senescence and aging in plants || calcium and senescence
TRANSCRIPT

Calcium and Senescence
B. W. Poovaiah Department of Horticulture
Washington State University Pullman, Washington
I. Introduction II. Calcium and Hormone Interactions
III. Cytosolic and Apoplastic Roles of Calcium A. Cell Walls and Membranes B. Calcium and Calmodulin C. Protein Phosphorylation
IV. Role of Inositol Phospholipids in Calcium Messenger System V. Conclusion
References
I. INTRODUCTION
A major re-evaluation is occurring in our thinking about the role of calcium ions in mediating diverse physiological processes in plants. It is clear that calcium has an important role in controlling senescence. The influence of calcium on cell wall structure and membrane integrity has been well documented for many years. Recent investigations suggest that calcium plays a central role in regulating various metabolic processes in plants (Reddy and Poovaiah, 1987).
The possible participation of calcium in senescence in plants may be inferred from the fact that calcium is widely known to play a major role in
369
Copyright © 1988 by Academic Press, Inc. SENESCENCE AND AGING IN PLANTS All rights of reproduction in any form reserved.

370 Β. W. Poovaiah
membrane structure and function (Jones and Lunt, 1967). Its importance in the regulation of ion transport is well established (Epstein, 1961). Its effects on the maintenance of RNA and protein levels have been described by Trewavas (1970, 1972), and these components are considered to be central indices of senescence. Each of the parameters of senescence, such as chlorophyll loss, protein decrease, and increases in apparent free space and hydraulic conductivity, is affected by changes in the calcium status of the tissue (Poovaiah and Leopold, 1973a; Poovaiah, 1987).
The function of C a 2 + as a second messenger in animal cells has been recognized for years. Only in the last 10 years have plant scientists come to recognize the importance of Ca2+ in the regulation of plant metabolism. The role of calcium in signal transduction and cell function is beginning to be understood at the molecular level. The discovery of the calcium-binding protein, calmodulin (Cheung, 1980; Anderson and Cormier, 1978), and recent investigations demonstrating changes in intracellular calcium concentrations in response to primary stimuli such as light and hormones have provided the basis for suggesting that calcium serves as a second messenger in plants as well (Roux and Slocum, 1982; Marme and Dieter, 1983; Veluth-ambi and Poovaiah, 1984a; Poovaiah, 1985; Hepler and Wayne, 1985; Poovaiah and Reddy, 1987; Poovaiah et al, 1987a).
Calcium distribution at the cellular level plays a critical role in cell function. A high level of free C a 2 + in the cytoplasm is injurious to the cell. At such a level, C a 2 + reacts with inorganic phosphate to form an insoluble precipitate. Thus, if cytosolic Ca2"1" concentrations were allowed to reach the millimolar levels present in the extracellular space, phosphate-based energy metabolism would be seriously inhibited. At submicromolar levels of C a 2 + , its reaction with inorganic phosphate is negligible (Kretsinger, 1977). To maintain these submicromolar levels of intracellular calcium in the presence of millimolar levels in the extracellular space plants require the active pumping of calcium out of the cytoplasm. It is believed that deterioration of this active pumping mechanism can lead to senescence and cell death.
The major barrier to the flow of information into the cell is the plasma membrane, where transduction mechanisms translate external signals, such as hormones and light, into internal signals. A transient flux of C a 2 + appears to be the trigger allowing the chemical messages to be relayed to the biochemical machinery within the cell. Cells have evolved an elaborate system of proteins that interact with the calcium ion, controlling the transmission and reception of the intracellular message. Unraveling the intricacies of these interactions may lead to better control over intracellular C a 2 + concentration, a possibility that has broad implications for the control of plant growth and development.

11. Calcium and Senescence 371
II. CALCIUM AND HORMONE INTERACTIONS
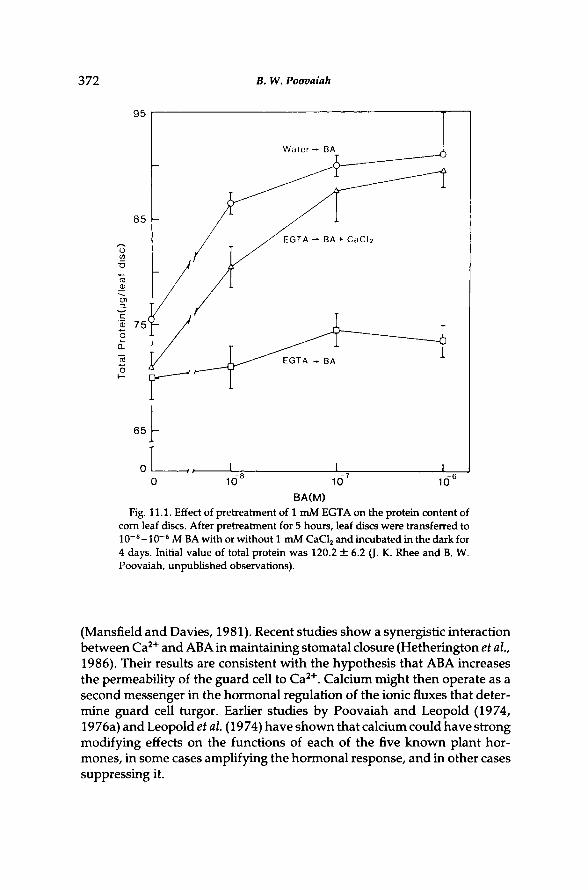
Since the discovery by Richmond and Lang (1957) that cytokinins could delay leaf senescence, it has become increasingly evident that each of the five known plant hormones is capable of altering senescence. A general concept of endogenous regulation of senescence by hormones is, however, made less attractive by the difficulties of producing major alterations in senescence development through application of hormones to intact plants. Earlier investigations have shown that calcium ions can significantly alter some aspects of senescence development and abscission in leaves (Poo-vaiah, 1979a, 1986; Poovaiah and Leopold, 1973a,b; Poovaiah and Ras-mussen, 1973a,b; Suwwan and Poovaiah, 1978). Recently, the role of calcium in mediating the cytokinin effect was studied by first depleting calcium from corn leaf discs by an EGTA pretreatment and then transferring them to a medium containing benzyladenine (BA) with or without calcium. After the EGTA-pretreatment, BA was no longer effective in delaying the loss of protein (Fig. 11.1). However, the cytokinin effect was restored by the addition of calcium. These results clearly point to a central role for calcium in hormone action as it relates to senescence.
Earlier investigations by Saunders and Hepler (1982) have shown that the calcium ionophore A 23187 could mimic the effect of cytokinin in the bud formation of Funaria. Furthermore, their results indicate that stimulus-response coupling involves an influx of calcium into the cytoplasm. Evidence for a calcium influx into the cytoplasm in response to a stimulus was obtained by Williamson and Ashley (1982), who demonstrated that inhibition of cytoplasmic streaming in Chara cells is associated with an increase in cytoplasmic C a 2 + concentration in response to action potentials. Recently, calcium redistribution during cell division (Keith et al, 1985) and tuberiza-tion (Balamani et al, 1986) have been observed.
Leakiness of membranes deprived of exogenous C a 2 + is observed in almost all kinds of plant tissues. The concept that membrane failure may be involved in leaf senescence was first suggested by Sacher (1957). Eilam (1965) showed that senescence of bean leaves was associated with a leakage of potassium from the leaves and an associated increase in apparent free space. The most conspicuous role for C a 2 + in the apoplast is the maintenance of the integrity of the plasma membrane. Poovaiah and Leopold (1973a) have shown an interaction of calcium and cytokinin in altering membrane permeability during the progression of senescence. Thimann and Satler (1979) suggest that stomatal apperture is the principal cytokinin-sensitive factor in delaying senescence. The induction of stomatal closure can be driven by abscisic acid (ABA) when water supply is suboptimal
372
9 5
Β. W. Poovaiah
Ο 10 10 10^
BA(M)
Fig. 11.1. Effect of pretreatment of 1 mM EGTA on the protein content of corn leaf discs. After pretreatment for 5 hours, leaf discs were transferred to 1 0 ~ 8 - 1 0 ~ 6 Μ Β A with or without 1 mM CaCl 2 and incubated in the dark for 4 days. Initial value of total protein was 120.2 ± 6.2 (J. K. Rhee and B. W. Poovaiah, unpublished observations).
(Mansfield and Davies, 1981). Recent studies show a synergistic interaction between C a 2 + and ABA in maintaining stomatal closure (Hetherington et al, 1986). Their results are consistent with the hypothesis that ABA increases the permeability of the guard cell to C a 2 + . Calcium might then operate as a second messenger in the hormonal regulation of the ionic fluxes that determine guard cell turgor. Earlier studies by Poovaiah and Leopold (1974, 1976a) and Leopold et al. (1974) have shown that calcium could have strong modifying effects on the functions of each of the five known plant hormones, in some cases amplifying the hormonal response, and in other cases suppressing it.

11. Calcium and Senescence 373
III. CYTOSOLIC AND APOPLASTIC ROLES OF CALCIUM
A. Cell Walls and Membranes
Calcium is essential for structural integrity of both membranes and cell walls (Jones and Lunt, 1967; Poovaiah, 1985). In plants, the major portion of the C a 2 + occurs in the apoplastic compartment, primarily complexed with cell wall moieties and the plasma membrane. Rossignol et al (1977) have estimated that at least 6 0 % of the total C a 2 + in plants is associated with the cell wall and the remaining with the membranes and soluble fraction. The importance of C a 2 + in cell-to-cell adhesion is well recognized (Demarty et al, 1984). The cementing effect is due primarily to the Ca-pectate of the middle lamella laid down during cytokinesis. The Ca-pectate complex is essential for the maintenance of cell wall structure, especially in fruits and other storage organs. Under conditions of calcium deficiency, membranes become leaky, cellular compartmentation is lost (Marinos, 1962), and C a 2 +
cross-linkage with the pectin in the middle lamella is weakened. The living cell surface is able to accommodate a rather wide variation in composition and pH of the apoplastic solution only in the presence of adequate C a 2 + in the extracellular space. Concentrations of 1 to 5 mM Ca2"4" are essential to protect the plasma membrane from the deleterious effects of low pH, salinity, toxic ions, and nutrient imbalance (Hanson, 1983). Without such protection, the plasma membrane fails to discriminate between ions, the active pumping mechanism fails, and senescence is accelerated.
There is evidence that calcium alters the actual architecture of membranes. Its introduction into natural (Paliyath et al, 1984) or artificial membranes (Gary-Bobo, 1970) results in an enormous change in fluidity and water permeability. Earlier studies in our laboratory showed that the fluorescence polarization of membrane-embedded diphenyl hexatriene increased in membranes from rapidly senescing apples as compared to normal cold-stored apples (Paliyath et al, 1984). This indicates an increase in mi-croviscosity of apple membranes with senescence.
There is now considerable evidence that the rate of senescence in some fruits and vegetables is influenced by the calcium content of the tissue. The calcium effect can be clearly seen in fruits such as apples, where senescence is gradual and spans a period of months. In addition, vacuum infiltration of apples with calcium chloride delays senescence (Poovaiah and Moulton, 1982). It has been suggested that in apples, the bulk of the infiltrated calcium remains in the cell wall region and results in firmer fruit (Poovaiah et al, 1988). To understand the process of softening in fruits, researchers have observed changes in cell wall composition and the activity of cell wall-degrading enzymes during ripening. The softening of the apple fruit appears to be due to the transfer of divalent cations, particularly calcium,

374
Fig. 11.2. Electron micrographs showing the ultrastructure of cell walls of control and calcium-treated apple fruits. Fruits were infiltrated with calcium chloride soon after harvest (for details refer to Poovaiah and Moulton, 1982) and stored for 6 to 8 months. (A) Cell wall of a control fruit showing the dissolution of middle lamella (arrow). (B) Cell wall of a calcium-treated fruit showing intact darkly stained middle lamella (arrow).

Fig. 11.2. (continued) (C) Cell wall of a control fruit showing severe breakdown of the middle lamella and the appearance of empty regions (arrow). (D) Cell wall of a calcium-treated fruit showing tightly packed and fibrillar middle lamella.
375

376 Β. W. Poovaiah
from the cell wall into storage compartments inside the cell. Calcium has long been thought to be important as a cross-linking component between polygalacturonide chains in plant cell walls (Knee, 1973; Demarty et al, 1984). Knee (1973) also found that ripening apples tend to lose galactose residues from the cell wall fractions, and this observation has since been confirmed by other workers (Bartley, 1974; Wallner, 1978). Doesburg (1957) reported the solubilization of pectin during apple ripening and proposed that it may be caused by reduced levels of C a 2 + in the cell walls. Mizrahi and Kopeliovitch (1983) found the pectin solubilization in tomatoes to be positively related to softening. Buescher and Hobson (1982) reported that calcium inhibits polygalacturonate degradation in tomatoes. Degradation was accelerated when EDTA or citrate was used to chelate calcium in cell wall tissue. Conway and Sams (1984) suggested that calcium treatment of apples results in increased firmness and less soluble pectin through cooperative binding with free carboxyl groups on polygalacturonate polymers in the cell wall and middle lamella. There are some indications that calcium actually promotes the synthesis of cell wall polymers. Ray and Baker (1965) observed increased incorporation of labeled glucose into cell wall components of plant tissues treated with calcium. Research in our laboratory indicates that calcium promotes the activity of /?-glucan synthase, a key enzyme involved in cell wall synthesis (Poovaiah and Veluth-ambi, 1986; Paliyath and Poovaiah, 1988). Cell wall synthesis and degradation may occur simultaneously during senescence. Cell wall synthesis may decline, while the net cell wall degradation leads to softening.
Postharvest calcium treatments were performed using apples in order to study their effects on changes in proteins and ultrastructure of the cell wall. Some qualitative differences in cell wall proteins were observed during prolonged storage periods, and dramatic changes in cell wall structure were noted (Fig. 11.2) (G. M. Glenn and B. W. Poovaiah, unpublished observations). During senescence, dissolution of the middle lamella of control fruits occurs, resulting in cell separation. Calcium-treated fruits, which remain much firmer during storage, have a densely stained middle lamella and greater cell cohesion. At higher magnifications, control fruits show disintegration of fibrillar material throughout the cell wall, whereas the cell wall structure is preserved in calcium-treated fruits. These results suggest that calcium has a major effect on cell wall changes during ripening and senescence.
B. Calcium and Calmodulin
In plants, the level of free calcium in the cytosol is relatively low, in the range of 0.01 to 1.0 μΜ. Vacuoles, mitochondria, chloroplasts, and endo-
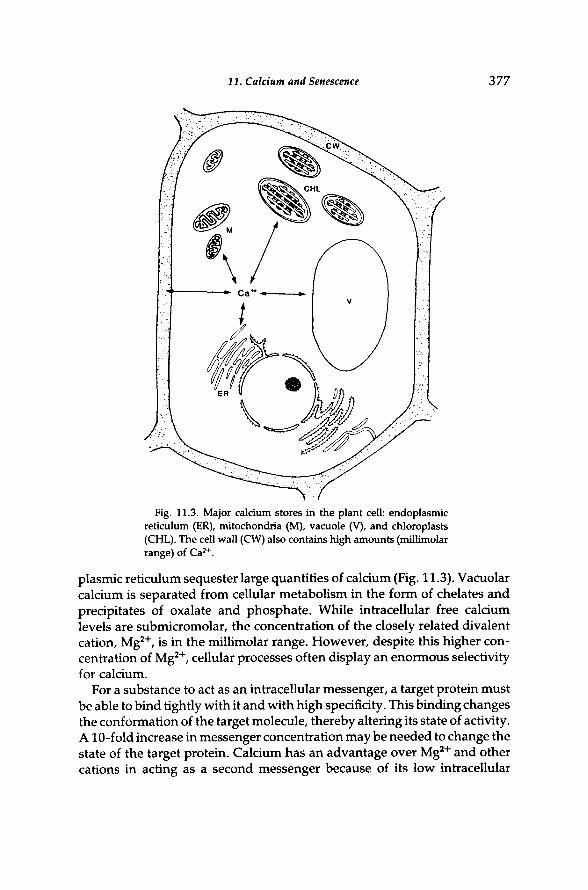
11. Calcium and Senescence 377
Fig. 11.3. Major calcium stores in the plant cell: endoplasmic reticulum (ER), mitochondria (M), vacuole (V), and chloroplasts (CHL). The cell wall (CW) also contains high amounts (millimolar range) of C a 2 + .
plasmic reticulum sequester large quantities of calcium (Fig. 11.3). Vacuolar calcium is separated from cellular metabolism in the form of chelates and precipitates of oxalate and phosphate. While intracellular free calcium levels are submicromolar, the concentration of the closely related divalent cation, M g 2 + , is in the millimolar range. However, despite this higher concentration of M g 2 + , cellular processes often display an enormous selectivity for calcium.
For a substance to act as an intracellular messenger, a target protein must be able to bind tightly with it and with high specificity. This binding changes the conformation of the target molecule, thereby altering its state of activity. A 10-fold increase in messenger concentration may be needed to change the state of the target protein. Calcium has an advantage over M g 2 + and other cations in acting as a second messenger because of its low intracellular

378 Β. W. Poovaiah
concentration. The low cytosolic calcium concentration is maintained by active C a 2 + pumping out of the cytoplasm by Ca-transporting ATPases on the plasma membrane. Recent investigations provide convincing evidence that Ca-transporting ATPase activities of microsomal membranes are controlled by the regulator protein, calmodulin, in a Ca-dependent manner (Marme and Dieter, 1983).
Calmodulin is a calcium-modulated protein that has multiple biochemical activities, including activation of enzymes involved in the regulation of a variety of cellular processes. While calmodulin is one member of the family of calcium-modulated proteins, its ubiquitous distribution in all higher plants as well as its highly conserved structure and function suggest that it may be serving a fundamental role in cellular homeostasis. This regulator protein is present in the cytoplasm and associated with membranes and organelles.
Stimuli such as light, gravity, and hormones have been shown to alter the C a 2 + concentration in the cytoplasm, thus allowing calcium to act as an intracellular signal (Fig. 11.4). This change in cytoplasmic C a 2 + concentration alters calmodulin activity. Each calmodulin molecule can bind up to four C a 2 + ions. The binding of C a 2 + activates calmodulin by altering its conformation and thereby exposing hydrophobic regions of the molecule. The Ca 2 + -calmodulin complex can then bind to target enzymes and activate them. The general mechanism by which calmodulin regulates these enzymes has been reviewed elsewhere (Cheung, 1980; Marme and Dieter, 1983; Poovaiah, 1985; Poovaiah and Reddy, 1987).
Calmodulin activity has been reported to be modulated in animal systems by a variety of compounds. The presence of a natural inhibitor of calmodulin action was detected in recent investigations on the role of calmodulin in the biochemical action of calcium in delaying senescence (Paliyath and Poovaiah, 1984, 1985a,b). Such an inhibitor could have a wide-ranging physiological role in developmental processes in plants. Since calmodulin regulates a multitude of processes, and many different classes of chemicals inhibit its activity, the question arises as to whether it could be possible to alter selectively calmodulin-dependent events in plants. It is conceivable that in the foreseeable future scientists might be able to regulate calcium-and calmodulin-mediated changes in order to induce or to delay senescence.
Inhibitors might block calmodulin action by a variety of mechanisms (Fig. 11.5): (1) by reducing the availability of C a 2 + [this is possible by inhibiting the entry of C a 2 + into cells, by inhibiting its release from intracellular stores (see Fig. 11.3), or by chelating it]; (2) by binding to calmodulin and altering its ability to bind C a 2 + ; (3) by binding to the Ca 2 + -calmodulin complex and altering its activity; (4) by binding to the calmodulin-recognition site on the calmodulin-sensitive enzyme, thereby preventing interaction of the C a 2 + -calmodulin complex with the enzyme; (5) by interacting with the catalytic

11. Calcium and Senescence 379
Stimulus
plasma membrane receptor
Calmodulin —
(nonfunctional)
Calmodulin
(functional)
Activation of enzymes
such as Protein Kinase,
NAD Kinase and ATPase
Response
Fig. 11.4. Calcium- and calmodulin-dependent regulation in higher plant cells. In the unstimulated cell, free calcium concentration in the cytoplasm remains low, in the submicromolar range. Thus, C a 2 + cannot bind to calmodulin. Following stimulation, the C a 2 + concentration in the cytoplasm increases and calcium binds to calmodulin, making it functional. The calcium-calmodulin complex binds to the enzyme, forming the functional calmodulin - C a - enzyme complex and induction of the response.
portion of the calmodulin-sensitive enzyme; and (6) by interacting with the ternary Ca2 +-calmodulin enzyme complex (Prozialeck and Weiss, 1985). The calmodulin inhibitors such as chlorpromazine, trifluoperazine, and N-(6-aminohexyl)5-chloro-l-naphthalene-sulfonamide hydrochloride (W-7) are known to block calmodulin action and have an effect on senescence (J. K. Rhee and B. W. Poovaiah, unpublished observations). However, since many calmodulin inhibitors have nonspecific effects, caution must be exercised in interpreting results when such chemicals are used.
C. Protein Phosphorylation
The molecular mechanisms by which cells communicate with each other are of central importance to the physiology of multicellular organisms. It is

380 Β. W. Poovaiah
RESPONSE Fig. 11.5. Diagram illustrating the mechanisms by which calmodulin action is inhibited. See
text for description.
now becoming clear that post-translational modification of enzymes may represent a key regulatory step in the molecular mechanism by which second messengers such as calcium respond to various external stimuli (Green-gard, 1978; Cohen, 1982). Covalent modification of enzymes between active and inactive forms is a key process involved in metabolic regulation. Covalent modifications caused by reversible phosphorylation of enzymes catalyzed by protein kinases and phosphatases are well studied (Schulman et al, 1980; Trewavas, 1976). The physiological roles of protein kinases are well known in animal systems (Greengard, 1978). Calcium- and calmodu-lin-promoted protein phosphorylation has been reported in many plant systems (Hetherington and Trewavas, 1982; Polya and Davies, 1982; Sali-math and Marme, 1983; Veluthambi and Poovaiah, 1984a,b; Poovaiah and Veluthambi, 1986). The promotion of C a 2 + and calmodulin-dependent protein kinases by C a 2 + and calmodulin provides a mechanism by which calcium can regulate diverse processes including senescence. The occurrence of C a 2 + and calmodulin-dependent protein kinases in membranes and soluble proteins from a wide variety of plant tissues (Fig. 11.6) suggests that such a phosphorylation system could be of general importance as a biochemical mechanism of calcium action. These observations offer a promising ap-
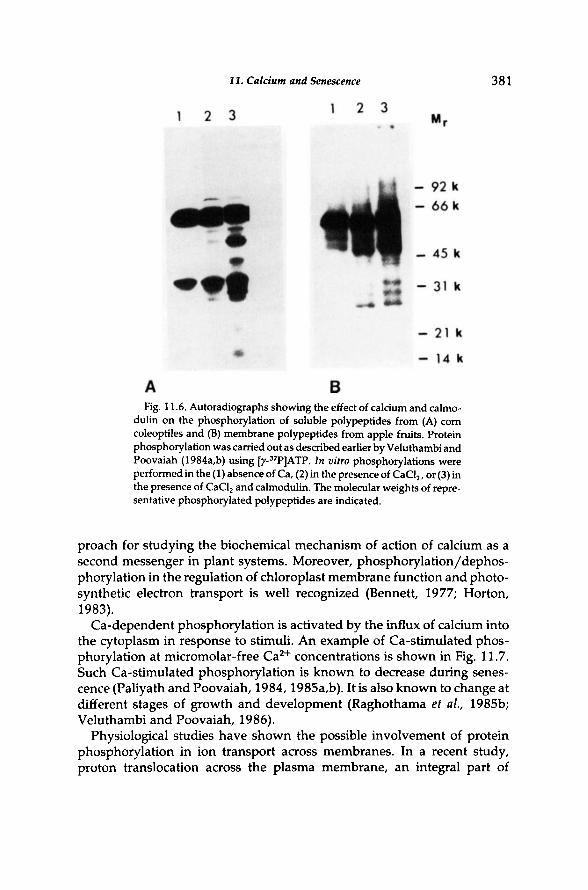
11. Calcium and Senescence 381
Fig. 11.6. Autoradiographs showing the effect of calcium and calmodulin on the phosphorylation of soluble polypeptides from (A) corn coleoptiles and (B) membrane polypeptides from apple fruits. Protein phosphorylation was carried out as described earlier by Veluthambi and Poovaiah (1984a,b) using [y-3 2P]ATP. In vitro phosphorylations were performed in the (1) absence of Ca, (2) in the presence of CaCl 2 , or (3) in the presence of CaCl 2 and calmodulin. The molecular weights of representative phosphorylated polypeptides are indicated.
proach for studying the biochemical mechanism of action of calcium as a second messenger in plant systems. Moreover, phosphorylation/dephos-phorylation in the regulation of chloroplast membrane function and photosynthetic electron transport is well recognized (Bennett, 1977; Horton, 1983).
Ca-dependent phosphorylation is activated by the influx of calcium into the cytoplasm in response to stimuli. An example of Ca-stimulated phosphorylation at micromolar-free C a 2 + concentrations is shown in Fig. 11.7. Such Ca-stimulated phosphorylation is known to decrease during senescence (Paliyath and Poovaiah, 1984,1985a,b). It is also known to change at different stages of growth and development (Raghothama et al., 1985b; Veluthambi and Poovaiah, 1986).
Physiological studies have shown the possible involvement of protein phosphorylation in ion transport across membranes. In a recent study, proton translocation across the plasma membrane, an integral part of
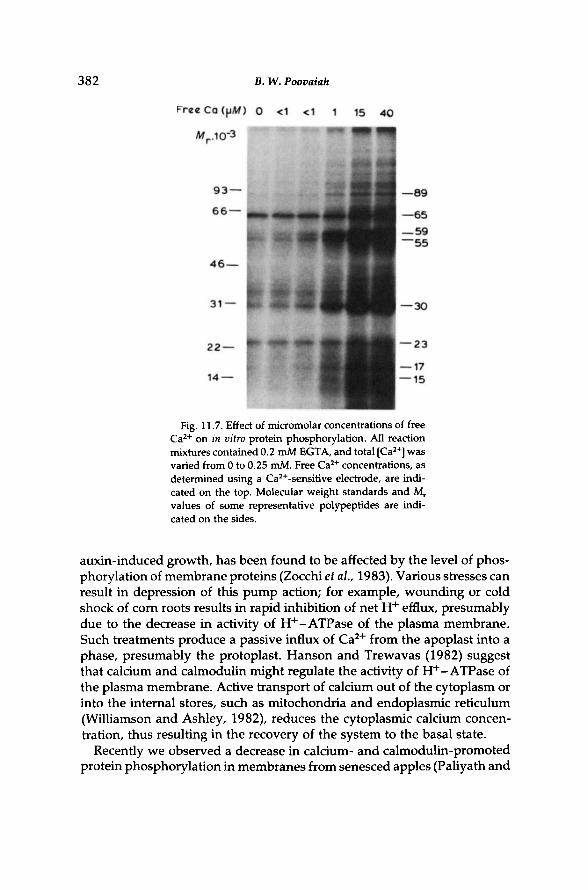
382 Β. W. Poovaiah
Fig. 11.7. Effect of micromolar concentrations of free C a 2 + on in vitro protein phosphorylation. All reaction mixtures contained 0.2 mM EGTA, and total [Ca 2 + ] was varied from 0 to 0.25 mM. Free C a 2 + concentrations, as determined using a Ca2 +-sensitive electrode, are indicated on the top. Molecular weight standards and M r
values of some representative polypeptides are indicated on the sides.
auxin-induced growth, has been found to be affected by the level of phosphorylation of membrane proteins (Zocchi et al, 1983). Various stresses can result in depression of this pump action; for example, wounding or cold shock of corn roots results in rapid inhibition of net H + efflux, presumably due to the decrease in activity of H + - A T P a s e of the plasma membrane. Such treatments produce a passive influx of C a 2 + from the apoplast into a phase, presumably the protoplast. Hanson and Trewavas (1982) suggest that calcium and calmodulin might regulate the activity of H + - A T P a s e of the plasma membrane. Active transport of calcium out of the cytoplasm or into the internal stores, such as mitochondria and endoplasmic reticulum (Williamson and Ashley, 1982), reduces the cytoplasmic calcium concentration, thus resulting in the recovery of the system to the basal state.
Recently we observed a decrease in calcium- and calmodulin-promoted protein phosphorylation in membranes from senesced apples (Paliyath and

11. Calcium and Senescence 383
Poovaiah, 1985b). Although equal amounts of membrane proteins were used from both normal and senesced apples, the phosphorylation of proteins promoted by C a 2 + and calmodulin was considerably reduced in membrane proteins from senesced apples. Similar results were observed when soluble proteins from ripening tomato fruits were used for phosphorylation (Raghothama et al, 1985b). Phosphoprotein phosphatase activity in membrane preparations from normal and senesced apples did not differ much, which suggests that a relatively high phosphoprotein phosphatase activity in senesced apple membranes could not be the major factor resulting in the decrease in calcium- and calmodulin-promoted protein phosphorylation. Thus, it appears that the decrease in membrane protein phosphorylation during senescence could be due to: (1) selective degradation of protein kinases and substrates or (2) inhibition of enzyme activities by an unfavorable microenvironment. The inhibition of calcium- and calmodulin-promoted protein phosphorylation could alter the biochemical processes in the cell and could participate in the onset and progress of senescence.
IV. ROLE OF INOSITOL PHOSPHOLIPIDS IN CALCIUM MESSENGER SYSTEM
The behavior of cells from one instant to another is controlled by signaling systems that translate external information into intracellular events. Recently, much attention has been given to the turnover of inositol phospholipids in the membrane (Poovaiah et al, 1987b). At present, there is no direct evidence that the turnover of inositol phospholipids is involved in senescence. However, inositol trisphosphate can release calcium from the intracellular stores and alter the free calcium status of the cell. This change in free calcium concentration could then play a major role in cellular regulation. A schematic diagram suggesting the turnover of inositol phospholipids and calcium mobilization leading to cellular response is shown in Fig. 11.8. This diagrammatic model presents an exciting area for further research. As shown in the figure, the primary stimulus, such as a hormone, interacts with the plasma membrane receptor. This results in the activation of phospho lipase C and the cleavage of the membrane lipid phosphatidylinositol 4,5-bisphosphate (PIP2) into diacylglycerol (DG) and inositol trisphosphate (IP3). Inositol trisphosphate stimulates the release of calcium from the endoplasmic reticulum. This released calcium can activate calmodulin-dependent enzymes, including protein kinases. Together with diacylglycerol, it can also activate protein kinase C, ultimately leading to protein phosphorylation and cellular response (Nishizuka, 1984; Berridge and Irvine, 1984). Recent evidence suggests the existence of similar pathways in trans-

384 Β. W. Poovaiah
P l a t m a . : ·
m e m b r a n e ,
A T P A D P A T P A D P
Enzyme Protein kinase Protein kinase C
I 1 Protein phosphorylation
Cellular r e s p o n s e
Fig. 11.8. Schematic illustration of stimulus-induced turnover of phosphatidylinositol 4,5-bisphosphate (PIP2) and the role of turnover products in the C a 2 + messenger system. This pathway is shown to occur in animal cells and evidence for its existence in plants is being accumulated. Interaction of extracellular signals with a receptor on the plasma membrane activates phospholipase C leading to hydrolysis of PIP 2 . Diacylglycerol (DG) and inositol 1,4,5-trisphosphate (IP3), the hydrolysis products of PIP 2 , play a key role in signal transduction. DG activates protein kinase C. IP 3 releases C a 2 + from endoplasmic reticulum, thereby raising cytosolic free C a 2 + and activating C a 2 + - and Ca2 +-calmodulin dependent enzymes. PI, phosphatidylinositol; PIP, phosphatidylinositol 4-phosphate; A, PI kinase; B, PIP kinase; ER, endoplasmic reticulum; IP 4 , inositol tetrakisphosphate; IP 3 inositol trisphosphate; IP 2 , inositol bisphosphate; IPX inositol monophosphate; MG, monoglyceride; AA, arachidonic acid; PA, phosphatidic acid. [From Poovaiah and Reddy (1987).]
ducing extracellular signals in plants (Reddy et al, 1987). The presence of polyphosphoinositides has been shown in plant tissue culture cells (Boss and Massel, 1985). Inositol trisphosphate-induced calcium mobilization from microsomal fractions has been observed in our laboratory (Reddy and Poovaiah, 1987) and elsewhere (Drobak and Ferguson, 1985). We also have preliminary evidence indicating that most of the calcium released by inositol trisphosphate is from the endoplasmic reticulum. Moreover, protein kinase C-like activity has been reported in plants (Schafer et al, 1985). Further investigations on this pathway are vital for a better understanding of signal transduction in plants. Additional information is needed to clarify the sig-

11. Calcium and Senescence 385
nificance of this pathway in plant growth and development in general and senescence in particular.
V. CONCLUSION
Calcium effects on various parameters of senescence, such as loss of chlorophyll and protein, increased membrane leakage, dissolution of middle lamella, and associated changes in the cell wall, are well established. However, the mechanism of calcium action is just beginning to be unraveled at the molecular level. Investigations on calcium- and calmodulin-mediated biochemical processes as well as the turnover of inositol phospholipids are in their infancy in plants. Additional information is necessary regarding the intracellular fluctuations in calcium concentration. Accurate and direct measurements of free cytoplasmic calcium concentrations are required to further clarify the involvement of calcium in senescence. There is potential for major breakthroughs. The prospects are promising.
ACKNOWLEDGMENTS
The author is indebted to all the former and current members of this laboratory who have made this chapter possible. This work was supported in part by the National Science Foundation grant DCB-8502215.
REFERENCES
Anderson, J. M., and Cormier, M. J. (1978). Calcium-dependent regulation of NAD kinase in higher plants. Biochem. Biophys. Res. Commun. 84, 5 9 5 - 6 0 2 .
Balamani, V., Veluthambi, K., and Poovaiah, B. W. (1986). Effect of calcium on tuberization in potato (Solanum tuberosum L.). Plant Physiol. 80, 8 5 6 - 8 5 8 .
Bartley, I. M. (1974). /?-Galactosidase activity in ripening apples. Phytochemistry 13, 2 1 0 7 -2111.
Bennett, J. (1977). Phosphorylation of chloroplast membrane polypeptides. Nature (London) 269, 3 4 4 - 3 4 6 .
Berridge, M. J., and Irvine, R. F. (1984). Inositol trisphosphate, a novel second messenger in cellular signal transduction. Nature (London) 312, 3 1 5 - 3 2 1 .
Boss, W. F„ and Massel, M. O. (1985). Polyphosphoinositides are present in plant tissue culture cells. Biochem. Biophys. Res. Commun. 132, 1 0 1 8 - 1 0 2 3 .
Buescher, R. W., and Hobson, G. E. (1982). Role of calcium and chelating agents in regulating the degradation of tomato fruit tissue by polygalacturonase. /. Plant Biochem. 6, 1 4 7 - 1 6 0 .
Cheung, W. Y. (1980). Calmodulin plays a pivotal role in cellular regulation. Science 207, 1 9 - 2 7 .
Cohen, P. (1982). The role of protein phosphorylation in neural and hormonal control of cellular activity. Nature (London) 296, 6 1 3 - 6 1 9 .

386 Β. W. Poovaiah
Conway, W. S., and Sams, C. E. (1984). Possible mechanisms by which postharvest calcium treatment reduces decay in apples. Phytopathology 74, 2 0 8 - 2 1 0 .
Demarty, M., Morvan, C , and Thellier, M. (1984). Calcium and the cell wall. Plant Cell Environ. 7, 4 4 1 - 4 4 8 .
Doesburg, J. J. (1957). Relation between the solubilization of pectin and the fate of organic acids during maturation of apples. /. Sci. Food Agric. 8, 2 0 6 - 2 1 6 .
Drobak, Β. K., and Ferguson, I. B. (1985). Release of C a 2 + from plant hypocotyl microsomes by inositol-4,5-trisphosphate. Biochem. Biophys. Res. Commun. 130, 1 2 4 1 - 1 2 4 6 .
Eilam, Y. (1965). Permeability changes in senescing tissue. /. Exp. Bot. 16, 6 1 4 - 6 2 7 . Epstein, E. (1961). The essential role of calcium in selective cation transport by plant cells. Plant
Physiol. 3 6 , 4 3 7 - 4 4 4 . Gary-Bobo, C. M. (1970). Effect of C a ^ on the water and non-electrolyte permeability of
phospholipid membranes. Nature (London) 228, 1 1 0 1 - 1 1 0 2 . Greengard, P. (1978). Phosphorylated proteins as physiological effectors. Science 199, 1 4 6 -
152. Hanson, J. B. (1983). The roles of calcium in plant growth. In "Current Topics in Plant
Biochemistry and Physiology" (D. D. Randall, D. G. Blevins, and R. Larson, eds.), Vol. 1, pp. 1 - 2 4 . Univ. of Missouri Press, Columbia.
Hanson, J. B., and Trewavas, A. J. (1982). Regulation of plant cell growth: The changing perspective. New Phytol. 90, 1 - 1 8 .
Hepler, P. K., and Wayne, R. O. (1985). Calcium and plant development. Annu. Rev. Plant Physiol. 36, 3 9 7 - 4 3 9 .
Hetherington, A. M., and Trewavas, A. (1982). Calcium-dependent protein kinase in pea shoot membranes. FEBS Lett. 145, 6 7 - 7 1 .
Hetherington, A. M., DeSilva, D. L. R., Cox, R. C , and Mansfield, T. A. (1986). Abscisic acid, calcium ions and stomatal function. In "Molecular and Cellular Aspects of Calcium in Plant Development" (A. J. Trewavas, ed.), pp. 3 8 7 - 3 8 8 . Plenum, New York.
Horton, P. (1983). Control of chloroplast electron transport by phosphorylation of thylakoid proteins. FEBS Lett. 152, 4 7 - 5 1 .
Jones, R. G. W., and Lunt, O. R. (1967). The function of calcium in plants. Bot. Rev. 33, 4 0 7 - 4 2 6 .
Keith, C. H., Ratan, R., Maxfield, F. R., Bajer, Α., and Shelanski, M. L. (1985). Local cytoplasmic calcium gradients in living mitotic cells. Nature (London) 316, 8 4 8 - 8 5 0 .
Knee, M. (1973). Polysaccharide changes in cell walls of ripening apples. Phytochemistry 12, 1 5 4 3 - 1 5 4 9 .
Kretsinger, R. H. (1977). Evolution of the informational role of calcium in eukaryotes. In "Calcium-Binding Proteins and Calcium Function" (R. H. Wasserman, R. A. Corradino, E. Carafoli, R. H. Kretsinger, D. H. MacLennan, and F. L. Siegel, eds.), pp. 6 3 - 7 2 . North-Holland Publ., New York.
Leopold, A. C , Poovaiah, B. W., Dela Fuente, R. K., and Williams, R. J. (1974). Regulation of growth with inorganic solutes. In "Plant Growth Substances," pp. 7 8 0 - 7 8 8 . Hirokawa Press, Tokyo.
Mansfield, Τ. Α., and Davies, W. J. (1981). Stomata and stomatal mechanisms. In "The Physiology and Biochemistry of Drought Resistance in Plants" (L. G. Paleg and D. Aspinall, eds.), pp. 3 1 5 - 3 4 6 . Academic Press, New York.
Marinos, N. G. (1962). Studies on submicroscopic aspects of mineral deficiencies. I. Calcium deficiency in the shoot apex of barley. Am. J. Bot. 49, 8 3 4 - 8 4 1 .
Marme, D., and Dieter, P. (1983). Role of Ca++ and calmodulin in plants. In "Calcium and Cell Function" (W. Y. Cheung, ed.), Vol. 4, pp. 2 6 3 - 3 1 1 . Academic Press, New York.

11. Calcium and Senescence 387
Mizrahi, Y., and Kopeliovitch, E. (1983). Pectic substances: changes in soft and firm tomato cultivars and in non-ripening mutants. /. Am. Soc. Hortic. Sci. 8 5 , 1 1 1 - 1 1 6 .
Nishizuka, Y. (1984). The role of protein kinase C in cell surface signal transduction and tumour promotion. Nature (London) 308, 6 9 3 - 6 9 8 .
Paliyath, G., and Poovaiah, B. W. (1984). Calmodulin inhibitor in senescing apples and its physiological and pharmacological significance. Proc. Natl. Acad. Sci. U.S.A. 8 1 , 2 0 6 5 - 2 0 6 9 .
Paliyath, G., and Poovaiah, B. W. (1985a). Identification of naturally occurring calmodulin inhibitors in plants and their effects on calcium- and calmodulin-promoted protein phosphorylation. Plant Cell Physiol. 26, 2 0 1 - 2 0 9 .
Paliyath, G., and Poovaiah, B. W. (1985b). Calcium- and calmodulin-promoted phosphorylation of membrane proteins during senescence in apples. Plant and Cell Physiol. 2 6 , 9 7 7 - 9 8 6 .
Paliyath, G., and Poovaiah, B. W. (1988). Promotion of /?-glucan synthase activity in corn microsomal membranes by calcium and protein phosphorylation. Plant Cell Physiol. 29, 6 7 - 7 3 .
Paliyath, G., Poovaiah, B. W., Munske, G. R., and Magnuson, J. (1984). Membrane fluidity in senescing apples: Effects of temperature and calcium. Plant Cell Physiol. 25, 1 0 8 3 - 1 0 8 7 .
Polya, G. M., and Davies, J. R. (1982). Resolution of Ca^-calmodulin activated protein kinase from wheat germ. FEBS Lett. 150, 1 6 7 - 1 7 1 .
Poovaiah, B. W. (1979a). Effects of inorganic cations on ethephon-induced increases in membrane permeability. /. Am. Soc. Hortic. Sci. 104, 1 6 4 - 1 6 6 .
Poovaiah, B. W. (1979b). Role of calcium in ripening and senescence. Commun. Soil Sci. Plant Anal. 1 0 , 8 3 - 8 8 .
Poovaiah, B. W. (1985). Role of calcium and calmodulin in plant growth and development. HortScience 20, 3 4 7 - 3 5 2 .
Poovaiah, B. W. (1986). Role of calcium in prolonging storage life of fruits and vegetables. Food Technol. 40, 8 6 - 8 9 .
Poovaiah, B. W. (1987). The role of calcium and calmodulin in senescence. In "Plant Senescence: Its Biochemistry and Physiology" (W. W. Thomson, E. A. Nothnagel, and R. C. Huffaker, eds.), pp. 1 8 2 - 1 8 9 . Am. Soc. Plant Physiol., Rockville, Maryland.
Poovaiah, B. W., and Leopold, A. C. (1973a). Deferral of leaf senescence with calcium. Plant Physiol. 52, 2 3 6 - 2 3 9 .
Poovaiah, B. W., and Leopold, A. C. (1973b). Inhibition of abscission by calcium. Plant Physiol. 51, 8 4 8 - 8 5 1 .
Poovaiah, B. W., and Leopold, A. C. (1974). Hormone-solute interactions in the lettuce hypo-cotyl hook. Plant Physiol. 54, 2 8 9 - 2 9 3 .
Poovaiah, B. W., and Leopold, A. C. (1976a). Effects of inorganic solutes on the binding of auxin. Plant Physiol. 58, 7 8 3 - 7 8 5 .
Poovaiah, B. W., and Leopold, A. C. (1976b). Effects of inorganic salts on tissue permeability. Plant Physiol. 58, 1 8 2 - 1 8 5 .
Poovaiah, B. W., and Moulton, G. A. (1982). Vacuum pressure infiltration process for fresh produce. U.S. Patent 4,331,691.
Poovaiah, B. W., and Rasmussen, H. P. (1973a). Effect of calcium, 2-chloroethylphosphonic acid, and ethylene on bean leaf abscission. Planta 113, 2 0 7 - 2 1 4 .
Poovaiah, B. W., and Rasmussen, H. P. (1973b). Calcium distribution in the abscission zone of bean leaves: electron microprobe X-ray analysis. Plant Physiol. 52, 6 8 3 - 6 8 4 .
Poovaiah, B. W., and Reddy, A. S. N. (1987). Calcium messenger system in plants. CRC Cut. Rev. Plant Sci. 6, 4 7 - 1 0 3 .
Poovaiah, B. W., and Veluthambi, K. (1986). The role of calcium and calmodulin in hormone action in plants: Importance of protein phosphorylation. In "Molecular and Cellular Aspects

388 Β. W. Poovaiah
of Calcium in Plant Development" (A. J. Trewavas, ed.), pp. 8 3 - 9 0 . Plenum, New York. Poovaiah, B. W., McFadden, J. J., and Reddy, A. S. N. (1987a). The role of calcium in gravity
signal perception and transduction. Physiol Plant 71, 4 0 1 - 4 0 7 . Poovaiah, B. W., Reddy, A. S. N., and McFadden, J. J. (1987b). Calcium messenger system: Role
of protein phosphorylation and inositol phospholipids. Physiol Plant 69, 5 6 9 - 5 7 3 . Poovaiah, B. W., Glenn, G. M., and Reddy, A. S. N. (1988). Calcium and fruit softening:
Physiology and biochemistry. Hortic. Rev. (in press). Prozialeck, W. C , and Weiss, B. (1985). Mechanisms of pharmacologically altering calmodulin
activity. In "Calcium in Biological Systems" (R. P. Rubin, G. B. Weiss, and J. W. Putney, Jr., eds.), pp. 2 5 5 - 2 6 4 . Plenum, New York.
Raghothama, K. G., Mizrahi, Y., and Poovaiah, B. W. (1985a). Effect of calmodulin antagonists on auxin-induced elongation. Plant Physiol. 79, 2 8 - 3 3 .
Raghothama, K. G., Veluthambi, K., and Poovaiah, B. W. (1985b). Stage-specific changes in calcium-regulated protein phosphorylation in developing fruits. Plant Cell Physiol. 26, 1 5 6 5 - 1 5 7 2 .
Raghothama, K. G., Reddy, A. S. N., Friedmann, M., and Poovaiah, B. W. (1988). Effect of calcium on in vivo protein phosphorylation in corn root tips. Plant Physiol (in press).
Ray, P. M., and Baker, D. B. (1965). The effect of auxin on synthesis of oat coleoptile cell wall constituents. Plant Physiol. 41, 3 5 3 - 3 6 0 .
Reddy, A. S. N., and Poovaiah, B. W. (1987). Inositol 1,4,5-triphosphate induced calcium release from corn coleoptile microsomes. /. Biochem. (Tokyo) 101, 5 6 9 - 5 7 3 .
Reddy, A. S. N., McFadden, J. J., Friedmann, M., and Poovaiah, B. W. (1987). Signal transduction in plants: Evidence for the involvement of calcium and turnover of inositol phospholipids. Biochem. Biophys. Res. Commun. 149, 3 3 4 - 3 3 9 .
Richmond, A. E., and Lang, A. (1957). Effect of kinetin on protein content and survival of detached Xanthium leaves. Science 125, 6 5 0 - 6 5 1 .
Rossignol, M., Lamant, D., Salsac, L., and Heller, R. (1977). Calcium fixation by the roots of calcicole and calcifuge plants: the importance of membrane systems and their lipid composition. In "Transmembrane Ionic Exchange in Plants" (M. Thellier, A. Monnier, M. Demarty, and J. Dainty, eds.), pp. 4 8 3 - 4 9 0 . CNRS, Paris and Editions Univ., Rouen.
Roux, S. J., and Slocum, R. D. (1982). Role of calcium in mediating cellular functions important for growth and development in higher plants. In "Calcium and Cell Function" (W. Y. Cheung, ed.), Vol. 3, pp. 4 0 9 - 4 5 3 . Academic Press, New York.
Sacher, J. A. (1957). Relationship between auxin and membrane-integrity in tissue senescence and abscission. Science 125, 1 1 9 9 - 1 2 0 0 .
Salimath, B. P., and Marme, D. (1983). Protein phosphorylation and its regulation by calcium and calmodulin in membrane fractions from zucchini hypocotyls. Planta 158, 5 6 0 - 5 6 8 .
Saunders, M. J., and Hepler, P. K. (1982). Calcium ionophore A23187 stimulates cytokinin-like mitosis in Funaria. Science 217, 9 4 3 - 9 4 5 .
Schafer, Α., Bygrave, F., Matzenauer, S., and Marme, D. (1985). Identification of a calcium-and phospholipid-dependent protein kinase in plant tissue. FEBS Lett. 187, 2 5 - 2 8 .
Schulman, H., Wieland, Β. H., and Greengard, P. (1980). Calcium-dependent protein phosphorylation in mammalian brain and other tissues. In "Calcium and Cell Function" (W. Y. Cheung, ed.), Vol. 1, pp. 2 2 0 - 2 4 8 . Academic Press, New York.
Suwwan, Μ. Α., and Poovaiah, B. W. (1978). Association between elemental content and fruit ripening in rin and normal tomatoes. Plant Physiol 61, 8 8 3 - 8 8 5 .
Thimann, Κ. V., and Satler, S. (1979). Interrelation between leaf senescence and stomatal aperture. Proc. Natl. Acad. Sci. U.S.A. 76, 2 2 9 5 - 2 2 9 8 , 2 7 7 0 - 2 7 7 3 .
Trewavas, A. (1970). The turnover of nucleic acids in Lemna minor. Plant Physiol 45, 7 4 2 - 7 5 1 . Trewavas, A. (1972). Control of protein turnover rates in Lemna minor. Plant Physiol 4 9 , 4 7 - 5 1 .

11. Calcium and Senescence 389
Trewavas, A. (1976). Post-translational modification of proteins by phosphorylation. Annu. Rev. Plant Physiol. 27, 3 4 9 - 3 7 4 .
Veluthambi, K., and Poovaiah, B. W. (1984a). Calcium-promoted protein phosphorylation in plants. Science 223, 1 6 7 - 1 6 9 .
Veluthambi, K., and Poovaiah, B. W. (1984b). Calcium- and calmodulin-regulated phosphorylation of soluble and membrane proteins from corn coleoptiles. Plant Physiol. 76 ,359 - 365.
Veluthambi, K., and Poovaiah, B. W. (1986). in vitro and in vivo protein phosphorylation in Avena sativa L. coleoptiles: Effects of Ca"4"*", calmodulin antagonists and auxin. Plant Physiol 81, 8 3 6 - 8 4 1 .
Wallner, S. J. (1978). Apple fruit/?-galactosidase and softening in storage. /. Am. Soc. Hortic. Sci. 1 0 3 , 3 6 4 - 3 6 6 .
Williamson, R. E., and Ashley, C. C. (1982). Free Ca++ and cytoplasmic streaming in algae Chara. Nature (London) 296, 6 4 7 - 6 5 1 .
Zocchi, G., Rogers, S. Α., and Hanson, J. B. (1983). Inhibition of proton pumping in corn roots is associated with increased phosphorylation of membrane proteins. Plant Sci. Lett. 81, 2 1 5 -221.