responses of sphagnum species to atmospheric nitrogen and sulphur deposition
TRANSCRIPT

Botanical Journal of the Linnean Society (1990), 104: 255-265. With 6 figures
Responses of Sphagnum species to atmospheric nitrogen and sulphur deposition
J. A. LEE, R. BAXTER
Department of Environmental Biology, The University, Manchester M 1 3 9PL
AND
M. J. EMES
Department of Cell and Structural Biology, The University, Manchester M 1 3 9PL
LEE, J. A., BAXTER, R. & EMES, M. J., 1990. Responses of Sphagnum species to atmospheric nitrogen and sulphur deposition. The responses of Sphagnum species to atmospheric pollution by sulphur and nitrogen oxides are reviewed. An explanation of the tolerance of sulphur dioxide in Sphagnum cuspidaturn is provided. This involves a n exclusion mechanism by the oxidation of bisulphite ions to less toxic sulphate ions catalysed by the presence of iron and possibly other metals on the cell wall cation-exchange sites. Tolerance is influenced by changing the iron content of the plants. A similar mechanism may account for sulphur dioxide tolerance in Sphagnum recurvum.
Evidence for a close coupling of nitrate assimilation with the atmospheric nitrogen supply in ombrotrophic Sphagnum species in unpolluted regions is reviewed, and this coupling is shown to be lost in polluted environments with high atmospheric nitrogen supply. The potential effects of a supra-optimal atmospheric nitrogen supply to Sphagnum species on the ecology of mires is discussed.
The effects of nitrogen and sulphur pollutants on Sphagnum species are used to make broad generalizations as to how other bryophyte species might respond to these pollutants and to explain known effects of atmospheric pollution on bryophyte communities.
ADDITIONAL KEYWORDS:- Atmospheric pollution - nitrate assimilation - nitrate deposition - SO, tolerance.
CONTENTS Introduction . . . . . . . . . . . . . . . . . . . 255 An explanation of SO, tolerance in Sphagnum . . . . . . . . . . . 256 Responses to nitrogen deposition in Sphagnum . . . . . . . . . . . 260 Discussion. . . . . . . . . . . . . . . . . . . . 263 References . . . . . . . . . . . . . . . . . . . 264
INTRODUCTION
Bryophytes are amongst the most sensitive plant species to atmospheric pollutants, but despite several decades of research our knowledge of the physiological responses of bryophytes to pollution is extremely limited (see e.g.
255 0 1990 The Linnean Society of London 0024-4074/90/090255 + 1 I $03.00/0

256 J. A. LEE E T AL.
Rao, 1982). Much circumstantial evidence from field surveys and transplant experiments has been accumulated to suggest that bryophyte species differ in their sensitivity to atmospheric pollution (see e.g. Gilbert, 1968, 1970; Rao & Leblanc, 1967). However, very few fumigation experiments have clearly demonstrated such differences at realistic concentrations of atmospheric pollutants (see e.g. Ferguson, Lee & Bell, 1978), and there have been no experimental investigations which unequivocally demonstrate how a species may survive on one substrate in a given region and pollution climate but not on another (see Rao, 1982 for examples).
In Britain, sulphur dioxide has been the major gaseous atmospheric pollutant for two centuries (see e.g. Smith, 1872), and the high concentrations of the gas which were once prevalent in the industrial towns and their surrounding rural regions (Ferguson & Lee, 1983) are most likely to have had profound effects on the bryophyte populations (Press, Ferguson & Lee, 1983). Although most historical records of vegetation changes as the putative result of atmospheric pollution are complicated by the subjectivity and approaches of different recorders, peatlands provide a means of examining vegetation change within an ecosystem without such constraints-since macrofossils preserved in the peat may allow changes in the abundance of bryophytes over recent centuries to be precisely determined. The southern Pennines of England are covered by an extensive series of blanket mires extending to greater than 50 000 hectares. This region is surrounded by towns which were early centres of the Industrial Revolution, and some of which were shown by Smith (1872) to be heavily polluted with sulphur dioxide and its solution products in the 19th century. These blanket mires can be shown to have peat in which the remains of Sphagnum species and other bryophytes are prominent, but the present-day vegetation is largely devoid of bryophyte cover. Tallis (1964) was able to correlate the disappearance of the once dominant Sphagnum species with the appearance of soot in these deposits, thus implicating atmospheric pollution as the cause of this widespread and important vegetation change. Ferguson et al. (1978) were able to demonstrate from a knowledge of the pollution climate that this change was most probably caused by sulphur dioxide since fumigation studies showed the sensitivity of Sphagnum species to this gas and its solution products. They also showed that the most widespread species in the region today, Sphagnum recuruum P. Beav., a minerotrophic species, was the most resistant to the gas. However, although they were able to show a rapid effect of bisulphite ions on the photosynthetic oxygen evolution of all species tested (Ferguson & Lee, 1979), a mechanism which could act as the basis of differences in species sensitivity to sulphur dioxide was not apparent.
AN EXPLANATION OF SO, TOLERANCE IN SPHAGNUM
The ability to demonstrate intra-specific differences in pollutant sensitivity provides a means of examining the precise causes of pollution tolerance. Studholme ( 1989) showed that southern Pennine populations of Sphagnum cuspidatum Hoffm. are apparently more tolerant of bisulphite ions than those populations of the species from regions without a long history of sulphur dioxide pollution (Fig. 1). The ability of a species to grow in a region with high

SPHAGNUM RESPONSES TO NITROGEN AND SULPHUR 257
M o r t Moor Berwyne
Ringinglow Butterburn
k 0 0.2 0.3 0 0.2 0.3
mM bisulphite
Figure 1. The growth (increase in length, ';A control in artificial rainwater) of Sphagnum cuspidaturn populations from the polluted southern Pennines (Alport Moor and Ringinglow j and from relatively unpolluted sites (Berwyn Mountains and Butterburn Flowej in artificial rainwater solutions containing a range of bisulphite concentrations.
concentrations of sulphur dioxide must result from either the avoidance of the gas and its toxic solution products (dissolved and unreacted sulphur dioxide and bisulphite ions) or from the ability of the plant to tolerate these solution products within its cells, or from some combination of these two phenomena. Although there is considerable interest in the mechanisms of tolerance to sulphur dioxide in higher plants, there are few data which show the importance of metabolic tolerance in this process. However, the scope for avoidance mechanisms in bryophytes is much more limited than in higher plants because many bryophyte species have leaves lacking cuticles in which the photosynthetic cells are directly and continuously exposed to the atmosphere. Avoidance mechanisms, if present, are therefore likely either to operate a t the cell wall or to involve the rapid conversion of the immediate sulphur dioxide solution products into less toxic or volatile forms.
Baxter, Emes & Lee (1989) have examined the former possibility in Sphagnum cuspidatum. The long-chain polymers containing uronic acids in the cell walls of Sphagnum species have long been recognized as the source of the important cation exchange capacity of these mosses (see e.g. Clymo & Hayward, 1982, for details) and these exchange sites play an important role in the mineral nutrition of the plants. Higher valence ions are taken up more readily by these sites than lower valence ions (Touffet, 1972), and ions which have been shown to be taken up include Fe(II1) and Mn(I1). These ions in solution are very active catalysts of the oxidation of aqueous sulphur dioxide or sulphur (IV) species such as bisulphite ions. Sulphate is much less toxic to Sphagnum than sulphur (IV) species (Ferguson et al., 1978), and thus if the concentrations of Fe(III) , Mn(I1) and

258
120
- c & loo- .- 0) 3 E 0 -
80- - z i 9 60-
E
2 4 0 -
- 0)
u)
c -
I-“
20
J. A. LEE ET AL.
I 4 O l -
- 1, - Copi t u b 0 -2cm 0-4 cm
Shoot segment
Figure 2. Total tissue Fe concentrations in segments of apical lengths of Sphagnum cuspidalum from a polluted site (open bars) and an unpolluted site (hatched bars). Vertical bars represent f I SE, n = 3. Data from Baxter, Emes & Lee (1989).
possibly other cations (e.g. Cu(I1)) on or around the cell walls of the mosses is sufficiently high, then it is possible that this could represent a sulphur dioxide avoidance mechanism for these mosses.
Figure 2 shows the iron content of stems of two populations differing in apparent tolerance to bisulphite (Fig. 1 ) . The southern Pennine population has a much higher total iron content than the ‘unpolluted’ population and Baxter et al. (1989) have shown that much of this iron is associated with the cell wall exchange sites. Under laboratory conditions bisulphite ions disappeared much more rapidly from a solution bathing southern Pennine Sphagnum cuspidatum than from a solution bathing the ‘unpolluted’ population of this species (Fig. 3) , and sulphate appeared more rapidly in the former bathing solution (Fig. 4). At the concentration of bisulphite used above, plants from the ‘unpolluted’ population died after seven days whereas those from the southern Pennines survived, and this is probably the result of the chlorophyllous cells of plants from the former population being exposed to high concentrations of bisulphite for long periods of time (fresh bisulphite solutions were supplied every second day). Importantly, the differential sensitivity of the populations to bisulphite could effectively be removed by two 15-minute washes of the plants from the two populations in 5 mM EDTA (Baxter et al., 1989). This treatment had no effect on the subsequent growth of the plants in an artificial rainwater solution, but growth was similarly strongly inhibited in both populations in the presence of bisulphite. Thus the apparent difference in tolerance was not due to genotypic differences in the Sphagnum cuspidatum populations but due to differences in cation loading on the plants in the two regions. The higher exchangeable metal content of southern
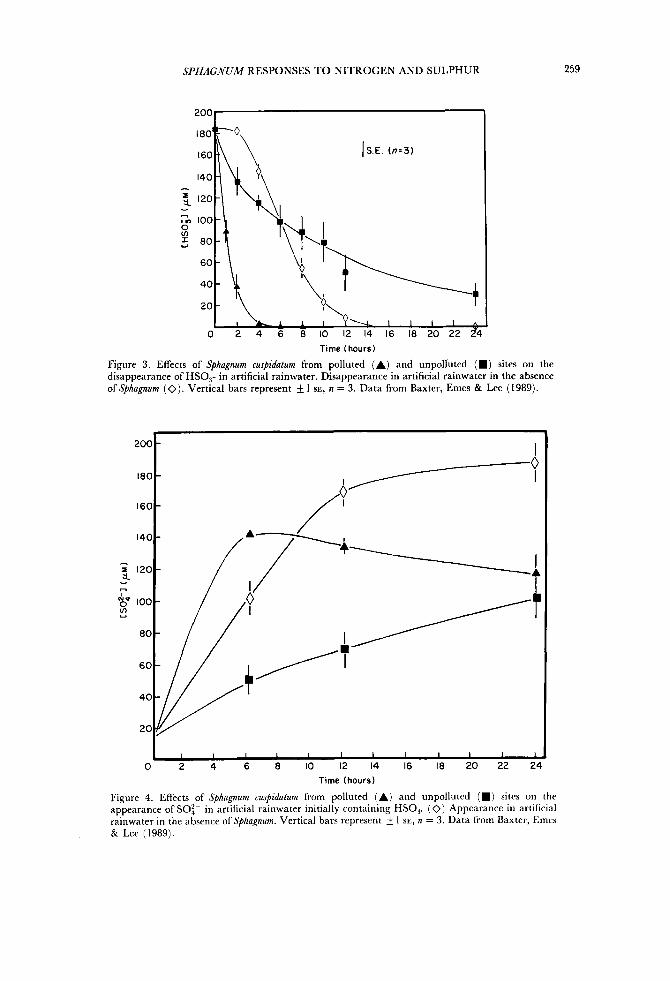
SPHAGNUM RESPONSES TO NITROGEN AND SULPHUR 259
Figure 3 . Effects of Sphagnum cuspidaturn from polluted (A) and unpolluted (m) sites on the disappearance of HS0,- in artificial rainwater. Disappearance in artificial rainwater in the absence of Sphagnum (0). Vertical bars represent 5 1 SE, n = 3. Data from Baxter, Emes & Lee (1989).
I I I I I I I I I I I I I
0 2 4 6 8 10 12 14 16 18 20 22 24 Time (hours)
Figure 4. Effects of Sphagnum cuspidaturn from polluted (A) and unpolluted (m) sites on the appearance of SO:- in artificial rainwater initially containing HSO,. (0) Appearance in artificial rainwater in the absence of Sphagnum. Vertical bars represent & 1 SE, n = 3. Data from Baxter, Emcs & Lee (1989).

260 J. A. LEE ET AL.
Pennine plants probably is the result of the long history of coal smoke pollution to which the mires in this region have been exposed. The surface peat contains very high contents of a number of metals including iron. In support of this explanation, the oxidation of bisulphite in a solution bathing plants from the ‘unpolluted’ population could be accelerated either by prior growth of the plants in an artificial rainwater solution enriched with 1 pg cm-3 FeCl, or by growth for 8 weeks in a southern Pennine bog pool (Baxter et al., 1989), and these treatments also increased the survival of plants in bisulphite solutions under laboratory conditions. Thus in these two populations at least, the differences in bisulphite tolerance are ’more apparent than real, and the exchangeable cation composition of the cell wall appears to be the critical factor in tolerance.
I t is unlikely that this explanation of the difference in tolerance to sulphur dioxide solution products is unique to these two populations of Sphagnum cuspidatum. One of the more remarkable features of the southern Pennines is that the only places where Sphagnum species appear to have thrived continuously since the Industrial Revolution are in the minerotrophic flushes. Sphagnum recuruum is the predominant species in these flushes, and has been shown to be more tolerant of sulphur dioxide in a fumigation experiment than several ombrotrophic species (Ferguson et al., 1978). But minerotrophic species have higher element concentrations than ombrotrophic species, and it could be again that the inter- specific differences in Sphagnum sensitivity to sulphur dioxide are more apparent than real. Although Sphagnum recurvum is predominantly a minerotrophic species, it can sometimes be found growing in ombrotrophic situations. Figure 5 compares the oxidation of bisulphite in solutions bathing S. recurvum taken from an ombrotrophic mire in the northern Pennines and from its minerotrophic margin. The plants from the minerotrophic site have a higher tissue iron content (7.9f0.7 pg dwt) compared to those at the ombrotrophic site (3.6rfr0.2 pg gdwtf 1). The former display an enhanced ability for the rapid oxidation of bisulphite and survive the bisulphite treatment. Thus, again it would appear that the cation content of the plants is critical to their tolerance of sulphur dioxide. I t is interesting to note that Ferguson & Lee (1979) showed that despite differences in the growth of Sphagnum species in atmospheres containing sulphur dioxide, there were no differences between species in the short term response ( < 5 min) of photosynthetic oxygen evolution or respiration to bisulphite ions. These experiments were conducted by the injection of bisulphite ions into aqueous medium in a well-stirred oxygen electrode chamber. Under these conditions, the exposure of the chlorophyllose cells to bisulphite may not have been influenced appreciably by the ‘normal’ cell wall phenomena, and thus differences between species were not apparent.
RESPONSES T O NITROGEN DEPOSITION IN SPHAGNUM
In recent decades, the importance of sulphur dioxide as an atmospheric pollutant in Britain has declined (see e.g. Ferguson & Lee, 1983), but the oxides of nitrogen have become more important. Thus in the southern Pennine region, nitrate deposition may have quadrupled since 1868 (Lee et al., 1987). After carbon, hydrogen and oxygen, nitrogen is the element required in greatest quantities by plants for growth. Concentrations of nitrate and ammonium in the precipitation of unpolluted regions are usually extremely low, and thus it could

SPHAGNUM RESPONSES TO NITROGEN AND SULPHUR 26 1
Time (hours)
Figure 5. Effects ofSphasnum recumurn from an ombrotrophic (m) and a minerotrophic (A) region of an unpolluted mire on the disappearance of HSO; in artificial rainwater. Vertical bars represent f 1 SE, n = 3.
be expected that ombrotrophic species of bryophytes would be adapted to utilize maximally the atmospheric supply. Woodin & Lee (1987a) showed that in a subarctic mire remote from pollution sources Sphagnum fuscum (Schimp.) Klinggr. plants apparently were able to immobilize the atmospheric nitrogen supply totally, and that if the throughflow contained nitrate and ammonium ions, then these were below the limits of detection. Woodin, Press & Lee (1985) were able to demonstrate that nitrate assimilation by Sphagnum furcum at this site was closely coupled to the atmospheric supply. Figure 6 shows this relationship for part of the summer of 1985. Nitrate ions in each precipitation event induce the activity of the primary assimilation enzyme nitrate reductase. This relationship has been demonstrated in four years at this site, and Woodin & Lee (1987b) were able to show that the maximum induced activity and the total induced activity were proportional to the log nitrate deposition. Thus, in sites remote from urban and industrial activity it would appear that in ombrotrophic Sphagnum species there is a very close coupling of nitrate uptake and assimilation with the atmospheric nitrate supply. However, Lee et al. (1987) showed that this coupling could be rapidly lost when Sphagnum species are transplanted from a relatively unpolluted to a polluted environment. Sphagnum cuspidaturn was transplanted from a Welsh mire into artificial bog pools in the southern Pennines

262
c o .- 0 0 .6 -
gz 3 IE 0.4-
c
T 5 0.2- Ic)
J. A. LEE E T AL.
- - -
4
z a L
receiving only an atmospheric supply of solutes. The plants rapidly accumulated total tissue nitrogen and lost their ability to show induction of nitrate reductase. Further Sphagnum cuspidatum indigenous to the southern Pennines showed no induction of nitrate reductase activity with deposition events, this aspect of metabolism being effectively uncoupled from the atmospheric supply. This results in the plants being less able to immobilize the atmospheric nitrogen supply (Press, Woodin & Lee, 1986). These workers concluded that the present supply of atmospheric combined nitrogen in the southern Pennines was supra- optimal for the growth of ombrotrophic Sphagnum species, and was contributing to the poor growth of these species in the region today.
Under unpolluted conditions the ombrotrophic Sphagnum species provide an almost perfect sink for the atmospheric combined nitrogen supply, capturing the nitrate and ammonium ions directly deposited onto the moss carpet and probably also assimilating much of the supply initially intercepted by the shoots of the angiosperms once this has percolated to the moss capitula. Thus it may be concluded that the Sphagnum carpet effectively controls the availability of nitrogen for the growth of higher plants since under unpolluted conditions nitrogen is only made available following the decomposition of the dominant Sphagnum species. However, if in polluted regions this Sphagnum sink becomes effectively saturated, then the atmospheric nitrogen supply becomes more available to the higher plants, and further, the rate of decay of dead Sphagnum material may be accelerated by its higher tissue nitrogen composition (Clymo & Hayward, 1982). The net result may be to stimulate the growth of the angiosperms and to change the competitive relationships between species in the mires. Thus, perhaps subtle changes in the ecology of ombrotrophic mires are

SPHAGNUM RESPONSES TO NITROGEN AND SULPHUR 263
occurring as the result of the widespread increase in the supply of atmospheric combined nitrogen even in regions relatively remote from major sources of atmospheric nitrogen pollutants.
DISCUSSION
A major question arising from the investigation of the relationship between Sphagnum species and their atmospheric supply of sulphur and nitrogen is to what extent are these responses unique to this genus or typical of bryophyte species in general?
The ombrotrophic nature of terrestrial bryophytes has been questioned (see e.g. Brown, 1982), and in terms of the acquisition of nitrogen and other elements from the atmosphere it could be expected that there would be considerable variation between species. Thus, endohydric species with leaf surfaces that are water repellent, and which take up water from the substratum and conduct it internally to the leaves might be expected to behave rather differently from ectohydric species which absorb water readily over their leaf surfaces. Indeed, when precipitation directly penetrates to the surface of the substratum its chemical composition will be to some extent modified by the activity of microorganisms and by ion exchange and solution processes. The degree of modification will vary from substrate to substrate and with, for example, the intensity of the precipitation and temperature. I t is quite possible that even in endohydric species the response of species to atmospheric nitrogen deposition is one of the rapid utilization of this source of supply. Many ectohydric and endohydric species show readily inducible nitrate reductase activity, and several species have been demonstrated to respond to the atmospheric supply by the induction of this enzyme (Lee and co-workers, unpublished). The mechanism controlling the response or lack of response of nitrogen assimilation to an enhanced atmospheric nitrogen supply resulting from atmospheric pollution is uncertain, but Schwoerbel & Tillmans (1974) observed a similar regulation of nitrate reductase by nitrogen supply in Fontinalis antipyretica Hedw. to that observed in Sphagnum species (Woodin & Lee, 198713). Thus the inhibition of nitrate reductase induction may be early evidence of supra-optimal atmospheric nitrogen supply in many bryophyte species.
A more speculative and so far unanswerable question is to what extent has the increased atmospheric nitrogen supply this century contributed, at least in part, to the success of species such as Dicranoweisia cirrata (Hedw.) Milde, Dicranum tauricum Sapehin, D . montanum Hedw. and Sphagnum recuruum which have either expanded their distributions dramatically or have become much more abundant in habitats than in the past? There is still relatively little known about the nitrogen nutrition of all but a few bryophyte species (see e.g. Brown, 1982) and about the deposition of atmospheric combined nitrogen to bryophyte canopies. Until this situation is rectified, it is impossible to ascribe any major role of nitrogen deposition in the success of these species, but our knowledge of the behaviour of nitrate assimilation reviewed above strongly suggests that the potential importance of the atmospheric nitrogen supply in these phenomena should not be ignored.
Although there is no evidence that at ambient concentrations the dry deposition of NO, is affecting bryophytes in rural areas of Britain, there have

264 J. A. LEE E l AL.
been few fumigation studies to support this view, and insufficient attempts have been made to examine the effects of NO, and sulphur dioxide mixtures. In general there is a great dearth of experiments involving the effects of mixtures of gaseous pollutants on bryophytes despite the evidence from higher plants that effects on growth in mixtures can be observed at lower concentrations than with single pollutants (see e.g. Mooi, 1984).
Despite the decline in sulphur dioxide concentrations in Britain in recent decades it still remains a major pollutant, and an understanding of sulphur dioxide tolerance in bryophytes is still important. The question raised by the present study is to what extent do bryophyes show differences in tolerance to sulphur dioxide; is it merely imposed by the physico-chemical nature of the habitats in which the species grow? Certainly, apparently sulphur dioxide- tolerant bryophytes such as Btyum argenteum Hedw. and Cerutodon purpureus (Hedw.) Brid. grow in situations where the milieu of their shoots and leaves must be rich in iron-containing particles. Thus a major part of the sulphur dioxide exclusion process might be environmentally imposed. Such a phenomenon could be used to explain the ability of some species to survive on some substrates but not on others in polluted districts. Thus, for example, Gilbert (1970) reported that a number of species showed enhanced growth and survival in a polluted habitat showing nutrient flushing. Similarly the generally poor survival of species growing on trees compared to other habitats (see e.g. Rao, 1982) might be explained by a generally poorer supply of iron, manganese or copper to the shoots of species growing epiphytically.
It would seem perhaps unlikely that this can provide a total explanation of sulphur dioxide tolerance in all bryophytes, but many species have high cation exchange capacities in their cell walls (see e.g. Brown, 1982) and these may be as high as those of some Sphagnum species. Almost certainly some physiological mechanisms of tolerance must exist in bryophytes including Sphagnum species, but the interaction of growth form, ion exchange properties of the leaves and the element supply from the substratum to provide a sulphur dioxide exclusion mechanism may be much more important in tolerance than has been hitherto realized.
REFERENCES
BAXTER, R., EMES, M. J. & LEE, J. A., 1989. The relationship between extracellular metal accumulation and bisulphite tolerance in S’ Nspidahtnr Hoffm. .New P&ologist, 111: 463-472.
BROWN, D. H., 1982. Mineral nutrition. In A. J. E. Smith (Ed.), Bryofhyt8 Ecoloo: 383444. London: Chapman & Hall.
CLYMO, R. S; & HAYWARD, P. M., 1982. The ecology of Sphagnum. In A. J. E. Smith (Ed.), Bryophytc Ecology: 229-289. London: Chapman & Hall.
FERGUSON, P. & LEE, J. A., 1979. The effects of bisulphite and sulphate upon photosynthesis in Sphagnum species. Ncu Plytologist, 82: 703-712.
FERGUSON, P. & LEE, J. A., 1983. Past and present sulphur pollution in the southern Pennines. Atmospheric Environmcnf, 17: 1131-1 137.
FERGUSON, P., LEE, J. A. & BELL, J. N. B., 1978. Effects of sulphur pollutants on the growth of Sphagnum species. Envirommkd Pollution, 16: 151-162.
GILBERT, 0. L., 1968. Bryophytes as indicators of air pollution in the Tyne Valley. Ncw Phytohgisl, 67: 15-30.
GILBERT, 0. L., 1970. Further studies on the effect of sulphur dioxide on lichens and bryophytes. .New Phytologisf, 69: 605-627.
LEE, J. A., PRESS, M. C., WOODIN, S. & FERGUSON, P., 1987. Responses to acidic depi t ion in ombmtmphic mires in the U.K. In T. C. Hutchinson & K. M. Meema (Eds), E f f h of A~~ Pdlu6ants on Forests, W c t l d and Agricurhaal Ecosysfm.~: 549-560. Berlin, Heidelberg: Springer-Verlag.

S P H A G N U M RESPONSES TO NITROGEN AND SULPHUR 265
LEE, J. A. Br WOODIN, S. J., 1988. Vegetation structure and the interception of acidic deposition by ombrotrophic mires. In J. T. A. Verhowen, G. W. Heil & M. J. Werger (Eds), Ycgdation Stnrchrrc: 137-148. The Hague: SPB Academic Publishing.
MOO& J., 1984. Wirkungen von SO,, NO,, O3 und ihrer Mixhungen auf Pappein und einige andere Pllanzenarten. Dcr Forst und Holzwirt, 3: 438-444.
PRESS, M. C., FERGUSON, P. & LEE, J. A., 1983. Two hundred years ofacid rain. Naturalist, 108: 125-129. PRESS, M. C., WOODIN, S. J. & LEE, J. A., 1986. The potential importance of an increased atmospheric
nitrogen supply to the growth of ombrotrophic Sphagnum species. New Phytologist, 105: 103-1 15. RAO, D. M., 1982. Responses of bryophytes to air pollution. In A. J. E. Smith (Ed.), BIyophyte Ecology: 445-
471. London: Chapman & Hall. RAO, D. N. & LEBLANC, F., 1967. Influence of an iron-sintering plant on corticolous epiphytes in Wawa,
Ontario. Bryologist, 70: 141-157. SCHWOERBEL, J. & TILLMANS, G. C., 1974. Assimilation of nitrogen from the medium and nitrate
reductase activity in submerged macrophytes: Fontinnlis anti&retica L. Archiv f? Hydrobiologic, 47: 282-294. SMITH, R. A., 1872. Air and f i n . The Beginnings Ofa Chemical Climatologv. London: Longmans, Green ik Co. STUDHOLME, C. J., 1989. The evolution of pollution tolcrancc in Sphagnum cuspidatum. Unpublished Ph.D.
TALLIS, J. H., 1964. Studies on southern Pennine peats. 111. The behaviour of Sphagnum. journal of Ecologv,
TOUFFET, J., 1972. Influence de la teneur en fer de milieu sur le dtveloppement des Sphaignes. Comptc rendu
WOODIN, S. J. & LEE, J. A., 1987a. The fate of some components of acidic deposition in ombrotrophic
WOODIN, S. J. & LEE, J. A., 1987b. The effects of nitrate, ammonium and temperature on nitrate reductase
WOODIN, S. J., PRESS, M. C. & LEE, J. A., 1985. Nitrate reductase activity in Sphagnum f a c u m in relation
thesis of the University of Manchester.
52: 345-353.
hebdomadaire des slances de I’Acadlmie dcs Sciences, Series d, 274: 2 175-2 177.
mires. Enuironmcntal Pollution, 45: 61-72.
activity in Sphagnum species. Nm Phytologisl, 105: 103-1 15.
to atmospheric nitrate deposition. New PhNologist, 99: 381-388.