seasonal climate effects on root colour and compounds of red radish
TRANSCRIPT

Seasonal climate effects on root colour andcompounds of red radishM Schreiner,1* S Huyskens-Keil,2 P Peters,1 I Schonhof,1 A Krumbein1 and S Widell11Institute of Vegetable and Ornamental Crops Großbeeren/Erfurt eV, Theodor-Echtermeyer-Weg 1, D-14979 Großbeeren, Germany2Humboldt-University Berlin, Institute for Horticultural Sciences, Section Quality Dynamics/Postharvest Physiology, Lentzeallee 75,D-14195 Berlin, Germany
Abstract: ‘Nevadar’ radishes were grown throughout the year. Climate parameters (mean
temperature, mean irradiation) and quality characteristics of radishes fulfilling consumer quality
requirements, such as root colour, glucosinolates, monosaccharides and pectic substances, were
determined. The quality characteristics strongly differed depending on the seasonal climate
conditions. The seasonal dependence ranged from a slight climate influence (alkenyl glucosinolates
r2=0.23), over a moderate climate effect (indolyl glucosinolates r2=0.40, glucose r2=0.50) up to a
strongly distinctive climate influence (hue angle r2=0.77, chroma r2=0.72, fructose r2=0.81, pectic
substances r2=0.99). Therefore, according to consumer-oriented quality production of radish, the
temperature and irradiation influence should be taken into account in the production process.
Recommendations for quality production of radish will be the selection of bright red cultivars marked
by a high photosynthetic capacity (yield>0.80mV) at relatively low mean irradiation intensities
(50–100mmol m2s�1) and lower mean temperatures (11–13°C). Thus sufficient photochemical energy
can be provided for the synthesis of quality-determining compounds. For the production of bioactive
radishes showing particularly relatively high contents of indolyl glucosinolates, cultivation should be
carried out in spring and autumn. In summer cultivation, consumer preferences in taste can
particularly be satisfied with the desired contents of alkenyl glucosinolates and monosaccharides.
# 2002 Society of Chemical Industry
Keywords: seasonal climate effects; temperature; photosynthetic photon flux density; colour; carbohydrates;glucosinolates; pectic substances; radish; Raphanus sativus L var sativus
INTRODUCTIONThe national and international market is characterised
by surplus production of vegetable products, resulting
in strong competition. Only those products that fulfil
consumer quality requirements have good sales poten-
tial.1 Therefore, regarding customer-oriented quality
production, one has to decisively determine:
. the quality characteristics of a vegetable product
that meet customer expectations;
. the parameters influencing these quality characteris-
tics.
In terms of consumer preference, root colour is one
of the main quality characteristics of radish and is
regarded as an indicator of radish quality. Hence
product colour strongly affects consumer perception
of quality and is one of the major characteristics in
making purchasing decisions on fresh vegetables.2–5
This is also valid for radish roots. A consumer
acceptance test in Germany (Berlin and Brandenburg)
showed that consumers preferred bright-reddish
radishes. Moreover, these investigations revealed that
taste and texture of the radish roots are also essential
quality attributes for consumers.6 Therefore, in the
present study, root colour and taste- and texture-
determining quality characteristics of radishes, such as
glucosinolates, monosaccharides and pectic sub-
stances, were analysed owing to their sensory impor-
tance. According to Widell et al,7 glucosinolates
provide mainly the taste and aftertaste attributes
‘pungent’, ‘intensive’ and ‘burning’. The monosac-
charides glucose and fructose also influence the taste
attribute ‘intensive’. In addition, the mouthfeel im-
pression ‘firm’ is predominantly determined by pectic
substances.8
Radishes are grown throughout the year under
varying climate conditions.9 Thus an understanding
of the climate factors modulating radish root colour
and composition during the production process is
essential for total quality management aiming at
consumer-oriented radish quality.
The objectives of these investigations were:
. the determination of seasonal effects, with special
emphasis on the effects of temperature and photo-
(Received 23 July 2001; accepted 24 April 2002)
* Correspondence to: M Schreiner, Institute of Vegetable and Ornamental Crops Großbeeren/Erfurt eV, Theodor-Echtermeyer-Weg 1,D-14979 Großbeeren, GermanyE-mail: [email protected]
# 2002 Society of Chemical Industry. J Sci Food Agric 0022–5142/2002/$30.00 1325
Journal of the Science of Food and Agriculture J Sci Food Agric 82:1325–1333 (online: 2002)DOI: 10.1002/jsfa.1189

synthetic photon flux density, on radish root colour
and texture- and taste-influencing compounds;
. the development of recommendations for radish
production characterised by consumer-oriented
quality.
MATERIALS AND METHODSPlant materialBased on the results of the consumer acceptance test
of radishes in Germany,6 radishes of the cultivar
‘Nevadar’ with the preferred bright red colour served
as investigation material. This cultivar forms a red,
round tuber consisting mainly of a thickened hypoco-
tyl. In this paper the term root is used as it is generally
done in the horticultural literature. The plant material
originated from 10 growth sets (Table 1). From
November until March the radishes were cultivated
in the greenhouse; from April until October they were
field grown. Fertilisation, irrigation and plant protec-
tion corresponded to standard cultivation procedures
for radishes.
The developmental stage influences the content of
glucosinolates,10 pectic substances11 and monosac-
charides.12 Thus the radishes were harvested in a
narrow range of physiological development marked by
the BBCH stages 41–43. The BBCH stages are
defined according to Bleiholder et al,13 where the first
number of the digit code describes the principal
growth stage (1–9), ie the number 4 representing the
growth stage ‘development of harvestable vegetative
plant parts’. The second number characterises the
secondary growth stage; for example, the number 5
means that 50% of the expected root diameter is
developed.
The radishes were harvested according to German
quality standards for radishes14 and selected for
uniformity of root diameter (BBCH 41: 22�1mm;
BBCH 43: 24�1mm), root shape (round, not oval)
and visual appearance (freedom from defects and
disorders).
Climate parametersAs climate parameters, the mean temperature and
mean photosynthetic photon flux density (PPFD)
were determined 12 times per hour and summarised as
means over the production period (Fig 1). Summaris-
ing the values of mean temperature and mean PPFD
as mean values over the last week or the last 2 weeks
before harvest respectively showed the same tendency
as the summarised values over the entire production
period. Therefore, for characterising the seasonal
climate influence preharvest on the colour and
compounds of ‘Nevadar’ radish, only the mean values
over the entire production period were used.
Determination of quality characteristicsFor determining the seasonal climate effects on radish
root quality, the root colour and important sensory
quality characteristics such as glucosinolates, mono-
saccharides and pectic substances were measured and
analysed at harvest. According to Schouten et al,15 thechlorophyll fluorescence values characterises the effi-
ciency of photosystem II and therefore the energy
supply of the plant as prerequisite for the synthesis of
compounds. Hence the determination of compound
Table 1. Dates of sowing and harvest, harvest parameters and investigated characteristics of ‘Nevadar’ radish
Growth
set Sowing date Harvest date
BBCH
stage
Diameter
(mm) Investigated characteristics
1 16 Dec 1997 6 Feb 1998 43 24�1 Glucosinolates, monosaccharides, colour, chlorophyll fluorescence
2 27 Jan 1998 23 Mar 1998 43 24�1 Glucosinolates, monosaccharides, colour, chlorophyll fluorescence
3 11 Mar 1998 22 Apr 1998 43 24�1 Glucosinolates, monosaccharides, pectic substances, colour
4 8 Apr 1998 11 May 1998 43 24�1 Glucosinolates, monosaccharides, colour, chlorophyll fluorescence
5 6 May 1998 2 Jun 1998 43 24�1 Glucosinolates, monosaccharides, pectic substances, colour, chlorophyll fluorescence
6 3 Jul 1998 27 Jul 1998 43 24�1 Glucosinolates, monosaccharides, colour, chlorophyll fluorescence
7 3 Aug 1998 27 Aug 1998 43 24�1 Glucosinolates, monosaccharides, pectic substances, colour, chlorophyll fluorescence
8 24 Aug 1998 1 Oct 1998 43 24�1 Glucosinolates, monosaccharides, pectic substances, colour, chlorophyll fluorescence
9 29 Sep 1998 26 Nov 1998 41 22�1 Glucosinolates, monosaccharides, colour, chlorophyll fluorescence
10 4 Nov 1998 21 Jan 1999 43 24�1 Pectic substances, colour, chlorophyll fluorescence
Figure 1. Mean temperature and mean PPFDduring experimental period.
1326 J Sci Food Agric 82:1325–1333 (online: 2002)
M Schreiner et al

composition was accompanied by that of chlorophyll
fluorescence.
For the analysis of internal quality characteristics, a
mixed sample comprising 30 radish roots was pre-
pared. For the determination of glucosinolates and
monosaccharides, whole radish roots (n=30 for each
treatment) were frozen (�28°C), then freeze-dried
and finely ground. Root material for pectin analysis
(n=30 for each treatment) was diced, and 10g of each
sample was frozen with liquid nitrogen and kept at
�28°C until further analysis.
GlucosinolatesA modified HPLC method of Lange et al16 was used
for the determination of glucosinolates. A 0.5g portion
of freeze-dried root tissue was immersed for 1min in a
water bath at 75°C and extracted with 10ml of
methanol/water mixture (7:3 v/v, T =70°C), then
centrifuged with the addition of 2ml of 0.4M barium
acetate. After a second treatment of the residue the
extracts were combined and refilled to 25ml. A 5ml
aliquot of extract was applied to a DEAE-Sephadex A-
25 anion exchanger (Sigma Chemie) and washed with
10ml of doubly distilled water. After application of
250ml of cleaned arylsulphatase solution (Boehringer
Mannheim) and an incubation period of 12h the
desulphonated glucosinolates were eluted with 3ml of
doubly distilled water.
The analysis of the desulphoglucosinolates was
carried out with HPLC equipment from Bischoff
(HPLC compact pump, UV-vis detector LAMBDA
1000, automatic sample generator model, HPCL soft-
ware Starlet) on a Spherisorb ODS2 column (5mm,
250mm�4mm). It was processed with a gradient of
0–20% acetonitrile in water from 2 to 34min, followed
by 20% acetonitrile in water for 6min and then 100%
acetonitrile for 10min. The analysis was carried out
with a flow of 1.3ml min�1 and at a wavelength of
229nm. A sample of 10ml was used for analysis after
clarification with a 0.45mm filter. The glucosinolate
content was calculated using sinigrin as external
standard and the response factor of each compound
relative to sinigrin. Analyses were performed in three
replications per treatment.
MonosaccharidesGlucose and fructose were analysed enzymatically17 in
freeze-dried root tissue. Analyses were performed in
three replications per treatment.
Pectic substancesCell wall extraction of radish roots was conducted as
described by McComb and McCready18 and
Blumenkrantz and Asboe-Hansen19 and modified by
Huyskens.20 The frozen material was blended in an
Ultra-Turrax with 95% acetone and then boiled for
20min. After boiling, the suspension was vacuum
filtered. The residue on the filter paper was resus-
pended subsequently in 95vol% acetone, in 70vol%
ethanol and finally again in acetone (95vol%). The
final white residue on the filter paper was dried
overnight at 70°C. This fraction, the alcohol-insoluble
solids (AIS), was weighed and stored in a vacuum
desiccator over silica gel until further analysis.
The isolated AIS was fractionated into three pectin
fractions, the water-soluble pectin fraction (WSP), the
EDTA-soluble pectin fraction (EDTA-SP) and the
insoluble pectin fraction (ISP), and total pectic
substances (TSP) according to the method described
by Blumenkrantz and Asboe-Hansen.19 The colori-
metric determination of the pectin fractions was
conducted using the carbazole method described by
McComb and McReady.18 The amount of galacturo-
nic acid was measured in each fraction photometrically
at 520nm. Analyses were performed in five replica-
tions for each treatment.
Colour measurementRoot colour was measured with a Minolta LR 321
colorimeter (Minolta Camera Co, Osaka, Japan) using
a white standard and standardised light type D65.
Colour measurements were expressed in the L*a*b*scale, where L* indicates the luminescence, a* repre-
sents the green–red colour axis and b* the blue–yellow
axis. The average of 10 equatorial measurements on
40 roots was recorded for each sample. Two derived
functions were computed from the recorded L*, a*and b* values as follows:
. chroma
C ¼ ½ða*Þ2 þ ðb*Þ2�1=2
. hue angle
H ¼ tan�1ðb*=a*Þ
The chroma characterises the colour saturation; the
hue angle marks the colouration.
Chlorophyll fluorescence measurementMeasurement of the chlorophyll fluorescence was
conducted with a Mini-Pam (Walz, Effeltrich, Ger-
many) on the first pair of leaves after a 20min
adaptation to darkness under standardised conditions
(20°C air temperature, 60–65% relative air humidity).
Four measurements in the middle part of the leaf area
of 40 radishes were taken per sample. A pulse-modu-
lated light source was used. The intensity of the
measuring actinic light was about 0.02mmolm2s�1.
This allowed an accurate assessment of the minimal
fluorescence (Fo).21 Applying a saturating light pulse
(duration 30s, intensity 3000mmolm2s�1), the maxi-
mal fluorescence (Fm) was measured. This procedure
was repeated after light adaptation, and the current
fluorescence (F) and maximal fluorescence (Fm’) weredetected. The following derived functions were calcu-
lated from these chlorophyll fluorescence values as
follows:22
. quantum yield
yield ¼ ðFm � FoÞ=Fm
J Sci Food Agric 82:1325–1333 (online: 2002) 1327
Seasonal effects on root colour and compounds of red radish
. photochemical quenching coefficient
qP ¼ ðFm0 � FÞ=ðFm0 � FoÞ
Statistical AnalysesThe results were analysed by analysis of variance
and calculation of correlations and regression with
STATISTICA23 and TableCurve24 respectively. Sig-
nificant differences are marked by different letters.
The number of samples necessary for the colour
measurement was determined with CADEMO25
(p=0.05; coefficient of variation 20%).
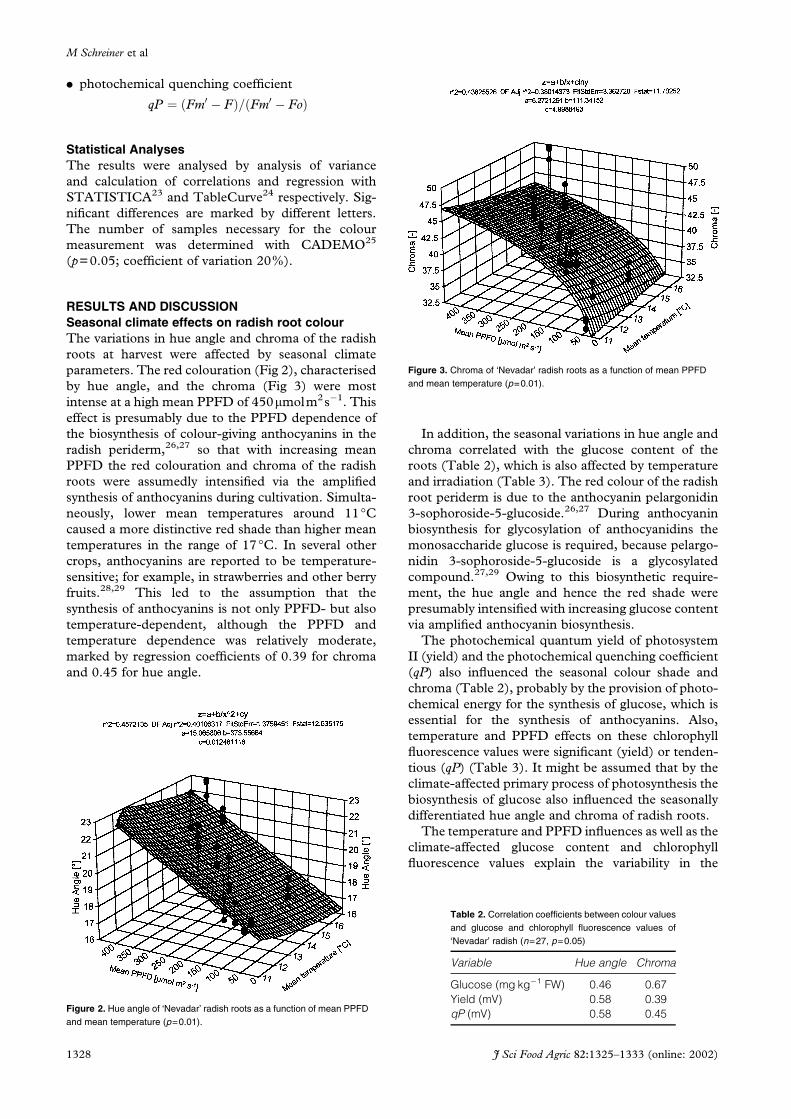
RESULTS AND DISCUSSIONSeasonal climate effects on radish root colourThe variations in hue angle and chroma of the radish
roots at harvest were affected by seasonal climate
parameters. The red colouration (Fig 2), characterised
by hue angle, and the chroma (Fig 3) were most
intense at a high mean PPFD of 450mmolm2s�1. This
effect is presumably due to the PPFD dependence of
the biosynthesis of colour-giving anthocyanins in the
radish periderm,26,27 so that with increasing mean
PPFD the red colouration and chroma of the radish
roots were assumedly intensified via the amplified
synthesis of anthocyanins during cultivation. Simulta-
neously, lower mean temperatures around 11°Ccaused a more distinctive red shade than higher mean
temperatures in the range of 17°C. In several other
crops, anthocyanins are reported to be temperature-
sensitive; for example, in strawberries and other berry
fruits.28,29 This led to the assumption that the
synthesis of anthocyanins is not only PPFD- but also
temperature-dependent, although the PPFD and
temperature dependence was relatively moderate,
marked by regression coefficients of 0.39 for chroma
and 0.45 for hue angle.
In addition, the seasonal variations in hue angle and
chroma correlated with the glucose content of the
roots (Table 2), which is also affected by temperature
and irradiation (Table 3). The red colour of the radish
root periderm is due to the anthocyanin pelargonidin
3-sophoroside-5-glucoside.26,27 During anthocyanin
biosynthesis for glycosylation of anthocyanidins the
monosaccharide glucose is required, because pelargo-
nidin 3-sophoroside-5-glucoside is a glycosylated
compound.27,29 Owing to this biosynthetic require-
ment, the hue angle and hence the red shade were
presumably intensified with increasing glucose content
via amplified anthocyanin biosynthesis.
The photochemical quantum yield of photosystem
II (yield) and the photochemical quenching coefficient
(qP) also influenced the seasonal colour shade and
chroma (Table 2), probably by the provision of photo-
chemical energy for the synthesis of glucose, which is
essential for the synthesis of anthocyanins. Also,
temperature and PPFD effects on these chlorophyll
fluorescence values were significant (yield) or tenden-
tious (qP) (Table 3). It might be assumed that by the
climate-affected primary process of photosynthesis the
biosynthesis of glucose also influenced the seasonally
differentiated hue angle and chroma of radish roots.
The temperature and PPFD influences as well as the
climate-affected glucose content and chlorophyll
fluorescence values explain the variability in the
Figure 2. Hue angle of ‘Nevadar’ radish roots as a function of mean PPFDand mean temperature (p=0.01).
Figure 3. Chroma of ‘Nevadar’ radish roots as a function of mean PPFDand mean temperature (p=0.01).
Table 2. Correlation coefficients between colour valuesand glucose and chlorophyll fluorescence values of‘Nevadar’ radish (n=27, p=0.05)
Variable Hue angle Chroma
Glucose (mg kg�1 FW) 0.46 0.67
Yield (mV) 0.58 0.39
qP (mV) 0.58 0.45
1328 J Sci Food Agric 82:1325–1333 (online: 2002)
M Schreiner et al
annual course of hue angle and chroma to the extent of
77 and 72% respectively (Table 4), demonstrating a
relatively large seasonal preharvest influence on the
colour expression of radish roots.
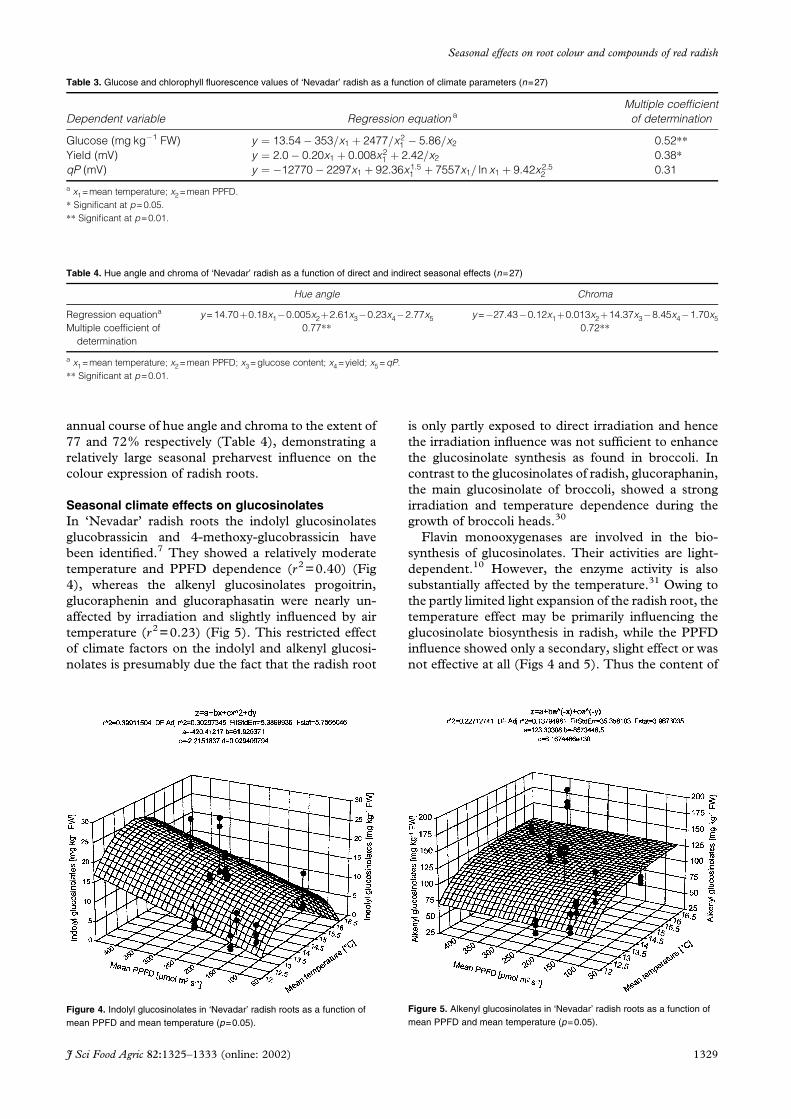
Seasonal climate effects on glucosinolatesIn ‘Nevadar’ radish roots the indolyl glucosinolates
glucobrassicin and 4-methoxy-glucobrassicin have
been identified.7 They showed a relatively moderate
temperature and PPFD dependence (r2=0.40) (Fig
4), whereas the alkenyl glucosinolates progoitrin,
glucoraphenin and glucoraphasatin were nearly un-
affected by irradiation and slightly influenced by air
temperature (r2=0.23) (Fig 5). This restricted effect
of climate factors on the indolyl and alkenyl glucosi-
nolates is presumably due the fact that the radish root
is only partly exposed to direct irradiation and hence
the irradiation influence was not sufficient to enhance
the glucosinolate synthesis as found in broccoli. In
contrast to the glucosinolates of radish, glucoraphanin,
the main glucosinolate of broccoli, showed a strong
irradiation and temperature dependence during the
growth of broccoli heads.30
Flavin monooxygenases are involved in the bio-
synthesis of glucosinolates. Their activities are light-
dependent.10 However, the enzyme activity is also
substantially affected by the temperature.31 Owing to
the partly limited light expansion of the radish root, the
temperature effect may be primarily influencing the
glucosinolate biosynthesis in radish, while the PPFD
influence showed only a secondary, slight effect or was
not effective at all (Figs 4 and 5). Thus the content of
Table 3. Glucose and chlorophyll fluorescence values of ‘Nevadar’ radish as a function of climate parameters (n=27)
Dependent variable Regression equation a
Multiple coefficient
of determination
Glucose (mg kg�1 FW) y ¼ 13:54� 353=x1 þ 2477=x21 � 5:86=x2 0.52**
Yield (mV) y ¼ 2:0� 0:20x1 þ 0:008x21 þ 2:42=x2 0.38*
qP (mV) y ¼ �12770� 2297x1 þ 92:36x1:51 þ 7557x1= ln x1 þ 9:42x2:5
2 0.31
a x1=mean temperature; x2=mean PPFD.
* Significant at p=0.05.
** Significant at p=0.01.
Table 4. Hue angle and chroma of ‘Nevadar’ radish as a function of direct and indirect seasonal effects (n=27)
Hue angle Chroma
Regression equationa y=14.70þ0.18x1�0.005x2þ2.61x3�0.23x4�2.77x5 y=�27.43�0.12x1þ0.013x2þ14.37x3�8.45x4�1.70x5Multiple coefficient of
determination
0.77** 0.72**
a x1=mean temperature; x2=mean PPFD; x3=glucose content; x4=yield; x5=qP.
** Significant at p=0.01.
Figure 4. Indolyl glucosinolates in ‘Nevadar’ radish roots as a function ofmean PPFD and mean temperature (p=0.05).
Figure 5. Alkenyl glucosinolates in ‘Nevadar’ radish roots as a function ofmean PPFD and mean temperature (p=0.05).
J Sci Food Agric 82:1325–1333 (online: 2002) 1329
Seasonal effects on root colour and compounds of red radish
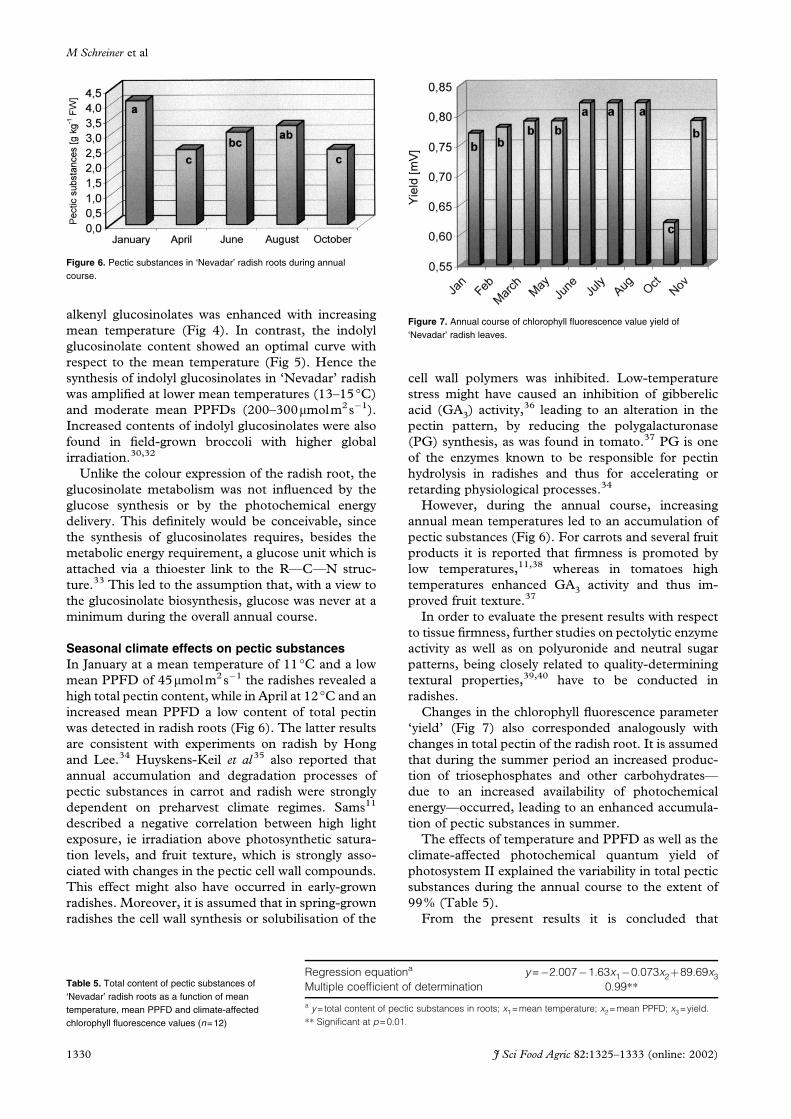
alkenyl glucosinolates was enhanced with increasing
mean temperature (Fig 4). In contrast, the indolyl
glucosinolate content showed an optimal curve with
respect to the mean temperature (Fig 5). Hence the
synthesis of indolyl glucosinolates in ‘Nevadar’ radish
was amplified at lower mean temperatures (13–15°C)
and moderate mean PPFDs (200–300mmolm2s�1).
Increased contents of indolyl glucosinolates were also
found in field-grown broccoli with higher global
irradiation.30,32
Unlike the colour expression of the radish root, the
glucosinolate metabolism was not influenced by the
glucose synthesis or by the photochemical energy
delivery. This definitely would be conceivable, since
the synthesis of glucosinolates requires, besides the
metabolic energy requirement, a glucose unit which is
attached via a thioester link to the R—C—N struc-
ture.33 This led to the assumption that, with a view to
the glucosinolate biosynthesis, glucose was never at a
minimum during the overall annual course.
Seasonal climate effects on pectic substancesIn January at a mean temperature of 11°C and a low
mean PPFD of 45mmolm2s�1 the radishes revealed a
high total pectin content, while in April at 12°C and an
increased mean PPFD a low content of total pectin
was detected in radish roots (Fig 6). The latter results
are consistent with experiments on radish by Hong
and Lee.34 Huyskens-Keil et al35 also reported that
annual accumulation and degradation processes of
pectic substances in carrot and radish were strongly
dependent on preharvest climate regimes. Sams11
described a negative correlation between high light
exposure, ie irradiation above photosynthetic satura-
tion levels, and fruit texture, which is strongly asso-
ciated with changes in the pectic cell wall compounds.
This effect might also have occurred in early-grown
radishes. Moreover, it is assumed that in spring-grown
radishes the cell wall synthesis or solubilisation of the
cell wall polymers was inhibited. Low-temperature
stress might have caused an inhibition of gibberelic
acid (GA3) activity,36 leading to an alteration in the
pectin pattern, by reducing the polygalacturonase
(PG) synthesis, as was found in tomato.37 PG is one
of the enzymes known to be responsible for pectin
hydrolysis in radishes and thus for accelerating or
retarding physiological processes.34
However, during the annual course, increasing
annual mean temperatures led to an accumulation of
pectic substances (Fig 6). For carrots and several fruit
products it is reported that firmness is promoted by
low temperatures,11,38 whereas in tomatoes high
temperatures enhanced GA3 activity and thus im-
proved fruit texture.37
In order to evaluate the present results with respect
to tissue firmness, further studies on pectolytic enzyme
activity as well as on polyuronide and neutral sugar
patterns, being closely related to quality-determining
textural properties,39,40 have to be conducted in
radishes.
Changes in the chlorophyll fluorescence parameter
‘yield’ (Fig 7) also corresponded analogously with
changes in total pectin of the radish root. It is assumed
that during the summer period an increased produc-
tion of triosephosphates and other carbohydrates—
due to an increased availability of photochemical
energy—occurred, leading to an enhanced accumula-
tion of pectic substances in summer.
The effects of temperature and PPFD as well as the
climate-affected photochemical quantum yield of
photosystem II explained the variability in total pectic
substances during the annual course to the extent of
99% (Table 5).
From the present results it is concluded that
Figure 6. Pectic substances in ‘Nevadar’ radish roots during annualcourse.
Figure 7. Annual course of chlorophyll fluorescence value yield of‘Nevadar’ radish leaves.
Table 5. Total content of pectic substances of‘Nevadar’ radish roots as a function of meantemperature, mean PPFD and climate-affectedchlorophyll fluorescence values (n=12)
Regression equationa y=�2.007�1.63x1�0.073x2þ89.69x3Multiple coefficient of determination 0.99**
a y=total content of pectic substances in roots; x1=mean temperature; x2=mean PPFD; x3=yield.
** Significant at p=0.01.
1330 J Sci Food Agric 82:1325–1333 (online: 2002)
M Schreiner et al
preharvest climate parameters, specifically the combi-
nation of temperature and PPFD, may evoke different
physiological responses in radish via hormonal and/or
enzymatic actions, leading to an alteration in pectin
metabolism.
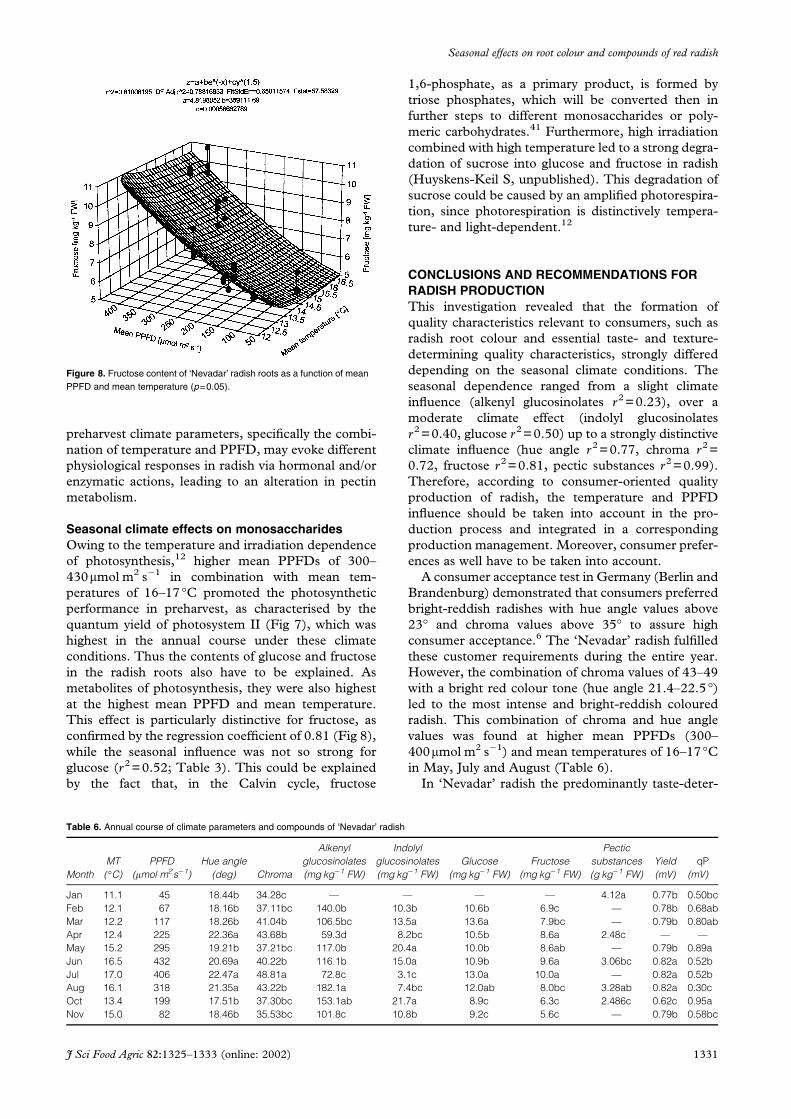
Seasonal climate effects on monosaccharidesOwing to the temperature and irradiation dependence
of photosynthesis,12 higher mean PPFDs of 300–
430mmol m2 s�1 in combination with mean tem-
peratures of 16–17°C promoted the photosynthetic
performance in preharvest, as characterised by the
quantum yield of photosystem II (Fig 7), which was
highest in the annual course under these climate
conditions. Thus the contents of glucose and fructose
in the radish roots also have to be explained. As
metabolites of photosynthesis, they were also highest
at the highest mean PPFD and mean temperature.
This effect is particularly distinctive for fructose, as
confirmed by the regression coefficient of 0.81 (Fig 8),
while the seasonal influence was not so strong for
glucose (r2=0.52; Table 3). This could be explained
by the fact that, in the Calvin cycle, fructose
1,6-phosphate, as a primary product, is formed by
triose phosphates, which will be converted then in
further steps to different monosaccharides or poly-
meric carbohydrates.41 Furthermore, high irradiation
combined with high temperature led to a strong degra-
dation of sucrose into glucose and fructose in radish
(Huyskens-Keil S, unpublished). This degradation of
sucrose could be caused by an amplified photorespira-
tion, since photorespiration is distinctively tempera-
ture- and light-dependent.12
CONCLUSIONS AND RECOMMENDATIONS FORRADISH PRODUCTIONThis investigation revealed that the formation of
quality characteristics relevant to consumers, such as
radish root colour and essential taste- and texture-
determining quality characteristics, strongly differed
depending on the seasonal climate conditions. The
seasonal dependence ranged from a slight climate
influence (alkenyl glucosinolates r2=0.23), over a
moderate climate effect (indolyl glucosinolates
r2=0.40, glucose r2=0.50) up to a strongly distinctive
climate influence (hue angle r2=0.77, chroma r2=0.72, fructose r2=0.81, pectic substances r2=0.99).
Therefore, according to consumer-oriented quality
production of radish, the temperature and PPFD
influence should be taken into account in the pro-
duction process and integrated in a corresponding
production management. Moreover, consumer prefer-
ences as well have to be taken into account.
A consumer acceptance test in Germany (Berlin and
Brandenburg) demonstrated that consumers preferred
bright-reddish radishes with hue angle values above
23° and chroma values above 35° to assure high
consumer acceptance.6 The ‘Nevadar’ radish fulfilled
these customer requirements during the entire year.
However, the combination of chroma values of 43–49
with a bright red colour tone (hue angle 21.4–22.5°)led to the most intense and bright-reddish coloured
radish. This combination of chroma and hue angle
values was found at higher mean PPFDs (300–
400mmol m2 s�1) and mean temperatures of 16–17°Cin May, July and August (Table 6).
In ‘Nevadar’ radish the predominantly taste-deter-
Figure 8. Fructose content of ‘Nevadar’ radish roots as a function of meanPPFD and mean temperature (p=0.05).
Table 6. Annual course of climate parameters and compounds of ‘Nevadar’ radish
Month
MT
(°C)PPFD
(mmol m2s�1)
Hue angle
(deg) Chroma
Alkenyl
glucosinolates
(mg kg�1 FW)
Indolyl
glucosinolates
(mg kg�1 FW)
Glucose
(mg kg�1 FW)
Fructose
(mg kg�1 FW)
Pectic
substances
(g kg�1 FW)
Yield
(mV)
qP
(mV)
Jan 11.1 45 18.44b 34.28c — — — — 4.12a 0.77b 0.50bc
Feb 12.1 67 18.16b 37.11bc 140.0b 10.3b 10.6b 6.9c — 0.78b 0.68ab
Mar 12.2 117 18.26b 41.04b 106.5bc 13.5a 13.6a 7.9bc — 0.79b 0.80ab
Apr 12.4 225 22.36a 43.68b 59.3d 8.2bc 10.5b 8.6a 2.48c — —
May 15.2 295 19.21b 37.21bc 117.0b 20.4a 10.0b 8.6ab — 0.79b 0.89a
Jun 16.5 432 20.69a 40.22b 116.1b 15.0a 10.9b 9.6a 3.06bc 0.82a 0.52b
Jul 17.0 406 22.47a 48.81a 72.8c 3.1c 13.0a 10.0a — 0.82a 0.52b
Aug 16.1 318 21.35a 43.22b 182.1a 7.4bc 12.0ab 8.0bc 3.28ab 0.82a 0.30c
Oct 13.4 199 17.51b 37.30bc 153.1ab 21.7a 8.9c 6.3c 2.486c 0.62c 0.95a
Nov 15.0 82 18.46b 35.53bc 101.8c 10.8b 9.2c 5.6c — 0.79b 0.58bc
J Sci Food Agric 82:1325–1333 (online: 2002) 1331
Seasonal effects on root colour and compounds of red radish

mining alkenyl glucosinolate is glucoraphasatin. The
majority of German consumers in Berlin and
Brandenburg preferred a glucoraphasatin content of
90–130mg kg�1 fresh weight, since this glucoraphasa-
tin concentration has the desired expression of the
taste attribute ‘pungent’.7 ‘Nevadar’ radishes pro-
duced in March, May, June and November fulfilled
these consumer requirements (Table 6). The different
climate conditions in these months also demonstrated
the slight climate dependence of the alkenyl gluco-
sinolates.
Consumer preferences for ‘Nevadar’ radish with
regard to colour expression and taste can be satisfied
best within the early summer months. However, to
produce radishes not only under summer climate
conditions, those bright red cultivars should be
preferred which are marked by a high photosynthetic
capacity (yield >0.80mV) at relatively low irradiation
intensities (50–100mmol m2 s�1) and lower mean
temperatures (11–13°C). Thus photochemical energy
can be delivered sufficiently for the synthesis of colour-
determining compounds. This potential would be
indicated by cultivars with distinctive formation of
chlorophyllrich leaves. This has particularly to be
taken into account for greenhouse cultivation in winter
or for cultivation with fleece or films.
These recommendations are valid not only for the
desired colour expression but also for the monosac-
charides, because the monosacharide content was also
highest under climate conditions in summer (mean
PPFD 300–400mmolm2s�1, mean temperature
16–17°C) (Table 6). Monosaccharides are not only
essential metabolites for the synthesis of anthocyanins
and glucosinolates, they also determine the taste of
radish. Increased monosaccharide contents led to a
more intense taste expression in radish.6
To satisfy the increasing health consciousness of
consumers and thus the demand for functional foods,
the content of bioactive indolyl glucosinolates should
be increased by creating lower mean temperatures
(13–15°C) with moderate mean PPFDs (200–
300mmolm2s�1) in the production process. Increased
contents of indolyl glucosinolates were also found in
broccoli with higher global irradiation.30,32
The relatively low contents of total pectin during the
entire year were associated with a weak cell wall
integrity of the radish root. Studies have demonstrated
that the sensory texture impression ‘firm’ is also
determined by the pectin content of the radish root.8
However, further investigations have to be conducted
in order to confirm quantitative relations between
pectic substances and consumer preferences.
With respect to the climate effects on quality
characteristics, various consumer preferences can be
satisfied. For the production of bioactive radishes
showing particularly relatively high contents of indolyl
glucosinolates, cultivation should be carried out in
spring and autumn. In summer cultivation, consumer
preferences in colour and taste can particularly be
satisfied with the desired contents of alkenyl glucosi-
nolates and monosaccharides as well as by the desired
colour expression.
Additionally, the type of soil should be taken into
account for consumer-oriented radish production. To
enhance the content of monosaccharides and alkenyl
glucosinolates in early summer and autumn, radishes
should be grown in heat-saving soils such as humic
loamy or loess soils comprising only a relatively low
humus content to avoid undesired colour changes. In
summer, sandy soils with a high potential for heat
emission should be selected in order to prevent low
indolyl glucosinolate contents. For early cultivation in
March and April, fleece or films could be used
independently of the soil type for enhancing the
temperature.
REFERENCES1 Geiger W, Qualitatslehre. Friedrich Vieweg, Braunschweig
(1994).
2 Gironi F and Testoni A, The relation between colour and quality
of vegetables. Acta Hort 259:141–167 (1990).
3 Clydesdale FM, Gover R, Philipsen D and Fugardi C, The effect
of color on thirst quenching, sweetness, acceptability and flavor
intensity in fruit punch flavored beverages. J Food Qual 15:19–
38 (1992).
4 Francis FJ, Quality as influenced by color. Food Qual Pref 6:149–
155 (1995).
5 Kays S, Preharvest factors affecting appearance. Postharv Biol
Technol 15:233–247 (1999).
6 Schreiner M, Schonhof I, Widell S, Krumbein A, Peters P,
Auerswald H, Huyskens-Keil S and Linke M, Qualitat von
Radies, in Annual Report 1998 of the Institute of Vegetable and
Ornamental Crops Großbeeren/Erfurt eV. Institute of Vegetable
and Ornamental Crops Großbeeren/Erfurt eV, Großbeeren,
pp 39–46 (1999).
7 Widell S, Krumbein A and AuerswaldH, Glucosinolate in Radies
und sensorische Bewertung. Wissenschaftlichen Arbeitstagung
der Deutschen Gartenbauwissenschaftlichen Gesellschaft und des
BDGL 35:151 (1998).
8 Huyskens-Keil S, Schreiner M, Widell S and Peters P, Zer-
storungsfreie Qualitatsbestimmung bei Radies. Gemuse 7:406–
408 (1998).
9 Vogel G, Handbuch des Speziellen Gemusebaus. Eugen Ulmer,
Stuttgart, pp 406–420 (1996).
10 Wallsgrove RM and Bennett RN, The biosynthesis of gluco-
sinolates in Brassicas, in Amino Acids and Their Derivatives in
Higher Plants. Society for Experimental Biology Seminar 56, Ed by
Wallsgrove RM, Cambridge University Press, Cambridge, pp
243–259 (1995).
11 Sams CE, Preharvest factors affecting postharvest texture.
Postharv Biol Technol 15:249–254 (1999).
12 Larcher W, Okophysiologie der Pflanzen. Ulmer, Stuttgart, pp 71–
112 (1994).
13 Bleiholder H, Buhr L, Feller C, Hack H, HessM, Klose R,Meier
U, Stauss R, van den Boom T and Weber E, Growth Stages of
Mono- and Dicotyledonous Plants. BBCH-Monograph. Black-
well, Berlin, pp 101–104 (1997).
14 Bickelmann U, Leitfaden fur die Anwendung der EG-Qualitatsnor-
men fur Frische Obst, Gemuse und Citrusfruchte. Rheinischer
Landwirtschaftsverlag, Bonn, pp 47–85 (1996).
15 Schouten RE, Otma EC, van Kooten O and Tijskens LM,
Keeping quality of cucumber fruits predicted by biological age.
Postharv Biol Technol 12:175–181 (1997).
16 Lange R, Petrzika M, Raab B and Linow F, Zur Kenntnis der
Schwefelverbindungen in Raps – (Brassica napus) Varietaten
und Verarbeitungsprodukten. Nahrung 35:385–389 (1991).
1332 J Sci Food Agric 82:1325–1333 (online: 2002)
M Schreiner et al

17 Boehringer Mannheim, Methoden der Enzymatischen Lebensmittel-
analytik. Boehringer Mannheim Gmb H, Mannheim (1986).
18 McComb E and McCready R, Colorimetric determination of
pectic substances. Anal Biochem 24:1630–1632 (1952).
19 Blumenkrantz N and Asboe-Hansen G, New method for quan-
titative determination of uronic acids. Anal Biochem 54:484–
489 (1973).
20 Huyskens S, Morphological, physiological and biochemical
aspects in the cultivation of two pantropical cucurbits: Luffa
acutangula L, Roxb and Momordica charantia L. Dissertation,
Rheinische Friedrich-Wilhelms-Universitat, Bonn (1991).
21 Van Kooten O and Snel J, The use of chlorophyll fluorescence
nomenclature in plant stress physiology. Photosynth Res
25:147–150 (1990).
22 Genty B, Briantais JM and Baker NR, The relationship between
the quantum yield of photosynthetic electron transport and
quenching of chlorophyll fluorescence. Biochim Biophys Acta
990:87–92 (1989).
23 StatSoft, Electronic Textbook StatSoft. StatSoft Inc, Tulsa, OK
(2001).
24 AISN Software, TableCurve 3D. Version 3.0. User’s Manual.
AISN Software Inc, Chicago, IL (1997).
25 Rasch R, Guiard V and Nurnberg G, Statistische Versuchsplanung:
Einfuhrung in die Methoden und Anwendungen des Dialogsystems
CADEMO. Gustav Fischer, Stuttgart (1992).
26 Mazza G and Miniati E, Radish, in Anthocyanins in Fruits,
Vegetables, and Grains, Ed by Mazza G and Miniati E, CRC
Press, Boca Raton, FL, pp 288–290 (1993).
27 Herrmann K, Inhaltsstoffe der Radieschen und Rettiche. Die
Industrielle Obst- und Gemuseverwertung 8:240–246 (1997).
28 Herrmann K, Vorkommen, Gehalte und Bedeutung von Inhalts-
stoffen des Obstes und Gemuses. II. Flavonoide: Catechine,
Proanthocyanide, Anthocyanide. Die Industrielle Obst- und
Gemuseverwertung 5/6:170–175 (1991).
29 Bohm H, Boeing H, Hempel J, Raab B and Kroke A, Flavonole,
Flavone und Anthocyane als naturliche Antioxidantien der
Nahrung und ihre mogliche Rolle bei der Pravention chron-
ischer Erkrankung. Z Emahrungswiss 37:147–163 (1998).
30 Schonhof I, Krumbein A and Gutezeit B, Vorerntemaßnahmen
zur Beeinflussung des Glucosinolatgehaltes in Brokkoli.
Wissenschaftlichen Arbeitstagung der Deutschen Gartenbauwissen-
schaftlichen Gesellschaft und des BDGL 36:56 (1999).
31 Ziegler H, Physiologie, in Lehrbuch der Botanik, Ed by Sitte P,
Noll F, Schenck H and Schimper AF, Gustav Fischer,
Stuttgart, pp 242–472 (1993).
32 Schonhof I, Krumbein A and Widell S, Glucosinolate in
Brassicaceae und Moglichkeiten ihrer Beeinflussung. Tagung
der Deutschen Gesellschaft fur Qualitatsforschung 35:47–56
(2000).
33 Fenwick GR and Heany RK, Glucosinolates and their break-
down products in cruciferous crops, foods and feedingstuffs.
Food Chem 11:249–271 (1983).
34 Hong SJ and Lee SK, Comparison of physiological characteris-
tics among radish cultivars after storage. J Korean Soc Hort Sci
36:812–817 (1995).
35 Huyskens-Keil S, Ulrichs C and Schreiner M, Cell wall carbo-
hydrate metabolism of perishable vegetables in pre- and post-
harvest. Acta Hort in preparation.
36 Libbert E, Lehrbuch der Pflanzenphysiologie. Gustav Fischer,
Stuttgart (1993).
37 Mignani I, Greve LC, Ben-Arie R, Stotz HU, Li C, Shackel KA
and Labavitch JM, The effect of GA3 and divalent cations on
aspects of pectin metabolism and tissue softening in ripening
tomato pericarp. Physiol Plant 93:108–115 (1995).
38 Ulrichs C and Huyskens-Keil S, Kohlenhydratmetabolismus von
Mohren mit Laub wahrend der Nachernte. Tagungsband der
Deutschen Gesellschaft fur Qualitatsforschung 33:22–26 (1997).
39 Tong C, Krueger D, Vickers Z and El-Shiekh A, Comparison of
softening-related changes during storage of ‘Honeycrisp’
apple, its parents, and ‘Delicious’. J Am Soc Hort Sci
124:407–415 (1999).
40 Sakurai N and Nevins DJ, Relationship between softening and
wall polysaccharides in avocado (Persea americana Mill) meso-
carp tissues. Plant Cell Physiol 38(5):43–49 (1998).
41 Heß D, Pflanzenphysiologie. Eugen Ulmer, Stuttgart (1999).
J Sci Food Agric 82:1325–1333 (online: 2002) 1333
Seasonal effects on root colour and compounds of red radish