sciencia acta xaveriana - st. xavier's college,...
TRANSCRIPT


SCIENCIA ACTA XAVERIANA
An International Research Journal of Basic and Applied Sciences
SCIENCIA ACTA XAVERIANA (SAX) is a referred biannual
research Journal published in March and September by
St. Xavier’s College (Autonomous), Palayamkottai - 627 002.
SAX carries research articles from Basic and Applied Sciences
(Biology, Chemistry, Computer Science, Mathematics, and Physics).
PATRONS
Rev. Dr. Danis Ponniah, S.J., Rector
Rev. Fr. Jesu Michael Das, S.J., Secretary
Dr. V. Gilburt Camillus S.J., Principal
St. Xavier’s College, Palayamkottai, India.
EDITORIAL BOARD
Editor- in-Chief
Dr. A. Lourdusamy,
Department of Mathematics, St.Xavier’s College, Palayamkottai, India.
[email protected], [email protected]
MEMBERS
Dr. Anton Ilango Department of Systems Physiology of Learning
Leibniz Institute for Neurobiology, Breeneckestraβe6,
39118, Magdeburg, Germany.
Email : [email protected]
Dr. O. N. Oigiangbe Department of Zoology, Ambrose Alli University, Nigeria.
Email : osawe_oigangbe @ hotmail.com
Dr. Antony Jeevarajan Deputy Division Chief,
Habitability and Environmental Factors Division,
Johnson Space Centre, NASA, USA.
Email : [email protected].
Dr. M. Venkatesan The University of Dublin, Trinity College,
Dublin, Ireland.
E-mail : [email protected]
Dr. Amsarani Ramamoorthy Textile Engineering, Chemistry and Science,
North Carolina State University, USA.
E-mail : [email protected]
Dr. M. Amsaveni Bhabha Atomic Research Centre
Trombay, Mumbai.
Email : [email protected]

Dr. Raja Angamuthu Department of Chemistry,
Indian Institute of Technology, Kanpur,
Uttarpradesh 208 016.
Email : [email protected]
Dr. S. Meheshwaran Indian Institute of Technology,
Powai, Mumbai.
Email : [email protected]
Dr. G. Raja Raman Department of Chemistry,
Indian Institute of Technology, Bombay.
Email : [email protected]
Dr. Satheesh Krishnamurthy Nanoscale Energy and Surface Engineering Group
The Open University, UK.
Email : [email protected]
Dr. R.Lakshminarayanan Senior Manager (Scientist E3), R & D Center
HLL Lifecare Limited (a Government of India Enterprise),
Akkulam, Sreekaryam (PO), Trivandrum - 695017, India.
Email : [email protected]
Dr. Sukumar Venkataramani Indian Institute of Science Education and Research
(IISER) - Mohali, Knowledge City, Sector 81,
SAS Nagar, Manauti, Punjabi - 140306,
Email : [email protected]
Dr. E.M. Mothi Centre for Scientific and Applied Research,
PSN College of Engineering and Technology,
Tirunelveli - 627 152, India.
Email : [email protected]
Dr. M. Sankar Cardiff Catalysis Institute
Cardiff University,
Cardiff CF103AT, UK.
E-mail : [email protected]
Dr. Enrico Marsili School of Biotechnology
Dublin City University,
Glasnevin, Dublin 9, Ireland,
E-mail : [email protected]

MEMBERS FROM ST.XAVIER’S COLLEGE
Dr. L. Henry Joseph Department of Botany
E-mail : [email protected]
Dr. A. John De Britto Department of Botany
E-mail : [email protected]
Dr. K. Sahayaraj Department of Zoology
E-mail : [email protected]
Dr. V. Jeyapaul Department of Chemistry
E-mail : [email protected]
Dr. M. Baby Maryatra Department of Chemistry
E-mail : [email protected]
Dr. S.P. Victor Department of Computer Science
E-mail : spvictor [email protected]
Dr. S. Athisayanathan Department of Mathematics
E-mail : [email protected]
Rev. Dr. A. Arockiasamy, S.J. Department of Mathematics
E-mail : [email protected]
Rev. Dr. Danis Ponniah, S.J. Department of Physics
E-mail : [email protected]
Dr. D. Prem Anand Department of Physics
E-mail : [email protected]
Subscription Rates
ANNUAL SUBSCRIPTION
(i) for individuals : Rs 300 (in India) US $50 (airmail)
(ii) for institutions: Rs 2000 (in India) US $ 100 (air mail)
Subscription orders along with a Demand Draft drawn in favour of “Sciencia Acta Xaveriana”
payable at Tirunelveli, may please be sent to the Editor-in-chief.
Disclaimer : Opinions expressed are those of their authors and not the responsibility of the
Board, the Editors or the publisher. All disputes arising, if any, are subjected to the exclusive
jurisdiction of competent court in Palayamkottai, Tamilnadu, India.
INFORMATION FOR AUTHORS
SCIENCIA ACTA XAVIERIANA - An International Research Journal of Basic and Applied
Sciences aims at publication of original research papers, comprehensive review articles, book-
reviews, dissertation abstracts etc. devoted to any area in science (Biology, Chemistry,
Computer Science, Mathematics, and Physics).

Manuscripts must be submitted electronically to the Editor-in-Chief through :
[email protected], [email protected]. Authors may submit manuscripts to any
member of the Editorial Board. No hard copy should be sent.
Submitted manuscripts must not have been published nor be under consideration for publication
elsewhere. Publication of paper in the journal automatically transfers the copyrights from the
authors to the publisher.
All manuscripts should be sent electronically in either .pdf or.doc format, type set in English
with double spacing. The file must be prepared using preferably by La-TeX processing system
or MS word. The paper should contain the following headings:
1. Title 8. Results/observations
2. Author/Authors 9. Discussion
3. Institution with address and E-mail 10. References
4. Summary with not more than 250 words 11. Appendix (if any)
5. Keywords – Maximum ten 12. Tables
6. Introduction 13. Figures - Illustrations or photos
7. Materials and Methods (B/W or colour, JPEG or
Photoshop files of all
photographs)
References : Citations within the text are given within square brackets like [5], [9, 11],
[22-25] etc. The List of references shall be given at the end of the manuscript in alphabetical
order :
[1] G. Chartrand and P. Zang, Distance in Graphs – Taking the long view, AKCE.J. Graphs,
1 No. 1 (2004), 1-13.
[2] M. Johnson, J. Vallinayagam, V.S. Manickam and S. Seeni, Multiplication of rhinacanthus
nasutus through micro propagation, Phytomorphology, 52, No.4 (2002), 331-336.
[3] M. Nielsen and I. Chuang, Quantum Computation and Quantum Information, Cambridge
University Press, 2000.
All manuscripts are subjected to anonymous review by two independent reviewers. The editorial
board will make the decision within two months after submission.
Once a paper has been accepted, send a .pdf or .doc file as well as the LATEX file to the
Editor-in-Chief. The file must be in 11 point Times or Computer modern font, 1.5 spaced,
with the text being 5.75 x 8.25 inches (14.60 x 20.96cm). On the first page leave 1 inch space
on the top for the journal’s headings.

1
Volume 5
No. 2
pp. 1-26
September 2014
Sciencia Acta XaverianaAn International Science JournalISSN. 0976-1152
Tests on Vermicomposts for their suitability to
vegetable (Abelmoschus esculentus) crops
G.Sumathi1, M.Kanchilakshmi2, Arockiam Thaddeus3, P.Chandrasekar4
, P.Porkodi5 and
J.Sureka6
1 Research Scholar2 Project fellow3 Associate professor
Jayaraj Annapackiam College for Women, Periyakulam,
Theni District - 625601.TamilNadu, India.4 Principal, Mano college, Nagampatti, Tuticorin District – 628 7185,6 Assistant professors, SACWC, Cumbum.* Corresponding authors
Email : [email protected] ; [email protected]
ABSTRACT : Vermicomposting is a low cost technology for processing or
treatment of organic wastes. It contains N, P, K and micronutrients in forms that
are readily taken up by plants. For the present study, six genus of earthworms
(Lampito mauritii (T1A) , Octochetona pattoni (T
1B), Priodocheta pellucida (T
1C),
Notoscolex palniensis (T1D), Lemnoscolex scutarius (T
1E), and Hoplochetella
stuarti (T1F) collected from different ecosystems in Theni District were tried to
raise vermicomposts. These six different (T1A-B-C-D-E-F) treatments
(vermicomposts) were applied in different dosages (100,200,300 gm) to Ladies
finger plant (Abelmoschus esculentus) to test their efficacy. Treatments along with
control (T1) plants (Ladies finger) were reared. Highest observations were recorded
in Notoscolex palniensis (T1D) on growth and yield in 100 gm of compost
application. Soil samples were collected before and after the cultivation of
vegetables (Abelmoschus esculentus). These were subjected to physico-chemical

2
analysis of N, P, K pH, EC and micronutrients. These values were compared
with the values of plant height, number of fruits, yield, and length of fruit to apply
the regression analysis; there is a significant changes in soil parameters after the
application of vermicompost compared with control.
Keywords : Notoscolex palniensis, Vermicompost, Abelmoschus esculentus,
N, P, K, and Yield.
Introduction
Earthworms are major components of the soil fauna in a wide variety of soils and climates and
are involved directly or indirectly in biodegradation, stabilization through humus formation and
various soil processes (Lavelle and Spain 2001). The disposal of wastes through the use of
earthworms also upgrades the value of original waste materials insitu and allows a final product
to be obtained free of chemical or biological pollutants (Divya 2001). Vermicomposting involves
bio-oxidation and stabilization of organic material through the interactions between earthworms
and microorganisms. Although microorganisms are mainly responsible for the biochemical
degradation of organic matter, earthworms play an important role in the process by fragmenting
and conditioning the substrate, increasing the surface area for growth of microorganisms, and
altering its biological activity (Dominguez 2004; Dominguez and Edwards 2004). Vermicompost
contains major and minor nutrients in plant- available forms, enzymes, vitamins and plant growth
hormones. It has a more beneficial impact on plants than normal compost (Gajalakshmi and
Abbasi, 2004). Earthworms have been described as a keystone species as their activity helps
regulate soil fertility, water infiltration and soil detachability in agro ecosystems (Lavelle and
Spain, 2001; Shipitalo and LeBayon, 2004). Casts deposited on the soil surface may also be
carried away in surface water runoff after rain, leading to increased on-site soil erosion and
perhaps also affecting soil properties downstream of the worm’s actual home (Shipitalo and
Le Bayon, 2004). Direct effect of plant species on soil organisms are caused by the plant’s
inputs of organic matter above and below the ground, while indirect effect of plants on biota
include shading, soil protection and uptake of water and nutrients by the root (Neher, 1999).
According to Chaudhuri et al. (2003), the quality and quantity of food material influences not
only the size but also the species composition, growth rate, fecundity of an earthworm population.
They reported differences in the rate of growth and reproduction of three vermicomposting
Tests on Vermicomposts for their suitability to vegetable (Abelmoschus esculentus) crops

3
species Perionyx excavates, Eudrilus Eugenia and Eisenia fetida in the Hevea leaf litters used
as vermiculture substrate. Earthworms play an important role in maintaining soil fertility,
ecosystem function, production and biodiversity conservation (Chaudhuri et al., 2012; Kavdir
and Ilay 2011). As global food production is already dependent on intensive agricultural
production and demands for food are likely to increase substantially, the future challenge is to
match demands for production with forms of soil management that are sensitive to maintaining
soil biodiversity(Giller et al., 1997).
In the present study, six genus of earthworm adults [Lampito mauritii (T1A), Octochetona
pattoni (T1B), Priodocheta pellucida (T
1C), Notoscolex palniensis (T
1D), Lemnoscolex scutarius
(T1E) and Hoplochetella stuarti (T
1F)] collected from various ecosystems of Theni District
were selected and inoculated in decompost of vegetable refuses T1 to identify the suitable
genus of earthworm for vermicomposting out of six. Their efficacy was tested with Ladies
finger plants (Abelmoschus esculentus) in different dosages (100, 200, 300 gm).
Materials and Methods
Experiments were conducted during 2012-2013 at a farm land of Periyakulam, Theni District,
Tamilndu, India, to study the application of vermicompost and thereby to investigate the effect
of different dosages of vermicompost on vegetables.
Preparation of vermicompost and Germination
From the 6 selected earthworms (Octochetona pattoni, Priodocheta pellucida, Notoscolex
palniensis, Lemnoscolex scutarius, Hoplochetella stuarti and Lampito mauritii) were chosen as
experimental animal. Pre-composts were prepared with leaf litters of Manilkara zapota (sapota)
and Spathodea campanulata (Nandi Flame) separately mixed with cow dung (3:1) at regular
interval of 7 days over the soil bed and were used later in the earthworms were allowed to
feed on and converted them into vermicompost. It was harvested every 45 days.
Seeds of A. esculentus (Ladies finger) was sowed in the plotted fields and germinated when
cotyledons project out through the surface of the soil. Seedlings were planted at a distance of
30 cm between two plants. Lady’s Finger (A. esculentus) plants were grown in plots and were
applied with different dosages (100g, 200g & 300gm) of vegetable vermicompost raised from
G.Sumathi, M.Kanchilakshmi, Arockiam Thaddeus, P.Chandrasekar, P.Porkodi and J.Sureka

4
leaf litter. All the necessary cultural practices and plant protection measures were followed
uniformly for all the treatments during the entire period of experimentation and were replicated
four times in a randomized complete block design.
Following treatments were organized
1. Lampito mauritii Vermicompost (T1A)
2. Octochetona pattoni Vermicompost (T1B)
3. Priodocheta pellucida Vermicompost (T1C)
4. Notoscolex palniensis Vermicompost (T1D)
5. Lemnoscolex scutarius Vermicompost (T1E)
6. Hoplochetella stuarti Vermicompost (T1F)
7. Control (Farm soil)
Plant Growth Parameters
Vegetable refuse (T1A-B-C-D-E- F) vermicompost at the dosage of 100gm, 200gm and
300gm. Time of flowering, Height of the plant (cm), length of the fruit (cm), weight of the fruit
(gm), number of fruits per plant, yield of fruit per plant (gm) were recorded on every 10th day
from the date of seed germination up to 90 days.
Physico-Chemical Analysis
Soil samples were collected before and after the harvest of vegetables. These were subjected
to physico-chemical analysis (pH, electrical conductivity, organic carbon, total Kjeldahl nitrogen)
[Jackson, 1958]. Soil and vermicompost pH were measured in deionised water (solids/solution
ratio of 1: 2.5) using pH meter. Electrical conductivity (EC) was measured in the effluent and
in a saturated solution extract of the vermicompost (Rhoades, et al., 1989). Organic carbon
was determined by the Walkley-Black method (Gaudette, et al., 1974). Methods of measuring
N, P, and K in soil by Diethen-triamin-penta-acetic acid (DTPA), extractable Zn, Fe, Cu and
Tests on Vermicomposts for their suitability to vegetable (Abelmoschus esculentus) crops

5
Mn were determined in soil samples by atomic absorption spectroscopy (Perkin Elmer, type
3041, series 3000) (Lindsay and Norvell, 1978).
Statistical analysis
Correlation and Regression analysis (SPSS computer version 17.0) was used to evaluate the
relationship between plant growth parameters and soil physicochemical parameter.
Result and Discussion
In the present study experiments were conducted to assess the requirements of vermicompost
by vegetable crops in bioremediation process to fulfil the needs of the farmers during the
transfer of technology at large scale. Among the different dosages (100, 200 & 300 gm) of
vegetable refuse vermicompost (T1A-B-C-D-E-F) applied (Abelmoschus esculentus) in the
present investigation, there has been a significant improvement in the soil quality of plots amended
with vermicompost @ 100 gm per plant (Table 1, 2, 3 and Fig 1, 2, 3). This is in concordance
with the results of the work of Edwards et al., 2000 that application of compost like
vermicompost enhances physical and chemical characteristics of soil in Bhendi (Abelmoschus
esculentus) cultivation. Vermicompost as an organic input has been applied to grow vegetables
and other crops succesfully (Ismail, S.A., 2005). Application of composts like vermicompost
could contribute to increased availablity of food (Ouedraogo, et al., 2001). The overall
productivity of vegetable crop Ladies finger during the 5 months of the trial was significantly
greater in plots treated with vegetable refuse (T1) @ 100 gm per plant.
Plant Height and Number of Branches
All treatments showed significant increase of plant height when compared to control plant
(grown in farm soil). Vegetative growths in all treatments were recorded till 90th day. The
tallest lady’s finger plant was observed in 100 gm/plant vermicompost of Notoscolex palniensis
(T1D) 60.41 ± 0.064 cm with good foliage and branching numbers 6.120 ± 0.113 (Table 1&
Fig.2). Gutierrez et al. (2007) reported that addition of vermicompost increased plant heights
and number of leaves in the yield of tomato (Lycopersicum esculentum) significantly which
confirms the results of the present study.
G.Sumathi, M.Kanchilakshmi, Arockiam Thaddeus, P.Chandrasekar, P.Porkodi and J.Sureka

6
The number of branches shows a significant (6.120 ± 0.113) variation over that of control
(2.115 ± 0.163) in all the treated plants (Table 3). According to Forde and Lorenzo (2001)
root growth and branching is favored in nutrient-rich environment and in the presence of
hormones like auxins that enable the plant to optimize the exploitation of the available resources
which are in turn transformed into photo assimilates and transported again to the root
consequently influencing plant growth and morphology.
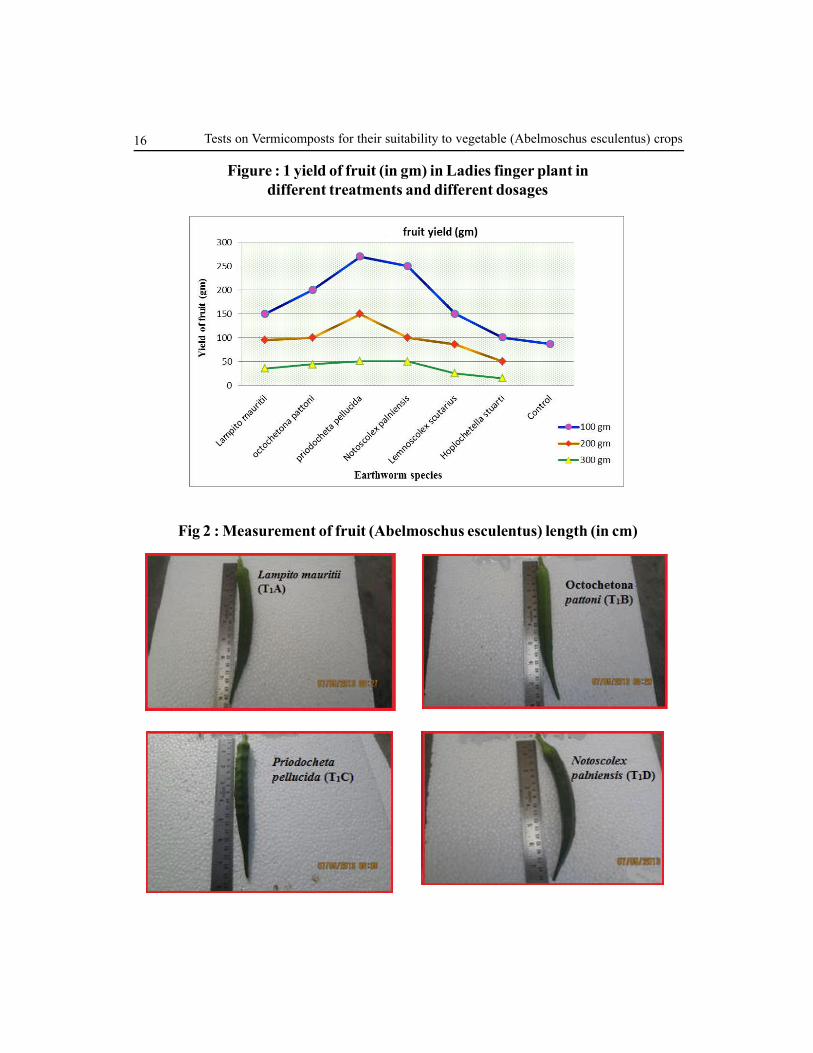
Yield
The weight and length of fruit and number of fruits / Bhendi plant is one of the most important
yield contributing traits in lady’s finger and was found maximum when treated with 100 gm
vegetable refuse (T1) vermicompost of Priodocheta pellucida 270.44 ± 0.134 kg fruit/plant
(T1C), Notoscolex palniensis 250.4 ± 0.163 (T
1D) followed by Lampito mauritii 150.6 ±
0.219 (T1A) and Octochetona pattoni 200.75 ± 0.176 (T
1B) vermicompost (Fig.1&3). Average
number of fruits/plant was Lemnoscolex scutarius 150.5 ± 0.141 (T1E) and Hoplochetella
stuarti 100.8 ± 0.134 (T1F) (Fig.1). Azarmi et al., (2008) studied on tomato (Lycopersicum
esculentum var. Super Beta) and the results of their study supported the findings of our study
that vermicompost has positive effect on growth, yield and elemental contents of plant as
compared to control. In the present study, number of fruits per plant was greatly influenced by
the treatment T1. Chand et al. (2008) experimented on tomato plants to find out the effect of
natural fertilizers on their yield and quality. They found that significantly highest yield was recorded
in the treatment receiving enriched vermicompost along with 3 sprays of liquid manure. Similar
trend was observed in development of fruit length and fruit girth. They also confirmed that
earthworms significantly improve plant growth.
Different doses of vermicompost produced different responses in A. esculentus and showed
maximum positive effect on growth parameters. The results corroborate the findings that different
doses of vermicompost caused different responses in growth parameters of L. esculentum
plant (Azarmi et al. 2008). Joshi and Vig (2010) reported significant increase in growth
parameters with application of vermicompost in L. esculentum. Vermicompost has influenced
the plant growth parameters like plant length, number of branches, number of flowers per
plant, number of fruits per plant, yield of fruit per plant, length of fruits were significantly
Tests on Vermicomposts for their suitability to vegetable (Abelmoschus esculentus) crops

7
influenced in plots receiving different doses of different genus of vermicompost. According to
Gonzalez et al. (1996) and Tien et al. (2000) tree plantations may influence earthworm
abundance by altering the physico-chemical properties of soils viz. temperature, moisture
regime, pH, organic matter content and litter inputs. Overall plant growth with vermicompost
application has been reported in different studies (Arancon et al., 2006; Zaller 2007; Bachman
and Metzger 2008; Singh et al. 2008). Arancon et al. (2004, 2006); Bachman and Metzger
(2008) also reported growth and yield improvement in different crops with vermicompost
application. The results clearly indicated that the plants receiving vermicompost had produced
more fruits/branches, branches/plant, large sized fruits with higher total yield than those of
control.
Plot soil tests for physico chemical parameters (N, P, K, pH, EC and micronutrients) was
done before and after cultivation and the results were statistically correlated (Table 4). The
vermicompost prepared by all the three earthworm species showed a substantial difference in
total N content, which could be attributed directly to the species specific feeding preference of
individual earthworm species and indirectly to mutualistic relationship between ingested
microorganisms and intestinal mucus (Suthar and singh 2008). The worms during
vermicomposting converted the insoluable P into soluable forms with the help of P-solublizing
microorganisms through phosphates present in the gut, making it more available to plants
(Suthar and singh 2008, Padmavathiamma et al., 2008). Vermicomposting proved to be an
efficient process for recovering higher K from organic waste (Suthar and singh 2008). The
present findings corroborated to those of Delgado et al., 1995, who demonstrated that higher
K concentration in the end product prepared from sewage sludge. pH was neutral being
around 7 and and increased gradually from substrate to compost to vermicompost
(Nagavallemma et al., 2006,). Fares et al., 2005, found the increased pH at the end of the
composting process, which was attributed to progressive utilization of organic acids and increase
in mineral constituents of waste. The increased EC during the period of the composting and
vermicomposting processes is in consistence with that of earlier workers (Jadia and Fulekar
2008), which was probably due to the degradation of organic matter releasing minerals such
as exchangeable Ca, Mg, K and P in the available forms, that is, in the form of cations in the
vermicompost and compost (Tognetti et al., 2005).
G.Sumathi, M.Kanchilakshmi, Arockiam Thaddeus, P.Chandrasekar, P.Porkodi and J.Sureka

8
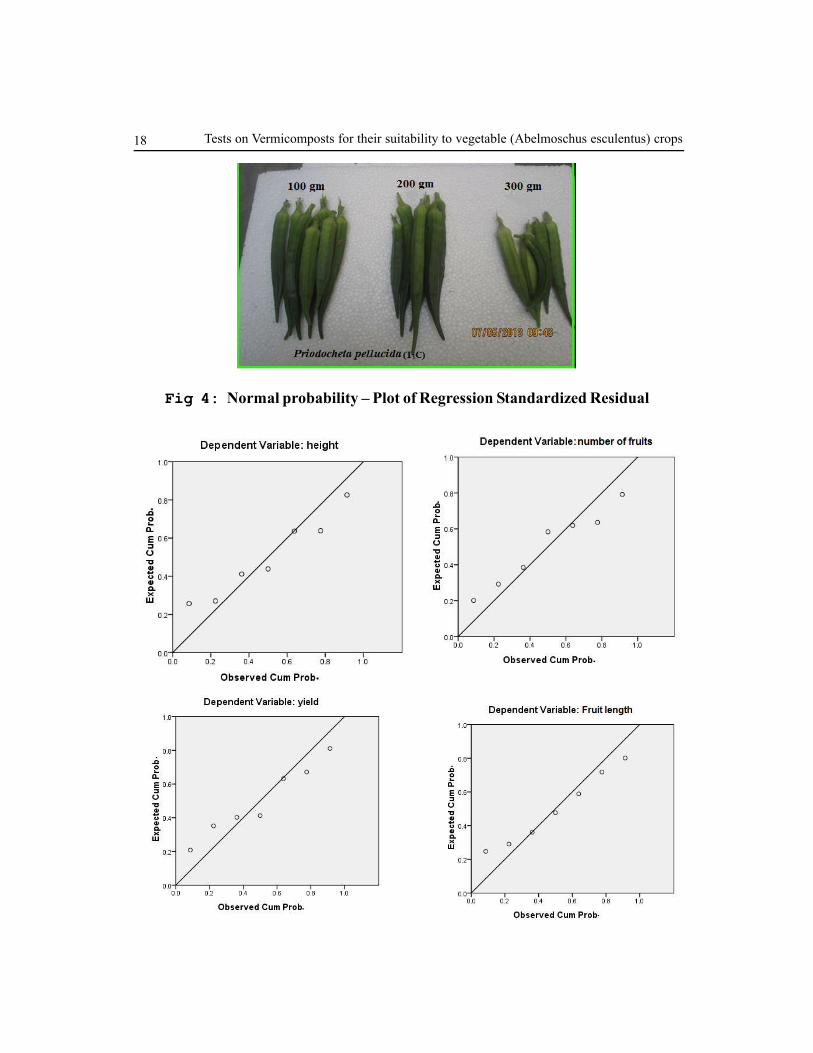
The values of plant height, number of fruits, yield, and length of fruit with the soil parameters
were statistically analysed using regression ‘B’ value is positive for all parameters, because of
the application of vermicompost, height, fruit length and yield of fruit also (positive result)
increased. The low earthworm diversity observed is consistent with other studies on invertebrate
ecology in urban areas (Paul and Meyer, 2001). According to Paoletti (1999) and Curry et al.
(2002), earthworm populations in cultivated land are generally lower than those found in
undisturbed habitats. Agricultural activities such as ploughing, several tillage operations, fertilizing
and application of chemical pesticides have dramatical effect on invertebrate animals. Any
management practices applied to soil are likely to have some (positive or negative) effects on
earthworm abundance and diversity. These effects are primarily the result of changes in soil
temperature, soil moisture and organic matter quantity or quality (Hendrix and Edwards, 2004).
The abundance of earthworms may increase due to some agricultural activities like liming,
organic fertilizing etc. (Kõlli, Lemetti, 1999). Lavelle and Spain (2001) admit that the regional
abundance of earthworms and the relative importance of the different ecological categories
are determined by large scale climatic factors (mainly temperature and rainfall) as well as by
their phylogenetic and bio geographical histories together with regional parameters such as
vegetation type and soil characteristics. According to Hole et al (2005) the evidence from
comparative studies under arable regimes indicates a general trend for higher earthworm
abundance under organic management.
Vermicompost contains more nutrients in plant available forms such as phosphates, exchangeable
calcium, soluble potassium and other macronutrients with huge quantity of beneficial
microorganisms, vitamins and hormones which have influence on the growth and yield of plants
(Theunissen et al. 2010). Kumari and Ushakumari (2002) reported that enriched vermicompost
was a superior treatment for enhancing uptake of N, P, K, Ca and Mg by cowpea.
Vermicompost have been recognized as having considerable potentials as soil amendments.
Vermicomposts are products of depredated organic matter through interactions of earthworms
and microorganisms. The process accelerates the rete of decomposition of the organic matter,
alters the physical and chemical properties of the material and lowers the C: N ratio leading to
a rapid humification process in which the unstable organic matter is fully oxidised and stabilized
(Albanell et al., 1998). The application of organic manures brings about structural improvement
Tests on Vermicomposts for their suitability to vegetable (Abelmoschus esculentus) crops

9
regeneration of soil structures and increasing the aeration within. It may cause the roots to
extend into a large volume of soil in addition to the increase of water retention in the soil profile
(Agarwal et al., 1995). The analysis of soil applied with fertilizer showed that it has all kinds of
nutrients needed for the better growth of the crop. The soil properties such as pH, EC, available
nitrogen, phosphorus, potassium, iron zinc, copper and manganese were found to vary in the
soils treated with vermicompost application (Chidambaram et al., 2013). This is also in
concordance with the present investigation that the soil nature differs entirely on the application
of vermicompost of various treatments. The results of this study indicate that incorporation of
vermicompost of plant origin into a traditional base medium of farm soil and sand enhanced
growth of bhendi plant through, at least in part, improved mineral nutrition.
References
1. Agarwal G.C, Sidhu A.S, Sekhon N.K, Sandhu K.S and Sur H.S.(1995) Soil Till. Res.,
Vol.36, pp.129-139.
2. Albanell E. Plaixats J and Cabrero T (1998) Biol.Fertil.Soils, Vol.6, pp. 266-269.
3. Arancon, N. Q., Edwards, C. A., Bierman, P., Welch, C., & Metzer, J. D. (2004).
Influence of Vermicomposts on field strawberries: effect on growth and yields. Bioresource
Technology, 93, 145-153.
4. Arancon, N. Q., Edwards, C. A., & Bierman, P. (2006). Influences of vermicomposts
on field strawberries: effects on soil microbial and chemical properties. Bioresource
Technology, 97, 831-840.
5. Azarmi, R., Giglou, M. T., & Taleshmikail, D. (2008). Influence of vermicompost on soil
chemical and physical properties in tomato (Lycopersicum esculentum) field. African
Journal of Biotechnology, 7(14), 2397-2401.
6. Bachman, G. R., and Metzger, J. D. (2008). Growth of bedding plants in commercial
potting substrate amended with vermicompost. Bioresource Technology, 99, 3155-3161.
7. Chand, S., Ali, T., Wani, J.A. and Masih, M.R., (2008). ‘Integrated Nutrient Resource
Management (INRM) for Sustainable Agriculture’, Proceedings of International
G.Sumathi, M.Kanchilakshmi, Arockiam Thaddeus, P.Chandrasekar, P.Porkodi and J.Sureka

10
Symposium on Natural Resource Management in Agriculture. December19-20, 2008,
Jaipur, Rajasthan, India, pp. 51.
8. Chaudhuri, P.S., T.K.Pal, G.Battachrjee and S.K. Dey: (2003). Rubber leaf litters (Hevea
brasilensis, var RRIM 600) as vermiculture substrate for epigeic earthworms. Perionyx
excavates, Eudrilus eugeniae and Eisenia fetida. Pedobiologia, 47, 796-800.
9. Chaudhuri, P.S., T.K. Pal, S.Nath and S.K.Dey: (2012). Effects of five earthworm
species on some physico-chemical properties of soil. J.Environ.Biol., 33, 713-716.
10. Chidambaram AL.A, Mathivanan S, Kalaikandhan R and Sundramoorthy P. (2013)
Asian Journal of Plant Science and Research, Vol. 3(2), pp.15-22.
11. Curry, J. P., Byrne, D., Schmidt, O. (2002) Intensive cultivation can drastically reduce
earthworm populations in arable land. – Eur. J. Soil Biol. 38, p. 127-130.
12. Delgado M. Bigeriego, I. Walter and Calbo R. (1995) “Use of california red worm in
sewage sludge transformation”, Turrialba, Vol.45, pp.33-41.
13. Divya UK (2001). Relevance of vermiculture in sustainable agriculture. Agriculture
General –World July 2001, 9-11.
14. Dominguez J (2004). State of the art and new prospectives on vermicomposting research.
In: Edwards CA (Ed) Earthworm Ecology (2nd Edn), CRC Press LLC, Boca Raton, FI
USA, pp 401-424.
15. Dominguez J, Edwards CA (2004). Vermicomposting organic wastes: a review. In: Hanna
SHS, Mikhail WZA (Eds) Soil Zoology for Sustainable Development in the 21st Century,
Cairo, pp 369-395.
16. Edwards, L., J.R. Burney, G. Richter and A.H. MacRae, (2000). Evaluation of compost
and straw mulching on soil-loss characteristics in erosion plots of potatoes in Prince
Edward Island, Canada. Ecosystem and Environment, 81: 217-222.
17. Fares F., Albalkhi A., Dec.J, Bruns. M.A and Bollag.J.M (2005) “Physicochemical
characteristics of animal and municipal wastes decomposed in arid soils,” Journal of
Environmental quality, vol.34, No.4, pp.1392-1403.
Tests on Vermicomposts for their suitability to vegetable (Abelmoschus esculentus) crops

11
18. Forde, B., and Lorenzo, H. (2001). The nutritional control of root development. Plant
Soil, 232, 51-68.
19. Gajalakshmi. S and Abbasi S.A (2004) Earthworms and vermicomposting, Indian J
Biotechnol, 3 486-494.
20. Gaudette, H.E., W.R. Flight, L. Toner and D.W. Folger, (1974). An inexpensive titration
method for the determination of organic carbon in recent sediments. J. Sediment Petrol.,
44: 249-253.
21. Giller, K.E, M.H.Beare, P. Lavelle, A.M.N. Izac and M.J. Swift: (1997). Agricultural
intensification, soil biodiversity and agroecosystem function. Applied soil Ecology, 6,
3-16.
22. Gonzalez, G., X.M. Zou and S. Borges: (1996). Earthworm abundance and species
composition in abandoned tropical crop lands: Comparisons of tree plantations and
secondary forests. Pedobioligia, 40, 385 -391.
23. Gutiérrez, M., Federico, A., Santiago-B., Jorge, M. M., Joaquin A., Carlos, N., Camerino,
A., Miguel, O. L., Maria, A., Rincón, R. and Dendooven, L. (2007) ‘Vermicompost as
a soil supplement to improve growth, yield and fruit quality of tomato (Lycopersicum
esculentum)’, Bioresource Technology, Vol.98, No .15 ,pp. 2781-2786.
24. Hendrix, P. F. and Edwards, C. A. (2004) Earthworms in Agro ecosystems: research
approaches. – Earthworm Ecology. Ed. C. A. Edwards, 2nd edition, CRC Press, Boca
Raton, London, New York, p. 287-295.
25. Hole, D.G., Perkins, A.J., Wilson, J.D., Alexander, I.H., Grice, P.V., Evans, A.D.(2005)
Does organic farming benefit biodiversity? Biological Conservation 122 113-130.
26. Ismail, S.A., (2005). The Earthworm Book. Other India Press, apusa, Goa, pp: 101.
27. Jackson, M.L., 1958. Soil Chemical Analysis, Prentice Hall Inc, Englewood Cliffs, New
Jersey, USA, pp: 498.
G.Sumathi, M.Kanchilakshmi, Arockiam Thaddeus, P.Chandrasekar, P.Porkodi and J.Sureka

12
28. Jadia C.D and Fulekar M.H, (2008) “Vermicomposting of vegetable wastes: a biophysico
chemical process based on hydro-operating bioreacter,” African journal of Biotechnology,
vol.7, pp.3723-3730.
29. Joshi, R., & Vig, A. P. (2010). Effect of vermicompost on growth, yield and quality of
tomato (Lycopersicum esculentum L). African Journal of Basic and Applied Science 2,
(3-4), 117-123.
30. Kavdir, Y and R.Ilay: (2011). Earthworms and soil structure. In: Biology of earthworms
(Ed. : A.Karaca). Spiringer, London, p.39.
31. Kõlli, R., Lemetti, I. (1999) Eesti muldade lühiiseloomustus. I. Normaalsed
mineraalmullad. – Eesti Põllumajandusülikool, Tartu, 122 lk.
32. Kumari, M.S. and K. Ushakumari, (2002). Effect of vermicompost enriched with rock
phosphate on the yield and uptake of nutrients in cowpea (Vigna unguinculata L. WALP).
J. Trop. Agric.,40: 27-30.
33. Lavelle, P. and Spain, A.V. (2001). Soil Ecology. Dordrecht, Netherlands: Kluwer
Academic Publishers.
34. Lindsay, W.L. and W.A. Norvell, (1978). Development of DTPA soil test for zinc, iron,
manganese and copper. Soil Sci. Soc. Am. J., 42: 421-428.
35. Nagavallemma K.P, Wani S.P and Stephane L (2006) “Vermicomposting: recycling
wastes into valuable organic fertilizer, ” Journal of SAT Agricultural Research, Vol.2,
No.1, pp.1-17.
36. Neher, D.A.: (1999). Soil community composition and ecosystem process: comparing
agricultural ecosystems with natural Ecosystems. Agroforestry systems, 45, 159.
37. Ouedraogo, E., A. Mando and N.P. Zombré, (2001). Use of compost to improve soil
properties and crop productivity under low input agricultural system in West Africa.
Agric. Ecosys. Environ., 84: 259-266.
38. Paoletti, M. G. (1999) The role of earthworms for assessment of sustainability and as
bioindicators. – Agriculture. Ecosystems, Environment. 74, p. 37-155.
Tests on Vermicomposts for their suitability to vegetable (Abelmoschus esculentus) crops

13
39. Padmavathiamma P.K, Li L.Y and Kumari U.R (2008) “An experimental study of
vermi-biowaste composting for agricultural soil improvement,” Bioresource technology,
Vol.99, No.6, pp.1672-1681.
40. Paul, M.J., Meyer, J.L. (2001). Streams in the urban landscape. Annu. Rev. Ecol.Syst.
32: 333-365.
41. Rhoades, J.D., N.A. Mantghi, P.J. Shause and W. Alves, (1989). Estimating soil salinity
from saturate soil-paste electrical conductivity. Soil Sci. Soc. Am. J., 53: 428-433.
42. Shipitalo, M. J. and Le Bayon, R.C. (2004). “Quantifying the effects of earthworms on
soil aggregation and porosity”. In Edwards, C.A. (ed.). Earthworm Ecology, second
edition. Boca raton, London, New York, Washington: CRC Press.
43. Singh, R., Sharma, R.R., Kumar, S., Gupta, R. K., & Patil, R. T. (2008). Vermicompost
substitution influences growth, physiological disorders, fruit yield and quality of strawberry
(Fragaria xananassa Duch). Bioresource Technology, 99(17), 8507-8511.
44. Suthar. S and Singh. S (2008) “Vermicomposting of domestic waste by using two epigeic
earthworms (Perionyx excavatus and Perionyx sansibaricus),” International Journal of
Environment Science and Technology, Vol.5, No.1, pp.99-106.
45. Theunissen, P. A., Ndakidemi, & Laubscher, C. P. (2010). Potential of vermicompost
produced from plant waste on the growth and nutrient status in vegetable production.
International Journal of Physical Science, 5(13), 1964-1973.
46. Tien, G, J.A. Olimah, G.O. Adeoye and B.T. Kang: (2000). Regeneration of earthworm
population in a degraded soil by natural and planted fallows under humid tropical condition.
Soil Sci. Sco. Am. J., 64, 222-228.
47. Tognetii C, Laos F, Mazzarino M.J and Hernandez M.T. (2005) “composting Vs
vermicomposting:a comparison of end product quality,” Compost Science and Utilization,
Vol.13, no.1,pp.6-13.
48. Zaller, J. G. (2007). Vermicompost as a substitute for peat in potting media: Effects on
germination, biomass allocation, yields and fruit quality of three tomato varieties. Science
of Horticulture, 112, 191-199.
G.Sumathi, M.Kanchilakshmi, Arockiam Thaddeus, P.Chandrasekar, P.Porkodi and J.Sureka
14
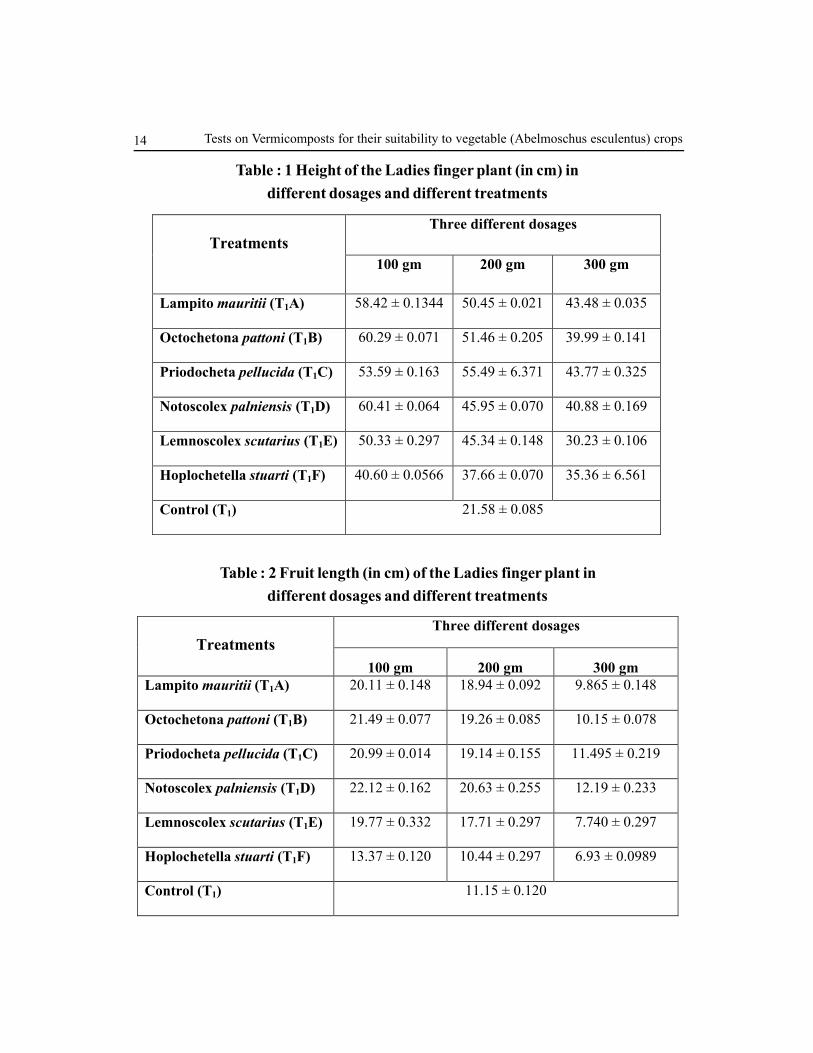
Table : 1 Height of the Ladies finger plant (in cm) in
different dosages and different treatments
Three different dosages
Treatments
100 gm 200 gm 300 gm
Lampito mauritii (T1A) 58.42 ± 0.1344 50.45 ± 0.021 43.48 ± 0.035
Octochetona pattoni (T1B) 60.29 ± 0.071 51.46 ± 0.205 39.99 ± 0.141
Priodocheta pellucida (T1C) 53.59 ± 0.163 55.49 ± 6.371 43.77 ± 0.325
Notoscolex palniensis (T1D) 60.41 ± 0.064 45.95 ± 0.070 40.88 ± 0.169
Lemnoscolex scutarius (T1E) 50.33 ± 0.297 45.34 ± 0.148 30.23 ± 0.106
Hoplochetella stuarti (T1F) 40.60 ± 0.0566 37.66 ± 0.070 35.36 ± 6.561
Control (T1) 21.58 ± 0.085
Table : 2 Fruit length (in cm) of the Ladies finger plant in
different dosages and different treatments
Three different dosages
Treatments
100 gm 200 gm 300 gm
Lampito mauritii (T1A) 20.11 ± 0.148 18.94 ± 0.092 9.865 ± 0.148
Octochetona pattoni (T1B) 21.49 ± 0.077 19.26 ± 0.085 10.15 ± 0.078
Priodocheta pellucida (T1C) 20.99 ± 0.014 19.14 ± 0.155 11.495 ± 0.219
Notoscolex palniensis (T1D) 22.12 ± 0.162 20.63 ± 0.255 12.19 ± 0.233
Lemnoscolex scutarius (T1E) 19.77 ± 0.332 17.71 ± 0.297 7.740 ± 0.297
Hoplochetella stuarti (T1F) 13.37 ± 0.120 10.44 ± 0.297 6.93 ± 0.0989
Control (T1) 11.15 ± 0.120
Tests on Vermicomposts for their suitability to vegetable (Abelmoschus esculentus) crops
15
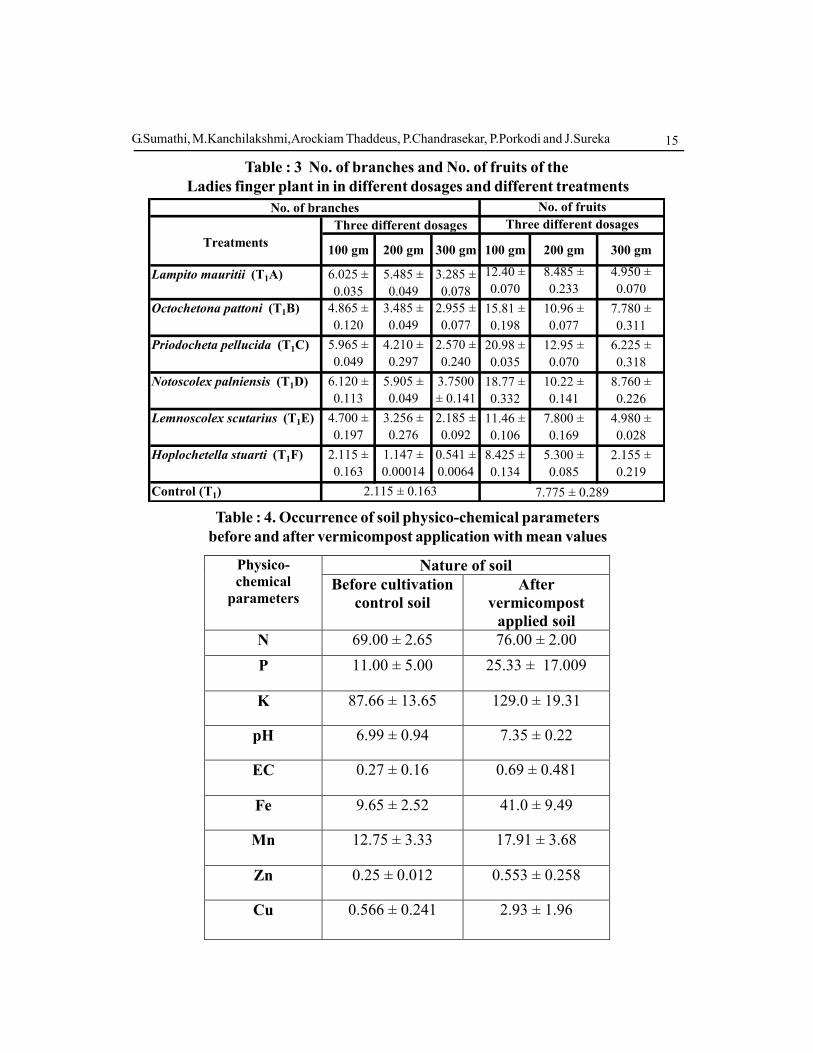
Table : 3 No. of branches and No. of fruits of the
Ladies finger plant in in different dosages and different treatments
Treatments100 gm 200 gm 300 gm 100 gm 200 gm 300 gm
Lampito mauritii (T1A) 6.025 ±
0.035
5.485 ±
0.049
3.285 ±
0.078
12.40 ±
0.070
8.485 ±
0.233
4.950 ±
0.070
Octochetona pattoni (T1B) 4.865 ±
0.120
3.485 ±
0.049
2.955 ±
0.077
15.81 ±
0.198
10.96 ±
0.077
7.780 ±
0.311
Priodocheta pellucida (T1C) 5.965 ±
0.049
4.210 ±
0.297
2.570 ±
0.240
20.98 ±
0.035
12.95 ±
0.070
6.225 ±
0.318
Notoscolex palniensis (T1D) 6.120 ±
0.113
5.905 ±
0.049
3.7500
± 0.141
18.77 ±
0.332
10.22 ±
0.141
8.760 ±
0.226
Lemnoscolex scutarius (T1E) 4.700 ±
0.197
3.256 ±
0.276
2.185 ±
0.092
11.46 ±
0.106
7.800 ±
0.169
4.980 ±
0.028
Hoplochetella stuarti (T1F) 2.115 ±
0.163
1.147 ±
0.00014
0.541 ±
0.0064
8.425 ±
0.134
5.300 ±
0.085
2.155 ±
0.219
Control (T1) 2.115 ± 0.163 7.775 ± 0.289
No. of branches No. of fruits
Three different dosages Three different dosages
Table : 4. Occurrence of soil physico-chemical parameters
before and after vermicompost application with mean values
Nature of soil Physico-
chemical
parameters Before cultivation
control soil
After
vermicompost
applied soil
N 69.00 ± 2.65 76.00 ± 2.00
P 11.00 ± 5.00 25.33 ± 17.009
K 87.66 ± 13.65 129.0 ± 19.31
pH 6.99 ± 0.94 7.35 ± 0.22
EC 0.27 ± 0.16 0.69 ± 0.481
Fe 9.65 ± 2.52 41.0 ± 9.49
Mn 12.75 ± 3.33 17.91 ± 3.68
Zn 0.25 ± 0.012 0.553 ± 0.258
Cu 0.566 ± 0.241 2.93 ± 1.96
G.Sumathi, M.Kanchilakshmi, Arockiam Thaddeus, P.Chandrasekar, P.Porkodi and J.Sureka
16
Figure : 1 yield of fruit (in gm) in Ladies finger plant in
different treatments and different dosages
Fig 2 : Measurement of fruit (Abelmoschus esculentus) length (in cm)
Tests on Vermicomposts for their suitability to vegetable (Abelmoschus esculentus) crops

17
Fig 3 : A.esculantus plant fruit yield on different treatments Lampito mauritii (T1A),
Octochaetona pattoni (T1B) and Priodocheta pellucida (T
1C) in 3 different dosages
G.Sumathi, M.Kanchilakshmi, Arockiam Thaddeus, P.Chandrasekar, P.Porkodi and J.Sureka
18
Fig 4: Normal probability – Plot of Regression Standardized Residual
Tests on Vermicomposts for their suitability to vegetable (Abelmoschus esculentus) crops

19
Volume 5
No. 2
pp. 19-30
September 2014
Sciencia Acta Xaveriana
An International Science Journal
ISSN. 0976-1152
Maximum Independent Set Cover
Pebbling Number of an m-ary Tree
A. Lourdusamy1, C. Muthulakhmi @ Sasikala2 and T. Mathivanan3
1, 3 Department of Mathematics
St. Xavier’s College,
Palayamkottai - 627 002.
2 Assistant Professor in mathematics,
Sri Paramakalyani College,
Alwarkurichi, India.
Abstract : A pebbling move is defined by removing two pebbles from
some vertex and placing one pebble on an adjacent vertex. A graph is said
to be cover pebbled if every vertex has a pebble on it after a series of
pebbling moves. The maximum independent set cover pebbling number of
a graph G is the minimum number, , of pebbles required so that any
initial configuration of pebbles can be transformed by a sequence of
pebbling moves so that after the pebbling moves the set of vertices that
contains pebbles form a maximum independent set S of G. In this paper, we
determine the maximum independent set cover pebbling number of an
m-ary tree.
Key words: Graph pebbling, cover pebbling, maximum independent set
cover pebbling, m-ary tree.

20
1. Introduction
Given a graph G, distribute k pebbles on its vertices in some configuration, call it as
C. Assume that G is connected in all cases. A pebbling move is defined by removing
two pebbles from some vertex and placing one pebble on an adjacent vertex. [1] The
pebbling number is the minimum number of pebbles that are sufficient, so that
for any initial configuration of pebbles, it is possible to move a pebble to any
root vertex v in G. [2] The cover pebbling number is defined as the minimum
number of pebbles needed to place a pebble on every vertex of the graph using a
sequence of pebbling moves, regardless of the initial configuration. A set S of
vertices in a graph G is said to be an independent set (or an internally stable set) if
no two vertices in the set S are adjacent. An independent set S is maximum if G has
no independent set with .
We introduce the concept maximum independent set cover pebbling number in [4].
The maximum independent set cover pebbling number, , of a graph , to be the
minimum number of pebbles that are placed on such that after a sequence of
pebbling moves, the set of vertices with pebbles forms a maximum independent set
S of G, regardless of their initial configuration. In this paper, we determine the
maximum independent set cover pebbling number for an m-ary tree.
Notation: denotes the number of pebbles placed at the vertex . Also
denotes the number of pebbles on the graph .
2. Maximum independent set cover pebbling number of an m-ary tree
Definition 2.1. A complete -ary tree, denoted by , is a tree of height with
vertices at distances i from the root. Each vertex of has children except
Maximum Independent Set Cover Pebbling Number of an m-ary Tree

21
for the set of vertices that are at distance away from the root, none of which
have children. The root is denoted by .
Or Simply a complete -ary tree with height , denoted by , is an -ary tree
satisfying that has children for each vertex not in the th level.
Theorem 2.2. (i) ρ(M0) = 1 (obvious).
(ii) ρ(M1) = 4m-3 (m ≥ 3) and if m = 2 then ρ(M1) = 6. Since, for m ≥ 3, M1 ≡
K1,m[4] and for m = 2, M1 ≡ P3, the path of length two[5].
(iii) ρ(M2) = 16m2-12m+1.
Proof of (iii). Note that M2 contains m-M1’s as subtrees which are all connected to
the root R2 of M2. Let R11, R12, … , R1m be the root of the m-M1’s (say M11, M12, …
, M1m). In general, Mn contains m-Mn-1’s as subtrees which are all connected to the
root Rn of Mn. Let R(n-1)1, R(n-1)2, … , R(n-1)m be the root of the m-M(n-1)’s. Choose the
rightmost vertex of this subtree, label it by v. Put 16m2-12m pebbles on this vertex.
Then we cannot cover the maximum independent set of M2. Thus ρ(M2) ≥ 16m2-
12m+1.
Now consider the distribution of 16m2-12m+1 pebbles on the vertices of M2.
According to the distribution of these amounts of pebbles, we find the following
cases:
Case 1 : f(M1i) ≥ 4m-3, where 1 ≤ i ≤ m.
Clearly we are done if f(R2) ≥ 1. So assume that, f(R2) = 0. This implies that
2
1
1
( ) 16 12 1.m
i
i
f M m m=
= − +∑ Any one of the m2
paths (of length two) leading
from the root R2 to the bottom of M2 must contain at least four pebbles and hence
we are done, since any one the subtree contains at least
A. Lourdusamy1, C. Muthulakhmi @ Sasikala2 and T. Mathivanan3

22
we are done, since any one the subtree contains at least 216 12 1
16 12 1m m
mm
− +≥ − +
pebbles.
Case 2: f(M1i) ≤ 4m-4, for all i (1 ≤ i ≤ m)
This implies that f(R2) ≥ 16m2-12m+1-m(4m-4)=12m
2-8m+1. We need 2m(4m-3)+1
pebbles at R2. But f(R2)- 2m(4m-3)- 1>0.
Case 3 : f(M1i) ≥ 4m-3 for some i (1 ≤ i ≤ m).
Let t ≥1 subtrees of M2 contains at least 4m-3 pebbles. Note that, for every subtree
(except one subtree that contains 4m-3 or more pebbles, we have 16m pebbles to
cover its maximum independent set.
Let f('
1 jM ) = aj where aj ≤ 4m-4. Thus, to cover the maximum independent set of
the subtree '
1 jM , we have another 16m-aj pebbles somewhere on the graph. So, we
can send 16
44 4
j jm a a
m−
≥ −
pebbles to the root R2 and then we move
28
ja
m − pebbles to the root '
1 jR of '
1 jM . Thus '
1 jM contains aj+2m-8
ja
=
2m+7
8ja . But these numbers of pebbles are enough to cover the maximum
independent set of '
1 jM , or the value of 2m+7
4 38
ja m≥ − , and hence we are
done. So using (m-t)(16m-aj)- 1
t
j
i
a=
∑ pebbles, we cover the maximum independent
set of the (m-t) subtrees that contains aj pebbles. So we have at least (t-1)16m+4m+1
Maximum Independent Set Cover Pebbling Number of an m-ary Tree

23
pebbles on the t-subtrees plus R2 that are all contains 4m-3 or more pebbles. If f(R2)
≥ 1 then we are done. Otherwise we can always move a pebble to R2 using at most
four pebbles from the remaining pebbles on the t-subtrees.
(iv) ρ(M3) = 64m3-
48m
2+4m-15 (m ≥ 3).
Proof of (iv). Clearly, M3 contains m-M2’s as subtrees which are all connected to
the root R3 of M3. Consider the rightmost bottom vertex, say v, of M3 and put 64m3-
48m2+4m-16 pebbles on the vertex v. Then we cannot cover the maximum
independent set of M3. Thus ρ(M3) ≥ 64m3- 48m
2+4m-15.
Now consider the distribution of 64m3-
48m
2+4m-15 pebbles on the vertices of M3.
According to the distribution of these amounts of pebbles, we find the following
cases:
Case 1 : f(M2i) ≥ ρ(M2) where 1 ≤ i ≤ m.
Clearly we are done if f(R2) =0, or 2 or f(R2) ≥ 4. So assume that f(R2) = 1 or 3. This
implies that, 3 2
2
1
( ) 64m - 48m +4m-18 .m
i
i
f M=
≥∑ pebbles. So, any one of the
path (of length three) leading from the root R3 to the bottom row of M3 must contain
at least eight pebbles. Thus we move a pebble to R3 and hence we are done.
Case 2 : f(M2i) < ρ(M2) where 1 ≤ i ≤ m.
We need 2m ρ(M2)+5 pebbles on the root vertex R3 of M3. We have ρ(M3)-m
ρ(M2)+m pebbles on the root vertex R3. But, ρ(M3)-m ρ(M2)+m-(2m ρ(M2)+5) ≥ 0.
Since, ρ(M3) = 64m3-
48m
2+4m-15, ρ(M2) = 16m
2-12m+1 and m ≥ 3.
Case 3 : f(M2i) ≥ ρ(M2) for some i (1 ≤ i ≤ m).
A. Lourdusamy1, C. Muthulakhmi @ Sasikala2 and T. Mathivanan3

24
Let t ≥ 1 subtrees contains ρ(M2) or more pebbles. Label those subtrees by M2i (1 ≤ i
≤ t) and label the other subtrees by '
2 jM (1 ≤ i ≤ m-t). Also, let f('
2 jM ) = aj where
aj < ρ(M2). Note that, we have usually (64m2+16)(m-1) pebbles each to cover the
maximum independent set of M2i’s and '
2 jM ’s, except one subtree M2k (1 ≤ k ≤ t)
that contains ρ(M2) or more pebbles.
Since aj < ρ(M2), we have another 64m2+16-aj pebbles that are in somewhere of the
graph M3 to cover the maximum independent set of '
2 jM . So we can send
2
j 264m +16-a
8 28 8
jam
≥ + −
pebbles to the root R3 and then we move
4m2+1- pebbles to the root
'
2 jR of '
2 jM . Thus, '
2 jM contains 2 15
4 116
jm a+ +
pebbles. But these number of pebbles are at least ρ(M2) or it is enough to cover the
maximum independent set of '
2 jM using the pebbles at
'
2 jR plus aj pebbles. Thus
the t-subtrees M2i plus R3 contains (64m2+16)(t-1)+ 16m
2-12m+1 or more pebbles.
We know that f(M2i) ≥ ρ(M2) where 1 ≤ i ≤ t. Let f(R3) =1 or 3 (Otherwise, we are
done). We can move a pebble to R3, using at most eight pebbles from the subtree
that contains 16m2-12m+9 pebbles or more. And hence we are done.
(v) .
Proof of (v): Consider the rightmost bottom vertex, say v, of M4, and put
pebbles. Then we cannot cover the maximum
independent set of M4. Thus, .
Maximum Independent Set Cover Pebbling Number of an m-ary Tree

25
Now consider the distribution of pebbles on the
vertices of M4. According to the distribution of these amounts of pebbles, we find
the following cases:
Case 1: for all i (1 ≤ i ≤ m).
Clearly we are done if . So assume that . This implies that
. So any one of the m4
paths (of length four) leading from the root R4 to the bottom row of M4 contains at
least sixteen ‘extra’ pebbles. Thus we can move a pebble to R4 and hence we are
done.
Case 2: for all i (1 ≤ i ≤ m).
We need pebbles on the root vertex R4 of M4. We have
on the root vertex R4. Since,
,
and we get and hence we are done.
Case 3: for some i.
Similar to Case (iii) of previous theorems; using the hints, from that
pebbles, we can send to the root R4 of M4.
Theorem 2.3: For a complete m-ary tree Mn (n ≥ 3), the maximum independent set
cover pebbling number is given by,
A. Lourdusamy1, C. Muthulakhmi @ Sasikala2 and T. Mathivanan3

26
where , and
.
Proof. Consider the rightmost vertex of Mn, say v, and put pebbles on the
vertex v. Then we cannot cover a maximum independent set of Mn. Thus the lower
bound follows.
We prove the upper bound of by induction on n. For n=3 and n=4, this
theorem is true by previous theorem (iv) and (v). So assume the result is true for the
complete m-ary tree Mn-1 (n ≥ 5).
Consider the distribution of pebbles on the vertices of Mn. According to the
distribution of these amounts of pebbles, we find the following cases:
Case (1): for all i (1 ≤ i ≤ m).
We need, pebbles on the root Rn, to cover the maximum
independent set of Mn. We have to prove that
. It is enough to prove that,
(for m ≥ 3).
------ (1)
From the 1st term, by considering k=0 we get,
------ (2)
Maximum Independent Set Cover Pebbling Number of an m-ary Tree
27
------ (3)
------ (4)
and ------- (5)
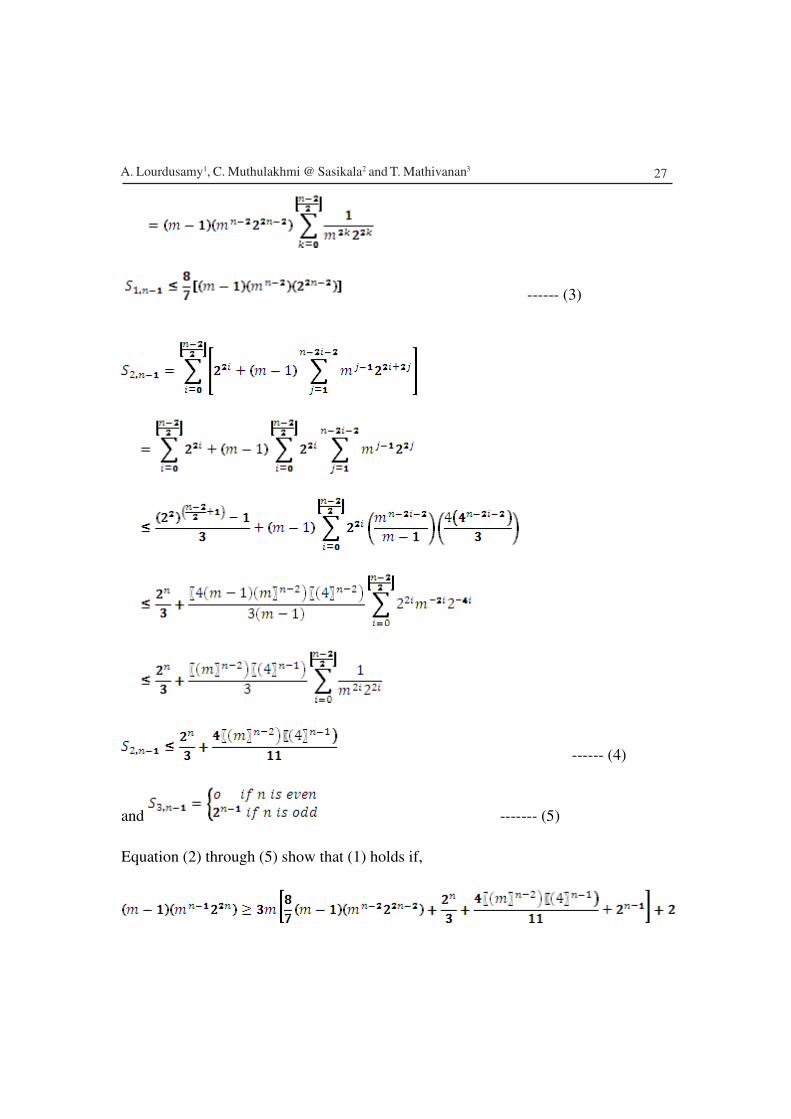
Equation (2) through (5) show that (1) holds if,
A. Lourdusamy1, C. Muthulakhmi @ Sasikala2 and T. Mathivanan3

28
which holds for and . Also, for and
.
Case (2): for all i (1 ≤ i ≤ m).
Subcase 2.1: n is odd.
If then clearly we are done. So assume that
. Then, or more pebbles on the . We know
that, and . We have,
extra pebbles on the vertices of . Thus at least one
subtree contains extra pebbles, so at least
one of the paths leading to the root from the bottom of the subtree has at
least 2n pebbles and hence we are done.
Subcase 2.2: n is even.
If then we are done. So assume that . Like, Subcase 2.1, at
least one of the paths has 2n or more pebbles and hence we are done.
Case (3): for some i.
Maximum Independent Set Cover Pebbling Number of an m-ary Tree

29
Let subtrees contain or more pebbles. Label those subtrees by
and label the other subtrees by . Also let
where . Clearly we can supply at
least one pebble to the root of for every 2n extra pebbles on
. Also, having one additional pebble in is
equivalent to have at least one pebble on the root vertex of .
Note that, we have usually used P pebbles each to cover the maximum independent
set of and , except one subtree, say
, that contains or more pebbles. Since , we have
extra pebbles, that are in somewhere of the graph , to cover the maximum
independent set of . So we can send pebbles to
the root vertex of . Thus contains
pebbles. But these amounts of pebbles are at least
or it is enough to cover the maximum independent set of , using the
pebbles at plus pebbles. Thus the t-subtrees plus
contains or more pebbles. We know that
where .
Subcase 3.1: n is odd.
Let (otherwise we are done easily). Then we can move a pebble to
A. Lourdusamy1, C. Muthulakhmi @ Sasikala2 and T. Mathivanan3

30
, using at most pebbles from the subtree that contains at least
pebbles and hence we are done [since ].
Subcase 3.2 : n is even.
Let (otherwise we are done). Like the Subcase 3.1, we can move a pebble
to , using at most pebbles (from the subtree that contains
pebbles or more).
References :
[1] F.R.K. Chung, Pebbling in hypercubes, SIAM J. Disc. Math 2(1989), 467-472.
[2] B.Crull, T.Cundiff, P.Feltman, G.H. Hurlbert, L.Pudwell, Z.Szaniszlo, Z.Tuza, The
cover pebbling number of Graphs, (2004).
[3] G.Hurlbert, A survey of Graph Pebbling, Congressus Numerantium 139 (1999)
41-64.
[4] A. Lourdusamy, C. Muthulakshmi @ Sasikala and T. Mathivanan, Maximum
independent set cover pebbling number of a Binary Tree, Sciencia Acta Xaveriana,
Vol. 3(2) (2012) , 9-20.
[5] A. Lourdusamy, C. Muthulakshmi @ Sasikala, Maximum independent set cover
pebbling number of a Star, International Journal of Mathematical Archive- 3(2),
2012, 616-618.
[6] A. Lourdusamy, C. Muthulakshmi @ Sasikala and T. Mathivanan, Maximum
independent set cover pebbling number of complete graphs and paths, submitted for
publication.
Maximum Independent Set Cover Pebbling Number of an m-ary Tree

31
Volume 5
No. 2
pp. 31-38
September 2014
Sciencia Acta Xaveriana
An International Science Journal
ISSN. 0976-1152
Generalized t-Pebbling Numbers of
Wheel and Complete r-partite graph
A. Lourdusamy
Department of Mathematics
St. Xavier’s College (Autonomous)
Palayamkottai - 627 002, India
C. Muthulakshmi@Sasikala
Department of Mathematics
Sri Paramakalyani College
Alwarkurichi - 627 412, India
Abstract : The generalized t-pebbling number of a graph G, fglt
(G), is the least
positive integer n such that however n pebbles are placed on the vertices of G, we
can move t-pebbles to any vertex by a sequence of moves, each move taking
p pebbles off one vertex and placing one on an adjacent vertex. In this paper, we
determine the generalized t-pebbling number of wheel Wn and complete r-partite
graph.
Key Words : Graph, wheel and complete r-partitle graph.
1 Introduction
Let G be a simple connected graph. The pebbling number of G is the smallest
number f(G) such that however these f(G) pebbles are placed on the vertices of G,

32
we can move a pebble to any vertex by a sequence of moves, each move taking two
pebbles off one vertex and placing one on an adjacent vertex [2]. Suppose n pebbles
are distributed on to the vertices of a graph G, a generalized p pebbling step [u,v]
consists of removing p pebbles from a vertex u, and then placing one pebble on an
adjacent vertex v, for any p ≥ 2. Is it possible to move a pebble to a root vertex r, if
we can repeatedly apply generalized p pebbling steps? It is answered in the
affirmative by Chung in [1]. The generalized pebbling number of a vertex v in a
graph G is the smallest number fgl(v,G) with the property that from every placement
of fgl(v,G) pebbles on G, it is possible to move a pebble to v by a sequence of
pebbling move consists of removing p pebbles from a vertex and placing one pebble
on an adjacent vertex. The generalized pebbling number of the graph G, denoted by
fgl(G), is the maximum fgl(G) over all vertices v in G.
Again the generalized t-pebbling number of a vertex v in a graph G is the smallest
number fglt(v,G) with the property that from every placement of fglt(v,G} pebbles on
G, it is possible to move t pebbles to v by a sequence of pebbling moves where a
pebbling move consists of the removal of p pebbles from a vertex and the placement
of one of these pebbles on an adjacent vertex. The generalized t-pebbling number
of the graph G, denoted by fglt(G) is the maximum fglt(v,G) over all vertices v of G.
Throughout this paper G denotes a simple connected graph with vertex set V(G) and
edge set E(G).
x denote the largest integer less than or equal to x and x denote the smallest
integer greater than or equal to x.
2 Known Results
We find the following results with regard to the generalized pebbling numbers of
graph in [2, 6] and their generalized t-pebbling numbers in [3].
Generalized t-Pebbling Numbers of Wheel and Complete r-partite graph

33
Theorem 2.1. For a complete graph Kn, fgl(Kn) = (p-1)n-(p-2) where p ≥ 2.
Theorem 2.2. For a path of length n, fgl(Pn) = pn where p ≥ 2.
Theorem 2.3. For a star K1,n, fgl(K1,n) = (p-1)n+(p2-2p+2) if n > 1 and p ≥ 2.
Theorem 2.4. The generalized t-pebbling number for a path of length n is
fglt(Pn)=tpn.
Theorem 2.5. The generalized t-pebbling number of a complete graph on n vertices
where n ≥ 3, p ≥ 2 is fglt(Kn) = pt+(p-1)(n-2).
Theorem 2.6. The generalized t-pebbling number for a star K1,n where n > 1 is
fglt(K1,n)=p2t+(p-1)(n-2) where p ≥ 2.
Theorem 2.7. For n ≥ 4, the generalized pebbling number of the wheel graph Wn is
fgl(Wn) = (p-1)+(p2-2p+1) where p ≥ 2.
Theorem 2.8. The generalized pebbling number of the fan graph Fn is fgl(Fn) = (p-
1)n+(p2-2p+1).
Theorem 2.9. For G = 1 2 rs ,s , ... ,s
K the generalized pebbling number is given by
fgl(G) =
2
1 1
1
p ( 1)( 2).
( 1)( 2)
p s if p n s
p p n if p n s
+ − − ≥ −
+ − − < −
We will now proceed to compute the genearlized t-pebbling numbers of wheel Wn
and complete r-partite graph.
3 Computation of Genearlized t-pebbling number
Definition 3.1. We define the wheel graph denoted by Wn to be the graph with
V(Wn)= {h,v1,v2, … ,vn} where h is called the hub of Wn and E(Wn)=E(Cn) ∪ {hv1,
hv2, … , hvn} where Cn denotes the cycle graph on n vertices.
A. Lourdusamy and C. Muthulakshmi@Sasikala

34
Theorem 3.2. Let K1 = {h}. Let Cn = {v1,v2, … ,vn} be a cycle of length n. Then the
generalized t-pebbling number of the wheel graph Wn is fglt(Wn) = p2(t-1)+(p-
1)n+(p2-2p+1).
Proof : By Theorem 2.5, fglt(h,Wn) = pt+(p-1)(n-1). Let us now find the generalized
t-pebbling number of v1. Assume that v1 has zero pebbles. Let us place (p2t-1)
pebbles at 2
nv
, (p-2) pebbles at vn and (p-1) pebbles at each of wn\{v1, 2
nv
, vn}.
Then t pebbles cannot be moved to v1.
So fglt(v1,Wn) ≥ p2(t-1)+(p-1)n+(p
2-2p+1).
Let us use induction on t to prove the fglt(v1,Wn) ≤ p2(t-1)+(p-1)n+(p
2-2p+1).
For t=1, the result is true by Theorem 2.7.
By distributing p2(m-2)+(p-1)n+(p
2-2p+1) pebbles on Wn \{v1}, then we can move
(m-1) pebbles to the target vertex v1.
That is, fgl(m-1)(Wn)=p2(m-2)+(p-1)n+(p
2-2p+1). Suppose p
2(m-1)+(p-1)n+(p
2-2p+1)
pebbles are distributed on to the vertices of Wn \ {v1}. Let the target vertex be v1 of
Cn.
0.
1
If there is a vertex in Cn with at least p2 pebbles, then a pebble can be moved to v1.
Using only p2 pebbles through h. The remaining p
2(m-2)+(p-1)n+(p
2-2p+1) pebbles
are sufficient to put (m-1) additional pebbles on v1 by using induction. Otherwise
any one of the vertices of Wn \ {v1} say
2
nv
receive at least p pebbles and each of
the vertices Wn\ {v1, 2
nv
} receive p-1 pebbles then from 2
nv
using a sequence of
Generalized t-Pebbling Numbers of Wheel and Complete r-partite graph

35
pebbling moves, 2
nv
, 1
2
nv
−
, … ,v1 we can move a pebble to v1. Remaining p2+(p-1)
(n-2
n
+2)+(p2-3p+1) > 0. So by induction, (m-1) pebbles can be moved to v1.
Hence in all cases fglm(v1,Wn) ≤ p2(m-1)+(p-1)n+(p
2-2p+1). Therefore fglt(Wn)=p
2(m-
1)+(p-1)n+(p2-2p+1).
Definition 3.3. A graph G = (V,E) is called an r-partite graph if V can be partitioned
into r non-empty subsets V1,V2, … ,Vr such that no edge of G joins vertices in the
same set. The sets V1,V2, … ,Vr are called partite sets or vertex classes of G. If G is
an r-partite graph having partite sets V1,V2, … ,Vr such that every vertex of Vi is
joined to every vertex of Vj where 1 ≤ i, j ≤ r and i ≠ j, then G is called a complete r-
partite graph. If |Vi|=si for i=1,2, … , r then we denote G by 1 2 rs ,s , ... ,s
K .
Notation 3.4. For s1 ≥ s2 ≥ … ≥ sr, s1 > 1 and if r = 2, s2 > 1, let 1 2 rs ,s , ... ,sK be the
complete r-partitle graph with s1, s2, … , sr vertices in vertex classes C1, C2, … , Cr
respectively. Let n = 1
r
i
i
s=
∑ .
Theorem 3.5. For G = 1 2 rs ,s , ... ,sK the generalized t-pebbling number for a complete r-
partite graph G is given by
fglt(G) = 1
2
1 1
( 1)( 2).
p ( 1)( 2)
pt p n if pt n s
t p s if pt n s
+ − − < −
+ − − ≥ −
Proof :
Case i: Assume pt < n - s1.
A. Lourdusamy and C. Muthulakshmi@Sasikala

36
Let us place pt+(p-1)(n-2)-1 pebbles on the vertices of G-{v} as follows. Let us
choose (t-1) vertices and we place p+(p-1) pebbles on each of the (t-1) vertices and
we place (p-1) pebbles each on the remaining vertices clearly t pebbles cannot be
moved to v.
Hence fglt(v,G) > (t-1)[(p+(p-1)]+(p-1)(n-t)
= pt+(p-1)(n-2)-1
≥ pt+(p-1)(n-2).
Next we will use induction to show that pt+(p-1)(n-2) pebbles are sufficient to move
t pebbles to any desired vertex. For t=1 results is true by Theorem 2.9. Suppose t >
s1, and pt+(p-1)(n-2) pebbles are placed on the vertices of G. Let the target vertex be
v of Ck for some k=1, 2, … , n. If there is a vertex w of Cj (j ≠ k) with at least p
pebbles then a pebble can be placed on v.
The remaining p(t-1)+(p-1)(n-2) pebbles are sufficient to put (t-1) additional pebbles
on v by induction. If not then every vertex of G\Ck wil have at most (p-1) pebbles on
it. Suppose among these n-sk vertices, q is the number of vertices with at least one
pebble. Therefore there will be pt+(p-1)(n-2)-q pebbles on the vertices of Ck. We
consider the following cases.
Subcase I : q ≥ t.
We use pebbling move from sk-1 vertices of Ck\{v} to put the remaining at most (p-
1) pebbles on each of the t of the q occupied vertices of v(G)-Ck. Using (p-1)t
pebbles we can pebble t vertices with (p-1) pebbles. Then remaining (p-1)(n-2)-(q-t)
pebbles are in Ck\{v}. From the t vertices with p pebbles we can move t pebbles to v.
Generalized t-Pebbling Numbers of Wheel and Complete r-partite graph

37
Subcase ii : q < t.
As in subcase (i) first we will put (p-1) more pebbles on each of these q vertices by
maiing (p-1)q moves from the vertices of Ck\{v} in order to put q pebbles on v. Then
we have to place t-q additional pebbles on v. So we use p2(t-q)+(p-1)pq=p
2t-pq
pebbles among pt+(p-1)(n-2)-q pebbles in the vertices of Ck\{v}. Hence in all the
cases fglt(v,G) ≤ pt+(p-1)(n-2).
Case ii: Assume pt ≥ n - s1.
Let the vertices of C1 be v1, v2, … , vn and let 1s
v be the target vertex. Let us place
p2t+(p-1)(s_1-2) pebbles on the vertices of C1 as follows. Let us place p
2t-1 pebbles
on v1 and place (p-1) pebbles each on (s1-2) vertices of C1 other than v1 and 1s
v . In
this case t-pebbles cannot be moved to 1s
v . Hence fglt(G) ≥ p2t+(p-1)(s1-2).
Next we will use induction on t to prove that p2t+(p-1)(s1-2) pebbles are sufficient to
put t pebbles on any desired vertex clearly the claim is true for pt=n-s1.
Since by case(i) fglt(G) = pt+(p-1)(n-2)
= pt+(p-1)(pt+s1-2)
= p2t+(p-1)(s1-2).
Suppose p(m-1) > n-s1 and fgl(m-1)(G) = p2t(m-1)+(p-1)(s1-2) = p
2m+(p-1)s1-
(p2+2p+2).
We prove the result is true for m where pm > n-s1. Suppoe p2m+(p-1)(s1-2) pebbles
are distributed on the vertices of G. Let the target vertex be v of Ck. If there is a
vertex in some Cj (j ≠ k) with at least p pebbles, then a pebble can be placed on v
2
A. Lourdusamy and C. Muthulakshmi@Sasikala

38
using only p pebbles. The remaining p2m+(p-1)s1-3p+2 pebbles are sufficient to put
(m-1) additional pebbles on v, since p2+2p-2-3p+2 > 0. If not then every vertex of
G\Ck will contain either zero or at least one pebble on it. If there is a vertex say w in
some Cj (j ≠ k) with at least one pebble on it, we use (p-1)p pebbles from the vertices
of Ck to put (p-1) pebbles on w and hence a pebble can be placed on v. Since p2+2p-
2-(p-1)(p+3) > 0, then remaining fgl(m-1)(G) pebbles would suffice to put (m-1)
additional pebbles on v. Otherwise, every vertex of G\Ck will have zero pebbles,
using p2 pebbles we can place a pebble on v in this case the remaining p
2(m-1)+(p-
1)(s1-2) pebbles would suffice to put (m-1) additional pebbles on v. Thus fglm(v,G) ≤
p2m+(p-1)(s1-2). Therefore by induction fglt(v,G) ≤ p
2t+(p-1)(s1-2) for all pt < n-s1.
Thus fglt(G) < p2t+(p-1)(s1-2) for all pt ≥ n-s1 and so the proof is over.
References :
[1] F.R.K.Chung, Pebbling in Hypercubes, SIAM J. Discrete Maths., Vol 2(4)(1989)
pp 467-472.
[2] G. Hurlbert, Recent Progress in graph pebbling, Graph Theory notes of New York
XLIX (2005), 25-34.
[3] A. Lourdusamy and C. Muthulakshmi@ Sasikala, Generalized Pebbling Number,
International Mathematical Forum, 5, 2010, No.27, pp.1331-1337.
[4] A. Lourdusamy and C. Muthulakshmi@ Sasikala, Generalized t-pebbling Number of
a Graph, Journal of Discrete Mathematical Sciences & Cryptography, Vol. 12 (2009),
No. 1, pp. 109-120.
[5] A. Lourdusamy and C. Muthulakshmi@ Sasikala, Generalized pebbling Numbers of
some Graphs, Sciencia Acta Xaveriana,Vol3, No.1, (20012), pp107-114.
[6] A. Lourdusamy and A. Punitha Tharani, On t-pebbling graphs, Utilitas Mathematica,Vol.
87,( 2012), pp.331-342.
Generalized t-Pebbling Numbers of Wheel and Complete r-partite graph

39
Volume 5
No. 2
pp. 39-38
September 2014
Sciencia Acta XaverianaAn International Science JournalISSN. 0976-1152
Super Vertex Mean Graphs
1. Introduction
A vertex labeling1 of a graph G is an assignment ƒ of labels to the vertices of G that
induces a label for each edge uv depending on the vertex labels. An edge labeling of a
graph G is assignment ƒ of labels to the edges of G that induces a label for each
vertex v depending on the edge labels. Let G = (V, E) be a simple graph with p
vertices and q edges. A mean labeling ƒ is an injection from V to the set (0,1,2,…, q)
that induces for each edge uv the label ( ) ( )
2
u v
such that the set of edge labels is
(1, 2,…, q). Mean labeling was introduced by Somasundaram and ponraj [8]. A graph
that accepts a mean labeling is known as mean graph. A super mean labeling ƒ is an
injection from V to the set {1,2,…, p+q} that induces for each edge uv the label
( ) ( )
2
u v
such that the set of all vertex labels and the induced edge labels is
{1,2,…,p+q}. In this paper we study super vertex behavior of certain classes of graph.
Super vertex mean labeling was introduced by R.Ponraj et al.[7]. A graph that
A.Lourdusamy1, M.Seenivasan2, Sherry George3 and R.revathy4
Abstract : An edge labeling of a graph G is an assignment ƒ of labels
to the edges of G that induces a label for each vertex depending on
the edges labels. A super vertex mean labeling ƒ is an injection from E
to the set {1,2,3,...,p+q} that induces for each vertex the label
Round such that the set of all edge labels and the induced
vertex labels is {1,2,3,...,p+q}. In this paper we study super vertex
behavior of certain classes of graphs.

40
(1,2,…,p+q}. In this paper we study super vertex behavior of certain classes of graph.
Super vertex mean labeling was introduced by R.Ponraj et al.[7]. A graph that
accepts a super mean labeling is known as super mean graph. Some results on mean
labeling and super mean labeling are given in [4, 5, 6, 7, 8, 9]. For a summary on
various graph labeling see the Dynamic survey of graph labeling by Gallian [2].
Lourdusamy and Seenivasan [3] introduced vertex mean labeling as an edge
analogue of mean labeling as follows: A vertex mean labeling of a (p,q) graph G(V,E)
is defined as an injection f :E {0,1,... , q*}, q* = max(p,q) such that the injection f :
V N defined by the rule v(V) = Round
( )
( )
ev e
d v
satisfies the property that
v(V) = {
v (u): u } = {1,2,..., p}, where v denotes the set of edges in G that
are incident at v and N denotes the set of all natural numbers. A graph that has a
vertex mean labeling is called a vertex mean graph or V-mean graph. For all
terminology and notations in graph theory, we refer the reader to the text book by
D.B.West [10]. All graphs considered in this paper are finite and simple. Motivated
by the concept of super mean labeling, we introduce super vertex mean labeling of
graphs as follows
Definition 1.1. A Super vertex mean labeling f of a graph G(V,E) is an injection from E
to the set {1,2,3,...,p+q} that induces for each vertex v the label v(v) = Round
( )
( )
EV e
d v
such that the set of all edge labels and the induced vertex labels is
{1,2,3,... , p+q}.
Henceforth we call super vertex mean as SVM. To initiate the investigation we obtain
certain classes of graphs which are SVM graphs. It is obvious that no tree is an SVM
graph. We also observe that C4 is not an SVM graph.
Super Vertex Mean Graphs

41
1. Some SVM Graphs
Theorem 2.1. The cycle Cn is an SVM graph if and only if n ? 4.
Proof: Let {e1, e2, ..., en} be the edge set of Cn such that ei =vivi+1, ,
en = vnv1. Checking each of the possibilities reveals that the cycle C4 is not an
SVM. So we assume that n 4.
Case 1: n 1 (mod 2). Let n = 2r+1. The edges of Cn are labeled as follows:
f (ei) =
It is easy to observe that f is injective. The induced vertex labels are given as
follows
v(vi) =
It is clear that f (E) U v(V) = {2i – 1: 1 } U {
= {1,2,3, ..., 2n}.
Case 2: n let n = 2r. The edges of Cn are labeled as follows:
A.Lourdusamy1, M.Seenivasan2, Sherry George3 and R.revathy4
42
Hence the theorem.
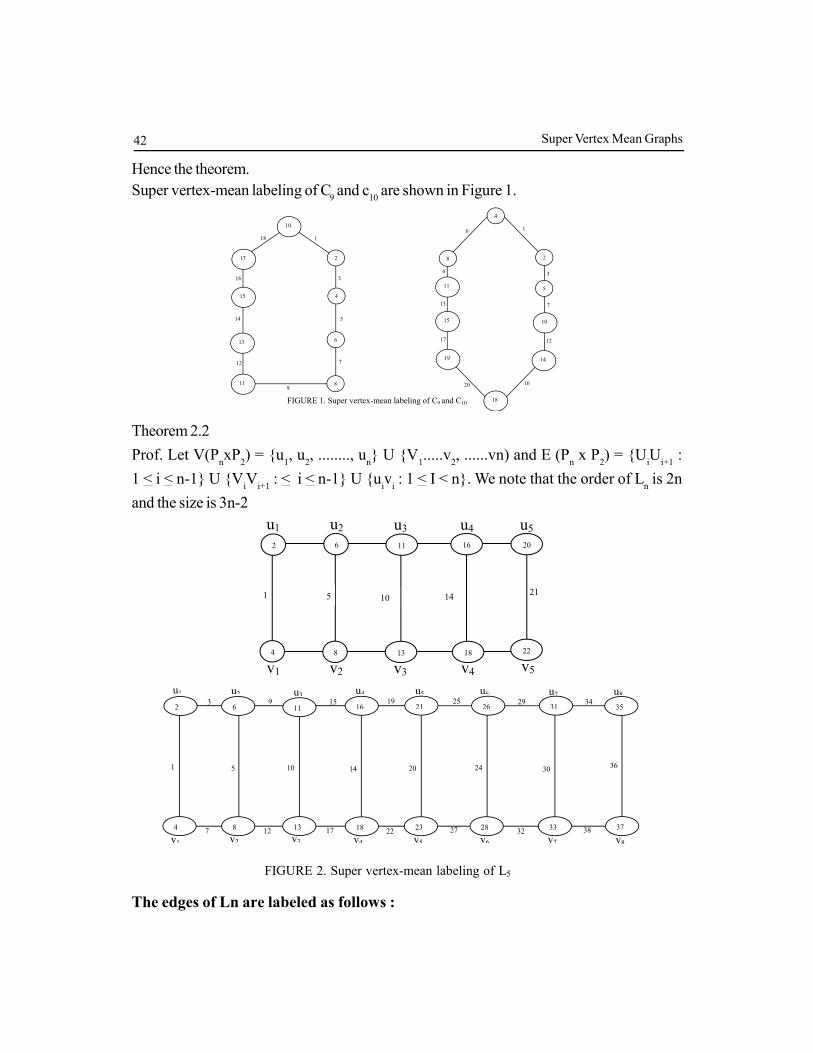
Super vertex-mean labeling of C9 and c
10 are shown in Figure 1.
FIGURE 1. Super vertex-mean labeling of C9 and C10
7
9
12
14 5
16
18
3
1
10
2 17
15
13
11
4
8
6
16 20
17 12
7 13
6 3
1 6
5
4
8 2
10
14 19
18
15
11
Theorem 2.2
Prof. Let V(PnxP
2) = {u
1, u
2, ........, u
n} U {V
1.....v
2, ......vn) and E (P
n x P
2) = {U
iU
i+1 :
1 < i < n-1} U {ViV
i+1 : < i < n-1} U {u
iv
i : 1 < I < n}. We note that the order of L
n is 2n
and the size is 3n-2
v1 v2 v3 v4 v5
u5 u4 u3 u2 u1
21 1
2 6
22 4 8 13
11
18
16 20
5 10 14
FIGURE 2. Super vertex-mean labeling of L5
and L8
3 9 15 19 25 29 34
38 32 27 22 17 12 7
u8 u7 u6 u5 u4 u3 u2 u1
1
v8 v7 v6 v5 v4 v3 v2 v1
5 10 14 20 24 30 36
4 8 13 18 23 28 33 37
35 31 26 21 16 11 6 2
The edges of Ln are labeled as follows :
Super Vertex Mean Graphs

43
f (uiui+1) =
f (vivi+1) =
f (uivi) =
it is easily observed that f is injective. The induced vertex labels are as follows:
v(ui) =
v(vi) =
It is easy to verify, in both cases, that the set of all edge labels and the induced
vertex labels is {1,2,....,5n-2}.
Hence the theorem.
SVM labeling of L5 and L8 are shown in Figure 2.
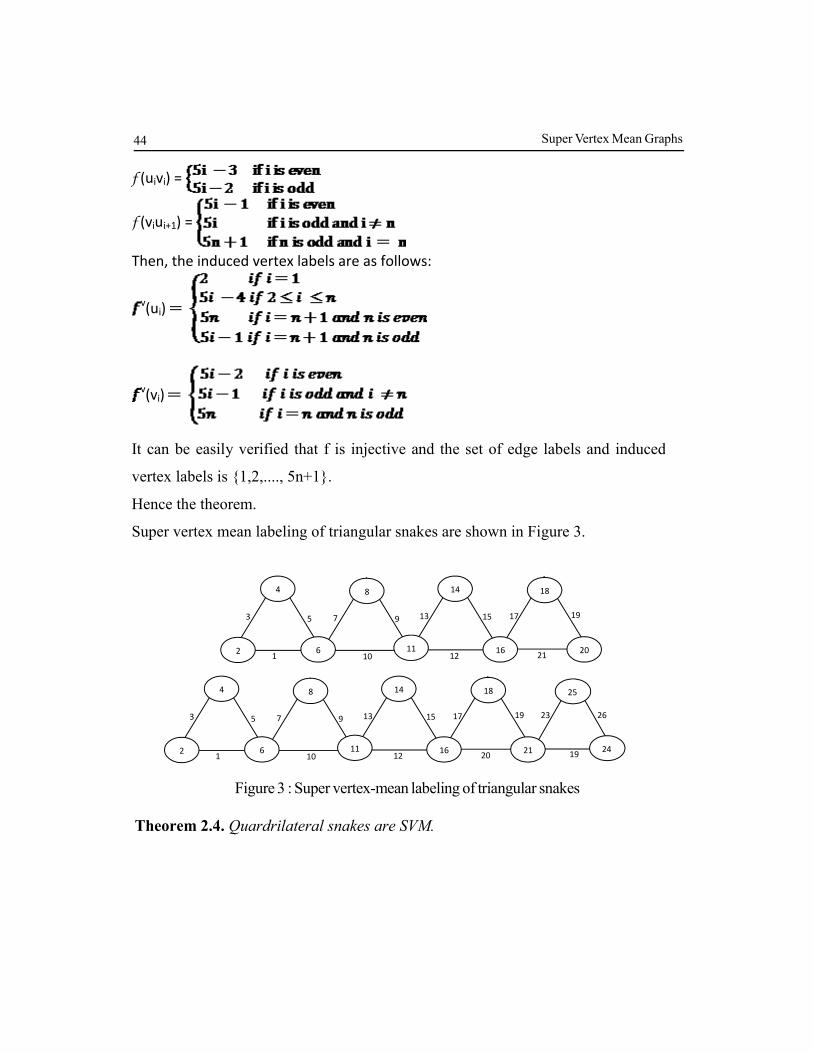
Theorem 2.3. A triangular snake with n blocks is SVM.
Proof. Let Gn be a triangular snake with n blocks on p vertices and q edges. Then p
= 2n + 1 and q = 3n.
Let V(Gn) = {ui : U vi : and V(Gn) = {uiui+1, uivi, ui+1vi :
The order of Gn is 2n+1 and size is 3n.
The edges of Gn are labeled as follows :
f (uiui+1) =
A.Lourdusamy1, M.Seenivasan2, Sherry George3 and R.revathy4
44
f (uivi) =
f (viui+1) =
Then, the induced vertex labels are as follows:
v(ui)
v(vi)
It can be easily verified that f is injective and the set of edge labels and induced
vertex labels is {1,2,...., 5n+1}.
Hence the theorem.
Super vertex mean labeling of triangular snakes are shown in Figure 3.
19
21
17 15
12
13 9
10
7 5
1
3
2
4
6
8
11
14
16
18
20
23
19
26 19
20
17 15
12
13 9
10
7 5
1
3
2
4
6
8
11
14
16
18
21 24
25
FIGURE 3. Super vertex-mean labeling of triangular snakes Figure 3 : Super vertex-mean labeling of triangular snakes
Theorem 2.4. Quardrilateral snakes are SVM.
Super Vertex Mean Graphs

45
Proof. Let Gn be a quadrilateral snake with V(Gn) = ui : U ui, wi :
and E(Gn) = uiui+1, uivi, ui+1wi, viwi : . Then p = 3n + 1 and q =
4n. Define f: E(Gn) {1,2,3,..., 7n+1} as follows:
i i+1) =
i i) =
i i) =
i i+1) =
Then the induced vertex labels are as follows :
vi) =
vi) =
vi) =
It can be easily verified that is injective and the set of edge labels and induced
vertex labels I {1,2,3,..., 7n+1}.
Hence the theorem.
A Super vertex-mean labeling of a Quadrilateral snake is shown in Figure 4.
A.Lourdusamy1, M.Seenivasan2, Sherry George3 and R.revathy4

46
27
25
22
29
20
18
13
11
8
15
14
6
3
1
7
2
4
5
9
10 12
16
17 19
23
24 26
28
FIGURE 4. A Super vertex-mean labeling of a Quadrilateral snake
References :
[1] B.D.Acharya and K.A.Germina. Vertex-graceful Graphs. Journal of Discrete
Mathematical Science & Chryptography, Vol.13(2010), No.5, pp.453-463.
[2] J.A.Gallian. A Dynamic Survey of Graph Labeling. The electronic journal of
combinatorics, 18, 2011.
[3] A.Lourdusamy and M.Seenivasan. Vertex-mean Graphs. International journal of
Mathematical Combinatorics, 3:114-120, 2011.
[4] A.Lourdusamy and M.Seenivasan. Mean Labelings of Cyclic Snakes. AKCE
International Journal of Graphs and Combinatorics, 8(2): 105-113, 2011.
[5] R.Ponraj, Studies in Labelings of Graphs. Ph.D.thesis, Manonmaniam Sundaranar
University, India (2004).
[6] R.Ponraj and D.Ramya. Super Mean Labeling of Graphs, Preprint.
[7] D.Ramya, R.Ponraj, and P.Jeyanthi. Super Mean Labeling of Graphs, Ars Combin.,
to appear.
[8] S.Somasundaram and R.Ponraj. Super Mean Labeling of Graphs, Natl Acad., Sci.Let.,
26(2003) 210-213.
[9] R.Vasuki and A.Nagrajan. Some Results on Super Mean Labeling of Graphs.
International Journal of Mathematical Combinatorics, Vol.3 (2009) 82-96.
[10] D.B.West. Introduction to Graph Theory. Prentice-Hall of India, Private Limited, New
Delhi, 1996.
Super Vertex Mean Graphs
