L.J. Shyr, F.A. Seiler, and B.R. Scott, The Combined Effects of Alpha- Particles and X-Rays on Cell Killing and Micronuclei Induction in Lung Epithelial Cells, Int. J. Radiat. Biol., 58, 799-811, 1990.
Durante, M., K. George, and T.C.Yang, Biological Dosimetry by Interphase Chromosome Painting, Radiat. Res., 145, 53-60, 1996.
Durante, M., Y. Furusawa, and E. Gotoh, A Simple Method for Simultaneous Interphase-Metaphase Chromosome Analysis in Biological Dosimetry, Int. J. Radiat. BioL, 74, 457-462, 1998.
Furusawa, Y., K. Fukutsu, M. Aoki, H. Itsukaichi, K. Eguchi-Kasai, H. Ohara, F. Yatagai, T. Kanai, and K. Ando, Inactivation of Aerobic and Hypoxic Cells from Three different Cell Lines by Accelerated 3He-, 12C- and 2°Ne-Ion Beams, Radiat. Res., 154, 485-496, 2000.
Higgins, P.D,, P.M. DeLuca, D.W. Pearson, and M.N. Gould, V79 Survival Following Simultaneous or Sequential Irradiation by 15-MeV Neutrons and COCo Photons, Radiat. Res., 95, 45-56, 1983.
Higgins, P.D., P.M. DeLuca, and M.N. Gould, Effect of Pulsed Dose in Simultaneous and Sequential Irradiation of V-79 Cells by 14.8-MeV Neutrons and coCo Photons, Radiat. Res., 99, 591-595, 1984.
Kraft, G., and M. Scholz, On the Parametrization of the Biological Effect in a Mixed Radiation Field, Adv. Space Res. 14, 234-244, 1994.
Lain, G.K.Y., The Definition of Damage Interaction in International Commission of Radiation Units and Measurement Report 30, Phys. Med. Biol., 40, 9-16, 1995.
McNally, N.J., J. de Ronde, and M. Hinchliffe, The Effect of Sequential Irradiation with X-rays and Fast Neutrons on the Survival of V79 Chinese Hamster Cells, Int. J. Radiat. Biol., 45, 301-310, 1984.
McNally, N.J., J. de Ronde, and M. Folkard, Interaction Between X-ray and or-Particle Damage in V79 Cells, Int. J. Radiat. Biol., 53, 917-920, 1988.
Ngo, F.Q.H., E.A. Blakeley, and C.A. Tobias, Sequential Exposure of Mammalian Cells to Low- and High-LET Radiations. I. Lethal Effects Following X-ray and Neon Irradiation, Radiat. Res., 87, 59- 78, 1981.
Ngo, F.Q.H., E.A. Blakeley, C.A. Tobias, P.Y. Chang and L. Lommel, Sequential Exposure of Mammalian Cells to Low- and High-LET Radiations. II. As a Function of Cell-Cycle Stages, Radiat. Res., 115, 54-69, 1988.
Suzuki, S., The "Synergistic" Action of Mixed Irradiation with High-LET and L0w-LET Radiation, Radiat. Res., 138, 297-301, 1994.
Suzuki, S,, A Theoretical Model for Simultaneous Mixed Irradiation with Multiple Types of Radiation, J. Radiat. Res., 39, 215-221, 1998.
Zaider, M., and H.H. Rossi, The Synergistic Effects of Different Radiations. Radiat. Res., 83, 732-739, 1980.