responses to drought and flooding in tropical forage grasses
TRANSCRIPT

Plant and Soil 164: 97-105, 1994. © 1994 Kluwer Academic Publishers. Printed in the Netherlands.
Responses to drought and flooding in tropical forage grasses II. L e a f water potential, photosynthesis rate and alcohol dehydrogenase activity
Z d r a v k o B a r uch Departamento de Estudios Ambientales, Universidad Sim6n Bol(var, Apartado 89000, Caracas 1080, Venezuela
Received 19 January 1994. Accepted in revised form 3 May 1994
Key words: alcohol dehydrogenase, drought, flooding, photosynthesis, tropical grasses, water potential
Abstract
Long dry seasons or permanent flooding, typical of tropical savannas, severely limit the growth of pasture plants. This study compares the responses of water relations, carbon assimilation and alcohol dehydrogenase (ADH) activity to drought and flooding in four perennial Ca grasses: the tufted or caespitose Hyparrhenia rufa and Andropogon gayanus (CIAT 621) and the stoloniferous Brachiaria mutica and Echinochloa polystachya.
Plants of the four species were subjected to medium term flooding (20-25 days) and moderate drought in a greenhouse. Leaf water potential (W), stomatal conductance (Gs) and photosynthesis rate (Pn) were measured throughout the experiment and ADH activity was measured in flooded and control plants.
Moderate drought produced similar effects in all grasses reducing Gs which caused reduced Pn. Net photosynthesis compensation point was reached at the lowest W in A. gayanus which was considered as the most drought tolerant. The responses to flooding varied across species. Andropogon gayanus and H. rufa showed early stomatal closure without concurrent decrease in W and leaf turgor. This low Gs was responsible of reduced Pn and growth rate. There was a slight increase of Gs in the middle of the flooding period and both grasses recovered pre-stress Gs and Pn after drainage. ADH activity increased markedly only in A. gayanus under flooding suggesting that this grass was the most flood-sensitive. Stomatal aperture, Pn and ADH activity in B. mutica and E. polystachya were not affected by flooding. The higher flood-tolerance in these grasses might be attributed to enhanced oxygen diffusion to the roots through the hollow stolons, development of adventitious rootlets and large aerenchyma in the roots which compensate for the reduction of soil oxygen and permit the maintenance of root activity.
Introduction
Forage plants in tropical savannas may be subjected to broad shifts in water availability throughout the year, from severe drought to permanent flooding (Sarmien- to, 1984). If the selection and cultivation of new forage grasses in these regions is to succeed and be produc- tive, it seems advisable to previously know the poten- tial responses of these species to the particular envi- ronmental conditions of the savannas, specially to the prevailing water regime.
Plant water relations and gas exchange are influ- enced by the excess and deficit of water. Drought affects water relations and gas exchange in tropi-
cal forage grasses mainly through reduced leaf water potential causing reduced turgor, stomatal closure and reduced carbon assimilation rates. Several types of response, comprising evasion and tolerance to drought, permit the survival of the adapted forage grasses (Baruch and Fisher, 1991; Baruch et al., 1985; Ludlow, 1980; Wilson et al., 1980).
Another limitation in tropical savannas is seasonal or permanent waterlogging. The overriding effect of soil flooding is to limit the diffusion of oxygen to the root zone (Ponnamperuma, 1984). Hypoxia or anoxia are the main limiting factors that reduce aerobic respi- ration and the absorption of minerals and water by the roots. As root conductance is reduced by anoxia, wilt-

98
ing occurs, especially under high transpiration rates (Bradford and Hsiao, 1982). In non-tolerant species, stomata close soon after flooding and can remain closed or reopen afterwards which may or may not be associat- ed with leaf dehydration and reduction of the leaf water potential (Jackson and Drew, 1984; Sojka, 1992). Car- bon assimilation and translocation also are reduced in plants sensitive to flooding. This is attributed, initial- ly, to stomatal closure but subsequently, changes in nutrient availability, accumulation of assimilates and hormonal unbalance are important causes for the reduc- tion of photosynthesis. Also, in species not tolerant to root anaerobiosis, rapid metabolic changes occur that imply the activation of the enzyme alcohol dehydro- genase (ADH) that converts acetaldehyde generated in anaerobic respiration to ethanol that is less tox- ic to the plant (Crawford, 1982; Drew, 1992; Hook, 1984). In wetland plants, stomatal closure generally does not occur and root metabolism is not altered con- siderably by anoxia due to an efficient oxygen trans- port and diffusion system to the roots (Armstrong et al., 1991; Jackson and Drew, 1984). Although there is ample literature on the responses of crops and wet- land plants to waterlogging (Jackson and Hall, 1987; Naidoo and Mundree, 1993; Thomson et al., 1990; Wadmanvan Schravendijk and van Andel, 1985), stud- ies on the response of tropical forages to flooding are scarce (Anderson, 1974; Baruch and Fisher, 1991; Humphreys, 1980; Medina and Motta, 1990).
A previous work determined the effect of drought and flooding on biomass production and allocation, leaf growth rate and leaf nutrient concentration in four tropical forage grasses that differ in their respec- tive tolerance to drought and flooding (Baruch, 1994). The objective of this study is to analyze the mech- anisms of drought and flooding tolerance related to gas exchange, water relations and ADH activity in the same four C4 perennial forage grasses: Andro- pogon gayanus Kunth (CIAT 621) and Hyparrhenia rufa (Nees) Stapf. are tufted or bunch grasses that were more drought than flood tolerant whereas the stolonif- erous grasses Brachiaria mutica (H. B. K.) Hitchc. and Echinochloa polystachya (Forsk.) Stapf. were more flood than drought tolerant (Baruch, 1994).
Materials and methods
The biological attributes of the grasses under study as well as the general cultivation conditions were specified in a previous study (Baruch, 1994). Briefly,
drought was imposed by suspending irrigation until the plants showed wilting symptoms which occurred at a predawn leaf water potential (~) close to - 2.0 MPa (measured with a pressure bomb). At this point, the plants were re-watered. This cycle was repeated twice during the experiment. Flooding was imposed by inundating the plants up to 5 cm above soil surface and control plants were free-draining and watered dai- ly. Eight replicates were measured in each treatment at the end of which drought plants were watered and flooded plants were drained. The experiment contin- ued for 32 days forA. gayanus and H. rufa and 35 days for the other grasses.
Throughout the experiment ~ was determined in the youngest fully expanded leaf between 6 and 7 am. Later, between 10 am, and 12 am, and only when the density of the photosynthetically active photon flux (PPFD) was higher than 1,000 #mol. m -2. s -1 , stom- atal conductance (Gs), transpiration rate (E) and photo- synthesis rate (Pn) were measured with a portable gas exchange system (ADC, Model LCA-2 and Parkinson leaf chamber). The mean environmental conditions in the greenhouse are given in Baruch (1994).
Alcohol dehydrogenase (ADH) activity was mea- sured according to Bergmeyer (1974) on 3 replicates of flooded and control plants from a parallel experiment where flooding lasted for 8-10 days before the mea- surements. Two grams of young roots were homoge- nized in 10 mL of Tris buffer at pH 7.4 and centrifuged at 13,000 rpm during 20 min under 4 °C. Alcohol dehydrogenase activity was determined spectrophoto- metrically by measuring the reduction ofNAD + under 340 nm and the results are expressed by unit of root protein determined according to Bradford (1976).
In the appropriate cases, the differences between means were analyzed for statistical significance with an ANOVA followed by a Tukey HSD test of ranks. Statistical significance was assumed at p < 0.05.
Results
The imposed drought originated the typical decrease of in all grasses that began soon after water withdrawal
(Fig. 1). In all species, wilting was evident at k~ near - 2.0 MPa which recovered its pre-stress values after irrigation. The lowest • were recorded in A. gayanus. Flooding did not cause significant decrease of • in any species (Fig. 1).
In all grasses and during both drying cycles, Gs followed the pattern imposed by the fluctuations of
0 - -
-25
-3.5
-4.5
o
-05
-I
- 1 . 5
a)
o
~ -0.5
. J -1.5
-2.5
0
-0.5
",,,-
\ / \ / t / X //
I i i i i I i
,, / -,,,/ , / b
H. FIUFA b ' ~ 4
I i I I i i 1
a o a
\ / X / /
B. MUTICA I ~blk
i l - i i i i i / i
o o o 4 a
-~ \ / X\ /
\ i \ / -1.s \ / 5,l -z b~,~" "~'-. q b
-2.5 F_ POLYSTACHYA b
3 , , i , , , # ,
D a y s
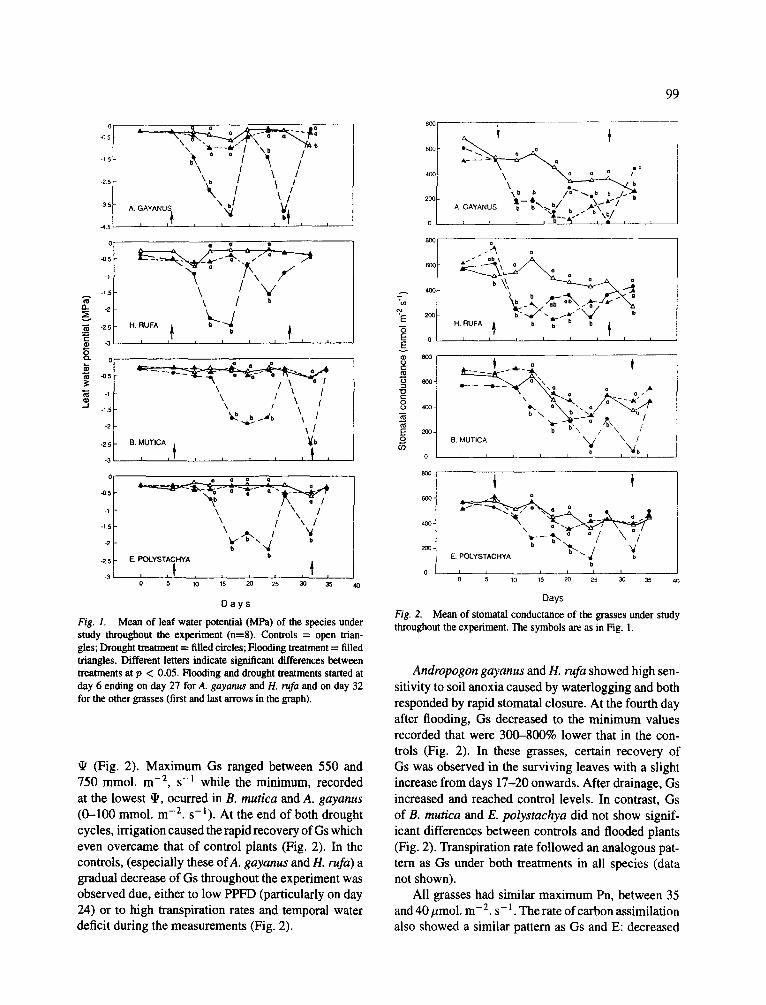
Fig. 1. Mean of leaf water potential (MPa) of the species under study throughout the experiment (n=8). Controls = open trian- gles; Drought treatment = filled circles; Flooding treatment = filled triangles. Different letters indicate significant differences between treatments at p < 0.05. Flooding and drought treatments started at day 6 ending on day 27 for A. gayanus and H. rufa and on day 32 for the other grasses (first and last arrows in the graph).
(Fig. 2). Maximum Gs ranged between 550 and 750 mmol. m -2, s -I while the minimum, recorded at the lowest tIs, ocurred in B. mutica and A. gayanus (0-100 mmol. m -2. s- l ) . At the end of both drought cycles, irrigation caused the rapid recovery of Gs which even overcame that of control plants (Fig. 2). In the controls, (especially these ofA. gayanus and H. rufa) a gradual decrease of Gs throughout the experiment was observed due, either to low PPFD (particularly on day 24) or to high transpiration rates and temporal water deficit during the measurements (Fig. 2).
99
E "6 E E
r-
8
E o 6O
800
6O(]
400
20£
0
800
6OO
400
20(
0
80O
400
200.
O
800
600-
400-
200.
0
, 4 < . ' / ,]L .-" kb/
A ~ ~ a b X \ a a a
I I ~ I I I I
b \ / N 1 B. MUTICA \ ~ / \ /
l i i l i bl i b i
1 i
* - - % . I I
b
0 5 0 5 20 25 30 35 40
Days
Fig. 2. Mean of stomatal conductance of the grasses under study throughout the experiment. The symbols are as in Fig. 1.
Andropogon gayanus and H. rufa showed high sen- sitivity to soil anoxia caused by waterlogging and both responded by rapid stomatal closure. At the fourth day after flooding, Gs decreased to the minimum values recorded that were 300-800% lower that in the con- trois (Fig. 2). In these grasses, certain recovery of Gs was observed in the surviving leaves with a slight increase from days 17-20 onwards. After drainage, Gs increased and reached control levels. In contrast, Gs of B. mutica and E. polystachya did not show signif- icant differences between controls and flooded plants (Fig. 2). Transpiration rate followed an analogous pat- tern as Gs under both treatments in all species (data not shown).
All grasses had similar maximum Pn, between 35 and 40/smol. m- 2. s - I. The rate of carbon assimilation also showed a similar pattern as Gs and E: decreased

100
°
\b 7 , \ , /
E "5 E :::L
j
I ! i I I i I I
¢ )
o (3. 10
0 i i i i i i i t
. . . . • a
v . ,,.-,,
\,~! b lO
F... P O L Y S T A C H Y A b
0 I i ~ I i I i 0 $ ~0 ~5 20 25 30 35 40
D a y s
Fig. 3. Mean of photosynthesis rate of the grasses under study throughout the experiment. The symbols are as in Fig. 1.
with the decrease of • under drought in all grasses but only in A. gayanus and H. rufa under flooding (Fig. 3). Exept in flooded A. gayanus, the instantaneous effi- ciency of water use (Pn/E) ranged between 2.0 and 3.5 x 10 -3 which indicates that stomatal closure caused by both treatments had equivalent effects on both pro- cesses. In A. gayanus under flooding Pn/E was c.a. 50 % lower than in the controls indicating a possible additional effect on Pn besides stomatal closure.
Drought affected analogously Gs and Pn and their relationship with q was non-linear (Fig. 4, only for Pn). The minimum values of Gs and Pn occurred at lower • in A. gayanus ( -3.5 to --4.0 MPa) than in the other grasses. Drought and flooding had similar effects on the relationship between Gs and Pn in all species (Fig. 5). They were related linearly until Pn saturation was reached at Gs c. a. 500--600 mmol. m -2. s - t
in all grasses under both treatments. There were not marked differences among the grasses under neither treatment. Andropogon gayanus was the only grass where flooding promoted significantly the activity of ADH, from 0.08 + 0.04 to 0.28 4- 0.01 #mol. mg root protein- 1. min- 1 which indicates that root metabolism in this species was the least tolerant to anoxia.
D i s c u s s i o n
Leaf water potential, carbon assimilation and ADH activity under drought and flooding varied across the species studied. Andropogon gayanus and H. rufa were appreciably more sensitive to waterlogging than B. mutica and E. polystachya. Considering the moderate drought stress applied, the four grasses showed similar responses although A. gayanus could be considered the most drought tolerant.
All grasses showed wilting symptoms at q near - 2.0 MPa accompanied by a perceptible reduction of Gs, E and Pn but all recovered after irrigation. This response to moderate water stress is typical of all plants (Baruch and Fisher, 1991; Bradford and Hsiao, 1982) and caused a marked decrease of total biomass (Baruch, 1994) due to Pn decrease. The anal- ogy between the relationship of Gs and Pn with suggests that the reduction of Pn was mediated chiefly by stomatal closure.
Andropogon gayanus and H. rufa are considered as drought tolerant grasses (Daubenmire, 1972; Tole- do and Fisher, 1989), more than B. mutica and E. polystachya as evidenced in a previous work (Baruch, 1994). The higher drought tolerance of A. gayanus could be attributed to its positive Pn rate under lower
than in the other species. However, this was not the case of H. rufa where other explanations are needed to sustain its reputation as drought tolerant.
There are few studies on the effects of flooding on the water relations and carbon assimilation in non- crop grasses (Pezeshki et al., 1988; Medina and Motta, 1990; Naidoo and Mundree, 1993) and none yet on tropical forage grasses. The results of this study show a clear contrast between the responses of species con- sidered flood-tolerant as B. mutica and E. polystachya and the non-tolerant A. gayanus and H. rufa. The dra- matic reduction of Gs under flooding in the latter is one of the earliest responses of dryland plants to flooding. This caused the steep reduction of Pn with a conse- quent decrease in total plant biomass (Baruch, 1994). However, stomatal closure occurred under high leaf

H. RUFA
Leaf water potential (MPa)
I I I
0 B. MUTICA
0
l
l l l -‘\, l ’
l
l e l e l
l :e -1 -2 -3 -
Leaf water potential (MPa) Fig. 4. Response of photosynthesis rate to leaf water potentials in the drought treatment. Lines were fitted by quadratic regression. The coefficients are: A. gayanus y = 33.4 - 16.62 + 2.0z2R2 = 0.84; H. rufa y = 46.9 - 33.5x + 5.9z2R2 = 0.73; B. mutica y = 39.3 - 25.1s + 4.4~~ + 4.4x2R2 = 0.80: E. potystachya y = 34.6 - 14.5x + 0.6x2R2 = 0.76.
turgor and q and there is agreement that stomata1 clo- increases the survival possibilities under flooding due sure under these conditions is caused by abscisic acid to reduced water and nutrient demands to the water- (ABA) accumulation but uncertainity persists about logged roots and a reduced absorption of the potential- whether this hormone originates in the roots or in the ly toxic reduced ions Fe and Mn (Jackson and Drew, leaves (Jackson andHall, 1987). Early stomata1 closure 1984). Also, if oxygen enters through the stomata,

102
A
'ttJ
E O E
t -
t , -
O O t - D.
50
40
30
20
10
0
I I " I I
• 0 0
0 7 Q • • / O I •
i/'" / H.RUFA (Drought)
/ , , e
f e l I I l 200 400 600 800
I I i
(3O
o qoP ~ O0 O/~0
O 0 ~/ 0
Oo /.o o o
o o H.RUFA (Flooded) O O
O
i O .c
O
O
I I I I
0 200 400 600 800 100C
Stomatal conductance (mmol m "2 s "1)
.__. 50
E 40 0 E
30
50
20 e-
N 10 0 e-
I I i I
b ° • O ~ , ~ " " ' ~ r ' , - , , , ~ I"
"/"
A. GAYANUS (Drought)
. - , , , ,
0 200 400 600 800 0
I I I I
?
aa 0 ~.-,-- ~ ~ "~,~0
o o.,~o 0 / 0
g ~ , b o Oop
O /
o / 8 0 A. GAYANUS (Flooded)
O
O I I I I
200 400 600 800 100C
Stomatal conductance (mmol m "2 s "1)
Fig. 5(a)-(b). Responses of photosynthesis rate to stomatal conductance under drought and flooding treatments (only drought for B. mutica and E. polystachya). All curves were fitted by quadratic regressions.
(a) H. rufa
(b) A. gayanus
(c) B. mufca (d) E. polystachya
Drought y = - 4 . 1 + 0 .13z - 0.01x2; R 2 = .84
Floodingy = - 8 . 6 + 0 .05z - 0.01x2; R 2 = .78
Drought y = - 1 . 4 + 0 .10x + 0.01x2; R 2 = .88
Floodingy = - 3 . 9 + 0 .10x - 0 .01z2; R E = .84
Drought y = 0.9 + 0 .10x - 0.01x2; R 2 = .95
Drought y = 0.2 + 0 .11x - 0.01x2; R E = .83

0 I I I I
¢ *59
40 • • - o _
E w I I ~ o
~=- 30 o O O O ~ e " ' ' " "
_~ 20 c~
+ ) . N ¢-
~'10 •
o ~ E. POLYSTACHYA (Drought) t ' - 13..
I 1 I I
200 400 600 800
d I I I I
O0 ° 0 O 0
[ , . ' / . " . .
I ~ B.MUTICA (Drought)
I " I 1 I I
0 200 400 600 800 100C
103
Stomatal conductance (mmol m "2 s "1)
Fig. 5(c)-(d)+
their closure in non-tolerant species may aggraviate root anoxia if they depend on internal aeration (Hook, 1984).
The partial stomatal opening observed in A. gayanus and H. rufa from day 17 onwards could have been caused by the simultaneous emergence of adven- titious roots in the latter (Baruch, 1984). The genera- tion of adventitious roots is considered as one of the most effective responses to waterlogging that allevi- ates the effects of anoxia by supplying oxygen to the roots and resuming limited absorption of water and mineral nutrients (Jackson and Drew, 1984; Kawase, 1981; Naidoo and Mundree, 1993; Wenkert et al., 1981). However this cause - effect relationship can not be invoked in A. gayanus which lacked adventi- tious roots. Perhaps, the simultaneous development of aerenchymatous tissue in the roots (up to 25% of root cross sectional area; M6rida and Baruch, unpublished results) was able to relieve oxygen deficiency and per- mited partial stomatal aperture.
Only A. gayanus, the species most affected by anoxia, showed a significant induction of ADH activity as a metabolic response to waterlogging which is typ- ical of flood intolerant plants (Crawford, 1982; Drew, 1992; Hook, 1984; Medina and Motta, 1990). Alcohol dehydrogenase activity indicates that morphological and metabolic adaptations, induced by waterlogging, were inadequate to provide sufficient oxygen for aer- obic respiration. Although the activity of ADH cannot
be taken as a direct quantitative measurement of the degree of anaerobiosis, it is one of the limited num- ber of proteins synthetized in hypoxic plant tissues (Drew, 1992). The lack of ADH activity in H. rufa suggests that its roots were under less severe anaero- bic conditions probably caused by the development of adventitious rootlets.
In the flood-tolerant B. mutica and E. polystachya, neither a decrease in Gs and Pn nor inducement of ADH activity under flooding was detected. In these grasses oxygen for the roots was probably supplied by a combination of the rapid growth of adventitious roots, the extensive amount of hollow culms (Baruch, 1994) and the generation of large aerenchymatous spaces up to 60% of root cross sectional area (M6rida and Baruch, unpublished results). These responses are typical of wetland plants (Burdick and Mendelssohn, 1990; Jackson and Drew, 1984; Justin and Armstrong, 1987; Medina and Motta, 1990; Naidoo and Mundree, 1993). Nevertheless, total yield was reduced even in these flood-tolerant plants (Baruch, 1994) as found in other wetland grasses, such as Echinochloa phyllo- pogon (Everard et al., 1991) and Sporobolus virginicus (Naidoo and Mundree, 1993) under severe soil anoxia when adventitious roots and aerenchyma may be still unable to supply enough oxygen to fully support aero- bic respiration which is inadequate for growth require- ments. (Burdick and Mendelsshon, 1990; Yu et al., 1969).

104
The analogy between stomatal response to drought in all grasses, to flooding in A. gayanus and H. rufa and the analogy of the relation between Gs and Pn under drought and flooding suggest that the respons- es to these stresses may be basically similar. Because ABA has been related to stomatal closure caused by either soil drought or oxygen deficiency (Blackman and Davies, 1985; Davies and Zhang, 1991) it has been proposed as the mediator in a centralized response system that, through early reduced growth, reduces the damage caused to the plant by these environmen- tal stresses (Chapin, 1991; Davies and Zhang, 1991; Jackson, 1993).
Acknowledgements
This work was supported by the National Council of Scientific and Technological Research of Venezuela (CONICIT) through Project S 1-1710. I wish to thank the Director and personnel of the Nticleo del Litoral of Universidad Sim6n Bolfvar for their help in the instal- lation and maintenance of the greenhouse. Also, I thank Pl~isticos Guarenas for providing the special polyethy- lene for greenhouse roofing. Liz Izquierdo performed the determinations of enzymatic activity at the labora- tory facilities and with reactives generously provided by Drs Z SuArez-Mata, E Medina and S Bahar. Arrigo Luchetti collaborated in the experiments. I thank two anonymous reviewers that improved the manuscript.
References
Anderson E R 1974 The reaction of seven Cenchrus ciliaris L. cultivars to flooding. Trop. Grassl. 8, 33-123.
Armstrong W, Justin S H F W, Beckett P M and Lythe S 1991 Root adaptation to soil waterlogging. Aquatic Bot. 39, 57-73.
Baruch Z 1994 Responses to drought and flooding in tropical forage grasses: 1. - Production and allocation of biomass, leaf growth and mineral nutrients. Plant and Soil 164, 87-96.
Baruch Z and Fisher M J 1991 Factores clim~ticos y de competencia que afectan el desarrollo de ias plantulas de las especies forrajeras. In Establecimiento y renovaci6n de pasturas. Eds. C E Lascano and J M Spain. pp 103-142, C.entro internacional de Agricultura Tropical. Cali, Colombia.
Baruch Z, Ludlow M M and Davis R 1985 Photosynthetic responses of native and introduced C4 grasses from Venezuelan savannas. Oecologia 67, 288-293.
Blackman P G and Davies W J 1985 Root to shoot communication in maize plants on the effects of soil drying. J. Exp. Bot. 36, 39-48.
Bergrneyer H U 1974 Methods of enzymatic Analysis. Vol. 4. Aca- demic Press. New York.
Bradford K J 1983 Involvement of plant growth substances in the alteration of leaf gas exchanges of flooded tomato plants. Plant Physiol. 73, 480-483.
Bradford K J and Hsiao T C 1982 Stomatal behavior and water relations of waterlogged tomato plants. Plant Physiol, 70, 1508- 1513.
Bradford M 1976 A rapid and sensitive method for the quantifica- tion of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 72, 248-258.
Burdick D M and Mendelssohn I A 1990 Relationship between anatomical and metabolic responses to soil waterlogging in the coastal grass Spartina patens. J. Exp. Bot. 41,223-228.
Chapin F S 1991 Integrated responses of plants to stress. BioScience 41, 29-36.
Crawford R M 1982 Physiological response to flooding. In Physio- logical Plant Ecology II. Encyclopedia of Plant Physiology. New Series. Vol. 12B. Eds. O L Lange, P S Nobel, C B Osmond and H Ziegler. pp 453-477. Springer-Verlag. Berlin.
Danbenmire R 1972 Ecology of Hyparrhenia rufa Nees in derived savanna in NW Costa Rica. J. Appl. Ecol. 9, 11-23.
Davies W J and Zhang J H 1991 Root signals and the regulation of growth and development of plants in drying soil. Ann. Rev. Plant Physiol. Plant Mol. Biol. 42, 55-76.
Drew M C 1992 Soil aeration and plant root metabolism. Soil Sci. 154, 259-268.
Everard J D, Lecain D R, Rumpho M E and Kennedy R A 1991 Mesictyl root formation in Echinochloa phyllogopon (Poaceae) in relation to root zone aeration. Am. J. Bot. 78, 462-469.
Hook D D 1984 Adaptations to flooding with fresh water. In Flooding and Plant Growth. Ed. T T Kozlowski. pp 265-294. Academic Press, New York.
Humphreys L R 1980 Environmental Adaptation of Tropical Pasture Plants. Macmillan, New York.
Jackson M B 1993 Are plant hormones involved in root to shoot communication? Adv. Bot. Res. 19, 103-187.
Jackson M B and Drew M C 1984 Effects of flooding on growth and metabolism of herbaceous plants. In Flooding and Plant Growth. Ed. T T Kozlowski. pp 47-128. Academic Press, New York.
Jackson M B and Hall K C 1987 Early stomatal closure in water- logged pea plants is mediated by abscisic acid in the absence of foliar water deficits. Plant, Cell Environm. 10, 121-130.
Justin S H and Armstrong W 1987 The anatomical characteristics of roots and plant response to soil flooding. New Phytol. 106, 465-496.
Kawase M 1981 Anatomical and morphological adaptation of plants to waterlogging. Hortscience 16, 8-12
Ludlow M M 1980 Stress physiology in tropical pasture plants. Trop. Grassi. 14, 136--145.
Medina E and Motta N 1990 Metabolism and distribution of grasses in tropical flooded savannas in Venezuela. J. Trop. Ecol. 6, 77-89.
Naidoo G and Mundree S G 1993 Relationships between morpho- logical and physiological responses to waterlogging and salinity in Sporobolus virginicus (L.) Kunth. Oecologia 93, 360-366.
Pezeshki S R, DeLaune R D and Lindau C W 1988 Interaction among sediment anaerobiosis, nitrogen uptake and photosynthesis of Spartina alterniflora. Physiol. Plant. 74, 561-565.
Ponnamperuma E N 1984 Effects of flooding on soils. In: Flooding and Plant Growth. Ed. T T Kozlowski. pp 265-294. Academic Press, New York.
Sarmiento G 1984 The ecology of Neotropical savannas. Harvard Univ. Press, Cambridge, Mass.
Sojka R E 1992 Stomatal closure in oxygen stressed plants. Soil Sci. 154, 269-280.

Thomson C J, Armstrong W, Waters I and Greenway H 1990 Aerenchyma formation and associated oxygen movement in sem- inal and radial roots of wheat. Plant Cell Environm. 13, 395-404.
Toledo J M and Fisher M J 1989 Aspectos fisiol6gicos de Andro- pogon gayanus y su compatibilidad con leguminosas forrajeras. In Andropogon gayanus Kunth un pasto para suelos ~icidos del tr6pico. Eds. J M Toledo, R Vera and C Lascano. pp 69-104. CIAT. Cali, Colombia.
Wadman-van Schravendijk S H and van Andel O M 1985 Inter- dependence of growth water relations and absicic acid level in Phaseolus vulgaris during waterlogging. Physiol. Plant. 63, 215- 220.
105
Wenkert W, Fausey N R and Watters H D 1981 Flooding responses in Zea mays L. Plant and Soil 62, 351-366.
Wilson J R, Ludlow M M, Fisher M J and Schulze E D 1980 Adap- tation to water stress of the leaf water relations of four tropical forage species. Aust. J. Plant Physiol. 7, 207-220.
Yu P T, Stolzy L H and Letey J 1969 Survival of plants under prolonged flooded conditions. Agron. J. 61,844-847.
Section editor: H Lambers