screening of plant growth promoting rhizobacteria as potential microbial inoculants
TRANSCRIPT

at SciVerse ScienceDirect
Crop Protection 40 (2012) 43e48
Contents lists available
Crop Protection
journal homepage: www.elsevier .com/locate/cropro
Screening of plant growth promoting rhizobacteria as potential microbialinoculants
Éva Laslo a,*, Éva György b, Gyöngyvér Mara b, Éva Tamás a, Beáta Ábrahámb, Szabolcs Lányi b
a Politehnica University of Bucharest, Faculty of Applied Chemistry and Material Science, Bucharest 060042, Splaiul Independentei, Nr. 313, Romaniab Sapientia University, Cluj-Napoca, Faculty of Sciences, Miercurea Ciuc 530104, Libert�atii Sq, Nr. 1, Romania
a r t i c l e i n f o
Article history:Received 1 November 2011Received in revised form30 April 2012Accepted 5 May 2012
Keywords:Phytopathogenic fungiAntimycogenic effectSiderophoresRhizobacteriaGrowth rate inhibitionPhosphate solubilizationAmmonia production
* Corresponding author.E-mail address: [email protected] (É. Laslo).
0261-2194/$ e see front matter � 2012 Elsevier Ltd.doi:10.1016/j.cropro.2012.05.002
a b s t r a c t
Plant growth promoting bacteria can enhance and promote plant growth and development in differentways. These mechanisms include solubilization of phosphorus, nitrogen fixation and biocontrollingeffects on phytopathogenic microorganisms. Suppression of phytopathogenic fungi by 47 differentbacteria, isolated from different monocotyledonic plants rhizosphere and soil, was tested against Fusa-rium oxysporum radicis-lycopersici, Sclerotium bataticola, Pythium ultimum, Fusarium graminearum, andAlternaria spp. The antifungal activity of these isolates was described based on the comparison of thegrowth rate inhibition. As the production of iron-chelating compounds is one of the mechanismsresponsible for the antimycotic effect, we tested the siderophore producing capacity of the isolatedstrains. Also, we assayed the ammonia production of these bacteria. This secondary metabolitecompound contributes to the biocontrolling property of these bacteria. Our examinations also includethe inorganic phosphate solubilization capacity of these isolates, which may improve the phosphorusuptake of plants. The results indicate that 17 bacterial isolates are able to produce siderophores and 30from them possess capacity of calcium-phosphate mobilization. The majority of the cultures were foundto have highly inhibitory effects against the mycelium growth of P. ultimum, F. oxysporum radicis-lyco-persici and F. graminearum, whereas others showed little activity. Only twelve bacteria showed no activityagainst the S. bataticola plant pathogen fungus.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Plant-associated bacteria may indirectly benefit the plants bypreventing the growth or activity of plant pathogens throughdifferent mechanisms (e.g. competition for space and nutrients,antibiosis, production of hydrolytic enzymes, inhibition ofpathogen-produced enzymes or toxins) and through inductionof plant defense mechanisms (Weyens et al., 2009). The diversity ofthe rhizospheric and nodule bacteria is very high (György et al.,2010).
Biological control of plant diseases is gaining attention due toincreased pollution concerns caused by the excessive use of pesti-cides for crop protection and development of pathogen resistance.The use of environmental friendlymicroorganisms has proved to beuseful in plant growth promotion due to their role in nutrientcycling (Bhattacharyya and Jha, 2012) and disease control. Plantgrowth promoting rhizobacteria (PGPR) inoculation has proven to
All rights reserved.
be a promising agricultural approach that plays an important rolein crop protection, growth promotion or biological disease control.The use of biocontrol bacteria isolated from the rhizosphere maypresent an alternative for plant disease prevention (Compant et al.,2005; Fernando et al., 2006; Fatima et al., 2009). In crop protection,integrated pest management involves the application of differentbacteria alone or in combination with other antagonistic agents(Spadaro and Gullino, 2005).
One of the plant growth promoting mechanisms of rhizobac-teria is the antagonism against phytopathogenic microorganismsdue to the production of antimicrobial metabolites like side-rophores, antibiotics, cyanides, fungal cell wall degrading enzymesand gaseous products including ammonia (Idris et al., 2007;Lugtenberg and Kamilova, 2009). The mechanism of antifungaleffects lies on the production of a variety of antimicrobialcompounds that act in different ways. The antagonistic effects arecaused by cytolysis, leakage of potassium ions, disruption of thestructural integrity of membranes, inhibition of mycelial growthand the protein biosynthesis (Quan et al., 2010).
One of the most popular bacteria studied and exploited asbiocontrol agent is the Pseudomonas species (Ahmad et al., 2008).

É. Laslo et al. / Crop Protection 40 (2012) 43e4844
Most of the identified Pseudomonas biocontrol strains produceantifungal metabolites such phenazines, pyrrolnitrin, pyoluteorinand cyclic lipopeptides like viscosinamide. It was demonstratedthat viscosinamide prevents the infection of sugarbeet by Pythiumultimum (Bloemberg and Lugtenberg, 2001). These bacterial strainsbeside the antagonistic effect also influence the defense system ofplants (Maksimov et al., 2011).
The siderophore-mediated competition for iron is one amongthe mechanisms responsible for the antagonistic activity of Pseu-domonas spp. The secreted iron-chelating compounds bind theferric ions (Fe3þ), and are taken up by microbial cells throughspecific recognition by membrane proteins (Srivastava and Shalini,2008). The presence of iron-chelating compounds makes thebacteria better competitors for iron, preventing this way thegrowth of the pathogen microorganisms. The Pseudomonas speciesproduce two different types of siderophore: pseudobactin andpyoverdin (Oldal et al., 2002).
Siderophores produced by biocontrol bacteria have a higheraffinity for iron than those produced by some fungal pathogens,allowing the former microbes to scavenge most of the availableiron, preventing the proliferation of fungal pathogens (Hillel, 2005).
Some authors have reported that Pseudomonas fluorescens e
belonging to the PGPR class e produces siderophores and havebiocontrol effect against P. ultimum, R. batatticola, Fusarium oxy-sporum. Other Pseudomonas species like Pythium stutzeri produceextracellular enzymes like chitinase and laminase capable of lysingthe mycelia of Fusarium solani (Kumar et al., 2002; Srivastava andShalini, 2008). Pseudomonas aeruginosa under iron-limitingconditions, produces three types of siderophores: pyoverdine,pyochelin and its precursor salicylic acid, and induces resistance toplant diseases caused by Botrytis cinerea on bean and tomato, Col-letotrichum lindemuthianum on bean (Höfte and Bakker, 2007).
F. oxysporum causes vascular wilt and foot-, root- and bulbrotdiseases in a wide variety of economically important crops. Alter-naria spp., Sclerotium spp. cause leaf spots, root rot and stem rot,which also leads to serious yield losses (Chaiharn et al., 2009).
The antifungal effect of PGPRs is influenced by a lot of envi-ronmental and genetic factors. Biotic and abiotic environmentalsignals may have an important input on the regulation of biocontrolgenes in pseudomonads, e.g. on the repression of siderophorebiosynthesis. Together with low oxygen concentrations, the avail-able carbon and nitrogen sources that influence the molecularmechanisms are involved in biocontrol activity (Haas and Défago,2005).
Another plant growth promoting activity of these bacteriaconsists in solubilization of inorganic insoluble phosphates, trans-forming them into bioavailable forms. This nutrient mobilizationmay enhance crop productivity because phosphorus is a macronu-trient for plants, required for growth and development. It is alsoinvolved in photosynthesis, energy transfer, signal transduction,macromolecular biosynthesis and respiration (Zaidi et al., 2009a,2010). The insoluble phosphate-content of the soils is high, due tothe excessive application of chemical fertilizers. A considerableamount of phosphorus is rapidly fixed into less available formstrough complexation with aluminium or iron (in acidic soils) orwith calcium (in calcareous soils), before plant roots have hada chance to absorb it in orthophosphate form (Malboobi et al.,2009).
Phosphate-solubilizing bacteria have been reported forpromoting plant growth and enhancing production yield (Rodrigezand Fraga, 1999; Khan et al., 2009). Secretion of organic acids andphosphatase enzymes are common mechanisms that facilitate theconversion of insoluble forms of phosphorous to plant accessibleforms (Kumari et al., 2009). The inorganic phosphate mobilizationis realised due to organic acid production, proton release or
production of chelating substances by the bacteria (Zaidi et al.,2009b). Some soil bacteria with phytase activity contribute to thephosphorus release from organic phosphates (Singh andSatyanarayana, 2011).
The application of phosphorus biofertilizers in the form of plantgrowth promoting microorganisms can facilitate the availability ofaccumulated phosphates for plant growth and development bysolubilization. The bacteria involved in phosphorus solubilizationas well as better scavenging of soluble forms can enhance plantgrowth by increasing the efficiency of biological nitrogen fixation,enhancing the availability of other trace elements (Gyaneshwaret al., 2002).
The broad aim of this study is the development of plant growthpromoting inoculants. To our best knowledge, only one recentresearch was made in the neighbouring geographical area (Djuricet al., 2011). In the present article, we screen bacteria for side-rophore production and antifungal activity against plant patho-genic soil borne fungi as follows: F. oxysporum radicis-lycopersici,Sclerotium bataticola, P. ultimum, Fusarium graminearum and Alter-naria spp. We assayed the ammonia production capacity andinorganic phosphate solubilization trait of these isolates.
2. Materials and methods
2.1. Isolation of the bacterial strains
The bacterial strains were isolated from different mono-cotyledonous plants (Carex spp., Zea mays L.) rhizosphere and soilon King’s B selective media for Pseudomonas species. From the soiland rhizosphere samples serial dilutions were prepared in gnoto-biotic conditions, and a volume of 0.1 ml was spread on King’s Bselective media. The inoculated Petri-dishes were incubated 48 h atthe temperature of 28 �C. On the basis of morphological andphysiological characteristics we worked with 47 selected cultures,12 bacterial pure cultures originated from rhizosphere and 35 fromsoil.
2.2. Evaluation method for siderophore production
Siderophore production was detected by the universal methodof Schwyn and Neilands (1987). This assay is based on a competi-tion for iron between the ferric (Fe3þ) complex of an indicator dye,chrome azurol S (CAS), and a chelator or siderophore produced bythe microorganism. The iron is removed from CAS by the side-rophore, which apparently has a higher affinity for ferric ions(Fe3þ). The positive reaction results in a colour change of CASreagent (usually from blue to orange).
The bacterial cultures were grown in King’s B broth (containingproteose peptone 20 g, glycerol 10 ml, K2HPO4 1.5 g, MgSO4 1.5 g, in1000 ml distilled water) for 24 h at 28 �C, followed by an inocula-tion with 5 ml bacterial suspension of the modified, CAS containingKing’s B agar plates and incubation for 72 h at 28 �C. For the side-rophore producing colonies the original blue colour of mediumwaschanged in orange.
2.3. Antifungal effect assay
Isolated bacterial cultures were tested for growth inhibitoryeffect on the mycelium growth of F. oxysporum radicis-lycopersici,S. bataticola, P. ultimum, F. graminearum, and Alternaria spp. oncomplex agar medium (containing peptone 10 g, D-dextrose 40 g,yeast extract 10 g, agar 18 g). The test fungi were grown andmaintained on Czapek Dox agar (containing sucrose 30 g, NaNO3,3 g, K2HPO4 1 g, KCL 0.5 g, MgSO4$7H2O 0.5 g, FeSO4$7H2O 0.01 g,agareagar 15 g in 1000 ml distilled water) (Atlas, 2010).
0
20
40
60
80
100
BC 1BC 2
BC 3BC 4
BC 5BC 6
BC 7BC 8
BC 9
BC 10
BC 11
BC 12
BC 13
BC 14
BC 15
BC 16
BC 17
BC 18
BC 19
BC 20
BC 21
BC 22
BC 23
BC 24
BC 25
BC 26
BC 27
BC 28
BC 29
BC 30
BC 31
BC 32
BC 33
BC 34
BC 35
BC 36
BC 37
BC 38
BC 39
BC 40
BC 41
BC 42
BC 43
BC 44
BC 45
BC 46
BC 47
Isolated bacteria
Inhi
biti
on r
ate
%
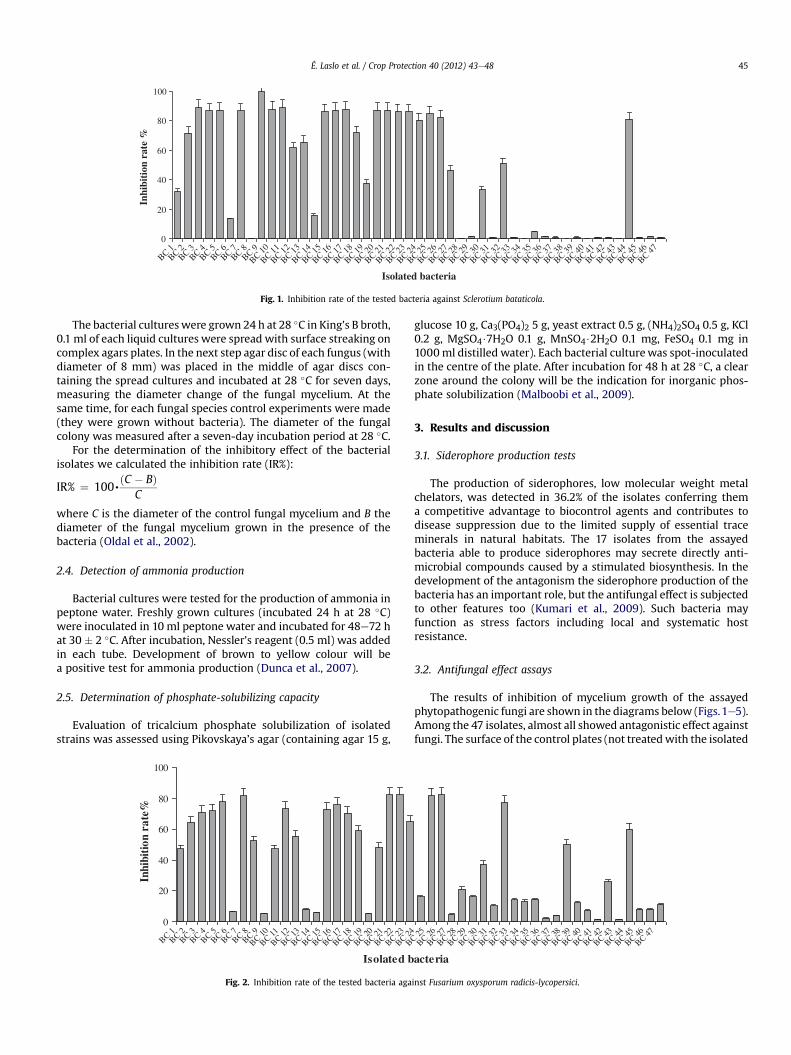
Fig. 1. Inhibition rate of the tested bacteria against Sclerotium bataticola.
É. Laslo et al. / Crop Protection 40 (2012) 43e48 45
The bacterial cultureswere grown 24 h at 28 �C in King’s B broth,0.1 ml of each liquid cultures were spread with surface streaking oncomplex agars plates. In the next step agar disc of each fungus (withdiameter of 8 mm) was placed in the middle of agar discs con-taining the spread cultures and incubated at 28 �C for seven days,measuring the diameter change of the fungal mycelium. At thesame time, for each fungal species control experiments were made(they were grown without bacteria). The diameter of the fungalcolony was measured after a seven-day incubation period at 28 �C.
For the determination of the inhibitory effect of the bacterialisolates we calculated the inhibition rate (IR%):
IR% ¼ 100,ðC � BÞ
C
where C is the diameter of the control fungal mycelium and B thediameter of the fungal mycelium grown in the presence of thebacteria (Oldal et al., 2002).
2.4. Detection of ammonia production
Bacterial cultures were tested for the production of ammonia inpeptone water. Freshly grown cultures (incubated 24 h at 28 �C)were inoculated in 10 ml peptone water and incubated for 48e72 hat 30 � 2 �C. After incubation, Nessler’s reagent (0.5 ml) was addedin each tube. Development of brown to yellow colour will bea positive test for ammonia production (Dunca et al., 2007).
2.5. Determination of phosphate-solubilizing capacity
Evaluation of tricalcium phosphate solubilization of isolatedstrains was assessed using Pikovskaya’s agar (containing agar 15 g,
0
20
40
60
80
100
BC 1BC 2
BC 3BC 4
BC 5BC 6
BC 7BC 8
BC 9
BC 10
BC 11
BC 12
BC 13
BC 14
BC 15
BC 16
BC 17
BC 18
BC 19
BC 20
BC 21
BC 22
BC 23
BC 2B
Isolated b
Inhi
biti
on r
ate%
Fig. 2. Inhibition rate of the tested bacteria aga
glucose 10 g, Ca3(PO4)2 5 g, yeast extract 0.5 g, (NH4)2SO4 0.5 g, KCl0.2 g, MgSO4$7H2O 0.1 g, MnSO4$2H2O 0.1 mg, FeSO4 0.1 mg in1000ml distilled water). Each bacterial culturewas spot-inoculatedin the centre of the plate. After incubation for 48 h at 28 �C, a clearzone around the colony will be the indication for inorganic phos-phate solubilization (Malboobi et al., 2009).
3. Results and discussion
3.1. Siderophore production tests
The production of siderophores, low molecular weight metalchelators, was detected in 36.2% of the isolates conferring thema competitive advantage to biocontrol agents and contributes todisease suppression due to the limited supply of essential traceminerals in natural habitats. The 17 isolates from the assayedbacteria able to produce siderophores may secrete directly anti-microbial compounds caused by a stimulated biosynthesis. In thedevelopment of the antagonism the siderophore production of thebacteria has an important role, but the antifungal effect is subjectedto other features too (Kumari et al., 2009). Such bacteria mayfunction as stress factors including local and systematic hostresistance.
3.2. Antifungal effect assays
The results of inhibition of mycelium growth of the assayedphytopathogenic fungi are shown in the diagrams below (Figs.1e5).Among the 47 isolates, almost all showed antagonistic effect againstfungi. The surface of the control plates (not treatedwith the isolated
4C 25
BC 26
BC 27
BC 28
BC 29
BC 30
BC 31
BC 32
BC 33
BC 34
BC 35
BC 36
BC 37
BC 38
BC 39
BC 40
BC 41
BC 42
BC 43
BC 44
BC 45
BC 46
BC 47
acteria
inst Fusarium oxysporum radicis-lycopersici.
0
20
40
60
80
100
BC 1BC 2
BC 3BC 4
BC 5BC 6
BC 7BC 8
BC 9
BC 10
BC 11
BC 12
BC 13
BC 14
BC 15
BC 16
BC 17
BC 18
BC 19
BC 20
BC 21
BC 22
BC 23
BC 24
BC 25
BC 26
BC 27
BC 28
BC 29
BC 30
BC 31
BC 32
BC 33
BC 34
BC 35
BC 36
BC 37
BC 38
BC 39
BC 40
BC 41
BC 42
BC 43
BC 44
BC 45
BC 46
BC 47
Isolated bacteria
Inhi
biti
on r
ate
%
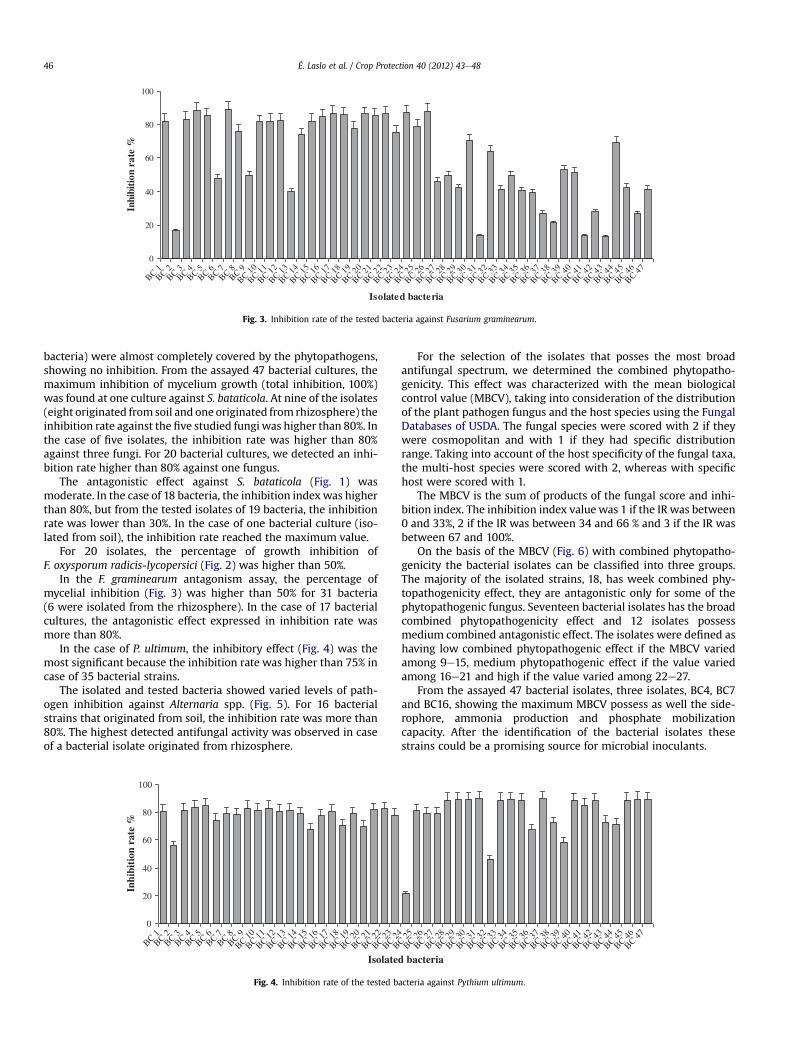
Fig. 3. Inhibition rate of the tested bacteria against Fusarium graminearum.
É. Laslo et al. / Crop Protection 40 (2012) 43e4846
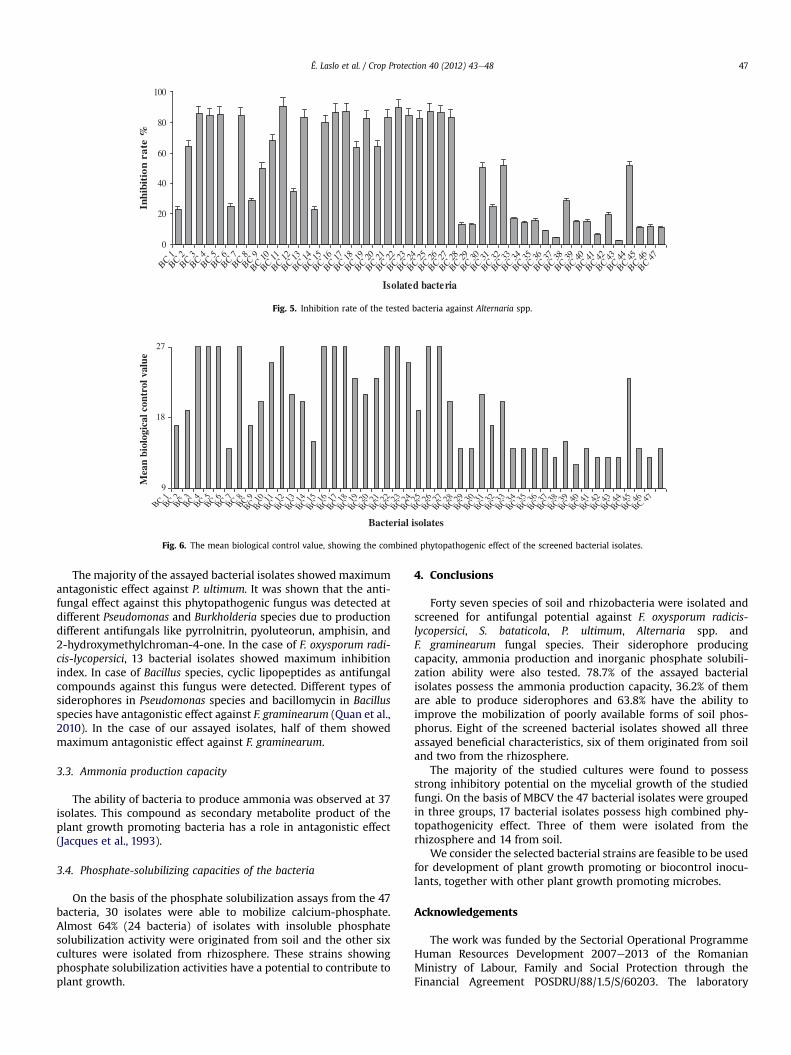
bacteria) were almost completely covered by the phytopathogens,showing no inhibition. From the assayed 47 bacterial cultures, themaximum inhibition of mycelium growth (total inhibition, 100%)was found at one culture against S. bataticola. At nine of the isolates(eight originated from soil and one originated from rhizosphere) theinhibition rate against the five studied fungi was higher than 80%. Inthe case of five isolates, the inhibition rate was higher than 80%against three fungi. For 20 bacterial cultures, we detected an inhi-bition rate higher than 80% against one fungus.
The antagonistic effect against S. bataticola (Fig. 1) wasmoderate. In the case of 18 bacteria, the inhibition indexwas higherthan 80%, but from the tested isolates of 19 bacteria, the inhibitionrate was lower than 30%. In the case of one bacterial culture (iso-lated from soil), the inhibition rate reached the maximum value.
For 20 isolates, the percentage of growth inhibition ofF. oxysporum radicis-lycopersici (Fig. 2) was higher than 50%.
In the F. graminearum antagonism assay, the percentage ofmycelial inhibition (Fig. 3) was higher than 50% for 31 bacteria(6 were isolated from the rhizosphere). In the case of 17 bacterialcultures, the antagonistic effect expressed in inhibition rate wasmore than 80%.
In the case of P. ultimum, the inhibitory effect (Fig. 4) was themost significant because the inhibition rate was higher than 75% incase of 35 bacterial strains.
The isolated and tested bacteria showed varied levels of path-ogen inhibition against Alternaria spp. (Fig. 5). For 16 bacterialstrains that originated from soil, the inhibition rate was more than80%. The highest detected antifungal activity was observed in caseof a bacterial isolate originated from rhizosphere.
0
20
40
60
80
100
BC 1BC 2
BC 3BC 4
BC 5BC 6
BC 7BC 8
BC 9
BC 10
BC 11
BC 12
BC 13
BC 14
BC 15
BC 16
BC 17
BC 18
BC 19
BC 20
BC 21
BC 22
BC 23
BC 2B
Isolate
Inhi
biti
on r
ate
%
Fig. 4. Inhibition rate of the tested b
For the selection of the isolates that posses the most broadantifungal spectrum, we determined the combined phytopatho-genicity. This effect was characterized with the mean biologicalcontrol value (MBCV), taking into consideration of the distributionof the plant pathogen fungus and the host species using the FungalDatabases of USDA. The fungal species were scored with 2 if theywere cosmopolitan and with 1 if they had specific distributionrange. Taking into account of the host specificity of the fungal taxa,the multi-host species were scored with 2, whereas with specifichost were scored with 1.
The MBCV is the sum of products of the fungal score and inhi-bition index. The inhibition index value was 1 if the IR was between0 and 33%, 2 if the IR was between 34 and 66 % and 3 if the IR wasbetween 67 and 100%.
On the basis of the MBCV (Fig. 6) with combined phytopatho-genicity the bacterial isolates can be classified into three groups.The majority of the isolated strains, 18, has week combined phy-topathogenicity effect, they are antagonistic only for some of thephytopathogenic fungus. Seventeen bacterial isolates has the broadcombined phytopathogenicity effect and 12 isolates possessmedium combined antagonistic effect. The isolates were defined ashaving low combined phytopathogenic effect if the MBCV variedamong 9e15, medium phytopathogenic effect if the value variedamong 16e21 and high if the value varied among 22e27.
From the assayed 47 bacterial isolates, three isolates, BC4, BC7and BC16, showing the maximum MBCV possess as well the side-rophore, ammonia production and phosphate mobilizationcapacity. After the identification of the bacterial isolates thesestrains could be a promising source for microbial inoculants.
4C 25
BC 26
BC 27
BC 28
BC 29
BC 30
BC 31
BC 32
BC 33
BC 34
BC 35
BC 36
BC 37
BC 38
BC 39
BC 40
BC 41
BC 42
BC 43
BC 44
BC 45
BC 46
BC 47
d bacteria
acteria against Pythium ultimum.
0
20
40
60
80
100
BC 1BC 2
BC 3BC 4
BC 5BC 6
BC 7BC 8
BC 9
BC 10
BC 11
BC 12
BC 13
BC 14
BC 15
BC 16
BC 17
BC 18
BC 19
BC 20
BC 21
BC 22
BC 23
BC 24
BC 25
BC 26
BC 27
BC 28
BC 29
BC 30
BC 31
BC 32
BC 33
BC 34
BC 35
BC 36
BC 37
BC 38
BC 39
BC 40
BC 41
BC 42
BC 43
BC 44
BC 45
BC 46
BC 47
Isolated bacteria
Inh
ibit
ion
ra
te %
Fig. 5. Inhibition rate of the tested bacteria against Alternaria spp.
9
18
27
BC 1BC 2
BC 3BC 4
BC 5BC 6
BC 7BC 8
BC 9BC 10
BC 11BC 12
BC 13BC 14
BC 15BC 16
BC 17BC 18
BC 19BC 20
BC 21BC 22
BC 23BC 24
BC 25BC 26
BC 27BC 28
BC 29BC 30
BC 31BC 32
BC 33BC 34
BC 35BC 36
BC 37BC 38
BC 39BC 40
BC 41BC 42
BC 43BC 44
BC 45BC 46
BC 47
Bacterial isolates
Mea
n bi
olog
ical
con
trol
val
ue
Fig. 6. The mean biological control value, showing the combined phytopathogenic effect of the screened bacterial isolates.
É. Laslo et al. / Crop Protection 40 (2012) 43e48 47
The majority of the assayed bacterial isolates showed maximumantagonistic effect against P. ultimum. It was shown that the anti-fungal effect against this phytopathogenic fungus was detected atdifferent Pseudomonas and Burkholderia species due to productiondifferent antifungals like pyrrolnitrin, pyoluteorun, amphisin, and2-hydroxymethylchroman-4-one. In the case of F. oxysporum radi-cis-lycopersici, 13 bacterial isolates showed maximum inhibitionindex. In case of Bacillus species, cyclic lipopeptides as antifungalcompounds against this fungus were detected. Different types ofsiderophores in Pseudomonas species and bacillomycin in Bacillusspecies have antagonistic effect against F. graminearum (Quan et al.,2010). In the case of our assayed isolates, half of them showedmaximum antagonistic effect against F. graminearum.
3.3. Ammonia production capacity
The ability of bacteria to produce ammonia was observed at 37isolates. This compound as secondary metabolite product of theplant growth promoting bacteria has a role in antagonistic effect(Jacques et al., 1993).
3.4. Phosphate-solubilizing capacities of the bacteria
On the basis of the phosphate solubilization assays from the 47bacteria, 30 isolates were able to mobilize calcium-phosphate.Almost 64% (24 bacteria) of isolates with insoluble phosphatesolubilization activity were originated from soil and the other sixcultures were isolated from rhizosphere. These strains showingphosphate solubilization activities have a potential to contribute toplant growth.
4. Conclusions
Forty seven species of soil and rhizobacteria were isolated andscreened for antifungal potential against F. oxysporum radicis-lycopersici, S. bataticola, P. ultimum, Alternaria spp. andF. graminearum fungal species. Their siderophore producingcapacity, ammonia production and inorganic phosphate solubili-zation ability were also tested. 78.7% of the assayed bacterialisolates possess the ammonia production capacity, 36.2% of themare able to produce siderophores and 63.8% have the ability toimprove the mobilization of poorly available forms of soil phos-phorus. Eight of the screened bacterial isolates showed all threeassayed beneficial characteristics, six of them originated from soiland two from the rhizosphere.
The majority of the studied cultures were found to possessstrong inhibitory potential on the mycelial growth of the studiedfungi. On the basis of MBCV the 47 bacterial isolates were groupedin three groups, 17 bacterial isolates possess high combined phy-topathogenicity effect. Three of them were isolated from therhizosphere and 14 from soil.
We consider the selected bacterial strains are feasible to be usedfor development of plant growth promoting or biocontrol inocu-lants, together with other plant growth promoting microbes.
Acknowledgements
The work was funded by the Sectorial Operational ProgrammeHuman Resources Development 2007e2013 of the RomanianMinistry of Labour, Family and Social Protection through theFinancial Agreement POSDRU/88/1.5/S/60203. The laboratory

É. Laslo et al. / Crop Protection 40 (2012) 43e4848
experiments were prepared with the financial support from the“BIOPREP eMicrobial biopreparates for increasing the productivityand crop protection” research funded by Sectorial OperationalProgramme, Increase of Economic Competitiveness Operation 2.1.1.of the Romanian Ministry of Labour, Family and Social Protection,through financial agreement POSCEE No. 469/11817.
References
Ahmad, F., Ahmad, I., Khan, M.S., 2008. Screening of free-living rhizosphericbacteria for their multiple plant growth promoting activities. Microbiol. Res.163, 173e181.
Atlas, R.M., 2010. Handbook of Microbiological Media, fourth ed. CRC Press, Taylor &Francis Group, Boca Raton.
Bhattacharyya, P.N., Jha, D.K., 2012. Plant growth-promoting rhizobacteria (PGPR):emergence in agriculture. World J. Microbiol. Biotechnol. 28, 1327e1350.
Bloemberg, G.V., Lugtenberg, B.J.J., 2001. Molecular basis of plant growth promotionand biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 4, 343e350.
Chaiharn, M., Chunhaleuchanon, S., Lumyong, S., 2009. Screening siderophoreproducing bacteria as potential biological control agent for fungal rice patho-gens in Thailand. World J. Microbiol. Biotechnol. 25, 1919e1928.
Compant, S., Duffy, B., Nowak, J., Clément, C., Barka, E.A., 2005. Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanism ofaction, and future prospects. Appl. Environ. Microbiol. 71, 4951e4959.
Djuric, S., Pavic, A., Jarak, M., Pavlovici, S., Starovici, M., Pivic, R., Josic, D., 2011.Selection of indigenous fluorescent pseudomonad isolates from maize rhizo-spheric soil in Vojvodina as possible PGPR. Rom. Biotechnol. Lett.16, 6580e6590.
Dunca, S., Nimitan, E., Ailisiei, O., Stefan, M., 2007. Microbiologie aplicat�a. Tehno-press, Iasi.
Fatima, Z., Saleemi, M., Zia, M., Sultan, T., Aslam, M., Rehman, R.U., Chaudhary, M.F.,2009. Antifungal activity of plant growth- rhizobacteria isolates againstRhizoctonia solani in wheat. Afr. J. Biotechnol. 8, 219e225.
Fernando, W.G.D., Nakkeeran, S., Zhang, Y., 2006. Biosynthesis of antibiotics byPGPR and its relation in biocontrol of plant diseases. In: Sidiqqui, Z.A. (Ed.),PGPR: Biocontrol and Biofertilization. Springer, Dordrecht, pp. 67e109.
Fungal Database of USDA, http://nt.ars-grin.gov/fungaldatabases/ (accessed10.04.12.).
Gyaneshwar, P., Kumar, G.N., Parekh, J., Poole, P.S., 2002. Role of soil microorganismsin improving P nutrition of plants. Plant Soil 245, 83e93.
György, É., Mara, Gy., Máthé, I., Laslo, É., Márialigeti, K., Albert, B., Oancea, F.,Lányi, Sz, 2010. Characterization and diversity of the nitrogen fixing microbiotafrom a specific grassland habitat in the Ciuc Mountains. Rom. Biotechnol. Lett.15, 5474e5480.
Haas, D., Défago, G., 2005. Biological control of soil-borne pathogens by fluorescentpseudomonads. Nat. Rev. Microbiol. 10, 1e13.
Hillel, D., 2005. Encyclopedia of Soils in the Environment. Academic Press, NewYork.
Höfte, M., Bakker, P.A., 2007. Competition for iron and induced systemic resistanceby siderophores of plant growth promoting. Soil Biol. 12, 121e135.
Idris, H., Labuschagne, A.N., Korsten, L., 2007. Screening rhizobacteria for biologicalcontrol of Fusarium root and crown rot of sorghum in Ethiopia. Biol. Control 40,97e106.
Jacques, P., Delfosse, P., Ongena, M., Lepoivre, P., Cornelis, P., Koedam, P.,Neirinckx, L., Thonart, P., 1993. Les mécanismes biochimiques développés parles Pseudomonas fluorescents dans la lutte biologique contre les maladies desplantes transmises par le sol. Cah. Agric. 2, 301e307.
Khan, A.A., Jilani, G., Akhtar, M.S., Naqvi, S.M.S., Rasheed, M., 2009. Phosphorussolubilizing bacteria: occurrence, mechanisms and their role in crop produc-tion. J. Agric. Biol. Sci. 1, 48e58.
Kumar, N., Arasu, R.V.T., Gunasekaran, P., 2002. Genotyping of antifungalcompounds producing plant growth-promoting rhizobacteria. Curr. Sci. 82,1463e1466.
Kumari, B., Pragash, M.G., Cletus, J., Raman, G., Sakthivel, N., 2009. Simultaneousphosphate solubilization potential and antifungal activity of new fluorescentpseudomonad strains, Pseudomona aeruginosa, P. plecoglossicida and P. mosselii.World J. Microbiol. Biotechnol. 25, 573e581.
Lugtenberg, B., Kamilova, F., 2009. Plant-growth-promoting rhizobacteria. Annu.Rev. Microbiol. 63, 541e556.
Maksimov, I.V., Abizgil’dina, R.R., Pusenkova, L.I., 2011. Plant growth promotingrhizobacteria as alternative to chemical crop protectors from pathogens(Review). Appl. Biochem. Microbiol. 47, 333e345.
Malboobi, M., Owlia, P., Behbahani, M., Sarokhani, E., Moradi, S., Yakhchali, B.,Deljou, A., Heravi, K.M., 2009. Solubilization of organic and inorganic phos-phates by three highly efficient soil bacterial isolates. World J. Microbiol. Bio-technol. 25, 1471e1477.
Oldal, B., Jevcsák, I., Kecskés, M., 2002. A sziderofortermel}o képesség szerepePseudomonas-törzsek növénypatogén-antagonista hatásának biológiai vizsgá-latában. Biokémia 26, 57e63.
Quan, C.S., Wang, X., Fan, S.D., 2010. Antifungal compounds of plant growthpromoting rhizobacteria and its action mode. In: Maheshwari, D.K. (Ed.), PlantGrowth and Health Promoting Bacteria. Springer Verlag, Berlin-Heidelberg,pp. 117e156.
Rodrigez, H., Fraga, R., 1999. Phosphate solubilizing bacteria and their role in plantgrowth promotion. Biotechnol. Adv. 17, 319e339.
Schwyn, B., Neilands, J.B., 1987. Universal chemical assay for the detection anddetermination of siderophores. Anal. Biochem. 160, 47e56.
Singh, B., Satyanarayana, T., 2011. Microbial phytases in phosphorus acquisition andplant growth promotion. Physiol. Mol. Biol. Plants 17, 93e103.
Spadaro, D., Gullino, M.L., 2005. Improving the efficacy of bio-control agents againstsoil-borne pathogens. Crop Prot. 24, 601e613.
Srivastava, R., Shalini, S., 2008. Antifungal activity of Pseudomonas fluorescensagainst different plant pathogenic fungi. Elec. J. Env. Agricult. Food Chem. 7,2789e2796.
Weyens, N., van der Lelie, D., Taghavi, S., Newman, L., Vangronsveld, J., 2009.Exploiting plantemicrobe partnerships to improve biomass production andremediation. Trends Biotechnol. 27, 591e598.
Zaidi, A., Khan, M.S., Ahemad, M., Oves, M., Wani, P.A., 2009a. Recent advances inplant growth promotion by phosphate-solubilizing microbes. In: Khan, M.S.,Zaidi, A., Musarrat, J. (Eds.), Microbial Strategies for Crop Improvement.Springer Verlag, Berlin-Heidelberg, pp. 23e51.
Zaidi, A., Khan, M.S., Ahemad, M., Oves, M., 2009b. Plant growth promotion byphosphate solubilizing bacteria. Acta Microbiol. Immunol. Hung. 56,263e284.
Zaidi, A., Ahemad, M., Oves, M., Ahmad, E., Khan, M.S., 2010. Role of phosphate-solubilizing bacteria in legume improvement. In: Khan, M.S., Zaidi, A.,Mussarat, J. (Eds.), Microbes for Legume Improvement. Springer Verlag, Berlin-Heidelberg, pp. 273e292.