seedling and clonal recruitment of the invasive tree psidium cattleianum: implications for...
TRANSCRIPT

Biological Conservation 53 (1990) 199-211 • , ~ L , 1 ~,,.
Seedling and Clonal Recruitment of the Invasive Tree PMdium cattleianum: Implications for Management of
Native Hawaiian Forests
Laura Foster Huenneke
Department of Biology, New Mexico State University, Box 30001, Las Cruces, New Mexico 88003, USA
&
Peter M. Vitousek
Biological Sciences, Stanford University, Stanford, California 94305, USA
(Received 15 June 1989; revised version received 30 November 1989; accepted 7 December 1989)
A BS TRA C T
Non-native plants present serious management problems in many preserves. Strawberry guava Psidium cattleianum (Myrtaceae), a small tree and aggressive invader of tropical areas, is rapidly spreading through many Hawaiian forests including those of the two US national parks in Hawaii. Feral pigs and non-native birds disperse Psidium seeds; pigs also create soil disturbances that may enhance the tree's spread. Our study of guava's reproductive biology focussed on its dependence on non-native animals. We found that the abundantly produced seed germinated rapidly under a wide range of conditions, without scarification. Psidium seedlings occur on the same substrates as do native seedlings, usually on undisturbed sites. Both seedlings and clonally produced suckers are common, but suckers contribute greater leaf areas. Guava's clonal growth may partially explain its success in dominating native forests. Apparently germination and establishment do not depend on animal dispersal, or on disturbances created by pigs; thus, control of the plant cannot rest entirely on control of non-native animals.
199 Biol. Conserv. 0006-3207/90/$03"50 © 1990 Elsevier Science Publishers Ltd, England. Printed in Great Britain

200 Laura Foster Huenneke, Peter M. Vitousek
INTRODUCTION
Invasive plants can pose a significant challenge to the management of preserves or conservation areas. Invading plants (those non-native species able to establish themselves and spread in relatively undisturbed natural ecosystems) flourish at the expense of natives; they may even alter the structure and function of the invaded ecosystem (Vitousek, 1986). The resulting changes may be of conservation concern, when the management goal is to maintain natural ecosystem processes or features, or when native plant or animal species are adversely affected. Over the past few years a number of reviews have addressed the nature, mechanisms, and conse- quences of plant and animal invasions (e.g. Mooney & Drake, 1986; Kornberg & Williamson, 1986; Joenje et al., 1987). There has been growing realization that invasive plants can represent substantial threats to natural ecosystems, including those in parks or other preserve areas.
Like many other oceanic islands, the Hawaiian Islands have proven vulnerable to invasive non-native plants. Some species were introduced by the Polynesians, and thousands more in the past 200 years. Of these, 86 are considered to have become serious problems in natural ecosystems (Smith, 1985). Psidium cattleianum Sabine (Myrtaceae), the strawberry guava, is a small tree believed to have been brought to Hawaii from Brazil (Wagner et al., in press). The species has invaded Hawaiian forests on several islands, presenting management problems in national parks and other natural areas. On Maui strawberry guava has come to dominate extensive areas of previously native forest (L. Cuddihy, pers. comm.) in Haleakala National Park; on the Big Island of Hawaii, the species has been found in a variety of native ecosystems, from quite dry to very wet forests, from 100 m to 1000 m in elevation (Jacobi & Warshauer, in press). While strawberry guava forms dense thickets along roadsides and in other disturbed areas, it is also successful in seemingly undisturbed intact forest.
Psidium cattleianum produces abundant, juicy fruits (red in forma cattleianum, yellow in forma lucidum) attractive to birds and mammals. Non-native birds and feral pigs Sus scr~?[il may represent important dispersal agents for the hard-coated seeds within the fruits. Pigs may serve another role, as they create extensive areas of soil disturbance by their rooting activities within native forest; seedling establishment for weedy plant species might be favored in such disturbed sites. Potential feedback loops between guava and non-native animals, then, may present critical targets for any control of guava's further spread and domination of native forests.
The primary objective of our research on the population biology of P. cattleianum was to supply information on its reproductive behavior, and

Strawberry guava encroachment in Hawaiian forests 201
particularly its relationship with animals such as feral pigs, that might guide management and control programs. Are reproductive behavior or growth related to elevation, suggesting an upper elevational limit to the species' potential range? Is guava dependent on scarification or dispersal of its seeds by animals for successful germination? Is seedling establishment dependent on or responsive to soil disturbance? Finally, what is the relative importance of seed reproduction versus clonal behavior in guava's ability to form dense thickets and exclude native plants?
METHODS
Phenology
Our studies of P. cattleianum in and near Hawaii Volcanoes National Park began in June 1987. Stem densities, size structure and phenology were observed in thickets along an elevational gradient outside park boundaries. The five stations range from 150m to 760m in elevation, on state forest reserve land west of Hilo (along the Stainback Highway), where strawberry guava dominates much of the roadsides. Scattered individuals of native woody plants (chiefly ohi'a, Metrosideros polymorpha, and tree ferns of the genus Cibotium) occurred in the guava thickets. Other non-native plants (shrubs, vines and grasses) were common. Numbers and diameters of strawberry guava and other woody stems were recorded for three 5 m x 5 m plots within the thicket interior at each station. Twenty-five interior and 25 thicket-edge stems were tagged at each station and monitored monthly for phenological stage (ripe fruit, unripe fruit, flowering, in bud, new leaf flush, green vegetative).
Germination
Seed germination physiology was investigated in laboratory trials. Fruits were collected in November 1987 from the canopy and forest floor of the 5 stands along the elevational gradient. The hard-coated seeds were removed from the fruit pulp and allowed to air-dry; numbers of seeds per fruit were counted for 15 fruits from each station. Seed mass (air-dry weight) was measured for 4 sets of 10 seeds each for each station.
After storage for a month at room temperature, seeds were germinated under various conditions (in light and in dark, 22°C; in dark, 18°C, 37°C and 22°C after 2 or 14 days pre-chilling treatment, after scarification with sandpaper, and with a drought period of one week imposed after 2 weeks of moisture, with and without prior scarification). Seeds were germinated on

202 Laura Foster Huenneke, Peter M. Vitousek
filter paper in covered Petri dishes, wetted with deionized water, in sets of 25 seeds per dish (4 replicate dishes per trial).
Germination experiments were also carried out in the field. In November 1987 cleaned seed was set out in groups of 10 on bryophyte mats, on cleared soil surface, and on the litter layer, at the 460 m station (10 groups on each substrate). Several intact ripe fruits were also placed on moss-covered substrates and tagged. In addition, 1-m 2 quadrats were cleared of all leaf litter (5 at the 460 m site, 5 at another site (Campbell) adjacent to the park boundary at 1200m) and the soil disturbed to simulate pig damage. All locations were revisited in late March 1988 and the number and condition of germinants noted. Seedling densities, and locations of seedlings on particular microsites such as bryophyte mats or decomposing wood, were noted in one population in the park (Thurston, l190m), at Campbell, and at the 460m Stainback station. In February 1988, one 25 cm x 25 cm × 10 cm deep block of soil, and another block of equal size from a dense mat of bryophytes, were collected at the 460 m Stainback site, where fruit production had been very heavy the previous November. These blocks were removed to the laboratory and all guava seeds (or fragments of seeds) separated from them. Seeds were examined under a dissecting microscope and separated into intact, fragmented and germinated-but-dead categories (germinated seeds had pores in the seed coat corresponding to those displayed by germinated seeds in the laboratory germination trials, and often retained a portion of dead seedling tissue as well).
Clonal behavior
The relative importance of seedling vs sucker or vegetative recruitment was estimated by sampling understory guava recruits (10-50cm height) at Thurston, Campbell, and three of the Stainback stations (150 m, 460 m and 762m). Recruits were pulled from the soil; true seedlings were readily distinguished by their branched root systems, while suckers were attached to horizontal roots or rhizomes by unbranched connections. Stems were cut at the level of the soil surface, height measured, and number and total surface area of leaves recorded. Regressions of leaf area against recruit height were used to compare leaf area production of seedlings and suckers.
Stem demography
Demographic data (recruitment, stem growth and mortality) were collected for 3 populations within the park--Thurston (1190 m, where P. cat t le ianum
is invading as scattered stems beneath a closed canopy of native trees and tree ferns); Upper Kalapana (830 m, a denser stand of guava in a wet ohi'a-
Strawberry guava encroachment in Hawaiian forests 203
tree fern forest); and Boundary (1260 m, in drier open ohi'a-koa forest east of the Kipuka Puaulu trail loop, adjacent to the eastern park boundary). Plot boundaries were flagged (40 m x 40 m at Thurston; 30 m × 30 m at the other sites), and all guava stems within the plots tagged and measured (diameter at breast height). Each population was revisited at least once by June 1988, and all stems relocated and their diameters remeasured. Survivorship and growth rates were estimated for size classes of stems in each site.
RESULTS
Phenology
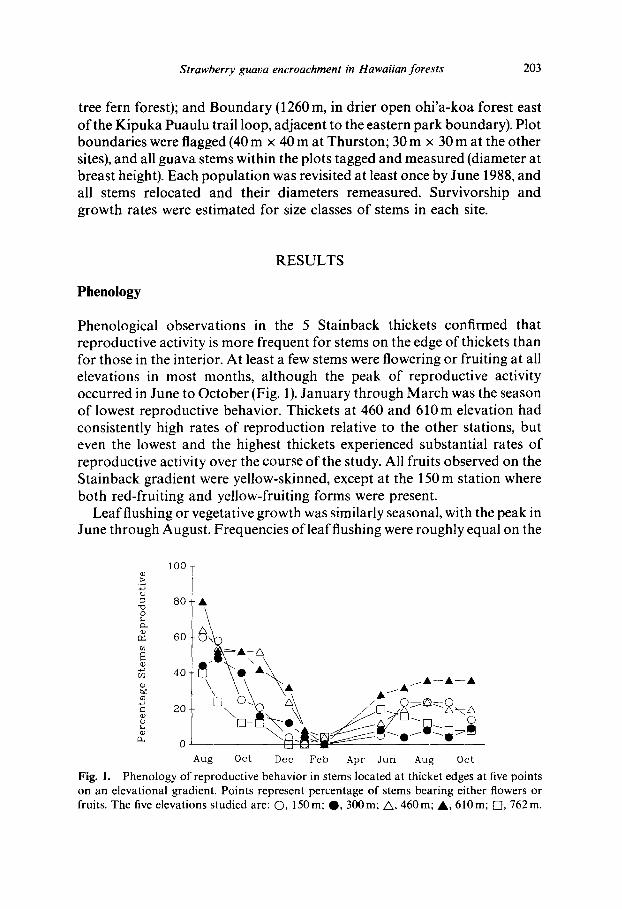
Phenological observations in the 5 Stainback thickets confirmed that reproductive activity is more frequent for stems on the edge of thickets than for those in the interior. At least a few stems were flowering or fruiting at all elevations in most months, although the peak of reproductive activity occurred in June to October (Fig. 1). January through March was the season of lowest reproductive behavior. Thickets at 460 and 610 m elevation had consistently high rates of reproduction relative to the other stations, but even the lowest and the highest thickets experienced substantial rates of reproductive activity over the course of the study. All fruits observed on the Stainback gradient were yellow-skinned, except at the 150 m station where both red-fruiting and yellow-fruiting forms were present.
Leaf flushing or vegetative growth was similarly seasonal, with the peak in June through August. Frequencies of leaf flushing were roughly equal on the
i00
o 80
o
0) c~ 60
m 40
~ 20
a~ 0 Aug Oct Dec Feb Apr Jun Aug Oct
Fig. 1. Phenology of reproductive behavior in stems located at thicket edges at five points on an elevational gradient. Points represent percentage of stems bearing either flowers or fruits. The five elevations studied are: O, 150m; Q, 300m; A , 460m; A , 610m; [], 762m.

204 Laura Foster Huenneke, Peter M. Vitousek
th icket edges and in thicket interiors, with 2 5 - 5 5 % o f stems flushing dur ing the peak season, and 0 - 1 2 % flushing dur ing the rest o f the year. The re were no consp icuous differences a m o n g elevat ional s ta t ions in f r equency o f leaf
flush. Stem densities were high at all elevations, ranging f ro m 3 to 9 stems m 2.
The re were significant differences be tween elevat ions in to ta l densi ty ( A N O V A , F4.14 = 8"00, p < 0"005), with highest densities at in te rmedia te e leva t ions (Table 1). T o t a l " s t e m area increased g radua l ly (but n o t significantly) with increasing e levat ion up to the highest s tat ion, which had significantly lower stem area (Table 1). A large percen tage o f stems at the highest e levat ion were recent recruits less than 1 m in height (Table 1); there were significant differences a m o n g s ta t ions in the relative a b u n d a n c e o f small stems, but on ly the 762 m s ta t ion differed f rom all the o thers ( A N O V A ,
F 4 , 1 4 = 6"80, p < 0"007).
Laboratory germination trials
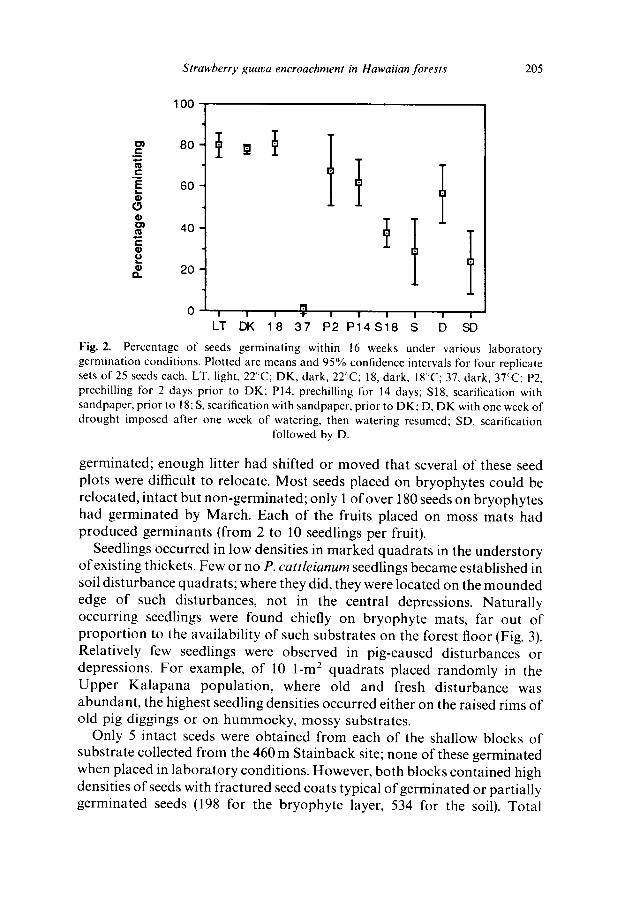
G e r m i n a t i o n rates were high (60 80%) under mos t condi t ions (Fig. 2), but h igh t e m p e r a t u r e s resu l ted in seed mor t a l i t y . Scar i f ied seeds had significantly lower ge rmina t ion than any o ther t rea tments . Chil l ing pr io r to ge rmina t ion had surpris ingly little effect on eventual germina t ion .
The re were significant differences be tween e leva t iona l s ta t ions in l a b o r a t o r y ge rmina t ion rates (Table 1; A N O V A , F5,~8 = 7"29, p < 0-001). G e r m i n a t i o n was not cor re la ted with elew~tion or seed weights. Seed n u m b e r s per fruit were highest for the in te rmedia te e levat ions (Table 1).
Field germination
G e r m i n a t i o n in the field was marked ly lower than tha t observed in the l abora to ry . N o n e o f the seeds placed on leaf li t ter or on cleared soil
TABLE 1 Stand, Seed, and Fruit Characteristics of Strawberry Guava Populations at Five Elevations
Elevation No. o f s tems % stems Sum steel area Seed mass Seed no. Germination (m) m 2 50- lO0 elgl (crn 2 m -2) (mg) per j?uit (%)
150 5"5 a 18"2 a 35.4 a 21"8 a 25"3 a 85-9 a 300 4.5 ab 16.2 a 44.6 a 15.0 b 59"6 bc 42"0 b 460 8"7 c 18"5 a 61"9 a 12"5 c 70"2 c 76"0 a 610 6"9 ac 14"5 a 66"6 a 15"9 bd 41"4 d 38"0 b 762 2'9 b 35'1 b 8"8 b 17"4 d 42"3 abd 40.5 b
Different letters within a column indicate significant differences between elevations.
Strawberry guava encroachment in Hawaiian forests 205
100
o . o f ._= III
' t t ° ~
60
o
• 2 0
0 I i I ~" I l I I I I
LT DK 18 37 P2 P14S18 S D SO
Fig. 2. Percentage of seeds germinating within 16 weeks under various laboratory germination conditions. Plotted are means and 95% confidence intervals for four replicate sets of 25 seeds each. LT, light, 22°C; DK, dark, 22°C; 18, dark, 18°C; 37, dark, 37°C; P2, prechilling for 2 days prior to DK; P14, prechilling for 14 days; S18, scarification with sandpaper, prior to 18; S, scarification with sandpaper, prior to DK; D, DK with one week of drought imposed after one week of watering, then watering resumed; SD, scarification
followed by D.
germinated; enough litter had shifted or moved that several of these seed plots were difficult to relocate. Most seeds placed on bryophytes could be relocated, intact but non-germinated; only 1 of over 180 seeds on bryophytes had germinated by March. Each of the fruits placed on moss mats had produced germinants (from 2 to 10 seedlings per fruit).
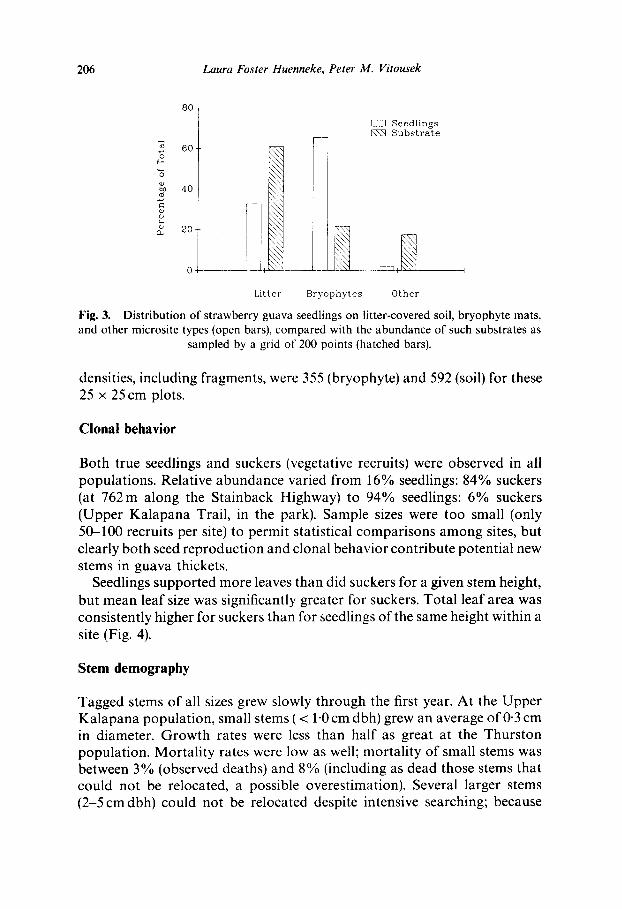
Seedlings occurred in low densities in marked quadrats in the understory of existing thickets. Few or no P. cattleianum seedlings became established in soil disturbance quadrats; where they did, they were located on the mounded edge of such disturbances, not in the central depressions. Natural ly occurring seedlings were found chiefly on bryophyte mats, far out of proport ion to the availability of such substrates on the forest floor (Fig. 3). Relatively few seedlings were observed in pig-caused disturbances or depressions. For example, of 10 1-m 2 quadrats placed randomly in the Upper Kalapana populat ion, where old and fresh disturbance was abundant , the highest seedling densities occurred either on the raised rims of old pig diggings or on hummocky, mossy substrates.
Only 5 intact seeds were obtained from each of the shallow blocks of substrate collected from the 460 m Stainback site; none of these germinated when placed in labora tory conditions. However, both blocks contained high densities o f seeds with fractured seed coats typical of germinated or partially germinated seeds (198 for the bryophyte layer, 534 for the soil). Total
206 Laura Foster Huenneke, Peter M. Vitousek
80
60. [....
's
40 r ' . q~
20
' ]]~ _ __
KSS] Seedlings E~5] Substrate
Litter Bryophytes Other
Fig. 3. Distribution of strawberry guava seedlings on litter-covered soil, bryophyte mats, and other microsite types (open bars), compared with the abundance of such substrates as
sampled by a grid of 200 points (hatched bars).
densities, including fragments, were 355 (bryophyte) and 592 (soil) for these 25 x 25 cm plots.
Clonal behavior
Both true seedlings and suckers (vegetative recruits) were observed in all populations. Relative abundance varied from 16% seedlings: 84% suckers (at 762m along the Stainback Highway) to 94% seedlings: 6% suckers (Upper Kalapana Trail, in the park). Sample sizes were too small (only 50-100 recruits per site) to permit statistical comparisons among sites, but clearly both seed reproduction and clonal behavior contribute potential new stems in guava thickets.
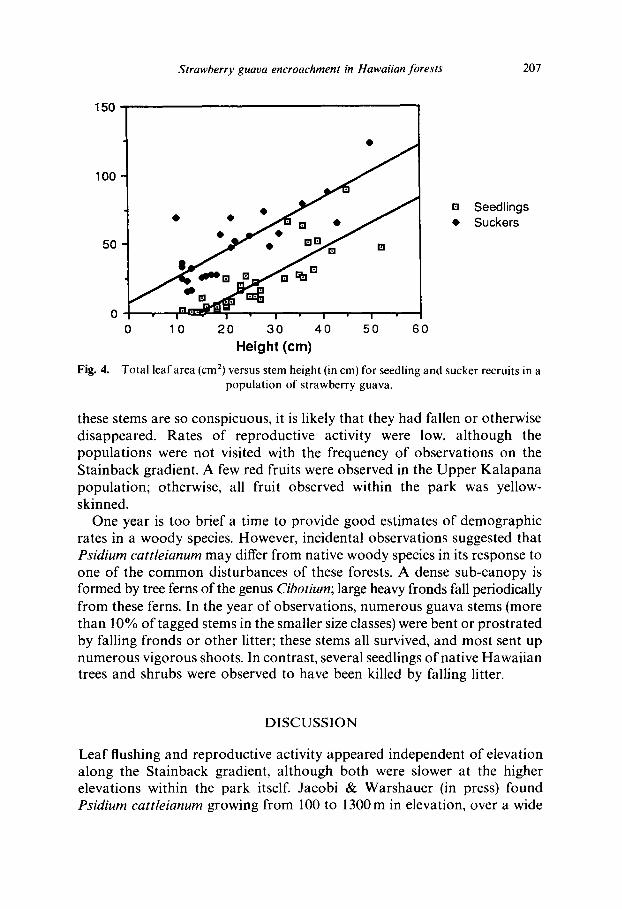
Seedlings supported more leaves than did suckers for a given stem height, but mean leaf size was significantly greater for suckers. Total leaf area was consistently higher for suckers than for seedlings of the same height within a site (Fig. 4).
Stem demography
Tagged stems of all sizes grew slowly through the first year. At the Upper Kalapana population, small stems ( < 1"0 cm dbh) grew an average of 0"3 cm in diameter. Growth rates were less than half as great at the Thurston population. Mortali ty rates were low as well; mortali ty of small stems was between 3% (observed deaths) and 8% (including as dead those stems that could not be relocated, a possible overestimation). Several larger stems (2-5 cm dbh) could not be relocated despite intensive searching; because
Strawberry guava encroachment in Hawaiian forests 207
150
1 O0
o
Fig. 4.
0 10 20 30 40 50 60
Height (era)
[] Seedlings • Suckers
Total leaf area (cm 2) versus stem height (in cm) for seedling and sucker recruits in a population of strawberry guava.
these stems are so conspicuous, it is likely that they had fallen or otherwise disappeared. Rates of reproductive activity were low, although the populations were not visited with the frequency of observations on the Stainback gradient. A few red fruits were observed in the Upper Kalapana population; otherwise, all fruit observed within the park was yellow- skinned.
One year is too brief a time to provide good estimates of demographic rates in a woody species. However, incidental observations suggested that Psidium cattleianum may differ from native woody species in its response to one of the common disturbances of these forests. A dense sub-canopy is formed by tree ferns of the genus Cibotium; large heavy fronds fall periodically from these ferns. In the year of observations, numerous guava stems (more than 10% of tagged stems in the smaller size classes) were bent or prostrated by falling fronds or other litter; these stems all survived, and most sent up numerous vigorous shoots. In contrast, several seedlings of native Hawaiian trees and shrubs were observed to have been killed by falling litter.
DISCUSSION
Leaf flushing and reproductive activity appeared independent of elevation along the Stainback gradient, although both were slower at the higher elevations within the park itself. Jacobi & Warshauer (in press) found Psidium cattleianum growing from 100 to 1300m in elevation, over a wide

208 Laura Foster Huenneke, Peter M. Vitousek
range in rainfall conditions. The plant has broad environmental tolerances; while there are obviously two genetic entities (the red- and the yellow-fruited forms), the contribution of genetic variability within each form to those broad tolerances has not been explored. It also remains unclear whether environmental conditions at higher elevations represent any barrier to guava's further spread.
Both the highest elevational station along the Stainback gradient, and most populations within the park boundaries, have lower stem densities and total stem area than do the densest thickets at intermediate elevations. It is not clear whether these stands are merely younger, and have not yet established a dense stand, or whether differences in native forest structure or in physical environment limit stand structure. The ability of strawberry guava to produce dense populations of seedlings and suckers, even in thickets with low light availability, suggests that most invading populations are eventually capable of reaching high densities.
Prolific fruit production, high rates of germination, and broad environmental tolerances frequently typify weedy plants (Baker, 1974; Bazzaz, 1986). In these respects P. cattleianum performs as expected. Interestingly, scarification lowered guava germination rates. Diong (1982) found that passage through the gut of feral pigs had little effect on total germination of Psidium cattleianum, but did shorten the time required for germination. Possibly scarification thins the tough seed coat enough to make the embryo more sensitive to minor or short-term drying events (as suggested by our laboratory results). At any rate, it is clear that high germination rates are possible without animal dispersal or scarification.
Seedling establishment in the field appears to be independent of soil disturbance, such as that provided by the rooting activities of feral pigs. Few germinants established in experimental soil disturbances, despite high seed inputs; naturally occurring seedlings were found disproportionately often on bryophyte mats and other undisturbed substrates. The same substrates support germination of many native forest plants, as well. In this regard strawberry guava is behaving no differently from native species. Much more information is needed on possible interactions between guava and seedlings of native plants.
Post-dispersal seed predation may also affect establishment rates. Potential seed predators include several species of non-native rodents and of ground-feeding birds. Removal by rats or by birds may explain the difficulty we had relocating seeds on the soil surface in the germination plots. However, the high densities of rotting fruits, and of seeds or seed fragments in the soil, indicate that many seeds escape removal by seed predators.
Our results suggest that germination and establishment do not depend on seed dispersal or 'processing' by animals. This does not imply, however, that

Strawberry guava encroachment in Hawaiian forests 209
animal dispersers are unimportant in strawberry guava's spread. The fruits are attractive to numerous vertebrates, from rodents to birds to pigs (and humans!). Effective dispersal of seeds over moderate distances is un- doubtedly a major component of the plant's invasiveness. This potential for dispersal also represents a major obstacle for control efforts.
Although we found no dependence of guava germination on soil disturbance by pigs, there are other possible links between feral pigs and P. cattleianum. Diong (1982) found high densities of viable guava seeds in pig droppings in Maui's Kipahulu Valley, where Psidium was spreading rapidly. Fewer than 5% of guava seeds in pig fecal material were fractured; germination totals were high and germination of gut-treated seed proceeded more rapidly than did that of untreated seed. Dung piles might also provide the same sort of protection from desiccation as do bryophyte-covered substrates. Diong (1982) pointed out that pig-dispersed seed might also contribute to a buried seed bank, as seed-laden droppings were often trampled into the soil.
While we have no quantitative information on the seed bank, our preliminary observations suggest that the majority of guava seeds germinate quickly (and often unsuccessfully). The few intact seeds found in the soil samples failed to germinate in the laboratory; it may be that seeds either germinate quickly, or die. Potential recruits may be concentrated in a 'seedling bank' rather than in a dormant stage. The presence or absence of viable seeds in the soil will influence the success of control strategies that emphasize the removal of above-ground plants.
Most guava populations apparently contain recruits produced both by seed reproduction and by clonal behavior. In many plants clonal 'offspring' have higher growth rates and survivorship than do true seedlings (Harper, 1977). Thus, suckers may be more effective at forming high leaf areas and rapidly dominating the forest floor, because of resources acquired from the parent. Detailed demographic observations, or else the use of genetic markers to identify ramets from particular genetic individuals, will be necessary to reconstruct the actual dynamics of seedlings and of suckers in thicket development.
There is little information on the demography of most native Hawaiian plants, except for the well-documented stand dynamics of Metrosideros explicated by Mueller-Dombois (Mueller-Dombois, 1987; Jacobi et al., 1988). The sprouting response of guava individuals to a common understory disturbance (falling tree fern fronds) constitutes an important difference between guava and at least some native species. A priority for future re- search is the investigation of differences in clonal behavior and disturbance response between strawberry guava and the native forest understory plants it is displacing.

210 Laura Foster Huenneke, Peter M. Vitousek
Our findings provide a preliminary basis for evaluating possible control methods. Prolific fruit production and high rates of suckering suggest that a dense stand could be generated (or regenerated) quickly from even a very small population. Measures which merely decrease local densities, therefore, are likely to have only temporary effects. Certainly an understanding of the physiological and environmental factors controlling suckering should be sought, with the objective of limiting vegetative spread. Dispersal is another process that needs to be studied; control of feral pigs, as currently emphasized in the park, will do little to slow the plant's spread if smaller vertebrates (birds and rodents) are also effective dispersal vectors. On the other hand, some aspects of strawberry guava's seed biology bode well for management's control efforts. Soil disturbances did not result in bursts of seedling recruitment, and in the one site studied there were few viable seeds present in the soil. Local eradication programs may be more successful if there is no pool of dormant seeds in the soil.
ACKNOWLEDGEMENTS
This research was supported by a grant from the National Geographic Society. The research staff of Hawaii Volcanoes National Park, especially Charles and Danielle Stone, Linda Cuddihy, and Steve Anderson, have provided logistic support as well as invaluable discussion and insights. Joyce Jacobson collected the phenological data and Melanie Florence supervised the germination tests. We also thank L. Loope, D. Mueller-Dombois, and L. Walker for useful discussions. C. Stone, L. Cuddihy, and C. Smith provided helpful comments on an earlier draft.
REFERENCES
Baker, H. G. (1974). The evolution of weeds. Ann. Rev. Ecol. & Systemat., 5, 1-24. Bazzaz, F. A. (1986). Life history of colonizing plants: some demographic, genetic,
and physiological features. In Ecology o]Biological Invasions of North America and Hawaii, ed. H. A. Mooney & J. A. Drake. Springer-Verlag, New York, pp. 96-110.
Diong, C. H. (1982). Population biology and management of the feral pig (Sus scroja) in Kipahulu Valley, Maui. PhD dissertation, University of Hawaii.
Harper, J. L. (1977). Population Biology of" Plants. Academic Press, London. Jacobi, J. D. & Warshauer, F. R. (in press). Distribution of six alien plant species in
upland habitats on the island of Hawaii. In Alien Plant Invasions in Native Ecosystems of Hawaii: Management and Research, ed. C. P. Stone, C. W. Smith & J. T. Tunison. University of Hawaii Press, Cooperative National Park Studies Unit, Honolulu.

Strawberry guava encroachment in Hawaiian forests 211
Jacobi, J. D., Gerrish, G., Mueller-Dombois, D. & Whiteaker, L. D. (1988). Stand- level dieback and Metrosideros regeneration in the montane rain forest of Hawaii. GeoJournal, 17, 193-200.
Joenje, W., Bakker, W. & Vlijm, L. (eds) (1987). The ecology of biological invasions. Proc. Kon. Ned. Akad. Wetensch., Series C, 90(1).
Kornberg, H. & Williamson, M. H. (eds)(1986). Quantitative Aspects of the Ecology of Biological Invasions. Royal Society, London.
Mooney, H. A. & Drake, J. A. (1986). Ecology of Biological Invasions of North America and Hawaii. Springer-Verlag, New York.
Mueller-Dombois, D. (1987). Forest dynamics in Hawaii. Trends Ecol. Evol., 2, 216-20.
Smith, C. W. (1985). Impact of alien plants on Hawaii's native biota. In Hawaii's Terrestrial Ecosystems: Preservation and Management, ed. C. P. Stone & J. M. Scott. Cooperative National Park Resources Studies Unit, Honolulu, pp. 180-250.
Vitousek, P. M. (1986). Biological invasions and ecosystem properties: can species make a difference? In Ecology of Biological Invasions of North America and Hawaii, ed. H. A. Mooney & J. A. Drake. Springer-Verlag, New York, pp. 163-78.
Wagner, W. L., Herbst, D. R. & Sohmer, S. H. (in press). Manual of the Flowering Plants of Hawaii. Bernice P. Bishop Museum and University of Hawaii Press, Honolulu.