research article rhoa controls wnt upregulation on...
TRANSCRIPT

Research ArticleRhoA Controls Wnt Upregulation on MicrostructuredTitanium Surfaces
Simone Lumetti1 Silvia Mazzotta2 Sara Ferrillo2 Maddalena Piergianni1
Marilina Piemontese1 Giovanni Passeri2 Guido Maria Macaluso13 and Carlo Galli1
1 Section of Periodontology and Implant Dentistry Centro di Odontoiatria University of Parma Via Gramsci 14 43126 Parma Italy2 Department of Clinical and Experimental Medicine University of Parma Via Gramsci 14 43126 Parma Italy3 Department of Biomedical Biotechnological and Translational Sciences University of Parma Via Gramsci 14 43126 Parma Italy
Correspondence should be addressed to Guido Maria Macaluso guidomariamacalusouniprit
Received 28 January 2014 Accepted 20 March 2014 Published 14 May 2014
Academic Editor H C Van der Mei
Copyright copy 2014 Simone Lumetti et alThis is an open access article distributed under theCreative CommonsAttribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Rough topography enhances the activation of Wnt canonical signaling in vitro and this mediates its effects on cell differentiationHowever the molecular mechanisms underlying topography-dependent control of Wnt signaling are still poorly understood Asthe small GTPase RhoA controls cytoskeletal reorganization and actomyosin-induced tensional forces we hypothesized that RhoAcould affect the activation ofWnt signaling in cells onmicropatterned titanium surfacesG-LISA assay revealed that RhoAactivationwas higher in C2C12 cells on rough (SLA) surfaces under basal conditions than on smooth (Polished) titanium Transfection withdominant negative RhoA decreasedWnt activation by normalized TCF-Luc activity on SLA whilst transfection with constitutivelyactive RhoA increased TCF-Luc activation on Polished titanium One mM Myosin II inhibitor Blebbistatin increased RhoAactivation but decreased Wnt activation on SLA surfaces indicating that tension-generating structures are required for canonicalWnt modulation on titanium surfaces Actin inhibitor Cytochalasin markedly enhanced RhoA and TCF-Luc activation on bothsurfaces and increased the expression of differentiation markers in murine osteoblastic MC3T3 cells Taken together these datashow that RhoA is upregulated in cells on rough surfaces and it affects the activation of Wnt canonical signaling through MyosinII modulation
1 Introduction
Surface profile is one of the major determinants of cell andtissue responses to endosseous implants A large amount ofexperimental evidence over the years has demonstratedthat rough topography can affect the differentiation ofmesenchymal and bone cells bolstering the expression ofosteoblast-specific genes [1ndash3] Several biochemical mecha-nisms have been advocated such as prostaglandin secretionand paracrine signaling and consensus has been reachedabout optimal roughness to support the differentiation ofbone cells but little insight is available as to how cells cansense the geometrical features of the implant surface andtransduce them into prodifferentiative stimuli Kilian et alhave shown that cell shape on micropatterned substrates canalter cytoskeletal organization and has dramatic effects on the
fate ofmesenchymal cells to specific lineages [4]More specif-ically star-like shapes were shown to promote osteoblasticdifferentiation in contrast to rounder morphologies associ-ated with adipocytic commitment The organization of thecytoskeleton is therefore central to the differentiation processand among the cellular pathways that control cell shape theWnt noncanonical planar cell polarity (PCP) pathway standsout as one of the most effective determinants of cytoskeletalarrangement As the Wnt canonical signaling cascade hasbeen shown to be affected by substrate microtexture [5ndash7]the present study investigated whether some of the maincontrollers of cytoskeletal organization the small GTPaseRhoA which is central to the PCP pathway and its down-stream targets actin andMyosin II are involved in topographymediated Wnt upregulation and how they contribute to cellsrsquoability to react to surface topography
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014 Article ID 401859 9 pageshttpdxdoiorg1011552014401859
2 BioMed Research International
2 Materials and Methods
21 Titanium Surfaces Polished and acid-etched sand-blasted (SLA) commercially pure titanium samples werekindly provided by Straumann Institut AG (Basel Switzer-land) These surfaces have been extensively described andcharacterized in the literature [8ndash10] The samples were pro-vided as sterile discs of 1mm thickness and 16mm diameterand were used in 24 well plates (Euroclone Pero MI Italy)for the biological assays
22 Cell Cultures The C2C12 cell line was obtained fromthe European Catalog of Cell Cultures (Health ProtectionAgencyCulture Collections Salisbury UK) andwas grown inDulbecco modified MEM (DMEM Paa Pasching Austria)10 fetal bovine serum (FBS Gibco Life Technologies ItalySan Giuliano Milanese MI Italy) 1 penicillin and strepto-mycin (Penstrep SigmaAldrich St LouisMOUSA) and 1Glutamine (SigmaAldrich) C2C12 cells are an uncommittedpromyoblastic murine cell line that displays the ability to dif-ferentiate along both the myocytic and osteoblastic lineagesin the presence of appropriate cues Due to their abundantexpression of Wnt molecular machinery they are a well-established in vitromodel to investigate the regulation ofWntcanonical signaling The MC3T3-E1 cell line was obtainedfrom the American Type Culture Collection (LGC StandardsSrL Sesto S Giovanni MI Italy) and cultured in DMEMmedium as described above MC3T3 are osteoblastic cellsfrom mouse calvaria They typically retain strong similaritieswith primary cells such as contact inhibition and are thusan established in vitro model of osteoblasts To perform theviability assay and for gene expression 100000 C2C12 cellswere plated on Polished or SLA discs in 1mL of completemedium in 24well plates (Corning TewksburyMAUSA) intriplicate and assayed 24 hours after plating For transfectionand reporter assays C2C12 cells were plated on titaniumsurfaces in 1mLwell Opti-MEM (Life Technologies Italy)5 FBS 1 Penstrep at the density of 120000 cellswell andthe cells were assayed after 24 hours For differentiation assay100000 MC3T3 cells were seeded on titanium surfaces incomplete DMEM Culture medium was replaced by freshmedium containing 250mM ascorbic acid (Sigma-AldrichSt Louis MO USA) 24 hours after plating and cells werecultured for 3 days prior to RNA extraction
23 Inhibitors For Blebbistatin and Cytochalasin experi-ments cells were plated as described above and stimulatedwith 02mM Cytochalasin or 1mM Blebbistatin (InalcoMilano Italy) four hours after plating for the whole durationof the experiments An equal amount of PBS was used as acontrol Pilot experiments were performed to choose thereagent concentrations which were chosen based on the lit-erature as the minimal dose to induce consistent and pre-dictable morphological changes at immunofluorescence
24 RhoA Measurement RhoA activation was measuredusing luminescence-based G-LISAActivation kit (Cytoskele-ton Inc Denver CO USA) according to the manufacturer
instructions Values were normalized by protein contentusing a colorimetric assay (Bio-Rad Laboratories HerculesCA USA) according to manufacturerrsquos recommendations
25 Constructs The TCF-Luc assay kit was purchased fromSABioscience (Frederick MD USA) The kits contained avector carrying the Firefly Luciferase gene under the controlof a TCF-binding regulatory element and a control plasmidcarrying a constitutively expressed Renilla Luciferase geneunder the control of the CMV promoter Luciferase activitywas calculated as the ratio between Firefly and RenillaLuciferase The dominant negative RhoA (plasmid 12152pTriEx-RhoA FLAREsc Biosensor T19N) and constitutivelyactive (plasmid 12151 pTriEx-RhoA FLAREsc BiosensorQ63L) isoforms were obtained from public Addgene plasmidrepository andwere kindly shared by theKlausHahnLab [11]
26 Reporter Assays The reporter assays were performedwith dual-Luciferase reporter assay system (Promega ItalyMilan) according to the manufacturerrsquos recommendationsFor background reading recombinant mouse Wnt3a wasused The Wnt3a recombinant protein was obtained fromRampD systems (Minneapolis MN USA)
The samples were read with a Glomax 2020 Luminome-ter (Promega) with double injectors
27 Real-Time PCR Total RNA was purified from cell cul-tures using Trizol (Life Technologies Italy San GiulianoMilanese MI Italy) according to the manufacturerrsquos direc-tions TaqMan quantitative RT-PCR was performed as pre-viously described using the following primer probe sets fromApplied Biosystems (Foster City CA USA) Alkaline Phos-phatase (Mm00475834 m1) Osteocalcin (for 51015840-GCTGCG-CTCTGTCTCTCTGA-31015840 rev 51015840-TGCTTGGACATGAAG-GCTTTG-31015840 probe 51015840-FAM-AAGCCCAGCGGCC-NFQ-31015840) Mouse ribosomal protein S2 ChoB (for 51015840-CCCAGG-ATGGCGACGAT-31015840 rev 51015840-CCGAATGCTGTAATGGCG-TAT-31015840 probe 51015840-FAM-TCCAGAGCAGGATCC-NFQ-31015840)was used as housekeeping gene
28 Immunofluorescence Cells were seeded at the concen-tration of 20000 cellswell in complete medium treated withvehicle or the inhibitor as described above and after 48hours they were fixed with 4 paraformaldehyde (Sigma-Aldrich StLouis MO USA) for 10min followed by threerinses with PBS They were then permeabilized with 01Triton-X100 (Sigma-Aldrich St Louis MO USA) for 5minfollowed by three rinses with PBS Nonspecific binding siteswere blocked by incubating the samples in 1 bovine serumalbumin in PBS for 20min Staining was then performedon each sample with TRITC-conjugated phalloidin (FAK100Chemicon Billerica MA USA) for 1 hour followed by threerinses with PBS Nuclear counterstaining was performedby incubation with DAPI (D1306 Molecular Probes LifeTechnologies) for 5min followed by three rinses with PBSAll the steps were carried out inside the culture well at roomtemperature The treated discs were then transferred tomicroscope slides and were mounted under glass cover
BioMed Research International 3
RhoA
Polished000
025
050
075
SLA
OD490to
tal p
rote
in co
nten
tlowast
(a)
TCF-Luc
000
025
050
075
100
Vehicle caRhoA
dnRhoA
Fire
flyR
enill
a lum
ines
cenc
e lowast
PolishedSLA
sect
(b)
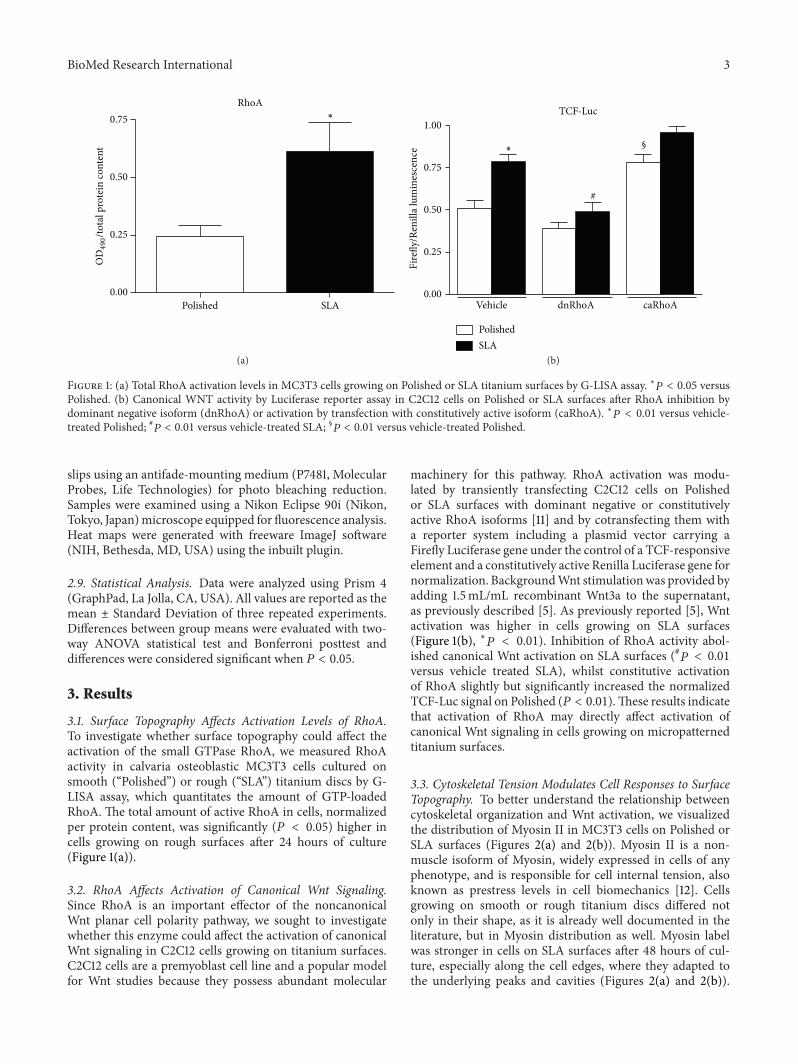
Figure 1 (a) Total RhoA activation levels in MC3T3 cells growing on Polished or SLA titanium surfaces by G-LISA assay lowast119875 lt 005 versusPolished (b) Canonical WNT activity by Luciferase reporter assay in C2C12 cells on Polished or SLA surfaces after RhoA inhibition bydominant negative isoform (dnRhoA) or activation by transfection with constitutively active isoform (caRhoA) lowast119875 lt 001 versus vehicle-treated Polished 119875 lt 001 versus vehicle-treated SLA sect119875 lt 001 versus vehicle-treated Polished
slips using an antifade-mounting medium (P7481 MolecularProbes Life Technologies) for photo bleaching reductionSamples were examined using a Nikon Eclipse 90i (NikonTokyo Japan)microscope equipped for fluorescence analysisHeat maps were generated with freeware ImageJ software(NIH Bethesda MD USA) using the inbuilt plugin
29 Statistical Analysis Data were analyzed using Prism 4(GraphPad La Jolla CA USA) All values are reported as themean plusmn Standard Deviation of three repeated experimentsDifferences between group means were evaluated with two-way ANOVA statistical test and Bonferroni posttest anddifferences were considered significant when 119875 lt 005
3 Results
31 Surface Topography Affects Activation Levels of RhoATo investigate whether surface topography could affect theactivation of the small GTPase RhoA we measured RhoAactivity in calvaria osteoblastic MC3T3 cells cultured onsmooth (ldquoPolishedrdquo) or rough (ldquoSLArdquo) titanium discs by G-LISA assay which quantitates the amount of GTP-loadedRhoA The total amount of active RhoA in cells normalizedper protein content was significantly (119875 lt 005) higher incells growing on rough surfaces after 24 hours of culture(Figure 1(a))
32 RhoA Affects Activation of Canonical Wnt SignalingSince RhoA is an important effector of the noncanonicalWnt planar cell polarity pathway we sought to investigatewhether this enzyme could affect the activation of canonicalWnt signaling in C2C12 cells growing on titanium surfacesC2C12 cells are a premyoblast cell line and a popular modelfor Wnt studies because they possess abundant molecular
machinery for this pathway RhoA activation was modu-lated by transiently transfecting C2C12 cells on Polishedor SLA surfaces with dominant negative or constitutivelyactive RhoA isoforms [11] and by cotransfecting them witha reporter system including a plasmid vector carrying aFirefly Luciferase gene under the control of a TCF-responsiveelement and a constitutively active Renilla Luciferase gene fornormalization BackgroundWnt stimulationwas provided byadding 15mLmL recombinant Wnt3a to the supernatantas previously described [5] As previously reported [5] Wntactivation was higher in cells growing on SLA surfaces(Figure 1(b) lowast119875 lt 001) Inhibition of RhoA activity abol-ished canonical Wnt activation on SLA surfaces (119875 lt 001versus vehicle treated SLA) whilst constitutive activationof RhoA slightly but significantly increased the normalizedTCF-Luc signal on Polished (119875 lt 001)These results indicatethat activation of RhoA may directly affect activation ofcanonical Wnt signaling in cells growing on micropatternedtitanium surfaces
33 Cytoskeletal Tension Modulates Cell Responses to SurfaceTopography To better understand the relationship betweencytoskeletal organization and Wnt activation we visualizedthe distribution of Myosin II in MC3T3 cells on Polished orSLA surfaces (Figures 2(a) and 2(b)) Myosin II is a non-muscle isoform of Myosin widely expressed in cells of anyphenotype and is responsible for cell internal tension alsoknown as prestress levels in cell biomechanics [12] Cellsgrowing on smooth or rough titanium discs differed notonly in their shape as it is already well documented in theliterature but in Myosin distribution as well Myosin labelwas stronger in cells on SLA surfaces after 48 hours of cul-ture especially along the cell edges where they adapted tothe underlying peaks and cavities (Figures 2(a) and 2(b))
4 BioMed Research International
Polished
(a)
SLA
(b)
RhoA
00
03
06
09
12
Vehicle Blebbistatin
OD490
lowast
PolishedSLA
(c)
TCF-Luc
000
005
010
015
020
Luci
fera
se ac
tivity
Vehicle Blebbistatin
lowast
PolishedSLA
(d)
Figure 2 (a)-(b) Immunocytochemistry ofMyosin II distribution inMC3T3 cells on Polished or SLA surfaces (20xmagnification bar 20mminsets 40xmagnification) Heatmapswere generated using ImageJ freeware software (c) Total RhoA activation levels inMC3T3 cells growingon Polished or SLA titanium surfaces by G-LISA assay after Myosin II inhibition by 1mM Blebbistatin lowast119875 lt 001 versus vehicle-treated SLA119875 lt 0001 versus Blebbistatin-treated Polished (d) Canonical WNT activity by Luciferase reporter assay in C2C12 cells on Polished or SLAsurfaces after Myosin II inhibition by 1mM Blebbistatin lowast119875 lt 001 versus vehicle-treated Polished and Blebbistatin-treated SLA
Interestingly inhibition of Myosin II activation by 1mMMyosin Light Chain Kinase inhibitor Blebbistatin signifi-cantly increasedRhoAactivity inMC3T3 cells asmeasured byG-LISA assay (Figure 2(c) lowast119875 lt 001 versus vehicle-treatedSLA) As RhoA controls Myosin II activation via the MyosinLight Chain Kinase enzyme this finding suggests a possiblefeedback loop control from Myosin or a tension-sensitivestructure back to RhoAHoweverMyosin inhibition by Bleb-bistatin decreased activation of Wnt canonical signaling inC2C12 cells on SLA surfaces (Figure 2(d) lowast119875 lt 001 versusvehicle treated Polished) in spite of the surge of RhoA activity(Figure 2(c)) thus indicating that cell contractility is requiredfor the increase in canonical Wnt activation observed onrough surfaces and that RhoA control of canonical Wntactivation requires cell tension
Immunofluorescence microscopy (Figures 3(a)ndash3(d))revealed that MC3T3 cells on Polished surfaces had fewerand less evident stress fibers after treatment with Blebbistatinwhile cells on rough surfaces appeared more elongated and
stretched presumably for their inability to retract their rearend during migration across the surface
34 Cytochalasin Increases RhoA Activation and Affects CellDifferentiation on Titanium We then investigated the effectsof microfilament structure on MC3T3 cell responses tomicropatterned titanium surfacesMicrofilaments are a densenetwork of actin fibers that permeate cells and can assemblealong the main tension lines to form stress fibers and wellvisible structures at transmission microscopy (Figure 3(a)arrowheads) These are more evident in cells growing onsmooth surfaces than on rough discs (Figures 3(a) and 3(b))where actin actually appears just as dots or short rodsCytochalasin is well known to disrupt actin polymerizationand 02mM Cytochalasin solution profoundly altered theconformation of the actin cytoskeleton on both titaniumsurfaces at least in part disrupting stress fibers whichappeared fragmented on both surfaces (Figures 3(e) and 3(f))Cytochalasin affected cell shape as well inducing irregular
BioMed Research International 5
Polished
Vehi
cle
(a)
SLA
(b)
Bleb
bista
tin
(c) (d)
Cyto
chal
asin
(e) (f)
Figure 3 Immunocytochemistry microphotographs (40x magnification bar 10mm) of MC3T3 cells growing on Polished (a c e) or SLA (bd f) titanium discs under basal conditions (a b) after addition of Blebbistatin (c d) or Cytochalasin (e f) Arrowheads indicate stress fibers
cell morphologies It has been previously reported thatCytochalasin increases RhoA activation and this was con-firmed in MC3T3 cells on Polished discs (Figure 4(a) lowast119875 lt001 versus vehicle-treated Polished 119875 lt 005 versus vehicle-treated Polished) Cytochalasin also significantly increasedthe activation of canonical Wnt signaling in C2C12 cells inboth titanium groups (Figure 4(b) lowast119875 lt 001 versus vehicle-treated POlished
119875 lt 005 versus vehicle treated SLA)When osteoblastic MC3T3 cells were treated with 02mMCytochalasin in osteogenic medium for 3 days an increase inthe expression of osteoblastic markers Alkaline Phosphatase(lowast119875 lt 005 versus vehicle treated Polished 119875 lt 005 versus
vehicle treated SLA) and Osteocalcin (lowast119875 lt 005 versusvehicle treated SLA) was observed (Figures 4(c)-4(d))
4 Discussion
Although there is a large consensus that rough titanium sur-faces promote higher expression levels of osteoblastic genes invitro and bone formation in vivo the underlyingmechanismsare still only partially known Our group and other researchteams have provided evidence that microtextured surfacescan affect the activation of Wnt canonical signaling [5 713] a pathway required for osteoblastic commitment and
6 BioMed Research International
RhoA
000
025
050
075
Vehicle
OD490to
tal p
rote
in co
nten
t
Cytochalasin
lowast
(a)
TCF-Luc
00
03
06
09
12
Vehicle Cytochalasin
Fire
flyR
enill
a lum
ines
cenc
e
lowast
(b)
lowast
ALP
00
04
08
12
Vehicle Cytochalasin
Relat
ive m
RNA
expr
essio
n
PolishedSLA
(c)
lowastOCN
0
10
20
30
40
Vehicle Cytochalasin
mRN
A re
lativ
e exp
ress
ion
PolishedSLA
(d)
Figure 4 Effects of 02mM Cytochalasin treatment on (a) total RhoA activation levels by G-LISA assay (lowast119875 lt 001 versus vehicle-treatedpolished 119875 lt 005 versus vehicle-treated Polished) in MC3T3 cells and (b) canonical WNT activity by Luciferase reporter assay in C2C12cells growing on Polished or SLA titanium surfaces lowast119875 lt 001 versus vehicle-treated Polished (c d) Real-time PCR analysis of AlkalinePhosphatase (ALP lowast119875 lt 001 versus vehicle-treated Polished) and Osteocalcin (OCN lowast119875 lt 001 versus vehicle-treated SLA) expression inmurine osteoblastic MC3T3 cells on Polished or SLA surfaces after Cytochalasin addition
differentiation [14 15] Understanding how cells can sensethe three-dimensional environment that surrounds them andtransform geometric cues into intracellular signals couldbe the key to create tailored implant surfaces to promotebone anabolism and to support bone formation aroundimplanted devices It is well known that cells can adaptto their culture substrate and conform to its profile cellshape can correspondently tune to the roughness degreeof the surface on which they grow thanks to an adaptablecytoskeleton According to Ingberrsquos tensegrity theory cellsreach a state of mechanical balance that maintains their con-formation by opposing forces [16 17] These are transmittedthrough rigid struts formed by cytoskeletal microtubulesand tensile wires the actin microfilaments anchored to thesubstrate or the extracellular matrix in a tissue throughintegrins [18 19] This structure is under a proper tension
or prestress provided by Myosin motor proteins that actby pulling on actin filaments [20] Myosin is activated bya kinase enzyme known as Myosin Light Chain Kinasewhich is controlled through a signaling cascade by the smallGTPase RhoA one of the target effectors of the noncanonicalWnt planar cell polarity (PCP) pathway [21] This signalingcascade plays an important role in orchestrating cell andtissue organization [22] and is triggered by binding of WNTfactors such asWNT5a to a FrizzledOrphan tyrosine kinaseRor2 receptor complex [23] This recruits the Disheveledprotein (Dvl) to the cell membrane activating it [24] Dvlcomprises three molecular domains which can alternativelytrigger the canonical b-catenin-dependent pathway andother noncanonical pathways such as the PCP pathway[25ndash27] Since cell shape adapts to rough implant surfacesand since the Wnt-related pathways controlling cytoskeletal
BioMed Research International 7
RhoA
MLCK
Actin
Integrins
Myosin II
TCF
Frizzled LRP4-6
Wnt
DvlAxin
Nucleus
Destructioncomplex
Frizzled Ror2C
Wnt
120573cat
120573cat
120573cat
GSK3120573
CK120572
Figure 5 Diagram depicting the proposed model for RhoA role in cell responses to substrate topography Noncanonical Wnt factors bindto receptor dimers Frizzled-Ror2C which activate RhoA Rho in turns activates downstream target Myosin Light Chain Kinase (MLCK)which phosphorylates Myosin II Acto-Myosin filaments create cell tension which facilitates the activation of canonical Wnt signaling andpromotes b catenin release from its destruction complex and translocation to the nucleus possibly by acting on tension sensitive structures(focal adhesions) A feedback loop from tension-activated structures controls RhoA activation levels Actin microfilaments can control RhoAtoo as their partial disruption increases RhoA activity and canonical Wnt activation
organization and osteoblastic commitment share some oftheir key molecular components we hypothesized thatcytoskeletal organization could at least in part cause theactivation of Wnt signaling observed on rough surfacesTo investigate the relation between cytoskeleton and Wntsignaling activation on implant surfaces we quantitatedactive RhoA small GTPase and found that its levels arehigher on rough SLA surfaces according to recent inde-pendent data [28] It can be speculated that cells need torearrange their cytoskeleton through RhoA activation toadapt to the irregular surface profile of SLA discs Interest-ingly hampering RhoA activity by transfecting cells with adominant negative isoformof RhoA (dnRhoA) reducedTCF-mediated transcriptional activity on rough surfaces whichwe previously showed to be higher than on smooth surfacesunder basal conditions [5] This clearly indicates that RhoAactivation is required for the enhancement of Wnt canonicalsignaling on rough surfaces However we also showed thatRhoA-mediated canonical Wnt activation required RhoAdownstream molecular effectors to be functional Myosininhibition by 1mM Blebbistatin did increase RhoA activitypossibly by a negative feedback loop but failed to enhanceWnt canonical signaling On the contrary Wnt activationdropped on SLA surfaces in the presence of Blebbistatinsimilar to what observed with dnRhoA This is consistentwith our observation of a complex Myosin distribution
in cells growing on SLA surfaces (Figures 2(a) and 2(b))presumably maintaining adequate cell adhesion by exertingproper cytoskeletal forces on focal adhesions and it isalso consistent with several reports showing that MyosinII function is required for cell differentiation [29 30] Itcan thus be hypothesized that RhoA enhances canonicalWnt signaling by increasing Myosin activation which inturn is known to activate cytoskeleton-associated signalingmolecules such as focal adhesion-associated Kinases (FAKs)[31] (Figure 5)These have been proven to affect the canonicalWnt signaling pathway in certain in vitro and in vivo models[32 33]This idea is consistent with recent findings indicatingthat focal adhesion formation and FAK phosphorylationare controlled by the RhoA pathway that focal adhesionmaturation and actin polymerization are promoted in cellscultured on micropatterned substrates [34] and that FAKphosphorylation is required for topography-induced differ-entiation of human mesenchymal stem cells [35]
In our study we then treated cells on both surfaceswith Cytochalasin Interestingly cells growing on Polishedsurfaces had thick stress fibers and a well-ordered actinnetwork under basal conditions whilst cells on SLA surfaceswhich do not have a plain uniform surface to adhere onlacked long actin filaments It can be speculated that shorteractin filaments can impart a more variegated array of forcevectors to the cell membrane and cytoplasm to better adapt to
8 BioMed Research International
the correspondently irregular titanium surface underneathCytochalasin at this concentration fragmented the actincytoskeleton by partially inhibiting its polymerization andsignificantly affected cell morphology Cytochalasin alsoincreased RhoA activity as previously reported and cor-respondently increased Wnt activation on both Polishedand SLA discs Under these conditions cell differentiationmarkers increased on both surfaces (Figures 4(c) and 4(d))consistently with reports by Higuchi et al [36]
Taken together our results indicate that cells growing onrough surfaces rearrange their cytoskeleton and this involvesthe activation of the small GTPase RhoA which controlsdownstream effectors that orchestrate cell contractility andshape Moreover RhoA can control and is required for theactivation ofWnt canonical signaling on rough surfacesThiseffect is not exerted directly but through its downstreameffector Myosin II When this is inhibited Wnt activation onrough surfaces is reversed in spite of RhoA activation Ourresults do not identify the specific mechanism of action forMyosin II although it may likely include focal adhesion-associated signaling which is known to be triggered by celltension and contractility Actin filaments can control RhoAactivity and their fragmentation increasesWnt activation andpromotes osteoblast differentiation (Figure 5)
Further studies will be required to better understand howcell contractility allows cells to sense their geometricalmicro-environment but the present study adds another and impor-tant piece of evidence to this elusive yet fundamental puzzlethat sustains the integration of osseointegrated implants inbone
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
S Mazzotta and S Ferrillo contributed equally to the presentstudy
Acknowledgments
The authors would like to thank Straumann Institut AG (andin particular Dr Appert and Dr Molenberg) Basel Switzer-land for kindly providing the titanium surfaces used in thepresent study The authors are also grateful to Dr FrancescaRavanetti and Professor Antonio Cacchioli for their preciousadvices and technical assistance The authors would liketo thank Dr Klaus Hahn for sharing his RhoA isoformsthrough the Addgene plasmid public repository The studywas funded by Grant no 839 2012 from the ITI Foundation(Basel Switzerland)
References
[1] C Galli G Guizzardi G Passeri et al ldquoComparison of humanmandibular osteoblasts grown on two commercially available
titanium implant surfacesrdquo Journal of Periodontology vol 76no 3 pp 364ndash372 2005
[2] B G Keselowsky L Wang Z Schwartz A J Garcia and BD Boyan ldquoIntegrin 1205725 controls osteoblastic proliferation anddifferentiation responses to titanium substrates presenting dif-ferent roughness characteristics in a roughness independentmannerrdquo Journal of BiomedicalMaterials Research A vol 80 no3 pp 700ndash710 2007
[3] I Wall N Donos K Carlqvist F Jones and P Brett ldquoModifiedtitanium surfaces promote accelerated osteogenic differentia-tion of mesenchymal stromal cells in vitrordquo Bone vol 45 no1 pp 17ndash26 2009
[4] K A Kilian B Bugarija B T Lahn and M Mrksich ldquoGeo-metric cues for directing the differentiation of mesenchymalstem cellsrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 11 pp 4872ndash4877 2010
[5] C Galli G Passeri F Ravanetti E Elezi M Pedrazzoni andG M MacAluso ldquoRough surface topography enhances theactivation of Wnt120573-catenin signaling in mesenchymal cellsrdquoJournal of Biomedical Materials Research A vol 95 no 3 pp682ndash690 2010
[6] R Olivares-Navarrete S L Hyzy D L Hutton et al ldquoDirectand indirect effects of microstructured titanium substrates onthe induction of mesenchymal stem cell differentiation towardsthe osteoblast lineagerdquo Biomaterials vol 31 no 10 pp 2728ndash2735 2010
[7] N Chakravorty S Hamlet A Jaiprakash et al ldquoPro-osteogenictopographical cues promote early activation of osteoprogenitordifferentiation via enhanced TGFbeta Wnt and Notch signal-ingrdquo Clinical Oral Implants Research 2013
[8] J Y Martin D D Dean D L Cochran J Simpson B DBoyan and Z Schwartz ldquoProliferation differentiation and pro-tein synthesis of human osteoblast-like cells (MG63) culturedon previously used titanium surfacesrdquo Clinical Oral ImplantsResearch vol 7 no 1 pp 27ndash37 1996
[9] S M Bang H J Moon Y D Kwon J Y Yoo A Pae and I KKwon ldquoOsteoblastic and osteoclastic differentiation on SLAand hydrophilic modified SLA titanium surfacesrdquo Clinical OralImplants Research 2013
[10] S Zinelis N Silikas A Thomas K Syres and G Eliades ldquoSur-face characterization of SLActive dental implantsrdquo EuropeanJournal of Esthetic Dentistry vol 7 no 1 pp 72ndash92 2012
[11] O Pertz L Hodgson R L Klemke and K M Hahn ldquoSpatio-temporal dynamics of RhoA activity in migrating cellsrdquoNaturevol 440 no 7087 pp 1069ndash1072 2006
[12] D Stamenovic and M F Coughlin ldquoThe role of prestress andarchitecture of the cytoskeleton and deformability of cytoskele-tal filaments in mechanics of adherent cells a quantitativeanalysisrdquo Journal of Theoretical Biology vol 201 no 1 pp 63ndash74 1999
[13] R Olivares-Navarrete S L Hyzy J H Park et al ldquoMediationof osteogenic differentiation of human mesenchymal stem cellson titanium surfaces by a Wnt-integrin feedback looprdquo Bioma-terials vol 32 no 27 pp 6399ndash6411 2011
[14] D A Glass II P Bialek J D Ahn et al ldquoCanonical Wnt signal-ing in differentiated osteoblasts controls osteoclast differentia-tionrdquo Developmental Cell vol 8 no 5 pp 751ndash764 2005
[15] S J Rodda and A P McMahon ldquoDistinct roles for Hedgehogand caronicalWnt signaling in specification differentiation andmaintenance of osteoblast progenitorsrdquo Development vol 133no 16 pp 3231ndash3244 2006
BioMed Research International 9
[16] N Wang and D E Ingber ldquoControl of cytoskeletal mechanicsby extracellular matrix cell shape and mechanical tensionrdquoBiophysical Journal vol 66 no 6 pp 2181ndash2189 1994
[17] C Galli S Guizzardi G Passeri G M Macaluso and R Scan-droglio ldquoLife on the wire on tensegrity and force balance incellsrdquo Acta Biomedica de lrsquoAteneo Parmense vol 76 no 1 pp5ndash62 2005
[18] D E Ingber ldquoTensegrity I Cell structure and hierarchical sys-tems biologyrdquo Journal of Cell Science vol 116 no 7 pp 1157ndash11732003
[19] C S Chen M Mrksich S Huang G M Whitesides and D EIngber ldquoGeometric control of cell life and deathrdquo Science vol276 no 5317 pp 1425ndash1428 1997
[20] D Stamenovi S M Mijailovich I M Toli-Noslashrrelykke J Chenand N Wang ldquoCell prestressmdashII Contribution of microtu-bulesrdquo American Journal of Physiology Cell Physiology vol 282no 3 pp C617ndashC624 2002
[21] M Endo M Nishita and Y Minami ldquoAnalysis of wntplanarcell polarity pathway in cultured cellsrdquo Methods in MolecularBiology vol 839 pp 201ndash214 2012
[22] C Gao and Y-G Chen ldquoDishevelled the hub ofWnt signalingrdquoCellular Signalling vol 22 no 5 pp 717ndash727 2010
[23] A J Mikels and R Nusse ldquoPurified Wnt5a protein activatesor inhibits beta-catenin-TCF signaling depending on receptorcontextrdquo PLoS Biology vol 4 no 4 Article ID e115 2006
[24] Y-N Lee Y Gao and H-Y Wang ldquoDifferential mediation ofthe Wnt canonical pathway by mammalian Dishevelleds-1 -2and -3rdquo Cellular Signalling vol 20 no 2 pp 443ndash452 2008
[25] J D Axelrod J R Miller J M Shulman R T Moon andN Perrimon ldquoDifferential recruitment of dishevelled providessignaling specificity in the planar cell polarity and Winglesssignaling pathwaysrdquo Genes and Development vol 12 no 16 pp2610ndash2622 1998
[26] K AWharton Jr ldquoRunninrsquo with the Dvl proteins that associatewith DshDvl and their significance to Wnt signal transduc-tionrdquo Developmental Biology vol 253 no 1 pp 1ndash17 2003
[27] W J Pan S Z Pang T Huang H Y Guo D Wu and L LildquoCharacterization of function of three domains in dishevelled1 DEP domain is responsible for membrane translocation ofdishevelled-1rdquo Cell Research vol 14 no 4 pp 324ndash330 2004
[28] A Calzado-Martın A Mendez-Vilas M Multigner et al ldquoOnthe role of RhoAROCK signaling in contact guidance ofbone-forming cells on anisotropic Ti6Al4V surfacesrdquo Acta Bio-materialia vol 7 no 4 pp 1890ndash1901 2011
[29] Y Sun L G Villa-Diaz R H Lam W Chen P H Krebsbachand J Fu ldquoMechanics regulates fate decisions of human embry-onic stem cellsrdquo PLoS ONE vol 7 no 5 Article ID e37178 2012
[30] H J Kwon ldquoChondrogenesis on sulfonate-coated hydrogelsis regulated by their mechanical propertiesrdquo Journal of theMechanical Behavior of BiomedicalMaterial vol 17 pp 337ndash3462013
[31] D W Dumbauld H Shin N D Gallant K E Michael HRadhakrishna and A J Garcıa ldquoContractility modulates celladhesion strengthening through focal adhesion kinase andassembly of vinculin-containing focal adhesionsrdquo Journal ofCellular Physiology vol 223 no 3 pp 746ndash756 2010
[32] Y Fonar Y E Gutkovich H Root et al ldquoFocal adhesion kinaseprotein regulates Wnt3a gene expression to control cell fatespecification in the developing neural platerdquoMolecular Biologyof the Cell vol 22 no 13 pp 2409ndash2421 2011
[33] M Despeaux G Chicanne E Rouer et al ldquoFocal adhesionkinase splice variants maintain primitive acute myeloid leuke-mia cells through alteredWnt signalingrdquo Stem Cells vol 30 no8 pp 1597ndash1610 2012
[34] C H Seo K Furukawa K Montagne H Jeong and T UshidaldquoThe effect of substrate microtopography on focal adhesionmaturation and actin organization via the RhoAROCK path-wayrdquo Biomaterials vol 32 no 36 pp 9568ndash9575 2011
[35] B K Teo S T Wong C K Lim et al ldquoNanotopographymodulates mechanotransduction of stem cells and inducesdifferentiation through focal adhesion kinaserdquo ACS Nano 2013
[36] C Higuchi N Nakamura H Yoshikawa and K Itoh ldquoTran-sient dynamic actin cytoskeletal change stimulates the osteo-blastic differentiationrdquo Journal of Bone andMineralMetabolismvol 27 no 2 pp 158ndash167 2009
Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials

2 BioMed Research International
2 Materials and Methods
21 Titanium Surfaces Polished and acid-etched sand-blasted (SLA) commercially pure titanium samples werekindly provided by Straumann Institut AG (Basel Switzer-land) These surfaces have been extensively described andcharacterized in the literature [8ndash10] The samples were pro-vided as sterile discs of 1mm thickness and 16mm diameterand were used in 24 well plates (Euroclone Pero MI Italy)for the biological assays
22 Cell Cultures The C2C12 cell line was obtained fromthe European Catalog of Cell Cultures (Health ProtectionAgencyCulture Collections Salisbury UK) andwas grown inDulbecco modified MEM (DMEM Paa Pasching Austria)10 fetal bovine serum (FBS Gibco Life Technologies ItalySan Giuliano Milanese MI Italy) 1 penicillin and strepto-mycin (Penstrep SigmaAldrich St LouisMOUSA) and 1Glutamine (SigmaAldrich) C2C12 cells are an uncommittedpromyoblastic murine cell line that displays the ability to dif-ferentiate along both the myocytic and osteoblastic lineagesin the presence of appropriate cues Due to their abundantexpression of Wnt molecular machinery they are a well-established in vitromodel to investigate the regulation ofWntcanonical signaling The MC3T3-E1 cell line was obtainedfrom the American Type Culture Collection (LGC StandardsSrL Sesto S Giovanni MI Italy) and cultured in DMEMmedium as described above MC3T3 are osteoblastic cellsfrom mouse calvaria They typically retain strong similaritieswith primary cells such as contact inhibition and are thusan established in vitro model of osteoblasts To perform theviability assay and for gene expression 100000 C2C12 cellswere plated on Polished or SLA discs in 1mL of completemedium in 24well plates (Corning TewksburyMAUSA) intriplicate and assayed 24 hours after plating For transfectionand reporter assays C2C12 cells were plated on titaniumsurfaces in 1mLwell Opti-MEM (Life Technologies Italy)5 FBS 1 Penstrep at the density of 120000 cellswell andthe cells were assayed after 24 hours For differentiation assay100000 MC3T3 cells were seeded on titanium surfaces incomplete DMEM Culture medium was replaced by freshmedium containing 250mM ascorbic acid (Sigma-AldrichSt Louis MO USA) 24 hours after plating and cells werecultured for 3 days prior to RNA extraction
23 Inhibitors For Blebbistatin and Cytochalasin experi-ments cells were plated as described above and stimulatedwith 02mM Cytochalasin or 1mM Blebbistatin (InalcoMilano Italy) four hours after plating for the whole durationof the experiments An equal amount of PBS was used as acontrol Pilot experiments were performed to choose thereagent concentrations which were chosen based on the lit-erature as the minimal dose to induce consistent and pre-dictable morphological changes at immunofluorescence
24 RhoA Measurement RhoA activation was measuredusing luminescence-based G-LISAActivation kit (Cytoskele-ton Inc Denver CO USA) according to the manufacturer
instructions Values were normalized by protein contentusing a colorimetric assay (Bio-Rad Laboratories HerculesCA USA) according to manufacturerrsquos recommendations
25 Constructs The TCF-Luc assay kit was purchased fromSABioscience (Frederick MD USA) The kits contained avector carrying the Firefly Luciferase gene under the controlof a TCF-binding regulatory element and a control plasmidcarrying a constitutively expressed Renilla Luciferase geneunder the control of the CMV promoter Luciferase activitywas calculated as the ratio between Firefly and RenillaLuciferase The dominant negative RhoA (plasmid 12152pTriEx-RhoA FLAREsc Biosensor T19N) and constitutivelyactive (plasmid 12151 pTriEx-RhoA FLAREsc BiosensorQ63L) isoforms were obtained from public Addgene plasmidrepository andwere kindly shared by theKlausHahnLab [11]
26 Reporter Assays The reporter assays were performedwith dual-Luciferase reporter assay system (Promega ItalyMilan) according to the manufacturerrsquos recommendationsFor background reading recombinant mouse Wnt3a wasused The Wnt3a recombinant protein was obtained fromRampD systems (Minneapolis MN USA)
The samples were read with a Glomax 2020 Luminome-ter (Promega) with double injectors
27 Real-Time PCR Total RNA was purified from cell cul-tures using Trizol (Life Technologies Italy San GiulianoMilanese MI Italy) according to the manufacturerrsquos direc-tions TaqMan quantitative RT-PCR was performed as pre-viously described using the following primer probe sets fromApplied Biosystems (Foster City CA USA) Alkaline Phos-phatase (Mm00475834 m1) Osteocalcin (for 51015840-GCTGCG-CTCTGTCTCTCTGA-31015840 rev 51015840-TGCTTGGACATGAAG-GCTTTG-31015840 probe 51015840-FAM-AAGCCCAGCGGCC-NFQ-31015840) Mouse ribosomal protein S2 ChoB (for 51015840-CCCAGG-ATGGCGACGAT-31015840 rev 51015840-CCGAATGCTGTAATGGCG-TAT-31015840 probe 51015840-FAM-TCCAGAGCAGGATCC-NFQ-31015840)was used as housekeeping gene
28 Immunofluorescence Cells were seeded at the concen-tration of 20000 cellswell in complete medium treated withvehicle or the inhibitor as described above and after 48hours they were fixed with 4 paraformaldehyde (Sigma-Aldrich StLouis MO USA) for 10min followed by threerinses with PBS They were then permeabilized with 01Triton-X100 (Sigma-Aldrich St Louis MO USA) for 5minfollowed by three rinses with PBS Nonspecific binding siteswere blocked by incubating the samples in 1 bovine serumalbumin in PBS for 20min Staining was then performedon each sample with TRITC-conjugated phalloidin (FAK100Chemicon Billerica MA USA) for 1 hour followed by threerinses with PBS Nuclear counterstaining was performedby incubation with DAPI (D1306 Molecular Probes LifeTechnologies) for 5min followed by three rinses with PBSAll the steps were carried out inside the culture well at roomtemperature The treated discs were then transferred tomicroscope slides and were mounted under glass cover
BioMed Research International 3
RhoA
Polished000
025
050
075
SLA
OD490to
tal p
rote
in co
nten
tlowast
(a)
TCF-Luc
000
025
050
075
100
Vehicle caRhoA
dnRhoA
Fire
flyR
enill
a lum
ines
cenc
e lowast
PolishedSLA
sect
(b)
Figure 1 (a) Total RhoA activation levels in MC3T3 cells growing on Polished or SLA titanium surfaces by G-LISA assay lowast119875 lt 005 versusPolished (b) Canonical WNT activity by Luciferase reporter assay in C2C12 cells on Polished or SLA surfaces after RhoA inhibition bydominant negative isoform (dnRhoA) or activation by transfection with constitutively active isoform (caRhoA) lowast119875 lt 001 versus vehicle-treated Polished 119875 lt 001 versus vehicle-treated SLA sect119875 lt 001 versus vehicle-treated Polished
slips using an antifade-mounting medium (P7481 MolecularProbes Life Technologies) for photo bleaching reductionSamples were examined using a Nikon Eclipse 90i (NikonTokyo Japan)microscope equipped for fluorescence analysisHeat maps were generated with freeware ImageJ software(NIH Bethesda MD USA) using the inbuilt plugin
29 Statistical Analysis Data were analyzed using Prism 4(GraphPad La Jolla CA USA) All values are reported as themean plusmn Standard Deviation of three repeated experimentsDifferences between group means were evaluated with two-way ANOVA statistical test and Bonferroni posttest anddifferences were considered significant when 119875 lt 005
3 Results
31 Surface Topography Affects Activation Levels of RhoATo investigate whether surface topography could affect theactivation of the small GTPase RhoA we measured RhoAactivity in calvaria osteoblastic MC3T3 cells cultured onsmooth (ldquoPolishedrdquo) or rough (ldquoSLArdquo) titanium discs by G-LISA assay which quantitates the amount of GTP-loadedRhoA The total amount of active RhoA in cells normalizedper protein content was significantly (119875 lt 005) higher incells growing on rough surfaces after 24 hours of culture(Figure 1(a))
32 RhoA Affects Activation of Canonical Wnt SignalingSince RhoA is an important effector of the noncanonicalWnt planar cell polarity pathway we sought to investigatewhether this enzyme could affect the activation of canonicalWnt signaling in C2C12 cells growing on titanium surfacesC2C12 cells are a premyoblast cell line and a popular modelfor Wnt studies because they possess abundant molecular
machinery for this pathway RhoA activation was modu-lated by transiently transfecting C2C12 cells on Polishedor SLA surfaces with dominant negative or constitutivelyactive RhoA isoforms [11] and by cotransfecting them witha reporter system including a plasmid vector carrying aFirefly Luciferase gene under the control of a TCF-responsiveelement and a constitutively active Renilla Luciferase gene fornormalization BackgroundWnt stimulationwas provided byadding 15mLmL recombinant Wnt3a to the supernatantas previously described [5] As previously reported [5] Wntactivation was higher in cells growing on SLA surfaces(Figure 1(b) lowast119875 lt 001) Inhibition of RhoA activity abol-ished canonical Wnt activation on SLA surfaces (119875 lt 001versus vehicle treated SLA) whilst constitutive activationof RhoA slightly but significantly increased the normalizedTCF-Luc signal on Polished (119875 lt 001)These results indicatethat activation of RhoA may directly affect activation ofcanonical Wnt signaling in cells growing on micropatternedtitanium surfaces
33 Cytoskeletal Tension Modulates Cell Responses to SurfaceTopography To better understand the relationship betweencytoskeletal organization and Wnt activation we visualizedthe distribution of Myosin II in MC3T3 cells on Polished orSLA surfaces (Figures 2(a) and 2(b)) Myosin II is a non-muscle isoform of Myosin widely expressed in cells of anyphenotype and is responsible for cell internal tension alsoknown as prestress levels in cell biomechanics [12] Cellsgrowing on smooth or rough titanium discs differed notonly in their shape as it is already well documented in theliterature but in Myosin distribution as well Myosin labelwas stronger in cells on SLA surfaces after 48 hours of cul-ture especially along the cell edges where they adapted tothe underlying peaks and cavities (Figures 2(a) and 2(b))
4 BioMed Research International
Polished
(a)
SLA
(b)
RhoA
00
03
06
09
12
Vehicle Blebbistatin
OD490
lowast
PolishedSLA
(c)
TCF-Luc
000
005
010
015
020
Luci
fera
se ac
tivity
Vehicle Blebbistatin
lowast
PolishedSLA
(d)
Figure 2 (a)-(b) Immunocytochemistry ofMyosin II distribution inMC3T3 cells on Polished or SLA surfaces (20xmagnification bar 20mminsets 40xmagnification) Heatmapswere generated using ImageJ freeware software (c) Total RhoA activation levels inMC3T3 cells growingon Polished or SLA titanium surfaces by G-LISA assay after Myosin II inhibition by 1mM Blebbistatin lowast119875 lt 001 versus vehicle-treated SLA119875 lt 0001 versus Blebbistatin-treated Polished (d) Canonical WNT activity by Luciferase reporter assay in C2C12 cells on Polished or SLAsurfaces after Myosin II inhibition by 1mM Blebbistatin lowast119875 lt 001 versus vehicle-treated Polished and Blebbistatin-treated SLA
Interestingly inhibition of Myosin II activation by 1mMMyosin Light Chain Kinase inhibitor Blebbistatin signifi-cantly increasedRhoAactivity inMC3T3 cells asmeasured byG-LISA assay (Figure 2(c) lowast119875 lt 001 versus vehicle-treatedSLA) As RhoA controls Myosin II activation via the MyosinLight Chain Kinase enzyme this finding suggests a possiblefeedback loop control from Myosin or a tension-sensitivestructure back to RhoAHoweverMyosin inhibition by Bleb-bistatin decreased activation of Wnt canonical signaling inC2C12 cells on SLA surfaces (Figure 2(d) lowast119875 lt 001 versusvehicle treated Polished) in spite of the surge of RhoA activity(Figure 2(c)) thus indicating that cell contractility is requiredfor the increase in canonical Wnt activation observed onrough surfaces and that RhoA control of canonical Wntactivation requires cell tension
Immunofluorescence microscopy (Figures 3(a)ndash3(d))revealed that MC3T3 cells on Polished surfaces had fewerand less evident stress fibers after treatment with Blebbistatinwhile cells on rough surfaces appeared more elongated and
stretched presumably for their inability to retract their rearend during migration across the surface
34 Cytochalasin Increases RhoA Activation and Affects CellDifferentiation on Titanium We then investigated the effectsof microfilament structure on MC3T3 cell responses tomicropatterned titanium surfacesMicrofilaments are a densenetwork of actin fibers that permeate cells and can assemblealong the main tension lines to form stress fibers and wellvisible structures at transmission microscopy (Figure 3(a)arrowheads) These are more evident in cells growing onsmooth surfaces than on rough discs (Figures 3(a) and 3(b))where actin actually appears just as dots or short rodsCytochalasin is well known to disrupt actin polymerizationand 02mM Cytochalasin solution profoundly altered theconformation of the actin cytoskeleton on both titaniumsurfaces at least in part disrupting stress fibers whichappeared fragmented on both surfaces (Figures 3(e) and 3(f))Cytochalasin affected cell shape as well inducing irregular
BioMed Research International 5
Polished
Vehi
cle
(a)
SLA
(b)
Bleb
bista
tin
(c) (d)
Cyto
chal
asin
(e) (f)
Figure 3 Immunocytochemistry microphotographs (40x magnification bar 10mm) of MC3T3 cells growing on Polished (a c e) or SLA (bd f) titanium discs under basal conditions (a b) after addition of Blebbistatin (c d) or Cytochalasin (e f) Arrowheads indicate stress fibers
cell morphologies It has been previously reported thatCytochalasin increases RhoA activation and this was con-firmed in MC3T3 cells on Polished discs (Figure 4(a) lowast119875 lt001 versus vehicle-treated Polished 119875 lt 005 versus vehicle-treated Polished) Cytochalasin also significantly increasedthe activation of canonical Wnt signaling in C2C12 cells inboth titanium groups (Figure 4(b) lowast119875 lt 001 versus vehicle-treated POlished
119875 lt 005 versus vehicle treated SLA)When osteoblastic MC3T3 cells were treated with 02mMCytochalasin in osteogenic medium for 3 days an increase inthe expression of osteoblastic markers Alkaline Phosphatase(lowast119875 lt 005 versus vehicle treated Polished 119875 lt 005 versus
vehicle treated SLA) and Osteocalcin (lowast119875 lt 005 versusvehicle treated SLA) was observed (Figures 4(c)-4(d))
4 Discussion
Although there is a large consensus that rough titanium sur-faces promote higher expression levels of osteoblastic genes invitro and bone formation in vivo the underlyingmechanismsare still only partially known Our group and other researchteams have provided evidence that microtextured surfacescan affect the activation of Wnt canonical signaling [5 713] a pathway required for osteoblastic commitment and
6 BioMed Research International
RhoA
000
025
050
075
Vehicle
OD490to
tal p
rote
in co
nten
t
Cytochalasin
lowast
(a)
TCF-Luc
00
03
06
09
12
Vehicle Cytochalasin
Fire
flyR
enill
a lum
ines
cenc
e
lowast
(b)
lowast
ALP
00
04
08
12
Vehicle Cytochalasin
Relat
ive m
RNA
expr
essio
n
PolishedSLA
(c)
lowastOCN
0
10
20
30
40
Vehicle Cytochalasin
mRN
A re
lativ
e exp
ress
ion
PolishedSLA
(d)
Figure 4 Effects of 02mM Cytochalasin treatment on (a) total RhoA activation levels by G-LISA assay (lowast119875 lt 001 versus vehicle-treatedpolished 119875 lt 005 versus vehicle-treated Polished) in MC3T3 cells and (b) canonical WNT activity by Luciferase reporter assay in C2C12cells growing on Polished or SLA titanium surfaces lowast119875 lt 001 versus vehicle-treated Polished (c d) Real-time PCR analysis of AlkalinePhosphatase (ALP lowast119875 lt 001 versus vehicle-treated Polished) and Osteocalcin (OCN lowast119875 lt 001 versus vehicle-treated SLA) expression inmurine osteoblastic MC3T3 cells on Polished or SLA surfaces after Cytochalasin addition
differentiation [14 15] Understanding how cells can sensethe three-dimensional environment that surrounds them andtransform geometric cues into intracellular signals couldbe the key to create tailored implant surfaces to promotebone anabolism and to support bone formation aroundimplanted devices It is well known that cells can adaptto their culture substrate and conform to its profile cellshape can correspondently tune to the roughness degreeof the surface on which they grow thanks to an adaptablecytoskeleton According to Ingberrsquos tensegrity theory cellsreach a state of mechanical balance that maintains their con-formation by opposing forces [16 17] These are transmittedthrough rigid struts formed by cytoskeletal microtubulesand tensile wires the actin microfilaments anchored to thesubstrate or the extracellular matrix in a tissue throughintegrins [18 19] This structure is under a proper tension
or prestress provided by Myosin motor proteins that actby pulling on actin filaments [20] Myosin is activated bya kinase enzyme known as Myosin Light Chain Kinasewhich is controlled through a signaling cascade by the smallGTPase RhoA one of the target effectors of the noncanonicalWnt planar cell polarity (PCP) pathway [21] This signalingcascade plays an important role in orchestrating cell andtissue organization [22] and is triggered by binding of WNTfactors such asWNT5a to a FrizzledOrphan tyrosine kinaseRor2 receptor complex [23] This recruits the Disheveledprotein (Dvl) to the cell membrane activating it [24] Dvlcomprises three molecular domains which can alternativelytrigger the canonical b-catenin-dependent pathway andother noncanonical pathways such as the PCP pathway[25ndash27] Since cell shape adapts to rough implant surfacesand since the Wnt-related pathways controlling cytoskeletal
BioMed Research International 7
RhoA
MLCK
Actin
Integrins
Myosin II
TCF
Frizzled LRP4-6
Wnt
DvlAxin
Nucleus
Destructioncomplex
Frizzled Ror2C
Wnt
120573cat
120573cat
120573cat
GSK3120573
CK120572
Figure 5 Diagram depicting the proposed model for RhoA role in cell responses to substrate topography Noncanonical Wnt factors bindto receptor dimers Frizzled-Ror2C which activate RhoA Rho in turns activates downstream target Myosin Light Chain Kinase (MLCK)which phosphorylates Myosin II Acto-Myosin filaments create cell tension which facilitates the activation of canonical Wnt signaling andpromotes b catenin release from its destruction complex and translocation to the nucleus possibly by acting on tension sensitive structures(focal adhesions) A feedback loop from tension-activated structures controls RhoA activation levels Actin microfilaments can control RhoAtoo as their partial disruption increases RhoA activity and canonical Wnt activation
organization and osteoblastic commitment share some oftheir key molecular components we hypothesized thatcytoskeletal organization could at least in part cause theactivation of Wnt signaling observed on rough surfacesTo investigate the relation between cytoskeleton and Wntsignaling activation on implant surfaces we quantitatedactive RhoA small GTPase and found that its levels arehigher on rough SLA surfaces according to recent inde-pendent data [28] It can be speculated that cells need torearrange their cytoskeleton through RhoA activation toadapt to the irregular surface profile of SLA discs Interest-ingly hampering RhoA activity by transfecting cells with adominant negative isoformof RhoA (dnRhoA) reducedTCF-mediated transcriptional activity on rough surfaces whichwe previously showed to be higher than on smooth surfacesunder basal conditions [5] This clearly indicates that RhoAactivation is required for the enhancement of Wnt canonicalsignaling on rough surfaces However we also showed thatRhoA-mediated canonical Wnt activation required RhoAdownstream molecular effectors to be functional Myosininhibition by 1mM Blebbistatin did increase RhoA activitypossibly by a negative feedback loop but failed to enhanceWnt canonical signaling On the contrary Wnt activationdropped on SLA surfaces in the presence of Blebbistatinsimilar to what observed with dnRhoA This is consistentwith our observation of a complex Myosin distribution
in cells growing on SLA surfaces (Figures 2(a) and 2(b))presumably maintaining adequate cell adhesion by exertingproper cytoskeletal forces on focal adhesions and it isalso consistent with several reports showing that MyosinII function is required for cell differentiation [29 30] Itcan thus be hypothesized that RhoA enhances canonicalWnt signaling by increasing Myosin activation which inturn is known to activate cytoskeleton-associated signalingmolecules such as focal adhesion-associated Kinases (FAKs)[31] (Figure 5)These have been proven to affect the canonicalWnt signaling pathway in certain in vitro and in vivo models[32 33]This idea is consistent with recent findings indicatingthat focal adhesion formation and FAK phosphorylationare controlled by the RhoA pathway that focal adhesionmaturation and actin polymerization are promoted in cellscultured on micropatterned substrates [34] and that FAKphosphorylation is required for topography-induced differ-entiation of human mesenchymal stem cells [35]
In our study we then treated cells on both surfaceswith Cytochalasin Interestingly cells growing on Polishedsurfaces had thick stress fibers and a well-ordered actinnetwork under basal conditions whilst cells on SLA surfaceswhich do not have a plain uniform surface to adhere onlacked long actin filaments It can be speculated that shorteractin filaments can impart a more variegated array of forcevectors to the cell membrane and cytoplasm to better adapt to
8 BioMed Research International
the correspondently irregular titanium surface underneathCytochalasin at this concentration fragmented the actincytoskeleton by partially inhibiting its polymerization andsignificantly affected cell morphology Cytochalasin alsoincreased RhoA activity as previously reported and cor-respondently increased Wnt activation on both Polishedand SLA discs Under these conditions cell differentiationmarkers increased on both surfaces (Figures 4(c) and 4(d))consistently with reports by Higuchi et al [36]
Taken together our results indicate that cells growing onrough surfaces rearrange their cytoskeleton and this involvesthe activation of the small GTPase RhoA which controlsdownstream effectors that orchestrate cell contractility andshape Moreover RhoA can control and is required for theactivation ofWnt canonical signaling on rough surfacesThiseffect is not exerted directly but through its downstreameffector Myosin II When this is inhibited Wnt activation onrough surfaces is reversed in spite of RhoA activation Ourresults do not identify the specific mechanism of action forMyosin II although it may likely include focal adhesion-associated signaling which is known to be triggered by celltension and contractility Actin filaments can control RhoAactivity and their fragmentation increasesWnt activation andpromotes osteoblast differentiation (Figure 5)
Further studies will be required to better understand howcell contractility allows cells to sense their geometricalmicro-environment but the present study adds another and impor-tant piece of evidence to this elusive yet fundamental puzzlethat sustains the integration of osseointegrated implants inbone
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
S Mazzotta and S Ferrillo contributed equally to the presentstudy
Acknowledgments
The authors would like to thank Straumann Institut AG (andin particular Dr Appert and Dr Molenberg) Basel Switzer-land for kindly providing the titanium surfaces used in thepresent study The authors are also grateful to Dr FrancescaRavanetti and Professor Antonio Cacchioli for their preciousadvices and technical assistance The authors would liketo thank Dr Klaus Hahn for sharing his RhoA isoformsthrough the Addgene plasmid public repository The studywas funded by Grant no 839 2012 from the ITI Foundation(Basel Switzerland)
References
[1] C Galli G Guizzardi G Passeri et al ldquoComparison of humanmandibular osteoblasts grown on two commercially available
titanium implant surfacesrdquo Journal of Periodontology vol 76no 3 pp 364ndash372 2005
[2] B G Keselowsky L Wang Z Schwartz A J Garcia and BD Boyan ldquoIntegrin 1205725 controls osteoblastic proliferation anddifferentiation responses to titanium substrates presenting dif-ferent roughness characteristics in a roughness independentmannerrdquo Journal of BiomedicalMaterials Research A vol 80 no3 pp 700ndash710 2007
[3] I Wall N Donos K Carlqvist F Jones and P Brett ldquoModifiedtitanium surfaces promote accelerated osteogenic differentia-tion of mesenchymal stromal cells in vitrordquo Bone vol 45 no1 pp 17ndash26 2009
[4] K A Kilian B Bugarija B T Lahn and M Mrksich ldquoGeo-metric cues for directing the differentiation of mesenchymalstem cellsrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 11 pp 4872ndash4877 2010
[5] C Galli G Passeri F Ravanetti E Elezi M Pedrazzoni andG M MacAluso ldquoRough surface topography enhances theactivation of Wnt120573-catenin signaling in mesenchymal cellsrdquoJournal of Biomedical Materials Research A vol 95 no 3 pp682ndash690 2010
[6] R Olivares-Navarrete S L Hyzy D L Hutton et al ldquoDirectand indirect effects of microstructured titanium substrates onthe induction of mesenchymal stem cell differentiation towardsthe osteoblast lineagerdquo Biomaterials vol 31 no 10 pp 2728ndash2735 2010
[7] N Chakravorty S Hamlet A Jaiprakash et al ldquoPro-osteogenictopographical cues promote early activation of osteoprogenitordifferentiation via enhanced TGFbeta Wnt and Notch signal-ingrdquo Clinical Oral Implants Research 2013
[8] J Y Martin D D Dean D L Cochran J Simpson B DBoyan and Z Schwartz ldquoProliferation differentiation and pro-tein synthesis of human osteoblast-like cells (MG63) culturedon previously used titanium surfacesrdquo Clinical Oral ImplantsResearch vol 7 no 1 pp 27ndash37 1996
[9] S M Bang H J Moon Y D Kwon J Y Yoo A Pae and I KKwon ldquoOsteoblastic and osteoclastic differentiation on SLAand hydrophilic modified SLA titanium surfacesrdquo Clinical OralImplants Research 2013
[10] S Zinelis N Silikas A Thomas K Syres and G Eliades ldquoSur-face characterization of SLActive dental implantsrdquo EuropeanJournal of Esthetic Dentistry vol 7 no 1 pp 72ndash92 2012
[11] O Pertz L Hodgson R L Klemke and K M Hahn ldquoSpatio-temporal dynamics of RhoA activity in migrating cellsrdquoNaturevol 440 no 7087 pp 1069ndash1072 2006
[12] D Stamenovic and M F Coughlin ldquoThe role of prestress andarchitecture of the cytoskeleton and deformability of cytoskele-tal filaments in mechanics of adherent cells a quantitativeanalysisrdquo Journal of Theoretical Biology vol 201 no 1 pp 63ndash74 1999
[13] R Olivares-Navarrete S L Hyzy J H Park et al ldquoMediationof osteogenic differentiation of human mesenchymal stem cellson titanium surfaces by a Wnt-integrin feedback looprdquo Bioma-terials vol 32 no 27 pp 6399ndash6411 2011
[14] D A Glass II P Bialek J D Ahn et al ldquoCanonical Wnt signal-ing in differentiated osteoblasts controls osteoclast differentia-tionrdquo Developmental Cell vol 8 no 5 pp 751ndash764 2005
[15] S J Rodda and A P McMahon ldquoDistinct roles for Hedgehogand caronicalWnt signaling in specification differentiation andmaintenance of osteoblast progenitorsrdquo Development vol 133no 16 pp 3231ndash3244 2006
BioMed Research International 9
[16] N Wang and D E Ingber ldquoControl of cytoskeletal mechanicsby extracellular matrix cell shape and mechanical tensionrdquoBiophysical Journal vol 66 no 6 pp 2181ndash2189 1994
[17] C Galli S Guizzardi G Passeri G M Macaluso and R Scan-droglio ldquoLife on the wire on tensegrity and force balance incellsrdquo Acta Biomedica de lrsquoAteneo Parmense vol 76 no 1 pp5ndash62 2005
[18] D E Ingber ldquoTensegrity I Cell structure and hierarchical sys-tems biologyrdquo Journal of Cell Science vol 116 no 7 pp 1157ndash11732003
[19] C S Chen M Mrksich S Huang G M Whitesides and D EIngber ldquoGeometric control of cell life and deathrdquo Science vol276 no 5317 pp 1425ndash1428 1997
[20] D Stamenovi S M Mijailovich I M Toli-Noslashrrelykke J Chenand N Wang ldquoCell prestressmdashII Contribution of microtu-bulesrdquo American Journal of Physiology Cell Physiology vol 282no 3 pp C617ndashC624 2002
[21] M Endo M Nishita and Y Minami ldquoAnalysis of wntplanarcell polarity pathway in cultured cellsrdquo Methods in MolecularBiology vol 839 pp 201ndash214 2012
[22] C Gao and Y-G Chen ldquoDishevelled the hub ofWnt signalingrdquoCellular Signalling vol 22 no 5 pp 717ndash727 2010
[23] A J Mikels and R Nusse ldquoPurified Wnt5a protein activatesor inhibits beta-catenin-TCF signaling depending on receptorcontextrdquo PLoS Biology vol 4 no 4 Article ID e115 2006
[24] Y-N Lee Y Gao and H-Y Wang ldquoDifferential mediation ofthe Wnt canonical pathway by mammalian Dishevelleds-1 -2and -3rdquo Cellular Signalling vol 20 no 2 pp 443ndash452 2008
[25] J D Axelrod J R Miller J M Shulman R T Moon andN Perrimon ldquoDifferential recruitment of dishevelled providessignaling specificity in the planar cell polarity and Winglesssignaling pathwaysrdquo Genes and Development vol 12 no 16 pp2610ndash2622 1998
[26] K AWharton Jr ldquoRunninrsquo with the Dvl proteins that associatewith DshDvl and their significance to Wnt signal transduc-tionrdquo Developmental Biology vol 253 no 1 pp 1ndash17 2003
[27] W J Pan S Z Pang T Huang H Y Guo D Wu and L LildquoCharacterization of function of three domains in dishevelled1 DEP domain is responsible for membrane translocation ofdishevelled-1rdquo Cell Research vol 14 no 4 pp 324ndash330 2004
[28] A Calzado-Martın A Mendez-Vilas M Multigner et al ldquoOnthe role of RhoAROCK signaling in contact guidance ofbone-forming cells on anisotropic Ti6Al4V surfacesrdquo Acta Bio-materialia vol 7 no 4 pp 1890ndash1901 2011
[29] Y Sun L G Villa-Diaz R H Lam W Chen P H Krebsbachand J Fu ldquoMechanics regulates fate decisions of human embry-onic stem cellsrdquo PLoS ONE vol 7 no 5 Article ID e37178 2012
[30] H J Kwon ldquoChondrogenesis on sulfonate-coated hydrogelsis regulated by their mechanical propertiesrdquo Journal of theMechanical Behavior of BiomedicalMaterial vol 17 pp 337ndash3462013
[31] D W Dumbauld H Shin N D Gallant K E Michael HRadhakrishna and A J Garcıa ldquoContractility modulates celladhesion strengthening through focal adhesion kinase andassembly of vinculin-containing focal adhesionsrdquo Journal ofCellular Physiology vol 223 no 3 pp 746ndash756 2010
[32] Y Fonar Y E Gutkovich H Root et al ldquoFocal adhesion kinaseprotein regulates Wnt3a gene expression to control cell fatespecification in the developing neural platerdquoMolecular Biologyof the Cell vol 22 no 13 pp 2409ndash2421 2011
[33] M Despeaux G Chicanne E Rouer et al ldquoFocal adhesionkinase splice variants maintain primitive acute myeloid leuke-mia cells through alteredWnt signalingrdquo Stem Cells vol 30 no8 pp 1597ndash1610 2012
[34] C H Seo K Furukawa K Montagne H Jeong and T UshidaldquoThe effect of substrate microtopography on focal adhesionmaturation and actin organization via the RhoAROCK path-wayrdquo Biomaterials vol 32 no 36 pp 9568ndash9575 2011
[35] B K Teo S T Wong C K Lim et al ldquoNanotopographymodulates mechanotransduction of stem cells and inducesdifferentiation through focal adhesion kinaserdquo ACS Nano 2013
[36] C Higuchi N Nakamura H Yoshikawa and K Itoh ldquoTran-sient dynamic actin cytoskeletal change stimulates the osteo-blastic differentiationrdquo Journal of Bone andMineralMetabolismvol 27 no 2 pp 158ndash167 2009
Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials
BioMed Research International 3
RhoA
Polished000
025
050
075
SLA
OD490to
tal p
rote
in co
nten
tlowast
(a)
TCF-Luc
000
025
050
075
100
Vehicle caRhoA
dnRhoA
Fire
flyR
enill
a lum
ines
cenc
e lowast
PolishedSLA
sect
(b)
Figure 1 (a) Total RhoA activation levels in MC3T3 cells growing on Polished or SLA titanium surfaces by G-LISA assay lowast119875 lt 005 versusPolished (b) Canonical WNT activity by Luciferase reporter assay in C2C12 cells on Polished or SLA surfaces after RhoA inhibition bydominant negative isoform (dnRhoA) or activation by transfection with constitutively active isoform (caRhoA) lowast119875 lt 001 versus vehicle-treated Polished 119875 lt 001 versus vehicle-treated SLA sect119875 lt 001 versus vehicle-treated Polished
slips using an antifade-mounting medium (P7481 MolecularProbes Life Technologies) for photo bleaching reductionSamples were examined using a Nikon Eclipse 90i (NikonTokyo Japan)microscope equipped for fluorescence analysisHeat maps were generated with freeware ImageJ software(NIH Bethesda MD USA) using the inbuilt plugin
29 Statistical Analysis Data were analyzed using Prism 4(GraphPad La Jolla CA USA) All values are reported as themean plusmn Standard Deviation of three repeated experimentsDifferences between group means were evaluated with two-way ANOVA statistical test and Bonferroni posttest anddifferences were considered significant when 119875 lt 005
3 Results
31 Surface Topography Affects Activation Levels of RhoATo investigate whether surface topography could affect theactivation of the small GTPase RhoA we measured RhoAactivity in calvaria osteoblastic MC3T3 cells cultured onsmooth (ldquoPolishedrdquo) or rough (ldquoSLArdquo) titanium discs by G-LISA assay which quantitates the amount of GTP-loadedRhoA The total amount of active RhoA in cells normalizedper protein content was significantly (119875 lt 005) higher incells growing on rough surfaces after 24 hours of culture(Figure 1(a))
32 RhoA Affects Activation of Canonical Wnt SignalingSince RhoA is an important effector of the noncanonicalWnt planar cell polarity pathway we sought to investigatewhether this enzyme could affect the activation of canonicalWnt signaling in C2C12 cells growing on titanium surfacesC2C12 cells are a premyoblast cell line and a popular modelfor Wnt studies because they possess abundant molecular
machinery for this pathway RhoA activation was modu-lated by transiently transfecting C2C12 cells on Polishedor SLA surfaces with dominant negative or constitutivelyactive RhoA isoforms [11] and by cotransfecting them witha reporter system including a plasmid vector carrying aFirefly Luciferase gene under the control of a TCF-responsiveelement and a constitutively active Renilla Luciferase gene fornormalization BackgroundWnt stimulationwas provided byadding 15mLmL recombinant Wnt3a to the supernatantas previously described [5] As previously reported [5] Wntactivation was higher in cells growing on SLA surfaces(Figure 1(b) lowast119875 lt 001) Inhibition of RhoA activity abol-ished canonical Wnt activation on SLA surfaces (119875 lt 001versus vehicle treated SLA) whilst constitutive activationof RhoA slightly but significantly increased the normalizedTCF-Luc signal on Polished (119875 lt 001)These results indicatethat activation of RhoA may directly affect activation ofcanonical Wnt signaling in cells growing on micropatternedtitanium surfaces
33 Cytoskeletal Tension Modulates Cell Responses to SurfaceTopography To better understand the relationship betweencytoskeletal organization and Wnt activation we visualizedthe distribution of Myosin II in MC3T3 cells on Polished orSLA surfaces (Figures 2(a) and 2(b)) Myosin II is a non-muscle isoform of Myosin widely expressed in cells of anyphenotype and is responsible for cell internal tension alsoknown as prestress levels in cell biomechanics [12] Cellsgrowing on smooth or rough titanium discs differed notonly in their shape as it is already well documented in theliterature but in Myosin distribution as well Myosin labelwas stronger in cells on SLA surfaces after 48 hours of cul-ture especially along the cell edges where they adapted tothe underlying peaks and cavities (Figures 2(a) and 2(b))
4 BioMed Research International
Polished
(a)
SLA
(b)
RhoA
00
03
06
09
12
Vehicle Blebbistatin
OD490
lowast
PolishedSLA
(c)
TCF-Luc
000
005
010
015
020
Luci
fera
se ac
tivity
Vehicle Blebbistatin
lowast
PolishedSLA
(d)
Figure 2 (a)-(b) Immunocytochemistry ofMyosin II distribution inMC3T3 cells on Polished or SLA surfaces (20xmagnification bar 20mminsets 40xmagnification) Heatmapswere generated using ImageJ freeware software (c) Total RhoA activation levels inMC3T3 cells growingon Polished or SLA titanium surfaces by G-LISA assay after Myosin II inhibition by 1mM Blebbistatin lowast119875 lt 001 versus vehicle-treated SLA119875 lt 0001 versus Blebbistatin-treated Polished (d) Canonical WNT activity by Luciferase reporter assay in C2C12 cells on Polished or SLAsurfaces after Myosin II inhibition by 1mM Blebbistatin lowast119875 lt 001 versus vehicle-treated Polished and Blebbistatin-treated SLA
Interestingly inhibition of Myosin II activation by 1mMMyosin Light Chain Kinase inhibitor Blebbistatin signifi-cantly increasedRhoAactivity inMC3T3 cells asmeasured byG-LISA assay (Figure 2(c) lowast119875 lt 001 versus vehicle-treatedSLA) As RhoA controls Myosin II activation via the MyosinLight Chain Kinase enzyme this finding suggests a possiblefeedback loop control from Myosin or a tension-sensitivestructure back to RhoAHoweverMyosin inhibition by Bleb-bistatin decreased activation of Wnt canonical signaling inC2C12 cells on SLA surfaces (Figure 2(d) lowast119875 lt 001 versusvehicle treated Polished) in spite of the surge of RhoA activity(Figure 2(c)) thus indicating that cell contractility is requiredfor the increase in canonical Wnt activation observed onrough surfaces and that RhoA control of canonical Wntactivation requires cell tension
Immunofluorescence microscopy (Figures 3(a)ndash3(d))revealed that MC3T3 cells on Polished surfaces had fewerand less evident stress fibers after treatment with Blebbistatinwhile cells on rough surfaces appeared more elongated and
stretched presumably for their inability to retract their rearend during migration across the surface
34 Cytochalasin Increases RhoA Activation and Affects CellDifferentiation on Titanium We then investigated the effectsof microfilament structure on MC3T3 cell responses tomicropatterned titanium surfacesMicrofilaments are a densenetwork of actin fibers that permeate cells and can assemblealong the main tension lines to form stress fibers and wellvisible structures at transmission microscopy (Figure 3(a)arrowheads) These are more evident in cells growing onsmooth surfaces than on rough discs (Figures 3(a) and 3(b))where actin actually appears just as dots or short rodsCytochalasin is well known to disrupt actin polymerizationand 02mM Cytochalasin solution profoundly altered theconformation of the actin cytoskeleton on both titaniumsurfaces at least in part disrupting stress fibers whichappeared fragmented on both surfaces (Figures 3(e) and 3(f))Cytochalasin affected cell shape as well inducing irregular
BioMed Research International 5
Polished
Vehi
cle
(a)
SLA
(b)
Bleb
bista
tin
(c) (d)
Cyto
chal
asin
(e) (f)
Figure 3 Immunocytochemistry microphotographs (40x magnification bar 10mm) of MC3T3 cells growing on Polished (a c e) or SLA (bd f) titanium discs under basal conditions (a b) after addition of Blebbistatin (c d) or Cytochalasin (e f) Arrowheads indicate stress fibers
cell morphologies It has been previously reported thatCytochalasin increases RhoA activation and this was con-firmed in MC3T3 cells on Polished discs (Figure 4(a) lowast119875 lt001 versus vehicle-treated Polished 119875 lt 005 versus vehicle-treated Polished) Cytochalasin also significantly increasedthe activation of canonical Wnt signaling in C2C12 cells inboth titanium groups (Figure 4(b) lowast119875 lt 001 versus vehicle-treated POlished
119875 lt 005 versus vehicle treated SLA)When osteoblastic MC3T3 cells were treated with 02mMCytochalasin in osteogenic medium for 3 days an increase inthe expression of osteoblastic markers Alkaline Phosphatase(lowast119875 lt 005 versus vehicle treated Polished 119875 lt 005 versus
vehicle treated SLA) and Osteocalcin (lowast119875 lt 005 versusvehicle treated SLA) was observed (Figures 4(c)-4(d))
4 Discussion
Although there is a large consensus that rough titanium sur-faces promote higher expression levels of osteoblastic genes invitro and bone formation in vivo the underlyingmechanismsare still only partially known Our group and other researchteams have provided evidence that microtextured surfacescan affect the activation of Wnt canonical signaling [5 713] a pathway required for osteoblastic commitment and
6 BioMed Research International
RhoA
000
025
050
075
Vehicle
OD490to
tal p
rote
in co
nten
t
Cytochalasin
lowast
(a)
TCF-Luc
00
03
06
09
12
Vehicle Cytochalasin
Fire
flyR
enill
a lum
ines
cenc
e
lowast
(b)
lowast
ALP
00
04
08
12
Vehicle Cytochalasin
Relat
ive m
RNA
expr
essio
n
PolishedSLA
(c)
lowastOCN
0
10
20
30
40
Vehicle Cytochalasin
mRN
A re
lativ
e exp
ress
ion
PolishedSLA
(d)
Figure 4 Effects of 02mM Cytochalasin treatment on (a) total RhoA activation levels by G-LISA assay (lowast119875 lt 001 versus vehicle-treatedpolished 119875 lt 005 versus vehicle-treated Polished) in MC3T3 cells and (b) canonical WNT activity by Luciferase reporter assay in C2C12cells growing on Polished or SLA titanium surfaces lowast119875 lt 001 versus vehicle-treated Polished (c d) Real-time PCR analysis of AlkalinePhosphatase (ALP lowast119875 lt 001 versus vehicle-treated Polished) and Osteocalcin (OCN lowast119875 lt 001 versus vehicle-treated SLA) expression inmurine osteoblastic MC3T3 cells on Polished or SLA surfaces after Cytochalasin addition
differentiation [14 15] Understanding how cells can sensethe three-dimensional environment that surrounds them andtransform geometric cues into intracellular signals couldbe the key to create tailored implant surfaces to promotebone anabolism and to support bone formation aroundimplanted devices It is well known that cells can adaptto their culture substrate and conform to its profile cellshape can correspondently tune to the roughness degreeof the surface on which they grow thanks to an adaptablecytoskeleton According to Ingberrsquos tensegrity theory cellsreach a state of mechanical balance that maintains their con-formation by opposing forces [16 17] These are transmittedthrough rigid struts formed by cytoskeletal microtubulesand tensile wires the actin microfilaments anchored to thesubstrate or the extracellular matrix in a tissue throughintegrins [18 19] This structure is under a proper tension
or prestress provided by Myosin motor proteins that actby pulling on actin filaments [20] Myosin is activated bya kinase enzyme known as Myosin Light Chain Kinasewhich is controlled through a signaling cascade by the smallGTPase RhoA one of the target effectors of the noncanonicalWnt planar cell polarity (PCP) pathway [21] This signalingcascade plays an important role in orchestrating cell andtissue organization [22] and is triggered by binding of WNTfactors such asWNT5a to a FrizzledOrphan tyrosine kinaseRor2 receptor complex [23] This recruits the Disheveledprotein (Dvl) to the cell membrane activating it [24] Dvlcomprises three molecular domains which can alternativelytrigger the canonical b-catenin-dependent pathway andother noncanonical pathways such as the PCP pathway[25ndash27] Since cell shape adapts to rough implant surfacesand since the Wnt-related pathways controlling cytoskeletal
BioMed Research International 7
RhoA
MLCK
Actin
Integrins
Myosin II
TCF
Frizzled LRP4-6
Wnt
DvlAxin
Nucleus
Destructioncomplex
Frizzled Ror2C
Wnt
120573cat
120573cat
120573cat
GSK3120573
CK120572
Figure 5 Diagram depicting the proposed model for RhoA role in cell responses to substrate topography Noncanonical Wnt factors bindto receptor dimers Frizzled-Ror2C which activate RhoA Rho in turns activates downstream target Myosin Light Chain Kinase (MLCK)which phosphorylates Myosin II Acto-Myosin filaments create cell tension which facilitates the activation of canonical Wnt signaling andpromotes b catenin release from its destruction complex and translocation to the nucleus possibly by acting on tension sensitive structures(focal adhesions) A feedback loop from tension-activated structures controls RhoA activation levels Actin microfilaments can control RhoAtoo as their partial disruption increases RhoA activity and canonical Wnt activation
organization and osteoblastic commitment share some oftheir key molecular components we hypothesized thatcytoskeletal organization could at least in part cause theactivation of Wnt signaling observed on rough surfacesTo investigate the relation between cytoskeleton and Wntsignaling activation on implant surfaces we quantitatedactive RhoA small GTPase and found that its levels arehigher on rough SLA surfaces according to recent inde-pendent data [28] It can be speculated that cells need torearrange their cytoskeleton through RhoA activation toadapt to the irregular surface profile of SLA discs Interest-ingly hampering RhoA activity by transfecting cells with adominant negative isoformof RhoA (dnRhoA) reducedTCF-mediated transcriptional activity on rough surfaces whichwe previously showed to be higher than on smooth surfacesunder basal conditions [5] This clearly indicates that RhoAactivation is required for the enhancement of Wnt canonicalsignaling on rough surfaces However we also showed thatRhoA-mediated canonical Wnt activation required RhoAdownstream molecular effectors to be functional Myosininhibition by 1mM Blebbistatin did increase RhoA activitypossibly by a negative feedback loop but failed to enhanceWnt canonical signaling On the contrary Wnt activationdropped on SLA surfaces in the presence of Blebbistatinsimilar to what observed with dnRhoA This is consistentwith our observation of a complex Myosin distribution
in cells growing on SLA surfaces (Figures 2(a) and 2(b))presumably maintaining adequate cell adhesion by exertingproper cytoskeletal forces on focal adhesions and it isalso consistent with several reports showing that MyosinII function is required for cell differentiation [29 30] Itcan thus be hypothesized that RhoA enhances canonicalWnt signaling by increasing Myosin activation which inturn is known to activate cytoskeleton-associated signalingmolecules such as focal adhesion-associated Kinases (FAKs)[31] (Figure 5)These have been proven to affect the canonicalWnt signaling pathway in certain in vitro and in vivo models[32 33]This idea is consistent with recent findings indicatingthat focal adhesion formation and FAK phosphorylationare controlled by the RhoA pathway that focal adhesionmaturation and actin polymerization are promoted in cellscultured on micropatterned substrates [34] and that FAKphosphorylation is required for topography-induced differ-entiation of human mesenchymal stem cells [35]
In our study we then treated cells on both surfaceswith Cytochalasin Interestingly cells growing on Polishedsurfaces had thick stress fibers and a well-ordered actinnetwork under basal conditions whilst cells on SLA surfaceswhich do not have a plain uniform surface to adhere onlacked long actin filaments It can be speculated that shorteractin filaments can impart a more variegated array of forcevectors to the cell membrane and cytoplasm to better adapt to
8 BioMed Research International
the correspondently irregular titanium surface underneathCytochalasin at this concentration fragmented the actincytoskeleton by partially inhibiting its polymerization andsignificantly affected cell morphology Cytochalasin alsoincreased RhoA activity as previously reported and cor-respondently increased Wnt activation on both Polishedand SLA discs Under these conditions cell differentiationmarkers increased on both surfaces (Figures 4(c) and 4(d))consistently with reports by Higuchi et al [36]
Taken together our results indicate that cells growing onrough surfaces rearrange their cytoskeleton and this involvesthe activation of the small GTPase RhoA which controlsdownstream effectors that orchestrate cell contractility andshape Moreover RhoA can control and is required for theactivation ofWnt canonical signaling on rough surfacesThiseffect is not exerted directly but through its downstreameffector Myosin II When this is inhibited Wnt activation onrough surfaces is reversed in spite of RhoA activation Ourresults do not identify the specific mechanism of action forMyosin II although it may likely include focal adhesion-associated signaling which is known to be triggered by celltension and contractility Actin filaments can control RhoAactivity and their fragmentation increasesWnt activation andpromotes osteoblast differentiation (Figure 5)
Further studies will be required to better understand howcell contractility allows cells to sense their geometricalmicro-environment but the present study adds another and impor-tant piece of evidence to this elusive yet fundamental puzzlethat sustains the integration of osseointegrated implants inbone
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
S Mazzotta and S Ferrillo contributed equally to the presentstudy
Acknowledgments
The authors would like to thank Straumann Institut AG (andin particular Dr Appert and Dr Molenberg) Basel Switzer-land for kindly providing the titanium surfaces used in thepresent study The authors are also grateful to Dr FrancescaRavanetti and Professor Antonio Cacchioli for their preciousadvices and technical assistance The authors would liketo thank Dr Klaus Hahn for sharing his RhoA isoformsthrough the Addgene plasmid public repository The studywas funded by Grant no 839 2012 from the ITI Foundation(Basel Switzerland)
References
[1] C Galli G Guizzardi G Passeri et al ldquoComparison of humanmandibular osteoblasts grown on two commercially available
titanium implant surfacesrdquo Journal of Periodontology vol 76no 3 pp 364ndash372 2005
[2] B G Keselowsky L Wang Z Schwartz A J Garcia and BD Boyan ldquoIntegrin 1205725 controls osteoblastic proliferation anddifferentiation responses to titanium substrates presenting dif-ferent roughness characteristics in a roughness independentmannerrdquo Journal of BiomedicalMaterials Research A vol 80 no3 pp 700ndash710 2007
[3] I Wall N Donos K Carlqvist F Jones and P Brett ldquoModifiedtitanium surfaces promote accelerated osteogenic differentia-tion of mesenchymal stromal cells in vitrordquo Bone vol 45 no1 pp 17ndash26 2009
[4] K A Kilian B Bugarija B T Lahn and M Mrksich ldquoGeo-metric cues for directing the differentiation of mesenchymalstem cellsrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 11 pp 4872ndash4877 2010
[5] C Galli G Passeri F Ravanetti E Elezi M Pedrazzoni andG M MacAluso ldquoRough surface topography enhances theactivation of Wnt120573-catenin signaling in mesenchymal cellsrdquoJournal of Biomedical Materials Research A vol 95 no 3 pp682ndash690 2010
[6] R Olivares-Navarrete S L Hyzy D L Hutton et al ldquoDirectand indirect effects of microstructured titanium substrates onthe induction of mesenchymal stem cell differentiation towardsthe osteoblast lineagerdquo Biomaterials vol 31 no 10 pp 2728ndash2735 2010
[7] N Chakravorty S Hamlet A Jaiprakash et al ldquoPro-osteogenictopographical cues promote early activation of osteoprogenitordifferentiation via enhanced TGFbeta Wnt and Notch signal-ingrdquo Clinical Oral Implants Research 2013
[8] J Y Martin D D Dean D L Cochran J Simpson B DBoyan and Z Schwartz ldquoProliferation differentiation and pro-tein synthesis of human osteoblast-like cells (MG63) culturedon previously used titanium surfacesrdquo Clinical Oral ImplantsResearch vol 7 no 1 pp 27ndash37 1996
[9] S M Bang H J Moon Y D Kwon J Y Yoo A Pae and I KKwon ldquoOsteoblastic and osteoclastic differentiation on SLAand hydrophilic modified SLA titanium surfacesrdquo Clinical OralImplants Research 2013
[10] S Zinelis N Silikas A Thomas K Syres and G Eliades ldquoSur-face characterization of SLActive dental implantsrdquo EuropeanJournal of Esthetic Dentistry vol 7 no 1 pp 72ndash92 2012
[11] O Pertz L Hodgson R L Klemke and K M Hahn ldquoSpatio-temporal dynamics of RhoA activity in migrating cellsrdquoNaturevol 440 no 7087 pp 1069ndash1072 2006
[12] D Stamenovic and M F Coughlin ldquoThe role of prestress andarchitecture of the cytoskeleton and deformability of cytoskele-tal filaments in mechanics of adherent cells a quantitativeanalysisrdquo Journal of Theoretical Biology vol 201 no 1 pp 63ndash74 1999
[13] R Olivares-Navarrete S L Hyzy J H Park et al ldquoMediationof osteogenic differentiation of human mesenchymal stem cellson titanium surfaces by a Wnt-integrin feedback looprdquo Bioma-terials vol 32 no 27 pp 6399ndash6411 2011
[14] D A Glass II P Bialek J D Ahn et al ldquoCanonical Wnt signal-ing in differentiated osteoblasts controls osteoclast differentia-tionrdquo Developmental Cell vol 8 no 5 pp 751ndash764 2005
[15] S J Rodda and A P McMahon ldquoDistinct roles for Hedgehogand caronicalWnt signaling in specification differentiation andmaintenance of osteoblast progenitorsrdquo Development vol 133no 16 pp 3231ndash3244 2006
BioMed Research International 9
[16] N Wang and D E Ingber ldquoControl of cytoskeletal mechanicsby extracellular matrix cell shape and mechanical tensionrdquoBiophysical Journal vol 66 no 6 pp 2181ndash2189 1994
[17] C Galli S Guizzardi G Passeri G M Macaluso and R Scan-droglio ldquoLife on the wire on tensegrity and force balance incellsrdquo Acta Biomedica de lrsquoAteneo Parmense vol 76 no 1 pp5ndash62 2005
[18] D E Ingber ldquoTensegrity I Cell structure and hierarchical sys-tems biologyrdquo Journal of Cell Science vol 116 no 7 pp 1157ndash11732003
[19] C S Chen M Mrksich S Huang G M Whitesides and D EIngber ldquoGeometric control of cell life and deathrdquo Science vol276 no 5317 pp 1425ndash1428 1997
[20] D Stamenovi S M Mijailovich I M Toli-Noslashrrelykke J Chenand N Wang ldquoCell prestressmdashII Contribution of microtu-bulesrdquo American Journal of Physiology Cell Physiology vol 282no 3 pp C617ndashC624 2002
[21] M Endo M Nishita and Y Minami ldquoAnalysis of wntplanarcell polarity pathway in cultured cellsrdquo Methods in MolecularBiology vol 839 pp 201ndash214 2012
[22] C Gao and Y-G Chen ldquoDishevelled the hub ofWnt signalingrdquoCellular Signalling vol 22 no 5 pp 717ndash727 2010
[23] A J Mikels and R Nusse ldquoPurified Wnt5a protein activatesor inhibits beta-catenin-TCF signaling depending on receptorcontextrdquo PLoS Biology vol 4 no 4 Article ID e115 2006
[24] Y-N Lee Y Gao and H-Y Wang ldquoDifferential mediation ofthe Wnt canonical pathway by mammalian Dishevelleds-1 -2and -3rdquo Cellular Signalling vol 20 no 2 pp 443ndash452 2008
[25] J D Axelrod J R Miller J M Shulman R T Moon andN Perrimon ldquoDifferential recruitment of dishevelled providessignaling specificity in the planar cell polarity and Winglesssignaling pathwaysrdquo Genes and Development vol 12 no 16 pp2610ndash2622 1998
[26] K AWharton Jr ldquoRunninrsquo with the Dvl proteins that associatewith DshDvl and their significance to Wnt signal transduc-tionrdquo Developmental Biology vol 253 no 1 pp 1ndash17 2003
[27] W J Pan S Z Pang T Huang H Y Guo D Wu and L LildquoCharacterization of function of three domains in dishevelled1 DEP domain is responsible for membrane translocation ofdishevelled-1rdquo Cell Research vol 14 no 4 pp 324ndash330 2004
[28] A Calzado-Martın A Mendez-Vilas M Multigner et al ldquoOnthe role of RhoAROCK signaling in contact guidance ofbone-forming cells on anisotropic Ti6Al4V surfacesrdquo Acta Bio-materialia vol 7 no 4 pp 1890ndash1901 2011
[29] Y Sun L G Villa-Diaz R H Lam W Chen P H Krebsbachand J Fu ldquoMechanics regulates fate decisions of human embry-onic stem cellsrdquo PLoS ONE vol 7 no 5 Article ID e37178 2012
[30] H J Kwon ldquoChondrogenesis on sulfonate-coated hydrogelsis regulated by their mechanical propertiesrdquo Journal of theMechanical Behavior of BiomedicalMaterial vol 17 pp 337ndash3462013
[31] D W Dumbauld H Shin N D Gallant K E Michael HRadhakrishna and A J Garcıa ldquoContractility modulates celladhesion strengthening through focal adhesion kinase andassembly of vinculin-containing focal adhesionsrdquo Journal ofCellular Physiology vol 223 no 3 pp 746ndash756 2010
[32] Y Fonar Y E Gutkovich H Root et al ldquoFocal adhesion kinaseprotein regulates Wnt3a gene expression to control cell fatespecification in the developing neural platerdquoMolecular Biologyof the Cell vol 22 no 13 pp 2409ndash2421 2011
[33] M Despeaux G Chicanne E Rouer et al ldquoFocal adhesionkinase splice variants maintain primitive acute myeloid leuke-mia cells through alteredWnt signalingrdquo Stem Cells vol 30 no8 pp 1597ndash1610 2012
[34] C H Seo K Furukawa K Montagne H Jeong and T UshidaldquoThe effect of substrate microtopography on focal adhesionmaturation and actin organization via the RhoAROCK path-wayrdquo Biomaterials vol 32 no 36 pp 9568ndash9575 2011
[35] B K Teo S T Wong C K Lim et al ldquoNanotopographymodulates mechanotransduction of stem cells and inducesdifferentiation through focal adhesion kinaserdquo ACS Nano 2013
[36] C Higuchi N Nakamura H Yoshikawa and K Itoh ldquoTran-sient dynamic actin cytoskeletal change stimulates the osteo-blastic differentiationrdquo Journal of Bone andMineralMetabolismvol 27 no 2 pp 158ndash167 2009
Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials
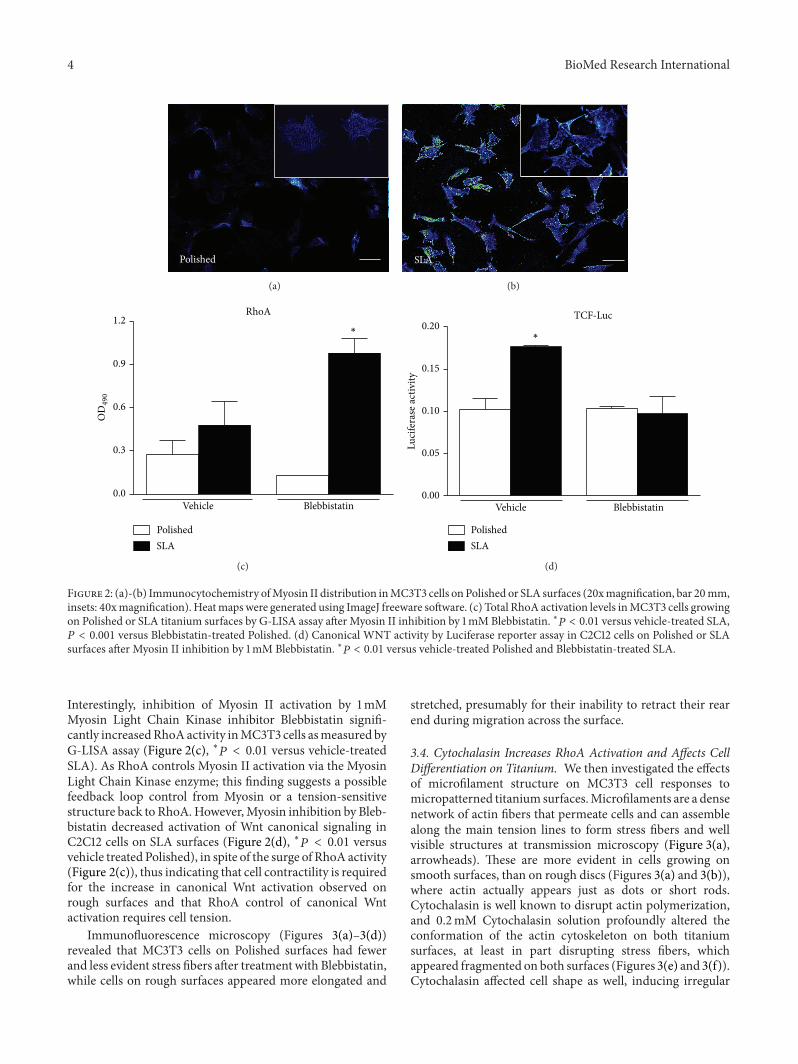
4 BioMed Research International
Polished
(a)
SLA
(b)
RhoA
00
03
06
09
12
Vehicle Blebbistatin
OD490
lowast
PolishedSLA
(c)
TCF-Luc
000
005
010
015
020
Luci
fera
se ac
tivity
Vehicle Blebbistatin
lowast
PolishedSLA
(d)
Figure 2 (a)-(b) Immunocytochemistry ofMyosin II distribution inMC3T3 cells on Polished or SLA surfaces (20xmagnification bar 20mminsets 40xmagnification) Heatmapswere generated using ImageJ freeware software (c) Total RhoA activation levels inMC3T3 cells growingon Polished or SLA titanium surfaces by G-LISA assay after Myosin II inhibition by 1mM Blebbistatin lowast119875 lt 001 versus vehicle-treated SLA119875 lt 0001 versus Blebbistatin-treated Polished (d) Canonical WNT activity by Luciferase reporter assay in C2C12 cells on Polished or SLAsurfaces after Myosin II inhibition by 1mM Blebbistatin lowast119875 lt 001 versus vehicle-treated Polished and Blebbistatin-treated SLA
Interestingly inhibition of Myosin II activation by 1mMMyosin Light Chain Kinase inhibitor Blebbistatin signifi-cantly increasedRhoAactivity inMC3T3 cells asmeasured byG-LISA assay (Figure 2(c) lowast119875 lt 001 versus vehicle-treatedSLA) As RhoA controls Myosin II activation via the MyosinLight Chain Kinase enzyme this finding suggests a possiblefeedback loop control from Myosin or a tension-sensitivestructure back to RhoAHoweverMyosin inhibition by Bleb-bistatin decreased activation of Wnt canonical signaling inC2C12 cells on SLA surfaces (Figure 2(d) lowast119875 lt 001 versusvehicle treated Polished) in spite of the surge of RhoA activity(Figure 2(c)) thus indicating that cell contractility is requiredfor the increase in canonical Wnt activation observed onrough surfaces and that RhoA control of canonical Wntactivation requires cell tension
Immunofluorescence microscopy (Figures 3(a)ndash3(d))revealed that MC3T3 cells on Polished surfaces had fewerand less evident stress fibers after treatment with Blebbistatinwhile cells on rough surfaces appeared more elongated and
stretched presumably for their inability to retract their rearend during migration across the surface
34 Cytochalasin Increases RhoA Activation and Affects CellDifferentiation on Titanium We then investigated the effectsof microfilament structure on MC3T3 cell responses tomicropatterned titanium surfacesMicrofilaments are a densenetwork of actin fibers that permeate cells and can assemblealong the main tension lines to form stress fibers and wellvisible structures at transmission microscopy (Figure 3(a)arrowheads) These are more evident in cells growing onsmooth surfaces than on rough discs (Figures 3(a) and 3(b))where actin actually appears just as dots or short rodsCytochalasin is well known to disrupt actin polymerizationand 02mM Cytochalasin solution profoundly altered theconformation of the actin cytoskeleton on both titaniumsurfaces at least in part disrupting stress fibers whichappeared fragmented on both surfaces (Figures 3(e) and 3(f))Cytochalasin affected cell shape as well inducing irregular
BioMed Research International 5
Polished
Vehi
cle
(a)
SLA
(b)
Bleb
bista
tin
(c) (d)
Cyto
chal
asin
(e) (f)
Figure 3 Immunocytochemistry microphotographs (40x magnification bar 10mm) of MC3T3 cells growing on Polished (a c e) or SLA (bd f) titanium discs under basal conditions (a b) after addition of Blebbistatin (c d) or Cytochalasin (e f) Arrowheads indicate stress fibers
cell morphologies It has been previously reported thatCytochalasin increases RhoA activation and this was con-firmed in MC3T3 cells on Polished discs (Figure 4(a) lowast119875 lt001 versus vehicle-treated Polished 119875 lt 005 versus vehicle-treated Polished) Cytochalasin also significantly increasedthe activation of canonical Wnt signaling in C2C12 cells inboth titanium groups (Figure 4(b) lowast119875 lt 001 versus vehicle-treated POlished
119875 lt 005 versus vehicle treated SLA)When osteoblastic MC3T3 cells were treated with 02mMCytochalasin in osteogenic medium for 3 days an increase inthe expression of osteoblastic markers Alkaline Phosphatase(lowast119875 lt 005 versus vehicle treated Polished 119875 lt 005 versus
vehicle treated SLA) and Osteocalcin (lowast119875 lt 005 versusvehicle treated SLA) was observed (Figures 4(c)-4(d))
4 Discussion
Although there is a large consensus that rough titanium sur-faces promote higher expression levels of osteoblastic genes invitro and bone formation in vivo the underlyingmechanismsare still only partially known Our group and other researchteams have provided evidence that microtextured surfacescan affect the activation of Wnt canonical signaling [5 713] a pathway required for osteoblastic commitment and
6 BioMed Research International
RhoA
000
025
050
075
Vehicle
OD490to
tal p
rote
in co
nten
t
Cytochalasin
lowast
(a)
TCF-Luc
00
03
06
09
12
Vehicle Cytochalasin
Fire
flyR
enill
a lum
ines
cenc
e
lowast
(b)
lowast
ALP
00
04
08
12
Vehicle Cytochalasin
Relat
ive m
RNA
expr
essio
n
PolishedSLA
(c)
lowastOCN
0
10
20
30
40
Vehicle Cytochalasin
mRN
A re
lativ
e exp
ress
ion
PolishedSLA
(d)
Figure 4 Effects of 02mM Cytochalasin treatment on (a) total RhoA activation levels by G-LISA assay (lowast119875 lt 001 versus vehicle-treatedpolished 119875 lt 005 versus vehicle-treated Polished) in MC3T3 cells and (b) canonical WNT activity by Luciferase reporter assay in C2C12cells growing on Polished or SLA titanium surfaces lowast119875 lt 001 versus vehicle-treated Polished (c d) Real-time PCR analysis of AlkalinePhosphatase (ALP lowast119875 lt 001 versus vehicle-treated Polished) and Osteocalcin (OCN lowast119875 lt 001 versus vehicle-treated SLA) expression inmurine osteoblastic MC3T3 cells on Polished or SLA surfaces after Cytochalasin addition
differentiation [14 15] Understanding how cells can sensethe three-dimensional environment that surrounds them andtransform geometric cues into intracellular signals couldbe the key to create tailored implant surfaces to promotebone anabolism and to support bone formation aroundimplanted devices It is well known that cells can adaptto their culture substrate and conform to its profile cellshape can correspondently tune to the roughness degreeof the surface on which they grow thanks to an adaptablecytoskeleton According to Ingberrsquos tensegrity theory cellsreach a state of mechanical balance that maintains their con-formation by opposing forces [16 17] These are transmittedthrough rigid struts formed by cytoskeletal microtubulesand tensile wires the actin microfilaments anchored to thesubstrate or the extracellular matrix in a tissue throughintegrins [18 19] This structure is under a proper tension
or prestress provided by Myosin motor proteins that actby pulling on actin filaments [20] Myosin is activated bya kinase enzyme known as Myosin Light Chain Kinasewhich is controlled through a signaling cascade by the smallGTPase RhoA one of the target effectors of the noncanonicalWnt planar cell polarity (PCP) pathway [21] This signalingcascade plays an important role in orchestrating cell andtissue organization [22] and is triggered by binding of WNTfactors such asWNT5a to a FrizzledOrphan tyrosine kinaseRor2 receptor complex [23] This recruits the Disheveledprotein (Dvl) to the cell membrane activating it [24] Dvlcomprises three molecular domains which can alternativelytrigger the canonical b-catenin-dependent pathway andother noncanonical pathways such as the PCP pathway[25ndash27] Since cell shape adapts to rough implant surfacesand since the Wnt-related pathways controlling cytoskeletal
BioMed Research International 7
RhoA
MLCK
Actin
Integrins
Myosin II
TCF
Frizzled LRP4-6
Wnt
DvlAxin
Nucleus
Destructioncomplex
Frizzled Ror2C
Wnt
120573cat
120573cat
120573cat
GSK3120573
CK120572
Figure 5 Diagram depicting the proposed model for RhoA role in cell responses to substrate topography Noncanonical Wnt factors bindto receptor dimers Frizzled-Ror2C which activate RhoA Rho in turns activates downstream target Myosin Light Chain Kinase (MLCK)which phosphorylates Myosin II Acto-Myosin filaments create cell tension which facilitates the activation of canonical Wnt signaling andpromotes b catenin release from its destruction complex and translocation to the nucleus possibly by acting on tension sensitive structures(focal adhesions) A feedback loop from tension-activated structures controls RhoA activation levels Actin microfilaments can control RhoAtoo as their partial disruption increases RhoA activity and canonical Wnt activation
organization and osteoblastic commitment share some oftheir key molecular components we hypothesized thatcytoskeletal organization could at least in part cause theactivation of Wnt signaling observed on rough surfacesTo investigate the relation between cytoskeleton and Wntsignaling activation on implant surfaces we quantitatedactive RhoA small GTPase and found that its levels arehigher on rough SLA surfaces according to recent inde-pendent data [28] It can be speculated that cells need torearrange their cytoskeleton through RhoA activation toadapt to the irregular surface profile of SLA discs Interest-ingly hampering RhoA activity by transfecting cells with adominant negative isoformof RhoA (dnRhoA) reducedTCF-mediated transcriptional activity on rough surfaces whichwe previously showed to be higher than on smooth surfacesunder basal conditions [5] This clearly indicates that RhoAactivation is required for the enhancement of Wnt canonicalsignaling on rough surfaces However we also showed thatRhoA-mediated canonical Wnt activation required RhoAdownstream molecular effectors to be functional Myosininhibition by 1mM Blebbistatin did increase RhoA activitypossibly by a negative feedback loop but failed to enhanceWnt canonical signaling On the contrary Wnt activationdropped on SLA surfaces in the presence of Blebbistatinsimilar to what observed with dnRhoA This is consistentwith our observation of a complex Myosin distribution
in cells growing on SLA surfaces (Figures 2(a) and 2(b))presumably maintaining adequate cell adhesion by exertingproper cytoskeletal forces on focal adhesions and it isalso consistent with several reports showing that MyosinII function is required for cell differentiation [29 30] Itcan thus be hypothesized that RhoA enhances canonicalWnt signaling by increasing Myosin activation which inturn is known to activate cytoskeleton-associated signalingmolecules such as focal adhesion-associated Kinases (FAKs)[31] (Figure 5)These have been proven to affect the canonicalWnt signaling pathway in certain in vitro and in vivo models[32 33]This idea is consistent with recent findings indicatingthat focal adhesion formation and FAK phosphorylationare controlled by the RhoA pathway that focal adhesionmaturation and actin polymerization are promoted in cellscultured on micropatterned substrates [34] and that FAKphosphorylation is required for topography-induced differ-entiation of human mesenchymal stem cells [35]
In our study we then treated cells on both surfaceswith Cytochalasin Interestingly cells growing on Polishedsurfaces had thick stress fibers and a well-ordered actinnetwork under basal conditions whilst cells on SLA surfaceswhich do not have a plain uniform surface to adhere onlacked long actin filaments It can be speculated that shorteractin filaments can impart a more variegated array of forcevectors to the cell membrane and cytoplasm to better adapt to
8 BioMed Research International
the correspondently irregular titanium surface underneathCytochalasin at this concentration fragmented the actincytoskeleton by partially inhibiting its polymerization andsignificantly affected cell morphology Cytochalasin alsoincreased RhoA activity as previously reported and cor-respondently increased Wnt activation on both Polishedand SLA discs Under these conditions cell differentiationmarkers increased on both surfaces (Figures 4(c) and 4(d))consistently with reports by Higuchi et al [36]
Taken together our results indicate that cells growing onrough surfaces rearrange their cytoskeleton and this involvesthe activation of the small GTPase RhoA which controlsdownstream effectors that orchestrate cell contractility andshape Moreover RhoA can control and is required for theactivation ofWnt canonical signaling on rough surfacesThiseffect is not exerted directly but through its downstreameffector Myosin II When this is inhibited Wnt activation onrough surfaces is reversed in spite of RhoA activation Ourresults do not identify the specific mechanism of action forMyosin II although it may likely include focal adhesion-associated signaling which is known to be triggered by celltension and contractility Actin filaments can control RhoAactivity and their fragmentation increasesWnt activation andpromotes osteoblast differentiation (Figure 5)
Further studies will be required to better understand howcell contractility allows cells to sense their geometricalmicro-environment but the present study adds another and impor-tant piece of evidence to this elusive yet fundamental puzzlethat sustains the integration of osseointegrated implants inbone
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
S Mazzotta and S Ferrillo contributed equally to the presentstudy
Acknowledgments
The authors would like to thank Straumann Institut AG (andin particular Dr Appert and Dr Molenberg) Basel Switzer-land for kindly providing the titanium surfaces used in thepresent study The authors are also grateful to Dr FrancescaRavanetti and Professor Antonio Cacchioli for their preciousadvices and technical assistance The authors would liketo thank Dr Klaus Hahn for sharing his RhoA isoformsthrough the Addgene plasmid public repository The studywas funded by Grant no 839 2012 from the ITI Foundation(Basel Switzerland)
References
[1] C Galli G Guizzardi G Passeri et al ldquoComparison of humanmandibular osteoblasts grown on two commercially available
titanium implant surfacesrdquo Journal of Periodontology vol 76no 3 pp 364ndash372 2005
[2] B G Keselowsky L Wang Z Schwartz A J Garcia and BD Boyan ldquoIntegrin 1205725 controls osteoblastic proliferation anddifferentiation responses to titanium substrates presenting dif-ferent roughness characteristics in a roughness independentmannerrdquo Journal of BiomedicalMaterials Research A vol 80 no3 pp 700ndash710 2007
[3] I Wall N Donos K Carlqvist F Jones and P Brett ldquoModifiedtitanium surfaces promote accelerated osteogenic differentia-tion of mesenchymal stromal cells in vitrordquo Bone vol 45 no1 pp 17ndash26 2009
[4] K A Kilian B Bugarija B T Lahn and M Mrksich ldquoGeo-metric cues for directing the differentiation of mesenchymalstem cellsrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 11 pp 4872ndash4877 2010
[5] C Galli G Passeri F Ravanetti E Elezi M Pedrazzoni andG M MacAluso ldquoRough surface topography enhances theactivation of Wnt120573-catenin signaling in mesenchymal cellsrdquoJournal of Biomedical Materials Research A vol 95 no 3 pp682ndash690 2010
[6] R Olivares-Navarrete S L Hyzy D L Hutton et al ldquoDirectand indirect effects of microstructured titanium substrates onthe induction of mesenchymal stem cell differentiation towardsthe osteoblast lineagerdquo Biomaterials vol 31 no 10 pp 2728ndash2735 2010
[7] N Chakravorty S Hamlet A Jaiprakash et al ldquoPro-osteogenictopographical cues promote early activation of osteoprogenitordifferentiation via enhanced TGFbeta Wnt and Notch signal-ingrdquo Clinical Oral Implants Research 2013
[8] J Y Martin D D Dean D L Cochran J Simpson B DBoyan and Z Schwartz ldquoProliferation differentiation and pro-tein synthesis of human osteoblast-like cells (MG63) culturedon previously used titanium surfacesrdquo Clinical Oral ImplantsResearch vol 7 no 1 pp 27ndash37 1996
[9] S M Bang H J Moon Y D Kwon J Y Yoo A Pae and I KKwon ldquoOsteoblastic and osteoclastic differentiation on SLAand hydrophilic modified SLA titanium surfacesrdquo Clinical OralImplants Research 2013
[10] S Zinelis N Silikas A Thomas K Syres and G Eliades ldquoSur-face characterization of SLActive dental implantsrdquo EuropeanJournal of Esthetic Dentistry vol 7 no 1 pp 72ndash92 2012
[11] O Pertz L Hodgson R L Klemke and K M Hahn ldquoSpatio-temporal dynamics of RhoA activity in migrating cellsrdquoNaturevol 440 no 7087 pp 1069ndash1072 2006
[12] D Stamenovic and M F Coughlin ldquoThe role of prestress andarchitecture of the cytoskeleton and deformability of cytoskele-tal filaments in mechanics of adherent cells a quantitativeanalysisrdquo Journal of Theoretical Biology vol 201 no 1 pp 63ndash74 1999
[13] R Olivares-Navarrete S L Hyzy J H Park et al ldquoMediationof osteogenic differentiation of human mesenchymal stem cellson titanium surfaces by a Wnt-integrin feedback looprdquo Bioma-terials vol 32 no 27 pp 6399ndash6411 2011
[14] D A Glass II P Bialek J D Ahn et al ldquoCanonical Wnt signal-ing in differentiated osteoblasts controls osteoclast differentia-tionrdquo Developmental Cell vol 8 no 5 pp 751ndash764 2005
[15] S J Rodda and A P McMahon ldquoDistinct roles for Hedgehogand caronicalWnt signaling in specification differentiation andmaintenance of osteoblast progenitorsrdquo Development vol 133no 16 pp 3231ndash3244 2006
BioMed Research International 9
[16] N Wang and D E Ingber ldquoControl of cytoskeletal mechanicsby extracellular matrix cell shape and mechanical tensionrdquoBiophysical Journal vol 66 no 6 pp 2181ndash2189 1994
[17] C Galli S Guizzardi G Passeri G M Macaluso and R Scan-droglio ldquoLife on the wire on tensegrity and force balance incellsrdquo Acta Biomedica de lrsquoAteneo Parmense vol 76 no 1 pp5ndash62 2005
[18] D E Ingber ldquoTensegrity I Cell structure and hierarchical sys-tems biologyrdquo Journal of Cell Science vol 116 no 7 pp 1157ndash11732003
[19] C S Chen M Mrksich S Huang G M Whitesides and D EIngber ldquoGeometric control of cell life and deathrdquo Science vol276 no 5317 pp 1425ndash1428 1997
[20] D Stamenovi S M Mijailovich I M Toli-Noslashrrelykke J Chenand N Wang ldquoCell prestressmdashII Contribution of microtu-bulesrdquo American Journal of Physiology Cell Physiology vol 282no 3 pp C617ndashC624 2002
[21] M Endo M Nishita and Y Minami ldquoAnalysis of wntplanarcell polarity pathway in cultured cellsrdquo Methods in MolecularBiology vol 839 pp 201ndash214 2012
[22] C Gao and Y-G Chen ldquoDishevelled the hub ofWnt signalingrdquoCellular Signalling vol 22 no 5 pp 717ndash727 2010
[23] A J Mikels and R Nusse ldquoPurified Wnt5a protein activatesor inhibits beta-catenin-TCF signaling depending on receptorcontextrdquo PLoS Biology vol 4 no 4 Article ID e115 2006
[24] Y-N Lee Y Gao and H-Y Wang ldquoDifferential mediation ofthe Wnt canonical pathway by mammalian Dishevelleds-1 -2and -3rdquo Cellular Signalling vol 20 no 2 pp 443ndash452 2008
[25] J D Axelrod J R Miller J M Shulman R T Moon andN Perrimon ldquoDifferential recruitment of dishevelled providessignaling specificity in the planar cell polarity and Winglesssignaling pathwaysrdquo Genes and Development vol 12 no 16 pp2610ndash2622 1998
[26] K AWharton Jr ldquoRunninrsquo with the Dvl proteins that associatewith DshDvl and their significance to Wnt signal transduc-tionrdquo Developmental Biology vol 253 no 1 pp 1ndash17 2003
[27] W J Pan S Z Pang T Huang H Y Guo D Wu and L LildquoCharacterization of function of three domains in dishevelled1 DEP domain is responsible for membrane translocation ofdishevelled-1rdquo Cell Research vol 14 no 4 pp 324ndash330 2004
[28] A Calzado-Martın A Mendez-Vilas M Multigner et al ldquoOnthe role of RhoAROCK signaling in contact guidance ofbone-forming cells on anisotropic Ti6Al4V surfacesrdquo Acta Bio-materialia vol 7 no 4 pp 1890ndash1901 2011
[29] Y Sun L G Villa-Diaz R H Lam W Chen P H Krebsbachand J Fu ldquoMechanics regulates fate decisions of human embry-onic stem cellsrdquo PLoS ONE vol 7 no 5 Article ID e37178 2012
[30] H J Kwon ldquoChondrogenesis on sulfonate-coated hydrogelsis regulated by their mechanical propertiesrdquo Journal of theMechanical Behavior of BiomedicalMaterial vol 17 pp 337ndash3462013
[31] D W Dumbauld H Shin N D Gallant K E Michael HRadhakrishna and A J Garcıa ldquoContractility modulates celladhesion strengthening through focal adhesion kinase andassembly of vinculin-containing focal adhesionsrdquo Journal ofCellular Physiology vol 223 no 3 pp 746ndash756 2010
[32] Y Fonar Y E Gutkovich H Root et al ldquoFocal adhesion kinaseprotein regulates Wnt3a gene expression to control cell fatespecification in the developing neural platerdquoMolecular Biologyof the Cell vol 22 no 13 pp 2409ndash2421 2011
[33] M Despeaux G Chicanne E Rouer et al ldquoFocal adhesionkinase splice variants maintain primitive acute myeloid leuke-mia cells through alteredWnt signalingrdquo Stem Cells vol 30 no8 pp 1597ndash1610 2012
[34] C H Seo K Furukawa K Montagne H Jeong and T UshidaldquoThe effect of substrate microtopography on focal adhesionmaturation and actin organization via the RhoAROCK path-wayrdquo Biomaterials vol 32 no 36 pp 9568ndash9575 2011
[35] B K Teo S T Wong C K Lim et al ldquoNanotopographymodulates mechanotransduction of stem cells and inducesdifferentiation through focal adhesion kinaserdquo ACS Nano 2013
[36] C Higuchi N Nakamura H Yoshikawa and K Itoh ldquoTran-sient dynamic actin cytoskeletal change stimulates the osteo-blastic differentiationrdquo Journal of Bone andMineralMetabolismvol 27 no 2 pp 158ndash167 2009
Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials
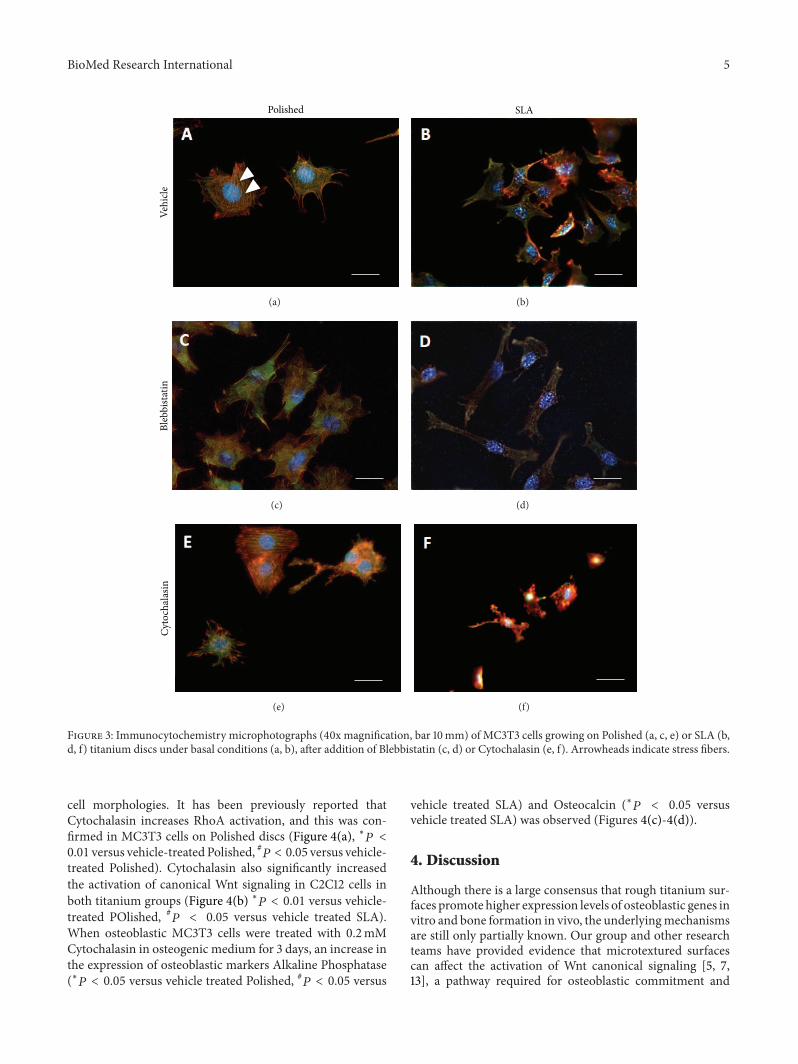
BioMed Research International 5
Polished
Vehi
cle
(a)
SLA
(b)
Bleb
bista
tin
(c) (d)
Cyto
chal
asin
(e) (f)
Figure 3 Immunocytochemistry microphotographs (40x magnification bar 10mm) of MC3T3 cells growing on Polished (a c e) or SLA (bd f) titanium discs under basal conditions (a b) after addition of Blebbistatin (c d) or Cytochalasin (e f) Arrowheads indicate stress fibers
cell morphologies It has been previously reported thatCytochalasin increases RhoA activation and this was con-firmed in MC3T3 cells on Polished discs (Figure 4(a) lowast119875 lt001 versus vehicle-treated Polished 119875 lt 005 versus vehicle-treated Polished) Cytochalasin also significantly increasedthe activation of canonical Wnt signaling in C2C12 cells inboth titanium groups (Figure 4(b) lowast119875 lt 001 versus vehicle-treated POlished
119875 lt 005 versus vehicle treated SLA)When osteoblastic MC3T3 cells were treated with 02mMCytochalasin in osteogenic medium for 3 days an increase inthe expression of osteoblastic markers Alkaline Phosphatase(lowast119875 lt 005 versus vehicle treated Polished 119875 lt 005 versus
vehicle treated SLA) and Osteocalcin (lowast119875 lt 005 versusvehicle treated SLA) was observed (Figures 4(c)-4(d))
4 Discussion
Although there is a large consensus that rough titanium sur-faces promote higher expression levels of osteoblastic genes invitro and bone formation in vivo the underlyingmechanismsare still only partially known Our group and other researchteams have provided evidence that microtextured surfacescan affect the activation of Wnt canonical signaling [5 713] a pathway required for osteoblastic commitment and
6 BioMed Research International
RhoA
000
025
050
075
Vehicle
OD490to
tal p
rote
in co
nten
t
Cytochalasin
lowast
(a)
TCF-Luc
00
03
06
09
12
Vehicle Cytochalasin
Fire
flyR
enill
a lum
ines
cenc
e
lowast
(b)
lowast
ALP
00
04
08
12
Vehicle Cytochalasin
Relat
ive m
RNA
expr
essio
n
PolishedSLA
(c)
lowastOCN
0
10
20
30
40
Vehicle Cytochalasin
mRN
A re
lativ
e exp
ress
ion
PolishedSLA
(d)
Figure 4 Effects of 02mM Cytochalasin treatment on (a) total RhoA activation levels by G-LISA assay (lowast119875 lt 001 versus vehicle-treatedpolished 119875 lt 005 versus vehicle-treated Polished) in MC3T3 cells and (b) canonical WNT activity by Luciferase reporter assay in C2C12cells growing on Polished or SLA titanium surfaces lowast119875 lt 001 versus vehicle-treated Polished (c d) Real-time PCR analysis of AlkalinePhosphatase (ALP lowast119875 lt 001 versus vehicle-treated Polished) and Osteocalcin (OCN lowast119875 lt 001 versus vehicle-treated SLA) expression inmurine osteoblastic MC3T3 cells on Polished or SLA surfaces after Cytochalasin addition
differentiation [14 15] Understanding how cells can sensethe three-dimensional environment that surrounds them andtransform geometric cues into intracellular signals couldbe the key to create tailored implant surfaces to promotebone anabolism and to support bone formation aroundimplanted devices It is well known that cells can adaptto their culture substrate and conform to its profile cellshape can correspondently tune to the roughness degreeof the surface on which they grow thanks to an adaptablecytoskeleton According to Ingberrsquos tensegrity theory cellsreach a state of mechanical balance that maintains their con-formation by opposing forces [16 17] These are transmittedthrough rigid struts formed by cytoskeletal microtubulesand tensile wires the actin microfilaments anchored to thesubstrate or the extracellular matrix in a tissue throughintegrins [18 19] This structure is under a proper tension
or prestress provided by Myosin motor proteins that actby pulling on actin filaments [20] Myosin is activated bya kinase enzyme known as Myosin Light Chain Kinasewhich is controlled through a signaling cascade by the smallGTPase RhoA one of the target effectors of the noncanonicalWnt planar cell polarity (PCP) pathway [21] This signalingcascade plays an important role in orchestrating cell andtissue organization [22] and is triggered by binding of WNTfactors such asWNT5a to a FrizzledOrphan tyrosine kinaseRor2 receptor complex [23] This recruits the Disheveledprotein (Dvl) to the cell membrane activating it [24] Dvlcomprises three molecular domains which can alternativelytrigger the canonical b-catenin-dependent pathway andother noncanonical pathways such as the PCP pathway[25ndash27] Since cell shape adapts to rough implant surfacesand since the Wnt-related pathways controlling cytoskeletal
BioMed Research International 7
RhoA
MLCK
Actin
Integrins
Myosin II
TCF
Frizzled LRP4-6
Wnt
DvlAxin
Nucleus
Destructioncomplex
Frizzled Ror2C
Wnt
120573cat
120573cat
120573cat
GSK3120573
CK120572
Figure 5 Diagram depicting the proposed model for RhoA role in cell responses to substrate topography Noncanonical Wnt factors bindto receptor dimers Frizzled-Ror2C which activate RhoA Rho in turns activates downstream target Myosin Light Chain Kinase (MLCK)which phosphorylates Myosin II Acto-Myosin filaments create cell tension which facilitates the activation of canonical Wnt signaling andpromotes b catenin release from its destruction complex and translocation to the nucleus possibly by acting on tension sensitive structures(focal adhesions) A feedback loop from tension-activated structures controls RhoA activation levels Actin microfilaments can control RhoAtoo as their partial disruption increases RhoA activity and canonical Wnt activation
organization and osteoblastic commitment share some oftheir key molecular components we hypothesized thatcytoskeletal organization could at least in part cause theactivation of Wnt signaling observed on rough surfacesTo investigate the relation between cytoskeleton and Wntsignaling activation on implant surfaces we quantitatedactive RhoA small GTPase and found that its levels arehigher on rough SLA surfaces according to recent inde-pendent data [28] It can be speculated that cells need torearrange their cytoskeleton through RhoA activation toadapt to the irregular surface profile of SLA discs Interest-ingly hampering RhoA activity by transfecting cells with adominant negative isoformof RhoA (dnRhoA) reducedTCF-mediated transcriptional activity on rough surfaces whichwe previously showed to be higher than on smooth surfacesunder basal conditions [5] This clearly indicates that RhoAactivation is required for the enhancement of Wnt canonicalsignaling on rough surfaces However we also showed thatRhoA-mediated canonical Wnt activation required RhoAdownstream molecular effectors to be functional Myosininhibition by 1mM Blebbistatin did increase RhoA activitypossibly by a negative feedback loop but failed to enhanceWnt canonical signaling On the contrary Wnt activationdropped on SLA surfaces in the presence of Blebbistatinsimilar to what observed with dnRhoA This is consistentwith our observation of a complex Myosin distribution
in cells growing on SLA surfaces (Figures 2(a) and 2(b))presumably maintaining adequate cell adhesion by exertingproper cytoskeletal forces on focal adhesions and it isalso consistent with several reports showing that MyosinII function is required for cell differentiation [29 30] Itcan thus be hypothesized that RhoA enhances canonicalWnt signaling by increasing Myosin activation which inturn is known to activate cytoskeleton-associated signalingmolecules such as focal adhesion-associated Kinases (FAKs)[31] (Figure 5)These have been proven to affect the canonicalWnt signaling pathway in certain in vitro and in vivo models[32 33]This idea is consistent with recent findings indicatingthat focal adhesion formation and FAK phosphorylationare controlled by the RhoA pathway that focal adhesionmaturation and actin polymerization are promoted in cellscultured on micropatterned substrates [34] and that FAKphosphorylation is required for topography-induced differ-entiation of human mesenchymal stem cells [35]
In our study we then treated cells on both surfaceswith Cytochalasin Interestingly cells growing on Polishedsurfaces had thick stress fibers and a well-ordered actinnetwork under basal conditions whilst cells on SLA surfaceswhich do not have a plain uniform surface to adhere onlacked long actin filaments It can be speculated that shorteractin filaments can impart a more variegated array of forcevectors to the cell membrane and cytoplasm to better adapt to
8 BioMed Research International
the correspondently irregular titanium surface underneathCytochalasin at this concentration fragmented the actincytoskeleton by partially inhibiting its polymerization andsignificantly affected cell morphology Cytochalasin alsoincreased RhoA activity as previously reported and cor-respondently increased Wnt activation on both Polishedand SLA discs Under these conditions cell differentiationmarkers increased on both surfaces (Figures 4(c) and 4(d))consistently with reports by Higuchi et al [36]
Taken together our results indicate that cells growing onrough surfaces rearrange their cytoskeleton and this involvesthe activation of the small GTPase RhoA which controlsdownstream effectors that orchestrate cell contractility andshape Moreover RhoA can control and is required for theactivation ofWnt canonical signaling on rough surfacesThiseffect is not exerted directly but through its downstreameffector Myosin II When this is inhibited Wnt activation onrough surfaces is reversed in spite of RhoA activation Ourresults do not identify the specific mechanism of action forMyosin II although it may likely include focal adhesion-associated signaling which is known to be triggered by celltension and contractility Actin filaments can control RhoAactivity and their fragmentation increasesWnt activation andpromotes osteoblast differentiation (Figure 5)
Further studies will be required to better understand howcell contractility allows cells to sense their geometricalmicro-environment but the present study adds another and impor-tant piece of evidence to this elusive yet fundamental puzzlethat sustains the integration of osseointegrated implants inbone
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
S Mazzotta and S Ferrillo contributed equally to the presentstudy
Acknowledgments
The authors would like to thank Straumann Institut AG (andin particular Dr Appert and Dr Molenberg) Basel Switzer-land for kindly providing the titanium surfaces used in thepresent study The authors are also grateful to Dr FrancescaRavanetti and Professor Antonio Cacchioli for their preciousadvices and technical assistance The authors would liketo thank Dr Klaus Hahn for sharing his RhoA isoformsthrough the Addgene plasmid public repository The studywas funded by Grant no 839 2012 from the ITI Foundation(Basel Switzerland)
References
[1] C Galli G Guizzardi G Passeri et al ldquoComparison of humanmandibular osteoblasts grown on two commercially available
titanium implant surfacesrdquo Journal of Periodontology vol 76no 3 pp 364ndash372 2005
[2] B G Keselowsky L Wang Z Schwartz A J Garcia and BD Boyan ldquoIntegrin 1205725 controls osteoblastic proliferation anddifferentiation responses to titanium substrates presenting dif-ferent roughness characteristics in a roughness independentmannerrdquo Journal of BiomedicalMaterials Research A vol 80 no3 pp 700ndash710 2007
[3] I Wall N Donos K Carlqvist F Jones and P Brett ldquoModifiedtitanium surfaces promote accelerated osteogenic differentia-tion of mesenchymal stromal cells in vitrordquo Bone vol 45 no1 pp 17ndash26 2009
[4] K A Kilian B Bugarija B T Lahn and M Mrksich ldquoGeo-metric cues for directing the differentiation of mesenchymalstem cellsrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 11 pp 4872ndash4877 2010
[5] C Galli G Passeri F Ravanetti E Elezi M Pedrazzoni andG M MacAluso ldquoRough surface topography enhances theactivation of Wnt120573-catenin signaling in mesenchymal cellsrdquoJournal of Biomedical Materials Research A vol 95 no 3 pp682ndash690 2010
[6] R Olivares-Navarrete S L Hyzy D L Hutton et al ldquoDirectand indirect effects of microstructured titanium substrates onthe induction of mesenchymal stem cell differentiation towardsthe osteoblast lineagerdquo Biomaterials vol 31 no 10 pp 2728ndash2735 2010
[7] N Chakravorty S Hamlet A Jaiprakash et al ldquoPro-osteogenictopographical cues promote early activation of osteoprogenitordifferentiation via enhanced TGFbeta Wnt and Notch signal-ingrdquo Clinical Oral Implants Research 2013
[8] J Y Martin D D Dean D L Cochran J Simpson B DBoyan and Z Schwartz ldquoProliferation differentiation and pro-tein synthesis of human osteoblast-like cells (MG63) culturedon previously used titanium surfacesrdquo Clinical Oral ImplantsResearch vol 7 no 1 pp 27ndash37 1996
[9] S M Bang H J Moon Y D Kwon J Y Yoo A Pae and I KKwon ldquoOsteoblastic and osteoclastic differentiation on SLAand hydrophilic modified SLA titanium surfacesrdquo Clinical OralImplants Research 2013
[10] S Zinelis N Silikas A Thomas K Syres and G Eliades ldquoSur-face characterization of SLActive dental implantsrdquo EuropeanJournal of Esthetic Dentistry vol 7 no 1 pp 72ndash92 2012
[11] O Pertz L Hodgson R L Klemke and K M Hahn ldquoSpatio-temporal dynamics of RhoA activity in migrating cellsrdquoNaturevol 440 no 7087 pp 1069ndash1072 2006
[12] D Stamenovic and M F Coughlin ldquoThe role of prestress andarchitecture of the cytoskeleton and deformability of cytoskele-tal filaments in mechanics of adherent cells a quantitativeanalysisrdquo Journal of Theoretical Biology vol 201 no 1 pp 63ndash74 1999
[13] R Olivares-Navarrete S L Hyzy J H Park et al ldquoMediationof osteogenic differentiation of human mesenchymal stem cellson titanium surfaces by a Wnt-integrin feedback looprdquo Bioma-terials vol 32 no 27 pp 6399ndash6411 2011
[14] D A Glass II P Bialek J D Ahn et al ldquoCanonical Wnt signal-ing in differentiated osteoblasts controls osteoclast differentia-tionrdquo Developmental Cell vol 8 no 5 pp 751ndash764 2005
[15] S J Rodda and A P McMahon ldquoDistinct roles for Hedgehogand caronicalWnt signaling in specification differentiation andmaintenance of osteoblast progenitorsrdquo Development vol 133no 16 pp 3231ndash3244 2006
BioMed Research International 9
[16] N Wang and D E Ingber ldquoControl of cytoskeletal mechanicsby extracellular matrix cell shape and mechanical tensionrdquoBiophysical Journal vol 66 no 6 pp 2181ndash2189 1994
[17] C Galli S Guizzardi G Passeri G M Macaluso and R Scan-droglio ldquoLife on the wire on tensegrity and force balance incellsrdquo Acta Biomedica de lrsquoAteneo Parmense vol 76 no 1 pp5ndash62 2005
[18] D E Ingber ldquoTensegrity I Cell structure and hierarchical sys-tems biologyrdquo Journal of Cell Science vol 116 no 7 pp 1157ndash11732003
[19] C S Chen M Mrksich S Huang G M Whitesides and D EIngber ldquoGeometric control of cell life and deathrdquo Science vol276 no 5317 pp 1425ndash1428 1997
[20] D Stamenovi S M Mijailovich I M Toli-Noslashrrelykke J Chenand N Wang ldquoCell prestressmdashII Contribution of microtu-bulesrdquo American Journal of Physiology Cell Physiology vol 282no 3 pp C617ndashC624 2002
[21] M Endo M Nishita and Y Minami ldquoAnalysis of wntplanarcell polarity pathway in cultured cellsrdquo Methods in MolecularBiology vol 839 pp 201ndash214 2012
[22] C Gao and Y-G Chen ldquoDishevelled the hub ofWnt signalingrdquoCellular Signalling vol 22 no 5 pp 717ndash727 2010
[23] A J Mikels and R Nusse ldquoPurified Wnt5a protein activatesor inhibits beta-catenin-TCF signaling depending on receptorcontextrdquo PLoS Biology vol 4 no 4 Article ID e115 2006
[24] Y-N Lee Y Gao and H-Y Wang ldquoDifferential mediation ofthe Wnt canonical pathway by mammalian Dishevelleds-1 -2and -3rdquo Cellular Signalling vol 20 no 2 pp 443ndash452 2008
[25] J D Axelrod J R Miller J M Shulman R T Moon andN Perrimon ldquoDifferential recruitment of dishevelled providessignaling specificity in the planar cell polarity and Winglesssignaling pathwaysrdquo Genes and Development vol 12 no 16 pp2610ndash2622 1998
[26] K AWharton Jr ldquoRunninrsquo with the Dvl proteins that associatewith DshDvl and their significance to Wnt signal transduc-tionrdquo Developmental Biology vol 253 no 1 pp 1ndash17 2003
[27] W J Pan S Z Pang T Huang H Y Guo D Wu and L LildquoCharacterization of function of three domains in dishevelled1 DEP domain is responsible for membrane translocation ofdishevelled-1rdquo Cell Research vol 14 no 4 pp 324ndash330 2004
[28] A Calzado-Martın A Mendez-Vilas M Multigner et al ldquoOnthe role of RhoAROCK signaling in contact guidance ofbone-forming cells on anisotropic Ti6Al4V surfacesrdquo Acta Bio-materialia vol 7 no 4 pp 1890ndash1901 2011
[29] Y Sun L G Villa-Diaz R H Lam W Chen P H Krebsbachand J Fu ldquoMechanics regulates fate decisions of human embry-onic stem cellsrdquo PLoS ONE vol 7 no 5 Article ID e37178 2012
[30] H J Kwon ldquoChondrogenesis on sulfonate-coated hydrogelsis regulated by their mechanical propertiesrdquo Journal of theMechanical Behavior of BiomedicalMaterial vol 17 pp 337ndash3462013
[31] D W Dumbauld H Shin N D Gallant K E Michael HRadhakrishna and A J Garcıa ldquoContractility modulates celladhesion strengthening through focal adhesion kinase andassembly of vinculin-containing focal adhesionsrdquo Journal ofCellular Physiology vol 223 no 3 pp 746ndash756 2010
[32] Y Fonar Y E Gutkovich H Root et al ldquoFocal adhesion kinaseprotein regulates Wnt3a gene expression to control cell fatespecification in the developing neural platerdquoMolecular Biologyof the Cell vol 22 no 13 pp 2409ndash2421 2011
[33] M Despeaux G Chicanne E Rouer et al ldquoFocal adhesionkinase splice variants maintain primitive acute myeloid leuke-mia cells through alteredWnt signalingrdquo Stem Cells vol 30 no8 pp 1597ndash1610 2012
[34] C H Seo K Furukawa K Montagne H Jeong and T UshidaldquoThe effect of substrate microtopography on focal adhesionmaturation and actin organization via the RhoAROCK path-wayrdquo Biomaterials vol 32 no 36 pp 9568ndash9575 2011
[35] B K Teo S T Wong C K Lim et al ldquoNanotopographymodulates mechanotransduction of stem cells and inducesdifferentiation through focal adhesion kinaserdquo ACS Nano 2013
[36] C Higuchi N Nakamura H Yoshikawa and K Itoh ldquoTran-sient dynamic actin cytoskeletal change stimulates the osteo-blastic differentiationrdquo Journal of Bone andMineralMetabolismvol 27 no 2 pp 158ndash167 2009
Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials
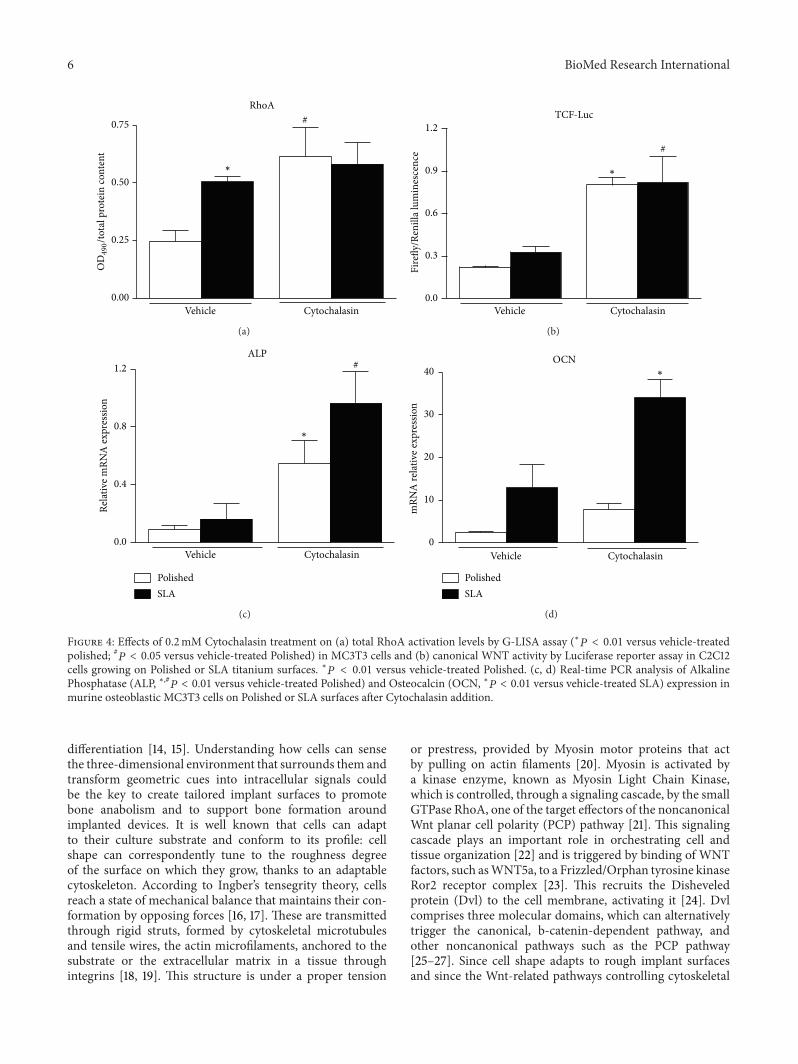
6 BioMed Research International
RhoA
000
025
050
075
Vehicle
OD490to
tal p
rote
in co
nten
t
Cytochalasin
lowast
(a)
TCF-Luc
00
03
06
09
12
Vehicle Cytochalasin
Fire
flyR
enill
a lum
ines
cenc
e
lowast
(b)
lowast
ALP
00
04
08
12
Vehicle Cytochalasin
Relat
ive m
RNA
expr
essio
n
PolishedSLA
(c)
lowastOCN
0
10
20
30
40
Vehicle Cytochalasin
mRN
A re
lativ
e exp
ress
ion
PolishedSLA
(d)
Figure 4 Effects of 02mM Cytochalasin treatment on (a) total RhoA activation levels by G-LISA assay (lowast119875 lt 001 versus vehicle-treatedpolished 119875 lt 005 versus vehicle-treated Polished) in MC3T3 cells and (b) canonical WNT activity by Luciferase reporter assay in C2C12cells growing on Polished or SLA titanium surfaces lowast119875 lt 001 versus vehicle-treated Polished (c d) Real-time PCR analysis of AlkalinePhosphatase (ALP lowast119875 lt 001 versus vehicle-treated Polished) and Osteocalcin (OCN lowast119875 lt 001 versus vehicle-treated SLA) expression inmurine osteoblastic MC3T3 cells on Polished or SLA surfaces after Cytochalasin addition
differentiation [14 15] Understanding how cells can sensethe three-dimensional environment that surrounds them andtransform geometric cues into intracellular signals couldbe the key to create tailored implant surfaces to promotebone anabolism and to support bone formation aroundimplanted devices It is well known that cells can adaptto their culture substrate and conform to its profile cellshape can correspondently tune to the roughness degreeof the surface on which they grow thanks to an adaptablecytoskeleton According to Ingberrsquos tensegrity theory cellsreach a state of mechanical balance that maintains their con-formation by opposing forces [16 17] These are transmittedthrough rigid struts formed by cytoskeletal microtubulesand tensile wires the actin microfilaments anchored to thesubstrate or the extracellular matrix in a tissue throughintegrins [18 19] This structure is under a proper tension
or prestress provided by Myosin motor proteins that actby pulling on actin filaments [20] Myosin is activated bya kinase enzyme known as Myosin Light Chain Kinasewhich is controlled through a signaling cascade by the smallGTPase RhoA one of the target effectors of the noncanonicalWnt planar cell polarity (PCP) pathway [21] This signalingcascade plays an important role in orchestrating cell andtissue organization [22] and is triggered by binding of WNTfactors such asWNT5a to a FrizzledOrphan tyrosine kinaseRor2 receptor complex [23] This recruits the Disheveledprotein (Dvl) to the cell membrane activating it [24] Dvlcomprises three molecular domains which can alternativelytrigger the canonical b-catenin-dependent pathway andother noncanonical pathways such as the PCP pathway[25ndash27] Since cell shape adapts to rough implant surfacesand since the Wnt-related pathways controlling cytoskeletal
BioMed Research International 7
RhoA
MLCK
Actin
Integrins
Myosin II
TCF
Frizzled LRP4-6
Wnt
DvlAxin
Nucleus
Destructioncomplex
Frizzled Ror2C
Wnt
120573cat
120573cat
120573cat
GSK3120573
CK120572
Figure 5 Diagram depicting the proposed model for RhoA role in cell responses to substrate topography Noncanonical Wnt factors bindto receptor dimers Frizzled-Ror2C which activate RhoA Rho in turns activates downstream target Myosin Light Chain Kinase (MLCK)which phosphorylates Myosin II Acto-Myosin filaments create cell tension which facilitates the activation of canonical Wnt signaling andpromotes b catenin release from its destruction complex and translocation to the nucleus possibly by acting on tension sensitive structures(focal adhesions) A feedback loop from tension-activated structures controls RhoA activation levels Actin microfilaments can control RhoAtoo as their partial disruption increases RhoA activity and canonical Wnt activation
organization and osteoblastic commitment share some oftheir key molecular components we hypothesized thatcytoskeletal organization could at least in part cause theactivation of Wnt signaling observed on rough surfacesTo investigate the relation between cytoskeleton and Wntsignaling activation on implant surfaces we quantitatedactive RhoA small GTPase and found that its levels arehigher on rough SLA surfaces according to recent inde-pendent data [28] It can be speculated that cells need torearrange their cytoskeleton through RhoA activation toadapt to the irregular surface profile of SLA discs Interest-ingly hampering RhoA activity by transfecting cells with adominant negative isoformof RhoA (dnRhoA) reducedTCF-mediated transcriptional activity on rough surfaces whichwe previously showed to be higher than on smooth surfacesunder basal conditions [5] This clearly indicates that RhoAactivation is required for the enhancement of Wnt canonicalsignaling on rough surfaces However we also showed thatRhoA-mediated canonical Wnt activation required RhoAdownstream molecular effectors to be functional Myosininhibition by 1mM Blebbistatin did increase RhoA activitypossibly by a negative feedback loop but failed to enhanceWnt canonical signaling On the contrary Wnt activationdropped on SLA surfaces in the presence of Blebbistatinsimilar to what observed with dnRhoA This is consistentwith our observation of a complex Myosin distribution
in cells growing on SLA surfaces (Figures 2(a) and 2(b))presumably maintaining adequate cell adhesion by exertingproper cytoskeletal forces on focal adhesions and it isalso consistent with several reports showing that MyosinII function is required for cell differentiation [29 30] Itcan thus be hypothesized that RhoA enhances canonicalWnt signaling by increasing Myosin activation which inturn is known to activate cytoskeleton-associated signalingmolecules such as focal adhesion-associated Kinases (FAKs)[31] (Figure 5)These have been proven to affect the canonicalWnt signaling pathway in certain in vitro and in vivo models[32 33]This idea is consistent with recent findings indicatingthat focal adhesion formation and FAK phosphorylationare controlled by the RhoA pathway that focal adhesionmaturation and actin polymerization are promoted in cellscultured on micropatterned substrates [34] and that FAKphosphorylation is required for topography-induced differ-entiation of human mesenchymal stem cells [35]
In our study we then treated cells on both surfaceswith Cytochalasin Interestingly cells growing on Polishedsurfaces had thick stress fibers and a well-ordered actinnetwork under basal conditions whilst cells on SLA surfaceswhich do not have a plain uniform surface to adhere onlacked long actin filaments It can be speculated that shorteractin filaments can impart a more variegated array of forcevectors to the cell membrane and cytoplasm to better adapt to
8 BioMed Research International
the correspondently irregular titanium surface underneathCytochalasin at this concentration fragmented the actincytoskeleton by partially inhibiting its polymerization andsignificantly affected cell morphology Cytochalasin alsoincreased RhoA activity as previously reported and cor-respondently increased Wnt activation on both Polishedand SLA discs Under these conditions cell differentiationmarkers increased on both surfaces (Figures 4(c) and 4(d))consistently with reports by Higuchi et al [36]
Taken together our results indicate that cells growing onrough surfaces rearrange their cytoskeleton and this involvesthe activation of the small GTPase RhoA which controlsdownstream effectors that orchestrate cell contractility andshape Moreover RhoA can control and is required for theactivation ofWnt canonical signaling on rough surfacesThiseffect is not exerted directly but through its downstreameffector Myosin II When this is inhibited Wnt activation onrough surfaces is reversed in spite of RhoA activation Ourresults do not identify the specific mechanism of action forMyosin II although it may likely include focal adhesion-associated signaling which is known to be triggered by celltension and contractility Actin filaments can control RhoAactivity and their fragmentation increasesWnt activation andpromotes osteoblast differentiation (Figure 5)
Further studies will be required to better understand howcell contractility allows cells to sense their geometricalmicro-environment but the present study adds another and impor-tant piece of evidence to this elusive yet fundamental puzzlethat sustains the integration of osseointegrated implants inbone
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
S Mazzotta and S Ferrillo contributed equally to the presentstudy
Acknowledgments
The authors would like to thank Straumann Institut AG (andin particular Dr Appert and Dr Molenberg) Basel Switzer-land for kindly providing the titanium surfaces used in thepresent study The authors are also grateful to Dr FrancescaRavanetti and Professor Antonio Cacchioli for their preciousadvices and technical assistance The authors would liketo thank Dr Klaus Hahn for sharing his RhoA isoformsthrough the Addgene plasmid public repository The studywas funded by Grant no 839 2012 from the ITI Foundation(Basel Switzerland)
References
[1] C Galli G Guizzardi G Passeri et al ldquoComparison of humanmandibular osteoblasts grown on two commercially available
titanium implant surfacesrdquo Journal of Periodontology vol 76no 3 pp 364ndash372 2005
[2] B G Keselowsky L Wang Z Schwartz A J Garcia and BD Boyan ldquoIntegrin 1205725 controls osteoblastic proliferation anddifferentiation responses to titanium substrates presenting dif-ferent roughness characteristics in a roughness independentmannerrdquo Journal of BiomedicalMaterials Research A vol 80 no3 pp 700ndash710 2007
[3] I Wall N Donos K Carlqvist F Jones and P Brett ldquoModifiedtitanium surfaces promote accelerated osteogenic differentia-tion of mesenchymal stromal cells in vitrordquo Bone vol 45 no1 pp 17ndash26 2009
[4] K A Kilian B Bugarija B T Lahn and M Mrksich ldquoGeo-metric cues for directing the differentiation of mesenchymalstem cellsrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 11 pp 4872ndash4877 2010
[5] C Galli G Passeri F Ravanetti E Elezi M Pedrazzoni andG M MacAluso ldquoRough surface topography enhances theactivation of Wnt120573-catenin signaling in mesenchymal cellsrdquoJournal of Biomedical Materials Research A vol 95 no 3 pp682ndash690 2010
[6] R Olivares-Navarrete S L Hyzy D L Hutton et al ldquoDirectand indirect effects of microstructured titanium substrates onthe induction of mesenchymal stem cell differentiation towardsthe osteoblast lineagerdquo Biomaterials vol 31 no 10 pp 2728ndash2735 2010
[7] N Chakravorty S Hamlet A Jaiprakash et al ldquoPro-osteogenictopographical cues promote early activation of osteoprogenitordifferentiation via enhanced TGFbeta Wnt and Notch signal-ingrdquo Clinical Oral Implants Research 2013
[8] J Y Martin D D Dean D L Cochran J Simpson B DBoyan and Z Schwartz ldquoProliferation differentiation and pro-tein synthesis of human osteoblast-like cells (MG63) culturedon previously used titanium surfacesrdquo Clinical Oral ImplantsResearch vol 7 no 1 pp 27ndash37 1996
[9] S M Bang H J Moon Y D Kwon J Y Yoo A Pae and I KKwon ldquoOsteoblastic and osteoclastic differentiation on SLAand hydrophilic modified SLA titanium surfacesrdquo Clinical OralImplants Research 2013
[10] S Zinelis N Silikas A Thomas K Syres and G Eliades ldquoSur-face characterization of SLActive dental implantsrdquo EuropeanJournal of Esthetic Dentistry vol 7 no 1 pp 72ndash92 2012
[11] O Pertz L Hodgson R L Klemke and K M Hahn ldquoSpatio-temporal dynamics of RhoA activity in migrating cellsrdquoNaturevol 440 no 7087 pp 1069ndash1072 2006
[12] D Stamenovic and M F Coughlin ldquoThe role of prestress andarchitecture of the cytoskeleton and deformability of cytoskele-tal filaments in mechanics of adherent cells a quantitativeanalysisrdquo Journal of Theoretical Biology vol 201 no 1 pp 63ndash74 1999
[13] R Olivares-Navarrete S L Hyzy J H Park et al ldquoMediationof osteogenic differentiation of human mesenchymal stem cellson titanium surfaces by a Wnt-integrin feedback looprdquo Bioma-terials vol 32 no 27 pp 6399ndash6411 2011
[14] D A Glass II P Bialek J D Ahn et al ldquoCanonical Wnt signal-ing in differentiated osteoblasts controls osteoclast differentia-tionrdquo Developmental Cell vol 8 no 5 pp 751ndash764 2005
[15] S J Rodda and A P McMahon ldquoDistinct roles for Hedgehogand caronicalWnt signaling in specification differentiation andmaintenance of osteoblast progenitorsrdquo Development vol 133no 16 pp 3231ndash3244 2006
BioMed Research International 9
[16] N Wang and D E Ingber ldquoControl of cytoskeletal mechanicsby extracellular matrix cell shape and mechanical tensionrdquoBiophysical Journal vol 66 no 6 pp 2181ndash2189 1994
[17] C Galli S Guizzardi G Passeri G M Macaluso and R Scan-droglio ldquoLife on the wire on tensegrity and force balance incellsrdquo Acta Biomedica de lrsquoAteneo Parmense vol 76 no 1 pp5ndash62 2005
[18] D E Ingber ldquoTensegrity I Cell structure and hierarchical sys-tems biologyrdquo Journal of Cell Science vol 116 no 7 pp 1157ndash11732003
[19] C S Chen M Mrksich S Huang G M Whitesides and D EIngber ldquoGeometric control of cell life and deathrdquo Science vol276 no 5317 pp 1425ndash1428 1997
[20] D Stamenovi S M Mijailovich I M Toli-Noslashrrelykke J Chenand N Wang ldquoCell prestressmdashII Contribution of microtu-bulesrdquo American Journal of Physiology Cell Physiology vol 282no 3 pp C617ndashC624 2002
[21] M Endo M Nishita and Y Minami ldquoAnalysis of wntplanarcell polarity pathway in cultured cellsrdquo Methods in MolecularBiology vol 839 pp 201ndash214 2012
[22] C Gao and Y-G Chen ldquoDishevelled the hub ofWnt signalingrdquoCellular Signalling vol 22 no 5 pp 717ndash727 2010
[23] A J Mikels and R Nusse ldquoPurified Wnt5a protein activatesor inhibits beta-catenin-TCF signaling depending on receptorcontextrdquo PLoS Biology vol 4 no 4 Article ID e115 2006
[24] Y-N Lee Y Gao and H-Y Wang ldquoDifferential mediation ofthe Wnt canonical pathway by mammalian Dishevelleds-1 -2and -3rdquo Cellular Signalling vol 20 no 2 pp 443ndash452 2008
[25] J D Axelrod J R Miller J M Shulman R T Moon andN Perrimon ldquoDifferential recruitment of dishevelled providessignaling specificity in the planar cell polarity and Winglesssignaling pathwaysrdquo Genes and Development vol 12 no 16 pp2610ndash2622 1998
[26] K AWharton Jr ldquoRunninrsquo with the Dvl proteins that associatewith DshDvl and their significance to Wnt signal transduc-tionrdquo Developmental Biology vol 253 no 1 pp 1ndash17 2003
[27] W J Pan S Z Pang T Huang H Y Guo D Wu and L LildquoCharacterization of function of three domains in dishevelled1 DEP domain is responsible for membrane translocation ofdishevelled-1rdquo Cell Research vol 14 no 4 pp 324ndash330 2004
[28] A Calzado-Martın A Mendez-Vilas M Multigner et al ldquoOnthe role of RhoAROCK signaling in contact guidance ofbone-forming cells on anisotropic Ti6Al4V surfacesrdquo Acta Bio-materialia vol 7 no 4 pp 1890ndash1901 2011
[29] Y Sun L G Villa-Diaz R H Lam W Chen P H Krebsbachand J Fu ldquoMechanics regulates fate decisions of human embry-onic stem cellsrdquo PLoS ONE vol 7 no 5 Article ID e37178 2012
[30] H J Kwon ldquoChondrogenesis on sulfonate-coated hydrogelsis regulated by their mechanical propertiesrdquo Journal of theMechanical Behavior of BiomedicalMaterial vol 17 pp 337ndash3462013
[31] D W Dumbauld H Shin N D Gallant K E Michael HRadhakrishna and A J Garcıa ldquoContractility modulates celladhesion strengthening through focal adhesion kinase andassembly of vinculin-containing focal adhesionsrdquo Journal ofCellular Physiology vol 223 no 3 pp 746ndash756 2010
[32] Y Fonar Y E Gutkovich H Root et al ldquoFocal adhesion kinaseprotein regulates Wnt3a gene expression to control cell fatespecification in the developing neural platerdquoMolecular Biologyof the Cell vol 22 no 13 pp 2409ndash2421 2011
[33] M Despeaux G Chicanne E Rouer et al ldquoFocal adhesionkinase splice variants maintain primitive acute myeloid leuke-mia cells through alteredWnt signalingrdquo Stem Cells vol 30 no8 pp 1597ndash1610 2012
[34] C H Seo K Furukawa K Montagne H Jeong and T UshidaldquoThe effect of substrate microtopography on focal adhesionmaturation and actin organization via the RhoAROCK path-wayrdquo Biomaterials vol 32 no 36 pp 9568ndash9575 2011
[35] B K Teo S T Wong C K Lim et al ldquoNanotopographymodulates mechanotransduction of stem cells and inducesdifferentiation through focal adhesion kinaserdquo ACS Nano 2013
[36] C Higuchi N Nakamura H Yoshikawa and K Itoh ldquoTran-sient dynamic actin cytoskeletal change stimulates the osteo-blastic differentiationrdquo Journal of Bone andMineralMetabolismvol 27 no 2 pp 158ndash167 2009
Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials

BioMed Research International 7
RhoA
MLCK
Actin
Integrins
Myosin II
TCF
Frizzled LRP4-6
Wnt
DvlAxin
Nucleus
Destructioncomplex
Frizzled Ror2C
Wnt
120573cat
120573cat
120573cat
GSK3120573
CK120572
Figure 5 Diagram depicting the proposed model for RhoA role in cell responses to substrate topography Noncanonical Wnt factors bindto receptor dimers Frizzled-Ror2C which activate RhoA Rho in turns activates downstream target Myosin Light Chain Kinase (MLCK)which phosphorylates Myosin II Acto-Myosin filaments create cell tension which facilitates the activation of canonical Wnt signaling andpromotes b catenin release from its destruction complex and translocation to the nucleus possibly by acting on tension sensitive structures(focal adhesions) A feedback loop from tension-activated structures controls RhoA activation levels Actin microfilaments can control RhoAtoo as their partial disruption increases RhoA activity and canonical Wnt activation
organization and osteoblastic commitment share some oftheir key molecular components we hypothesized thatcytoskeletal organization could at least in part cause theactivation of Wnt signaling observed on rough surfacesTo investigate the relation between cytoskeleton and Wntsignaling activation on implant surfaces we quantitatedactive RhoA small GTPase and found that its levels arehigher on rough SLA surfaces according to recent inde-pendent data [28] It can be speculated that cells need torearrange their cytoskeleton through RhoA activation toadapt to the irregular surface profile of SLA discs Interest-ingly hampering RhoA activity by transfecting cells with adominant negative isoformof RhoA (dnRhoA) reducedTCF-mediated transcriptional activity on rough surfaces whichwe previously showed to be higher than on smooth surfacesunder basal conditions [5] This clearly indicates that RhoAactivation is required for the enhancement of Wnt canonicalsignaling on rough surfaces However we also showed thatRhoA-mediated canonical Wnt activation required RhoAdownstream molecular effectors to be functional Myosininhibition by 1mM Blebbistatin did increase RhoA activitypossibly by a negative feedback loop but failed to enhanceWnt canonical signaling On the contrary Wnt activationdropped on SLA surfaces in the presence of Blebbistatinsimilar to what observed with dnRhoA This is consistentwith our observation of a complex Myosin distribution
in cells growing on SLA surfaces (Figures 2(a) and 2(b))presumably maintaining adequate cell adhesion by exertingproper cytoskeletal forces on focal adhesions and it isalso consistent with several reports showing that MyosinII function is required for cell differentiation [29 30] Itcan thus be hypothesized that RhoA enhances canonicalWnt signaling by increasing Myosin activation which inturn is known to activate cytoskeleton-associated signalingmolecules such as focal adhesion-associated Kinases (FAKs)[31] (Figure 5)These have been proven to affect the canonicalWnt signaling pathway in certain in vitro and in vivo models[32 33]This idea is consistent with recent findings indicatingthat focal adhesion formation and FAK phosphorylationare controlled by the RhoA pathway that focal adhesionmaturation and actin polymerization are promoted in cellscultured on micropatterned substrates [34] and that FAKphosphorylation is required for topography-induced differ-entiation of human mesenchymal stem cells [35]
In our study we then treated cells on both surfaceswith Cytochalasin Interestingly cells growing on Polishedsurfaces had thick stress fibers and a well-ordered actinnetwork under basal conditions whilst cells on SLA surfaceswhich do not have a plain uniform surface to adhere onlacked long actin filaments It can be speculated that shorteractin filaments can impart a more variegated array of forcevectors to the cell membrane and cytoplasm to better adapt to
8 BioMed Research International
the correspondently irregular titanium surface underneathCytochalasin at this concentration fragmented the actincytoskeleton by partially inhibiting its polymerization andsignificantly affected cell morphology Cytochalasin alsoincreased RhoA activity as previously reported and cor-respondently increased Wnt activation on both Polishedand SLA discs Under these conditions cell differentiationmarkers increased on both surfaces (Figures 4(c) and 4(d))consistently with reports by Higuchi et al [36]
Taken together our results indicate that cells growing onrough surfaces rearrange their cytoskeleton and this involvesthe activation of the small GTPase RhoA which controlsdownstream effectors that orchestrate cell contractility andshape Moreover RhoA can control and is required for theactivation ofWnt canonical signaling on rough surfacesThiseffect is not exerted directly but through its downstreameffector Myosin II When this is inhibited Wnt activation onrough surfaces is reversed in spite of RhoA activation Ourresults do not identify the specific mechanism of action forMyosin II although it may likely include focal adhesion-associated signaling which is known to be triggered by celltension and contractility Actin filaments can control RhoAactivity and their fragmentation increasesWnt activation andpromotes osteoblast differentiation (Figure 5)
Further studies will be required to better understand howcell contractility allows cells to sense their geometricalmicro-environment but the present study adds another and impor-tant piece of evidence to this elusive yet fundamental puzzlethat sustains the integration of osseointegrated implants inbone
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
S Mazzotta and S Ferrillo contributed equally to the presentstudy
Acknowledgments
The authors would like to thank Straumann Institut AG (andin particular Dr Appert and Dr Molenberg) Basel Switzer-land for kindly providing the titanium surfaces used in thepresent study The authors are also grateful to Dr FrancescaRavanetti and Professor Antonio Cacchioli for their preciousadvices and technical assistance The authors would liketo thank Dr Klaus Hahn for sharing his RhoA isoformsthrough the Addgene plasmid public repository The studywas funded by Grant no 839 2012 from the ITI Foundation(Basel Switzerland)
References
[1] C Galli G Guizzardi G Passeri et al ldquoComparison of humanmandibular osteoblasts grown on two commercially available
titanium implant surfacesrdquo Journal of Periodontology vol 76no 3 pp 364ndash372 2005
[2] B G Keselowsky L Wang Z Schwartz A J Garcia and BD Boyan ldquoIntegrin 1205725 controls osteoblastic proliferation anddifferentiation responses to titanium substrates presenting dif-ferent roughness characteristics in a roughness independentmannerrdquo Journal of BiomedicalMaterials Research A vol 80 no3 pp 700ndash710 2007
[3] I Wall N Donos K Carlqvist F Jones and P Brett ldquoModifiedtitanium surfaces promote accelerated osteogenic differentia-tion of mesenchymal stromal cells in vitrordquo Bone vol 45 no1 pp 17ndash26 2009
[4] K A Kilian B Bugarija B T Lahn and M Mrksich ldquoGeo-metric cues for directing the differentiation of mesenchymalstem cellsrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 11 pp 4872ndash4877 2010
[5] C Galli G Passeri F Ravanetti E Elezi M Pedrazzoni andG M MacAluso ldquoRough surface topography enhances theactivation of Wnt120573-catenin signaling in mesenchymal cellsrdquoJournal of Biomedical Materials Research A vol 95 no 3 pp682ndash690 2010
[6] R Olivares-Navarrete S L Hyzy D L Hutton et al ldquoDirectand indirect effects of microstructured titanium substrates onthe induction of mesenchymal stem cell differentiation towardsthe osteoblast lineagerdquo Biomaterials vol 31 no 10 pp 2728ndash2735 2010
[7] N Chakravorty S Hamlet A Jaiprakash et al ldquoPro-osteogenictopographical cues promote early activation of osteoprogenitordifferentiation via enhanced TGFbeta Wnt and Notch signal-ingrdquo Clinical Oral Implants Research 2013
[8] J Y Martin D D Dean D L Cochran J Simpson B DBoyan and Z Schwartz ldquoProliferation differentiation and pro-tein synthesis of human osteoblast-like cells (MG63) culturedon previously used titanium surfacesrdquo Clinical Oral ImplantsResearch vol 7 no 1 pp 27ndash37 1996
[9] S M Bang H J Moon Y D Kwon J Y Yoo A Pae and I KKwon ldquoOsteoblastic and osteoclastic differentiation on SLAand hydrophilic modified SLA titanium surfacesrdquo Clinical OralImplants Research 2013
[10] S Zinelis N Silikas A Thomas K Syres and G Eliades ldquoSur-face characterization of SLActive dental implantsrdquo EuropeanJournal of Esthetic Dentistry vol 7 no 1 pp 72ndash92 2012
[11] O Pertz L Hodgson R L Klemke and K M Hahn ldquoSpatio-temporal dynamics of RhoA activity in migrating cellsrdquoNaturevol 440 no 7087 pp 1069ndash1072 2006
[12] D Stamenovic and M F Coughlin ldquoThe role of prestress andarchitecture of the cytoskeleton and deformability of cytoskele-tal filaments in mechanics of adherent cells a quantitativeanalysisrdquo Journal of Theoretical Biology vol 201 no 1 pp 63ndash74 1999
[13] R Olivares-Navarrete S L Hyzy J H Park et al ldquoMediationof osteogenic differentiation of human mesenchymal stem cellson titanium surfaces by a Wnt-integrin feedback looprdquo Bioma-terials vol 32 no 27 pp 6399ndash6411 2011
[14] D A Glass II P Bialek J D Ahn et al ldquoCanonical Wnt signal-ing in differentiated osteoblasts controls osteoclast differentia-tionrdquo Developmental Cell vol 8 no 5 pp 751ndash764 2005
[15] S J Rodda and A P McMahon ldquoDistinct roles for Hedgehogand caronicalWnt signaling in specification differentiation andmaintenance of osteoblast progenitorsrdquo Development vol 133no 16 pp 3231ndash3244 2006
BioMed Research International 9
[16] N Wang and D E Ingber ldquoControl of cytoskeletal mechanicsby extracellular matrix cell shape and mechanical tensionrdquoBiophysical Journal vol 66 no 6 pp 2181ndash2189 1994
[17] C Galli S Guizzardi G Passeri G M Macaluso and R Scan-droglio ldquoLife on the wire on tensegrity and force balance incellsrdquo Acta Biomedica de lrsquoAteneo Parmense vol 76 no 1 pp5ndash62 2005
[18] D E Ingber ldquoTensegrity I Cell structure and hierarchical sys-tems biologyrdquo Journal of Cell Science vol 116 no 7 pp 1157ndash11732003
[19] C S Chen M Mrksich S Huang G M Whitesides and D EIngber ldquoGeometric control of cell life and deathrdquo Science vol276 no 5317 pp 1425ndash1428 1997
[20] D Stamenovi S M Mijailovich I M Toli-Noslashrrelykke J Chenand N Wang ldquoCell prestressmdashII Contribution of microtu-bulesrdquo American Journal of Physiology Cell Physiology vol 282no 3 pp C617ndashC624 2002
[21] M Endo M Nishita and Y Minami ldquoAnalysis of wntplanarcell polarity pathway in cultured cellsrdquo Methods in MolecularBiology vol 839 pp 201ndash214 2012
[22] C Gao and Y-G Chen ldquoDishevelled the hub ofWnt signalingrdquoCellular Signalling vol 22 no 5 pp 717ndash727 2010
[23] A J Mikels and R Nusse ldquoPurified Wnt5a protein activatesor inhibits beta-catenin-TCF signaling depending on receptorcontextrdquo PLoS Biology vol 4 no 4 Article ID e115 2006
[24] Y-N Lee Y Gao and H-Y Wang ldquoDifferential mediation ofthe Wnt canonical pathway by mammalian Dishevelleds-1 -2and -3rdquo Cellular Signalling vol 20 no 2 pp 443ndash452 2008
[25] J D Axelrod J R Miller J M Shulman R T Moon andN Perrimon ldquoDifferential recruitment of dishevelled providessignaling specificity in the planar cell polarity and Winglesssignaling pathwaysrdquo Genes and Development vol 12 no 16 pp2610ndash2622 1998
[26] K AWharton Jr ldquoRunninrsquo with the Dvl proteins that associatewith DshDvl and their significance to Wnt signal transduc-tionrdquo Developmental Biology vol 253 no 1 pp 1ndash17 2003
[27] W J Pan S Z Pang T Huang H Y Guo D Wu and L LildquoCharacterization of function of three domains in dishevelled1 DEP domain is responsible for membrane translocation ofdishevelled-1rdquo Cell Research vol 14 no 4 pp 324ndash330 2004
[28] A Calzado-Martın A Mendez-Vilas M Multigner et al ldquoOnthe role of RhoAROCK signaling in contact guidance ofbone-forming cells on anisotropic Ti6Al4V surfacesrdquo Acta Bio-materialia vol 7 no 4 pp 1890ndash1901 2011
[29] Y Sun L G Villa-Diaz R H Lam W Chen P H Krebsbachand J Fu ldquoMechanics regulates fate decisions of human embry-onic stem cellsrdquo PLoS ONE vol 7 no 5 Article ID e37178 2012
[30] H J Kwon ldquoChondrogenesis on sulfonate-coated hydrogelsis regulated by their mechanical propertiesrdquo Journal of theMechanical Behavior of BiomedicalMaterial vol 17 pp 337ndash3462013
[31] D W Dumbauld H Shin N D Gallant K E Michael HRadhakrishna and A J Garcıa ldquoContractility modulates celladhesion strengthening through focal adhesion kinase andassembly of vinculin-containing focal adhesionsrdquo Journal ofCellular Physiology vol 223 no 3 pp 746ndash756 2010
[32] Y Fonar Y E Gutkovich H Root et al ldquoFocal adhesion kinaseprotein regulates Wnt3a gene expression to control cell fatespecification in the developing neural platerdquoMolecular Biologyof the Cell vol 22 no 13 pp 2409ndash2421 2011
[33] M Despeaux G Chicanne E Rouer et al ldquoFocal adhesionkinase splice variants maintain primitive acute myeloid leuke-mia cells through alteredWnt signalingrdquo Stem Cells vol 30 no8 pp 1597ndash1610 2012
[34] C H Seo K Furukawa K Montagne H Jeong and T UshidaldquoThe effect of substrate microtopography on focal adhesionmaturation and actin organization via the RhoAROCK path-wayrdquo Biomaterials vol 32 no 36 pp 9568ndash9575 2011
[35] B K Teo S T Wong C K Lim et al ldquoNanotopographymodulates mechanotransduction of stem cells and inducesdifferentiation through focal adhesion kinaserdquo ACS Nano 2013
[36] C Higuchi N Nakamura H Yoshikawa and K Itoh ldquoTran-sient dynamic actin cytoskeletal change stimulates the osteo-blastic differentiationrdquo Journal of Bone andMineralMetabolismvol 27 no 2 pp 158ndash167 2009
Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials

8 BioMed Research International
the correspondently irregular titanium surface underneathCytochalasin at this concentration fragmented the actincytoskeleton by partially inhibiting its polymerization andsignificantly affected cell morphology Cytochalasin alsoincreased RhoA activity as previously reported and cor-respondently increased Wnt activation on both Polishedand SLA discs Under these conditions cell differentiationmarkers increased on both surfaces (Figures 4(c) and 4(d))consistently with reports by Higuchi et al [36]
Taken together our results indicate that cells growing onrough surfaces rearrange their cytoskeleton and this involvesthe activation of the small GTPase RhoA which controlsdownstream effectors that orchestrate cell contractility andshape Moreover RhoA can control and is required for theactivation ofWnt canonical signaling on rough surfacesThiseffect is not exerted directly but through its downstreameffector Myosin II When this is inhibited Wnt activation onrough surfaces is reversed in spite of RhoA activation Ourresults do not identify the specific mechanism of action forMyosin II although it may likely include focal adhesion-associated signaling which is known to be triggered by celltension and contractility Actin filaments can control RhoAactivity and their fragmentation increasesWnt activation andpromotes osteoblast differentiation (Figure 5)
Further studies will be required to better understand howcell contractility allows cells to sense their geometricalmicro-environment but the present study adds another and impor-tant piece of evidence to this elusive yet fundamental puzzlethat sustains the integration of osseointegrated implants inbone
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
S Mazzotta and S Ferrillo contributed equally to the presentstudy
Acknowledgments
The authors would like to thank Straumann Institut AG (andin particular Dr Appert and Dr Molenberg) Basel Switzer-land for kindly providing the titanium surfaces used in thepresent study The authors are also grateful to Dr FrancescaRavanetti and Professor Antonio Cacchioli for their preciousadvices and technical assistance The authors would liketo thank Dr Klaus Hahn for sharing his RhoA isoformsthrough the Addgene plasmid public repository The studywas funded by Grant no 839 2012 from the ITI Foundation(Basel Switzerland)
References
[1] C Galli G Guizzardi G Passeri et al ldquoComparison of humanmandibular osteoblasts grown on two commercially available
titanium implant surfacesrdquo Journal of Periodontology vol 76no 3 pp 364ndash372 2005
[2] B G Keselowsky L Wang Z Schwartz A J Garcia and BD Boyan ldquoIntegrin 1205725 controls osteoblastic proliferation anddifferentiation responses to titanium substrates presenting dif-ferent roughness characteristics in a roughness independentmannerrdquo Journal of BiomedicalMaterials Research A vol 80 no3 pp 700ndash710 2007
[3] I Wall N Donos K Carlqvist F Jones and P Brett ldquoModifiedtitanium surfaces promote accelerated osteogenic differentia-tion of mesenchymal stromal cells in vitrordquo Bone vol 45 no1 pp 17ndash26 2009
[4] K A Kilian B Bugarija B T Lahn and M Mrksich ldquoGeo-metric cues for directing the differentiation of mesenchymalstem cellsrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 107 no 11 pp 4872ndash4877 2010
[5] C Galli G Passeri F Ravanetti E Elezi M Pedrazzoni andG M MacAluso ldquoRough surface topography enhances theactivation of Wnt120573-catenin signaling in mesenchymal cellsrdquoJournal of Biomedical Materials Research A vol 95 no 3 pp682ndash690 2010
[6] R Olivares-Navarrete S L Hyzy D L Hutton et al ldquoDirectand indirect effects of microstructured titanium substrates onthe induction of mesenchymal stem cell differentiation towardsthe osteoblast lineagerdquo Biomaterials vol 31 no 10 pp 2728ndash2735 2010
[7] N Chakravorty S Hamlet A Jaiprakash et al ldquoPro-osteogenictopographical cues promote early activation of osteoprogenitordifferentiation via enhanced TGFbeta Wnt and Notch signal-ingrdquo Clinical Oral Implants Research 2013
[8] J Y Martin D D Dean D L Cochran J Simpson B DBoyan and Z Schwartz ldquoProliferation differentiation and pro-tein synthesis of human osteoblast-like cells (MG63) culturedon previously used titanium surfacesrdquo Clinical Oral ImplantsResearch vol 7 no 1 pp 27ndash37 1996
[9] S M Bang H J Moon Y D Kwon J Y Yoo A Pae and I KKwon ldquoOsteoblastic and osteoclastic differentiation on SLAand hydrophilic modified SLA titanium surfacesrdquo Clinical OralImplants Research 2013
[10] S Zinelis N Silikas A Thomas K Syres and G Eliades ldquoSur-face characterization of SLActive dental implantsrdquo EuropeanJournal of Esthetic Dentistry vol 7 no 1 pp 72ndash92 2012
[11] O Pertz L Hodgson R L Klemke and K M Hahn ldquoSpatio-temporal dynamics of RhoA activity in migrating cellsrdquoNaturevol 440 no 7087 pp 1069ndash1072 2006
[12] D Stamenovic and M F Coughlin ldquoThe role of prestress andarchitecture of the cytoskeleton and deformability of cytoskele-tal filaments in mechanics of adherent cells a quantitativeanalysisrdquo Journal of Theoretical Biology vol 201 no 1 pp 63ndash74 1999
[13] R Olivares-Navarrete S L Hyzy J H Park et al ldquoMediationof osteogenic differentiation of human mesenchymal stem cellson titanium surfaces by a Wnt-integrin feedback looprdquo Bioma-terials vol 32 no 27 pp 6399ndash6411 2011
[14] D A Glass II P Bialek J D Ahn et al ldquoCanonical Wnt signal-ing in differentiated osteoblasts controls osteoclast differentia-tionrdquo Developmental Cell vol 8 no 5 pp 751ndash764 2005
[15] S J Rodda and A P McMahon ldquoDistinct roles for Hedgehogand caronicalWnt signaling in specification differentiation andmaintenance of osteoblast progenitorsrdquo Development vol 133no 16 pp 3231ndash3244 2006
BioMed Research International 9
[16] N Wang and D E Ingber ldquoControl of cytoskeletal mechanicsby extracellular matrix cell shape and mechanical tensionrdquoBiophysical Journal vol 66 no 6 pp 2181ndash2189 1994
[17] C Galli S Guizzardi G Passeri G M Macaluso and R Scan-droglio ldquoLife on the wire on tensegrity and force balance incellsrdquo Acta Biomedica de lrsquoAteneo Parmense vol 76 no 1 pp5ndash62 2005
[18] D E Ingber ldquoTensegrity I Cell structure and hierarchical sys-tems biologyrdquo Journal of Cell Science vol 116 no 7 pp 1157ndash11732003
[19] C S Chen M Mrksich S Huang G M Whitesides and D EIngber ldquoGeometric control of cell life and deathrdquo Science vol276 no 5317 pp 1425ndash1428 1997
[20] D Stamenovi S M Mijailovich I M Toli-Noslashrrelykke J Chenand N Wang ldquoCell prestressmdashII Contribution of microtu-bulesrdquo American Journal of Physiology Cell Physiology vol 282no 3 pp C617ndashC624 2002
[21] M Endo M Nishita and Y Minami ldquoAnalysis of wntplanarcell polarity pathway in cultured cellsrdquo Methods in MolecularBiology vol 839 pp 201ndash214 2012
[22] C Gao and Y-G Chen ldquoDishevelled the hub ofWnt signalingrdquoCellular Signalling vol 22 no 5 pp 717ndash727 2010
[23] A J Mikels and R Nusse ldquoPurified Wnt5a protein activatesor inhibits beta-catenin-TCF signaling depending on receptorcontextrdquo PLoS Biology vol 4 no 4 Article ID e115 2006
[24] Y-N Lee Y Gao and H-Y Wang ldquoDifferential mediation ofthe Wnt canonical pathway by mammalian Dishevelleds-1 -2and -3rdquo Cellular Signalling vol 20 no 2 pp 443ndash452 2008
[25] J D Axelrod J R Miller J M Shulman R T Moon andN Perrimon ldquoDifferential recruitment of dishevelled providessignaling specificity in the planar cell polarity and Winglesssignaling pathwaysrdquo Genes and Development vol 12 no 16 pp2610ndash2622 1998
[26] K AWharton Jr ldquoRunninrsquo with the Dvl proteins that associatewith DshDvl and their significance to Wnt signal transduc-tionrdquo Developmental Biology vol 253 no 1 pp 1ndash17 2003
[27] W J Pan S Z Pang T Huang H Y Guo D Wu and L LildquoCharacterization of function of three domains in dishevelled1 DEP domain is responsible for membrane translocation ofdishevelled-1rdquo Cell Research vol 14 no 4 pp 324ndash330 2004
[28] A Calzado-Martın A Mendez-Vilas M Multigner et al ldquoOnthe role of RhoAROCK signaling in contact guidance ofbone-forming cells on anisotropic Ti6Al4V surfacesrdquo Acta Bio-materialia vol 7 no 4 pp 1890ndash1901 2011
[29] Y Sun L G Villa-Diaz R H Lam W Chen P H Krebsbachand J Fu ldquoMechanics regulates fate decisions of human embry-onic stem cellsrdquo PLoS ONE vol 7 no 5 Article ID e37178 2012
[30] H J Kwon ldquoChondrogenesis on sulfonate-coated hydrogelsis regulated by their mechanical propertiesrdquo Journal of theMechanical Behavior of BiomedicalMaterial vol 17 pp 337ndash3462013
[31] D W Dumbauld H Shin N D Gallant K E Michael HRadhakrishna and A J Garcıa ldquoContractility modulates celladhesion strengthening through focal adhesion kinase andassembly of vinculin-containing focal adhesionsrdquo Journal ofCellular Physiology vol 223 no 3 pp 746ndash756 2010
[32] Y Fonar Y E Gutkovich H Root et al ldquoFocal adhesion kinaseprotein regulates Wnt3a gene expression to control cell fatespecification in the developing neural platerdquoMolecular Biologyof the Cell vol 22 no 13 pp 2409ndash2421 2011
[33] M Despeaux G Chicanne E Rouer et al ldquoFocal adhesionkinase splice variants maintain primitive acute myeloid leuke-mia cells through alteredWnt signalingrdquo Stem Cells vol 30 no8 pp 1597ndash1610 2012
[34] C H Seo K Furukawa K Montagne H Jeong and T UshidaldquoThe effect of substrate microtopography on focal adhesionmaturation and actin organization via the RhoAROCK path-wayrdquo Biomaterials vol 32 no 36 pp 9568ndash9575 2011
[35] B K Teo S T Wong C K Lim et al ldquoNanotopographymodulates mechanotransduction of stem cells and inducesdifferentiation through focal adhesion kinaserdquo ACS Nano 2013
[36] C Higuchi N Nakamura H Yoshikawa and K Itoh ldquoTran-sient dynamic actin cytoskeletal change stimulates the osteo-blastic differentiationrdquo Journal of Bone andMineralMetabolismvol 27 no 2 pp 158ndash167 2009
Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials

BioMed Research International 9
[16] N Wang and D E Ingber ldquoControl of cytoskeletal mechanicsby extracellular matrix cell shape and mechanical tensionrdquoBiophysical Journal vol 66 no 6 pp 2181ndash2189 1994
[17] C Galli S Guizzardi G Passeri G M Macaluso and R Scan-droglio ldquoLife on the wire on tensegrity and force balance incellsrdquo Acta Biomedica de lrsquoAteneo Parmense vol 76 no 1 pp5ndash62 2005
[18] D E Ingber ldquoTensegrity I Cell structure and hierarchical sys-tems biologyrdquo Journal of Cell Science vol 116 no 7 pp 1157ndash11732003
[19] C S Chen M Mrksich S Huang G M Whitesides and D EIngber ldquoGeometric control of cell life and deathrdquo Science vol276 no 5317 pp 1425ndash1428 1997
[20] D Stamenovi S M Mijailovich I M Toli-Noslashrrelykke J Chenand N Wang ldquoCell prestressmdashII Contribution of microtu-bulesrdquo American Journal of Physiology Cell Physiology vol 282no 3 pp C617ndashC624 2002
[21] M Endo M Nishita and Y Minami ldquoAnalysis of wntplanarcell polarity pathway in cultured cellsrdquo Methods in MolecularBiology vol 839 pp 201ndash214 2012
[22] C Gao and Y-G Chen ldquoDishevelled the hub ofWnt signalingrdquoCellular Signalling vol 22 no 5 pp 717ndash727 2010
[23] A J Mikels and R Nusse ldquoPurified Wnt5a protein activatesor inhibits beta-catenin-TCF signaling depending on receptorcontextrdquo PLoS Biology vol 4 no 4 Article ID e115 2006
[24] Y-N Lee Y Gao and H-Y Wang ldquoDifferential mediation ofthe Wnt canonical pathway by mammalian Dishevelleds-1 -2and -3rdquo Cellular Signalling vol 20 no 2 pp 443ndash452 2008
[25] J D Axelrod J R Miller J M Shulman R T Moon andN Perrimon ldquoDifferential recruitment of dishevelled providessignaling specificity in the planar cell polarity and Winglesssignaling pathwaysrdquo Genes and Development vol 12 no 16 pp2610ndash2622 1998
[26] K AWharton Jr ldquoRunninrsquo with the Dvl proteins that associatewith DshDvl and their significance to Wnt signal transduc-tionrdquo Developmental Biology vol 253 no 1 pp 1ndash17 2003
[27] W J Pan S Z Pang T Huang H Y Guo D Wu and L LildquoCharacterization of function of three domains in dishevelled1 DEP domain is responsible for membrane translocation ofdishevelled-1rdquo Cell Research vol 14 no 4 pp 324ndash330 2004
[28] A Calzado-Martın A Mendez-Vilas M Multigner et al ldquoOnthe role of RhoAROCK signaling in contact guidance ofbone-forming cells on anisotropic Ti6Al4V surfacesrdquo Acta Bio-materialia vol 7 no 4 pp 1890ndash1901 2011
[29] Y Sun L G Villa-Diaz R H Lam W Chen P H Krebsbachand J Fu ldquoMechanics regulates fate decisions of human embry-onic stem cellsrdquo PLoS ONE vol 7 no 5 Article ID e37178 2012
[30] H J Kwon ldquoChondrogenesis on sulfonate-coated hydrogelsis regulated by their mechanical propertiesrdquo Journal of theMechanical Behavior of BiomedicalMaterial vol 17 pp 337ndash3462013
[31] D W Dumbauld H Shin N D Gallant K E Michael HRadhakrishna and A J Garcıa ldquoContractility modulates celladhesion strengthening through focal adhesion kinase andassembly of vinculin-containing focal adhesionsrdquo Journal ofCellular Physiology vol 223 no 3 pp 746ndash756 2010
[32] Y Fonar Y E Gutkovich H Root et al ldquoFocal adhesion kinaseprotein regulates Wnt3a gene expression to control cell fatespecification in the developing neural platerdquoMolecular Biologyof the Cell vol 22 no 13 pp 2409ndash2421 2011
[33] M Despeaux G Chicanne E Rouer et al ldquoFocal adhesionkinase splice variants maintain primitive acute myeloid leuke-mia cells through alteredWnt signalingrdquo Stem Cells vol 30 no8 pp 1597ndash1610 2012
[34] C H Seo K Furukawa K Montagne H Jeong and T UshidaldquoThe effect of substrate microtopography on focal adhesionmaturation and actin organization via the RhoAROCK path-wayrdquo Biomaterials vol 32 no 36 pp 9568ndash9575 2011
[35] B K Teo S T Wong C K Lim et al ldquoNanotopographymodulates mechanotransduction of stem cells and inducesdifferentiation through focal adhesion kinaserdquo ACS Nano 2013
[36] C Higuchi N Nakamura H Yoshikawa and K Itoh ldquoTran-sient dynamic actin cytoskeletal change stimulates the osteo-blastic differentiationrdquo Journal of Bone andMineralMetabolismvol 27 no 2 pp 158ndash167 2009
Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials

Submit your manuscripts athttpwwwhindawicom
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CorrosionInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Polymer ScienceInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CeramicsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CompositesJournal of
NanoparticlesJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Biomaterials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
NanoscienceJournal of
TextilesHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Journal of
NanotechnologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
CrystallographyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
CoatingsJournal of
Advances in
Materials Science and EngineeringHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Smart Materials Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MetallurgyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
MaterialsJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Nano
materials
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal ofNanomaterials