role of superoxide dismutase in the oxidation of n-alkanes by yeasts
TRANSCRIPT

Free Radical Biology & Medicine, Vol. 11, pp. 263-268, 1991 0891-5849191 $3.00 + .00 Printed in the USA. All rights reserved. Copyright ~ 1991 Pergamon Press plc
Original Contribution
R O L E O F S U P E R O X I D E D I S M U T A S E IN T H E O X I D A T I O N O F
N - A L K A N E S B Y Y E A S T S
ANNA V. KUJUMDZIEVA-SAvoVA,* VALENTIN A . SAVOV, ELENA I. GEORGIEVAt
University of Sofia, Center of Biotechnology, Bul. "D. Tzankov" N 8, Sofia, Bulgaria; tBulgarian Academy of Sciences, Institute of Genetics "D. Kostov", Molecular Genetics Laboratory, 1113 Sofia, Bulgaria
(Received 29 September 1989; Revised 21 August 1990; Rerevised 7 March 1991; Accepted 21 March 1991)
A b s t r a c t - - Y e a s t microorganisms from Candida genus are investigated for their superoxide dismutase (SOD) and catalase activ- ity during cultivation on N-alkanes. The later caused a considerable increase of Cu/Zn SOD activity of yeast cells in comparison with glucose. A correlation between SOD and catalase activity existed. It is further observed that cells of Candida lipolytica 68-72 which contain a high level of Cu/Zn SOD were more resistant to lethality of exogenous O - . An over-production of Cu/Zn SOD
• 2 . .
during the assimilation of N-alkanes by yeasts is also connected to their considerable resistance to increased concentratmns of Cu 2+ and Zn 2+ ions in the nutrient medium. The results are consistent with the assumption that the enhanced resistance of yeast cells to O - and high concentrations of Cu 2+ and Zn2+-ions are due to the increased activity of Cu/Zn SOD and that SOD is • . 2
revolved m the protection of some cellular components. Polyacrylamide gel electrophoresis of Candida lipolytica cell-free extracts revealed the same chromatic bands of SOD activity
under growth on glucose and N-alkanes. The type of the carbon source used from yeast cells as a single source of carbon and energy had no influence on the SOD profile of the cell•
Keywords--N-Alkanes, Catalase, Oxidation, Superoxide Dismutase, Yeast, Free radicals
INTRODUCTION
In recent years attention has been increasingly paid to alkane-utilizing microorganisms with respect to the syn- thesis of wide range of microbially derived products. 1,2 Particularly, the metabolism of N-alkanes by microor- ganisms has been extensively investigated. 3 As it is known, alkanes uptaken by yeast cells are subjected to initial oxidation, followed by further metabolic process- ing. For this initial oxidation of N-alkanes three differ- ent mechanisms are proposed. Alkane molecules are susceptible to such oxidation in mono-, di- terminal and internal positions of the carbon chain. The most probable alkane oxidation system in yeasts is the hy- droxylational involving cytochrome P450 which oper- ates under aerobic conditions. 4
The product of this hydroxylating reaction is the corresponding alcohol. It is further metabolized by yeast cells through an oxidation reaction. As a result of oxidation of the obtained primary alcohols superox- ide radical is formed. 5
In this work we have studied the activities of super- oxide dismutase (SOD) and catalase during the growth
of yeasts on N-alkanes in dependence on the superox- ide radicals production during oxidation.
MATERIALS AND METHODS
Microorganisms
The following strains were used: Candida lipolytica 68-72, Candida lipolytica 68-16, Candida tropicalis Kd-14, and Candida guilliermondii 63-8.
Culture media
Rider medium was used in all experiments. The tested carbon sources were: glucose 20 g/L; N-alkanes Clo - C2o--1% (w/v). Instead of yeast extract the medium was supplemented with biotine -- 20 mkg/L and tiamine--40 mkg/L.
Cultivation
Cultivation was carried out at 28°C in 500 mL Er- lenmeyer flasks containing 100 mL glucose or N-al-
*Author to whom correspondence should be addressed.
263

264 A .V. KUJUMDZIEVA-SAVOVA et al.
kane supplemented medium. The inoculum was precultured at 28°C for 24 h on slant agar and one tube of start culture was added to the nutrient medium. After culti- vation in a reciprocal shaker, the culture was harvested in its late exponential phase by centrifugation at 5000 x g for 15 min at 4°C. The strain Candida lipolytica 68-72 was cultivated in a 10L working volume fermen- tor (New Brunswick) at 28°C. pH was maintained at 4,5 by titration with 25% NH4OH. The culture was permanently aerated and stirred at a constant rate of 300 rpm.
Preparation of cell-free extracts
For the preparation of cell-free extracts the harvested cells were washed twice with 0,05 M potassium phos- phate buffer, containing 0,1 mM EDTA pH 7,8 and were suspended in a small amount of the same buffer. The cell suspension was disrupted in a Vibration Ho- mogenizator VHG with glass beads. The homogenates were centrifuged at 3000 x g, and after freezing and thawing--at 13000 x g for 15 min at 4°C. The super- natant obtained was used as a cell-free extract for en- zyme determination.
Enzyme assays
SOD determination was performed on dialyzed cell- free extracts as described. 6 One unit of SOD was de- fined as the quantity of enzyme causing 50% inhibition of the nitrotetrazoliumblau chloride (NBT) reduction and was expressed as U x mg protein -1
Catalase activity was determined spectrophotometri- cally according to Aebi. 7 Enzyme activity was ex- pressed as AE x min- I x mg protein-1.
Analysis of glucose and N-alkanes
All assays were performed on samples with cell mass removed by centrifugation. Glucose was mea- sured by the method of Somogy 8 and Nelson. 9 The concentration of N-alkanes was determined by gas chro- matography.
Assay methods
Growth measurements. Growth was followed by mea- suring the absorbance at 540 n m (A540).
Determination of growth yield. The growth yield with respect to glucose (Yglu) was calculated as a dry weight of cell mass produced per gram of glucose con- sumed.
Protein determination. The protein content in the cell free extracts was measured according to Lowry et al. 1° with bovine serum albumin as a standard.
Polyacrylamide gel electrophoresis. Electrophoresis was performed in 10% polyacrylamide slab gel according to the method of Davis.ll Bromphenolblue was used as a tracing dye. SOD activity was localized by soaking the gels in 2 mM dianisidine, 0, 1 mM riboflavin, 10 mM potassium phosphate pH 7,2 for 1 h at room tempera- ture, followed by a brief rinsing with water and by il- lumination for 5-15 min as described by Misra and Fridovich. 12 As a result of SOD activity, a brown band appeared against a pale yellow background. The pres- ence of Cu/Zn SOD was detected by inhibition caused by 2 mM KCN. Human blood SOD from Serva was used as a control.
RESULTS
Effect of glucose and N-alkanes on the synthesis of SOD and catalase
The specific activity of SOD enzymes in the crude extracts from Candida lipolytica 68-72, 68-16, Candida tropicalis Kd-14, and Candida guilliermondii 63-8 grown to their late exponential phase, were estimated to be 70-80 U x mg protein-1 for cultures grown on N-al- kane and 2-5 U x mg protein-l if grown on glucose (Table 1). It should be noted that the increase in SOD activity during cultivation with N-alkanes was 15-20- fold higher in comparison with glucose-grown culture. A similar effect was observed in the activity of cata- lase, which marked a 10 to 15-fold increase. The SOD from all tested strains was sensitive to an inhibition caused by 1 mM KCN and insensitive to 5-10 mM NaN 3. These findings suggested the presence of Cu, Zn-containing SOD in N-alkane grown cells.
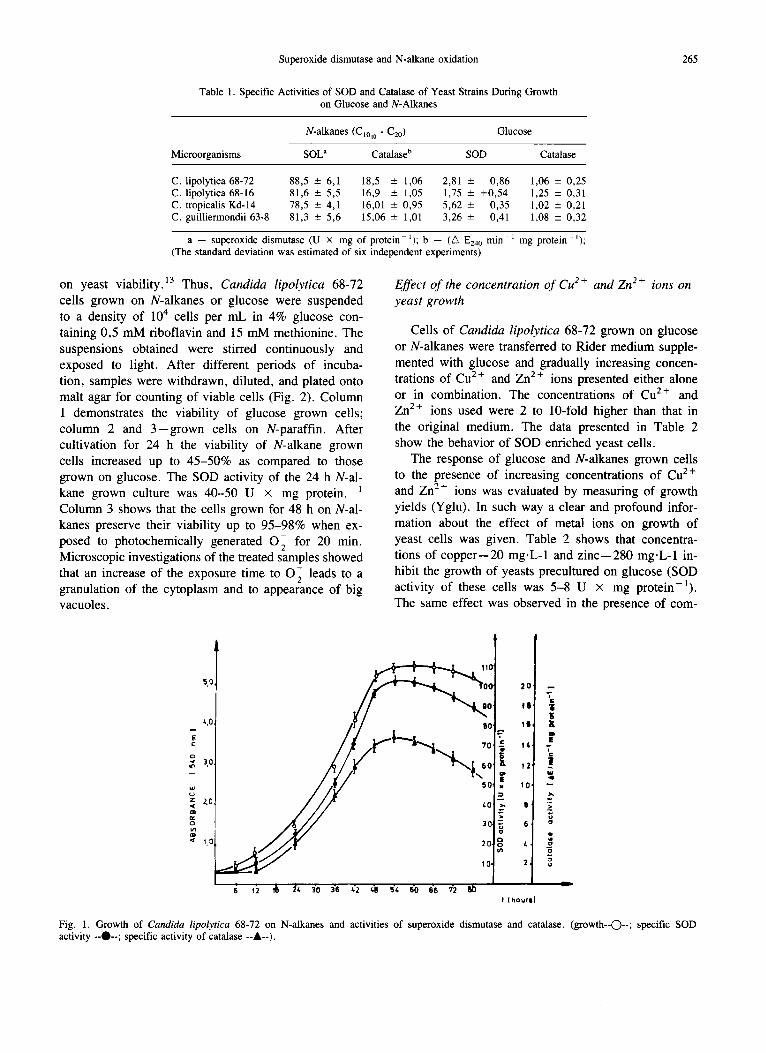
In order to investigate the dynamics of SOD pro- duction under conditions of constant aeration and pH, Candida lipolytica 68-72 was grown batchwise in a fermentor. Tested strains started to remove actively N-alkanes from the nutrient medium with a lag period of 8-10 h. Culture turbidity, as measured by the ab- sorbance at 540 nm, increased approximately from 0,03 to 5,0 for 48 h (Fig. 1). The initial SOD activity was 2,0-7,2 U x mg protein-l , and after 48-50 h of cultivation on N-alkanes mixture it increased up to 90- 100 U x mg protein-1. The same effect was observed for the catalase activity, as compared to the initial value, it marked a 10-15-fold increase.
Effect of the exogenous O- 2 on the viability of yeast cells
Photochemically assayed SOD activity was used as a basis for investigation of the effect of exogenous O
Superoxide dismutase and N-alkane oxidation 265
Table 1. Specific Activities of SOD and Catalase of Yeast Strains During Growth on Glucose and N-Alkanes
N-alkanes (C~o~0 - C2o ) Glucose
Microorganisms SOL ~ Catalase b SOD Catalase
C. lipolytica 68-72 88,5 - 6,1 18,5 - 1,06 2,81 +- 0,86 1,06 ± 0,25 C. lipolytica 68-16 81,6 ± 5,5 16,9 ± 1,05 1,75 ± +0,54 1,25 +- 0,31 C. tropicalis Kd-14 78,5 _+ 4,1 16,01 ± 0,95 5,62 ± 0,35 1,02 +- 0,21 C. guilliermondii 63-8 81,3 -+ 5,6 15,06 ± 1,01 3,26 ± 0,41 1,08 +__ 0,32
a - - superoxide dismutase (U × mg of protein-~); b - - (Zk E240 min -1 mg protein-l); (The standard deviation was estimated of six independent experiments)
on yeast viability. ]3 Thus, Candida lipolytica 68-72 cells grown on N-alkanes or glucose were suspended to a density of 104 cells per mL in 4% glucose con- taining 0,5 mM riboflavin and 15 mM methionine. The suspensions obtained were stirred continuously and exposed to light. After different periods of incuba- tion, samples were withdrawn, diluted, and plated onto malt agar for counting of viable cells (Fig. 2). Column 1 demonstrates the viability of glucose grown cells; column 2 and 3--grown cells on N-paraffin. After cultivation for 24 h the viability of N-alkane grown cells increased up to 45-50% as compared to those grown on glucose. The SOD activity of the 24 h N-al- kane grown culture was 40-50 U x mg protein.- ] Column 3 shows that the cells grown for 48 h on N-al- kanes preserve their viability up to 95-98% when ex- posed to photochemically generated 0 2 for 20 min. Microscopic investigations of the treated samples showed that an increase of the exposure time to O2 leads to a granulation of the cytoplasm and to appearance of big vacuoles.
Effect of the concentration of C u 2 + and Zn 2 + ions on yeast growth
Cells of Candida lipolytica 68-72 grown on glucose or N-alkanes were transferred to Rider medium supple- mented with glucose and gradually increasing concen- trations of Cu 2 + and Zn 2 ÷ ions presented either alone or in combination. The concentrations of Cu 2+ and Zn 2÷ ions used were 2 to 10-fold higher than that in the original medium. The data presented in Table 2 show the behavior of SOD enriched yeast cells.
The response of glucose and N-alkanes grown cells to the presence of increasing concentrations of Cu 2+ and Zn 2 ÷ ions was evaluated by measuring of growth yields (Yglu). In such way a clear and profound infor- mation about the effect of metal ions on growth of yeast cells was given. Table 2 shows that concentra- tions of copper--20 mg.L-1 and zinc--280 mg.L-1 in- hibit the growth of yeasts precultured on glucose (SOD activity of these cells was 5-8 U × mg protein-]). The same effect was observed in the presence of com-
l ~ 110
_,o //.
! 30,
/ ,g ' ",o ~ 30.
<~ 10, 20. 10.
!0' rS '
,21
01
s l
61
24
I
t ( ho~rl}
i¢
"6
t
|
>
sa
Fig. 1. Growth of Candida lipolytica 68-72 on N-alkanes and activities of superoxide dismutase and catalase. (growth--C)--; specific SOD activity --O--; specific activity of catalase --A--).

Metal Ion Concentration in Rider's Medium mg.L- l
A. V. KUJUMDZIEVA-SAvoVA et al.
Table 2. Influence of Different Concentrations of Cu 2+ and Zn 2+ Ions in Nutrient Medium on Growth Yields (Yglu) of Candida lipolytica 68-72
Zn 2+
Growth Yields (Yglu)
Y g l u a Y g l u b Y g l u c
C u 2+
%
100'
70 0,485 -+ 0,021 0,478 ± 0,018 0,481 ± 0,019 140 0,470 ± 0,019 0,480 -+ 0,021 0,480 ± 0,020 280 0,338 ± 0,015 0,478 ± 0,019 0,478 + 0,018 420 0,301 ± 0,016 0,400 ± 0,015 0,479 ± 0,019 560 0,248 • 0,011 0,350 ± 0,015 0,472 ± 0,017 700 0,200 --- 0,010 0,321 --- 0,013 0,461 ± 0,019
10 0,481 -+ 0,018 0,476 + 0,018 0,461 ±- 0,018 20 0,380 ± 0,015 0,470 + 0,018 0,458 --- 0,018 40 0,305 ± 0,014 0,478 ± 0,019 0,463 ± 0,020 60 0,251 ± 0,011 0,310 ± 0,014 0,458 ± 0,017 80 0,211 ± 0,010 0,300 ± 0,014 0,448 ± 0,017
100 0,140 ± 0,007 0,263 ± O,Oll 0,450 ± 0,017 Cu 2+ + Zn 2. 10 70 0,478 ± 0,019 0,481 ± 0,018 0,480 ± 0,019 20 140 0,356 ± 0,015 0,480 ± 0,020 0,478 ± 0,020 40 280 0,248 ± 0,010 0,468 ± 0,017 0,479 ± 0,019 60 420 0,200 -+ 0,010 0,301 -- 0,014 0,305 -- 0,014 80 560 0,131 ± 0,007 0,090 ± 0,005 0,256 ± 0,012 100 700 0,110 ± 0,007 0,078 ± 0,005 0,200 ± 0,010
Note. a--Candid_a lipolytica cells were precultured 24 h on glucose (SOD activity was 5-8 U × mg of protein ); b -- Candida lipolytica cells were precultured 24 h on N-alkane (SOD activity was 40--45 U × mg of protein- i); c -- Candida lipolytica cells were precultured 48 h on N-alkane (SOD activity was 92-98 U × mg of protein-1); (The standard deviation was estimated of six independent experiments).
b ina t i on o f Cu 2+ and Z n 2+ ions , too (20 m g - L - l and
140 m g . L - t , r espec t ive ly ) . T he dec rease in Yglu f rom
0 .48- -0 .47 to 0 . 1 5 - 0 . 1 1 (g cel ls /g g lucose c o n s u m e d )
o b s e r v e d du r ing cu l t iva t ion o f yeas ts on g radua l ly in-
c reas ing Cu 2 + and Z n 2+ concen t r a t i ons sugges t ed that
the to le rance to these ions o f g lucose g r o w n cel ls was
ve ry low.
80-
I 60. o
s-~ 40. E~
>
~ 20.
I0 t (rain)
266
1 2b
Fig. 2. Effect of superoxide dismutase on the lethality of photochem- ically generated O- to Candida lipolytica 68-72 (1-glucose grown
2 . cells; 2-yeast cells culnvated on N-alkanes 24h; 3-yeast cells culti- vated on N-alkanes for 48 h.
Cul tu re g r o w n for 24 h on N-a lkanes cou ld wi th-
s tand wel l a two to four - fo ld increase in the concen t ra -
t ion of these me ta l ions ( S O D act iv i ty o f this i n o c u l u m
was 4 0 - 4 5 U × m g p r o t e i n - 1). A dec rease o f Yglu to
4 0 - 5 0 % at 10-fold inc reased ions concen t r a t i ons was
d e m o n s t r a t e d . In the case o f c o m b i n a t i o n of the two
ions tes ted a s t rong decrease o f Yglu was obse rved at
8 - 1 0 fold inc reased concen t r a t i ons .
Tab le 2 shows that the h ighes t res i s tance o f yeasts
to ion concen t r a t i ons tes ted was o b s e r v e d w h e n a 48-h
i n o c u l u m g r o w n on N-a lkanes was used ( S O D act iv i ty
o f this cu l ture was 9 2 - 9 8 U × m g p r o t e i n - l ) . Thus ,
the Yglu va lues d id not c h a n g e cons ide rab ly at 2 to
10-fold h i g h e r ion concen t ra t ions . On ly the c o m b i n e d
p re sence of a s ix-fold inc reased concen t r a t i ons o f Cu 2 ÷
and Z n 2÷ ions caused g r o w t h inh ib i t ion (Yglu de-
c reased up to 0 ,35 + 0 ,02) . D u r i n g the cu l t iva t ion of
yeas t cel ls at 8 - 1 0 t imes h i g h e r concen t r a t i on of me ta l
ions in the nu t r i en t m e d i u m part ia l lysis o f the cul ture
cou ld be obse rved . It shou ld be po in ted out tha t the
toxic i ty o f the Z n z+ ions ( for yeas t cel ls tes ted) was
less than the toxic i ty found for the Cu 2 + ions.
Visualization and inhibition sensit ivi ty o f S O D activi ty
T h e S O D act iv i ty o f Candida lipolytica 68-72 was
v i sua l i zed by e l ec t rophores i s o f cel l free ext rac ts on
10% p o l y a c r y l a m i d e gel , f o l lowed by a speci f ic s tain-

Superoxide dismutase and N-alkane oxidation 267
1 2 3
m
mama
top
"~mum
m
bottom
Fig. 3. Electrophoretic migration patterns of SOD of: 1--Candida lipolytica 68-72 cultivated on glucose, 2--Candida lipolytica 68-72 grown on N-alkanes, and 3--control-human blood SOD. Crude cell- free extracts were subjected to 10% polyacrylamide slab gel and af- ter electrophoresis the gel was stained for SOD (see Materials and Methods).
ing for the enzyme's activity. Figure 3 shows the elec- trophoretic profiles of the SOD enzymes obtained from crude cell extracts, compared to human blood SOD as a control. No differences were observed between the SOD profiles obtained with extracts from Candida li- polytica 68-72 grown on glucose and on N-alkanes. Two clear bands and a minor one with different mobil- ities may be visualized. The control (human blood SOD) displayed two bands. After staining the gels with dianizidine in the presence of 2mM KCN, only the mi- nor band could be visualized, indicating that this en- zyme was an Mn-SOD type. Alkanes--and glucose-- grown Candida lipolytica 68-72 exhibited three bands, only two of which were inhibited by KCN.
DISCUSSION
Yeast strains of genus Candida utilizing N-alkanes have SOD and catalase activities incomparable as a whole to those of glucose-grown cells. Suppression of the SOD synthesis in the presence of glucose is due to the glucose effect. It could be effected by a catabolic repression or by a reduction of the concentration of the true inducer. 14 In the case of N-alkane utilization there was a positive correlation between the cell growth, SOD, and catalase activities of the culture. If the rate of SOD synthesis corresponds to the production rate of O2, a continuously increasing intracellular flux of O
would be generated during the oxidation of N-alkanes by yeasts.
As Strobell and Coon 15 suggested, superoxide is generated by molecular oxygen as an essential step in the hydroxylation of hydrocarbons in liver microsomes, and superoxide dismutase interferes with this reaction by lowering the concentration of this free radical. A possible interpretation of the increased SOD activity during hydrocarbon utilization by yeasts could be the scavenging of 0 2 produced by the hydrocarbon alco- hols oxidation system and the high rate of respir- ation. 5 The correlation between the SOD activity and the culture growth cycle supports the suggestion that superoxide dismutase may have a regulatory role in maintaining the low intracellular concentration of O 2 generated during the exponential growth of yeasts on hydrocarbons.
SOD, which takes part in the oxidation of N-alkanes by yeast cells, is of the Cu/Zn type. These results sup- port the hypothesis that the concentration of Cu/Zn SOD in the cells is modulated to correlate with the in- tracellular flux of O 2 generated during the growth of yeasts on N-alkanes. Such an effect has been observed during cultivation of yeasts on poor nutrient medium supplemented with methanol or ethanol as a carbon and an energy source, too . 16"17 These findings led to the conclusion that the yeast cells are capable to modulate their content of Cu/Zn SOD to meet the changing needs for scavenging of 0 2. However, the level of Cu/Zn SOD was manipulated by specific pathways of the car- bon sources mentioned above. So, the carbon source utilized by yeast cells may be considered an effective tool for manipulation of Cu/Zn SOD in these microor- ganisms. Therefore, these carbon sources may be ac- cepted as intracellular generators of O 2' whose specificity is due to their unique metabolism and correlate with the growth of the yeast cells.
SOD-enriched yeast cells obtained during cultivation on N-alkanes are a good model for studying cytotoxic- ity of oxygen free radicals because the presence of SOD normally prevented this effect. It has been already shown that the high activity of Cu/Zn SOD in yeast cells correlated with the increased resistance to photo- chemically generated O 2' During the photoreduction of riboflavine by L-methionine 90% from the oxygen free radicals obtained are represented as 0 2 and 10% as other active oxygen forms. 6 Thus, the death of the yeast cells in this system could be due predominantly to 0 2 , but other mechanisms may also take part. In such way a protective action of Cu/Zn SOD against O 2 was observed when SOD enriched yeast cells were tested in the system mentioned above.
All these data indicate that the increase in viability of hydrocarbon grown yeast cells could be probably due to effective scavenging of extracellular 0 2 by su-

268 A.V. KUJUMDZIEVA-SAVOVA et al.
peroxide dismutase. Thus, the high protection of SOD in 48 h grown culture on N-alkanes was demonstrated by 100% viability saving and budding of the cells after
20-min exposure to O 2" In the case of a glucose-grown
culture, a steady lethal effect could be observed. In order to investigate the potential role of SOD in
cell resistance to increasing concentrations of metal
ions, two test systems with different level of Cu/Zn SOD were exposed to increased concentrations of Cu 2 + and Zn 2+ ions. The results obtained clearly showed
that under the conditions of overproduction of Cu/Zn SOD, the resistance of yeast cells to high concentra- tions of Cu 2 + and Zn 2 + increased considerably. The
physiological effect of these ions on yeasts depends upon their essential role for the growth, on the one hand, and beneficial/deleterious balance that exists in
living cells, on the other. This balance is connected with the rate of metallothionein biosynthesis. Yeast, however, is known to produce metal-binding metal-
lothioneins (MT) in response to copper and zinc in the medium. 18 A steady state level of MT in the cells con-
fers a much higher metal resistance. The addition of Cu 2 +, Zn 2+, Cd 2+ and other transition metals to yeast
cells induces MT accumulation, 19 which binds and
detoxifies them. Since in our experiments one and the same concentration of Cu 2÷ and Zn 2 + ions in the nu-
trient medium exerts a growth inhibition only on glu- cose precultured cells, the possibility that an induction of MT biosynthesis is occurring as a single defensive mechanism lacks credence. These results confer the
suggestion that the copper and zinc tolerance in N-al- kane grown yeasts is due not only to common cellular components responsible for detoxification, but to the high level of SOD, too. These findings supported the hypothesis that a considerable part of metal toxicity is due to activation of nonspecific oxidative processes in the cells. As a consequence, a formation of active ox- ygen radicals occurs because of interaction of heavy metals with thyol compounds and pigment systems of the cells. 2° Because of such effect a higher production of active oxygen species may be occurring during ex- posure of yeast cells in increased concentrations of Cu 2÷ and Zn 2+ ions. Higher SOD activity of N-alkane grown cells required their much better survival than of
glucose-grown ones. The above data strongly suggest that SOD is in-
volved in the protection of some cellular components crucial to the overall process of cellular growth. As SOD apparently correlates with superoxide anion pro- duction, the high level of this enzyme could result from an increased production of this specific substrate dur- ing the metabolism of N-alkanes by yeasts and their active respiration.
Polyacrylamide gel electrophoresis of C a n d i d a li-
p o l y t i c a cell-free extracts have revealed the same chro- matic bands of SOD activity under all physiological
conditions tested. Inhibition by exposure to KCN indi- cates that SOD is of the Mn and Cu/Zn type. Undoubt- edly, the type of carbon source used for the cultivation
of yeast strain C a n d i d a l i p o l y t i c a 68-72 has had no in- fluence on the SOD profile of the cells.
Acknowledgements-The authors wish to thank Dr. Z. Avramova for critical reading of the manuscript.
REFERENCES
1. Tanaka, A.; Osumi, M.; Fukui, S. Peroxisomes of alkane-grown yeast: fundamental and practical aspects. Ann. N.Y. Acad. Sci. 386:186-199; 1982.
2. Boulton, C.; Ratlege, C. The physiology of hydrocarbon-utiliz- ing microorganisms. In: Topics in enzyme and Fermentation Biotechnology 9:10-77; 1984.
3. Fukui, S.; Tanaka, A. Metabolism of N-alkanes by yeasts. Adv. Biochem. Eng. 19:217-237; 1981.
4. Reum, H. J.; Reiff, I. Mechanism and occurrence of microbial oxidation of long-chain alkanes. Adv. Biochem. Eng. 19:175- 215; 1981.
5. Krauzova, B. I.; Ilchenko, H. P.; Sharishev, H. M; Lozinov, A. B. Possible pathways of oxidation of higher alcohols by membrane fractions of yeast grown on hexadecane and hexade- canol. Biochemistry 50:726-732; 1985.
6. Beauchamp, C.; Fridovich, I. Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Anal. Bio- chem. 44:276-287; 1971.
7. Aebi, H. In: Bergemeyer, H. V., ed. Methoden der Enzyma- tishen Analyse. Berlin: Acad. Verl.; 1970:634-641.
8. Somogy, M. Notes on sugar determination. J. Biol. Chem. 195: 19-23; 1952.
9. Nelson, N. A photometric adaptation of the Somogy method for the determination of glucose. J. Biol. Chem. 153:375-379; 1944.
10. Lowry, O. H.; Rosebrough, N. J.; Farr, O. L.; Randle, R. J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193:265-275; 1951.
11. Davis, B. J. Disc electrophoresis II. Method and application to human serum proteins. Ann. N.Y. Acad. Sci. 121:404-427; 1964.
12. Misra, H. P.; Fridovich, I. Superoxide dismutase and peroxi- dase: a positive activity stain applicable to polyacrylamide gel electrophoresis. Arch. Biochem. Biophys. 183:511-513; 1977.
13. Gregory, E. M.; Yost, E. J.; Fridovich, I. Superoxide dismu- tase in E.coli. Intracellular localization and function. J. Bacte- riol. 115:987-991; 1973.
14. Hassan, H. M.; Fridovich, I. Regulation and role of superoxide dismutase. Biochem. Soc. Trans. 573rd Meet. Hull. 6:356-361; 1978.
15. Strobell, H. W.; Conn, M. J. Effect of superoxide generation and dismutation of hydroxylation reactions catalyzed by liver microsomal Cytochrome P450. J. Biol. Chem. 246:7826-7829; 1971.
16. Kujumdzieva-Savova, A. V.; Savov, V. A.; Genova, L. K.; Peikova, S. P. Superoxide dismutase in methylotrophic yeasts. FEMF Microbiol. Lett. 127:103-105; 1985.
17. Kujumdzieva-Savova, A. V.; Savov, V. A.; Kehlibarova, L. I.; Georgieva, E. I. Influence of the carbon source on the activity of the superoxide-dismutase in yeasts. Compt. Rend. Acad. Bulg. Sci. 41:113-116; 1988.
18. Foulkes, E. C. ed. Biological role of metallothionein. New York: Elsevier; 1982.
19. Kagi, J. H. R.; Schaffer, A. Biochemistry of metallothionein. Biochemistry 27:8509-8515; 1988.
20. Naiki, N. Role of superoxide dismutase in copper-resistant strain of yeast. Plant Cell Physiol. 21:775-783; 1980.