shellfish aquaculture and the environment (shumway/shellfish aquaculture and the environment) ||...
TRANSCRIPT

Chapter 9
Expanding s hellfi sh a quaculture: a r eview of the e cological s ervices p rovided by and i mpacts of n ative and c ultured b ivalves in s hellfi sh - d ominated e cosystems Loren D. Coen , Brett R. Dumbauld , and Michael L. Judge
Introduction
Aquaculture is making an ever - increasing con-tribution to the worldwide demand for shell-fi sh at the same time that native “ wild stock ” populations have been or are signifi cantly declining (Naylor et al. 2000 ; Beck et al. 2009, in press ). At the same time, there has been increasing awareness of the ecosystem services that native bivalves provide as habitat, biofi l-ters, and shoreline stabilizers (reviewed in Newell 2004 ; Coen et al. 2007 ; Grabowski
and Peterson 2007 ; see also Chapter 1 in this book). Habitat - forming bivalve species as those that are (1) reef - forming; (2) aggregation - forming; or (3) shell - accumulating species (ASMFC 2007 ). Given (1) the worldwide decline of native species ’ populations (Kirby 2004 ; Airoldi and Beck 2007 ; Beck et al. 2009 ; Gillespie 2009 ); (2) the associated loss of criti-cal ( “ nursery ” ) habitats; and (3) the ever - increasing worldwide contribution of cultured native and nonnative species, the benefi cial services and negative impacts associated with
Shellfi sh Aquaculture and the Environment, First Edition. Edited by Sandra E. Shumway.© 2011 John Wiley & Sons, Inc. Published 2011 by John Wiley & Sons, Inc.
239

240 Shellfi sh Aquaculture and the Environment
commercial culture. We use indirect evidence where available to gauge these effects for other cultured species, especially oysters (see also Heffernan 1999 ; Deal 2005 ; Dumbauld et al. 2009 ; Forrest et al. 2009 ; NRC 2009, 2010 ) and provide a brief overview of the state of our knowledge regarding natural bivalve com-munities, reefs and aggregations, “ ecosystem services, ” justifi cation for shellfi sh protection, enhancement, and restoration efforts. We then include several case studies of shellfi sh aqua-culture and discuss the direct and indirect, biotic and abiotic, and both positive and nega-tive effects in an estuarine or marine context. For shellfi sh these impacts vary by (1) location of habitat (intertidal vs. subtidal); (2) extent of cultured versus natural acreage (scale); (3) other elements associated with the proposed area to be farmed (e.g., existing seagrasses, competitors, benthos, available hard substrate, exotics); and (4) latitudinal/geographical dif-ferences (cf. native or introduced species, regu-lations, western Atlantic, Gulf of Mexico, or Eastern Pacifi c). For some systems there is a wealth of general information that is transfer-able in part to specifi c sites (e.g., carrying capacity, lost habitat structure); for others site - specifi c sampling and related research is required (e.g., marine mammals, fl ow, com-petitors, submerged aquatic vegetation [SAV]). All of these factors should be addressed before deciding whether bivalve aquaculture can partly or fully replace lost natural systems and the resultant effects (NRC 2009, 2010 ), espe-cially given our limited understanding of these natural, and in many cases, “ relic ” native shellfi sh populations (Beck et al. 2009, in press ).
Shellfi sh h abitats, e cosystem s ervices, and e ngineers
Shellfi sh habitats — whether a living assem-blage or an accumulation of dead shells — provide the necessary hard substrate for the
native shellfi sh should be examined. This eval-uation is critical since expanding aquaculture may provide many of the same services as native shellfi sh populations, while reducing fi shing pressure on native “ wild stocks ” (e.g., Shumway and Kraeuter 2004 ; NRC 2010 ). Most or all native oyster species on the U.S. West Coast are signifi cantly depressed or extir-pated (e.g., U.S. West Coast, Canada, and Mexico oysters, Ostrea lurida and Ostrea con-chaphila , reviewed in McGraw 2009 ), whereas Crassostrea virginica is either still harvested directly on the Gulf of Mexico and eastern U.S. coasts because of suffi cient larval supply and settlement substrates (e.g., Coen et al. 2007 ; Brumbaugh and Coen 2009 ).
In contrast to fi nfi sh aquaculture (numerous reviews), the use of molluscs as “ farmed ” species is generally perceived as benign, except perhaps with regard to the production of mussels on an industrial scale where signifi -cant impacts have been demonstrated quite often (e.g., Grant et al. 1995 ; Black 2001 ; Alongi et al. 2003 ; Asmus and Asmus 2005 ; Cranford et al. 2007 ; McKindsey et al. 2009 ; Paraskevi et al. 2009 ). In addition to the above - mentioned services, there are numerous examples where the shellfi sh aquaculture industry has fought for improved water quality standards, and helped to regulate and enhance waste water treatment, septic system upgrades (see also Lindahl et al. 2005 ; Gren et al. 2009 ) and the impacts of upland runoff (Chapters 8 and 9 in this book). Although numerous studies have focused on mussels grown in rope culture, associated carrying capacities, and related community and ecosystem effects, few if any directed studies have assessed molluscan aquaculture ’ s impacts or its parallel services to native - dominated systems (cf. NRC 2009, 2010 ).
In this chapter, we summarize what is known about ecosystem services provided by bivalves, particularly in the United States, as well as any potential ecosystem impacts (both negative and positive), resulting from their

Impacts of native and cultured bivalves 241
biology and ecology of most shellfi sh species has primarily focused, until recently, on resource - related questions (e.g., Coen et al. 1999a, 1999b ; Lenihan 1999 ; Luckenbach et al. 1999 ; Coen and Luckenbach 2000 ; Beck et al. 2003 ). Shellfi sh, traditionally, have only been considered a resource to be extracted (e.g., Lenihan and Peterson 1998 ; Coen et al. 1999b, 2007 ; Beck et al. 2009 ), and most state and federal fi sheries management agencies did not view shell bottom habitats as essential fi sh habitat ( EFH ) until the late 1990s (e.g., Thayer 1992 ; Luckenbach et al. 1999 ; Coen et al. 1999b ). The broader ecological value of shell-fi sh recently has begun to gain more universal recognition (Luckenbach et al. 1999 ; Coen et al. 2007 ; Grabowski and Peterson 2007 ; Beck et al. 2009 ). The result has been new and broadened shellfi sh research and restoration perspectives (Luckenbach et al. 1999 ; Coen and Luckenbach 2000 ; Thayer et al. 2003, 2005 ; Grabowski and Peterson 2007 ; Schulte et al. 2009 ).
Complex, h abitat - f orming s hellfi sh s pecies
For bivalves and gastropod molluscs, the cal-careous shell functions as the protective exo-skeleton forming the foundation for the above - outlined ecosystem services. Some shell-fi sh (e.g., oysters and mussels) support signifi -cant commercial and recreational fi sheries, with a subset creating unique and important three - dimensional habitats, particularly when in high densities. Using a previously developed classifi cation (ASMFC 2007 ), we segregated these shellfi sh - dominated habitats into one of three major habitat - forming “ morphotypes ” : (1) reefs (a three - dimensional structure with a veneer of living and dead animals only paral-leling coral reefs); (2) shell aggregations (living and dead shell); and (3) shell accumulations (dead; often referred to as shell hash) (reviewed in ASMFC 2007 ; Beck et al. 2009 ). Both
attachment of many epifaunal species that would not be present otherwise in areas con-sisting of soft sediments . Along most of the world ’ s temperate to tropical shores, shellfi sh populations currently occur or once occurred in estuaries, as well as near - shore and offshore seafl oor of the continental shelf (e.g., Kirby 2004 ; Shumway and Kraeuter 2004 ; Airoldi and Beck 2007 ; Beck et al. 2009, in press ). One of the most well studied and widely occurring, habitat - forming species is the eastern oyster, Crassostrea virginica . Crassostrea virginica occurs from the St. Lawrence River, Canada, to the Atlantic coast of Argentina (Kennedy et al. 1996 ). Human activities, in concert with natural phenomena, have signifi cantly affected the distribution and abundance of this oyster throughout its range as with many native habitat - forming bivalve species that are also widely harvested as a resource (see reviews in Kirby 2004 ; Kirby and Miller 2005 ; NRC 2007 ). Thus, they are unique in that they form “ the habitat, ” but are a “ resource ” and are impacted signifi cantly by harvesting (e.g., Kaufman and Dayton 1997 ; Breitburg et al. 2000 ; Coen and Luckenbach 2000 ; Lenihan and Micheli 2000 ; Smyth et al. 2009 ). Such bivalve - generated habitats have declined precipitously due to many causes including (1) overharvesting; (2) physical dis-turbance by natural and anthropogenic impacts including waves and boat wakes; (3) diseases; (4) nutrient enrichment through runoff; (5) predation; (6) alteration of natural fl ow regimes and salinity patterns; and (7) loss of appropriate substrate for new recruits (e.g., Kennedy et al. 1996 ; Luckenbach et al. 1999 ; Coen and Luckenbach 2000 ; Beck et al. 2009 ; Volety et al. 2009 ).
In contrast to other estuarine and marine habitats such as seagrasses, marshes, and man-groves, whose importance, functioning, and protection has been long studied (e.g., Nixon 1980 ; Boesch and Turner 1984 ; Costanza et al. 1997 ; Beck et al. 2001, 2003 ; Williams and Heck 2001 ; Faunce and Serafy 2006 ), the

242 Shellfi sh Aquaculture and the Environment
represented by scallops such as Placopecten magellanicus , a species that although not a true reef - former in the strictest sense, often occurs in suffi cient densities to generate scallop shell - dominated habitats utilized by many other species (Langton and Robinson 1990 ; Stokesbury 2002 ; Talman et al. 2004 ). In many areas, pen shells (e.g., genera Atrina , Pinna ), and other densely aggregating infaunal bivalves whose shells often protrude above the substrate, generate complex habitats with numerous species using both living animals or dead articulated shells as habitat (e.g., Perry 1936 ; Keough 1984 ; Cummings et al. 1998 ; Kuhlmann 1998 ; Norkko et al. 2006 ; Munguia 2007 ). Many estuarine and brackish water bivalve clams also occur in dense accumula-tions to form unique habitat assemblages (e.g., Rangia , Polymesoda , and other members of the corbiculid family). Mussels and pen shells also affect the abiotic realm. By protruding a few centimeters above the bottom, these shells create signifi cant topographic roughness ele-ments that substantially affect fl ow, creating larger areas of turbulence, thereby changing local hydrodynamics and particle transport (e.g., Offi cer et al. 1982 ; Keough 1984 ; Dame 1996 ; Cummings et al. 1998 ; Coen et al. 1999b ; Norkko et al. 2006 ; ASMFC 2007 ; see also Chapter 10 in this book).
A third shellfi sh habitat type, the “ Shell Accumulation ” group includes ocean quahogs ( Arctica islandica ), surf clams ( Spisula solidis-sima ), and other abundant large bivalves (e.g., Mercenaria spp., Tresus spp., and Mya are-naria ) whose shells often persist long after the living organism is gone (NRC 2010 ). Shells of these species often accumulate on the seabed of the continental shelf and in estuaries in quantities suffi cient to provide signifi cant structure and habitat for a variety of organ-isms (e.g., Dumbauld et al. 1993, 2000 ; Auster et al. 1995 ; Palacios et al. 2000 ; Steimle and Zetlin 2000 ; Stoner and Titgen 2003 ). Recent studies and modeling efforts have focused on the decrease of available shell stocks in areas
bivalve and gastropod molluscs form these types of shellfi sh habitats, with some species falling into one or more of the above (classes 2 or 3) depending on the relative abundance of “ dead ” shell versus live individuals. A fourth “ assemblage ” type that potentially par-allels many of the above - mentioned attributes are the high - density shellfi sh farms and associ-ated structural hardware utilized by the aqua-culture industry, along with its extensive netting and anchoring structures and devices.
Members of two widespread genera, Crassostrea and Mytilus , are examples of reef - forming shellfi sh that occur globally through-out many coastal areas (Galtsoff 1964 ; Sellers and Stanley 1984 ; Seed 1992 ; Seed and Suchanek 1992 ). Before hand and mechanized harvesting, subtidal oyster reefs in many estu-aries extended meters to tens of meters above the bottom, forming extensive and complex three - dimensional reef structures that pro-vided thousands to tens of thousands of hect-ares of habitat for various fi nfi sh and invertebrates (reviewed in Hargis and Haven 1999 ; Coen et al. 1999b ; Peterson et al. 2003 ; ASMFC 2007 ; Grabowski and Peterson 2007 ; Dumbauld et al. 2009 ). Many bivalves (espe-cially oysters such as Crassostrea virginica ) have even more direct impacts, in addition to their signifi cant fi ltering at higher turbidity levels (discussed in Coen 1995 ; Harsh and Luckenbach 1999 ; Hewitt and Norkko 2007 ; NRC 2010 ). These often very localized impacts infl uence one or all of the following: (1) recruitment; (2) growth; and (3) other organ-isms that live on or near the reef that seek shelter or food (Suchanek 1985 ; Zimmerman et al. 1989 ; Breitburg et al. 1995 ; Cummings et al. 1998 ; Kuhlmann 1998 ; Breitburg 1999 ; Kennedy and Sanford 1999 ; Lenihan 1999 ; Coen et al. 1999b ; Beadman et al. 2004 ; Luckenbach et al. 2005 ; Tolley and Volety 2005 ; Rodney and Paynter 2006 ; ASMFC 2007 ; Munguia 2007 ; Stunz et al. 2010 ).
Other habitat - forming species are those in the “ Shell Aggregation ” group. These may be

Impacts of native and cultured bivalves 243
where shell is “ mined ” for restoration efforts. These studies have shown that extant shellfi sh populations are no longer generating enough new shell, and that ocean acidifi cation may be impacting the survival of live animals and the longevity of nonliving shell (discussed in Powell et al. 2006 ; Mann and Powell 2007 ; Powell and Klinck 2007 ; NRC 2010 ). Aquaculture may be one solution for supply-ing the increasing amounts of shell required for restoration (NRC 2010 ), addressing buff-ering by shell dissolution from increasing global CO 2 emissions, as well as declining estuarine pH (NRC 2010 ; Waldbusser et al. 2011 ). Accumulated shell also has important consequences for critical habitat of many off-shore and inshore species. One hypothesis for the crash of Florida ’ s calico scallop ( Argopecten gibbus ) fi shery in the 1980s is the loss of for-merly signifi cant shell accumulations at off-shore nursery sites (ASMFC 2007 ; W. Arnold and W. Lyons, pers. comm.).
A fourth habitat category (ASMFC 2007 ), “ Shellfi sh Aquaculture, ” has received little or no previous attention, but potential services and associated ecosystem values include most of those generated by wild stock bivalve popu-lations (summarized in Coen et al. 2007 ; Grabowski and Peterson 2007 ; and discussed further below), and may include other “ poten-tial ” and “ realized ” services or negative impacts that natural shellfi sh populations do not provide or cause. The diversity of bivalve shellfi sh species cultured worldwide is rela-tively high, with Crassostrea gigas (often referred to as the Pacifi c or Japanese oyster) having the widest global distribution through both deliberate and accidental introductions (reviewed in Ruesink et al. 2005 ; Molnar et al. 2008 ; Dumbauld et al. 2009 ; Smaal et al. 2009 ; Wrange et al. 2010 ; Chapter 14 ). Given the widespread occurrence and ever - increasing scale of molluscan aquaculture in intertidal and subtidal waters (see Naylor et al. 2000 ; NRC 2010 ; FAO 2009a ), and the signifi cant structural elements they provide in coastal and
estuarine waters, this fourth habitat type needs to be assessed in much greater detail as pointed out in several recent reviews (Dumbauld et al. 2009 ; NRC 2009, 2010 , and references therein).
Together, these four groups of shellfi sh habitat provide signifi cant amounts of “ above - bottom ” structure not only for many mobile fi sh and invertebrate species but also for sessile species (Forrest et al. 2007 ; McKindsey et al. 2007 ; D ’ Amours et al. 2008 ; Mallet et al. 2009 ; Watson et al. 2009 ; NRC 2010 ). All four shellfi sh habitat reefs (aggregations, shell accumulations, and cultured species with asso-ciated “ gear ” ) have been poorly studied (e.g., Safriel 1975 ; Suchanek 1985 ; Sebens 1991 ; Holt et al. 1998 ; ASMFC 2007 ), especially since many of these habitats are today only found as relics of past and widely occurring distributions (e.g., Kirby 2004 ; Lotze et al. 2006 ; Beck et al. 2009, in press ). In addition to introducing new individuals to the system, bivalve mariculture operations utilize a great diversity of habitat - forming gear (i.e., ropes, rebar, netting; Figs. 9.1 and 9.2 ; see also Dealteris et al. 2004 ; Powers et al. 2007 ; Tallman and Forrester 2007 ; Munroe and McKinley 2007a ; Coen et al. 2010 ; NRC 2010 ). Bivalve mariculture has other implica-tions, such as changes in sedimentation and fl ow rates that are less understood (reviewed in Dumbauld et al. 2009 ; NRC 2009, 2010 ).
For the remainder of this review, we focus on systems dominated by either native or cul-tured bivalves. We highlight parallels between native and cultured ecosystem services, while stressing direct and indirect impacts, as well as potential or demonstrated positive and nega-tive effects, on the surrounding systems.
Shellfi sh e cosystem s ervices
Recently, scientists have begun to attribute numerous important ecological functions to molluscan shellfi sh - dominated systems

244 Shellfi sh Aquaculture and the Environment
vices ” include (1) signifi cant fi ltering capacity; (2) enhanced benthic - pelagic coupling (Porter et al. 2004 ); (3) sediment/shoreline stabiliza-tion; (4) indirect habitat provisioning; and (5) juvenile and adult feeding areas (e.g., nursery habitats, areas of intense predation), among others. Perhaps three of the most important of these services include refuges for juvenile species (nursery areas), benthic - pelagic
(reviewed in Newell 1988, 2004 ; Coen et al. 1999b, 2007 ; Grabowski and Peterson 2007 ; Beck et al. 2009, in press ). The ecological pro-cesses that depend on the shellfi sh habitat described above can be thought of as “ ecosys-tem services ” ( sensu Jones et al. 1994, 1997 ). In addition to their direct habitat - related value, shellfi sh provide important services for the ecosystem as a whole. These “ ecosystem ser-
Figure 9.1 Fouled subtidal clam growout cages in North Carolina. (Adapted from M. Power, Dauphin Island Sea Lab, and Powers et al. 2007 .)
Figure 9.2 ( A) A planted oyster restoration site in Charleston, SC. Oyster shell ( Crassostrea virginica ) stabilized with polypropylene mesh and rebar enforcing rods. (B) Note grunt ( Orthopristis chrysopterus ) caught in the deployed stabiliz-ing mesh in Beaufort County, SC; a negative side effect of the use of larger meshes.
(A) (B)

Impacts of native and cultured bivalves 245
of over 300 species of plants and animals. Coen et al. ( 2006 , unpublished data) observed over 140 fi nfi sh and macroinvertebrates species on South Carolina intertidal reefs. In the southeastern United States (portions of South Carolina, North Carolina, Georgia, and Florida), oyster reefs are a conspicuous inter-tidal rather than subtidal feature in most estu-aries (e.g., Dame 1979 ; Bahr and Lanier 1981 ; Dame et al. 1984a, 1984b ; Stanley and Sellers 1986 ; Coen et al. 1999a, 1999b ; Van Dolah et al. 1999 ; Coen et al. 2004 ; Stunz et al. 2010 ). Shellfi sh also provide signifi cant habitat struc-ture for often larger, mobile ( “ transient ” ) species (e.g., Coen et al. 1999a, 1999b ; Nestlerode 2004 ; ASMFC 2007 ). Given the total absence of any seagrasses in many of these southeastern U.S. estuaries, the presence of oyster reefs and related functioning is even more critical in these productive systems (Beck et al. 2001, 2003 ).
In most estuaries, shellfi sh habitats exist in a landscape of more than one habitat (e.g., oysters, seagrasses, and salt marsh), together potentially providing complex utilization pat-terns and interactions among the numerous species (e.g., Bell et al. 1991 ; Eggleston et al. 1999 ; Micheli and Peterson 1999 ). We are just beginning to understand the broader ecologi-cal importance of this association (Valentine and Heck 1993 ; Peterson and Heck 1999, 2001a, 2001b ; Wall et al. 2008 ; Booth and Heck 2009 ; NRC 2009 ). For example, oysters can fi lter up to 200 L of water per individual per day through direct ingestion, feces ejec-tion, and particle rejection (pseudofeces). Oysters remove both algae and suspended par-ticles while processing water, thereby improv-ing water clarity and quality (Dame et al. 1984b ; Cressman et al. 2003 ; Nelson et al. 2004 ; Newell 2004 ; Grizzle et al. 2006, 2008 ; Newell et al. 2007 ; NRC 2010 ). As a result, bivalves can positively impact light penetra-tion and seagrass photosynthetic rates, which can enhance nutrients and potentially benefi t seagrasses in shallow waters (Peterson and
coupling, and shoreline protection through the reduction of erosion where operations/habitats are intertidal.
Often only a single or a limited number of bivalve species form a dominant feature of inshore (estuarine) or near - shore (coastal) marine ecosystems. For example, the eastern oyster Crassostrea virginica forms living sub-tidal and intertidal reefs in many Atlantic and Gulf coast estuaries. Although Crassostrea vir-ginica can occur to a depth of 30 m, they are found primarily in shallow waters less than 6 m deep to intertidal (Galtsoff 1964 ; Bahr and Lanier 1981 ; Burrell 1986 ; Kennedy et al. 1996 ; Coen et al. 1999b ; ASMFC 2007 ). Oyster reefs, like many other reef - forming invertebrate species, are unique in that they form the actual living reef structure (e.g., Lenihan et al. 2001 ; J.B.C. Jackson et al. 2001 ; Newell 2004 ; Coen et al. 1999b, 2007 ) in estuaries, supporting a host of other organisms generally not found in surrounding sand or mud habitats (e.g., Wells 1961 ; Stanley and Sellers 1986 ; Coen et al. 1999b ; ASMFC 2007 ; Stunz et al. 2010 ). We know very little about the unique ecosystem that native oysters on the Pacifi c coast of the United States (Olympia oyster, Ostrea equestris or Ostrea lurida ) sup-ported when they were suffi ciently abundant, that is, prior to the late 1800s and early 1900s (McGraw 2009 , and related papers in this Special Issue of the Journal of Shellfi sh Research vol. [28]). Recent work has attempted to quantify the contribution of oyster habitat to ecosystem functioning (e.g., Peterson et al. 2003 ; Peterson and Lipcius 2003 ; Grabowski and Peterson 2007 ). Oysters create complex habitats utilized by fi sh, crustaceans, bivalves, numerous other invertebrates, birds, and mammals (reviewed in Coen et al. 1999b ; ASMFC 2007 ), potentially rivaling the role of salt marshes, seagrasses, and mangroves in nursery habitats (e.g., Glancy et al. 2003 ; Coen et al. 2006 , unpublished data; Stunz et al. 2010 ). Wells (1961) surveyed oyster reefs in North Carolina and observed the presence

246 Shellfi sh Aquaculture and the Environment
2004 ; Bain et al. 2007 ; Brumbaugh and Coen 2009 ).
Importance of o yster r eef h abitats
The eastern oyster Crassostrea virginica , once a dominant feature of most Atlantic and Gulf coast estuaries, has drastically declined in many areas across the United States (Lenihan 1999 ; J.B.C. Jackson et al. 2001 ; Kirby 2004 ). Once valued primarily as a resource, oysters are now also recognized as key elements of many estuarine ecosystems (Coen et al. 1999a, 1999b ; Luckenbach et al. 1999 ; Grabowski and Peterson 2007 ). Oysters create complex habitats utilized by fi sh, crustaceans, bivalves, numerous other invertebrates, birds, and mammals (Coen et al. 1999b ; Lehnert and Allen 2002 ; Tolley and Volety 2005 ; ASMFC 2007 ; Stunz et al. 2010 ). During feeding, oysters and other bivalve molluscs can fi lter large quantities of water (Lindahl et al. 2005 ; Borthagaray and Carranza 2007 ; Buschbaum et al. 2009 ; Gren et al. 2009 ), improving water clarity and quality while transferring nutrients from the water column to the benthos (Newell 1988, 2004 ; Dame 1996 ; Dame et al. 2001 ). Declines in oyster populations are associated with decline in critical habitat, shifts from benthic to pelagically dominated communities, adverse effects on other species, reduced water quality, and changes in ecosystem dynamics (Rothschild et al. 1994 ; Newell et al. 2007 ).
Complex reef - forming invertebrate species have been centers of biodiversity throughout geologic time (Kiessling et al. 2010 ). It is now widely recognized that oyster reefs are valu-able habitat for a wide variety of organisms (e.g., Luckenbach et al. 1999 ; Coen et al. 1999b, 2007 ; Rodney and Paynter 2006 ; ASMFC 2007 ; Grabowski and Peterson 2007 ; Stunz et al. 2010 ) and that oyster resources/habitats can be enhanced through directed oyster reef restoration programs (e.g., Coen and Luckenbach 2000 ; Luckenbach et al.
Heck 1999, 2001a, 2001b ; Newell and Koch 2004 ; Valentine and Heck 1993 ; Cerrato et al. 2004 ; Wall et al. 2008 ; Booth and Heck 2009 ).
Habitat c onsiderations
In contrast to the extensive subtidal oyster beds in estuaries such as in the Chesapeake Bay, Delaware Bay, off the Mississippi River (LA), or Apalachicola Bay (FL) in the south-eastern United States (e.g., southern North Carolina, South Carolina, Georgia), oysters are found primarily in the intertidal zone to depths of 1 – 3 m (3 – 9 ft), which is in part dic-tated by local tidal ranges, among other factors (Bahr 1976 ; Bahr and Lanier 1981 ; Burrell 1986 ). Intertidal oyster beds typically become established where salinity is moderately high, food supply is suffi cient, and siltation is not excessive. Intertidal oyster reefs consist of ver-tical clusters built upon a fragile matrix of live and dead shell surrounded by fi ne sediments (Anderson et al. 1979 ; Bahr and Lanier 1981 ; Dame et al. 1984a, 1984b ; Burrell 1986 ; Coen et al. 1999a ; Lenihan and Micheli 2000 ; Lenihan and Peterson 2004 ). Since many oyster larvae remain in the water column for up to 3 weeks after spawning occurs, larvae are subject to distribution throughout an estuary by tidal currents. Since the fi nal larval stage of reef - forming species crawl on the bottom to search for suitable hard substrate upon which to permanently attach, its location is critical to the continued survival and regen-eration of these complex reef habitats. Attachment sites include almost any hard surface, for example, other living oysters, oyster shell, rocks, docks, tree limbs, and pilings. It is during this fi nal process that aqua-culture can fl ourish in areas where suffi cient natural larval supplies are available. However, in many areas, settlement is insuffi cient and requires hatcheries to supplement or totally supply larval recruits (Kennedy 1996 ; Luckenbach et al. 1999 ; Mann and Evans

Impacts of native and cultured bivalves 247
(see ASMFC 2007 ; Grabowski and Peterson 2007 ; Beck et al. 2009, in press ).
While there is a dearth of information on vertebrate or invertebrate use of shellfi sh - dominated reefs (reviewed in Coen et al. 1999b ; ASMFC 2007 ; Grabowski and Peterson 2007 ; NRC 2009, 2010 ), new quantitative information on species associated with natural reefs or other complex, three - dimensional structure is growing (Luckenbach et al. 2005 ; NRC 2010 ; Coen et al., unpublished data). Observational data from intertidal and sub-tidal shellfi sh habitats in North America and Europe has also been summarized into the worldwide status of oysters (Airoldi and Beck 2007 ; Beck et al. 2009, 2011 ; Buschbaum et al. 2009 ; Reise et al. 2009 ; Smaal et al. 2009 ). More information is currently available regarding the impacts of avian predators and associated negative impacts on fi nfi sh aquacul-ture than for intertidal or subtidal shellfi sh operations (see Comeau et al. 2009 ).
Impacts to s horebirds and m ammals
Several studies have been conducted to examine the effects of shorebird foraging or human dis-turbance in intertidal habitats on mariculture. In Canada, Yasue (2005) found that human disturbance of migratory shorebirds causes subtle, but important, changes in foraging by semipalmated plovers ( Charadrius semipalma-tus ) and least sandpipers ( Calidris minutilla ). In NewJersey, Burger (1994) has found that human disturbance can have negative effects on piping plover ( Charadrius melodus ) forag-ing and suggested that it is important to main-tain the historical diversity of coastal habitats in order to alleviate the negative effects of humans on piping plover foraging. In northern California, Oregon, and Washington, seasonal monitoring of shorebird and waterfowl densi-ties has provided invaluable baseline informa-tion, which allows evaluation of the effects of humans on these species (e.g., Colwell 1993 ;
2005 ). As a result, oyster reef restoration pro-grams have expanded rapidly throughout the United States in recent years. Unfortunately, the shell hash needed to rebuild reefs is gener-ally scarce in those areas where restoration is needed most (Bushek et al. 2004 ; Coen et al. 2007 ). Intertidal oysters bordering salt marshes can prevent erosion of tidal edge habitats (dis-cussed in Meyer et al. 1997 ; Piazza et al. 2005 ; ASMFC 2007 ; Grabowski and Peterson 2007 ; Beck et al. 2009, in press ; Coen et al., unpub-lished data) such as mangroves or marshes reducing wave impacts in tropical to subtropi-cal zones (e.g., Danielsen et al. 2005 ; Kathiresan and Rajendran 2005 ; NRC 2007 ; Feigin et al. 2009 ). For example, in the southeastern United States, reefs often fringe Spartina and Juncus marsh edges in creeks and rivers, forming a unique association that protects these critical marsh habitats from natural (e.g., tidal or wind - driven waves) and anthropogenically derived (e.g., boat wakes) erosion (e.g., Meyer et al. 1997 ; Meyer and Townsend 2000 ; Piazza et al. 2005 ; ASMFC 2007 ; Goodwin 2007 ; Beck et al. 2009, in press ; Coen et al., unpub-lished data).
Use of s hellfi sh h abitats by i nvertebrates and fi nfi sh
We now know that shellfi sh - generated reefs have signifi cantly greater vertical, three - dimensional relief when compared with the surrounding two - dimensional bottom, thereby greatly enhancing biodiversity (reviewed in ASMFC 2007 ; NRC 2010 ). As discussed above, eastern oyster Crassostrea virginica reefs harbor diverse communities largely restricted to the reef structure or other hard - bottom habitats, versus adjacent nonreef habitat. This enhanced vertical relief is of major importance, with implications for assessing habitat value for managed species, and is responsible for a revised management policy regarding these important biogenic reefs

248 Shellfi sh Aquaculture and the Environment
sideration for the eastern oyster on the U.S. East and Gulf coasts, perhaps because oyster aquaculture is only now being taken up in these areas. These potential issues need to be addressed as part of the Environmental Impact Statement (EIS) assessment and permitting process in areas where a potential “ confl ict ” may arise between naturally occurring species and bivalve aquaculture (FAO 2009b ). The ecology of these systems is too variable and the diversity of impacts too diverse and complex to transpose the limited results from other areas to new sites without additional assess-ments, especially in areas where “ user ” con-fl icts may occur (e.g., Forrest et al. 2009 ; NRC 2010 ).
Restoration of n atural h abitats and a quaculture ’ s p otential r ole
Most subtidal oyster reefs currently found in “ Approved ” shellfi sh harvesting areas (i.e., open to direct harvesting based on human health standards) rarely extend even a meter off the bottom while still providing one or more services for economically and ecologi-cally important species (Bahr and Lanier 1981 ; Sellers and Stanley 1984 ; Dame 1996 ; Breitburg 1999 ; Coen et al. 1999a, 1999b ; Lenihan 1999 ; Posey et al. 1999 ; Harding and Mann 2001 ; Peterson et al. 2003 ; Nestlerode 2004 ; Powell et al. 2008 ; Schulte et al. 2009 ). Intertidal oyster reefs are still common in many areas, in contrast to subtidal popula-tions of conspecifi cs within the same ecoregion ( sensu Beck et al. 2009, in press ). This is mostly due to the fact that individuals from these intertidal habitats are often not sought after for the raw bar trade.
Large - and small - scale restoration of both subtidal and intertidal oyster habitats is ongoing along most Atlantic and Gulf coast states (discussed in Coen and Luckenbach 2000 ; Burrows et al. 2005 ; Beck et al. 2009, in press ; Brumbaugh and Coen 2009 ; ORET
Colwell and Landrum 1993 ; Dodd and Colwell 1996 ; Warnock et al. 1998 ; Stempien 2007 ). Other studies have attempted to associate prey availability with shorebird abundances on intertidal mudfl ats in South America, and throughout Europe (e.g., Pienkowski 1983 ; Backwell et al. 1998 ; Triplet et al. 1999 ; Sanchez et al. 2006 ).
Numerous birds use intertidal sand - or mudfl ats as feeding and resting sites during their normal activities or migrations (e.g., Quammen 1981, 1982, 1984 ; Baird et al. 1985 ; Norris et al. 1998 ; Caldow et al. 2004 ; Goss - Custard et al. 2004 ; Connolly and Colwell 2005 ; Stempien 2007 ; Stillman et al. 2007 ; Kraan et al. 2009 ). For shorebirds asso-ciated with aquaculture, concerns have included one or more of the following: (1) limited access to feeding due to cages and other gear; (2) changes in benthic sediments and associated communities; (3) potential entanglements (see Fig. 9.2 B above) or inges-tion of mesh or other materials; (4) noise related to activities during growout (i.e., plant-ing, harvesting, and related maintenance); and (5) potential for migratory disruptions (e.g., Kelly et al. 1996 ; Kaiser et al. 1998 ; Hilgerloh et al. 2001 ; Connolly and Colwell 2005 ; Forrest et al. 2009 ; Godet et al. 2009 ; Atkinson et al. 2010 ). Unfortunately, too little is known at this point to draw any general conclusions.
For marine and terrestrial mammals, we know even less about their positive or negative impacts related to aquaculture than for shore-birds as discussed above (e.g., Nash et al. 2000 ; Kemper et al. 2003 ; Lloyd 2003 ; Read et al. 2006 ; Roycroft et al. 2006 ; Forrest et al. 2009 ; NRC 2010 ). Recently, this has become an important consideration related to permit-ting, as National Park Service and National Research Council studies were undertaken to assess the impacts of intertidal oyster maricul-ture in California (NRC 2009, 2010 ), where marine mammals were an important factor in the deliberations. This rarely has been a con-

Impacts of native and cultured bivalves 249
by adding adult broodstock to reefs. Additionally, oysters are moved or “ relayed ” into growout areas with lower disease or higher growth and survival rates as in the Delaware Bay (Kennedy 1996 ; Southworth and Mann 1998 ; Luckenbach et al. 1999 ; Coen and Luckenbach 2000 ; Smith et al. 2005 ; Brumbaugh et al. 2006 ; Powell et al. 2008 ; Brumbaugh and Coen 2009 ).
Aquaculture - b ased s ystems
Bivalve s pecies c ommonly c ultured in the U nited S tates
Current w est c oast i ntertidal o yster and c lam m ariculture
Bivalve shellfi sh have been harvested for sub-sistence from estuaries along the U.S. West Coast for thousands of years (Naylor et al. 2000 ; Beck et al. 2009, in press ), but extensive harvest of native oysters ( Ostrea lurida ) began with European colonialism in the mid - 1800s (Baker 1995 ). Similar to the fate of the East Coast ’ s Crassostrea virginica , these oysters were gradually depleted due to a combination of overharvesting, extreme natural events, and the lack of shell replacement to maintain a substrate for natural recruitment (e.g., Kirby 2004 ; Ruesink et al. 2005 ). Eastern oysters ( Crassostrea virginica ) and later Pacifi c oysters ( Crassostrea gigas ) were introduced from the U.S. East Coast and Japan, respectively, in order to overcome this decline (Townsend 1896 ; Steele 1964 ; Lindsay and Simons 1997 ; Robinson 1997 ; Shaw 1997 ). Of particular interest for this chapter and related ones (see Chapter 14 in this book) is the potential impact (both obvious and more subtle) of a novel oyster ’ s (e.g., Pacifi c oyster) ecosystem services and resulting positive and negative effects after its introduction and expansion locally, as well as worldwide (Reise 1998 ; Drinkwaard 1999 ; Black 2001 ; Naylor et al. 2001 ; Ruesink et al.
2009 ; Powers et al. 2009 ; Schulte et al. 2009 ). Determining the success of these restoration efforts is critical to optimizing our use of the requisite limited shell resource, and to be cost - effective for restoring reef habitats. Consensus, however, on what constitutes a successful reef restoration project currently does not exist (e.g., Coen and Luckenbach 2000 ; Thayer et al. 2003, 2005 ; Coen et al. 2004 ; Brumbaugh et al. 2006 ; Powers et al. 2009 ; Hadley et al. 2010 ). The most commonly used metric of success has been the presence of market - sized (e.g., 3 - in. - long by shell height) oysters (Luckenbach et al. 2005 ; Schulte et al. 2009 ).
Use of a size metric derived from a com-mercial harvesting focus (and associated fi shery regulation) is inappropriate if the goal of restoration is to restore ecological function and not a marketable resource (see Breitburg et al. 2000 ; Coen et al. 2004 ; Luckenbach et al. 2005 ; Thayer et al. 2005 ). State agencies have planted shell or relayed oysters to enhance fi sheries for many years (Coen and Luckenbach 2000 ; Coen et al. 2004, 2007, 2010 ), but current large - scale (e.g., Army Corps of Engineers in the Chesapeake Bay) to small - scale community restoration efforts are nearly all targeted at reestablishing the associated ecological value of oyster reefs. Recent reef restoration efforts have also focused on the educational benefi ts derived from the process at the community - based scale and not the numbers of bushels that may be harvested from the reefs at some later time. In the future, restoration efforts need to focus on success criteria that are connected explicitly to the implicit goals of the restoration effort (Coen et al., unpublished data).
Small - or large - scale oyster enhancement/restoration approaches can create tens to hun-dreds of hectares of shellfi sh beds by adding material above an otherwise shell - less, soft - bottom habitat. This may increase settlement of larvae when recruits are not limiting, and can be enhanced by relaying “ cultch ” with juvenile oyster spat that is already attached, or

250 Shellfi sh Aquaculture and the Environment
this role as well as their interactions with and impacts upon other estuarine resources (reviewed in Dumbauld et al. 2009 ). The native oyster Ostrea lurida prefers to live in low intertidal and subtidal areas, where it is less exposed to extreme temperatures and probably formed extensive low relief aggrega-tions before it succumbed to overexploitation and other anthropogenic infl uences (see Fig. 9.3 ; Baker 1995 ; Gillespie 2009 ; White et al. 2009 ). In contrast, Crassostrea gigas tolerates a wider range of temperatures and can with-stand greater turbidity resulting in survival at much higher tidal elevations and over a broader spatial range, from more riverine areas to the estuary mouth (Fig. 9.3 ).
Cultured Crassostrea gigas successfully grow where they have been planted. In loca-tions such as Willapa Bay, they cover expan-sive areas of the upper intertidal zone (over 4625 ha or 20.4% of the bay ’ s intertidal area). Growers typically plant seed at relatively low densities and do not allow individuals to aggregate through thinning clusters in order to achieve optimal growth for later harvesting (Fig. 9.4 C). Nevertheless, this oyster still forms extensive reefl ike habitat in areas where it is left to “ naturalize. ” The resulting reefs are three - dimensionally complex with some verti-cal relief, much like the reefs formed by Crassostrea virginica on the East and Gulf coast estuaries of the United States (Fig. 9.4 G). Oysters are harvested in a 2 - to 4 - year cycle, with harvesting practices themselves also directly infl uencing the environment (Tallis et al. 2009 ). Culture practices for this oyster also involve deploying artifi cial structures within which the deployed oysters are placed in bags on racks, clustered on stakes, or strung along polypropylene ropes supported by PVC stakes in the expansive tidal fl ats (Fig. 9.4 D). These structures can enhance erosion and/or sediment deposition depending on local water fl ow conditions. They also provide numerous attachment sites for other sessile fouling organ-isms (see NRC 2009, 2010 ).
2005 ; Thieltges et al. 2006 ; McKindsey et al. 2007 ; Kochmann et al. 2008 ; Molnar et al. 2008 ; Sousa et al. 2009 ; Markert et al. 2010 ).
On the U.S. West Coast, the introduction of Pacifi c oysters initiated widespread aquacul-ture and farming operations for this species and other bivalves. Recognizing the impor-tance of the shellfi sh industry in the state of Washington, tidelands were deeded directly to growers for use. This encouraged potentially “ sustainable ” farming instead of a traditional fi shery and Washington became the leading shellfi sh producer on the West Coast. Eastern oysters never reproduced successfully and were essentially replaced with the Pacifi c oyster ( Crassostrea gigas ). Juvenile Pacifi c oysters or seed were imported annually from Japan (NRC 2004 ), but this oyster also became natu-ralized in some locations where water tem-peratures were suffi cient for spawning (e.g., Willapa and Dabob Bays in Washington, USA; Pendrell Sound and Ladysmith Harbor in British Columbia, Canada).
Oyster farming operations became inte-grated with the advent of hatchery technology in the early 1980s (Nosho and Chew 1991 ; Conte et al. 1994 ). The Manila clam, Tapes philippinarum , which was inadvertently intro-duced with the Pacifi c oyster, and the much larger native geoduck, Panope generosa , are currently two of the most actively farmed clams on the West Coast. Both species are raised in hatcheries and seeded into estuarine intertidal culture grounds. It should be noted that mussels are also farmed on the West Coast, but here we restrict our discussion to oysters and clams because they co - occur in intertidal estuarine areas and mussels are mostly grown in suspended culture.
The functional role of these cultured inter-tidal bivalves is similar to that of native bivalves in West Coast estuaries, and those described for Eastern oysters and others else-where, that is, they infl uence material pro-cesses and physical structure. Culture practices shape both the spatial and temporal scale of

Impacts of native and cultured bivalves 251
(e.g., netting and tubes; Fig. 9.4 E and 9.4 F; Powers et al. 2007 ; Tallman and Forrester 2007 ), substrate modifi cation (beach gravel-ling), and the recurring disturbance related to planting, maintenance, and harvesting oper-ations (Simenstad and Fresh 1995 ; NRC 2010 ).
Numerous models have been built to assess the carrying capacity of localized systems for bivalve aquaculture (see Chapters 1 and 6 in this book), and where they have been tested
Native littleneck ( Leucoma staminea ) and butter clams ( Saxidomus gigantea ) are also generally found at lower intertidal elevation than the cultured, nonnative Manila clams ( Tapes philippinarum ). All infaunal clams infl uence material processes in a similar way, but in contrast to oysters, they provide signifi -cantly less, physical structure above the sediment. A signifi cant difference in cultured clam operations is the added physical feature provided by predator - exclusion devices
Figure 9.3 Intertidal distribution of eelgrass and oysters in a hypothetical U.S. West Coast estuary in 1800 versus present. Native oysters Ostrea lurida formed low - profi le reefs in low intertidal and subtidal areas in 1800 where native eelgrass Zostera marina was also distributed (top). Pacifi c oysters Crassostrea gigas are now typically spread out in beds slightly higher in the intertidal where they intermingle with native eelgrass and in some cases the introduced eelgrass Zostera japonica at a higher elevation. This is depicted at the estuarine landscape scale (bottom) in an estuary where Pacifi c oysters have become naturalized and also form reefs closer to the head of the estuary where their larvae are retained.
Zostera japonica
Crassostrea gigas
Ostrea lurida
Zostera marina
Eelgrass
Oyster reef
Oyster culture
1800
1800
2010
2010

Figures 9.4 Intertidal habitats and shellfi sh culture in U.S. West Coast estuaries. (A) Open unstructured mudfl at often dominated by burrowing shrimp; (B) structured eelgrass, Zostera marina , habitat; (C) bottom culture of Pacifi c oysters Crassostrea gigas forming structured habitat intermingled with eelgrass; (D) longline culture of Pacifi c oysters; (E) Manila clam Tapes philippinarum culture with predator netting; (F) geoduck Panope generosa culture with tubes for predator protection; and (G) Pacifi c oyster reef where they have become naturalized in Willapa Bay, WA.
(A)
(C) (D)
(F)(E)
(G)
(B)
252

Impacts of native and cultured bivalves 253
(Dumbauld et al. 2009 ). Banas et al. (2007) found that phytoplankton concentrations declined more rapidly with distance from the mouth of Willapa Bay than would be expected due to simple mixing, and that the difference was consistent with the capacity of oysters to fi lter out signifi cant portions of the phyto-plankton community. This is possible because a large portion ( > 80%) of the water never fully circulates over the intertidal fl at, but is exported and reimported into the estuary due to the local complexities of tidal circulation, and the extent of the tidal fl ats in this estuary (Banas et al. 2007 ).
More recent fi eld measurements confi rm that oysters are in fact able to remove > 10% of the phytoplankton that circulated over a 100 - m distance across a tidal fl at versus a control, where no oysters were present (W. Wheat and J.L. Ruesink, unpublished data) . Thus, hydrography and plankton dynamics of the estuarine system must be accounted for when defi ning carrying capacity and quantita-tively characterizing the impact of adding or maintaining bivalve aquaculture. Comparing the historical situation in Willapa Bay with current conditions is also interesting as native Olympia oysters were less abundant or at least represented a lower biomass (i.e., 2.5 × lower than current Crassostrea gigas populations; Ruesink et al. 2005 ) and occupied a different location within the estuary (Fig. 9.3 , lower intertidal and generally further within the estuary). Another unknown component is the abundance of native clams and other fi lter - feeders that were historically present on the tidal fl ats where Crassostrea gigas is currently cultivated. Anecdotal information suggests past densities of native clams were also much higher than current population densities (L. Bennett and R. Sheldon, pers. comm.). Recent anthropogenic nutrient inputs have caused sig-nifi cant eutrophication within many estuaries such as the Chesapeake Bay, making the fi lter-ing capability of bivalves a potentially attrac-tive “ ecosystem service ” (Chapters 8 and 9 in
and verifi ed, results generally suggest that cul-tured bivalves have measurable effects on water properties (e.g., Heral 1993 ; Grant et al. 1995, 2005 ; Leguerrier et al. 2004 ; Dowd 2005 ; Duarte et al. 2005, 2008 ; Drapeau et al. 2006 ; McKindsey et al. 2006 ; Ferreira et al. 2007 ; Fulford et al. 2007, 2010 ; Newell 2007 ; Ferreira et al. 2009 ; North et al. 2010 ; NRC 2010 ). The presence and magnitude of this effect depends on fi ltration ability of the bivalves (i.e., clearance rates of individuals and population size), location, water properties, and residence time of the water mass being fi ltered. These models have rarely been applied to intertidal shellfi sh aquaculture in West Coast estuaries (primarily Oregon and Washington, USA, and Canada) where water properties are largely driven by the coastal ocean and tidal forcing. These estuaries gener-ally have large tidal prisms and relatively short resident times (i.e., are rapidly fl ushed) relative to the fi ltration capacity of the native bivalves present or for nonnative, cultured bivalves, and thus only very localized effects of fi lter feeding have been demonstrated (reviewed in NRC 2010 ).
An exception is Willapa Bay, where the extensive culture of oysters and clams has the potential to regulate phytoplankton produc-tion in the system (Banas et al. 2007 ; Wheat et al. in review ). Hydrography and corre-sponding phytoplankton concentrations in Willapa Bay and in many other West Coast estuaries are different from those of larger and perhaps more typical river - dominated systems on the U.S. East Coast. This results in a gradi-ent of high food concentrations near the mouth of the estuary and lower values in the upper reaches of that estuary. Three competing hypotheses for these West Coast systems could explain this pattern: (1) physical mixing of nutrient - rich oceanic and poorer, river waters; (2) a longer residence time in the upper estuary relative to estuaries on the Gulf or East coasts; or (3) signifi cant grazing by farmed oysters localized near the mouth of these estuaries

254 Shellfi sh Aquaculture and the Environment
shellfi sh and associated structures, as well as the effects of “ pulsed ” disturbances such as harvesting - or maintenance - related activities.
A signifi cant amount of research has been conducted on the effects of adding shellfi sh, particularly oysters, to open, unstructured tidal fl ats in U.S. West Coast estuaries or on SAV such as eelgrass ( Zostera marina ; Dumbauld et al. 2009 ). The direct impact of such activities on eelgrass is a primary concern to management and regulatory agencies because this habitat has been shown to be so important for numerous estuarine fi nfi sh and invertebrates, and because seagrass densities are declining on a global basis (E.L. Jackson et al. 2001 ; Heck et al. 2003 ; Orth et al. 2006 ; Heck et al. 2008 ; Hughes et al. 2009 ; Waycott et al. 2009 ). That said, the ecosystem services of shellfi sh aquaculture, both as a structured habitat and other attendant services, have yet to be examined suffi ciently (but see Harbin - Ireland 2004 ; Brumbaugh and Toropova 2008 ; Beck et al. 2009 ; Dumbauld et al. 2009 ; Forrest et al. 2009 ; NRC 2010 ). However, given time and more rigorous experimental studies, we may see more examples that elucidate these positive ecosystem services for the shellfi sh aquaculture industry, especially given shellfi sh growout sites may also provide optimal growth conditions for eelgrasses by adding suffi cient nutrients in otherwise oligotrophic estuaries or by removing suffi cient seston to improve light conditions in areas too turbid to allow sea-grasses to take hold and thrive (reviewed in NRC 2010 , and references therein).
Although enhanced eelgrass ( Zostera marina ) growth due to enhanced bivalve biode-position has been documented along the U.S. East Coast (cf. Peterson and Heck 2001a, 2001b ; Wall et al. 2008 ), this same positive result does not seem to be the case for what has been observed in aquaculture systems on the West Coast, at least as has been examined to date. In Willapa Bay, the presence of oysters appears to have no effect on eelgrass growth (Ruesink et al. 2009 ; Tallis et al. 2009 ; Wagner
this book; Coen et al. 2007 ; Newell et al. 2007 ; Huang et al. 2008 ). To date, bivalve fi ltration has not been demonstrated as a direct benefi t of aquaculture on the West Coast (Harbin - Ireland 2004 ; NRC 2010 ), in part due to high fl ushing rates, and also the lack of overlap between aquaculture and areas where eutrophication is now becoming an issue. This could change as the services provided by fi lter - feeders become more valuable, allowing aqua-culture to be extended into areas whose waters are currently classifi ed as nonshellfi sh - growing areas (see Brumbaugh and Toropova 2008 ; Beck et al. 2009 , and discussed elsewhere here).
Measurable effects of cultured bivalves on sediment properties depend on the density of the organisms and local system characteristics. Effects are expected to be local, depending on water fl ow, which disperses biodeposits (e.g., Callier et al. 2006 ; Callier et al. 2008 ; Weise et al. 2009 ), and on existing sediment charac-teristics at the aquaculture site (e.g., grain size; see Nizzoli et al. 2006 ; Mesnage et al. 2007 ; see also Chapter 10 ). Intertidal culture of clams and oysters typically occurs in areas with sandier substrates and relatively high fl ow, but it is more diffi cult to quantify the effects of biodeposition and to distinguish it from other sources of sediment deposition and nutrient addition than in suspended culture. This is in part due to the fact that most existing studies are based on suspended culture. Minimal evidence collected to date suggests that the mechanisms are similar to those docu-mented for suspended culture and that high densities of oysters can cause reduced grain size and increased organic content (Rumrill and Poulton 2004 ; E.L. Wagner, unpublished data). Geoduck clams have been shown to raise pore water ammonia concentrations (J.L. Ruesink and K. Rowell, unpublished data). When evaluating the effects of localized biode-position on the surrounding benthic inverte-brate community, it is diffi cult to separate these effects from those of simply adding the

Impacts of native and cultured bivalves 255
2009 ; E.L. Wagner, J.L. Ruesink, and B.R. Dumbauld, unpublished data).
Studies have shown that recruitment of eel-grass seedlings varies by location within Willapa Bay and with different aquaculture methods (Wisehart et al. 2007 ). Seedling recruitment was lowest in areas that had just been dredged, low in longline culture areas, but surprisingly high in dredged areas 1 year later, suggesting fairly robust recovery rates and that competition with adult plants may likely play a role in seedling growth and sur-vival. Evidence suggests that asexual reproduc-tion (rhizome branching), which is important for local - scale recovery from disturbance, was relatively high at most sites, whereas sexual reproduction, which could be important for recovery of larger areas, varied substantially in locations across Washington State (see Ruesink et al. 2009 ). Eelgrass is less abundant overall in aquaculture beds (from 30% to 35% cover; Tallis et al. 2009 ); however, based on either individual shoot or ramet metrics and at the coarser population level, it appears to be able to coexist and survive with oyster aquaculture at multiple sites and over multiple years in Willapa Bay.
Use of U.S. West Coast estuarine habitats by fi sh and invertebrates is much more limited than that published for both the East and Gulf coasts (reviewed in Coen et al. 1999b ; ASMFC 2007 , and papers therein) and appears to depend on spatial scale and organismal mobil-ity. Researchers have generally found abun-dant and diverse communities of both benthic and epibenthic invertebrates associated with on - bottom oyster aquaculture (Fig. 9.4 C), similar to those documented in adjacent natural eelgrass habitats (Fig. 9.4 B). In Pacifi c Northwest estuaries, aquaculture - associated assemblages are also more diverse than those found in open, unstructured mud - or sand - fl at areas, which are often dominated by burrow-ing shrimp (Fig. 9.4 A; Simenstad and Fresh 1995 ; Trianni 1995 ; Hosack et al. 2006 ; Ferraro and Cole 2007 ).
et al., unpublished data), in part because nutri-ents are already close to optimum levels for seagrasses, but also because oysters do not substantially infl uence light levels, which have been shown to be the primary limiting factor for eelgrass in most West Coast estuaries studied (Thom et al. 2008 ). In another study, Japanese littleneck clam ( Tapes philippinarum ) production areas are generally found higher in the intertidal zone than native eelgrass. However, the introduced eelgrass Zostera japonica does interact with the cultured clams in Willapa Bay, WA (Tsai et al. 2010) . There the presence of Zostera japonica reduces clam “ condition ” (or the ratio of meat weight to shell size) by altering water fl ow and thus food supply, but clams have no demonstrable recip-rocal effect on eelgrass growth. Another cul-tured bivalve, the geoduck ( Panope generosa ), does have seasonal effects on eelgrass growth in south Puget Sound, WA. This observed impact may be due to the reduced seasonal density of eelgrass in the presence of clams during the summer and not biodeposition from clams per se (Ruesink and Rowell 2010 ).
Some harvest techniques associated with oyster aquaculture such as harvest - related dredging reduce eelgrass densities directly by removing plant biomass. Additionally, observed seagrass recovery rates vary spatially within these estuaries, with slower rates in areas with softer, muddy sediments (up to 4 years) than for those in areas that have natu-rally sandier (coarser) sediments or for that matter those that are left undisturbed by dredging. Surveys and manipulative dredging studies suggest a gradient of impact from har-vesting (recovery) technique on eelgrass. Ranked from worst to least impact: harvest dredging > narrow - spaced longlines > widely spaced longlines > handpicked ground culture (Tallis et al. 2009 ). Additionally, simply plant-ing oysters at dense concentrations ( > 20% ground cover) will displace some eelgrass and change both cover and density, but results appear to be site specifi c (e.g., Tallis et al.

256 Shellfi sh Aquaculture and the Environment
(e.g., Semmens 2008 ; Dumbauld et al. 2009 ), but these functional associations with inter-tidal benthic habitat, especially shellfi sh aqua-culture habitat, are less studied for other species (reviewed in NRC 2010 ). Structured habitat, such as aquaculture sites, can be used for feeding sites or as protection from larger predators (e.g., Bell et al. 1991 ; Heck and Crowder 1991 ; Coen et al. 1999b ).
As introduced above, both oyster and clam aquaculture involve the addition of signifi cant structures such as longlines, poles, and bags for raising the shellfi sh off - bottom and tubes and netting for predator protection (e.g., Hecht and Britz 1992 ; Everett et al. 1995 ; Kaiser et al. 1998 ; Heise and Bortone 1999 ; Dealteris et al. 2004 ; O ’ Beirn et al. 2004 ; Munroe and McKinley 2007a ; Powers et al. 2007 ; Tallman and Forrester 2007 ). Studies suggest that these affect water fl ow and biode-position, while providing novel attachment sites for organisms, increasing the shading of seagrasses, and infl uencing the behavior of nekton, particularly those that are structure oriented and/or feed on fouling organisms (e.g., pipefi sh, pile perch, kelp surfperch; Everett et al. 1995 ; Simenstad and Fresh 1995 ; Thompson 1995 ; Rumrill and Poulton 2004 ; Weschler 2004 ; Munroe and McKinley 2007a, 2007b ; Whiteley and Bendell - Young 2007 ; D ’ Amours et al. 2008 ; Dumbauld et al. 2009 ).
Bivalves have been actively cultured in many U.S. West Coast estuaries such as Willapa Bay for nearly 100 years or more. At the present scale, shellfi sh aquaculture seems more sustainable than other human activities such as coastal development and pollution, which degrade and can even eliminate estuarine func-tion and potentially undermine resilience (reviewed in Dumbauld et al. 2009 ). Management decisions about how to classify estuaries and maintain ecosystem services as provided by both native and cultured bivalves in West Coast estuaries and other areas should therefore consider both temporal and spatial scales. Cultured Crassostrea gigas have
A special case exists in Willapa Bay and Grays Harbor, WA, where the pesticide carba-ryl (better known as “ Sevin, ” 1 - naphthyl methylcarbamate) is currently used to control two species of burrowing shrimp that are viewed as signifi cant “ pests ” by the shellfi sh aquaculture industry. This practice has been shown to act as a signifi cant, short - term dis-turbance by causing direct mortality to many of the benthic invertebrates present, but having few long - term ( > 60 days) effects, other than removing the shrimp. Burrowing shrimp them-selves may be viewed as “ ecosystem engineers ” because they signifi cantly rework the structure of the associated benthic community via their extensive “ bioturbation ” (e.g., Posey 1986 ; Dumbauld et al. 2001 ). When treated with pesticide, the shrimp - dominated community is then replaced, at least for a culture cycle or two, with above - sediment structure in the form of oysters and even eelgrass (e.g., Dumbauld and Wyllie - Echeverria 2003 ).
Among large, more mobile nekton, some species show loose habitat associations due to the presence of aquaculture operations, while others appear to be uneffected. In Willapa Bay, juvenile Chinook salmon ( Oncorhynchus tshawytscha) and English sole ( Parophrys vetulis ) were found across habitats (eelgrass, oyster aquaculture, and mudfl ats), while other fi nfi sh such as tubesnouts ( Aulorhynchus fl a-vidus ) were clearly associated with eelgrass (e.g., Hosack et al. 2006 ; Dumbauld et al. 2009 ). Separate studies conducted in Willapa Bay and Grays Harbor estuaries have shown that mature Dungeness crab ( Metacarcinus magister ) utilize unstructured muddy areas to feed and rock crab ( Cancer productus ) utilized cultured oysters, while young crab of both species prefer shell deposits and oyster aqua-culture areas over eelgrass and especially unstructured, shrimp - dominated habitat for protection (e.g., Dumbauld et al. 2000 ; Holsman et al. 2006 ). Data from fi eld enclo-sures and laboratory mesocosms suggests that juvenile salmon seek refuge in eelgrass habitats

Impacts of native and cultured bivalves 257
grass. Forty - fi ve percent of the area currently covered by eelgrass was once native oyster habitat (Fig. 9.5 A).
These hypotheses assume that estuarine bathymetry has not changed. Borde et al. (2003) suggest that the area where eelgrass can now grow has increased by 22% through time. The functional value of large, undisturbed eel-grass meadows and unvegetated tidal fl ats versus “ mixed ” landscapes of patchy habitats, including shellfi sh beds with edges and corri-dors, needs to be examined at the appropriate landscape scale. This may be an area where innovative practices and best management practice s ( BMP s) developed by growers in association with scientists can be applied to conserve and even enhance the functional value of these shrinking estuarine habitats.
Associated i mpacts ( p ositive and n egative)
Most mariculture operations remove all or most of the shell, along with the meats derived from the waters in which the bivalves were farmed. On the U.S. West Coast, the aquacul-ture industry operates nearly all bivalve har-vesting operations, with little or no wild stock - based operations, so that shell is most often recycled effi ciently for reuse. In contrast, during most of the nineteenth through twenti-eth centuries a great deal of the Crassostrea virginica shellstock was removed from estuar-ies in the Gulf of Mexico and Atlantic coast of the United States and not necessarily replanted. These operations removed the critical carbonate - based shell habitat (oysters, scal-lops, clams, etc.) that, under pre - European settlement accumulated, dissolved or was reduced through normal taphonomic pro-cesses (discussed in Tevesz and McCall 1983 ; Donovan 1991 ; Andersson et al. 2003 ; Guti é rrez et al. 2003 ; Powell et al. 2006 ; NRC 2010 ). In many ways, harvesting of the entire
assumed a similar role to that played by the native Ostrea lurida prior to human interven-tion, but key differences exist, including plant-ing and harvesting cycles, associated management practices and structures, as well as scale and location of these culture opera-tions in the estuary. While there have been few landscape - level approaches to studies of bivalve shellfi sh aquaculture in West Coast estuaries (but see Carswell et al. 2006 ), we hypothesize about the case for Willapa Bay, WA, where shellfi sh culture operations are relatively extensive at nearly 13% of the total estuarine area (Dumbauld et al. 2009 ).
In comparison, native (Olympia) oysters covered 7.5% of the total estuarine area (based on historical maps; Collins 1892 ), but their extrapolated occurrence was at a lower tidal elevation and distance further from the mouth of the estuary than the majority of the current shellfi sh aquaculture areas (Fig. 9.5 A). Though they likely formed vertical aggregations of “ clustered ” oysters that might be called “ reefs, ” native oysters may have had a similar profi le to current aquaculture beds because individuals and even clusters were much smaller. Thus, the role of cultured bivalves as material processors is potentially similar to that of native oysters (NRC 2009, 2010 ). However, their location closer to the estuary ’ s mouth and higher within the intertidal zone suggests that they may be processing phyto-plankton before it reaches locations further up the estuary where native oysters were once abundant (Fig. 9.5 B). The role of other fi lter - feeders such as native clams and burrowing shrimp, which exclude bivalves at some loca-tions due to their “ bioturbation, ” is unknown but likely fl uctuated over time as well. Cultured oyster habitat overlaps with approximately 43% of eelgrass habitat (mostly native but also nonnative Zostera japonica ). This potentially provides more habitat for recruitment of species, such as juvenile Dungeness crab and English sole ( Parophrys vetulus ), than did native oysters that also overlapped with eel-
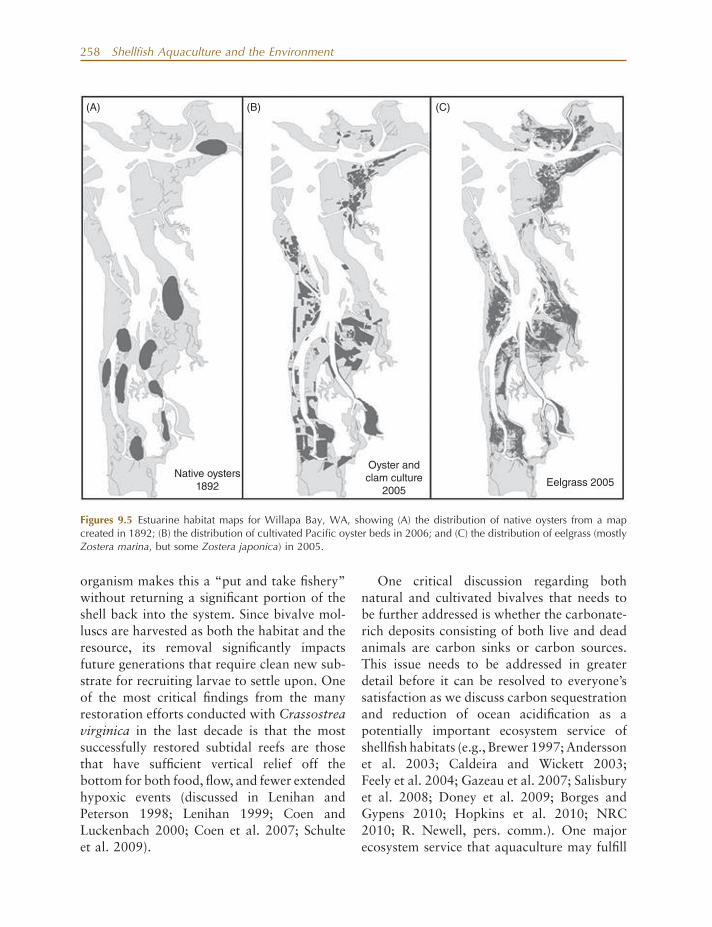
258 Shellfi sh Aquaculture and the Environment
One critical discussion regarding both natural and cultivated bivalves that needs to be further addressed is whether the carbonate - rich deposits consisting of both live and dead animals are carbon sinks or carbon sources. This issue needs to be addressed in greater detail before it can be resolved to everyone ’ s satisfaction as we discuss carbon sequestration and reduction of ocean acidifi cation as a potentially important ecosystem service of shellfi sh habitats (e.g., Brewer 1997 ; Andersson et al. 2003 ; Caldeira and Wickett 2003 ; Feely et al. 2004 ; Gazeau et al. 2007 ; Salisbury et al. 2008 ; Doney et al. 2009 ; Borges and Gypens 2010 ; Hopkins et al. 2010 ; NRC 2010 ; R. Newell, pers. comm.). One major ecosystem service that aquaculture may fulfi ll
organism makes this a “ put and take fi shery ” without returning a signifi cant portion of the shell back into the system. Since bivalve mol-luscs are harvested as both the habitat and the resource, its removal signifi cantly impacts future generations that require clean new sub-strate for recruiting larvae to settle upon. One of the most critical fi ndings from the many restoration efforts conducted with Crassostrea virginica in the last decade is that the most successfully restored subtidal reefs are those that have suffi cient vertical relief off the bottom for both food, fl ow, and fewer extended hypoxic events (discussed in Lenihan and Peterson 1998 ; Lenihan 1999 ; Coen and Luckenbach 2000 ; Coen et al. 2007 ; Schulte et al. 2009 ).
Figures 9.5 Estuarine habitat maps for Willapa Bay, WA, showing (A) the distribution of native oysters from a map created in 1892; (B) the distribution of cultivated Pacifi c oyster beds in 2006; and (C) the distribution of eelgrass (mostly Zostera marina , but some Zostera japonica ) in 2005.
Native oysters1892
Oyster andclam culture
2005Eelgrass 2005
(A) (B) (C)

Impacts of native and cultured bivalves 259
positive effects of bivalve biodeposits on sea-grass “ fertilization ” are more likely to play a role in oligotrophic waters than on those with relatively high available nutrients (Castel et al. 1989 ; Reusch et al. 1994 ; ASMFC 2007 ; Carroll et al. 2008 ; Dumbauld et al. 2009 ; Tallis et al. 2009 ; NRC 2010 ). Mariculture operations generally take place in “ Approved ” waters, away from coastal development and are associated high bacterial and nutrient levels. These waters are approved for direct shellfi sh harvest, that is, where shellfi sh are primarily grown for sale, while restoration for other “ ecosystem services ” such as fi ltration and habitat and broodstock enhancement can take place often in more eutrophic locations (cf. the Nature Conservancy ’ s Long Island Sound and Great South Bay goals; C. LoBue, pers. comm.). Though there are exceptions, on the U.S. East Coast where rivers supply most of the nutrients, mariculture locations occur in more oligotrophic waters where the fertiliza-tion effect could take place, while on the West Coast background nutrients are already high and supplied by the near - shore coastal ocean so the fertilization effect is less likely.
Although many positive interactions have already been discussed for shellfi sh maricul-ture and associated seagrasses, and to a lesser extent macroalgae, many negative impacts or concerns have been expressed by seagrass researchers and permitting agencies charged with the protection of “ potential ” or “ real-ized ” seagrass populations. These have been discussed previously for shellfi sh aquaculture in general (Pillay 1992 ; Kaiser et al. 1998 ; Heffernan 1999 ; Kaiser 2000, 2001 ; Black 2001 ; E.L. Jackson et al. 2001 ; Ruesink et al. 2005, 2006 ; Hosack et al. 2006 ; ASMFC 2007 ; Tallman and Forrester 2007 ; Wisehart et al. 2007 ; Richardson et al. 2008 ; Dumbauld et al. 2009 ; Forrest et al. 2009 ; NRC 2009, 2010 ; Tallis et al. 2009 ) and include (1) co - opting space where seagrasses might other-wise expand their coverage through time (Everett et al. 1995 ); (2) extreme reduction of
is required shell for wild stock and related restoration efforts (NRC 2010 ). Ocean acidi-fi cation may become a serious impediment to potential shell “ stores ” in estuarine waters (Waldbusser et al. 2011 ).
Bivalve aquaculture often has positive effects for the surrounding seagrass communi-ties, enhancing light through reduction in tur-bidity, and adding nutrient - rich biodeposits (reviewed in Haven and Morales - Alamo 1966 ; Luckenbach and Orth 1999 ; Williams and Heck 2001 ; Newell et al. 2002, 2005 ; Mallet et al. 2006, 2009 ; ASMFC 2007 ; NRC 2009, 2010 , although some disagree with this con-clusion; Smith et al. 2009 ). Seagrasses alone can enhance water clarity through removal of suspended sediments as water is baffl ed through blades (e.g., Hemminga and Duarte 2000 ; Beck et al. 2001, 2003 ; Williams and Heck 2001 ; Agawin and Duarte 2002 ; Larkum et al. 2006 ; McGlathery et al. 2007 ). Through their effi cient fi ltering process, bivalve mol-luscs can remove signifi cant portions of all of the total suspended sediments carried in the overlying water column. However, in some estuaries where suspended sediments are low relative to colored dissolved organic matter ( CDOM ), the impact of fi lter - feeding bivalves may have little or no impact on light levels (Corbett 2007 ; L. Coen and E. Milbrandt, pers. obs.). Additionally, seagrass seeds or veg-etative shoots may be entrained directly or indirectly captured in both natural bivalve systems or in and around aquaculture opera-tions where currents are often reduced.
The presence of native bivalve populations also has been shown to have positive effects on submerged vegetation in temperate and subtropical - tropical systems (discussed in Castel et al. 1989 ; ASMFC 2007 ; Dumbauld et al. 2009 ; NRC 2009, 2010 ) where many seagrass - epiphyte communities are phospho-rus and nitrogen limited (e.g., Johnson et al. 2006 ). Shellfi sh may enhance the supply of these nutrients through processes such as biodeposition. It has been suggested that the

260 Shellfi sh Aquaculture and the Environment
tem structure (e.g., Crassostrea Gigas ; see “ Uniqueness of West Coast Aquaculture ” section above; see also Michael and Chew 1976 ; Chew 1990 ; Carlton and Mann 1996 ; Reise 1998 ; Ruiz et al. 2000 ; Black 2001 ; Ruesink et al. 2005 ; Thieltges et al. 2006 ; McKindsey et al. 2007 ; Kochmann et al. 2008 ; Molnar et al. 2008 ; Cohen and Zabin 2009 ; Dumbauld et al. 2009 ; Sousa et al. 2009 ; Markert et al. 2010 ).
Problems related to biodeposits (as fi rst noted and reviewed by Dame 1996 ) have gen-erally been associated with subtidal mussel operations in areas with poor fl ushing rates (e.g., Grant et al. 1995 ; Miller et al. 2002 ; Crawford et al. 2003 ; Cranford et al. 2007 ; Mallet et al. 2009 ; McKindsey et al. 2009 ; Chapter 10 in this book). Rarely have biode-posit accumulations been a problem in inter-tidal growout activities in well - fl ushed areas (e.g., discussed in the “ Uniqueness of West Coast Aquaculture ” section above; Dumbauld et al. 2009 ; NRC 2010 ). Given the extensive umbrella of knowledge under which large - scale aquaculture now operates, site selection criteria and permitting most likely reduce the likelihood of such negative effects (e.g., Tenore and Gonzalez 1975 ; Tenore et al. 1982 ; Dame 1996 ; Wildish and Kristmanson 1997 ; Callier et al. 2008 , but see Deal 2005 ), with most mariculturists striving to operate under more sustainable practices (see Chapter 3 in this book).
Case s tudy: p otential s hort - and l ong - t erm i mpacts of h igh - d ensity i ntertidal h ard c lam a quaculture in s outheastern U . S . t idal c reeks
Background
At present, culture of the northern quahog ( Mercenaria mercenaria ) occurs in more U.S. states than any other native bivalve species under culture. Although market prices fl uctu-ate greatly, overall U.S. market value for these clams is currently among the highest. Direct
fertilization rates caused by cultured bivalve biodeposition (Huang et al. 2008 ) and epi-phytic fouling or excessive macroalgal growth associated with enhanced nutrient availability (e.g., DeCasabianca et al. 1997 ; Hauxwell et al. 2001 ; Munroe and McKinley 2007a, 2007b ; Powers et al. 2007 ); (3) physical dis-turbance from planting and harvesting associ-ated with normal fi shing or mariculture operations (Tallis et al. 2009 ); (4) enhanced competition from biofouling and other species as a result of the introduction of novel hard substrates related to aquaculture operations and deployment or accumulation of dead shell from these extended operations (Beal and Kraus 2002 ; Costa - Pierce and Bridger 2002 ; Orth et al. 2002 ; Erbland and Ozbay 2008 ; Lu and Grant 2008 ; Kimbro et al. 2009 ); (5) changes in both physical and chemical sedi-ment characteristics resulting from changes in fl ow due to the extensive outplanting of aquaculture - related gear (Soniat et al. 2004 ; Kelly et al. 2008 ; see also Fig. 9.4 and the case study in this chapter); (6) past and present spraying of insecticides related to the control of burrowing decapod shrimp populations (e.g., reviewed in Feldman et al. 2000 ; Dumbauld et al. 2001, 2006, 2009 ); (7) intro-duction of direct seagrass competitors ( Zostera japonica , probably the only documented inva-sive seagrass; Harrison and Bigley 1982 ; Posey 1988 ; Baldwin and Lovvorn 1994 ); (8) enhanced structure may cause trophic cascades and negatively affect seagrass populations (e.g., Heck et al. 2000a, 2000b, 2006 ; Duffy et al. 2003 ; Inglis and Gust 2003 ); (9) intro-duction of novel species through the use of oyster shell (discussed in Bushek et al. 2004 ; Cohen and Zabin 2009 ); (10) relocation of shellfi sh stocks and transport of harmful algae (e.g., Carriker 1992 ; H é garet et al. 2008 ; Heil 2009 ; Lewitus and Coen, pers. obs.); (11) impacting access of birds, marine mammals, and other species to areas that would be simple, two - dimensional mudfl ats otherwise ; and (12) potential shifts in dominant ecosys-

Impacts of native and cultured bivalves 261
and indirect ecosystem effects of raising these clams at the elevated densities necessary for commercial success have not been well studied (e.g., Doering et al. 1986 ; Nugues et al. 1996 ; Cerrato et al. 2004 ; Tyler 2007 ). Some studies suggest that elevated bivalve densities can have important impacts on many of the above - mentioned system attributes (e.g., Cloern 1982 ; Offi cer et al. 1982 ; Cohen et al. 1984 ; Asmus and Asmus 1991, 2005 ; Dame, 1993 , 1996 ; Dame and Libes 1993 ; Newell 2004 ; ASMFC 2007 ; Cerco and Noel 2007 ). This species is typically grown in shallow subtidal or intertidal seafl oor areas that are highly visible, and often adjacent to seagrass habitats, which impacts site selection, local permitting, and confl icts over seagrass issues (NRC 2009, 2010 ). Intertidal aquaculture areas with tidal ranges ≥ 1 – 2 m are complex benthic systems and diffi cult to study since sediments are subject to daily aerial exposure where micro-phytobenthos respond to light and other cues. For example, intertidal marine sediments are “ biostabilized ” by diatom fi lms within the surface (Stal 2010 ).
Although not the primary goal of a past research effort in South Carolina, one objec-tive was to examine the direct and indirect effects of high - density hard clam mariculture on the adjacent inshore benthic ecosystem (Coen et al. 2000 ). Results showed that the deployment of hundreds to thousands of clam culture pens, each with tens of thousands of clams, had the potential to affect (1) local hydrodynamics, (2) sediment characteristics, (3) associated benthos, (4) food quality and quantity, and (5) the carrying capacity of the surrounding ecosystem. At that time, hard clam culture occurred on low intertidal mud-fl ats in small tidal creeks where dense oyster reefs ( Crassostrea virginica ) co - occurred (Coen et al. 2000 ; Judge et al. 2000 ).
In addition to clam growth experiments, several other experimental fi eld studies were conducted to (1) evaluate food and fl ow regimes and the effects of localized food deple-
tion on observed clam growth; (2) examine how clam culture pens and related activities directly and indirectly affect ecosystem attri-butes of tidal creeks (e.g., C/N ratios, total organic carbon [TOC], sediment % organics, sediment grain size); and (3) examine how clam culture might affect creek communities (e.g., water column and benthic chlorophyll a , infauna, and stable isotope ratios of deployed seed clams over time).
The study site was located just off the Kiawah River, Kiawah, SC (32 ° 37.41 ′ N, 80 ° 06.60 ′ W; Fig. 9.6 ) in an intertidal creek with shallow mudfl ats ( < 0.5 m in depth during low tide), interspersed with intertidal oysters on either side of the creek bank. Experimental plots were adjacent to and within a large commercial operation with approximately 600,000 clams in 400 circular predator - exclusion pens (0.30 - m high, diameter = 2.44 m, area = 4.68 m 2 ). They were typically exposed during mean low tide. At the site overall water depths at high tide often exceeded 1.8 m or more. Free stream tidal (fl ow) rates were typi-cally high, driven by a 100% change every 12 h. Pens were approximately 0.5 – 2.5 m from one another within a given area. Densities of newly outplanted seed clams (5 – 10 mm in shell length) were approximately 4274 clams m − 2 (approximately 20,000 individuals per pen). At a size of around 20 mm, clams were typi-cally thinned out to 855 clams m − 2 (4000 indi-viduals per pen). During the study, the larger overall area often had over 41 million clams in 2300 pens, with other areas having up to 17 million additional clams in over 800 pens. Overall, Atlantic LittleNeck ClamFarms, Inc. (referred to subsequently as “ ALC ” ) at one time had more than 7000 pens in South Carolina waters, with over 70 million clams planted within them (Judge et al. 2000 ; J. Manzi, pers. comm., 1997).
This study focused on four locations with increasing distance from the above ALC site: (1) between and within pens; (2) 1 – 50 m away (three sites seaward and three sites

262 Shellfi sh Aquaculture and the Environment
Effects on s ediment and i nfauna
Sediment attributes were impacted by the pres-ence of the pens, but the effects tended to be evident only within pens (Figs. 9.7 and 9.8 ). Even at short distances from the pens, sedi-ment changes were not evident or predictable. Sediment organics (% at ignition), % silt, and
downstream); (3) a control site at the creek entrance that was 340 m upstream of the above ALC site; and (4) a reference site without any clam aquaculture. At locations 1 and 2 (listed above), small experimental cages (0.25 m 2 , with 6.4 - mm mesh) were also deployed to examine clam growth at three different densities.
Figure 9.6 Large - scale hard clam growout in South Carolina. Note hundreds of pens at one site near Kiawah, SC (each 4.68 m 2 in area).
Figure 9.7 Small experimental cages (0.25 m 2 , with 6.4 - mm mesh) adjacent to clam pens (see also Fig. 9.6 ).
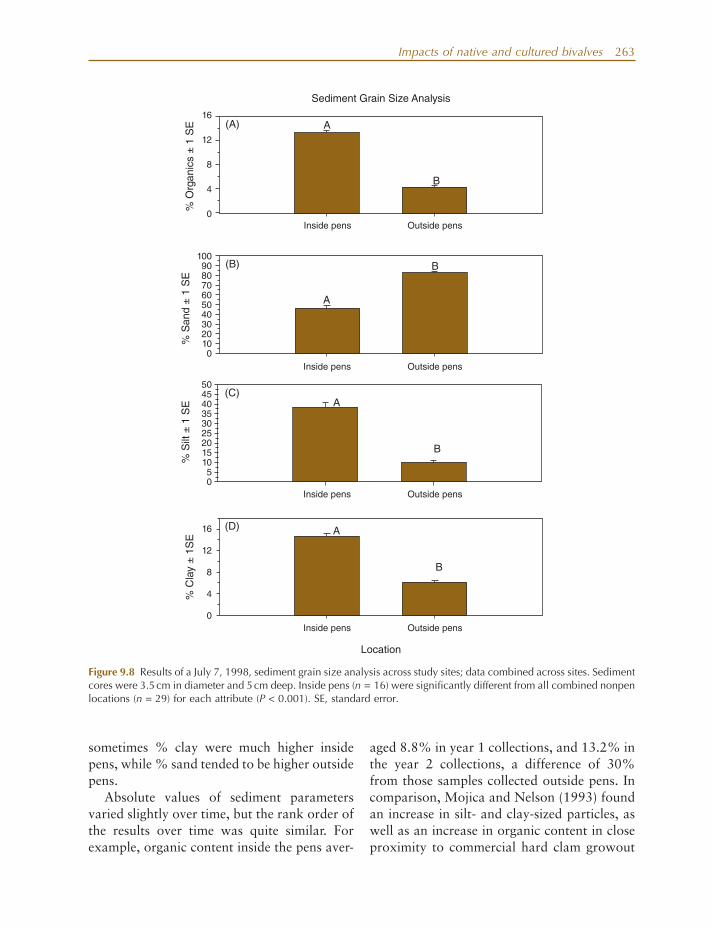
Impacts of native and cultured bivalves 263
aged 8.8% in year 1 collections, and 13.2% in the year 2 collections, a difference of 30% from those samples collected outside pens. In comparison, Mojica and Nelson (1993) found an increase in silt - and clay - sized particles, as well as an increase in organic content in close proximity to commercial hard clam growout
sometimes % clay were much higher inside pens, while % sand tended to be higher outside pens.
Absolute values of sediment parameters varied slightly over time, but the rank order of the results over time was quite similar. For example, organic content inside the pens aver-
Figure 9.8 Results of a July 7, 1998, sediment grain size analysis across study sites; data combined across sites. Sediment cores were 3.5 cm in diameter and 5 cm deep. Inside pens ( n = 16) were signifi cantly different from all combined nonpen locations ( n = 29) for each attribute ( P < 0.001). SE, standard error.
% O
rgan
ics
± 1
SE
% S
and
± 1
SE
% S
ilt ±
1 S
E%
Cla
y ±
1S
E
16
12
8
4
0
1009080706050403020100
50454035302520151050
16
12
8
4
0
Sediment Grain Size Analysis
(A)
(B)
(C)
(D)
Inside pens Outside pens
Outside pensInside pens
Outside pensInside pens
Outside pensInside pens
Location
A
A
A
A
B
B
B
B

264 Shellfi sh Aquaculture and the Environment
proportion of silts or clays and organic matter as reported by Light and Beardall ( 1998 , see also Cahoon et al. 1999 ).
Effects on p hytoplankton a bundance
Concentrations of chlorophyll a in the water column were used as a surrogate for food quantity. Water samples were collected at slack high tide at all of the study sites at approxi-mately monthly intervals using a horizontal Wildco water sampler, then fi ltered following standard fl uorometric methods for quantifying chlorophyll a ( μ g L − 1 ; see Coen et al. 2000 ; Grizzle et al. 2006, 2008 ). Water column chlo-rophyll a levels among sites increased and decreased with seasonal spring and summer phytoplankton blooms. Highest chlorophyll a levels were observed during the summer at all sites. Despite small - scale spatial variability, no consistent differences among adjacent loca-tions were found.
In order to determine within - site effects on variability of suspended food, the potential for depletion of water column phytoplankton at four different spatial scales was assessed by collecting water from different locations simul-taneously within our fi eld site using a vertical sampling apparatus or “ sipper ” (Judge et al. 1993 ). Experiments included (1) food deple-tion (or enhancement) as a parcel of water traveled the entire creek length from the control (creek entrance) site to within the center of the pens (340 m downstream) and as sampled at fi xed heights above the substrate (1, 5, 15, and 45 cm); (2) food depletion as a parcel of water traveled 150 m across the entire ALC clam pen site (over approximately 400,000 clams); (3) food depletion as a parcel of water traveled through an individual clam pen housing approximately 3000 clams (sampled 1 m upstream from a pen and within that same pen); and fi nally, (4) small - scale (ca. 50 cm) food depletion, sampling over our smaller growth cages at densities of 5, 50, and 200 clams per 0.25 - m 2 cage (Coen et al. 2000 ; Judge et al. 2000 ).
bags. The shift in particle size may affect clam growth indirectly, as Grizzle (1988) noted that growth was slowest in fi ner sediments and fastest in coarser, sand - dominated sediments. Although nutrient characteristics of the sedi-ments were not examined, investigators have found both positive and negative impacts from various shellfi sh farming practices (e.g., Holmer 1991 ; Gilbert et al. 1997 ).
Within a given location, no small - scale ( < 50 cm) differences in total benthic biomass were detected between insides and outsides of pens. To examine larger - scale differences in total benthic abundance, data were pooled to compare among locations. One - way analysis of variance (ANOVA) tests revealed no signifi -cant differences among locations ( P = 0.58, degrees of freedom [d.f.] = 38). Linear regres-sion analysis, examining the relationship between total biomass and distance from pens, also revealed no signifi cant relationship ( r 2 = 0.02, P = 0.43). Although total biomass of benthos does not appear to be related to proximity to pens, difference in the abundance of faunal taxa may be infl uenced by the pres-ence of pens or associated changes in local microhabitat (e.g., sediment type or organic content, fl ow), but they were beyond the scope of this project (see Weston 1990 ; Hammel and Webb 1999 ).
Chlorophyll a levels of intertidal sediment were several orders of magnitude higher than water column values. Results from both sedi-ment chlorophyll samplings revealed signifi -cantly higher levels of chlorophyll a inside versus outside the pen area. Levels were approx-imately 20% higher inside pens than outside pens in May (6000 μ g mL − 1 sediment) and 50% higher in July (11,000 μ g mL − 1 samples). This difference corresponds to the time period during which site water column temperature and salinity were highest and dissolved oxygen levels were lowest. Seasonal variability in water column chlorophyll levels was observed with chlorophyll peaking in May, probably a result of initial spring blooms. Overall, levels of chlo-rophyll were highest in sediments with a high
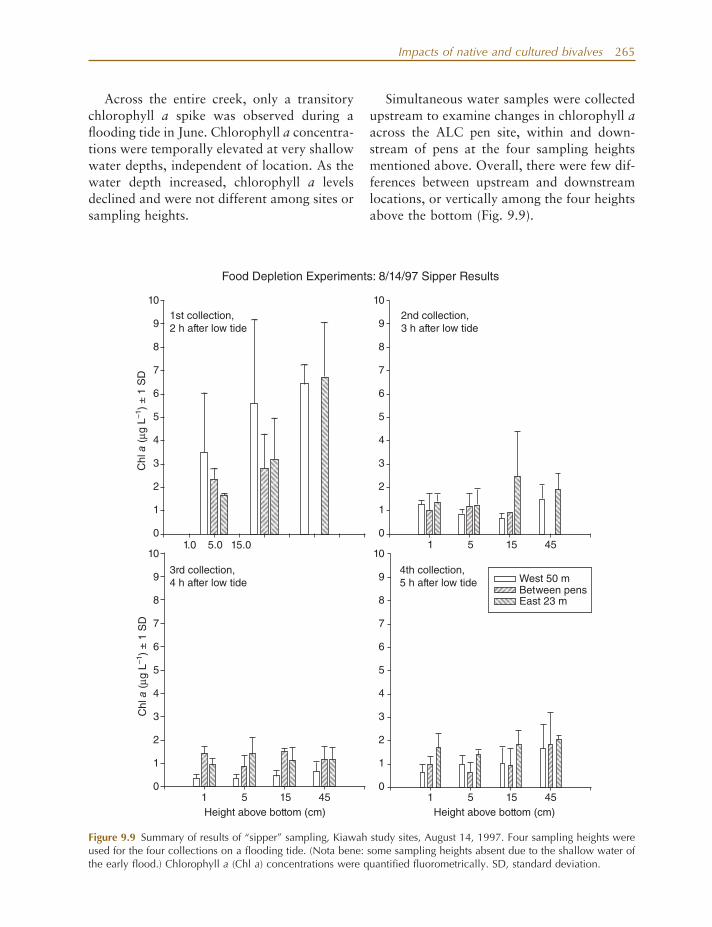
Impacts of native and cultured bivalves 265
Simultaneous water samples were collected upstream to examine changes in chlorophyll a across the ALC pen site, within and down-stream of pens at the four sampling heights mentioned above. Overall, there were few dif-ferences between upstream and downstream locations, or vertically among the four heights above the bottom (Fig. 9.9 ).
Across the entire creek, only a transitory chlorophyll a spike was observed during a fl ooding tide in June. Chlorophyll a concentra-tions were temporally elevated at very shallow water depths, independent of location. As the water depth increased, chlorophyll a levels declined and were not different among sites or sampling heights.
Figure 9.9 Summary of results of “ sipper ” sampling, Kiawah study sites, August 14, 1997. Four sampling heights were used for the four collections on a fl ooding tide. (Nota bene: some sampling heights absent due to the shallow water of the early fl ood.) Chlorophyll a (Chl a ) concentrations were quantifi ed fl uorometrically. SD, standard deviation.
10
9
8
7
6
5
4
3
2
1
0
10
9
8
7
6
5
4
3
2
1
0
10
9
8
7
6
5
4
3
2
1
0
10
9
8
7
6
5
4
3
2
1
0
Food Depletion Experiments: 8/14/97 Sipper Results
1st collection,2 h after low tide
2nd collection,3 h after low tide
3rd collection,4 h after low tide
4th collection,5 h after low tide West 50 m
Between pensEast 23 m
1.0 5.0 15.0
1 5 15 45 1 5 15 45
1 5 15 45
Height above bottom (cm) Height above bottom (cm)
Chl
a (
μg L
–1)
± 1
SD
Chl
a (
μg L
–1)
± 1
SD

266 Shellfi sh Aquaculture and the Environment
Water fl ow speed measurements were gen-erally conducted on fl ooding tides, occasion-ally extending into the early ebb tide. On each sampling date, up to 13 heights above the bottom were sampled to characterize the benthic boundary layer conditions. These ver-tical velocity profi les allowed for the genera-tion of shear velocity ( u * ) calculations via logarithmic profi ling (Eq. 9.1 ) and Reynolds shear stress (Eq. 9.2 ) methods. When the fl ow direction varied within a benthic boundary layer profi le (e.g., adjacent to obstruction created by the pens), only the Reynolds shear stress method could be calculated.
u s u k s d so o( ) = −( )[ ]∗ / ln / (9.1)
u u wR R∗ = [ ] = ′ ′τ ρ τ ρ/ ,.0 5 where (9.2)
Benthic boundary layer sampling was con-ducted at two locations (340 m and 50 m) upstream of the ALC study site for 3 h during a fl ooding tide. Maximum near - surface water fl ow speeds approached or exceeded 30 cm s − 1 at both locations. Vertical velocity profi les were constructed at the control creek entrance site and at a location 50 m away. At both loca-tions, fl ow direction did not vary with sam-pling height above the bottom. Shear velocities ( u * ) for the control creek location at 5 cm above the bottom (Reynolds shear stress method) ranged from 1.205 cm s − 1 (initial fl ood) to 1.715 cm s − 1 (peak fl ood). Shear velocities ( u * ) at the site 50 m upstream (log profi le method) ranged from 0.306 cm s − 1 (late fl ood) to 1.557 cm s − 1 (peak fl ood). Benthic boundary layer sampling between the pens (i.e., within the ALC study site) was conducted on three occasions for a total of 5.5 h during two fl ooding tides and an additional 6.5 h from the initial fl ood through early ebb tide. Maximum near - surface water approached 20 cm s − 1 at this site. Vertical velocity profi les were constructed for three components of the tidal cycle, including peak fl ood. At this loca-tion, fl ow direction varied dramatically with
A second experiment, conducted in May, quantifi ed chlorophyll a concentrations (1) within a single pen; (2) in the pen area; and (3) upstream of pens. As before, no consistent depletion of chlorophyll a concentrations was observed. A third comparison of food levels within a pen and an area 1 m upstream of that pen was run in June and only a transitory elevation of chlorophyll a was documented during the initial fl ood. In general, no sus-tained reduction within a pen or among mul-tiple sampling heights was observed. A fourth comparison, based on samples collected in June, measured potential food reduction across the smaller experimental cages (5, 50, or 200 clams per 0.25 m 2 ), and similar to the experi-ments described above results exhibited no upstream versus downstream differences. In all cases, no consistent depletion of food levels arose at any of these scales, with the exception of the transitory elevation during the initial fl ood, most likely due to a resuspension of surfi cial sediments and benthic microalgae (Coen et al. 2000 ; Judge et al. 2000 ).
Effects on w ater fl ow
Flow regimes in aquatic systems can be complex, especially when associated with three - dimensional structures (reviewed in Nowell and Jumars 1984 ; Denny 1988 ; Denny and Wethey 2001 ). Water fl ow in the fi eld was measured at two temporal scales. First, a Sontek (YSI, Inc., San Diego, CA) acoustic Doppler velocimeter ( ADV ; fi eld model) was used to characterize current speed and direction at our fi eld site via high frequency ( ≤ 25 MHz) readings of water fl ow and vertical mixing ( x - , y - , z - axes). Current velocity was measured inside individual clam pens, aisles between pens, and at various dis-tances upstream of pens. The ADV sampled variations in water fl ow speed along each dimensional axis up to 25 per second, within a small sampling cylinder (9 - mm length × 6 - mm diameter) whose height above the substratum could be positioned by the experimenter.
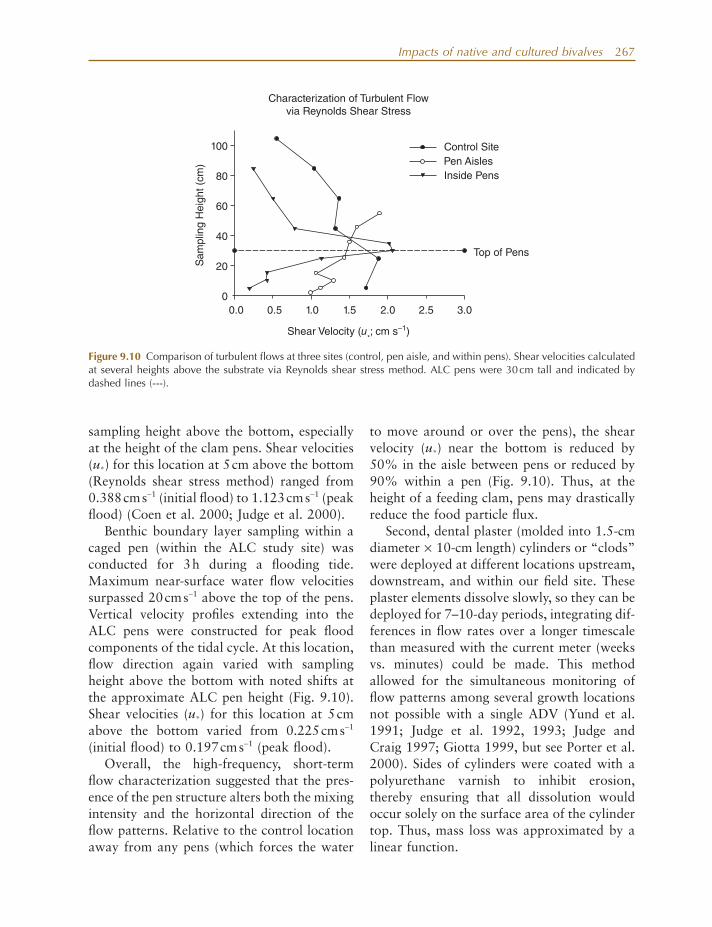
Impacts of native and cultured bivalves 267
to move around or over the pens), the shear velocity ( u * ) near the bottom is reduced by 50% in the aisle between pens or reduced by 90% within a pen (Fig. 9.10 ). Thus, at the height of a feeding clam, pens may drastically reduce the food particle fl ux.
Second, dental plaster (molded into 1.5 - cm diameter × 10 - cm length) cylinders or “ clods ” were deployed at different locations upstream, downstream, and within our fi eld site. These plaster elements dissolve slowly, so they can be deployed for 7 – 10 - day periods, integrating dif-ferences in fl ow rates over a longer timescale than measured with the current meter (weeks vs. minutes) could be made. This method allowed for the simultaneous monitoring of fl ow patterns among several growth locations not possible with a single ADV (Yund et al. 1991 ; Judge et al. 1992, 1993 ; Judge and Craig 1997 ; Giotta 1999 , but see Porter et al. 2000 ). Sides of cylinders were coated with a polyurethane varnish to inhibit erosion, thereby ensuring that all dissolution would occur solely on the surface area of the cylinder top. Thus, mass loss was approximated by a linear function.
sampling height above the bottom, especially at the height of the clam pens. Shear velocities ( u * ) for this location at 5 cm above the bottom (Reynolds shear stress method) ranged from 0.388 cm s − 1 (initial fl ood) to 1.123 cm s − 1 (peak fl ood) (Coen et al. 2000 ; Judge et al. 2000 ).
Benthic boundary layer sampling within a caged pen (within the ALC study site) was conducted for 3 h during a fl ooding tide. Maximum near - surface water fl ow velocities surpassed 20 cm s − 1 above the top of the pens. Vertical velocity profi les extending into the ALC pens were constructed for peak fl ood components of the tidal cycle. At this location, fl ow direction again varied with sampling height above the bottom with noted shifts at the approximate ALC pen height (Fig. 9.10 ). Shear velocities ( u * ) for this location at 5 cm above the bottom varied from 0.225 cm s − 1 (initial fl ood) to 0.197 cm s − 1 (peak fl ood).
Overall, the high - frequency, short - term fl ow characterization suggested that the pres-ence of the pen structure alters both the mixing intensity and the horizontal direction of the fl ow patterns. Relative to the control location away from any pens (which forces the water
Figure 9.10 Comparison of turbulent fl ows at three sites (control, pen aisle, and within pens). Shear velocities calculated at several heights above the substrate via Reynolds shear stress method. ALC pens were 30 cm tall and indicated by dashed lines ( - - - ).
Characterization of Turbulent Flowvia Reynolds Shear Stress
Shear Velocity (u*; cm s–1)
Sam
plin
g H
eig
ht (c
m)
100
80
60
40
20
0
0.0 0.5 1.0 1.5 2.0 2.5 3.0
Control Site
Pen Aisles
Inside Pens
Top of Pens

268 Shellfi sh Aquaculture and the Environment
Long - term patterns differences in water fl ow among locations where clam growth was also measured were estimated by dissolution of dental plaster cylinders. A total of 270 clods were deployed at multiple locations upstream, downstream, and within our fi eld site, includ-ing within clam pens. Replicate clod cards were always deployed in pairs, glued with sili-cone cement atop a rectangular PVC plate (10 cm × 5 cm) and positioned 5 cm above the substrata. After retrieval, cylinders were removed from their holders, rinsed of sedi-ment, dried at 60 ° C for 48 h, allowed to accli-mate to ambient lab temperature and humidity for 48 h, and reweighed for mass lost (Coen et al. 2000 ; Judge et al. 2000 ).
Variability in fl ow among locations (Fig. 9.11 ) were analyzed as an incomplete block ANOVA with deployment date as the blocking variable and mass lost as the dependent vari-able. Overall, there was a signifi cant difference in mass loss among locations (ANOVA, P < 0.001), although several intermediate locations overlapped in plaster loss. The three locations common to all (control, 50 m, and between pens), food supply, sediment analysis, and ADV measurements proved to be signifi -cantly different (Coen et al. 2000 ; Judge et al. 2000 ).
Moreover, the rank orders of these sites both qualitatively and quantitatively matched the shear velocity differences illustrated by the ADV. In addition, the plaster dissolution within pens exhibited a 25 – 33% reduction relative to outside (ANOVA, P < 0.001), dem-onstrating the extent of fl ow speed loss by the cage mesh itself. The use of the ADV and the plaster cylinders allowed water fl ow patterns to be quantifi ed at two different temporal scales (minutes vs. weeks) and both measures exhibited strong concordance. Both current meter and clod card data revealed that clam farm pens signifi cantly altered fl ow (and other site characteristics) within the tidal creek.
Overall, the structural presence of cages imparted profound changes in the hydrody-
namic regimes within and around clam pens, thereby altering numerous sediment attributes (i.e., grain size and chlorophyll a concentra-tions). Cage - induced mixing caused a localized decoupling of the benthic boundary layer, which can dramatically affect the temporal variation of resuspended algal food supplies. Sediments generally were enriched in the organic fraction, but lower in sand, silt, and clay adjacent to the aquaculture pens. Sediment chlorophyll a (an indirect metric for food quantity) was signifi cantly enhanced within the pens. Despite differences in mixing and sediment attributes induced by the clam pens, however, total sediment infaunal biomass (exclusive of Mercenaria ) was not adversely impacted by high - density clam pens. Availability of suspended food varied among sampling dates, but there was little evidence for food depletion within the water column or across the growth cages. The only regular pattern was a transitory chlorophyll a spike via resuspension during the initial stages of a fl ooding tide (i.e., water depths less than 20 cm). Hydrodynamic patterns (measured at timescales from seconds to weeks) affecting delivery of suspended food showed that clam farm pens reduced mean water speed and mixing by an order of magnitude within a cage, while the caging material itself created localized regions of intense shear stress (Coen et al. 2000 ; Judge et al. 2000 ).
Longer - t erm and l arger - s cale c oncerns
The case study was completed in 2000 at which time ALC had deployed more than 7000 pens on local South Carolina mudfl ats, with over 7 × 10 7 clams planted among them (Coen et al. 2000 ; ALC, pers. comm., 1997). The relevant fi ndings included differences in sediment inside versus outside of growout pens, with interiors generally enriched in the organic fraction, but with lower coarse sands
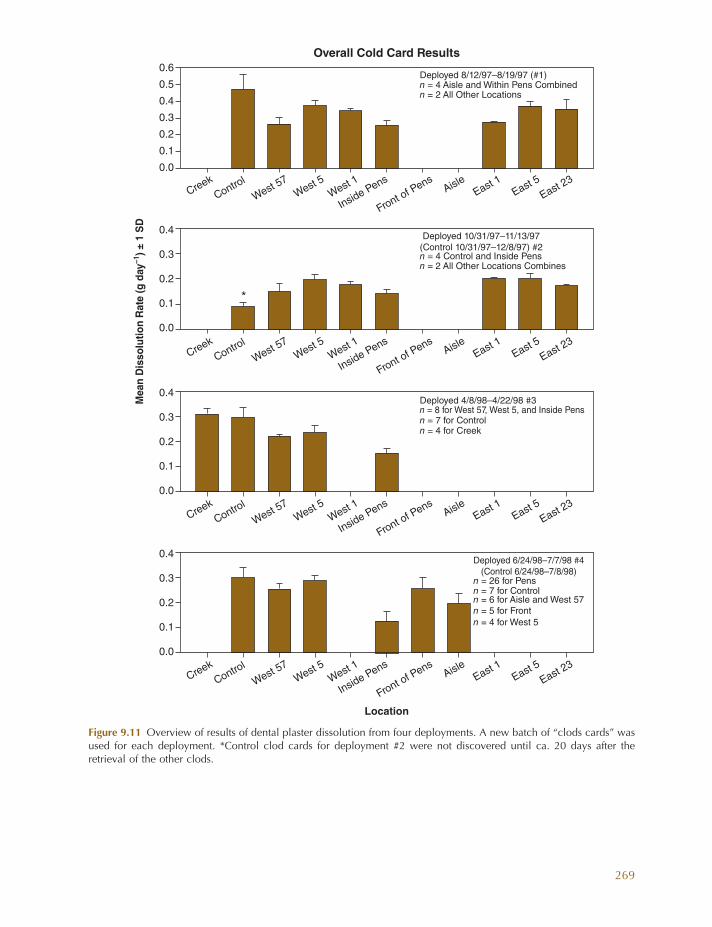
Figure 9.11 Overview of results of dental plaster dissolution from four deployments. A new batch of “ clods cards ” was used for each deployment. * Control clod cards for deployment #2 were not discovered until ca. 20 days after the retrieval of the other clods.
0.6
0.5
0.4
0.3
0.2
0.1
0.0
Creek
Control
West 57
West 5
West 1
Inside Pens
Front of P
ensAisle
East 1East 5
East 23
Creek
Control
West 57
West 5
West 1
Inside Pens
Front of P
ensAisle
East 1East 5
East 23
Creek
Control
West 57
West 5
West 1
Inside Pens
Front of P
ensAisle
East 1East 5
East 23
Creek
Control
West 57
West 5
West 1
Inside Pens
Front of P
ensAisle
East 1East 5
East 23
Overall Cold Card Results
Deployed 8/12/97–8/19/97 (#1)
Deployed 10/31/97–11/13/97
(Control 10/31/97–12/8/97) #2
Deployed 4/8/98–4/22/98 #3
Deployed 6/24/98–7/7/98 #4(Control 6/24/98–7/8/98)
n = 4 Aisle and Within Pens Combinedn = 2 All Other Locations
n = 4 Control and Inside Pens
n = 8 for West 57, West 5, and Inside Pensn = 7 for Controln = 4 for Creek
n = 26 for Pensn = 7 for Control
n = 5 for Front
n = 4 for West 5
n = 6 for Aisle and West 57
n = 2 All Other Locations Combines
0.4
0.3
0.2
0.1
0.0
0.4
0.3
0.2
0.1
0.0
0.4
0.3
0.2
0.1
0.0
*
Mea
n D
isso
lutio
n R
ate
(g d
ay–1
) ± 1
SD
Location
269

270 Shellfi sh Aquaculture and the Environment
habitat for nontarget species (e.g., oysters, mussels; see Figs. 9.12 and 9.13 ; see also Costa - Pierce and Bridger 2002 ; Dealteris et al. 2004 ; Munroe and McKinley 2007a ; Powers et al. 2007 ; D ’ Amours et al. 2008 ; Erbland and Ozbay 2008 ; Dumbauld et al. 2009 ). The maintenance and ultimate removal of such materials presents an additional concern as does the possible impacts of excess nutrients related to death and decay of the associated fouling organisms removed in situ and left to decompose.
In 2004, the South Carolina Department of Natural Resources (SCDNR) (through L. Coen) provided a research plan to address questions related to the remaining 4000, 8 - ft - diameter pens abandoned in South Carolina ’ s waters. Given that insuffi cient escrow funds were set aside with the two South Carolina state agencies (SCDNR and the South Carolina Department of Health and Environmental Control [SCDHEC]) tasked with pen removal and related materials (Figs. 9.12 and 9.13 ) in the event of a fi nancial failure, these two agen-cies were required to determine how to remove all of the remaining abandoned pens and asso-ciated materials. The plaintiff suggested that they be left as functioning critical habitat for numerous fi sh and invertebrate species. The abandonment of aquaculture - related gear is a potentially major problem and one that needs to be addressed more explicitly (see Fig. 9.14 ).
and more silt - clay fractions versus areas adja-cent to the aquaculture pens. Disturbance of mudfl ats during sampling, removal of pens, human disturbance (e.g., noise, viewscape impacts), or other resident organisms had been expressed as a potential concern (see discus-sions and reviews by Brenchley 1981 ; Mojica and Nelson 1993 ; Addessi 1994 ; Brosnan and Crumrine 1994 ; Burger 1994 ; Hall 1994 ; Rodgers and Smith 1997 ; DeGrave et al. 1998 ; Dernie et al. 2003 ; Yasue 2005 ; Kimbro and Grosholz 2006 ; Becker et al. 2009 ; NRC 2009, 2010 ). Although a lot of work has been expended on the impacts of predators (e.g., rays, Orth 1975 ), other disturbances or bio-turbation (e.g., Brenchley 1981 ; Hall 1994 ; Dahlgren et al. 1999 ), and human events (i.e., fi shing; Glude and Landers 1953 ; Godcharles 1971 ; Mojica and Nelson 1993 ; Addessi 1994 ; Brosnan and Crumrine 1994 ; Coen 1995 ; Brown and Wilson 1997 ; Eckrich and Holmquist 2000 ; Kaiser 2000, 2001 ; Neckles et al. 2005 ; Forrest et al. 2009 ; Macleod et al. 2009 ), little direct research has simulated and assessed the impacts related to intertidal or subtidal shellfi sh culture (NRC 2009, 2010 ).
As discussed above and elsewhere, growout structures and gear are generally cleaned or removed on an annual or multiyear basis. If left exposed over longer timescales (years) without attention, the cage pens themselves might provide additional hard substrate
Figure 9.12 Abandoned cage pens left out for over six years near Kaiwah, SC. These intertidal substrates were heavily fouled by Crassostrea virginica and other bivalve species.

Figure 9.13 Abandoned cages and other pen parts (mesh, PVC, etc.) left on the leased mudfl ats in South Carolina, USA.
Figure 9.14 High - resolution aerial imagery of an area near Kiawah, SC (lease M194). Clam pens (see lower - altitude and ground - level images, Figs. 9.6 and 9.12 ) were removed in 2004. Note the cage impacts still recognizable (delineated by red lines) years after their removal. (Courtesy of K. Schulte, SCDNR and GeoVantage and PhotoScience, Inc.)
271

272 Shellfi sh Aquaculture and the Environment
(3) development of necessary markets for attendant shellfi sh services; and (4) related nonmarket challenges (e.g., Freeman 1995 ; Costanza et al. 1997 ; Daily 1997 ; Meyer et al. 1997 ; Heal 2000 ; Meyer and Townsend 2000 ; Farber et al. 2002 ; Millennium Ecosystem Assessment 2003 ; Peterson et al. 2003 ; Cerrato et al. 2004 ; Lipton 2004 ; Newell 2004 ; NRC 2004 ; Newell et al. 2005 ; Cerco and Noel 2007 ; Grabowski and Peterson 2007 ; Brumbaugh and Toropova 2008 ; Wei et al. 2009 ).
The continued use and expansion of BMPs (see Chapter 3 ) in the fi rst, second, and third world, coupled with the application of research fi ndings, will better focus shellfi sh mariculture on (1) where it can be commercially and eco-logically sustainable; (2) site selection; (3) appropriate densities; (4) multiuse operations; and (5) where restoration using scaled - up aquaculture may be applied to areas where harvesting cannot occur as a result of admin-istrative closures, (e.g., initiatives in places such as New York City) but where “ farmed ” species on an industrial scale may be used to treat eutrophication (e.g., Lindahl et al. 2005 ; Gren et al. 2009 ; Chapter 8 in this book). When one compares fi nfi sh aquaculture, which requires large inputs of feed often with associ-ated negative impacts (e.g., Grant et al. 1995 ; Hastings and Heinle 1995 ; Kaiser et al. 1998 ; Heffernan 1999 ; Kaiser 2000, 2001 ; Crawford et al. 2003 ; Forrest et al. 2009 ; NRC 2010 ), use of bivalve (especially oyster) aquaculture is generally considered to be more sustainable, itself requiring excellent water quality, often with few demonstrable negative impacts at this time (e.g., Pillay 1992 ; Rice 2000 ; Read and Fernandes 2003 ; Newell 2004 ; Whiteley and Bendell - Young 2007 ; Dumbauld et al. 2009 ; NRC 2009, 2010 ). There is less consensus, however, among researchers on the use of non-native species regardless of one ’ s following of the International Council for the Exploration of the Sea ( ICES ) protocols (e.g., Simenstad and Fresh 1995 ; Trianni 1995 ; Rumrill and
Remaining q uestions
Aquaculture will continue to increase world-wide while native “ wild stock ” populations decline to near unsustainable levels (Beck et al. 2009, in press ). Living shoreline projects and native shellfi sh restoration projects conducted to provide one or more of the ecosystem ser-vices discussed above will also continue to expand, especially given sea level rise and the search for more ecofriendly approaches. One question that needs to be further addressed is whether structure alone is suffi cient to enhance biodiversity around either natural biogenic shellfi sh reefs with multiple age classes of mol-luscs (e.g., Luckenbach et al. 2005 ), or whether the act of substrate placement alone is suffi -cient? Recent work with intertidal reefs sug-gests that signifi cant resident and transient associates (e.g., Harding and Mann 1999, 2001 ; Coen et al. 2000 ; Coen et al. 2006 ) are attracted to shell even without live recruited oysters (Coen et al. 2006 , unpublished data).
More is known about the impacts of water column - based aquaculture (e.g., mussel culture) than we do about other bivalve species (e.g., oysters). While the number of large - scale shellfi sh aquaculture operations throughout the world, especially in Europe, the northwest-ern United States, and the Indo - West Pacifi c, continues to increase, few studies in those areas have examined the impacts on ecosystem properties at the landscape level. One excep-tion is the spread and impact of Crassostrea gigas replacing soft - sediment dominated com-munities with intertidal oyster reefs (e.g., see Chapter 14 in this book and discussion above), which will continue to be critically debated and evaluated for the benefi cial services and associated negative impacts of aquaculture practices. The underpinnings of an economic basis for restoration need to be resolved: (1) identifi cation and quantifi cation of ecosystem services (direct, indirect, and existence use); (2) determination of a clear defi nition of asso-ciated valuation and supporting services;

Impacts of native and cultured bivalves 273
tifi cation for shellfi sh protection, enhance-ment, and restoration efforts using several specifi c case studies that discuss the direct and indirect, biotic and abiotic, and positive and negative effects of shellfi sh aquaculture in estuarine and marine contexts. For the various shellfi sh species, these impacts vary by location of habitat (intertidal vs. subtidal), extent of cultured versus natural acreage (scale), other elements (e.g., existing seagrasses, competi-tors, benthos, available hard substrate, exotics) associated with the proposed area to be farmed, and by latitudinal/geographical differ-ences (cf. native species, regulations, western Atlantic, Gulf of Mexico, or Eastern Pacifi c). For some systems there is a wealth of general information that is particularly transferable in part to specifi c sites (e.g., carrying capacity, lost habitat structure); for others site - specifi c sampling and related research is required (e.g., interactions with marine mammals, fl ow, com-petitors, SAV). All of these factors need to be addressed to assess whether bivalve aquacul-ture can even partly replace lost natural systems. Finally, general statements as to whether aquaculture has benign, relatively neutral, or even negative effects (NRC 2009, 2010 ) is still unclear, especially given our limited understanding of the natural, often “ relic, ” or “ functionally extinct ” populations historically dominating estuarine and marine ecosystems (Beck et al. 2009, in press ).
Although polyculture has been practiced for thousands of years in Asia, and the concept of producing seaweed and bivalves as dual prod-ucts in estuaries is also well entrenched, recent interest in integrated multitrophic aquaculture has revolved around specifi cally using these organisms to process effl uents produced by shrimp and fi sh culture in both onshore and offshore systems where feed is added (dis-cussed in Folke and Kautsky 1989, 1992 ; McVey et al. 2002 ; Tomasso 2002 ; Neori et al. 2004 ; Shumway and Kraeuter 2004 ; Barrington et al. 2009 ; Pitta et al. 2009 ). Shellfi sh have been cocultured with other
Poulton 2004 ; Ruesink et al. 2005 ; Kochmann et al. 2008 ; Molnar et al. 2008 ; NRC 2009, 2010 ).
Although there is insuffi cient space to review the issues related to the recent fi shery options (including the potential introduction of Crassostrea ariakensis , a nonnative oyster from China and Japan) for the nearly lost native oyster population in the Chesapeake Bay (U.S. states of Maryland and Virginia) and North Carolina, there is much to be learned from the associated 5 - year draft EIS process (discussed in depth in NRC 2004 ; a recent dedicated Journal of Shellfi sh Research vol. 27[3] 2008; Blankenship 2009 ). The NRC (2004) report was over 300 pages long, and the fi nal EIS document exceeded 1500 pages, within which numerous potential oyster “ man-agement ” options were examined, such as 2n or 3n Crassostrea ariakensis , and numerous outcomes and fruitful discussions resulted. Comments from individual state and local governments, federal agencies, the scientifi c community, industry groups, lawmakers, and countless others drew diverse opinions. In con-trast to the numerous and earlier worldwide decisions to introduce (directed) nonnative bivalves into local waters (reviewed in Carlton and Mann 1996 ; NRC 2004 ; Ruesink et al. 2005 ; McKindsey et al. 2007 ; Molnar et al. 2008 ), the decision for the Chesapeake Bay was quite different. There was nearly a con-sensus conclusion to not introduce a nonnative oyster into the bay given the perceived or dem-onstrated risks versus benefi ts; whether this will follow elsewhere, only time will tell.
Since all shellfi sh habitats consist of living and dead individuals, and given the lack of suffi cient information currently available for any one site or multiple sites, it is necessary to consider the impacts of harvesting, aquacul-ture, and activities that ultimately impact estu-arine and marine habitats. This chapter provides an overview of the state of our knowl-edge regarding natural bivalve communities, reefs and aggregations, ecosystem services, jus-

274 Shellfi sh Aquaculture and the Environment
tion (e.g., Olympia oyster, Polson et al. 2009 ; use of Crassostrea virginica triploids in Chesapeake Bay and elsewhere, Beck et al. 2009 ). A great deal more site - specifi c research (e.g., NRC 2009, 2010 ) needs to be directed at examining potential sustainable practices (FAO 2009b ) and a greater emphasis needs to be placed on the impacts, both positive and negative, of large - scale bivalve aquaculture at the landscape or ecosystem scales.
Literature cited
Addessi , L. 1994 . Human disturbance and long - term changes on a rocky intertidal community . Ecological Applications 4 : 786 – 797 .
Agawin , N.S.R. , and Duarte , C.M. 2002 . Evidence of direct particle trapping by a tropical seagrass meadow . Estuaries 25 : 1205 – 1209 .
Airoldi , L. , and Beck , M.W. 2007 . Loss, status and trends for coastal marine habitats of Europe . Oceanography Marine Biology: An Annual Review 45 : 345 – 405 .
Alongi , D.M. , Chong , V.C. , Dixon , P. , Sasekumar , A. , and Tirendi , F. 2003 . The infl uence of cage aquaculture on pelagic carbon fl ow and water chemistry in tidally dominated mangrove estuar-ies of peninsular Malaysia . Marine Environmental Research 55 : 313 – 333 .
Anderson , W.D. , Keith , W.J. , Tuten , W.R. , and Mills , F.H. 1979 . A survey of South Carolina ’ s washed shell resource . South Carolina Wildlife and Marine Resources Department Technical Rpt. No. 36.
Andersson , A.J. , Mackenzie , F.T. , and Ver , L.M. 2003 . Solution of shallow - water carbonates . Geology 31 : 513 – 516 .
ASMFC . 2007 . The importance of habitat created by shellfi sh and shell beds along the Atlantic coast of the U.S. . Prepared by L.D. Coen, and R. Grizzle, with contributions by J. Lowery and K.T. Paynter, Jr. Habitat Management Series #8.
Asmus , R.M. , and Asmus , H. 1991 . Mussel beds: limiting or promoting phytoplankton? Journal of Experimental Marine Biology and Ecology 148 : 215 – 232 .
Asmus , H. , and Asmus , R.M. 2005 . Signifi cance of suspension - feeders systems on different spatial
invertebrates or algae in numerous marine systems, though this is usually more the case in suspended culture operations (e.g., Tenore et al. 1974 ; Paltzat et al. 2008 ). In other exam-ples, it is simply important to recognize and quantify the role that bivalves play in provid-ing these services for existing near - shore eco-systems (see McVey et al. 2002 ; Nelson et al. 2004 ; Lindahl et al. 2005 ; Gren et al. 2009 ) and yet recognize variability in system charac-teristics, bivalve species being cultured, and the possibility of overloading systems with bivalves that can then become serious sources of self - pollution (e.g., Cranford et al. 2007 ; Yuan et al. 2010 ). New technology such as remote sensing and geographic information system (GIS) will continue to be used to exam-ining aquaculture effects at the ecosystem level (e.g., Ward et al. 2003 ; Carswell et al. 2006 ; Grant et al. 2007 ).
Despite the fact that numerous studies have focused on mussels grown in rope culture, the associated carrying capacities, and related community and ecosystem effects (above cited references), more directed studies need to be conducted that assess the impact of molluscan aquaculture, along with its potential parallel “ services ” with native ( “ wild stock ” ) reef systems (cf. NRC 2009, 2010 ). Currently, most information is derived from the results of studies in other areas and only indirect evi-dence is generally available, often for other species, thereby making it diffi cult to evaluate positive and negative potential (positive or negative) or actual impacts (see also Heffernan 1999 ; Deal 2005 ; Dumbauld et al. 2009 ; Forrest et al. 2009 ; NRC 2009, 2010 ).
In conclusion, we suggest that aquaculture can provide enhanced structure paralleling that observed for natural communities. The use of native species is desirable because it reduces the chance of other unwanted effects while potentially restoring native bivalve eco-system functioning, reduces the fi shing pres-sure on native “ wild stocks, ” and at least partially contributes to native species restora-

Impacts of native and cultured bivalves 275
scales . In: Dame , R.F. , and Olenin , S. (eds.), The Comparative Roles of Suspension - Feeders in Ecosystems . Springer - Verlag , Dordrecht, Netherlands , pp. 199 – 219 .
Atkinson , P.W. , Maclean , I.M.D. , and Clark , N.A. 2010 . Impacts of shellfi sheries and nutrient inputs on waterbird communities in the Wash, England . The Journal of Applied Ecology 47 : 191 – 199 .
Auster , P.J. , Malatesta , R.J. , and LaRosa , S.C. 1995 . Patterns of microhabitat utilization by mobile megafauna on the southern New England (USA) continental shelf and slope . Marine Ecology Progress Series 127 : 77 – 85 .
Backwell , P.R.Y. , P.D.O. Hara , and Christy J.H. 1998 . Prey availability and selective foraging in shorebirds . Animal Behaviour 55 : 1659 – 1667 .
Bahr , L.M. , Jr. 1976 . Energetic aspects of the inter-tidal oyster reef community at Sapelo Island, Georgia, USA . Ecology 57 : 121 – 131 .
Bahr , L.M. , and Lanier , W.P. 1981 . The ecology of intertidal oyster reefs of the South Atlantic coast: a community profi le . U.S. Fish and Widlife Service, FWS/OBS - 81/15, Washington, D.C.
Bain , M. , Suszkowski , D. , Lodge , J. , and Xu , L. 2007 . Setting Targets for Restoration of the Hudson - Raritan Estuary . Hudson River Foundation , New York .
Baird , D. , Evans , P.R. , Milne , H. , and Pienkowski , M.W. 1985 . Utilization by shorebirds of benthic invertebrate production in intertidal areas . Oceanography and Marine Biology: An Annual Review 23 : 573 – 597 .
Baker , P. 1995 . Review of ecology and fi shery of the Olympia oyster, Ostrea lurida with annotated bibliography . Journal of Shellfi sh Research 14 : 501 – 518 .
Baldwin , J.R. , and Lovvorn , J.R. 1994 . Expansion of seagrass habitat by the exotic Zostera japon-ica , and its use by dabbling ducks and brant in Boundary Bay, British Columbia . Marine Ecology Progress Series 103 : 119 – 127 .
Banas , N.S. , Hickey , B.M. , Newton , J.A. , and Ruesink , J.L. 2007 . Tidal exchange, bivalve grazing, and patterns of primary production in Willapa Bay, Washington, USA . Marine Ecology Progress Series 341 : 123 – 139 .
Barrington , K. , Chopin , T. , and Robinson , S. 2009 . Integrated multitrophic aquaculture (IMTA) in
marine temperate waters . In: Soto , D. (ed.), Integrated Mariculture: A Global Review . FAO Fisheries and Aquaculture Technical Paper No. 529. FAO , Rome , pp. 7 – 46 .
Beadman , H.A. , Kaiser , M.J. , Galanidi , M. , Shucksmith , R. , and Willows , R.I. 2004 . Changes in species richness with stocking density of marine bivalves . The Journal of Applied Ecology 41 : 464 – 475 .
Beal , B.F. , and Kraus , M.G. 2002 . Interactive effects of initial size, stocking density, and type of pred-ator deterrent netting on survival and growth of cultured juveniles of the soft - shell clam, Mya arenaria L., in eastern Maine . Aquaculture 208 : 81 – 111 .
Beck , M.W. , Heck , K.L. , Able , K.W. , Childers , D.L. , Eggleston , D.B. , Gillanders , B.M. , Halpern , B. , Hays , C.G. , Hoshino , K. , Minello , T.J. , Orth , R.J. , Sheridan , P.F. , and Weinstein , M.R. 2001 . The identifi cation, conservation, and manage-ment of estuarine and marine nurseries for fi sh and invertebrates . Bioscience 51 : 633 – 641 .
Beck , M.W. , Heck , K.L. , Jr. , Able , K.W. , Childers , D.L. , Eggleston , D.B. , Gillanders , B.M. , Halpern , B.S. , Hays , C.G. , Hoshino , K. , Minello , T.J. , Orth , R.J. , Sheridan , P.F. , and Weinstein , M.P. 2003 . The Role of Nearshore Ecosystems as Fish and Shellfi sh Nurseries . Issues in Ecology, Number 11. Ecological Society of America , Washington, DC .
Beck , M.W. , Brumbaugh , R.D. , Airoldi , L. , Carranza , A. , Coen , L.D. , Crawford , C. , Defeo , O. , Edgar , G.J. , Hancock , B. , Kay , M. , Lenihan , H. , Luckenbach , M.W. , Toropova , C.L. , and Zhang , G. 2009 . Shellfi sh reefs at risk: a global analysis of problems and solutions . The Nature Conservancy , Arlington, VA , p. 52 .
Beck , M. , Brumbaugh , R. , Airoldi , L. , Carranza , A. , Coen , L. , Crawford , C. , Defeo , O. , Edgar , G. , Hancock , B. , Kay , M. , Lenihan , H. , Luckenbach , M. , Toropova , C. , Shepard , C. , and Zhang , G. 2011 . Oyster reefs at risk globally and recom-mendations for ecosystem revitalization . Bioscience 61 : 107 – 116 .
Becker , B.H. , Press , D.T. , and Allen , S.G. 2009 . Modeling the effects of El Ni ñ o, density depen-dence, and disturbance on harbor seal ( Phoca vitulina ) counts in Drakes Estero, California: 1997 – 2007 . Marine Mammal Science 25 : 1 – 18 .

276 Shellfi sh Aquaculture and the Environment
Bell , S.S. , McCoy , E.D. , and Mushinsky , H.R. (eds.). 1991 . Habitat Structure: The Physical Arrangement of Objects in Space . Chapman and Hall , London .
Black , K.D. 2001 . Environmental Impacts of Aquaculture . Sheffi eld Academic Press , Sheffi eld .
Blankenship , K. 2009 . All sides weigh in on C. ariak-ensis introduction to Bay . February 2009 issue.
Boesch , D.F. , and Turner , R.E. 1984 . Dependence of fi shery species on salt marshes: the role of food and refuge . Estuaries 7 : 460 – 468 .
Booth , D.M. , and Heck , K.L. , Jr. 2009 . Impacts of the American oyster ( Crassostrea virginica ) on growth rates of the seagrass Halodule wrightii . Marine Ecology Progress Series 389 : 117 – 126 .
Borde , A.B. , Thom , R.M. , Rumrill , S. , and Miller , L.M. 2003 . Geospatial habitat change analysis in Pacifi c Northwest coastal estuaries . Estuaries 26 : 1104 – 1116 .
Borges , A.V. , and Gypens , N. 2010 . Carbonate chemistry in the coastal zone responds more strongly to eutrophication than to ocean acidifi -cation . Limnology and Oceanography 55 : 346 – 353 .
Borthagaray , A. , and Carranza , A. 2007 . Mussels as ecosystem engineers: their contribution to species richness in a rocky littoral community . Acta Oecologica 31 : 243 – 250 .
Breitburg , D.L. 1999 . Are 3 - dimensional structure and healthy oyster populations the keys to an ecologically interesting and important fi sh com-munity? Luckenbach , M.W. , Mann , R. , and Wesson , J.E. (eds.), Oyster Reef Habitat Restoration: A Synopsis and Synthesis of Approaches . VIMS Press , Gloucester Point, VA , pp. 239 – 249 .
Breitburg , D.L. , Palmer , M.A. , and Loher , T. 1995 . Larval distributions and the spatial patterns of settlement of an oyster reef fi sh: responses to fl ow and structure . Marine Ecology Progress Series 125 : 45 – 60 .
Breitburg , D. , Coen , L.D. , Luckenbach , M.W. , Mann , R. , Posey , M. , and Wesson , J.A. 2000 . Oyster reef restoration: convergence of harvest and conservation strategies . Journal of Shellfi sh Research 19 : 371 – 377 .
Brenchley , G.A. 1981 . Disturbance and community structure: an experimental study of bioturbation in marine soft - bottom environments . Journal of Marine Research 39 : 767 – 790 .
Brewer , P.G. 1997 . Ocean chemistry of the fossil fuel CO 2 signal: the haline signature of “ business as usual. ” Geophysical Research Letters 24 : 1367 – 1369 .
Brosnan , D.M. , and Crumrine , L.L. 1994 . Effects of human trampling on marine rocky shore com-munities . Journal Experimental Marine Biology and Ecology 177 : 79 – 97 .
Brown , B. , and Wilson , W.H. , Jr. 1997 . The role of commercial digging of mudfl ats as an agent for change of infaunal intertidal populations . Journal Experimental Marine Biology and Ecology 218 : 49 – 61 .
Brumbaugh , R.D. , and Coen , L.D. 2009 . Contemporary approaches for small - scale oyster reef restoration to address substrate versus recruitment limitation: a review and comments relevant for the Olympia oyster, Ostrea lurida (Carpenter, 1864) . Journal of Shellfi sh Research 28 : 147 – 161 .
Brumbaugh , R.D. , and Toropova , C. 2008 . Economic valuation of ecosystem services: a new impetus for shellfi sh restoration . Basins and Coasts 2 : 8 – 15 .
Brumbaugh , R.D. , Beck , M.W. , Coen , L.D. , Craig , L. , and Hicks , P. 2006 . A Practitioners ’ Guide to the Design and Monitoring of Shellfi sh Restoration Projects: An Ecosystem Services Approach . The Nature Conservancy , Arlington, VA .
Burger , J. 1994 . The effect of human disturbance on foraging behavior and habitat use in Piping Plover ( Charadrius melodus ) . Estuaries 17 : 695 – 701 .
Burrell , V.G. , Jr. 1986 . Species profi les: life histories and environmental requirements of coastal fi shes and invertebrates (South Atlantic) — American oyster . U.S. Fish Wildl. Serv. Biological Report 82 (11.57), U.S. Army Corps of Engineers TR EL - 82 - 4, 17pp.
Burrows , F. , Harding , J.M. , Mann , R. , Dame , R. , and Coen , L. 2005 . Chapter 4, Restoration mon-itoring of oyster reefs . In: Thayer , G.W. , McTigue , T.A. , Salz , R.J. , Merkey , D.H. , Burrows , F.M. , and Gayaldo , P.F. (eds.), Science - Based Restoration Monitoring of Coastal Habitats, Volume Two: Tools for Monitoring Coastal Habitats . NOAA Coastal Ocean Program Decision Analysis Series No. 23. NOAA National Centers for Coastal Ocean Science , Silver Spring, MD , pp. 4.2 – 4.73 .

Impacts of native and cultured bivalves 277
Buschbaum , C. , Dittmann , S. , Hong , J.S. , Hwang , I.S. , Strasser , M. , Thiel , M. , Valdivia , N. , Yoon , S.P. , and Reise , K. 2009 . Mytilid mussels: global habitat engineers in coastal sediments . Helgoland Marine Research 63 : 47 – 58 .
Bushek , D. , Richardson , D. , Bobo , M.Y. , and Coen , L.D. 2004 . Short - term shell pile quarantine reduces the abundance of Perkinsus marinus remaining in tissues attached to oyster shell . Journal of Shellfi sh Research 23 : 369 – 373 .
Cahoon , L.B. , Nearhoof , J.E. , and Tilton , C.L. 1999 . Sediment grain size effect on benthic microalgal biomass in shallow aquatic ecosys-tems . Estuaries 22 : 735 – 741 .
Caldeira , K. , and Wickett , M.E. 2003 . Oceanography: anthropogenic carbon and ocean pH . Nature 425 : 365 – 365 .
Caldow , R.W.G. , Beadman , H.A. , McGrorty , S. , Stillman , R.A. , Goss - Custard , J.D. , le V. dit Durell , S.E.A. , West , A.D. , Kaiser , M.J. , Mould , K. , and Wilson , A. 2004 . A behavior - based mod-eling approach to reducing shorebird - shellfi sh confl icts . Ecological Applications 14 : 1411 – 1427 .
Callier , M.D. , Weise , A.M. , McKindsey , C.W. , and Desrosiers , G. 2006 . Sedimentation rates in a suspended mussel farm (Great - Entry Lagoon, Canada): biodeposit production and dispersion . Marine Ecology Progress Series 322 : 129 – 141 .
Callier , M.D. , McKindsey , C.W. , and Desrosiers , G. 2008 . Evaluation of indicators used to detect mussel farm infl uence on the benthos: two case studies in the Magdalen Islands, Eastern Canada . Aquaculture 278 : 77 – 88 .
Carlton , J.T. , and Mann , R.H. 1996 . Transfers and world wide introductions . In: Kennedy , V.S. , Newell , R.I.E. , and Eble , A.F. (eds.), The Eastern Oyster Crassostrea Virginica . Maryland Sea Grant , College Park, MD , pp. 691 – 705 .
Carriker , M.R. 1992 . Introductions and transfers of molluscs: risk consideration and implications . Journal of Shellfi sh Research 11 : 507 – 510 .
Carroll , J. , Gobler , C.J. , and Peterson , B.J. 2008 . Resource limitation of eelgrass in New York estuaries: light limitation and nutrient stress alle-viation by hard clams . Marine Ecology Progress Series 369 : 39 – 50 .
Carswell , B. , Cheesman , S. , and Anderson , J. 2006 . The use of spatial analysis for environmental assessment of shellfi sh aquaculture in Baynes
Sound, Vancouver Island, British Columbia, Canada . Aquaculture 253 : 408 – 414 .
Castel , J. , Labourg , P.J. , Escaravage , V. , Auby , I. , and Garcia , M.E. 1989 . Infl uence of seagrass beds and oyster parks on the abundance and biomass patterns of meio - and macrobenthos intertidal fl ats . Estuarine, Coastal and Shelf Science 28 : 71 – 85 .
Cerco , C.F. , and Noel , M.R. 2007 . Can oyster res-toration reverse cultural eutrophication in Chesapeake Bay? Estuaries and Coasts 30 : 331 – 343 .
Cerrato , R.M. , Caron , D.A. , Lonsdale , D.J. , Rose , J.M. , and Schaffner , R.A. 2004 . Effect of the northern quahog Mercenaria mercenaria on the development of blooms of the brown tide alga Aureococcus anophagefferens . Marine Ecology Progress Series 281 : 93 – 108 .
Chew , K.K. 1990 . Global bivalve shellfi sh introduc-tions . World Aquaculture 21 : 9 – 22 .
Cloern , J.E. 1982 . Does the benthos control phyto-plankton biomass in south San Francisco Bay? Marine Ecology Progress Series 9 : 191 – 202 .
Coen , L.D. 1995 . A review of the potential impacts of mechanical harvesting on subtidal and inter-tidal shellfi sh resources . SCDNR - MRRI, 46 pp. + three Appendices.
Coen , L.D. , and Luckenbach , M.W. 2000 . Developing success criteria and goals for evalu-ating oyster reef restoration: ecological function or resource exploitation? Ecological Engineering 15 : 323 – 343 .
Coen , L.D. , Knott , D.M. , Wenner , E.L. , Hadley , N.H. , and Ringwood , A.H. 1999a . Intertidal oyster reef studies in South Carolina: design, sampling and experimental focus for evaluating habitat value and function . In: Luckenbach , M.W. , Mann , R. , and Wesson , J.A. (eds.), Oyster Reef Habitat Restoration: A Synopsis and Synthesis of Approaches . Virginia Institute of Marine Science Press , Gloucester Point, VA , pp. 131 – 156 .
Coen , L.D. , Luckenbach , M.W. , and Breitburg , D.L. 1999b . The role of oyster reefs as essential fi sh habitat: a review of current knowledge and some new perspectives . In: Benaka , L.R. (ed.), Fish Habitat: Essential Fish Habitat and Rehabilitation . American Fisheries Society , Symposium 22, Bethesda, MD , pp. 438 – 454 .

278 Shellfi sh Aquaculture and the Environment
Coen , L.D. , Judge , M. , Moncreiff , C. , and Hammerstrom , K. 2000 . Final S - K Project Report, Hard clam ( Mercenaria mercenaria ) mariculture in U.S. waters: evaluating the effects of large - scale fi eld grow - out practices on clam growth, nutrition and inshore estuarine creek communities . 119 .
Coen , L.D. , Walters , K. , Wilber , D. , and Hadley , N. 2004 . A South Carolina Sea Grant Report of A 2004 workshop to examine and evaluate oyster restoration metrics to assess ecological function, sustainability and success: results and related information . Report to the South Carolina Sea Grant Consortium, 27pp. www.oyster - restora-tion.org/scsg04/SCSG04.pdf
Coen , L.D. , Bolton - Warberg , M. , and Stephen , J.A. 2006 . An examination of oyster reefs as “ bio-logically - critical ” estuarine ecosystems . Final Report, Grant R/ER - 10, Submitted to the South Carolina Sea Grant Consortium, 214 pages, plus appendices.
Coen , L.D. , Brumbaugh , R.D. , Bushek , D. , Grizzle , R. , Luckenbach , M.W. , Posey , M.H. , Powers , S.P. , and Tolley , G. 2007 . As we see it. A broader view of ecosystem services related to oyster restoration . Marine Ecology Progress Series 341 : 303 – 307 .
Coen , L.D. , Hadley , N. , Shervette , V. , and Anderson , W. 2010 . Managing oysters in South Carolina: a fi ve year program to enhance/restore shellfi sh stocks and reef habitats on through shell plant-ing and technology improvements . SC Saltwater Recreational Fisheries License Program Final Report, 124pp.
Cohen , A.N. , and Zabin , C.J. 2009 . Oyster shells as vectors for exotic organisms . Journal of Shellfi sh Research 28 : 163 – 167 .
Cohen , R.R.H. , Dresler , P.V. , Philips , E.J.P. , and Cory , R.L. 1984 . The effect of the Asiatic clam, Corbicula fl uminea on phytoplankton of the Potomac River, Maryland . Limnology and Oceanography 29 : 170 – 180 .
Collins , J.W. 1892 . Report on the fi sheries of the Pacifi c coast of the United States . Report of the Commisioner for 1888, pp. 3 – 209. Washington, DC: United States Commission of Fish and Fisheries.
Colwell , M.A. 1993 . Shorebird community patterns in a seasonally dynamic estuary . The Condor 95 : 104 – 114 .
Colwell , M.A. , and Landrum , S.L. 1993 . Nonrandom shorebird distribution and fi ne - scale variation in prey abundance . The Condor 95 : 94 – 103 .
Comeau , L.A. , St. - Onge , P. , Pernet , F. , and Lanteigne , L. 2009 . Deterring coastal birds from roosting on oyster culture gear in eastern New Brunswick, Canada . Aquacultural Engineering 40 : 87 – 94 .
Connolly , L.M. , and Colwell , M.A. 2005 . Comparative use of longline oysterbeds and adjacent tidal fl ats by waterbirds . Bird Conservation International 15 : 237 – 255 .
Conte , F.S. , Harbell , S.C. , and RaLonde , R.L. 1994 . Oyster Culture: Fundamentals and Technology of the West Coast Industry . Western Regional Aquaculture Center , Seattle, WA .
Corbett , C.A. 2007 . Colored dissolved organic matter (CDOM), workshop summary . Technical Document 07 - 3, CHNEP, FL, 82pp.
Costanza , R. , d ’ Arge , R. , de Groot , R. , Farber , S. , Grasso , M. , Hannon , B. , Naeem , S. , Limburg , K. , Paruelo , J. , O ’ Neill , R.V. , Raskin , R. , Sutton , P. , and van den Belt , M. 1997 . The value of the world ’ s ecosystem services and natural capital . Nature 387 : 253 – 260 .
Costa - Pierce , B.A. , and Bridger , C.J. 2002 . The role of marine aquaculture facilities as habitats and ecosystems . In: Stickney , R.R. , and McVey , J.P. (eds.), Ch. 8, Responsible Marine Aquaculture . CABI Publishing , Wallingford, UK , pp. 105 – 144 .
Cranford , P.J. , Strain , P.M. , Dowd , M. , Hargrave , B.T. , Grant , J. , and Archambault , M.C. 2007 . Infl uence of mussel aquaculture on nitrogen dynamics in a nutrient enriched coastal embay-ment . Marine Ecology Progress Series 347 : 61 – 78 .
Crawford , C.M. , Macleod , C.K.A. , and Mitchell , I.M. 2003 . Effects of shellfi sh farming on the benthic environment . Aquaculture 224 : 117 – 140 .
Cressman , K.A. , Posey , M.H. , Mallin , M.A. , Leonard , L.A. , and Alphin , T.D. 2003 . Effects of oyster reefs on water quality in a tidal creek estuary . Journal of Shellfi sh Research 22 : 753 – 762 .
Cummings , V.J. , Thrush , S.F. , Hewitt , J.E. , and Turner , S.J. 1998 . The infl uence of the pinnid

Impacts of native and cultured bivalves 279
bivalve Atrina zelandica (Gray) on benthic mac-roinvertebrate communities on soft - sediment habitats . Journal of Experimental Marine Biology and Ecology 228 : 227 – 240 .
D ’ Amours , O. , Archambault , P. , Christopher , W. , McKindsey , C.W. , and Johnson , L.E. 2008 . Local enhancement of epibenthic macrofauna by aquaculture activities . Marine Ecology Progress Series 371 : 73 – 84 .
Dahlgren , C.P. , Posey , M.H. , and Hulbert , A.W. 1999 . The effects of bioturbation on the infaunal community adjacent to an offhore hardbottom reef . Bulletin of Marine Science 64 : 21 – 34 .
Daily , G.E. 1997 . Nature ’ s Services — Societal Dependence on Natural Ecosystems . Island Press , Washington, DC .
Dame , R.F. 1979 . The abundance, diversity, and biomass of macrobenthos on North Inlet, South Carolina, intertidal oyster reefs . Proceedings of the National Shellfi sh Association 69 : 6 – 10 .
Dame , R.F. 1993 . Bivalve Filter Feeders and Coastal and Estuarine Ecosystem Processes . Springer - Verlag , Heidelberg, Germany .
Dame , R. 1996 . Ecology of Marine Bivalves: An Ecosystem Approach . CRC Marine Science Series , Boca Raton, FL .
Dame , R.F. , and Libes , S. 1993 . Oyster reefs and nutrient retention in tidal creeks . Journal of Experimental Marine Biology and Ecology 171 : 251 – 258 .
Dame , R.F. , Haskin , E. , and Kjerfve , B. 1984a . Water fl ow over an intertidal oyster reef and its relationship to nutrient dynamics . Journal of Shellfi sh Research 4 : 86 – 87 .
Dame , R.F. , Zingmark , R.G. , and Haskin , E. 1984b . Oyster reefs as processors of estuarine materials . Journal of Experimental Marine Biology and Ecology 83 : 239 – 247 .
Dame , R. , Bushek , D. , and Prins , T. 2001 . The role of suspension feeders as ecosystem transformers in shallow coastal environments . In: Reise , K. (ed.), The Ecology of Sedimentary Coasts . Springer - Verlag , Berlin , pp. 11 – 37 .
Danielsen , F. , S ø rensen , M.K. , Olwig , M.F. , Vaithilingam , S. , Parish , F. , Burgess , N.D. , Hiraishi , T. , Karunagaran , V.M. , Rasmussen , M.S. , Hansen , L.B. , Quarto , A. , and Suryadiputra , N. 2005 . The Asian tsunami: a protective role for coastal vegetation . Science 310 : 643 .
Deal , H. 2005 . Sustainable shellfi sh: recommenda-tions for responsible aquaculture . Report for the David Suzuki Foundation, BC, Canada, 41pp.
Dealteris , J.T. , Kilpatrick , B.D. , and Rheault , R.B. 2004 . A comparative evaluation of the habitat value of shellfi sh aquaculture gear, submerged aquatic vegetation and a non - vegetated seabed . Journal of Shellfi sh Research 23 : 867 – 874 .
DeCasabianca , M. - L. , Laugier , T. , and Collart , D. 1997 . Impact of shellfi sh farming eutrophication on benthic macrophyte communities in the Thau lagoon, France . Aquaculture International 5 : 301 – 314 .
DeGrave , S. , Moore , S.J. , and Burnell , G. 1998 . Changes in benthic macrofauna associated with intertidal oyster, Crassostrea gigas (Thunberg) culture . Journal of Shellfi sh Research 17 : 1137 – 1142 .
Denny , M.W. 1988 . Biology and the Mechanics of the Wave - Swept Environment . Princeton University Press , Princeton, NJ .
Denny , M. , and Wethey , D. 2001 . Physical pro-cesses that generate patterns in marine commu-nities . In: Bertness , M.D. , Gaines , S.M. , and Hixon , M.E. (eds.), Marine Community Ecology . Sinauer Associates , Sunderland, MA, USA , pp. 3 – 37 .
Dernie , K.M. , Kaiser , M.J. , Richardson , E.A. , and Warwick , R.M. 2003 . Recovery of soft sediment communities and habitats following physical disturbance . Journal of Experimental Marine Biology and Ecology 285 : 415 – 434 . Check citation.
Dodd , S.L. , and Colwell , M.A. 1996 . Seasonal variation in diurnal and nocturnal distributions of nonbreeding shorebirds at North Humboldt Bay, California . The Condor 98 : 196 – 207 .
Doering , P.H. , Oviatt , C.A. , and Kelly , J.R. 1986 . The effects of the fi lter - feeding clam Mercenaria mercenaria on carbon cycling in experimental marine mesocosms . Journal of Marine Research 44 : 839 – 861 .
Doney , S.C. , Fabry , V.J. , Feely , R.A. , and Kleypas , J.A. 2009 . Ocean acidifi cation: the other CO 2 problem . Annual Review of Marine Science 1 : 169 – 192 .
Donovan , S.K. 1991 . The Processes of Fossilization . Columbia University Press , New York .
Dowd , M. 2005 . A bio - physical coastal ecosystem model for assessing environment effects of

280 Shellfi sh Aquaculture and the Environment
marine bivalve aquaculture . Ecological Modelling 183 : 323 – 346 .
Drapeau , A. , Comeau , L.A. , Landry , T. , Stryhn , H. , and Davidson , J. 2006 . Association between longline design and mussel productivity in Prince Edward Island, Canada . Aquaculture 261 : 879 – 889 .
Drinkwaard , A.C. 1999 . Introductions and devel-opments of oysters in the North Sea area: a review . Helgol ä nder Meeresunters 52 : 301 – 308 .
Duarte , P. , Hawkins , A.J.S. , and Pereira , A. 2005 . How does estimation of environmental carrying capacity for bivalve culture depend upon spatial and temporal scales? In Dame , R.F. (ed.), Bivalve Filter Feeders in Estuarine and Marine Ecosystem Processes , NATO ASI Series, Vol. 47 , Springer, Dordrecht , The Netherlands .
Duarte , P. , Labarta , U. , and Fern á ndez - Reiriz , M.J. 2008 . Modelling local food depletion effects in mussel rafts of Galician Rias . Aquaculture 274 : 300 – 312 .
Duffy , J.E. , Richardson , J.P. , and Canuel , E.A. 2003 . Grazer diversity effects on ecosystem func-tioning in seagrass beds . Ecology Letters 6 : 637 – 645 .
Dumbauld , B.R. , and Wyllie - Echeverria , S. 2003 . The infl uence of burrowing thalassinid shrimps on the distribution of intertidal seagrasses in Willapa Bay, Washington, USA . Aquatic Botany 77 : 27 – 42 .
Dumbauld , B.R. , Armstrong , D.A. , and McDonald , T.L. 1993 . Use of oyster shell to enhance inter-tidal habitat and mitigate loss of dungenous crab ( Cancer magister ) caused by dredging . Canadian Journal of Fisheries and Aquatic Sciences 50 : 381 – 390 .
Dumbauld , B. , Visser , E. , Armstrong , D.A. , Cole - Warner , L. , Feldman , K. , and Kauffman , B. 2000 . Use of oyster shell to create habitat for juvenile Dungeness crab in Washington coastal estuaries: status and prospects . Journal of Shellfi sh Research 19 : 379 – 386 .
Dumbauld , B.R. , Brooks , K.M. , and Posey , M.H. 2001 . Response of an estuarine benthic com-munity to application of the pesticide carbaryl and cultivation of Pacifi c oysters ( Crassostrea gigas ) in Willapa Bay, Washington . Marine Pollution Bulletin 42 : 826 – 844 .
Dumbauld , B.R. , Booth , S. , Cheney , D. , Suhrbier , A. , and Beltran , H. 2006 . An integrated pest
management program for burrowing shrimp control in oyster aquaculture . Aquaculture 261 : 976 – 992 .
Dumbauld , B.R. , Ruesink , J.L. , and Rumrill , S.S. 2009 . The ecological role of bivalve shellfi sh aquaculture in the estuarine environment: a review with application to oyster and clam culture in West Coast (USA) estuaries . Aquaculture 290 : 196 – 223 .
Eckrich , C.E. , and Holmquist , J.G. 2000 . Trampling in a seagrass assemblage: direct effects, response of associated fauna, and the role of substrate characteristics . Marine Ecology Progress Series 201 : 199 – 209 .
Tomasso , J.R. (ed.) 2002 . Aquaculture and the environment in the United States . U.S. Aquaculture Society, A chapter of the World Aquaculture Society, Baton Rouge, LA. U.S.A., 277pp.
Eggleston , D.B. , Elis , W.E. , Etherington , L.L. , Dahlgren , C.P. , and Posey , M.H. 1999 . Organism responses to habitat fragmentation and diver-sity: habitat colonization by estuarine macro-fauna . Journal of Experimental Marine Biology and Ecology 236 : 107 – 132 .
Erbland , P.J. , and Ozbay , G. 2008 . Comparison of the macrofaunal communities inhabiting a Crassostrea virginica oyster reef and oyster aquaculture gear in Indian River Bay, Delaware . Journal of Shellfi sh Research 27 : 757 – 768 .
Everett , R.A. , Ruiz , G.M. , and Carlton , J.T. 1995 . Effect of oyster mariculture on submerged aquatic vegetation: an experimental test in a Pacifi c Northwest estuary . Marine Ecology Progress Series 125 : 205 – 217 .
FAO . 2009a . The State of World Fisheries and Aquaculture 2008 . Food and Agriculture Organization of the United Nations , Rome, Italy .
FAO . 2009b . Environmental impact assessment and monitoring in aquaculture . FAO Fisheries and Aquaculture Technical Paper No. 527. Rome, FAO. 2009. 57pp (CDROM full document is 648 pp).
Farber , S.C. , Costanza , R. , and Wilson , M.A. 2002 . Economic and ecological concepts for valuing ecosystem services . Ecological Economics 41 : 375 – 392 .
Faunce , C.H. , and Serafy , J.E. 2006 . Mangroves as fi sh habitat: fi fty years of fi eld studies . Marine Ecology Progress Series 318 : 1 – 18 .

Impacts of native and cultured bivalves 281
Feely , R.A. , Sabine , C.L. , Lee , K. , Berelson , W. , Kleypas , J. , Fabry , V.J. , and Millero , F.J. 2004 . Impact of anthropogenic CO 2 on the CaCO 3 system in the oceans . Science 305 : 362 – 366 .
Feigin , R.A. , Lozada - Bernhard , S.M. , Ravens , T.M. , M ö ller , I. , Yeager , K.M. , and Baird , A.H. 2009 . Does vegetation prevent wave erosion of salt marsh edges? PNAS 106 : 10109 – 10113 .
Feldman , K.L. , Armstrong , D.A. , Dumbauld , B.R. , DeWitt , T.H. , and Doty , D.C. 2000 . Oysters, crabs, and burrowing shrimp: review of an envi-ronmental confl ict over aquatic resources and pesticide use in Washington State ’ s (USA) coastal estuaries . Estuaries 23 : 141 – 176 .
Ferraro , S.P. , and Cole , F.A. 2007 . Benthic macro-fauna - habitat associations in Willapa Bay, Washington, USA . Estuarine, Coastal and Shelf Science 71 : 491 – 507 .
Ferreira , J.G. , Hawkins , A.J.S. , and Bricker , S.B. 2007 . Management of productivity, environ-mental effects and profi tability of shellfi sh aqua-culture — the Farm Aquaculture Resource Management (FARM) model . Aquaculture 264 : 160 – 174 .
Ferreira , J.G. , Sequeira , A. , Hawkins , A.J.S. , Newton , A. , Nickell , T.D. , Pastres , R. , Forte , J. , Bodoy , A. , and Bricker , S.B. 2009 . Analysis of coastal and offshore aquaculture: application of the FARM model to multiple systems and shell-fi sh species . Aquaculture 289 : 32 – 41 .
Folke , C. , and Kautsky , N. 1989 . The role of eco-system for a sustainable development of aqua-culture . Ambio 18 : 234 – 243 .
Folke , C. , and Kautsky , N. 1992 . Aquaculture with environment: prospects for sustainability . Ocean Coastal Management 17 : 5 – 24 .
Forrest , B. , Keeley , N. , Gillespie , P. , Hopkins , G. , Knight , B. , and Govier , D. 2007 . Review of the ecological effects of marine fi nfi sh aquaculture . Cawthron Report 1285 : 1 – 73 .
Forrest , B.M. , Keeley , N.B. , Hopkins , G.A. , Webb , S.C. , and Clement , D.M. 2009 . Bivalve aquaculture in estuaries: review and synthesis of oyster cultivation effects . Aquaculture 298 : 1 – 15 .
Freeman , M. 1995 . The benefi ts of water quality improvements for the marine recreation: a review of the empirical evidence . Marine Resource Economics 10 : 385 – 406 .
Fulford , R.S. , Breitburg , D.L. , Newell , R.I.E. , Kemp , W.M. , and Luckenbach , M.W. 2007 . Effects of oyster population restoration strate-gies on phytoplankton biomass in Chesapeake Bay: a fl exible modeling approach . Marine Ecology Progress Series 336 : 43 – 61 .
Fulford , R.S. , Breitburg , D.L. , Luckenbach , M.W. , and Newell , R.I.E. 2010 . Evaluating responses of estuarine food webs to oyster restoration and nutrient load reduction with a multi - species bio-energetics model . Ecological Applications 20 : 915 – 934 .
Galtsoff , P.S. 1964 . The American oyster Crassostrea virginica Gmelin . U.S Fish Widl Serv Fishery Bulletin 64 : 1 – 480 .
Gazeau , F. , Quiblier , C. , Jansen , J.M. , Gattuso , J. - P. , Middelburg , J.J. , and Heip , C.H.R. 2007 . Impact of elevated CO 2 on shellfi sh calcifi cation . Geophysical Research Letters 34 : L07603 . doi:10.1029/2006GL028554
Gilbert , F. , Souchu , P. , Bianchi , M. , and Bonin , P. 1997 . Infl uence of shellfi sh farming activities on nitrifi cation, nitrate reduction to ammonium and denitrifi cation at the water sediment inter-face of the Thau lagoon, France . Marine Ecology Progress Series 151 : 143 – 153 .
Gillespie , G. 2009 . Status of the Olympia oyster, Ostrea lurida , (Carpenter 1864) in British Columbia, Canada . Journal of Shellfi sh Research 28 : 59 – 68 .
Giotta , R.E. 1999 . Distribution of the American oyster ( Crassostrea virginica ) in South Carolina: interacting effects of predation, sedimentation and water fl ow at varying tidal elevations . M.S. Thesis, College of Charleston, South Carolina, 98pp.
Glancy , T.P. , Frazer , K.T.K. , Cichra , C.E. , and Lindberg , W.J. 2003 . Comparative patterns of occupancy by decapod crustaceans in seagrass, oyster, and marsh - edge habitats in a northeast Gulf of Mexico estuary . Estuaries 26 : 1291 – 1301 .
Glude , J.B. , and Landers , W.S. 1953 . Biological effects on hard clams of hand raking and power dredging . U.S. Fish Wildl. Serv. Spec. Rep. Fish. 110, 43pp.
Godcharles , M.F. 1971 . A study of the effects of a commercial hydraulic clam dredge on benthic communities in estuarine areas . State of Fla. Dept. of Nat. Res. Tech. Ser. No. 64:1 – 51.

282 Shellfi sh Aquaculture and the Environment
Godet , L. , Toupoint , N. , Fournier , J. , Le Mao , P. , Retiere , C. , and Olivier , F. 2009 . Clam farmers and oystercatchers: effects of the degradation of Lanice conchilega beds by shellfi sh farming on the spatial distribution of shorebirds . Marine Pollution Bulletin 58 : 589 – 595 .
Goodwin , L. 2007 . Evaluating the impacts of envi-ronmental parameters on shoreline erosion and related aspects: assessing the current status of vegetation, sediments, and biota . M.S. thesis, College of Charleston, SC 117pp.
Goss - Custard , J.D. , Stillman , R.A. , West , A.D. , Caldow , R.W.G. , Triplet , P. , le V. dit Durell , S.E.A. , and McGrorty , S. 2004 . When enough is not enough: shorebirds and shellfi shing . Proceedings of the Royal Society of London. Series B. Biological Sciences 271 : 233 – 237 .
Grabowski , J.H. , and Peterson , C.H. 2007 . Restoring oyster reefs to recover ecosystem ser-vices . In: Ch. 15, Cuddington , K. , Byers , J.E. , Wilson , W.G. , and Hastings , A. (eds.), Ecosystem Engineers: Concepts, Theory and Applications . Elsevier/Academic Press , Burlington, MA , pp. 281 – 298 .
Grant , J. , Hatcher , A. , Scott , D.B. , Pocklington , P. , Schafer , C.T. , and Winters , G.V. 1995 . A multi-disciplinary approach to evaluating impacts of shellfi sh aquaculture on benthic communities . Estuaries 18 : 124 – 144 .
Grant , J. , Cranford , P. , Hargrave , B. , Carreau , M. , Schofi eld , B. , Armsworthy , S. , Burdett - Coutts , V. , and Ibarra , D. 2005 . A model of aquaculture biodeposition for multiple estuaries and fi eld validation at blue mussel (Mytilus edulis) culture sites in eastern Canada . Canadian Journal of Fisheries and Aquatic Sciences 62 : 1271 – 1285 .
Grant , J. , Bugden , G. , Horne , E. , Archambault , M.C. , and Carreau , M. 2007 . Remote sensing of particle depletion by coastal suspension - feeders . Canadian Journal of Fisheries and Aquatic Sciences 64 : 387 – 390 .
Gren , I. - M. , Lindahl , O. , and Lindqvist , M. 2009 . Values of mussel farming for combating eutro-phication: an application to the Baltic Sea . Ecological Engineering 35 : 935 – 945 .
Grizzle , R.E. 1988 . The relative effects of seston fl ux and sediments on individual growth rates of Mercenaria mercenaria : results of a factorial fi eld experiment . Journal of Shellfi sh Research 7 : 160 – 161 .
Grizzle , R.E. , Greene , J.K. , Luckenbach , M.W. , and Coen , L.D. 2006 . Measuring and modeling seston uptake by suspension feeding bivalve mol-luscs . Journal of Shellfi sh Research 25 : 643 – 649 .
Grizzle , R.E. , Greene , J.K. , and Coen , L.D. 2008 . Seston removal by natural and constructed inter-tidal eastern oyster ( Crassostrea virginica ) reefs: a comparison with previous laboratory studies, and the value of in situ methods . Estuaries and Coasts 31 : 1208 – 1220 .
Guti é rrez , J.L. , Jones , C.G. , Strayer , D.L. , and Iribarne , O.O. 2003 . Molluscs as ecosystem engineers: the role of shell production in aquatic habitats . Oikos 101 : 79 – 90 .
Hadley , N.H. , Hodges , M. , Wilber , D.H. , and Coen , L.D. 2010 . Evaluating intertidal oyster reef development in South Carolina using associated faunal indicators . Restoration Ecology 18 : 691 – 701 .
Hall , S.J. 1994 . Physical disturbance and marine benthic communities: life in unconsolidated sedi-ments . Oceanography and Marine Biology Annual Review 32 : 179 – 239 .
Hammel , A. , and Webb , D. 1999 . Ecological impli-cations of Mercenaria aquaculture in the Cape Cod National Seashore . Abstract and poster pre-sented at the Annual Benthic Ecology Meeting, Baton Rouge, LA.
Harbin - Ireland , A.C. 2004 . Effects of oyster mari-culture on the benthic invertebrate community in Drakes Estero, Pt. Reyes Peninsula, California . M.S. thesis, University of California, Davis, California.
Harding , J.M. , and Mann , R. 1999 . Fish species richness in relation to restored oyster reefs, Piankatank River, Virginia . Bulletin of Marine Science 65 : 289 – 300 .
Harding , J.M. , and Mann , R. 2001 . Oyster reefs as fi sh habitat: opportunistic use of restored reefs by transient fi shes . Journal of of Shellfi sh Research 20 : 951 – 959 .
Hargis , W.J. , Jr. , and Haven , D.S. 1999 . Chesapeake oyster reefs, their importance, destruction and guidelines for restoring them . In: Luckenbach , M.W. , Mann , R. , and Wesson , J.A. (eds.), Oyster Reef Habitat Restoration: A Synopsis and Synthesis of Approaches . Virginia Institute of Marine Science Press , Gloucester Point, VA , pp. 329 – 358 .
Harrison , P.G. , and Bigley , R.E. 1982 . The recent introduction of the seagrass Zostera japonica to

Impacts of native and cultured bivalves 283
the Pacifi c coast of North America . Canadian Journal of Fisheries and Aquatic Sciences 39 : 1642 – 1648 .
Harsh , D.A. , and Luckenbach , M.W. 1999 . Materials processing by oysters in patches: inter-active roles of current speed and seston composi-tion . In: Luckenbach , M.W. , Mann , R. , and Wesson , J.A. (eds.), Oyster Reef Habitat Restoration. A Synopsis and Synthesis of Approaches . Virginia Institute of Marine Science Press , Gloucester Point, VA , pp. 251 – 265 .
Hastings , R.W. , and Heinle , D.R. 1995 . The effects of aquaculture in estuarine environments: intro-duction to the dedicated issue . Estuaries 18 : 1 – 284 .
Hauxwell , J. , Cebrian , J. , Furlong , C. , and Valiela , I. 2001 . Macroalgal canopies contribute to eel-grass ( Zostera marina ) decline in temperate estuarine ecosystems . Ecology 82 : 1007 – 1022 .
Haven , D.S. , and Morales - Alamo , R. 1966 . Aspects of biodeposition by oysters and other inverte-brate fi lter feeders . Limnology and Oceanography 11 : 487 – 498 .
Heal , G. 2000 . Nature and the Marketplace: Capturing the Value of Ecosystem Services . Island Press , Washington, DC .
Hecht , T. , and Britz , P. 1992 . The current status, future prospects and environmental implications of mariculture in South Africa . South African Journal of Science 88 : 335 – 342 .
Heck , K.L. , Jr. , and Crowder , L.B. 1991 . Habitat structure and predator - prey interactions in veg-etated aquatic systems . In: Bell , S.S. , McCoy , E.D. , and Mushinsky , H.R. (eds.), Habitat Structure: the Physical Arrangement of Objects in Space . Chapman and Hall , London , pp. 281 – 295 .
Heck , K.L. , Jr. , Pennock , J.R. , Valentine , J.F. , Coen , L.D. , and Sklenar , S.A. 2000a . Effects of nutrient enrichment and large predator removal on sea-grass nursery habitats: an experimental assess-ment . Limnology and Oceanography 45 : 1041 – 1057 .
Heck , K.L. , Jr. , Pennock , J.R. , Valentine , J.F. , Coen , L.D. , and Sklenar , S.A. 2000b . Effects of nutri-ent enrichment and overfi shing on seagrass eco-systems: an experimental assessment . Biologia Marina Mediterranea 7 : 220 – 222 .
Heck , K.L. , Jr. , Hays , C. , and Orth , R.J. 2003 . A critical evaluation of the nursery role hypothesis
for seagrass meadows . Marine Ecology Progress Series 253 : 123 – 136 .
Heck , K.L. , Jr. , Valentine , J.F. , Pennock , J.R. , Chaplin , G. , and Spitzer , P.M. 2006 . Effects of nutrient enrichment and grazing on shoal grass ( Halodule wrightii ) and its epiphytes: results of a fi eld experiment . Marine Ecology Progress Series 326 : 145 – 156 .
Heck , K.L. , Carruthers , T.J.B. , Duarte , C.M. , Hughes , A.R. , Kendrick , G. , Orth , R.J. , and Williams , S.W. 2008 . Trophic transfers from seagrass meadows subsidize diverse marine and terrestrial consumers . Ecosystems 11 : 1198 – 1210 .
Heffernan , M.L. 1999 . A review of the ecological implications of mariculture and intertidal har-vesting in Ireland. Irish Wildlife Manuals, No. 7 . D ú chas, The Heritage Service, Department of Arts, Heritage, Gaeltacht and the Islands, Dublin, Ireland, 156pp. www.npws.ie/en/media/Media,3762,en.pdf
H é garet , H. , Shumway , S.E. , Wikfors , G.H. , Pate , S. , and Burkholder , J.M. 2008 . Potential trans-port of harmful algae through relocation of bivalve molluscs . Marine Ecology Progress Series 361 : 169 – 179 .
Heil , D.C. 2009 . Karenia brevis monitoring, man-agement, and mitigation for Florida molluscan shellfi sh harvesting areas . Harmful Algae 8 : 608 – 610 .
Heise , R.J. , and Bortone , S.A. 1999 . Estuarine arti-fi cial reefs to enhance seagrass planting and provide fi sh habitat . Gulf of Mexico Science 17 : 59 – 74 .
Hemminga , M.A. , and Duarte , C.M. 2000 . Seagrass Ecology . Cambridge University Press , Cambridge .
Heral , M. 1993 . Why carrying capacity models are useful tools for management of bivalve molluscs culture . In: Dame , R.F. (ed.), Bivalve Filter Feeders in Estuarine and Coastal Ecosystem Processes . Springer - Verlag , Berlin, Heidelberg , pp. 455 – 477 .
Hewitt , J.E. , and Norkko , J. 2007 . Incorporating temporal variability of stressors into studies: an example using suspension - feeding bivalves and elevated suspended sediment concentrations . Journal of Experimental Marine Biology and Ecology 341 : 131 – 141 .
Hilgerloh , G. , O ’ Halloran , J. , Kelly , T.C. , and Burnell , G.M. 2001 . A preliminary study on the

284 Shellfi sh Aquaculture and the Environment
effects of oyster culturing structures on birds in a sheltered Irish estuary . Hydrobiologia 465 : 175 – 180 .
Holmer , M. 1991 . Impacts of aquaculture on sur-rounding sediments: generation of organic - rich sediments . In: De Pauw , N. , and Joyce , J. (eds.), Aquaculture and the Environment , Vol. 16 . European Aquaculture Soc. Spec. Publ., pp. 155 – 175 .
Holsman , K.K. , McDonald , P.S. , and Armstrong , D.A. 2006 . Intertidal migration and habitat use by subadult Dungeness crab Cancer magister in a NE Pacifi c estuary . Marine Ecology Progress Series 308 : 183 – 195 .
Holt , T.J. , Rees , E.I. , Hawkins , S.J. , and Seed , R. 1998 . Biogenic reefs, Volume 9: an overview of dynamic and sensitivity characteristics for con-servation management of marine SACs . Scottish Association for Marine Science, Port Erin Marine Laboratory, University of Liverpool, Scotland.
Hopkins , F.E. , Turnera , S.M. , Nightingale , P.D. , Steinke , M. , Bakkera , D. , and Lissa , P.S. 2010 . Ocean acidifi cation and marine trace gas emis-sions . PNAS 107 : 760 – 765 .
Hosack , G.R. , Dumbauld , B.R. , Ruesink , J.L. , and Armstrong , D.A. 2006 . Habitat associations of estuarine species: comparisons of intertidal mudfl at, seagrass ( Zostera marina ), and oyster ( Crassostrea gigas ) habitats . Estuaries Coasts 29 : 1150 – 1160 .
Huang , C.H. , Lin , H.J. , Huang , T.C. , Su , H.M. , and Hung , J.J. 2008 . Responses of phytoplankton and periphyton to system - scale removal of oys-ter - culture racks from a eutrophic tropical lagoon . Marine Ecology Progress Series 358 : 1 – 12 .
Hughes , A.R. , Williams , S.L. , Duarte , C.M. , Heck , K.L. , and Waycott , M. 2009 . Associations of concern: declining seagrasses and threatened dependent species . Frontiers in Ecology and the Environment 7 : 242 – 246 .
Inglis , G.J. , and Gust , N. 2003 . Potential indirect effects of shellfi sh culture on the reproductive success of benthic predators . The Journal of Applied Ecology 40 : 1077 – 1089 .
Jackson , J.B.C. , Kirby , M.X. , Berger , W.H. , Bjorndal , K.A. , Botsford , L.W. , Bourque , B.J. , Bradbury , R.H. , Cooke , R. , Erlandson , J. , Estes , J.A. , Hughes , T.P. , Kidwell , S. , Lange , C.B. , Lenihan , H.S. , Pandolfi , J.M. , Peterson , C.H. ,
Steneck , R.S. , Tegner , M.J. , and Warner , R.R. 2001 . Historical overfi shing and the recent col-lapse of coastal ecosystems . Science 293 : 629 – 638 .
Jackson , E.L. , Rowden , A.A. , Attrill , M.J. , Bossey , S.J. , and Jones , M.B. 2001 . The importance of seagrass beds as a habitat for fi shery species . Oceanography Marine Biology: An Annual Review 39 : 269 – 303 .
Johnson , M.W. , Heck , K.L. , Jr. , and Fourqurean , J.W. 2006 . Nutrient content of seagrasses and epiphytes in the northern Gulf of Mexico: evi-dence of phosphorus and nitrogen limitation . Aquatic Botany 85 : 103 – 111 .
Jones , C.G. , Lawton , J.H. , and Shachak , M. 1994 . Organisms as ecosystem engineers . Oikos 69 : 373 – 386 .
Jones , C.G. , Lawton , J.H. , and Shackak , M. 1997 . Positive and negative effects of organisms as physical ecosystem engineers . Ecology 78 : 1946 – 1957 .
Judge , M.L. , and Craig , S.F. 1997 . Positive fl ow dependence in the initial colonization of a fouling community: results from in situ water current manipulations . Journal of Experimental Marine Biology and Ecology 210 : 209 – 222 .
Judge , M.L. , Coen , L.D. , and Heck , K.L. , Jr. 1992 . The effect of long - term alteration of in situ cur-rents on the growth of Mercenaria mercenaria in the northern Gulf of Mexico . Limnology and Oceanography 37 : 1550 – 1559 .
Judge , M.L. , Coen , L.D. , and Heck , K.L. , Jr. 1993 . Does Mercenaria mercenaria encounter elevated food levels in seagrass beds? Results from a novel technique to collect suspended food resources . Marine Ecology Progress Series 92 : 141 – 150 .
Judge , M.L. , Coen , L.D. , and Hammerstrom , K. 2000 . The ecological implications of high density hard clam ( Mercenaria mercenaria ) mariculture on tidal creek environments . Journal of Shellfi sh Research 19 : 609 – 610 .
Kaiser , M.J. 2000 . Ecological effects of shellfi sh cultivation . In: Black , K.D. (ed.), Environmental Impacts of Aquaculture . Sheffi eld Academic Press , Sheffi eld, UK , pp. 51 – 75 .
Kaiser , M.J. 2001 . Ecological effects of shellfi sh cultivation . In: Black , K.D. (ed.), Environmental Impacts of Aquaculture . CRC Press , Boca Raton, FL .

Impacts of native and cultured bivalves 285
Kaiser , M.J. , Laing , I. , Utting , S.D. , and Burnell , G.M. 1998 . Environmental impacts of bivalve mariculture . Journal of Shellfi sh Research 17 : 59 – 66 .
Kathiresan , K. , and Rajendran , N. 2005 . Coastal mangrove forests mitigated tsunami . Estuarine, Coastal and Shelf Science 65 : 601 – 606 .
Kaufman , L. , and Dayton , P.K. 1997 . Impacts of marine resource extraction on ecosystem services and sustainability . In: Daily , G.C. (ed.), Nature ’ s Services: Societal Dependence on Natural Ecosystems . Island Press , Washington, DC , pp. 275 – 293 .
Kelly , J.P. , Evens , J.G. , Stallcup , R.W. , and Wimpfheimer , D. 1996 . Effects of oyster culture on habitat use by wintering shorebirds in Tomales Bay, California . California Fish Game 82 : 160 – 174 .
Kelly , J.R. , Proctor , H. , and Volpe , J.P. 2008 . Intertidal community structure differs signifi -cantly between substrates dominated by native eelgrass ( Zostera marina L.) and adjacent to the introduced oyster Crassostrea gigas (Thunberg) in British Columbia, Canada . Hydrobiologia 596 : 57 – 66 .
Kemper , C. , Pemberton , D. , Cawthorn , M. , Heinrich , S. , Mann , J. , W ü rsig , B. , Shaughnessy , P.D. , and Gales , R. 2003 . Aquaculture and marine mammals: coexistence or confl ict? Gales , N. , Hindell , M. , and Kirkwood , R. (eds.), Marine Mammals: Fisheries, Tourism and Management Issues . CSIRO Publishing , Collingwood, NSW, Australia .
Kennedy , V.S. 1996 . Biology of larvae and spat . In: Kennedy , V.S. , Newell , R.I.E. , and Eble , A.F. (eds.), The Eastern Oyster: Crassostrea Virginica . Maryland Sea Grant , College Park, MD , pp. 371 – 421 .
Kennedy , V.S. , and Sanford , L.P. 1999 . Characteristics of relatively unexploited beds of the eastern oyster, Crassostrea virginica , and early restora-tion programs . In: Luckenbach , M.W. , Mann , R. , and Wesson , J.A. (eds.), Oyster Reef Habitat Restoration: A Synopsis and Synthesis of Approaches . Virginia Institute of Marine Science Press , Gloucester Point, VA , pp. 25 – 46 .
Kennedy , V.S. , Newell , R.I.E. , and Eble , A.F. (eds.). 1996 . The Eastern Oyster: Crassostrea Virginica . Maryland Sea Grant , College Park, MD .
Keough , M.J. 1984 . Dynamics of the epifauna of the bivalve Pinna bicolor : interactions among recruitment, predation and competition . Ecology 65 : 677 – 688 .
Kiessling , W. , Simpson , C. , and Foote , M. 2010 . Reefs as cradles of evolution and sources of bio-diversity in the Phanerozoic . Science 327 : 196 – 198 .
Kimbro , D.L. , and Grosholz , E.D. 2006 . Disturbance infl uences oyster community richness and even-ness, but not diversity . Ecology 87 : 2378 – 2388 .
Kimbro , D.L. , Largier , J. , and Grosholz , E.D. 2009 . Coastal oceanographic processes infl uence the growth and size of a key estuarine species, the Olympia oyster . Limnology and Oceanography 54 : 1425 – 1437 .
Kirby , M.X. 2004 . Fishing down the coast: histori-cal expansion and collapse of oyster fi sheries along coastal margins . Proceedings of the National Shellfi sh Association 101 : 13096 – 13099 .
Kirby , M.X. , and Miller , H.M. 2005 . Response of a benthic suspension feeder ( Crassostrea virgin-ica Gmelin) to three centuries of anthropogenic eutrophication in Chesapeake Bay . Estuarine, Coastal and Shelf Science 62 : 679 – 689 .
Kochmann , J. , Buschbaum , C. , Volkenborn , N. , and Reise , K. 2008 . Shift from native mussels to alien oysters: differential effects of ecosystem engineers . Journal of Experimental Marine Biology and Ecology 364 : 1 – 10 .
Kraan , C. , van Gils , J.A. , Spaans , B. , Dekinga , A. , Bijleveld , A.I. , van Roomen , M. , Kleefstra , R. , and Piersma , T. 2009 . Landscape - scale experi-ment demonstrates that Wadden Sea intertidal fl ats are used to capacity by molluscivore migrant shorebirds . The Journal of Animal Ecology 78 : 1259 – 1268 .
Kuhlmann , M.L. 1998 . Spatial and temporal patterns in the dynamics and use of pen shells ( Atrina rigida ) as shelters in St. Joseph Bay, Florida . Bulletin of Marine Science 62 : 157 – 179 .
Langton , R.W. , and Robinson , W.E. 1990 . Faunal associations on scallop grounds in the western Gulf of Maine . Journal of Experimental Marine Biology and Ecology 144 : 157 – 171 .
Larkum , A.W.D. , Orth , R.J. , and Duarte , C. (eds.). 2006 . Seagrasses: Biology, Ecology and

286 Shellfi sh Aquaculture and the Environment
Conservation . Springer , Dordrecht, The Netherlands .
Leguerrier , D. , Niquil , N. , Petiau , A. , and Bodoy , A. 2004 . Modeling the impact of oyster culture on a mudfl at food web in Marennes - Oleron Bay (France) . Marine Ecology Progress Series 273 : 147 – 161 .
Lehnert , R.L. , and Allen , D.M. 2002 . Nekton use of subtidal oyster shell habitats in a southeastern U.S. estuary . Estuaries 25 : 1015 – 1024 .
Lenihan , H.S. 1999 . Physical - biological coupling on oyster reefs: how habitat structure infl uences individual performance . Ecological Monographs 69 : 251 – 275 .
Lenihan , H.S. , and Micheli , F. 2000 . Biological effects of shellfi sh harvesting on oyster reefs: resolving a fi shery confl ict by ecological experi-mentation . Fishery Bulletin 98 : 86 – 95 .
Lenihan , H.S. , and Peterson , C.H. 1998 . How habitat degradation through fi shery disturbance enhances impacts of hypoxia on oyster reefs . Ecological Applications 8 : 128 – 140 .
Lenihan , H.S. , and Peterson , C.H. 2004 . Conserving oyster reef habitat by switching from dredging and tonging to diver hand - harvesting . Fishery Bulletin 102 : 298 – 305 .
Lenihan , H.S. , Peterson , C.H. , Byers , J.E. , Grabowski , J.H. , Thayer , G.W. , and Colby , D.R. 2001 . Cascading of habitat degradation: oyster reefs invaded by refugee fi shes escaping stress . Ecological Applications 11 : 748 – 764 .
Light , B.R. , and Beardall , J. 1998 . Distribution and spatial variation of benthic microalgal biomass in a temperate, shallow - water marine system . Aquatic Botany 61 : 39 – 54 .
Lindahl , O. , Hart , R. , Hernroth , B. , Kollberg , S. , Loo , L. , Olrog , L. , and Rehnstam - Holm , A. 2005 . Improving marine water quality by mussel farming: a profi table solution for Swedish Society . Ambio 34 : 131 – 138 .
Lindsay , C.E. , and Simons , D. 1997 . The fi sheries for Olympia oysters, Ostreola conchaphila ; Pacifi c oysters, Crassostrea gigas ; and Pacifi c razor clams, Siliqua patula , in the State of Washington . In: Mackenzie , C.L.J. , Burrell , V.G.J. , Rosenfi eld , A. , and Hobart , W.L. (eds.), The History, Present Condition, and Future of the Molluscan Fisheries of North and Central America and Europe , Vol. 2 , Pacifi c Coast and Supplemental Topics. Seattle, WA: NOAA , U.S. Department of Commerce , pp. 89 – 113 .
Lipton , D. 2004 . The value of improved water quality to Chesapeake Bay boaters . Marine Resource Economics 19 : 265 – 270 .
Lloyd , B.D. 2003 . Potential Effects of Mussel Farming on New Zealand ’ s Marine Mammals and Seabirds: A Discussion Paper . Department of Conservation , Wellington, New Zealand .
Lotze , H.K. , Leniham , H.S. , Bourque , B.J. , Bradbury , R.H. , Cooke , R.G. , Kay , M.C. , Kidwell , S.M. , Kirby , M.X. , Peterson , C.H. , and Jackson , J.B.C. 2006 . Depletion, degredation, and recovery potential of estuaries and coastal seas . Science 312 : 1806 – 1809 .
Lu , L. , and Grant , J. 2008 . Recolonization of inter-tidal infauna in relation to organic deposition at an oyster farm in Atlantic Canada — a fi eld experiment . Estuaries and Coasts 31 : 767 – 775 .
Luckenbach , M.W. , and Orth , R.J. 1999 . Interactions between benthic infauna and the transport and burial of Zostera marina seeds . Aquatic Botany 62 : 235 – 247 .
Luckenbach , M.W. , Mann , R. , and Wesson , J.A. (eds.). 1999 . Oyster Reef Habitat Restoration. A Synopsis and Synthesis of Approaches . Virginia Institute of Marine Science Press , Gloucester Point, VA .
Luckenbach , M.W. , Coen , L.D. , Ross , P.G. , Jr. , and Stephen , J.A. 2005 . Oyster reef habitat restora-tion: relationships between oyster abundance and community development based on two studies in Virginia and South Carolina . Journal of Coastal Research Special Issue ( 40 ): 64 – 78 .
Macleod , C.K. , Forbes , S.E. , Shepherd , C.J. , and Crawford , C. 2009 . Effects of oyster farming service vehicles on an intertidal sand fl at . Aquaculture Research 40 : 772 – 780 .
Mallet , A.L. , Carver , C.E. , and Landry , T. 2006 . Impact of suspended and off - bottom eastern oyster culture on the benthic environment in eastern Canada . Aquaculture 255 : 362 – 373 .
Mallet , A.L. , Carver , C.E. , and Hardy , M. 2009 . The effect of fl oating bag management strategies on biofouling, oyster growth and biodeposition levels . Aquaculture 287 : 315 – 323 .
Mann , R. , and Evans , E.A. 2004 . Site selection for oyster habitat rehabilitation in the Virginia portion of the Chesapeake Bay: a commentary . Journal of Shellfi sh Research 23 : 41 – 49 .
Mann , R. , and Powell , E.N. 2007 . Why oyster res-toration goals in the Chesapeake Bay are not and

Impacts of native and cultured bivalves 287
probably cannot be achieved . Journal of Shellfi sh Research 26 : 905 – 917 .
Markert , A. , Wehrmann , A. , and Kr ö ncke1 , I. 2010 . Recently established Crassostrea - reefs versus native Mytilus - beds: differences in ecosys-tem engineering affects the macrofaunal com-munities (Wadden Sea of Lower Saxony, southern German Bight) . Biological Invasions 12 : 15 – 32 .
McGlathery , K.J. , Sundb ä ck , K. , and Anderson , I.C. 2007 . Eutrophication in shallow coastal bays and lagoons: the role of plants in the coastal fi lter . Marine Ecology Progress Series 348 : 1 – 18 .
McGraw , K.A. 2009 . The Olympia oyster, Ostrea lurida Carpenter, 1864 along the west coast of North America . Journal of Shellfi sh Research 28 : 5 – 10 .
McKindsey , C.W. , Thetmeyer , H. , Landry , T. , and Silvert , W. 2006 . Review of recent carrying capacity models for bivalve culture and recom-mendations for research and management . Aquaculture 261 : 451 – 462 .
McKindsey , C.W. , Landry , T. , O ’ Beirn , F.X. , and Davies , I.M. 2007 . Bivalve aquaculture and exotic species: a review of ecological considerations and management issues . Journal of Shellfi sh Research 26 : 281 – 294 .
McKindsey , C.W. , Lecuona , M. , Huot , M. , and Weise , A.M. 2009 . Biodeposit production and benthic loading by farmed mussels and associ-ated tunicate epifauna in Prince Edward Island . Aquaculture 295 : 44 – 51 .
McVey , J.P. , Stickney , R. , Yarish , C. , and Chopin , T. 2002 . Aquatic polyculture and balanced eco-system management: new paradigms for seafood production . In: Stickney , R.R. , and McVey , J.P. (eds.), Responsible Marine Aquaculture . CABI Publishing , Oxon, UK , pp. 91 – 104 .
Mesnage , V. , Ogier , S. , Bally , G. , Disnar , J.R. , Lottier , N. , Dedieu , K. , Rabouille , C. , and Copard , Y. 2007 . Nutrient dynamics at the sed-iment - water interface in a Mediterranean lagoon (Thau, France): infl uence of biodeposition by shellfi sh farming activities . Marine Environmental Research 63 : 257 – 277 .
Meyer , D.L. , and Townsend , E.C. 2000 . Faunal uti-lization of created intertidal eastern oyster ( Crassostrea virginica ) reefs in the southeastern United States . Estuaries 23 : 34 – 45 .
Meyer , D.L. , Townsend , E.C. , and Thayer , G.W. 1997 . Stabilization and erosion control value of
oyster cultch for intertidal marsh . Restoration Ecology 5 : 93 – 99 .
Michael , P. , and Chew , K.K. 1976 . Growth of Pacifi c oyster Crassostrea gigas and related fouling problems under tray culture at Seabeck Bay, Washington . Proceedings of the National Shellfi sh Association 66 : 36 – 41 .
Micheli , F. , and Peterson , C.H. 1999 . Estuarine vegetated habitats as corridors for predator movements . Conservation Biology 13 : 869 – 881 .
Millennium Ecosystem Assessment (MA) (ed.) 2003 . Ecosystems and Human Well - Being: A Framework for Assessment . Island Press , Washington, DC .
Miller , D.C. , Norkko , A. , and Pilditch , C.A. 2002 . Infl uence of diet on dispersal of horse mussel Atrina zelandica biodeposits . Marine Ecology Progress Series 242 : 153 – 167 .
Mojica , R. , Jr. , and Nelson , W.G. 1993 . Environmental effects of a hard clam ( Mercenaria mercenaria ) aquaculture site in the Indian River Lagoon, Florida . Aquaculture 113 : 313 – 329 .
Molnar , J.L. , Gamboa , R.L. , Revenga , C. , and Spalding , M.D. 2008 . Assessing the global threat of invasive species to marine biodiversity . Frontiers in Ecology and the Environment 6 : 485 – 492 .
Munguia , P. 2007 . Spatial structure of pen shell ( Atrina rigida ) communities . Marine Biology 152 : 149 – 156 .
Munroe , D. , and McKinley , R.S. 2007a . Effects of predator netting on recruitment and growth of Manila clams ( Venerupis philippinarum ) on soft substrate intertidal plots in British Columbia, Canada . Journal of Shellfi sh Research 26 : 1035 – 1044 .
Munroe , D. , and McKinley , R.S. 2007b . Commercial Manila clam ( Tapes philippinarum ) culture in British Columbia, Canada: the effects of preda-tor netting on intertidal sediment characteristics . Estuarine, Coastal and Shelf Science 72 : 319 – 328 .
Nash , C.E. , Iwamoto , R.N. , and Mahnken , C.V.W. 2000 . Aquaculture risk management and marine mammal interactions in the Pacifi c Northwest . Aquaculture 183 : 307 – 323 .
National Research Council (NRC) . 2004 . Nonnative Oysters in the Chesapeake Bay . National Academies Press , Washington, DC .

288 Shellfi sh Aquaculture and the Environment
National Research Council (NRC) . 2007 . Mitigating Shore Erosion along Sheltered Coasts . National Academies Press , Washington, DC .
National Research Council (NRC) . 2009 . Shellfi sh mariculture in Drakes Estero, Point Reyes National Seashore, California . Committee on Best Practices for Shellfi sh Mariculture and the Effects of Commercial Activities in Drakes Estero, Pt. Reyes National Seashore, California, 139pp.
National Research Council (NRC) . 2010 . Ecosystem concepts for sustainable bivalve mariculture . Committee on Best Practices for Shellfi sh Mariculture and the Effects of Commercial Activities in Drakes Estero, Pt. Reyes National Seashore, California, 179pp.
Naylor , R.L. , Goldburg , R.J. , Primavera , J.H. , Kautsky , N. , Beveridge , M.C.M. , Clay , J. , Folke , C. , Lubchenco , J. , Mooney , H. , and Troell , M. 2000 . Effect of aquaculture on world fi sh sup-plies . Nature 405 : 1017 – 1024 .
Naylor , R.L. , Williams , S.L. , and Strong , D.R. 2001 . Aquaculture — a gateway for exotic species . Science 294 : 1655 – 1666 .
Neckles , H.A. , Short , F.T. , Barker , S. , and Kopp , B.S. 2005 . Disturbance of eelgrass Zostera marina by commercial mussel Mytilus edulis harvesting in Maine: dragging impacts and habitat recovery . Marine Ecology Progress Series 285 : 57 – 73 .
Nelson , K.A. , Leonard , L.A. , Posey , M.H. , Alphin , T.D. , and Mallin , M.A. 2004 . Using transplanted oyster beds to improve water quality in small tidal creeks: a pilot study . Journal of Experimental Marine Biology and Ecology 298 : 347 – 368 .
Neori , A. , Chopin , T. , Troell , M. , Buschmann , A.H. , Kraemer , G.P. , Halling , C. , Shpigel , M. , and Yarish , C. 2004 . Integrated aquaculture: rationale, evolution and state of the art empha-sizing seaweed biofi ltration in modern maricul-ture . Aquaculture 231 : 361 – 391 .
Nestlerode , J.A. 2004 . Evaluating restored oyster reefs in Chesapeake Bay: how habitat structure infl uences ecological function . Ph.D. Dissertation, VIMS, 262pp.
Newell , R.I.E. 1988 . Ecological changes in Chesapeake Bay: are they the result of overhar-vesting the eastern oyster (Crassostrea virgin-ica)? Lynch , M.P. , and Krome , E.C. (eds.), Understanding the Estuary: Advances in Chesapeake Bay Research . Chesapeake Research
Consortium Publication 129 , Gloucester Point, VA , pp. 536 – 546 .
Newell , R.I.E. 2004 . Ecosystem infl uences of natural and cultivated populations of suspen-sion - feeding bivalve molluscs: a review . Journal of Shellfi sh Research 23 : 51 – 61 .
Newell , R.I.E. 2007 . A framework for developing “ ecological carrying capacity ” mathematical models for bivalve mollusc aquaculture . Bulletin of Fisheries Research Agency 19 : 41 – 52 .
Newell , R.I.E. , and Koch , E.W. 2004 . Modeling seagrass density and distribution in response to changes in turbidity stemming from bivalve fi l-tration and seagrass sediment stabilization . Estuaries 27 : 793 – 806 .
Newell , R.I.E. , Cornwell , J.C. , and Owens , M.S. 2002 . Infl uence of simulated bivalve biodeposi-tion and microphytobenthos on sediment nitro-gen dynamics: a laboratory study . Limnology and Oceanography 47 : 1367 – 1379 .
Newell , R. , Fisher , T. , Holyoke , R. , and Cornwell , J. 2005 . Infl uence of Eastern oysters on nitrogen and phosphorus regeneration in Chesapeake Bay, USA . In: Dame , R. , and Olenin , S. (eds.), The Comparative Roles of Suspension Feeders in Ecosystems , Vol. 47 , NATO Science Series: IV — Earth and Environmental Sciences. Springer , Dordrecht, the Netherlands , pp. 93 – 120 .
Newell , R.I.E. , Kemp , W.M. , Hagy , J.D. , III , Cerco , C.F. , Testa , J.M. , and Boynton , W.R. 2007 . Top - down control of phytoplankton by oysters in Chesapeake Bay, USA: comment on Pomeroy et al. (2006) . Marine Ecology Progress Series 341 : 293 – 298 .
Nixon , S.W. 1980 . Between coastal marshes and coastal waters: a review of twenty years of spec-ulation and research on the role of salt marshes in estuarine productivity and water chemistry . In: Hamilton , P. , MacDonald , P. , and MacDonald , K.B. (eds.), Estuarine and Wetland Processes . Plenum Press , New York , pp. 437 – 525 .
Nizzoli , D. , Welsh , D.T. , Fano , E.A. , and Viaroli , P. 2006 . Impact of clam and mussel farming on benthic metabolism and nitrogen cycling, with emphasis on nitrate reduction pathways . Marine Ecology Progress Series 315 : 151 – 165 .
Norkko , A. , Hewitt , J.E. , Thrush , S.F. , and Funnell , G.A. 2006 . Conditional outcomes of facilitation by a habitat - modifying subtidal bivalve . Ecology 87 : 226 – 234 .

Impacts of native and cultured bivalves 289
Norris , K. , Bannister , R.C.A. , and Walker , P.W. 1998 . Changes in the number of oystercatchers Haematopus ostralegus wintering in the Burry Inlet in relation to the biomass of cockles Cerastoderma edule and its commercial exploi-tation. The Journal of Applied Ecology 35 : 75 – 85 .
North , E.W. , King , D.M. , Xu , J. , Hood , R.R. , Newell , R.I.E. , Paynter , K.T. , Kellogg , M.L. , Liddel , M.K. , and Boesch , D.F. 2010 . Linking optimization and ecological models in a decision support tool for oyster restoration and manage-ment . Ecological Applications 20 : 851 – 866 .
Nosho , T.Y. , and Chew , K.K. 1991 . Remote Setting and Nursery Culture for Shellfi sh Growers, Workshop Record, February 19, 1991 . Washington Sea Grant , Seattle, WA .
Nowell , A.R.M. , and Jumars , P.A. 1984 . Flow envi-ronments of aquatic benthos . Annual Review of Ecology and Systematics 15 : 303 – 328 .
Nugues , M.M. , Kaiser , M.J. , Spencer , B.E. , and Edwards , D.B. 1996 . Benthic community changes associated with intertidal oyster cultiva-tion . Aquaculture Research 27 : 913 – 924 .
O ’ Beirn , F.X. , Ross , P.G. , and Luckenbach , M.W. 2004 . Organisms associated with oysters cultured in fl oating systems in Virginia, USA . Journal of Shellfi sh Research 23 : 825 – 829 .
Offi cer , C.B. , Smayda , T.J. , and Mann , R. 1982 . Benthic fi lter feeding: a natural eutrophication control . Marine Ecology Progress Series 9 : 203 – 210 .
ORET (Oyster Restoration Evaluation Team) . 2009 . Metadata analysis of restoration and monitoring activity database . Kramer , J.G. , and Sellner , K.G. (eds.), Native Oyster (Crassostrea virginica) Restoration in Maryland and Virginia . An evaluation of lessons learned 1990 – 2007 . Maryland Sea Grant Publication #UM - SG - TS - 2009 - 02; CRC Publ. No. 09 - 168. College Park, MD . Available at http://www.oyster - restoration.org/reports/OysterResReport_web.pdf .
Orth , R.J. 1975 . Destruction of eelgrass, Zostera marina , by the cownose ray, Rhinoptera bonasus , in the Chesapeake Bay . Chesapeake Science 16 : 205 – 208 .
Orth , R.J. , Fishman , J.R. , Wilcox , D.J. , and Moore , K.A. 2002 . Identifi cation and management of fi shing gear impacts in a recovering seagrass
system in the coastal bays of the Delmarva Peninsula, USA . Journal of Coastal Research , Special Issue ( 37 ): 111 – 129 .
Orth , R.J. , Carruthers , T.J.B. , Dennison , W.C. , Duarte , C.M. , Fourqurean , J.W. , Heck , K.L. , Jr. , Hughes , R. , Kendrick , G. , Kenworthy , W.J. , Olyarnik , S. , Short , F.T. , Waycott , M. , and Williams , S.L. 2006 . A global crisis for seagrass ecosystems . Bioscience 56 : 987 – 996 .
Palacios , R. , Armstrong , D.A. , and Orensanz , J. 2000 . Fate and legacy of an invasion: extinct and extant populations of the soft - shell clam ( Mya arenaria ) in Grays Harbor (Washington) . Aquatic Conservation: Marine and Freshwater Ecosystems 10 : 279 – 303 .
Paltzat , D.L. , Pearce , C.M. , Barnes , P.A. , and McKinley , R.S. 2008 . Growth and production of California sea cucumbers ( Parastichopus califor-nicus Stimpson) co - cultured with suspended Pacifi c oysters ( Crassostrea gigas Thunberg) . Aquaculture 275 : 124 – 137 .
Paraskevi , P. , Tsapkias , M. , Apostolaki , E.T. , Tsagaraki , T. , Holmer , M. , and Karakassis , I. 2009 . Ghost nutrients from fi sh farms are trans-ferred up the food web by phytoplankton grazers . Marine Ecology Progress Series 374 : 1 – 6 .
Perry , L.M. 1936 . A marine tenement . Science 84 : 156 – 157 .
Peterson , B.J. , and Heck , K.L. 1999 . The potential for suspension feeding bivalves to increase sea-grass productivity . Journal of Experimental Marine Biology and Ecology 240 : 37 – 52 .
Peterson , B.J. , and Heck , K.L. , Jr. 2001a . Positive interactions between suspension - feeding bivalves and seagrass - a facultative mutualism . Marine Ecology Progress Series 213 : 143 – 155 .
Peterson , B.J. , and Heck , K.L. 2001b . An experi-mental test of the mechanism by which suspension feeding bivalves elevate seagrass pro-ductivity . Marine Ecology Progress Series 218 : 115 – 125 .
Peterson , C.H. , and Lipcius , R.N. 2003 . Conceptual progress towards predicting quantitative ecosys-tem benefi ts of ecological restorations . Marine Ecology Progress Series 264 : 297 – 307 .
Peterson , C.H. , Grabowski , J.H. , and Powers , S.P. 2003 . Estimated enhancement of fi sh production resulting from restoring oyster reef habitat: quantitative valuation . Marine Ecology Progress Series 264 : 251 – 256 .

290 Shellfi sh Aquaculture and the Environment
Piazza , B.P. , Banks , P.D. , and La Peyre , M.K. 2005 . The potential for created oyster shell reefs as a sustainable shoreline protection strategy in Louisiana . Restoration Ecology 13 : 499 – 506 .
Pienkowski , M.W. 1983 . Surface activity of some intertidal invertebrates in relation to tempera-ture and the foraging behaviour of their shore-bird predators . Marine Ecology Progress Series 11 : 141 – 150 .
Pillay , T.V.R. 1992 . Aquaculture and the Environment . Halsted Press , New York .
Pitta , P. , Manolis , T. , Apostolaki , E.T. , Tsagaraki , T. , Holmer , M. , and Karakassis , I. 2009 . “ Ghost nutrients ” from fi sh farms are transferred up the food web by phytoplankton grazers . Marine Ecology Progress Series 374 : 1 – 6 .
Polson , M. , Hewson , W.E. , Eemisse , D.J. , Baker , P.K. , and Zacherl , D.C. 2009 . You say con-chaphila , I say lurida : molecular evidence for restricting the Olympia oyster ( Ostrea lurida Carpenter 1864) to temperate Western North America. Journal of Shellfi sh Research 28 : 11 – 21 .
Porter , E.T. , Sanford , L.P. , and Suttles , S.E. 2000 . Gypsum dissolution is not a universal integrator of “ water motion ” . Limnology and Oceanography 45 : 145 – 158 .
Porter , E.T. , Cornwell , J.C. , Sanford , L.P. , and Newell , R.I.E. 2004 . Effect of oysters Crassostrea virginica and bottom shear velocity on benthic - pelagic coupling and estuarine water quality . Marine Ecology Progress Series 271 : 61 – 75 .
Posey , M.H. 1986 . Predation on a burrowing shrimp: distribution and community conse-quences . Journal of Experimental Marine Biology and Ecology 103 : 143 – 161 .
Posey , M.H. 1988 . Community changes associated with the spread of an introduced seagrass, Zostera japonica . Ecology 69 : 974 – 983 .
Posey , M.H. , Alphin , T.D. , Powell , C.M. , and Townsend , E. 1999 . Use of oyster reefs as habitat for epibenthic fi sh and decapods . In: Luckenbach , M.W. , Mann , R. , and Wesson , J.E. (eds.), Oyster Reef Habitat Restoration: A Synopsis and Synthesis of Approaches . VIMS Press , Gloucester Point, VA , pp. 229 – 237 .
Powell , E.N. , and Klinck , J.M. 2007 . Is oyster shell a sustainable estuarine resource? Journal of Shellfi sh Research 26 : 181 – 194 .
Powell , E.N. , Kraeuter , J.N. , and Ashton - Alcox , K.A. 2006 . How long does oyster shell last on an oyster reef? Estuarine, Coastal and Shelf Science 69 : 531 – 542 .
Powell , E.N. , Ashton - Alcox , K.A. , Kraeuter , J.N. , Ford , S.E. , and Bushek , D. 2008 . Long - term trends in oyster population dynamics in Delaware Bay: regime shifts and response to disease . Journal of Shellfi sh Research 27 : 729 – 755 .
Powers , M.J. , Peterson , C.H. , Summerson , H.C. , and Powers , S.P. 2007 . Macroalgal growth on bivalve aquaculture netting enhances nursery habitat for mobile invertebrates and juvenile fi shes . Marine Ecology Progress Series 339 : 109 – 122 .
Powers , S.P. , Peterson , C.H. , Grabowski , J.H. , and Lenihan , H.S. 2009 . Success of constructed oyster reefs in no - harvest sanctuaries: implica-tions for restoration . Marine Ecology Progress Series 389 : 159 – 170 .
Quammen , M.L. 1981 . Use of exclosures in studies of predation by shorebirds on intertidal mud-fl ats . The Auk 98 : 812 – 817 .
Quammen , M.L. 1982 . Infl uence of subtle substrate differences on feeding by shorebirds on intertidal mudfl ats . Marine Biology 71 : 339 – 343 .
Quammen , M.L. 1984 . Predation by shorebirds, fi sh, and crabs on invertebrates in intertidal mud-fl ats: an experimental test . Ecology 65 : 529 – 537 .
Read , P. , and Fernandes , T.F. 2003 . Management of environmental impacts of marine aquaculture in Europe . Aquaculture 226 : 139 – 163 .
Read , A.J. , Drinker , P. , and Northridge , S. 2006 . Bycatch of marine mammals in U.S. and global fi sheries . Conservation Biology 20 : 163 – 169 .
Reise , K. 1998 . Pacifi c oysters invade mussel beds in the European Wadden Sea . Senckenbergiana Maritima 28 : 167 – 175 .
Reise , K. , Bouma , T.J. , Olenin , S. , and Ysebaert , T. 2009 . Coastal habitat engineers and the biodi-versity in marine sediments . Helgoland Marine Research 63 : 1 – 2 .
Reusch , T.B.H. , Chapman , A.R.O. , and Groger , J.P. 1994 . Blue mussels Mytilus edulis do not inter-fere with eelgrass Zostera marina but fertilize shoot growth through biodeposition . Marine Ecology Progress Series 108 : 265 – 282 .
Rice , M.A. 2000 . A review of shellfi sh restoration as a tool for coastal water quality management . Environment Cape Cod 3 : 1 – 8 .

Impacts of native and cultured bivalves 291
Richardson , N.F. , Ruesink , J.L. , Naeem , S. , Hacker , S.D. , Tallis , H.M. , Dumbauld , B.R. , and Wisehart , L.M. 2008 . Bacterial abundance and aerobic microbial activity across natural and oyster aquaculture habitats during summer con-ditions in a northeastern Pacifi c estuary . Hydrobiologia 596 : 269 – 278 .
Robinson , A.M. 1997 . Molluscan fi sheries in Oregon: past, present and future . In: Mackenzie , C.L.J. , Burrell , V.G.J. , Rosenfi eld , A. , and Hobart , W.L. (eds.), The History, Present Condition, and Future of the Molluscan Fisheries of North and Central America and Europe , Vol. 2 , Pacifi c Coast and Supplemental Topics. U.S. Dept. Commerce, NOAA Tech. Rep. NMFS 128. Seattle, WA: NOAA , U.S. Department of Commerce , pp. 75 – 87 .
Rodgers , J.A. , and Smith , H.T. 1997 . Buffer zone distances to protect foraging and loafi ng water-birds from human disturbance in Florida . Wildlife Society Bulletin 25 : 139 – 145 .
Rodney , W.S. , and Paynter , K.T. 2006 . Comparisons of macrofaunal assemblages on restored and non - restored oyster reefs in mesohaline regions of Chesapeake Bay in Maryland . Journal of Experimental Marine Biology and Ecology 335 : 39 – 51 .
Rothschild , B.J. , Ault , J.S. , Goulletquer , P. , and H é ral , M. 1994 . Decline of the Chesapeake Bay oyster population: a century of habitat destruc-tion and overfi shing . Marine Ecology Progress Series 111 : 29 – 39 .
Roycroft , D. , Kelly , T.C. , and Lewis , L.J. 2006 . Behavioural interactions of seabirds with sus-pended mussel longlines . Aquaculture International 15 : 25 – 36 .
Ruesink , J.L. , Lenihan , H.S. , Trimble , A.C. , Heiman , K.W. , Micheli , F. , Byers , J.E. , and Kay , M.C. 2005 . Introduction of non - native oysters: ecosystem effects and restoration implications . Annual Review of Ecology and Systematics 36 : 643 – 689 .
Ruesink , J.L. , Feist , B.E. , Harvey , C.J. , Hong , J.S. , Trimble , A.C. , and Wisehart , L.M. 2006 . Changes in productivity associated with four introduced species: ecosystem transformation of a “ pristine ” estuary . Marine Ecology Progress Series 311 : 203 – 215 .
Ruesink , J.L. , Hacker , S.D. , Dumbauld , B.R. , and Harbell , S. 2009 . Scale - dependent and indirect
effects of fi lter feeders on eelgrass: understand-ing complex ecological interactions to improve environmental impacts of aquaculture . Dept. of Biology, University of Washington. Final Report to the Western Regional Aquaculture Center Seattle, Washington, 25pp.
Ruiz , G.M. , Fofonoff , P.W. , Carlton , J.T. , Wonham , W.J. , and Hines , A.H. 2000 . Invasion of coastal marine communities in North America: apparent patterns, processes, and biases . Annual Review of Ecology and Systematics 31 : 481 – 531 .
Rumrill , S.S. , and Poulton , V.K. 2004 . Ecological role and potential impacts of molluscan shellfi sh culture in the estuarine environment of Humboldt Bay, Ca . Oregon Department of State Lands, Final Annual Report to the Western Regional Aquaculture Center, 22pp.
Safriel , U.N. 1975 . The role of vermetid gastropods in the formation of Mediterranean and Atlantic reefs . Oecologia 20 : 85 – 101 .
Salisbury , J. , Green , M. , Hunt , C. , and Campbell , J. 2008 . Coastal acidifi cation by rivers: a new threat to shellfi sh? Eos Transactions AGU 89 : 513 – 514 .
Sanchez , M.I. , Green , A.J. , and Castellanos , E.M. 2006 . Spatial and temporal fl uctuations in pres-ence and use of chironomid prey by shorebirds in the Odiel saltpans, south - west Spain . Hydrobiologia 567 : 329 – 340 .
Schulte , D.M. , Burke , R.P. , and Lipcius , R.N. 2009 . Unprecedented restoration of a native oyster metapopulation . Science 325 : 1124 – 1128 .
Sebens , K.P. 1991 . Habitat structure and commu-nity dynamics in marine benthic systems . In: Bell , S.S. , McCoy , E.D. , and Mushinsky , H.R. (eds.), Habitat Structure: the Physical Arrangement of Objects in Space . Chapman and Hall , London , pp. 211 – 234 .
Seed , R. 1992 . Systematics evolution and distribu-tion of mussels belonging to the genus Mytilus: an overview . American Malacological Bulletin 9 : 123 – 137 .
Seed , R. , and Suchanek , T.H. 1992 . Population and community ecology of Mytilus . In: Gosling , E.M. (ed.), The Mussel Mytilus: Ecology, Physiology, Genetics, and Culture . Elsevier Science , Amsterdam , pp. 87 – 169 .
Sellers , M.A. , and Stanley , J.G. 1984 . Species pro-fi les: life histories and environmental require-ments of coastal fi shes and invertebrates (north

292 Shellfi sh Aquaculture and the Environment
Atlantic) - American Oyster . U.S. F.W.S. Offi ce of Biological Services Report No. FWS/OBS - 82/11.23, and USACE Report No. TR EL - 82 - 4, Washington, DC.
Semmens , B.X. 2008 . Acoustically derived fi ne - scale behaviors of juvenile Chinook salmon associated with intertidal benthic habitats in an estuary . Canadian Journal of Fisheries and Aquatic Sciences 65 : 2053 – 2062 .
Shaw , W.N. 1997 . The shellfi sh industry of California - Past, present and future . In: Mackenzie , C.L.J. , Burrell , V.G.J. , Rosenfi eld , A. , and Hobart , W.L. (eds.), The History, Present Condition, and Future of the Molluscan Fisheries of North and Central America and Europe , Vol. 2 , Pacifi c Coast and Supplemental Topics. U.S. Dept. Commerce, NOAA Tech. Rep. NMFS 128. Seattle, WA: NOAA , U.S. Department of Commerce , pp. 57 – 74 .
Shumway , S.E. , and Kraeuter , J.N. (eds.) 2004 . Molluscan shellfi sh research and management: charting a course for the future . Final proceed-ings from the Cooperative Research and Information Institute (CRII) Workshop, Charleston, SC, 156pp .
Simenstad , C.A. , and Fresh , K.L. 1995 . Infl uence of intertidal aquaculture on benthic communities in Pacifi c Northwest estuaries: scales of distur-bance . Estuaries 18 : 43 – 70 .
Smaal , A.C. , Kater , B.J. , and Wijsman , J. 2009 . Introduction, establishment and expansion of the Pacifi c oyster Crassostrea gigas in the Oosterschelde (SW Netherlands) . Helgoland Marine Research 63 : 75 – 83 .
Smith , G.F. , Bruce , D.G. , Roach , E.B. , Hansen , A. , Newell , R.I.E. , and McManus , A.M. 2005 . Habitat conditions of mesohaline oyster bars in the Maryland Chesapeake Bay: an assessment of 40 years of oyster management . North American Journal of Fisheries Management 25 : 1569 – 1590 .
Smith , K.A. , North , E.W. , Shi , F. , Chen , S. , Hood , R.R. , Koch , E.W. , and Newell , R.I.E. 2009 . Modeling the effects of oyster reefs and break-waters on seagrass growth . Estuaries and Coasts 32 : 748 – 757 .
Smyth , D. , Roberts , D. , and Browne , L. 2009 . Impacts of unregulated harvesting on a recover-ing stock of native oysters ( Ostrea edulis ) . Marine Pollution Bulletin 58 : 916 – 922 .
Soniat , T.M. , Finelli , C.M. , and Ruiz , J.T. 2004 . Vertical structure and predator refuge mediate oyster reef development and community dynam-ics . Journal of Experimental Marine Biology and Ecology 310 : 163 – 182 .
Sousa , R. , Guti é rrez , J.L. , and Aldridge , D.C. 2009 . Non - indigenous invasive bivalves as ecosystem engineers . Biology Invasions 11 : 2367 – 2385 .
Southworth , M. , and Mann , R. 1998 . Oyster reef broodstock enhancement in the Great Wicomico River, Virginia , Journal of Shellfi sh Research 17 : 1101 – 1114 .
Stal , L.J. 2010 . Microphytobenthis as a biogeomor-phological force in intertidal sediment stabiliza-tion . Ecological Engineering 36 : 236 – 245 .
Stanley , D.W. , and Sellers , M.A. 1986 . Species profi le: life histories and environmental require-ments of coastal fi shes and invertebrates (Gulf of Mexico) - American Oyster . U.S. Fish Wildl. Serv. Biol Rep. 82(11.64) U.S. Army Corps of Engineers, TR EL - 82 - 4, 25pp.
Steele , E.N. 1964 . The Immigrant Oyster (Ostrea Gigas) Now Known as the Pacifi c Oyster . Warren ’ s Quick Print , Olympia, WA .
Steimle , F.W. , and Zetlin , C. 2000 . Reef habitats in the Middle Atlantic Bight: abundance, distribu-tion, associated biological communities, and fi shery resource use . Marine Fisheries Review 62 : 24 – 42 .
Stempien , J.A. 2007 . Detecting avian predation on bivalve assemblages using indirect methods . Journal of Shellfi sh Research 26 : 271 – 280 .
Stillman , R.A. , West , A.D. , Caldow , R.W.G. , and le V. dit Durell , S.E.A. 2007 . Predicting the, effect of disturbance on coastal birds . Ibis 149 : 73 – 81 .
Stokesbury , K.D.E. 2002 . Estimation of sea scallop abundance in closed areas of Georges Bank, USA . Transactions of the American Fisheries Society 131 : 1081 – 1092 .
Stoner , A.W. , and Titgen , R.H. 2003 . Biological structures and bottom type infl uence habitat choices made by Alaska fl atfi shes . Journal of Experimental Marine Biology and Ecology 292 : 43 – 59 .
Stunz , G.W. , Minello , T.J. , and Rozas , L.R. 2010 . Relative value of oyster reef as habitat for estuarine nekton in Galveston Bay, Texas . Marine Ecology Progress Series 406 : 147 – 159 .

Impacts of native and cultured bivalves 293
Suchanek , T.H. 1985 . Mussels and their role in structuring rocky shore communities . In: Moore , P.G. , and Seed , R. (eds.), The Ecology of Rocky Coasts: Essays Presented to J.R. Lewis . Columbia Press , New York , pp. 70 – 96 .
Tallis , H.M. , Ruesink , J.L. , Dumbauld , B. , Hacker , S. , and Wisehart , L.M. 2009 . Oysters and aqua-culture practices affect eelgrass density and pro-ductivity in a Pacifi c Northwest estuary . Journal of Shellfi sh Research 28 : 251 – 261 .
Tallman , J.C. , and Forrester , G.E. 2007 . Oyster grow - out cages function as artifi cial reefs for temperate fi shes . Transactions of the American Fisheries Society 136 : 790 – 799 .
Talman , S.G. , Norkko , A. , Thrush , S.F. , and Hewitt , J.E. 2004 . Habitat structure and the survival of juvenile scallops Pecten novaezelandiae : com-paring predation in habitats with varying com-plexity . Marine Ecology Progress Series 269 : 197 – 207 .
Tenore , K.R. , and Gonzalez , N. 1975 . Food chain patterns in the R í a de Arosa, Spain: an area of intense mussel aquaculture . In: Persoone , G. , and Jaspers , E. (eds.), 10 th European Symposium on Marine Biology , Vol. 2 . Universal Press , Wetteren, Belgium , pp. 601 – 619 .
Tenore , K.R. , Browne , M.G. , and Chesney , E.J. 1974 . Polyspecies aquaculture systems: the detri-tal trophic level . Journal of Marine Research 32 : 425 – 432 .
Tenore , K.R. , Boyer , L.F. , Cal , R.M. , Corral , J. , Garcia - Fernandez , C. , Gonzalez , N. , Gonzalez - Gurriaran , E. , Hanson , R.B. , Iglesias , J. , Krom , M. , Lopez - Jamar , E. , McClain , J. , Pamatmat , M.M. , Perez , A. , Rhoads , D.C. , de Santiago , G. , Tietjen , J. , Westrich , J. , and Windom , H.L. 1982 . Coastal upwelling in the Rias Bajas, NW Spain: contrasting the benthic regimes of the Rias de Arosa and de Muros . Journal of Marine Research 40 : 701 – 772 .
Tevesz , M.J.S. , and McCall , P.L. (eds.). 1983 . Biotic Interactions in Recent and Fossil Benthic Communities . Plenum Press , New York .
Thayer , G.W. (ed.) 1992 . Restoring the Nation ’ s Marine Environment . Maryland Sea Grant College Program , College Park, MD .
Thayer , G.W. , McTigue , T.A. , Bellmer , R.J. , Burrows , F.M. , Merkey , D.H. , Nickens , A.D. , Lozano , S.J. , Gayaldo , P.F. , Polmateer , P.J. , and Pinit , P.T. 2003 . Science - Based Restoration
Monitoring of Coastal Habitats, Volume 1: A Framework for Monitoring Plans under the Estuaries and Clean Waters Act of 2000 (Public Law 160 - 457) . NOAA Coastal Ocean Program Decision Analysis Series No. 23. NOAA National Centers for Coastal Ocean Science , Silver Spring, MD .
Thayer , G.W. , McTigue , T.A. , Salz , R.J. , Merkey , D.H. , Burrows , F.M. , and Gayaldo , P.F. (eds.) 2005 . Science - Based Restoration Monitoring of Coastal Habitats, Volume 2: Tools for Monitoring Coastal Habitats . NOAA Coastal Ocean Program Decision Analysis Series No. 23. NOAA National Centers for Coastal Ocean Science , Silver Spring, MD .
Thieltges , D.W. , Strasser , M. , and Reise , K. 2006 . How bad are invaders in coastal waters? The case of the American slipper limpet Crepidula fornicata in western Europe . Biological Invasions 8 : 1673 – 1680 .
Thom , R.M. , Southard , S.L. , Borde , A.B. , and Stoltz , P. 2008 . Light requirements for growth and survival of eelgrass ( Zostera marina L.) in Pacifi c Northwest (USA) estuaries . Estuaries and Coasts 31 : 969 – 980 .
Thompson , D.S. 1995 . Substrate additive studies for the development of hardshell clam habitat in Washington State: an analysis of effects on recruitment, growth and survival of the Manila clam, Tapes philippinarum , and on the species diversity and abundance of existing benthic organisms . Estuaries 18 : 91 – 107 .
Tolley , S.G. , and Volety , A.K. 2005 . The role of oysters in habitat use of oyster reefs by resident fi shes and decapod crustaceans . Journal of Shellfi sh Research 24 : 1007 – 1012 .
Townsend , C.H. 1896 . The transplanting of eastern oysters to Willapa Bay, Washington with notes on the native oyster industry . U.S. Commisioner of Fisheries, 193 – 202pp.
Trianni , M.S. 1995 . The infl uence of commercial oyster culture activities on the benthic infauna of Arcata Bay . M.S. thesis, Humboldt State University, Arcata, CA.
Triplet , P. , Stillman , R.A. , and Goss - Custard , J.D. 1999 . Prey abundance and the strength of inter-ference in a foraging shorebird . The Journal of Animal Ecology 68 : 254 – 265 .
Tsai , C. , Trimble , A.C. , and Ruesink , J.L. 2010 . Interactions between two introduced species:

294 Shellfi sh Aquaculture and the Environment
Zostera japonica (dwarf eelgrass) facilitates itself and reduces condition of Ruditapes phillipina-rum (Manila clam) on intertidal fl ats . Marine Biology 157 : 1929 – 1936 .
Tyler , R.M. 2007 . Effects of coverage by benthic seaweed mats on (northern quahog = hard clam) Mercenaria mercenaria in a eutrophic estuary . Journal of Shellfi sh Research 26 : 1021 – 1028 .
Valentine , J.F. , and Heck , K.L. , Jr. 1993 . Mussels in seagrass meadows: their infl uence on macroin-vertebrate abundance and secondary production in the Northern Gulf of Mexico . Marine Ecology Progress Series 96 : 63 – 74 .
Van Dolah , R.F. , Holland , A.F. , Coen , L.D. , Ringwood , A.H. , Levisen , M.V. , Maier , P.P. , Scott , G.I. , Leight , A.K. , Bobo , Y. , and Richardson , D. 1999 . Biological resources, report on the status of Broad Creek/Okatee River Systems . DHEC - MRRI - NOAA - Charleston.
Volety , A.K. , Savarese , M. , Tolley , S.G. , Arnold , W.S. , Sime , P. , Goodman , P. , Chamberlain , R.H. , and Doering , P.H. 2009 . Eastern oysters ( Crassostrea virginica ) as an indicator for resto-ration of Everglades ecosystems . Ecological Indicators 9 ( Suppl. 1 ): S120 – S136 .
Waldbusser , G.G. , Voigt , E.P. , Bergschneider , H. , Green , M.A. , and Newell , R.I.E. 2011 . Biocalcifi cation in the Eastern oyster ( Crassostrea virginica ) in relation to long - term trends in Chesapeake Bay pH. Estuaries and Coasts 34 : 221 – 231 .
Wall , C.C. , Peterson , B.J. , and Gobler , C.J. 2008 . Facilitation of seagrass Zostera marina produc-tivity by suspension - feeding bivalves . Marine Ecology Progress Series 357 : 165 – 174 .
Ward , D.H. , Morton , A. , Tibbitts , T.L. , Douglas , D.C. , and Carrera - Gonzalez , E. 2003 . Long - term change in eelgrass distribution at Bahia San Quintin, Baja California, Mexico, using satellite imagery . Estuaries 26 : 1529 – 1539 .
Warnock , N. , Haig , S.M. , and Oring , L.W. 1998 . Monitoring species richness and abundance of shorebirds in the Western Great Basin . The Condor 100 : 589 – 600 .
Watson , D. , Shumway , S.E. , and Whitlatch , R.B. 2009 . Biofouling and the shellfi sh industry . In: Shumway , S.E. , and Rodrick , G.E. (eds.), Shellfi sh Quality and Safety . Woodhead Publishing , Cambridge, England , pp. 317 – 336 .
Waycott , M. , Duarte , C.M. , Carruthers , T.J.B. , Orth , R.J. , Dennison , W.C. , Olyarnik , S. , Calladine , A. , Fourqurean , J.W. , Heck , K.L. , Jr. , Hughes , A.R. , Kendrick , G.A. , Kenworthy , J.W. , Short , F.T. , and Williams , S.L. 2009 . Accelerating loss of seagrasses across the globe threatens coastal ecosystems . PNAS 106 : 12377 – 12381 .
Wei , Z. , Shi , H. , Chen , S. , and Zhu , M. 2009 . Benefi t and cost analysis of mariculture based on ecosystem services . Ecological Economics 68 : 1626 – 1632 .
Weise , A.M. , Cromey , C.J. , Callier , M.D. , Archambault , P. , Chamberlain , J. , and McKindsey , C.W. 2009 . Shell fi sh - DEPOMOD: modelling the biodeposition from suspended shellfi sh aquaculture and assessing benthic effects . Aquaculture 288 : 239 – 253 .
Wells , H.W. 1961 . The fauna of oyster beds, with special reference to the salinity factor . Ecological Monographs 31 : 266 – 329 .
Weschler , J.F. 2004 . Assessing the relationship between the ichthyofauna and oyster maricul-ture in a shallow embayment, Drakes Estero, Point Reyes National Seashore . M.S. thesis, University of California, Davis.
Weston , D.P. 1990 . Quantitative examination of macrobenthic community changes along an organic enrichment gradient . Marine Ecology Progress Series 61 : 233 – 244 .
White , J. , Ruesink , J.L. , and Trimble , A.C. 2009 . The nearly forgotten oyster: Ostrea lurida Carpenter 1864 (Olympia oyster) history and management in Washington state . Journal of Shellfi sh Research 28 : 43 – 49 .
Whiteley , J. , and Bendell - Young , L. 2007 . Ecological implications of intertidal mariculture: observed differences in bivalve community structure between farm and reference sites . The Journal of Applied Ecology 44 : 495 – 505 .
Wildish , D.J. , and Kristmanson , D.D. 1997 . Benthic Suspension Feeders and Flow . Cambridge University Press , New York .
Williams , S.W. , and Heck , K.L. , Jr. 2001 . Seagrass communities . In: Bertness , M. , Gaines , S. , and Hay , M. (eds.), Marine Community Ecology . Sinauer Press , Sunderland, MA , pp. 317 – 337 .
Wisehart , L.M. , Dumbauld , B.R. , Ruesink , J.L. , and Hacker , S.D. 2007 . Importance of eelgrass early life history stages in response to oyster

Impacts of native and cultured bivalves 295
aquaculture disturbance . Marine Ecology Progress Series 344 : 71 – 80 .
Wrange , A. - L.J.V. , Harkestad , L.S. , Strand , Ø . , Lindegarth , S. , Christensen , H.T. , Dolmer , P. , Kristensen , P.S. , and Mortensen , S. 2010 . Massive settlements of the Pacifi c oyster, Crassostrea gigas , in Scandinavia . Biology Invasions 12 : 1145 – 1152 .
Yasue , M. 2005 . The effects of human presence, fl ock size and prey density on shorebird foraging rates . Journal of Ethology 23 : 199 – 204 .
Yuan , X. , Zhang , M. , Liang , Y. , Liu , D. , and Guan , D. 2010 . Self - pollutant loading from a suspen-
sion aquaculture system of Japanese scallop ( Patinopecten yessoensis ) in the Changhai sea area, Northern Yellow Sea of China . Aquaculture 304 : 79 – 87 .
Yund , P.O. , Gaines , S.D. , and Bertness , M.D. 1991 . Cylindrical tube traps for larval sampling . Limnology and Oceanography 36 : 1167 – 1177 .
Zimmerman , R. , Minello , T. , Baumer , T. , and Castiglione , M. 1989 . Oyster reef as habitat for estuarine macrofauna . NOAA Tech. Mem. NMFS - SEFC - 249, 16pp.