research article androgen receptors expression in...
TRANSCRIPT

Research ArticleAndrogen Receptors Expression in Pituitary of Male Viscachain relation to Growth and Reproductive Cycle
Veroacutenica Palmira Filippa12 Gabriela Judith Rosales1
Albana Andrea Marina Crucentildeo1 and Fabian Heber Mohamed1
1Histologıa Facultad de Quımica Bioquımica y Farmacia Universidad Nacional de San LuisAvenida Ejercito de los Andes 950 Bloque I Piso No 1 5700 San Luis Argentina2Consejo Nacional de Investigaciones Cientıficas y Tecnicas (CONICET) 5700 San Luis Argentina
Correspondence should be addressed to Fabian Heber Mohamed fhmounsleduar
Received 26 November 2014 Revised 6 March 2015 Accepted 8 March 2015
Academic Editor Małgorzata Kotula-Balak
Copyright copy 2015 Veronica Palmira Filippa et alThis is an open access article distributed under theCreativeCommonsAttributionLicense which permits unrestricted use distribution and reproduction in anymedium provided the originalwork is properly cited
The aim of this work was to study the androgen receptors (AR) expression in pituitary pars distalis (PD) of male viscachas inrelation to growth and reproductive cycle AR were detected by immunocytochemistry and quantified by image analysis Pituitaryglands from fetus immature prepubertal and adult viscachas during their reproductive cycle were used In the fetal PD theimmunoreactivity (ir) was mainly cytoplasmic In immature and prepubertal animals AR-ir was cytoplasmic (ARc-ir) and nuclear(ARn-ir) in medial region In adult animals ARn-ir cells were numerous at caudal end AR regionalization varied between thePD zones in relation to growth In immature animals the ARn-ir increased whereas the cytoplasmic expression decreased inrelation to the fetal glandsThe percentage of ARc-ir cells increased in prepubertal animals whereas the nuclear AR expression waspredominant in adult viscachas The AR expression changed in adults showing minimum percentage in the gonadal regressionperiod The variation of nuclear AR expression was directly related with testosterone concentration These results demonstratedvariations in the immunostaining pattern regionalization and number of AR-ir cells throughout development growth andreproductive cycle suggesting the involvement of AR in the regulation of the pituitary activity of male viscacha
1 Introduction
Androgens are important steroidal hormones involved inthe sexual development and reproduction of male mammalsThey are essential during development and for the mainte-nance of secondary male characteristics initiation and con-tinuation of spermatogenesis and gonadotropin regulation[1] Testosterone and dihydrotestosterone bind to specificcytosolic receptors the androgen receptors (AR) resultingin a conformational change in receptor protein dimeriza-tion nuclear translocation association with cofactors andinteraction with specific regions of the genome [2] to initiatea cellular response [3] The existence of the AR expressionin the cytoplasm of pituitary cells has been described inother species [4 5] It has been demonstrated that there isan inverse relationship between the cytoplasmic and nuclearlocalization and testosterone administration increases thenuclear expression of AR [6]
Several studies have demonstrated that the pituitary glandis an androgen target tissue and serves as a potential siteof androgen feedback on hormone secretion [4 6 7] Thelocalization and function of AR have been studied in thepituitary gland in some mammalian species in relation to ageor gender [8ndash10] AR expression has been studied during thefetal development of the hypothalamus hippocampus andpituitary in monkeys [11 12] and sheep [13] Studies of theontogeny of the AR expression in the developing pituitarygland have been performed in monkeys [14] chickens [15]mice [16] rats [17] and sheep [18] and it has been demon-strated that pituitary AR in mammals is conservative
Some species use environmental cues themost importantbeing the photoperiod to synchronize their endogenouscycles and to reproduce at the optimal time of the year [19]The pituitary gland is under control of the hypothalamusand the gonadal hormones which exert feedback effects on
Hindawi Publishing CorporationInternational Journal of EndocrinologyVolume 2015 Article ID 168047 11 pageshttpdxdoiorg1011552015168047
2 International Journal of Endocrinology
the hypothalamic-pituitary-gonadal axis [20] Variations inthe androgen serum concentration and their receptors in dif-ferent tissues are responsible for biological effects throughoutthe life cycle of adult animals Testosterone plays a highlysignificant role in the transition from the reproductive activ-ity phase to gonadal regression and gonadal recrudescence[21] Seasonal variations in testosterone serum concentrationare related to the reproductive status of male birds [22] andhamsters [23] The central mechanisms responsible for thephotoperiod negative effects upon the testosterone and thepossible influence of day length on AR expression have beenreported [24] Tetel et al [25] have indicated that day-length-induced fluctuations in AR expression may contribute toseasonal variations of testosterone
Our experimental model the viscacha (Lagostomus max-imus maximus) is a hystricomorph rodent of seasonalreproductive patternsThe annual reproductive cycle of adultmale animals has three periods reproductive (summer andearly autumn) gonadal regression (winter) and gonadalrecovery (spring) Previous studies have demonstrated thatthe gonadal regression period is characterized by an increaseof the secretory ability of the pineal gland [26] andmaximumlevels of melatonin in blood [27] In pituitary PD the activityof LH- and FSH-cells was different during the annual repro-ductive cycle demonstrating that these cells do not secrete inparallel andmelatonin acts differentially on the gonadotrophactivity [28] In addition corticotrophs somatotrophs thy-rotrophs and lactotrophs vary throughout the reproductivecycle [29ndash32] A reduction of the testicular weight and ofthe seminiferous tubule diameters seminiferous epitheliumwith Sertoli cells spermatogonia and a few primary sperma-tocytes and hypotrophic Leydig cells was observed in winter[33 34] The testicular activity was slowly recovered duringspring reaching its maximum during summer and earlyautumn in the reproductive period More recent results fromadult male viscachas have demonstrated seasonal changes inthe structures of seminal vesicles according to the AR expres-sion and testosterone serum concentrations [35] Crucenoet al [36] reported that the epididymal sperm morphologyunderwent changes during the reproductive cycle with thehighest quantity of abnormal gametes during the gonadalregression period
Earlier histological studies have described different pitu-itary PD zones called ventral medial and dorsal regionsand rostral and caudal ends It has been demonstratedthat pituitary cell types changed in relation to age andgonadal activity of viscachas [29 30 32] Changes in theimmunostaining for AR and LH- and FSH-cells have beenrecently reported in the pituitary of castrated adult maleviscachasThe immunocolocalization study showed that cellsexpressing AR and FSH were more affected by the lack ofgonadal androgens suggesting that some gonadotroph sub-populations may exist with different regulatory mechanismsfor hormonal synthesis storage and secretion [37]
The expression of AR in the pituitary of fetal immatureand prepubertal viscachas has not been examined yet andthere is no data about the seasonal expression of AR inthe pituitary gland of this rodent Based on the abovedata and our previous knowledge about the viscacha it
is reasonable to hypothesize that pituitary AR expressionchanges throughout development growth and reproductivecycle of male viscacha To test this hypothesis we performedan immunohistochemical and morphometrical study of ARin the pituitary PD of male viscachas during developmentgrowth and the annual reproductive cycle This immunohis-tochemical study might be useful for discussing the probablerole of androgens in the pituitary development of viscachasand their participation in the reproductive stage along theyear
2 Materials and Methods
21 Animals The viscachas were captured in their habitatnear San Luis Argentina (33∘ 201015840 south latitude 760maltitude) using traps placed in their burrows In San Luis insummer the light phase is up to 14 h light daily (14L 10D)with an average temperature of 25∘C Inwinter the light phasedecreases to 10 h (10L 14D) and the average temperature is10∘C In spring the light phase increases to 12 h (12L 12D)and the average temperature is 15∘C
A total of eight fetal male pituitary glands were collectedThe fetal pituitaries were collected from fetuses at mid- (119899 =4) and late (119899 = 4) pregnant females classified on thebasis of fetal weight and crown-heel length [38 39] Themale animals were carefully classified into immature (1-2 kg119899 = 4) and prepubertal (3-4 kg 119899 = 4) according to bodyweight and light microscopy observations of testes [30 40]Twelve adult male viscachas weighing 5ndash7 kg were capturedduring the most representative months of their reproductivecycle for a period of over 1 year 4 animals during thereproductive period in summer to early autumn (Februaryto April) 4 animals in the gonadal regression period inwinter (July) and 4 animals in the gonadal recovery period inspring (September) The reproductive condition of viscachaswas carefully assessed on the basis of observations by lightmicroscopy of testes
The animals were intramuscularly anesthetized with acombination of ketamine (Ketamina 50 Holliday-Scott SA)and xylazine (Vetanarcol Konig SA) at a dose of 12 and04mgkg respectively The blood was collected for cardiacpuncture for the evaluation of serum hormone concentrationand quickly sacrificed by intracardiac injection of Euthanyle(025mLKg body weight sodium pentobarbital sodiumdiphenylhydantoin Brouwer SA) The brain was rapidlyexposed and the pituitary processed for lightmicroscopyThepituitaries were fixed in Bouinrsquos fluid embedded in paraffinand sagittally sectioned (5 120583m thick) following the designused in previously reported studies [30 32]
The experimental design was approved by the local ethicscommittee and was in agreement with the National Instituteof Health (NIH USA) guidelines for the use of experimentalanimals Moreover the Biodiversity Control Area of the SanLuis Ministry of the Environment (Argentina) approved astudy protocol for conducting scientific research within theterritory of this province (Resolution number 45PBD2014)
22 Immunohistochemistry of AR in the Pituitary The tis-sue sections were stained using the streptavidin-biotin
International Journal of Endocrinology 3
(A) (B)
(C) (D)
Figure 1 (A) Rat prostate a positive control of immunoperoxidase staining with anti-AR (N-20) without hematoxylin staining Insetsimmunolabeled nuclei of epithelial cells (arrows) without (left) and with (right) counterstained hematoxylin (B) Pituitary of viscacha anegative control of immunohistochemistry counterstaining with hematoxylin ((C) (D)) Pituitary of viscacha staining with anti-AR (N-20)without hematoxylin staining (C) and counterstaining with hematoxylin (D) ARn-ir cells (arrows) Blood vessels v follicular structures f(A) Scale bar 100120583m (B) (C) and (D) and insets scale bar 25 120583m
peroxidase complex method at 20∘C The sections werefirst deparaffinized with xylene hydrated through decreasingconcentrations of ethanol and rinsed with distilled waterand phosphate-buffered saline (PBS 001M pH 74) Anti-gen retrieval was performed by microwaving the sectionsfor 6min (2 times 3min) at full power in a sodium citratebuffer (001M pH 60) Endogenous peroxidase activitywas inhibited with 3 H
2O2in water for 20min Non-
specific binding sites for immunoglobulins were blockedby incubation for 20min with normal serum diluted inPBS containing 1 bovine serum albumin 009 sodiumazide and 01 Tween-20 Sections were incubated withthe primary antibody 6 hours in a humidified chamber at20∘C with rabbit polyclonal anti-human (h) AR (N-20 SantaCruz Biotechnology Santa Cruz CA USA) After rinsingwith PBS for 10min immunohistochemical visualizationwas carried out using the Super Sensitive Ready-to-UseImmunostaining Kit (BioGenex San Ramon CA USA)whichwas used as follows sectionswere incubated for 30minwith diluted biotinylated anti-IgG and after being washed inPBS were incubated for 30min with horseradish peroxidaseconjugated streptavidin and were finally washed in PBS ARimmunoreactivity cells (AR-ir cells) were visualized usingfreshly prepared solution with 100 120583L 331015840-diaminobenzidinetetrahydrochloride chromogen in 25mL PBS and 50 120583LH2O2substrate solution In all cases two experiments for
controlling the specificity of the primary antibodyweremade
(i) omission of primary antibody and (ii) adsorption ofprimary antibody with a homologous antigen No positivestructures or cells were found in these sections Rat prostatewas used as positive control (Figure 1)
23 Morphometric Analysis A computer-assisted imageanalysis system was used for the morphometric analysisas previously reported [37 41] The image was displayedon a color monitor a standard area of 1814182 120583m2 (refer-ence area) was defined on the monitor distance calibrationwas performed using a slide with a micrometric scale formicroscopy (Reichert Austria) The morphometric studywas carried out as follows 3-4 regularly spaced serial tissuesections (100120583m each) from a pituitary were used andmicroscopic fields were examined under a 40x objective Ineach section 25 microscopic fields were randomly selectedthroughout the PD five from each region (ventral medialand dorsal regions) or end (rostral and caudal ends) ofthe PD In each image the percentage of immunoreactive(-ir) cells in the PD (percentages of ARn-ir ARc-ir) wasobtained according to the formula 119860(119860 + 119861) times 100 Eachimage contained approximately 250ndash280 cells The numberof immunoreactive cells (119860) and the number of nuclei inimmunonegative cells (119861) were counted
24 Testosterone Serum Concentrations Blood samples wereincubated at 4∘C for 30min and centrifuged at 5000 g for
4 International Journal of Endocrinology
005
115
225
335
445
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
Caudal ERostral ECentral R
Dorsal RVentral R
ARc
-ir ce
lls (
)lowastlowast
lowastlowast
(a)
02468
101214161820
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
Caudal ERostral ECentral R
Dorsal RVentral R
ARn
-ir ce
lls (
) lowastlowast
lowast lowast
lowast
lowast
(b)
Figure 2 Morphometry of AR-ir cells in pituitary of male viscacha in different pituitary PD zones (a) ARc-ir cells LP-FETUS lowastlowast119875 lt 001dorsal R versus ventral R rostral E and caudal E Prepubertal lowastlowast119875 lt 001medial R versus ventral R and caudal E Adults the percentageof ARc-ir cells did not reveal statistically significant changes in PD zones (b) ARn-ir cells Immature lowast119875 lt 005medial R versus dorsal Rand rostral E Prepubertal lowast119875 lt 005medial R versus dorsal R and ventral R Adult-Rep P lowast119875 lt 005 caudal E versus the other PD zonesAdult-Reg P lowast119875 lt 005medial R versus ventral R and rostral E Adult-Rec P lowastlowast119875 lt 001 caudal E versus rostral E and medial R Fetusesfrom midpregnancy MP-FETUS fetuses from late pregnancy LP-FETUS reproductive period Rep P gonadal regression period Reg Pgonadal recovery period Rec P region R end E
5min and serum was removed Testosterone serum concen-trations were measured using the total testosterone test aspreviously reported by Chaves et al [35] This test is a solid-phase competitive chemiluminescent enzyme immunoassayrun on a SiemenslowastMedical IMMULITElowast 1000 ImmunoassayAnalyzer (Siemens Medical Solutions Diagnostics)
25 Statistical Analysis The results were expressed asmean plusmnthe standard error of the mean (SEM) for all morphologicaldata sets The data were analyzed by nonparametric testprocedure the Kruskal-Wallis test for overall significanceof group followed by the Mann-Whitney test for individualcomparisons using the Infostat 2008 version A 119875 value ofless than 005 was considered statistically significant
3 Results
AR-ir cells were studied by immunohistochemistry on eachzone of the pituitary PD parenchyma of fetal immatureprepubertal and adult male viscachas The AR-ir cells wereobserved throughout the PD parenchyma and immunohis-tochemical results revealed their presence in the nuclei (ARn-ir) and in the cytoplasm (ARc-ir)
In fetal pituitary from midpregnancy scarce ARn-ir cellsand intensely stained ARc-ir cells were located in the medialregion and extended towards the ventral and dorsal regionsHowever the percentages of ARn-ir cells (caudal end 004 plusmn002 rostral end 005 plusmn 003 central region 055 plusmn 015dorsal region 015 plusmn 007 ventral region 016 plusmn 007)and the percentages of ARc-ir cells did not differ significantly
among different zones of the PD (119875 gt 005) (Figures 2(a)3(A) and 3(B))
In fetal pituitary from late pregnancy ARc-ir cellswere found around blood vessels or among other pituitaryparenchymal cellsThepercentage ofARc-ir cells in the dorsalregion was significantly higher (119875 lt 001) than in the ventralregion and at the rostral and caudal ends (Figures 2(a) 3(C)and 3(D)) ARn-ir cells were few and frequently sphericaland some of them were irregular in shape The percentage ofthese cells did not vary among different PD zones (119875 gt 005caudal end 054 plusmn 023 rostral end 021 plusmn 002 centralregion 025plusmn010 dorsal region 040plusmn010 ventral region028 plusmn 010)
In pituitary PD of immature animals the ARc-ir cellswere located in the medial region and extended towardsthe caudal end The cytoplasmic immunostaining was lessintense than in fetal glands cells Several ARc-ir cells wereoval and their spherical nucleus was located at one end ofthe cytoplasm The percentage of ARc-ir cells did not differsignificantly (119875 gt 005) in different PD zones (Figure 2(a))Intensely stained ARn-ir cells were mainly observed inthe medial region where their percentage was significantlyhigher than in the dorsal region and rostral end (119875 lt 005Figures 2(b) and 3(E)ndash3(H))
In prepubertal viscachas numerous immunopositive AR-ir cells were distributed throughout the PD parenchymaThese cells were arranged around blood vessels at the rostralend and in the medial and dorsal regions In the ventralregion there were some irregular AR-ir cells with a cyto-plasmic prolongation surrounding blood vessels or betweenother parenchymal cells The ARc-ir cells showed significant
International Journal of Endocrinology 5
(A) (B)
(C)
(E) (F) (H)
(I) (J) (K)
(G)
(D)
Figure 3 AR immunohistochemical staining of viscacha pituitaries ((A) (B)) Sections of a fetal pituitary from midpregnancy showing thedifferent zones of PD (A) In the medial region ARn-ir and ARc-ir cells were present (B) Inset ARc-ir cells (c) with a negative nucleus((C) (D)) Images of a fetal pituitary from late pregnancy Isolated ARc-ir cells around blood vessel (D) ((E) (F) (G) and (H)) AR-ir cellsin a pituitary gland of immature viscacha There are cells throughout the PD (E) The ARn-ir cells are increased in the medial region (F)Magnification of ARn-ir cells (G) and a pituitary cell with heterogeneous cytoplasmic staining (H) ((I) (J) and (K)) Pituitary of a prepubertalviscacha ARc-ir cells in groups in contact with blood vessel (J) and others with a cytoplasmic prolongation (K) ARn-ir cells (arrows) ARc-ircells (arrowheads) PD pars distalis PI pars intermedia PN pars nervosa R Rathkersquos pouch ce caudal end re rostral end vr ventral regionmr medial region dr dorsal region blood vessels v ((A) (C) and (I)) Scale bar 500 120583m (E) Scale bar 250 120583m ((B) (D) (F) and (J)) Scalebar 25 120583m Inset of (B) (G) (H) and (K) scale bar 10 120583m Images counterstaining with hematoxylin
6 International Journal of Endocrinology
Table 1 AR expression in pituitary PD of male viscachas
Distribution Immunostaining pattern Morphological characteristicsMP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cellsLP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cells
Immature Isolated or in small groups Homogeneous and heterogeneous cytoplasmsome intensely stained nuclei Cells oval in shape
Prepubertal Isolated or in groups Homogeneous cytoplasmintensely nuclear staining Irregular cells
Adult-Rep P Isolated or in small groups Nuclear immunoreactivityscarce immunostained cytoplasms Oval spherical or irregular cells
Adult-Reg P Isolated Nuclear immunoreactivityscarce immunostained cytoplasms Small and spherical cytoplasm
Adult-Rec P Isolated or in small groups Nuclear immunoreactivityintense cytoplasmic staining
Spherical oval or irregular cytoplasmsand nuclei
short cytoplasmic prolongation
variations the highest value was found in the medial regionin relation to the ventral region and caudal end (119875 lt001) whereas the lowest percentage was found at the caudalend (Figure 2(a)) The immunostaining of ARn-ir cells wasintense in the dorsal region and the highest percentage ofthese cells was found in the medial region in relation to thedorsal and ventral regions and rostral end (119875 lt 005 Figures2(b) and 3(I)ndash3(K))
In PD of adult animals the AR expression mainly hada nuclear localization In the reproductive period severalpositive nuclei were found at the caudal end and dorsalregion as well as on the dorsal and ventral edges of PDThe percentage of ARn-ir cells was significantly higher atthe caudal end in relation to the other PD zones (119875 lt005 Figure 2(b)) A small number of ARc-ir cells werefrequently located in the medial region The cytoplasmicimmunolabeling pattern was homogeneous and sometimesintense In addition some cells presented heterogeneouscytoplasmic staining and positivity on the nuclear periphery(Figures 4(A)ndash4(E)) No significant changes were observedin the percentage of ARc-ir cells between different PD zones(119875 gt 005 Figure 2(a))
In the gonadal regression period AR-ir cells were mainlyfound in the medial region of PD The immunostainedcytoplasms were small with medial or eccentric nucleusThere were no significant differences in ARc-ir cells betweendifferent PD zones (119875 gt 005 Figure 2(a)) whereas ARn-ircells were more numerous in the medial region in relation tothe ventral region and the rostral end (119875 lt 005 Figure 2(b))
In the gonadal recovery period an increase of AR-ir cellsthroughout PD parenchyma was observed In this periodthe cytoplasmic labeling was more intense and homogeneousin relation to the gonadal regression period (Figures 4(F)ndash4(I)) The percentage of ARc-ir cells was not significantlydifferent between the PD zones (119875 gt 005 Figure 2(a)) Thepercentage of ARn-ir cells was higher at the caudal end inrelation to the rostral end and the medial region (119875 lt 001Figure 2(b)) Table 1 shows a summary of themain cytologicalcharacteristics
The morphometric variations of the AR expression inrelation to development and growth in viscachas PD werestudied by comparing the data obtained from fetal pituitaryduring mid- and late pregnancy and from immature prepu-bertal and adult pituitary in the reproductive period Therewas an increase in ARn-ir cells percentage and a significantdecrease in the percentage of ARc-ir cells in immatureanimals as compared to fetus and in adult animals duringthe reproductive period as compared to prepubertal animalsThe percentage of ARc-ir cells increased significantly inprepubertal in relation to immature viscachas Statisticalanalysis is shown in Figure 5
In order to study the seasonal expression of AR the threeperiods of the reproductive cycle were analyzed The totalpercentages of ARn-ir and ARc-ir cells obtained from threeperiods of the reproductive cycle of adult male viscachaswere statistically analyzed (Figure 5)The percentage of ARn-ir cells showed a significant decrease during the gonadalregression period (119875 lt 0001) No variations were observedin the percentage of ARc-ir cells throughout the reproductivecycle (119875 gt 005)
The testosterone serum concentrations of immature ani-mals were below the detection limit (lt020 ngmL) and toolow to reliably reveal differences from the other groupsThe testosterone serum concentrations of prepubertal andadult viscachas in the gonadal regression period were lowerthan those in adult viscachas during their reproductive andgonadal recovery periods (119875 lt 0001 Figure 6)
4 Discussion
The analysis of the AR expression in the viscacha pituitarygland is essential for a better understanding of the regulatorysystems since androgens are involved in the control ofthe reproductive function This study represents the firstdescription of variations in the regionalization and in theimmunostaining pattern of pituitary AR in relation to thedevelopment growth and reproductive cycle of male vis-cachas
International Journal of Endocrinology 7
(A) (B)
(C) (D) (E)
(F) (G)
(H) (I)
Figure 4 Seasonal expression of AR in pituitary during the reproductive cycle of adult viscachas ((A)ndash(E)) Reproductive period ((A) (B))Images of caudal end the AR were expressed mainly in the nuclei of PD cells (arrows) ((C) (D) and (E)) AR expression at the rostral endventral region and dorsal region respectively The arrowhead indicated a cell with cytoplasmic labeling for AR ((F) (G)) A few AR-ir cellsat caudal end during the gonadal regression period ((H) (I)) AR-ir cells at caudal end during the gonadal recovery period Caudal endce blood vessels v follicular structures f ((A) (F) and (H)) Scale bar 100 120583m ((B) (C) (D) (E) (G) and (I)) Scale bar 25 120583m Imagescounterstaining with hematoxylin
Themain findings of this study can be summarized as fol-lows (1) the regionalization and immunostaining pattern ofpituitary AR vary in relation to growth (2) the AR expressionwas mainly cytoplasmic (ARc-ir) in fetuses (3) a significant
increase of ARc-ir cells was observed in prepubertal animals(4) the nuclear AR (ARn-ir) expression increased in relationto development reaching the highest values in adult animals(5) the ARn-ir cells varied seasonally with maximum values
8 International Journal of Endocrinology
0123456789
10
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
ARn-ir cells ()ARc-ir cells ()
Pitu
itary
PD
()
lowastlowastlowastlowastlowastlowast
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 5 Total percentage of ARn-ir cells and ARc-ir cells infetus (from mid- and late pregnancy) immature prepubertaland adult viscachas (during reproductive gonadal regression andgonadal recovery periods) The result revealed statistically signifi-cant changes lowastlowastlowast119875 lt 0001 immature versus LP-FETUS prepu-bertal versus immature Adult-Rep P versus prepubertal Adult-Reg P versus Adult-Rep P Adult-Rec P versus Adult-Reg P
0
1
2
3
4
5
6
7
Prepubertal Adult-Rep P Adult-Reg P Adult-Rec P
Seru
m te
stoste
rone
conc
entr
atio
n (n
gm
L)
lowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 6 Testosterone serum concentrations The values areexpressed as mean plusmn SEM (119899 = 4) lowastlowast119875 lt 001 Adult-Rep Pversus Adult-Reg P lowastlowastlowast119875 lt 0001 prepubertal versus Adult-RepP Adult-Reg P versus Adult-Rec P
during the reproductive period and minimum values duringthe gonadal regression period (6) the amount of ARn-ir cellswas directly related to testosterone serum concentration
There are still many questions about the relations ofAR expression with the pituitary development and hormonesecretion Wu et al [12] did not observe AR in the pituitarygland during late gestation and suggested that the lack of
feedback by way of AR at pituitary levels may have importantconsequences on the fetal pituitary function More recentlyYuan et al [18] have reported in sheep that AR emerges inthe anterior pituitary before day 60 of gestation and thepercentage of the total anterior pituitary cells expressing thereceptor did not change throughout gestation In additionother studies have concluded that the fetal brain and pitu-itary are target sites for circulating androgens or androgenprecursors in fetal plasma and the increase of androgensin hypothalamic action immediately prior to birth might becritical to the timing of parturition [13] Huang and Yuan[42] have reported that AR expression reflects the functionalstatus of androgen in the pituitary of male sheep lambTheseauthors suggest that during sexual maturity the feedback ofsex hormones on the pituitary is at low level and stimulatesgonadotropin secretion
Some reproductive characteristics of female viscachassuch as polyovulation implantation and offspring birth havebeen described in previous studies Gestation lasts approx-imately 154 days an unusually long period for a rodentand it finishes in spring when the survival probabilities areideal for the mother and offspring [38 43 44] The resultsobtained in this work demonstrate AR in fetal pituitary frommidpregnancy indicating that AR are present around days75ndash80 of gestation in the pituitary cells These receptors arepotentially in position to participate in the regulation of thefetal androgen function Furthermore the analysis of thedistribution morphometry and immunostaining pattern ofAR-ir cells in the parenchyma of fetal glands suggests thatthese cells contain variable amounts of AR and they aremorphologically related to blood supply and other endocrinecells of PD parenchyma In late pregnancy of viscacha thepercentages of ARc-ir and ARn-ir cells did not change withrespect to the values found during midpregnancy Theseresults suggested that fetal pituitary AR might have anyspecific function since midgestation in viscacha
Thehypothalamic-pituitary-gonadal axis is activated dur-ing puberty when hormonalmodulation of sexual behavior isorganized [45] During male development the masculiniza-tion is given by the hormones produced by the fetal testesprimarily testosterone [46 47] The gonadal steroids feedback to the central nervous system to exert a key role inthe central control of the reproductive axis by regulatingthe release of the gonadotropin-releasing hormone (GnRH)[48] However the neurons that synthesize GnRH did notexpress AR and there are sex-steroids sensitive neuronsupstream the GnRH cells where the feedback signals ofthe steroid might be processed to the GnRH cells [49] Theneurons of hypothalamic nuclei that expressed kisspeptin(K) and neurokinin B (NKB) may sense circulating levelsof testosterone through the expression of AR [49 50] Inrodents sheep and primates K and NKB have recently beenimplicated in the control of puberty and the maintenanceof the reproductive function in adulthood [51ndash53] In thepituitary the expression of AR has been mainly observedin gonadotrophs [18 54 55] and gonadal steroid hormonesaffect the gonadotroph population in order to regulate theiractivity
International Journal of Endocrinology 9
In immature male viscachas a significant increase ofnuclear AR and an important decrease of cytoplasmic ARwere observed In prepubertal viscachas more nuclei andcytoplasm appear and there is a clear increase of ARc-ir cellsmainly in the medial region suggesting that more cells beginto express AR in preparation for sexual maturation In adultthe immunostaining is mainly nuclear and regionalized atthe caudal end of PD suggesting that AR are active in thePD cells The present results demonstrate variations in theregionalization and immunostaining pattern of AR duringsexual maturity probably in relation to the contribution ofandrogens reaching the different pituitary zones throughblood vessels Thus the nuclear AR expression in pituitaryPD of viscacha varies according to gonadal developmentand testosterone serum concentration Our results are inagreement with those found by Wood and Newman [24]suggesting that a subset of unoccupied receptors is located inthe cell cytoplasm in the absence of ligand
Bibliographic reports referring to seasonal variations ofRA are scarce Lu et al [56] have recently demonstrated astronger expression ofAR in scented glands during the breed-ing season suggesting that androgens may directly influ-ence the glandular function of the muskrats and courtshipbehavior The viscacha is a seasonal breeding rodent andprevious studies have demonstrated that testosterone acts ongonadotrophs whether directly or indirectly regulating theiractivity in the pituitary gland [28] Castrated viscachamodelshave been used to examine the withdrawal effects of circulat-ing androgens on pituitary cells mainly on the gonadotrophsdue to their involvement in the reproductive processes Inaddition castration resulted in a significant decrease in thepercentage of cells expressing AR throughout the PD [37]The present results demonstrate changes in the amount ofpituitary AR-ir cells during the annual reproductive cycleof adult male viscacha which might be due to variations inthe supply of ligands across blood irrigation The maximumvalues of AR expression were found in the reproductiveperiod and the minimum values in the regression period inrelation to androgen concentrationsThus ARmight regulatethe activity of the pituitary cells involved in the reproductivecycle of adult male Lagostomus
In conclusion our results demonstrate thatAR expressionis related to the pituitary histophysiological changes duringthe growth and sexual maturity of male viscacha Moreoverthey are likely to participate in the seasonal reproductioncontrol in adulthoodHowever the precise actionmechanismof AR in the pituitary viscacha needs further study
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Mrs A Bernardi and Mr JArroyuelo for their technical participation This work wassupported by Project 2-2614 Secretaria de Ciencia y TecnicaUniversidad Nacional de San Luis
References
[1] A K Roy Y Lavrovsky C S Song et al ldquoRegulation ofandrogen actionrdquo Vitamins amp Hormones vol 55 pp 309ndash3521999
[2] S Denayer C Helsen L Thorrez A Haelens and F ClaessensldquoThe rules of DNA recognition by the androgen receptorrdquoMolecular Endocrinology vol 24 no 5 pp 898ndash913 2010
[3] E V Jensen G L Greene L E Closs E R DeSombre and MNadji ldquoReceptors reconsidered a 20-year perspectiverdquo RecentProgress in Hormone Research vol 38 pp 1ndash40 1982
[4] B D SchanbacherM P Johnson andD J Tindall ldquoAndrogenicregulation of luteinizing hormone secretion relationship toandrogen binding in sheep pituitaryrdquo Biology of Reproductionvol 36 no 2 pp 340ndash350 1987
[5] R J Handa D L Reid and J A Resko ldquoAndrogen receptorsin brain and pituitary of female rats cyclic changes andcomparisons with the malerdquo Biology of Reproduction vol 34no 2 pp 293ndash303 1986
[6] J V A Choate and J A Resko ldquoEffects of androgen on brain andpituitary androgen receptors and LH secretion of male guineapigsrdquoThe Journal of Steroid Biochemistry andMolecular Biologyvol 59 no 3-4 pp 315ndash322 1996
[7] T J Spady R Shayya V GThackray L Ehrensberger J S Bai-ley and P L Mellon ldquoAndrogen regulates follicle-stimulatinghormone 120573 gene expression in an activin-dependent manner inimmortalized gonadotropesrdquo Molecular Endocrinology vol 18no 4 pp 925ndash940 2004
[8] P B Connolly J V A Choate and J A Resko ldquoEffects ofexogenous androgen on brain androgen receptors of the fetalrhesus monkeyrdquoNeuroendocrinology vol 59 no 3 pp 271ndash2761994
[9] J A Resko A Perkins C E Roselli J A Fitzgerald J V AChoate and F Stormshak ldquoEndocrine correlates of partnerpreference behavior in ramsrdquo Biology of Reproduction vol 55no 1 pp 120ndash126 1996
[10] S E Abdelgadir C E Roselli J V A Choate and J A ReskoldquoAndrogen receptor messenger ribonucleic acid in brains andpituitaries of male Rhesus monkeys studies on distributionhormonal control and relationship to luteinizing hormonesecretionrdquo Biology of Reproduction vol 60 no 5 pp 1251ndash12561999
[11] R W Bonsall H D Rees and R P Michael ldquoThe distributionnuclear uptake and metabolism of [3H]dihydrotestosterone inthe brain pituitary gland and genital tract of the male rhesusmonkeyrdquo Journal of Steroid Biochemistry vol 23 no 4 pp 389ndash398 1985
[12] S S Wu P W Nathanielsz and T J McDonald ldquoImmunocy-tochemical distribution of androgen receptors in the hypotha-lamus and pituitary of the fetal baboon in late gestationrdquoDevelopmental Brain Research vol 84 no 2 pp 278ndash281 1995
[13] C E Wood and M Keller-Wood ldquoOntogeny of androgenreceptor expression in the ovine fetal central nervous systemand pituitaryrdquo Neuroscience Letters vol 439 no 2 pp 153ndash1562008
[14] J V A Choate O D Slayden and J A Resko ldquoImmuno-cytochemical localization of androgen receptors in brains ofdeveloping and adult male rhesus monkeysrdquo Endocrine vol 8no 1 pp 51ndash60 1998
10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 International Journal of Endocrinology
the hypothalamic-pituitary-gonadal axis [20] Variations inthe androgen serum concentration and their receptors in dif-ferent tissues are responsible for biological effects throughoutthe life cycle of adult animals Testosterone plays a highlysignificant role in the transition from the reproductive activ-ity phase to gonadal regression and gonadal recrudescence[21] Seasonal variations in testosterone serum concentrationare related to the reproductive status of male birds [22] andhamsters [23] The central mechanisms responsible for thephotoperiod negative effects upon the testosterone and thepossible influence of day length on AR expression have beenreported [24] Tetel et al [25] have indicated that day-length-induced fluctuations in AR expression may contribute toseasonal variations of testosterone
Our experimental model the viscacha (Lagostomus max-imus maximus) is a hystricomorph rodent of seasonalreproductive patternsThe annual reproductive cycle of adultmale animals has three periods reproductive (summer andearly autumn) gonadal regression (winter) and gonadalrecovery (spring) Previous studies have demonstrated thatthe gonadal regression period is characterized by an increaseof the secretory ability of the pineal gland [26] andmaximumlevels of melatonin in blood [27] In pituitary PD the activityof LH- and FSH-cells was different during the annual repro-ductive cycle demonstrating that these cells do not secrete inparallel andmelatonin acts differentially on the gonadotrophactivity [28] In addition corticotrophs somatotrophs thy-rotrophs and lactotrophs vary throughout the reproductivecycle [29ndash32] A reduction of the testicular weight and ofthe seminiferous tubule diameters seminiferous epitheliumwith Sertoli cells spermatogonia and a few primary sperma-tocytes and hypotrophic Leydig cells was observed in winter[33 34] The testicular activity was slowly recovered duringspring reaching its maximum during summer and earlyautumn in the reproductive period More recent results fromadult male viscachas have demonstrated seasonal changes inthe structures of seminal vesicles according to the AR expres-sion and testosterone serum concentrations [35] Crucenoet al [36] reported that the epididymal sperm morphologyunderwent changes during the reproductive cycle with thehighest quantity of abnormal gametes during the gonadalregression period
Earlier histological studies have described different pitu-itary PD zones called ventral medial and dorsal regionsand rostral and caudal ends It has been demonstratedthat pituitary cell types changed in relation to age andgonadal activity of viscachas [29 30 32] Changes in theimmunostaining for AR and LH- and FSH-cells have beenrecently reported in the pituitary of castrated adult maleviscachasThe immunocolocalization study showed that cellsexpressing AR and FSH were more affected by the lack ofgonadal androgens suggesting that some gonadotroph sub-populations may exist with different regulatory mechanismsfor hormonal synthesis storage and secretion [37]
The expression of AR in the pituitary of fetal immatureand prepubertal viscachas has not been examined yet andthere is no data about the seasonal expression of AR inthe pituitary gland of this rodent Based on the abovedata and our previous knowledge about the viscacha it
is reasonable to hypothesize that pituitary AR expressionchanges throughout development growth and reproductivecycle of male viscacha To test this hypothesis we performedan immunohistochemical and morphometrical study of ARin the pituitary PD of male viscachas during developmentgrowth and the annual reproductive cycle This immunohis-tochemical study might be useful for discussing the probablerole of androgens in the pituitary development of viscachasand their participation in the reproductive stage along theyear
2 Materials and Methods
21 Animals The viscachas were captured in their habitatnear San Luis Argentina (33∘ 201015840 south latitude 760maltitude) using traps placed in their burrows In San Luis insummer the light phase is up to 14 h light daily (14L 10D)with an average temperature of 25∘C Inwinter the light phasedecreases to 10 h (10L 14D) and the average temperature is10∘C In spring the light phase increases to 12 h (12L 12D)and the average temperature is 15∘C
A total of eight fetal male pituitary glands were collectedThe fetal pituitaries were collected from fetuses at mid- (119899 =4) and late (119899 = 4) pregnant females classified on thebasis of fetal weight and crown-heel length [38 39] Themale animals were carefully classified into immature (1-2 kg119899 = 4) and prepubertal (3-4 kg 119899 = 4) according to bodyweight and light microscopy observations of testes [30 40]Twelve adult male viscachas weighing 5ndash7 kg were capturedduring the most representative months of their reproductivecycle for a period of over 1 year 4 animals during thereproductive period in summer to early autumn (Februaryto April) 4 animals in the gonadal regression period inwinter (July) and 4 animals in the gonadal recovery period inspring (September) The reproductive condition of viscachaswas carefully assessed on the basis of observations by lightmicroscopy of testes
The animals were intramuscularly anesthetized with acombination of ketamine (Ketamina 50 Holliday-Scott SA)and xylazine (Vetanarcol Konig SA) at a dose of 12 and04mgkg respectively The blood was collected for cardiacpuncture for the evaluation of serum hormone concentrationand quickly sacrificed by intracardiac injection of Euthanyle(025mLKg body weight sodium pentobarbital sodiumdiphenylhydantoin Brouwer SA) The brain was rapidlyexposed and the pituitary processed for lightmicroscopyThepituitaries were fixed in Bouinrsquos fluid embedded in paraffinand sagittally sectioned (5 120583m thick) following the designused in previously reported studies [30 32]
The experimental design was approved by the local ethicscommittee and was in agreement with the National Instituteof Health (NIH USA) guidelines for the use of experimentalanimals Moreover the Biodiversity Control Area of the SanLuis Ministry of the Environment (Argentina) approved astudy protocol for conducting scientific research within theterritory of this province (Resolution number 45PBD2014)
22 Immunohistochemistry of AR in the Pituitary The tis-sue sections were stained using the streptavidin-biotin
International Journal of Endocrinology 3
(A) (B)
(C) (D)
Figure 1 (A) Rat prostate a positive control of immunoperoxidase staining with anti-AR (N-20) without hematoxylin staining Insetsimmunolabeled nuclei of epithelial cells (arrows) without (left) and with (right) counterstained hematoxylin (B) Pituitary of viscacha anegative control of immunohistochemistry counterstaining with hematoxylin ((C) (D)) Pituitary of viscacha staining with anti-AR (N-20)without hematoxylin staining (C) and counterstaining with hematoxylin (D) ARn-ir cells (arrows) Blood vessels v follicular structures f(A) Scale bar 100120583m (B) (C) and (D) and insets scale bar 25 120583m
peroxidase complex method at 20∘C The sections werefirst deparaffinized with xylene hydrated through decreasingconcentrations of ethanol and rinsed with distilled waterand phosphate-buffered saline (PBS 001M pH 74) Anti-gen retrieval was performed by microwaving the sectionsfor 6min (2 times 3min) at full power in a sodium citratebuffer (001M pH 60) Endogenous peroxidase activitywas inhibited with 3 H
2O2in water for 20min Non-
specific binding sites for immunoglobulins were blockedby incubation for 20min with normal serum diluted inPBS containing 1 bovine serum albumin 009 sodiumazide and 01 Tween-20 Sections were incubated withthe primary antibody 6 hours in a humidified chamber at20∘C with rabbit polyclonal anti-human (h) AR (N-20 SantaCruz Biotechnology Santa Cruz CA USA) After rinsingwith PBS for 10min immunohistochemical visualizationwas carried out using the Super Sensitive Ready-to-UseImmunostaining Kit (BioGenex San Ramon CA USA)whichwas used as follows sectionswere incubated for 30minwith diluted biotinylated anti-IgG and after being washed inPBS were incubated for 30min with horseradish peroxidaseconjugated streptavidin and were finally washed in PBS ARimmunoreactivity cells (AR-ir cells) were visualized usingfreshly prepared solution with 100 120583L 331015840-diaminobenzidinetetrahydrochloride chromogen in 25mL PBS and 50 120583LH2O2substrate solution In all cases two experiments for
controlling the specificity of the primary antibodyweremade
(i) omission of primary antibody and (ii) adsorption ofprimary antibody with a homologous antigen No positivestructures or cells were found in these sections Rat prostatewas used as positive control (Figure 1)
23 Morphometric Analysis A computer-assisted imageanalysis system was used for the morphometric analysisas previously reported [37 41] The image was displayedon a color monitor a standard area of 1814182 120583m2 (refer-ence area) was defined on the monitor distance calibrationwas performed using a slide with a micrometric scale formicroscopy (Reichert Austria) The morphometric studywas carried out as follows 3-4 regularly spaced serial tissuesections (100120583m each) from a pituitary were used andmicroscopic fields were examined under a 40x objective Ineach section 25 microscopic fields were randomly selectedthroughout the PD five from each region (ventral medialand dorsal regions) or end (rostral and caudal ends) ofthe PD In each image the percentage of immunoreactive(-ir) cells in the PD (percentages of ARn-ir ARc-ir) wasobtained according to the formula 119860(119860 + 119861) times 100 Eachimage contained approximately 250ndash280 cells The numberof immunoreactive cells (119860) and the number of nuclei inimmunonegative cells (119861) were counted
24 Testosterone Serum Concentrations Blood samples wereincubated at 4∘C for 30min and centrifuged at 5000 g for
4 International Journal of Endocrinology
005
115
225
335
445
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
Caudal ERostral ECentral R
Dorsal RVentral R
ARc
-ir ce
lls (
)lowastlowast
lowastlowast
(a)
02468
101214161820
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
Caudal ERostral ECentral R
Dorsal RVentral R
ARn
-ir ce
lls (
) lowastlowast
lowast lowast
lowast
lowast
(b)
Figure 2 Morphometry of AR-ir cells in pituitary of male viscacha in different pituitary PD zones (a) ARc-ir cells LP-FETUS lowastlowast119875 lt 001dorsal R versus ventral R rostral E and caudal E Prepubertal lowastlowast119875 lt 001medial R versus ventral R and caudal E Adults the percentageof ARc-ir cells did not reveal statistically significant changes in PD zones (b) ARn-ir cells Immature lowast119875 lt 005medial R versus dorsal Rand rostral E Prepubertal lowast119875 lt 005medial R versus dorsal R and ventral R Adult-Rep P lowast119875 lt 005 caudal E versus the other PD zonesAdult-Reg P lowast119875 lt 005medial R versus ventral R and rostral E Adult-Rec P lowastlowast119875 lt 001 caudal E versus rostral E and medial R Fetusesfrom midpregnancy MP-FETUS fetuses from late pregnancy LP-FETUS reproductive period Rep P gonadal regression period Reg Pgonadal recovery period Rec P region R end E
5min and serum was removed Testosterone serum concen-trations were measured using the total testosterone test aspreviously reported by Chaves et al [35] This test is a solid-phase competitive chemiluminescent enzyme immunoassayrun on a SiemenslowastMedical IMMULITElowast 1000 ImmunoassayAnalyzer (Siemens Medical Solutions Diagnostics)
25 Statistical Analysis The results were expressed asmean plusmnthe standard error of the mean (SEM) for all morphologicaldata sets The data were analyzed by nonparametric testprocedure the Kruskal-Wallis test for overall significanceof group followed by the Mann-Whitney test for individualcomparisons using the Infostat 2008 version A 119875 value ofless than 005 was considered statistically significant
3 Results
AR-ir cells were studied by immunohistochemistry on eachzone of the pituitary PD parenchyma of fetal immatureprepubertal and adult male viscachas The AR-ir cells wereobserved throughout the PD parenchyma and immunohis-tochemical results revealed their presence in the nuclei (ARn-ir) and in the cytoplasm (ARc-ir)
In fetal pituitary from midpregnancy scarce ARn-ir cellsand intensely stained ARc-ir cells were located in the medialregion and extended towards the ventral and dorsal regionsHowever the percentages of ARn-ir cells (caudal end 004 plusmn002 rostral end 005 plusmn 003 central region 055 plusmn 015dorsal region 015 plusmn 007 ventral region 016 plusmn 007)and the percentages of ARc-ir cells did not differ significantly
among different zones of the PD (119875 gt 005) (Figures 2(a)3(A) and 3(B))
In fetal pituitary from late pregnancy ARc-ir cellswere found around blood vessels or among other pituitaryparenchymal cellsThepercentage ofARc-ir cells in the dorsalregion was significantly higher (119875 lt 001) than in the ventralregion and at the rostral and caudal ends (Figures 2(a) 3(C)and 3(D)) ARn-ir cells were few and frequently sphericaland some of them were irregular in shape The percentage ofthese cells did not vary among different PD zones (119875 gt 005caudal end 054 plusmn 023 rostral end 021 plusmn 002 centralregion 025plusmn010 dorsal region 040plusmn010 ventral region028 plusmn 010)
In pituitary PD of immature animals the ARc-ir cellswere located in the medial region and extended towardsthe caudal end The cytoplasmic immunostaining was lessintense than in fetal glands cells Several ARc-ir cells wereoval and their spherical nucleus was located at one end ofthe cytoplasm The percentage of ARc-ir cells did not differsignificantly (119875 gt 005) in different PD zones (Figure 2(a))Intensely stained ARn-ir cells were mainly observed inthe medial region where their percentage was significantlyhigher than in the dorsal region and rostral end (119875 lt 005Figures 2(b) and 3(E)ndash3(H))
In prepubertal viscachas numerous immunopositive AR-ir cells were distributed throughout the PD parenchymaThese cells were arranged around blood vessels at the rostralend and in the medial and dorsal regions In the ventralregion there were some irregular AR-ir cells with a cyto-plasmic prolongation surrounding blood vessels or betweenother parenchymal cells The ARc-ir cells showed significant
International Journal of Endocrinology 5
(A) (B)
(C)
(E) (F) (H)
(I) (J) (K)
(G)
(D)
Figure 3 AR immunohistochemical staining of viscacha pituitaries ((A) (B)) Sections of a fetal pituitary from midpregnancy showing thedifferent zones of PD (A) In the medial region ARn-ir and ARc-ir cells were present (B) Inset ARc-ir cells (c) with a negative nucleus((C) (D)) Images of a fetal pituitary from late pregnancy Isolated ARc-ir cells around blood vessel (D) ((E) (F) (G) and (H)) AR-ir cellsin a pituitary gland of immature viscacha There are cells throughout the PD (E) The ARn-ir cells are increased in the medial region (F)Magnification of ARn-ir cells (G) and a pituitary cell with heterogeneous cytoplasmic staining (H) ((I) (J) and (K)) Pituitary of a prepubertalviscacha ARc-ir cells in groups in contact with blood vessel (J) and others with a cytoplasmic prolongation (K) ARn-ir cells (arrows) ARc-ircells (arrowheads) PD pars distalis PI pars intermedia PN pars nervosa R Rathkersquos pouch ce caudal end re rostral end vr ventral regionmr medial region dr dorsal region blood vessels v ((A) (C) and (I)) Scale bar 500 120583m (E) Scale bar 250 120583m ((B) (D) (F) and (J)) Scalebar 25 120583m Inset of (B) (G) (H) and (K) scale bar 10 120583m Images counterstaining with hematoxylin
6 International Journal of Endocrinology
Table 1 AR expression in pituitary PD of male viscachas
Distribution Immunostaining pattern Morphological characteristicsMP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cellsLP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cells
Immature Isolated or in small groups Homogeneous and heterogeneous cytoplasmsome intensely stained nuclei Cells oval in shape
Prepubertal Isolated or in groups Homogeneous cytoplasmintensely nuclear staining Irregular cells
Adult-Rep P Isolated or in small groups Nuclear immunoreactivityscarce immunostained cytoplasms Oval spherical or irregular cells
Adult-Reg P Isolated Nuclear immunoreactivityscarce immunostained cytoplasms Small and spherical cytoplasm
Adult-Rec P Isolated or in small groups Nuclear immunoreactivityintense cytoplasmic staining
Spherical oval or irregular cytoplasmsand nuclei
short cytoplasmic prolongation
variations the highest value was found in the medial regionin relation to the ventral region and caudal end (119875 lt001) whereas the lowest percentage was found at the caudalend (Figure 2(a)) The immunostaining of ARn-ir cells wasintense in the dorsal region and the highest percentage ofthese cells was found in the medial region in relation to thedorsal and ventral regions and rostral end (119875 lt 005 Figures2(b) and 3(I)ndash3(K))
In PD of adult animals the AR expression mainly hada nuclear localization In the reproductive period severalpositive nuclei were found at the caudal end and dorsalregion as well as on the dorsal and ventral edges of PDThe percentage of ARn-ir cells was significantly higher atthe caudal end in relation to the other PD zones (119875 lt005 Figure 2(b)) A small number of ARc-ir cells werefrequently located in the medial region The cytoplasmicimmunolabeling pattern was homogeneous and sometimesintense In addition some cells presented heterogeneouscytoplasmic staining and positivity on the nuclear periphery(Figures 4(A)ndash4(E)) No significant changes were observedin the percentage of ARc-ir cells between different PD zones(119875 gt 005 Figure 2(a))
In the gonadal regression period AR-ir cells were mainlyfound in the medial region of PD The immunostainedcytoplasms were small with medial or eccentric nucleusThere were no significant differences in ARc-ir cells betweendifferent PD zones (119875 gt 005 Figure 2(a)) whereas ARn-ircells were more numerous in the medial region in relation tothe ventral region and the rostral end (119875 lt 005 Figure 2(b))
In the gonadal recovery period an increase of AR-ir cellsthroughout PD parenchyma was observed In this periodthe cytoplasmic labeling was more intense and homogeneousin relation to the gonadal regression period (Figures 4(F)ndash4(I)) The percentage of ARc-ir cells was not significantlydifferent between the PD zones (119875 gt 005 Figure 2(a)) Thepercentage of ARn-ir cells was higher at the caudal end inrelation to the rostral end and the medial region (119875 lt 001Figure 2(b)) Table 1 shows a summary of themain cytologicalcharacteristics
The morphometric variations of the AR expression inrelation to development and growth in viscachas PD werestudied by comparing the data obtained from fetal pituitaryduring mid- and late pregnancy and from immature prepu-bertal and adult pituitary in the reproductive period Therewas an increase in ARn-ir cells percentage and a significantdecrease in the percentage of ARc-ir cells in immatureanimals as compared to fetus and in adult animals duringthe reproductive period as compared to prepubertal animalsThe percentage of ARc-ir cells increased significantly inprepubertal in relation to immature viscachas Statisticalanalysis is shown in Figure 5
In order to study the seasonal expression of AR the threeperiods of the reproductive cycle were analyzed The totalpercentages of ARn-ir and ARc-ir cells obtained from threeperiods of the reproductive cycle of adult male viscachaswere statistically analyzed (Figure 5)The percentage of ARn-ir cells showed a significant decrease during the gonadalregression period (119875 lt 0001) No variations were observedin the percentage of ARc-ir cells throughout the reproductivecycle (119875 gt 005)
The testosterone serum concentrations of immature ani-mals were below the detection limit (lt020 ngmL) and toolow to reliably reveal differences from the other groupsThe testosterone serum concentrations of prepubertal andadult viscachas in the gonadal regression period were lowerthan those in adult viscachas during their reproductive andgonadal recovery periods (119875 lt 0001 Figure 6)
4 Discussion
The analysis of the AR expression in the viscacha pituitarygland is essential for a better understanding of the regulatorysystems since androgens are involved in the control ofthe reproductive function This study represents the firstdescription of variations in the regionalization and in theimmunostaining pattern of pituitary AR in relation to thedevelopment growth and reproductive cycle of male vis-cachas
International Journal of Endocrinology 7
(A) (B)
(C) (D) (E)
(F) (G)
(H) (I)
Figure 4 Seasonal expression of AR in pituitary during the reproductive cycle of adult viscachas ((A)ndash(E)) Reproductive period ((A) (B))Images of caudal end the AR were expressed mainly in the nuclei of PD cells (arrows) ((C) (D) and (E)) AR expression at the rostral endventral region and dorsal region respectively The arrowhead indicated a cell with cytoplasmic labeling for AR ((F) (G)) A few AR-ir cellsat caudal end during the gonadal regression period ((H) (I)) AR-ir cells at caudal end during the gonadal recovery period Caudal endce blood vessels v follicular structures f ((A) (F) and (H)) Scale bar 100 120583m ((B) (C) (D) (E) (G) and (I)) Scale bar 25 120583m Imagescounterstaining with hematoxylin
Themain findings of this study can be summarized as fol-lows (1) the regionalization and immunostaining pattern ofpituitary AR vary in relation to growth (2) the AR expressionwas mainly cytoplasmic (ARc-ir) in fetuses (3) a significant
increase of ARc-ir cells was observed in prepubertal animals(4) the nuclear AR (ARn-ir) expression increased in relationto development reaching the highest values in adult animals(5) the ARn-ir cells varied seasonally with maximum values
8 International Journal of Endocrinology
0123456789
10
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
ARn-ir cells ()ARc-ir cells ()
Pitu
itary
PD
()
lowastlowastlowastlowastlowastlowast
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 5 Total percentage of ARn-ir cells and ARc-ir cells infetus (from mid- and late pregnancy) immature prepubertaland adult viscachas (during reproductive gonadal regression andgonadal recovery periods) The result revealed statistically signifi-cant changes lowastlowastlowast119875 lt 0001 immature versus LP-FETUS prepu-bertal versus immature Adult-Rep P versus prepubertal Adult-Reg P versus Adult-Rep P Adult-Rec P versus Adult-Reg P
0
1
2
3
4
5
6
7
Prepubertal Adult-Rep P Adult-Reg P Adult-Rec P
Seru
m te
stoste
rone
conc
entr
atio
n (n
gm
L)
lowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 6 Testosterone serum concentrations The values areexpressed as mean plusmn SEM (119899 = 4) lowastlowast119875 lt 001 Adult-Rep Pversus Adult-Reg P lowastlowastlowast119875 lt 0001 prepubertal versus Adult-RepP Adult-Reg P versus Adult-Rec P
during the reproductive period and minimum values duringthe gonadal regression period (6) the amount of ARn-ir cellswas directly related to testosterone serum concentration
There are still many questions about the relations ofAR expression with the pituitary development and hormonesecretion Wu et al [12] did not observe AR in the pituitarygland during late gestation and suggested that the lack of
feedback by way of AR at pituitary levels may have importantconsequences on the fetal pituitary function More recentlyYuan et al [18] have reported in sheep that AR emerges inthe anterior pituitary before day 60 of gestation and thepercentage of the total anterior pituitary cells expressing thereceptor did not change throughout gestation In additionother studies have concluded that the fetal brain and pitu-itary are target sites for circulating androgens or androgenprecursors in fetal plasma and the increase of androgensin hypothalamic action immediately prior to birth might becritical to the timing of parturition [13] Huang and Yuan[42] have reported that AR expression reflects the functionalstatus of androgen in the pituitary of male sheep lambTheseauthors suggest that during sexual maturity the feedback ofsex hormones on the pituitary is at low level and stimulatesgonadotropin secretion
Some reproductive characteristics of female viscachassuch as polyovulation implantation and offspring birth havebeen described in previous studies Gestation lasts approx-imately 154 days an unusually long period for a rodentand it finishes in spring when the survival probabilities areideal for the mother and offspring [38 43 44] The resultsobtained in this work demonstrate AR in fetal pituitary frommidpregnancy indicating that AR are present around days75ndash80 of gestation in the pituitary cells These receptors arepotentially in position to participate in the regulation of thefetal androgen function Furthermore the analysis of thedistribution morphometry and immunostaining pattern ofAR-ir cells in the parenchyma of fetal glands suggests thatthese cells contain variable amounts of AR and they aremorphologically related to blood supply and other endocrinecells of PD parenchyma In late pregnancy of viscacha thepercentages of ARc-ir and ARn-ir cells did not change withrespect to the values found during midpregnancy Theseresults suggested that fetal pituitary AR might have anyspecific function since midgestation in viscacha
Thehypothalamic-pituitary-gonadal axis is activated dur-ing puberty when hormonalmodulation of sexual behavior isorganized [45] During male development the masculiniza-tion is given by the hormones produced by the fetal testesprimarily testosterone [46 47] The gonadal steroids feedback to the central nervous system to exert a key role inthe central control of the reproductive axis by regulatingthe release of the gonadotropin-releasing hormone (GnRH)[48] However the neurons that synthesize GnRH did notexpress AR and there are sex-steroids sensitive neuronsupstream the GnRH cells where the feedback signals ofthe steroid might be processed to the GnRH cells [49] Theneurons of hypothalamic nuclei that expressed kisspeptin(K) and neurokinin B (NKB) may sense circulating levelsof testosterone through the expression of AR [49 50] Inrodents sheep and primates K and NKB have recently beenimplicated in the control of puberty and the maintenanceof the reproductive function in adulthood [51ndash53] In thepituitary the expression of AR has been mainly observedin gonadotrophs [18 54 55] and gonadal steroid hormonesaffect the gonadotroph population in order to regulate theiractivity
International Journal of Endocrinology 9
In immature male viscachas a significant increase ofnuclear AR and an important decrease of cytoplasmic ARwere observed In prepubertal viscachas more nuclei andcytoplasm appear and there is a clear increase of ARc-ir cellsmainly in the medial region suggesting that more cells beginto express AR in preparation for sexual maturation In adultthe immunostaining is mainly nuclear and regionalized atthe caudal end of PD suggesting that AR are active in thePD cells The present results demonstrate variations in theregionalization and immunostaining pattern of AR duringsexual maturity probably in relation to the contribution ofandrogens reaching the different pituitary zones throughblood vessels Thus the nuclear AR expression in pituitaryPD of viscacha varies according to gonadal developmentand testosterone serum concentration Our results are inagreement with those found by Wood and Newman [24]suggesting that a subset of unoccupied receptors is located inthe cell cytoplasm in the absence of ligand
Bibliographic reports referring to seasonal variations ofRA are scarce Lu et al [56] have recently demonstrated astronger expression ofAR in scented glands during the breed-ing season suggesting that androgens may directly influ-ence the glandular function of the muskrats and courtshipbehavior The viscacha is a seasonal breeding rodent andprevious studies have demonstrated that testosterone acts ongonadotrophs whether directly or indirectly regulating theiractivity in the pituitary gland [28] Castrated viscachamodelshave been used to examine the withdrawal effects of circulat-ing androgens on pituitary cells mainly on the gonadotrophsdue to their involvement in the reproductive processes Inaddition castration resulted in a significant decrease in thepercentage of cells expressing AR throughout the PD [37]The present results demonstrate changes in the amount ofpituitary AR-ir cells during the annual reproductive cycleof adult male viscacha which might be due to variations inthe supply of ligands across blood irrigation The maximumvalues of AR expression were found in the reproductiveperiod and the minimum values in the regression period inrelation to androgen concentrationsThus ARmight regulatethe activity of the pituitary cells involved in the reproductivecycle of adult male Lagostomus
In conclusion our results demonstrate thatAR expressionis related to the pituitary histophysiological changes duringthe growth and sexual maturity of male viscacha Moreoverthey are likely to participate in the seasonal reproductioncontrol in adulthoodHowever the precise actionmechanismof AR in the pituitary viscacha needs further study
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Mrs A Bernardi and Mr JArroyuelo for their technical participation This work wassupported by Project 2-2614 Secretaria de Ciencia y TecnicaUniversidad Nacional de San Luis
References
[1] A K Roy Y Lavrovsky C S Song et al ldquoRegulation ofandrogen actionrdquo Vitamins amp Hormones vol 55 pp 309ndash3521999
[2] S Denayer C Helsen L Thorrez A Haelens and F ClaessensldquoThe rules of DNA recognition by the androgen receptorrdquoMolecular Endocrinology vol 24 no 5 pp 898ndash913 2010
[3] E V Jensen G L Greene L E Closs E R DeSombre and MNadji ldquoReceptors reconsidered a 20-year perspectiverdquo RecentProgress in Hormone Research vol 38 pp 1ndash40 1982
[4] B D SchanbacherM P Johnson andD J Tindall ldquoAndrogenicregulation of luteinizing hormone secretion relationship toandrogen binding in sheep pituitaryrdquo Biology of Reproductionvol 36 no 2 pp 340ndash350 1987
[5] R J Handa D L Reid and J A Resko ldquoAndrogen receptorsin brain and pituitary of female rats cyclic changes andcomparisons with the malerdquo Biology of Reproduction vol 34no 2 pp 293ndash303 1986
[6] J V A Choate and J A Resko ldquoEffects of androgen on brain andpituitary androgen receptors and LH secretion of male guineapigsrdquoThe Journal of Steroid Biochemistry andMolecular Biologyvol 59 no 3-4 pp 315ndash322 1996
[7] T J Spady R Shayya V GThackray L Ehrensberger J S Bai-ley and P L Mellon ldquoAndrogen regulates follicle-stimulatinghormone 120573 gene expression in an activin-dependent manner inimmortalized gonadotropesrdquo Molecular Endocrinology vol 18no 4 pp 925ndash940 2004
[8] P B Connolly J V A Choate and J A Resko ldquoEffects ofexogenous androgen on brain androgen receptors of the fetalrhesus monkeyrdquoNeuroendocrinology vol 59 no 3 pp 271ndash2761994
[9] J A Resko A Perkins C E Roselli J A Fitzgerald J V AChoate and F Stormshak ldquoEndocrine correlates of partnerpreference behavior in ramsrdquo Biology of Reproduction vol 55no 1 pp 120ndash126 1996
[10] S E Abdelgadir C E Roselli J V A Choate and J A ReskoldquoAndrogen receptor messenger ribonucleic acid in brains andpituitaries of male Rhesus monkeys studies on distributionhormonal control and relationship to luteinizing hormonesecretionrdquo Biology of Reproduction vol 60 no 5 pp 1251ndash12561999
[11] R W Bonsall H D Rees and R P Michael ldquoThe distributionnuclear uptake and metabolism of [3H]dihydrotestosterone inthe brain pituitary gland and genital tract of the male rhesusmonkeyrdquo Journal of Steroid Biochemistry vol 23 no 4 pp 389ndash398 1985
[12] S S Wu P W Nathanielsz and T J McDonald ldquoImmunocy-tochemical distribution of androgen receptors in the hypotha-lamus and pituitary of the fetal baboon in late gestationrdquoDevelopmental Brain Research vol 84 no 2 pp 278ndash281 1995
[13] C E Wood and M Keller-Wood ldquoOntogeny of androgenreceptor expression in the ovine fetal central nervous systemand pituitaryrdquo Neuroscience Letters vol 439 no 2 pp 153ndash1562008
[14] J V A Choate O D Slayden and J A Resko ldquoImmuno-cytochemical localization of androgen receptors in brains ofdeveloping and adult male rhesus monkeysrdquo Endocrine vol 8no 1 pp 51ndash60 1998
10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
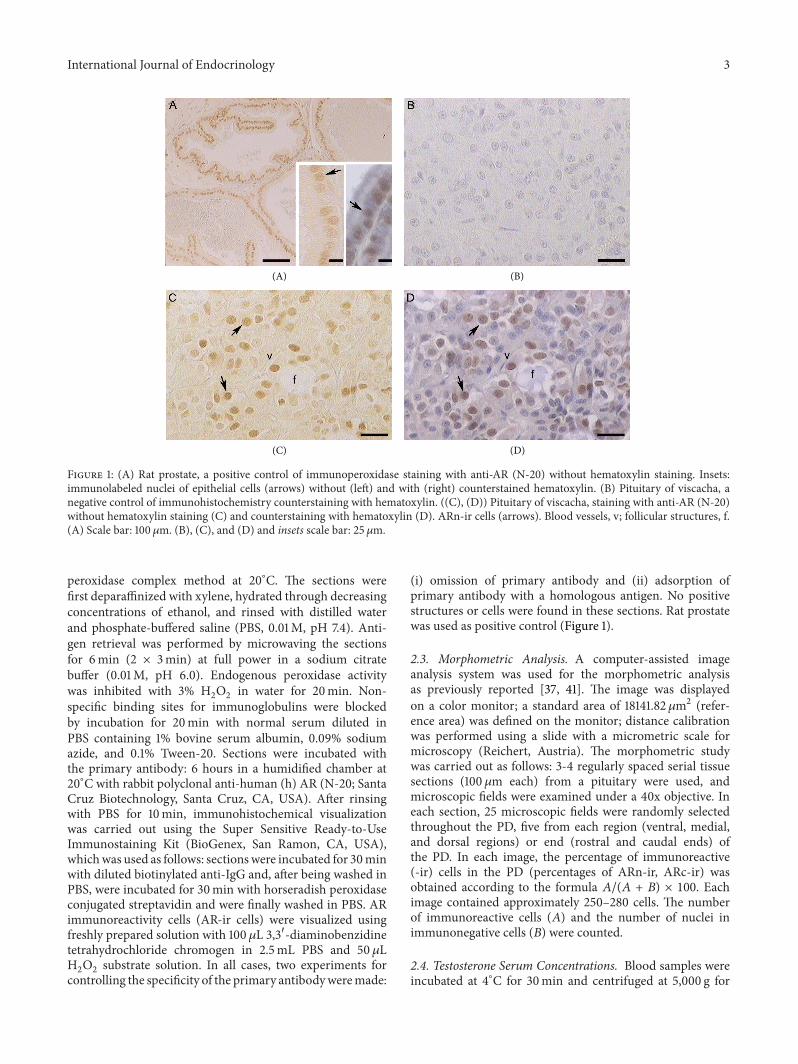
International Journal of Endocrinology 3
(A) (B)
(C) (D)
Figure 1 (A) Rat prostate a positive control of immunoperoxidase staining with anti-AR (N-20) without hematoxylin staining Insetsimmunolabeled nuclei of epithelial cells (arrows) without (left) and with (right) counterstained hematoxylin (B) Pituitary of viscacha anegative control of immunohistochemistry counterstaining with hematoxylin ((C) (D)) Pituitary of viscacha staining with anti-AR (N-20)without hematoxylin staining (C) and counterstaining with hematoxylin (D) ARn-ir cells (arrows) Blood vessels v follicular structures f(A) Scale bar 100120583m (B) (C) and (D) and insets scale bar 25 120583m
peroxidase complex method at 20∘C The sections werefirst deparaffinized with xylene hydrated through decreasingconcentrations of ethanol and rinsed with distilled waterand phosphate-buffered saline (PBS 001M pH 74) Anti-gen retrieval was performed by microwaving the sectionsfor 6min (2 times 3min) at full power in a sodium citratebuffer (001M pH 60) Endogenous peroxidase activitywas inhibited with 3 H
2O2in water for 20min Non-
specific binding sites for immunoglobulins were blockedby incubation for 20min with normal serum diluted inPBS containing 1 bovine serum albumin 009 sodiumazide and 01 Tween-20 Sections were incubated withthe primary antibody 6 hours in a humidified chamber at20∘C with rabbit polyclonal anti-human (h) AR (N-20 SantaCruz Biotechnology Santa Cruz CA USA) After rinsingwith PBS for 10min immunohistochemical visualizationwas carried out using the Super Sensitive Ready-to-UseImmunostaining Kit (BioGenex San Ramon CA USA)whichwas used as follows sectionswere incubated for 30minwith diluted biotinylated anti-IgG and after being washed inPBS were incubated for 30min with horseradish peroxidaseconjugated streptavidin and were finally washed in PBS ARimmunoreactivity cells (AR-ir cells) were visualized usingfreshly prepared solution with 100 120583L 331015840-diaminobenzidinetetrahydrochloride chromogen in 25mL PBS and 50 120583LH2O2substrate solution In all cases two experiments for
controlling the specificity of the primary antibodyweremade
(i) omission of primary antibody and (ii) adsorption ofprimary antibody with a homologous antigen No positivestructures or cells were found in these sections Rat prostatewas used as positive control (Figure 1)
23 Morphometric Analysis A computer-assisted imageanalysis system was used for the morphometric analysisas previously reported [37 41] The image was displayedon a color monitor a standard area of 1814182 120583m2 (refer-ence area) was defined on the monitor distance calibrationwas performed using a slide with a micrometric scale formicroscopy (Reichert Austria) The morphometric studywas carried out as follows 3-4 regularly spaced serial tissuesections (100120583m each) from a pituitary were used andmicroscopic fields were examined under a 40x objective Ineach section 25 microscopic fields were randomly selectedthroughout the PD five from each region (ventral medialand dorsal regions) or end (rostral and caudal ends) ofthe PD In each image the percentage of immunoreactive(-ir) cells in the PD (percentages of ARn-ir ARc-ir) wasobtained according to the formula 119860(119860 + 119861) times 100 Eachimage contained approximately 250ndash280 cells The numberof immunoreactive cells (119860) and the number of nuclei inimmunonegative cells (119861) were counted
24 Testosterone Serum Concentrations Blood samples wereincubated at 4∘C for 30min and centrifuged at 5000 g for
4 International Journal of Endocrinology
005
115
225
335
445
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
Caudal ERostral ECentral R
Dorsal RVentral R
ARc
-ir ce
lls (
)lowastlowast
lowastlowast
(a)
02468
101214161820
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
Caudal ERostral ECentral R
Dorsal RVentral R
ARn
-ir ce
lls (
) lowastlowast
lowast lowast
lowast
lowast
(b)
Figure 2 Morphometry of AR-ir cells in pituitary of male viscacha in different pituitary PD zones (a) ARc-ir cells LP-FETUS lowastlowast119875 lt 001dorsal R versus ventral R rostral E and caudal E Prepubertal lowastlowast119875 lt 001medial R versus ventral R and caudal E Adults the percentageof ARc-ir cells did not reveal statistically significant changes in PD zones (b) ARn-ir cells Immature lowast119875 lt 005medial R versus dorsal Rand rostral E Prepubertal lowast119875 lt 005medial R versus dorsal R and ventral R Adult-Rep P lowast119875 lt 005 caudal E versus the other PD zonesAdult-Reg P lowast119875 lt 005medial R versus ventral R and rostral E Adult-Rec P lowastlowast119875 lt 001 caudal E versus rostral E and medial R Fetusesfrom midpregnancy MP-FETUS fetuses from late pregnancy LP-FETUS reproductive period Rep P gonadal regression period Reg Pgonadal recovery period Rec P region R end E
5min and serum was removed Testosterone serum concen-trations were measured using the total testosterone test aspreviously reported by Chaves et al [35] This test is a solid-phase competitive chemiluminescent enzyme immunoassayrun on a SiemenslowastMedical IMMULITElowast 1000 ImmunoassayAnalyzer (Siemens Medical Solutions Diagnostics)
25 Statistical Analysis The results were expressed asmean plusmnthe standard error of the mean (SEM) for all morphologicaldata sets The data were analyzed by nonparametric testprocedure the Kruskal-Wallis test for overall significanceof group followed by the Mann-Whitney test for individualcomparisons using the Infostat 2008 version A 119875 value ofless than 005 was considered statistically significant
3 Results
AR-ir cells were studied by immunohistochemistry on eachzone of the pituitary PD parenchyma of fetal immatureprepubertal and adult male viscachas The AR-ir cells wereobserved throughout the PD parenchyma and immunohis-tochemical results revealed their presence in the nuclei (ARn-ir) and in the cytoplasm (ARc-ir)
In fetal pituitary from midpregnancy scarce ARn-ir cellsand intensely stained ARc-ir cells were located in the medialregion and extended towards the ventral and dorsal regionsHowever the percentages of ARn-ir cells (caudal end 004 plusmn002 rostral end 005 plusmn 003 central region 055 plusmn 015dorsal region 015 plusmn 007 ventral region 016 plusmn 007)and the percentages of ARc-ir cells did not differ significantly
among different zones of the PD (119875 gt 005) (Figures 2(a)3(A) and 3(B))
In fetal pituitary from late pregnancy ARc-ir cellswere found around blood vessels or among other pituitaryparenchymal cellsThepercentage ofARc-ir cells in the dorsalregion was significantly higher (119875 lt 001) than in the ventralregion and at the rostral and caudal ends (Figures 2(a) 3(C)and 3(D)) ARn-ir cells were few and frequently sphericaland some of them were irregular in shape The percentage ofthese cells did not vary among different PD zones (119875 gt 005caudal end 054 plusmn 023 rostral end 021 plusmn 002 centralregion 025plusmn010 dorsal region 040plusmn010 ventral region028 plusmn 010)
In pituitary PD of immature animals the ARc-ir cellswere located in the medial region and extended towardsthe caudal end The cytoplasmic immunostaining was lessintense than in fetal glands cells Several ARc-ir cells wereoval and their spherical nucleus was located at one end ofthe cytoplasm The percentage of ARc-ir cells did not differsignificantly (119875 gt 005) in different PD zones (Figure 2(a))Intensely stained ARn-ir cells were mainly observed inthe medial region where their percentage was significantlyhigher than in the dorsal region and rostral end (119875 lt 005Figures 2(b) and 3(E)ndash3(H))
In prepubertal viscachas numerous immunopositive AR-ir cells were distributed throughout the PD parenchymaThese cells were arranged around blood vessels at the rostralend and in the medial and dorsal regions In the ventralregion there were some irregular AR-ir cells with a cyto-plasmic prolongation surrounding blood vessels or betweenother parenchymal cells The ARc-ir cells showed significant
International Journal of Endocrinology 5
(A) (B)
(C)
(E) (F) (H)
(I) (J) (K)
(G)
(D)
Figure 3 AR immunohistochemical staining of viscacha pituitaries ((A) (B)) Sections of a fetal pituitary from midpregnancy showing thedifferent zones of PD (A) In the medial region ARn-ir and ARc-ir cells were present (B) Inset ARc-ir cells (c) with a negative nucleus((C) (D)) Images of a fetal pituitary from late pregnancy Isolated ARc-ir cells around blood vessel (D) ((E) (F) (G) and (H)) AR-ir cellsin a pituitary gland of immature viscacha There are cells throughout the PD (E) The ARn-ir cells are increased in the medial region (F)Magnification of ARn-ir cells (G) and a pituitary cell with heterogeneous cytoplasmic staining (H) ((I) (J) and (K)) Pituitary of a prepubertalviscacha ARc-ir cells in groups in contact with blood vessel (J) and others with a cytoplasmic prolongation (K) ARn-ir cells (arrows) ARc-ircells (arrowheads) PD pars distalis PI pars intermedia PN pars nervosa R Rathkersquos pouch ce caudal end re rostral end vr ventral regionmr medial region dr dorsal region blood vessels v ((A) (C) and (I)) Scale bar 500 120583m (E) Scale bar 250 120583m ((B) (D) (F) and (J)) Scalebar 25 120583m Inset of (B) (G) (H) and (K) scale bar 10 120583m Images counterstaining with hematoxylin
6 International Journal of Endocrinology
Table 1 AR expression in pituitary PD of male viscachas
Distribution Immunostaining pattern Morphological characteristicsMP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cellsLP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cells
Immature Isolated or in small groups Homogeneous and heterogeneous cytoplasmsome intensely stained nuclei Cells oval in shape
Prepubertal Isolated or in groups Homogeneous cytoplasmintensely nuclear staining Irregular cells
Adult-Rep P Isolated or in small groups Nuclear immunoreactivityscarce immunostained cytoplasms Oval spherical or irregular cells
Adult-Reg P Isolated Nuclear immunoreactivityscarce immunostained cytoplasms Small and spherical cytoplasm
Adult-Rec P Isolated or in small groups Nuclear immunoreactivityintense cytoplasmic staining
Spherical oval or irregular cytoplasmsand nuclei
short cytoplasmic prolongation
variations the highest value was found in the medial regionin relation to the ventral region and caudal end (119875 lt001) whereas the lowest percentage was found at the caudalend (Figure 2(a)) The immunostaining of ARn-ir cells wasintense in the dorsal region and the highest percentage ofthese cells was found in the medial region in relation to thedorsal and ventral regions and rostral end (119875 lt 005 Figures2(b) and 3(I)ndash3(K))
In PD of adult animals the AR expression mainly hada nuclear localization In the reproductive period severalpositive nuclei were found at the caudal end and dorsalregion as well as on the dorsal and ventral edges of PDThe percentage of ARn-ir cells was significantly higher atthe caudal end in relation to the other PD zones (119875 lt005 Figure 2(b)) A small number of ARc-ir cells werefrequently located in the medial region The cytoplasmicimmunolabeling pattern was homogeneous and sometimesintense In addition some cells presented heterogeneouscytoplasmic staining and positivity on the nuclear periphery(Figures 4(A)ndash4(E)) No significant changes were observedin the percentage of ARc-ir cells between different PD zones(119875 gt 005 Figure 2(a))
In the gonadal regression period AR-ir cells were mainlyfound in the medial region of PD The immunostainedcytoplasms were small with medial or eccentric nucleusThere were no significant differences in ARc-ir cells betweendifferent PD zones (119875 gt 005 Figure 2(a)) whereas ARn-ircells were more numerous in the medial region in relation tothe ventral region and the rostral end (119875 lt 005 Figure 2(b))
In the gonadal recovery period an increase of AR-ir cellsthroughout PD parenchyma was observed In this periodthe cytoplasmic labeling was more intense and homogeneousin relation to the gonadal regression period (Figures 4(F)ndash4(I)) The percentage of ARc-ir cells was not significantlydifferent between the PD zones (119875 gt 005 Figure 2(a)) Thepercentage of ARn-ir cells was higher at the caudal end inrelation to the rostral end and the medial region (119875 lt 001Figure 2(b)) Table 1 shows a summary of themain cytologicalcharacteristics
The morphometric variations of the AR expression inrelation to development and growth in viscachas PD werestudied by comparing the data obtained from fetal pituitaryduring mid- and late pregnancy and from immature prepu-bertal and adult pituitary in the reproductive period Therewas an increase in ARn-ir cells percentage and a significantdecrease in the percentage of ARc-ir cells in immatureanimals as compared to fetus and in adult animals duringthe reproductive period as compared to prepubertal animalsThe percentage of ARc-ir cells increased significantly inprepubertal in relation to immature viscachas Statisticalanalysis is shown in Figure 5
In order to study the seasonal expression of AR the threeperiods of the reproductive cycle were analyzed The totalpercentages of ARn-ir and ARc-ir cells obtained from threeperiods of the reproductive cycle of adult male viscachaswere statistically analyzed (Figure 5)The percentage of ARn-ir cells showed a significant decrease during the gonadalregression period (119875 lt 0001) No variations were observedin the percentage of ARc-ir cells throughout the reproductivecycle (119875 gt 005)
The testosterone serum concentrations of immature ani-mals were below the detection limit (lt020 ngmL) and toolow to reliably reveal differences from the other groupsThe testosterone serum concentrations of prepubertal andadult viscachas in the gonadal regression period were lowerthan those in adult viscachas during their reproductive andgonadal recovery periods (119875 lt 0001 Figure 6)
4 Discussion
The analysis of the AR expression in the viscacha pituitarygland is essential for a better understanding of the regulatorysystems since androgens are involved in the control ofthe reproductive function This study represents the firstdescription of variations in the regionalization and in theimmunostaining pattern of pituitary AR in relation to thedevelopment growth and reproductive cycle of male vis-cachas
International Journal of Endocrinology 7
(A) (B)
(C) (D) (E)
(F) (G)
(H) (I)
Figure 4 Seasonal expression of AR in pituitary during the reproductive cycle of adult viscachas ((A)ndash(E)) Reproductive period ((A) (B))Images of caudal end the AR were expressed mainly in the nuclei of PD cells (arrows) ((C) (D) and (E)) AR expression at the rostral endventral region and dorsal region respectively The arrowhead indicated a cell with cytoplasmic labeling for AR ((F) (G)) A few AR-ir cellsat caudal end during the gonadal regression period ((H) (I)) AR-ir cells at caudal end during the gonadal recovery period Caudal endce blood vessels v follicular structures f ((A) (F) and (H)) Scale bar 100 120583m ((B) (C) (D) (E) (G) and (I)) Scale bar 25 120583m Imagescounterstaining with hematoxylin
Themain findings of this study can be summarized as fol-lows (1) the regionalization and immunostaining pattern ofpituitary AR vary in relation to growth (2) the AR expressionwas mainly cytoplasmic (ARc-ir) in fetuses (3) a significant
increase of ARc-ir cells was observed in prepubertal animals(4) the nuclear AR (ARn-ir) expression increased in relationto development reaching the highest values in adult animals(5) the ARn-ir cells varied seasonally with maximum values
8 International Journal of Endocrinology
0123456789
10
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
ARn-ir cells ()ARc-ir cells ()
Pitu
itary
PD
()
lowastlowastlowastlowastlowastlowast
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 5 Total percentage of ARn-ir cells and ARc-ir cells infetus (from mid- and late pregnancy) immature prepubertaland adult viscachas (during reproductive gonadal regression andgonadal recovery periods) The result revealed statistically signifi-cant changes lowastlowastlowast119875 lt 0001 immature versus LP-FETUS prepu-bertal versus immature Adult-Rep P versus prepubertal Adult-Reg P versus Adult-Rep P Adult-Rec P versus Adult-Reg P
0
1
2
3
4
5
6
7
Prepubertal Adult-Rep P Adult-Reg P Adult-Rec P
Seru
m te
stoste
rone
conc
entr
atio
n (n
gm
L)
lowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 6 Testosterone serum concentrations The values areexpressed as mean plusmn SEM (119899 = 4) lowastlowast119875 lt 001 Adult-Rep Pversus Adult-Reg P lowastlowastlowast119875 lt 0001 prepubertal versus Adult-RepP Adult-Reg P versus Adult-Rec P
during the reproductive period and minimum values duringthe gonadal regression period (6) the amount of ARn-ir cellswas directly related to testosterone serum concentration
There are still many questions about the relations ofAR expression with the pituitary development and hormonesecretion Wu et al [12] did not observe AR in the pituitarygland during late gestation and suggested that the lack of
feedback by way of AR at pituitary levels may have importantconsequences on the fetal pituitary function More recentlyYuan et al [18] have reported in sheep that AR emerges inthe anterior pituitary before day 60 of gestation and thepercentage of the total anterior pituitary cells expressing thereceptor did not change throughout gestation In additionother studies have concluded that the fetal brain and pitu-itary are target sites for circulating androgens or androgenprecursors in fetal plasma and the increase of androgensin hypothalamic action immediately prior to birth might becritical to the timing of parturition [13] Huang and Yuan[42] have reported that AR expression reflects the functionalstatus of androgen in the pituitary of male sheep lambTheseauthors suggest that during sexual maturity the feedback ofsex hormones on the pituitary is at low level and stimulatesgonadotropin secretion
Some reproductive characteristics of female viscachassuch as polyovulation implantation and offspring birth havebeen described in previous studies Gestation lasts approx-imately 154 days an unusually long period for a rodentand it finishes in spring when the survival probabilities areideal for the mother and offspring [38 43 44] The resultsobtained in this work demonstrate AR in fetal pituitary frommidpregnancy indicating that AR are present around days75ndash80 of gestation in the pituitary cells These receptors arepotentially in position to participate in the regulation of thefetal androgen function Furthermore the analysis of thedistribution morphometry and immunostaining pattern ofAR-ir cells in the parenchyma of fetal glands suggests thatthese cells contain variable amounts of AR and they aremorphologically related to blood supply and other endocrinecells of PD parenchyma In late pregnancy of viscacha thepercentages of ARc-ir and ARn-ir cells did not change withrespect to the values found during midpregnancy Theseresults suggested that fetal pituitary AR might have anyspecific function since midgestation in viscacha
Thehypothalamic-pituitary-gonadal axis is activated dur-ing puberty when hormonalmodulation of sexual behavior isorganized [45] During male development the masculiniza-tion is given by the hormones produced by the fetal testesprimarily testosterone [46 47] The gonadal steroids feedback to the central nervous system to exert a key role inthe central control of the reproductive axis by regulatingthe release of the gonadotropin-releasing hormone (GnRH)[48] However the neurons that synthesize GnRH did notexpress AR and there are sex-steroids sensitive neuronsupstream the GnRH cells where the feedback signals ofthe steroid might be processed to the GnRH cells [49] Theneurons of hypothalamic nuclei that expressed kisspeptin(K) and neurokinin B (NKB) may sense circulating levelsof testosterone through the expression of AR [49 50] Inrodents sheep and primates K and NKB have recently beenimplicated in the control of puberty and the maintenanceof the reproductive function in adulthood [51ndash53] In thepituitary the expression of AR has been mainly observedin gonadotrophs [18 54 55] and gonadal steroid hormonesaffect the gonadotroph population in order to regulate theiractivity
International Journal of Endocrinology 9
In immature male viscachas a significant increase ofnuclear AR and an important decrease of cytoplasmic ARwere observed In prepubertal viscachas more nuclei andcytoplasm appear and there is a clear increase of ARc-ir cellsmainly in the medial region suggesting that more cells beginto express AR in preparation for sexual maturation In adultthe immunostaining is mainly nuclear and regionalized atthe caudal end of PD suggesting that AR are active in thePD cells The present results demonstrate variations in theregionalization and immunostaining pattern of AR duringsexual maturity probably in relation to the contribution ofandrogens reaching the different pituitary zones throughblood vessels Thus the nuclear AR expression in pituitaryPD of viscacha varies according to gonadal developmentand testosterone serum concentration Our results are inagreement with those found by Wood and Newman [24]suggesting that a subset of unoccupied receptors is located inthe cell cytoplasm in the absence of ligand
Bibliographic reports referring to seasonal variations ofRA are scarce Lu et al [56] have recently demonstrated astronger expression ofAR in scented glands during the breed-ing season suggesting that androgens may directly influ-ence the glandular function of the muskrats and courtshipbehavior The viscacha is a seasonal breeding rodent andprevious studies have demonstrated that testosterone acts ongonadotrophs whether directly or indirectly regulating theiractivity in the pituitary gland [28] Castrated viscachamodelshave been used to examine the withdrawal effects of circulat-ing androgens on pituitary cells mainly on the gonadotrophsdue to their involvement in the reproductive processes Inaddition castration resulted in a significant decrease in thepercentage of cells expressing AR throughout the PD [37]The present results demonstrate changes in the amount ofpituitary AR-ir cells during the annual reproductive cycleof adult male viscacha which might be due to variations inthe supply of ligands across blood irrigation The maximumvalues of AR expression were found in the reproductiveperiod and the minimum values in the regression period inrelation to androgen concentrationsThus ARmight regulatethe activity of the pituitary cells involved in the reproductivecycle of adult male Lagostomus
In conclusion our results demonstrate thatAR expressionis related to the pituitary histophysiological changes duringthe growth and sexual maturity of male viscacha Moreoverthey are likely to participate in the seasonal reproductioncontrol in adulthoodHowever the precise actionmechanismof AR in the pituitary viscacha needs further study
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Mrs A Bernardi and Mr JArroyuelo for their technical participation This work wassupported by Project 2-2614 Secretaria de Ciencia y TecnicaUniversidad Nacional de San Luis
References
[1] A K Roy Y Lavrovsky C S Song et al ldquoRegulation ofandrogen actionrdquo Vitamins amp Hormones vol 55 pp 309ndash3521999
[2] S Denayer C Helsen L Thorrez A Haelens and F ClaessensldquoThe rules of DNA recognition by the androgen receptorrdquoMolecular Endocrinology vol 24 no 5 pp 898ndash913 2010
[3] E V Jensen G L Greene L E Closs E R DeSombre and MNadji ldquoReceptors reconsidered a 20-year perspectiverdquo RecentProgress in Hormone Research vol 38 pp 1ndash40 1982
[4] B D SchanbacherM P Johnson andD J Tindall ldquoAndrogenicregulation of luteinizing hormone secretion relationship toandrogen binding in sheep pituitaryrdquo Biology of Reproductionvol 36 no 2 pp 340ndash350 1987
[5] R J Handa D L Reid and J A Resko ldquoAndrogen receptorsin brain and pituitary of female rats cyclic changes andcomparisons with the malerdquo Biology of Reproduction vol 34no 2 pp 293ndash303 1986
[6] J V A Choate and J A Resko ldquoEffects of androgen on brain andpituitary androgen receptors and LH secretion of male guineapigsrdquoThe Journal of Steroid Biochemistry andMolecular Biologyvol 59 no 3-4 pp 315ndash322 1996
[7] T J Spady R Shayya V GThackray L Ehrensberger J S Bai-ley and P L Mellon ldquoAndrogen regulates follicle-stimulatinghormone 120573 gene expression in an activin-dependent manner inimmortalized gonadotropesrdquo Molecular Endocrinology vol 18no 4 pp 925ndash940 2004
[8] P B Connolly J V A Choate and J A Resko ldquoEffects ofexogenous androgen on brain androgen receptors of the fetalrhesus monkeyrdquoNeuroendocrinology vol 59 no 3 pp 271ndash2761994
[9] J A Resko A Perkins C E Roselli J A Fitzgerald J V AChoate and F Stormshak ldquoEndocrine correlates of partnerpreference behavior in ramsrdquo Biology of Reproduction vol 55no 1 pp 120ndash126 1996
[10] S E Abdelgadir C E Roselli J V A Choate and J A ReskoldquoAndrogen receptor messenger ribonucleic acid in brains andpituitaries of male Rhesus monkeys studies on distributionhormonal control and relationship to luteinizing hormonesecretionrdquo Biology of Reproduction vol 60 no 5 pp 1251ndash12561999
[11] R W Bonsall H D Rees and R P Michael ldquoThe distributionnuclear uptake and metabolism of [3H]dihydrotestosterone inthe brain pituitary gland and genital tract of the male rhesusmonkeyrdquo Journal of Steroid Biochemistry vol 23 no 4 pp 389ndash398 1985
[12] S S Wu P W Nathanielsz and T J McDonald ldquoImmunocy-tochemical distribution of androgen receptors in the hypotha-lamus and pituitary of the fetal baboon in late gestationrdquoDevelopmental Brain Research vol 84 no 2 pp 278ndash281 1995
[13] C E Wood and M Keller-Wood ldquoOntogeny of androgenreceptor expression in the ovine fetal central nervous systemand pituitaryrdquo Neuroscience Letters vol 439 no 2 pp 153ndash1562008
[14] J V A Choate O D Slayden and J A Resko ldquoImmuno-cytochemical localization of androgen receptors in brains ofdeveloping and adult male rhesus monkeysrdquo Endocrine vol 8no 1 pp 51ndash60 1998
10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 International Journal of Endocrinology
005
115
225
335
445
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
Caudal ERostral ECentral R
Dorsal RVentral R
ARc
-ir ce
lls (
)lowastlowast
lowastlowast
(a)
02468
101214161820
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
Caudal ERostral ECentral R
Dorsal RVentral R
ARn
-ir ce
lls (
) lowastlowast
lowast lowast
lowast
lowast
(b)
Figure 2 Morphometry of AR-ir cells in pituitary of male viscacha in different pituitary PD zones (a) ARc-ir cells LP-FETUS lowastlowast119875 lt 001dorsal R versus ventral R rostral E and caudal E Prepubertal lowastlowast119875 lt 001medial R versus ventral R and caudal E Adults the percentageof ARc-ir cells did not reveal statistically significant changes in PD zones (b) ARn-ir cells Immature lowast119875 lt 005medial R versus dorsal Rand rostral E Prepubertal lowast119875 lt 005medial R versus dorsal R and ventral R Adult-Rep P lowast119875 lt 005 caudal E versus the other PD zonesAdult-Reg P lowast119875 lt 005medial R versus ventral R and rostral E Adult-Rec P lowastlowast119875 lt 001 caudal E versus rostral E and medial R Fetusesfrom midpregnancy MP-FETUS fetuses from late pregnancy LP-FETUS reproductive period Rep P gonadal regression period Reg Pgonadal recovery period Rec P region R end E
5min and serum was removed Testosterone serum concen-trations were measured using the total testosterone test aspreviously reported by Chaves et al [35] This test is a solid-phase competitive chemiluminescent enzyme immunoassayrun on a SiemenslowastMedical IMMULITElowast 1000 ImmunoassayAnalyzer (Siemens Medical Solutions Diagnostics)
25 Statistical Analysis The results were expressed asmean plusmnthe standard error of the mean (SEM) for all morphologicaldata sets The data were analyzed by nonparametric testprocedure the Kruskal-Wallis test for overall significanceof group followed by the Mann-Whitney test for individualcomparisons using the Infostat 2008 version A 119875 value ofless than 005 was considered statistically significant
3 Results
AR-ir cells were studied by immunohistochemistry on eachzone of the pituitary PD parenchyma of fetal immatureprepubertal and adult male viscachas The AR-ir cells wereobserved throughout the PD parenchyma and immunohis-tochemical results revealed their presence in the nuclei (ARn-ir) and in the cytoplasm (ARc-ir)
In fetal pituitary from midpregnancy scarce ARn-ir cellsand intensely stained ARc-ir cells were located in the medialregion and extended towards the ventral and dorsal regionsHowever the percentages of ARn-ir cells (caudal end 004 plusmn002 rostral end 005 plusmn 003 central region 055 plusmn 015dorsal region 015 plusmn 007 ventral region 016 plusmn 007)and the percentages of ARc-ir cells did not differ significantly
among different zones of the PD (119875 gt 005) (Figures 2(a)3(A) and 3(B))
In fetal pituitary from late pregnancy ARc-ir cellswere found around blood vessels or among other pituitaryparenchymal cellsThepercentage ofARc-ir cells in the dorsalregion was significantly higher (119875 lt 001) than in the ventralregion and at the rostral and caudal ends (Figures 2(a) 3(C)and 3(D)) ARn-ir cells were few and frequently sphericaland some of them were irregular in shape The percentage ofthese cells did not vary among different PD zones (119875 gt 005caudal end 054 plusmn 023 rostral end 021 plusmn 002 centralregion 025plusmn010 dorsal region 040plusmn010 ventral region028 plusmn 010)
In pituitary PD of immature animals the ARc-ir cellswere located in the medial region and extended towardsthe caudal end The cytoplasmic immunostaining was lessintense than in fetal glands cells Several ARc-ir cells wereoval and their spherical nucleus was located at one end ofthe cytoplasm The percentage of ARc-ir cells did not differsignificantly (119875 gt 005) in different PD zones (Figure 2(a))Intensely stained ARn-ir cells were mainly observed inthe medial region where their percentage was significantlyhigher than in the dorsal region and rostral end (119875 lt 005Figures 2(b) and 3(E)ndash3(H))
In prepubertal viscachas numerous immunopositive AR-ir cells were distributed throughout the PD parenchymaThese cells were arranged around blood vessels at the rostralend and in the medial and dorsal regions In the ventralregion there were some irregular AR-ir cells with a cyto-plasmic prolongation surrounding blood vessels or betweenother parenchymal cells The ARc-ir cells showed significant
International Journal of Endocrinology 5
(A) (B)
(C)
(E) (F) (H)
(I) (J) (K)
(G)
(D)
Figure 3 AR immunohistochemical staining of viscacha pituitaries ((A) (B)) Sections of a fetal pituitary from midpregnancy showing thedifferent zones of PD (A) In the medial region ARn-ir and ARc-ir cells were present (B) Inset ARc-ir cells (c) with a negative nucleus((C) (D)) Images of a fetal pituitary from late pregnancy Isolated ARc-ir cells around blood vessel (D) ((E) (F) (G) and (H)) AR-ir cellsin a pituitary gland of immature viscacha There are cells throughout the PD (E) The ARn-ir cells are increased in the medial region (F)Magnification of ARn-ir cells (G) and a pituitary cell with heterogeneous cytoplasmic staining (H) ((I) (J) and (K)) Pituitary of a prepubertalviscacha ARc-ir cells in groups in contact with blood vessel (J) and others with a cytoplasmic prolongation (K) ARn-ir cells (arrows) ARc-ircells (arrowheads) PD pars distalis PI pars intermedia PN pars nervosa R Rathkersquos pouch ce caudal end re rostral end vr ventral regionmr medial region dr dorsal region blood vessels v ((A) (C) and (I)) Scale bar 500 120583m (E) Scale bar 250 120583m ((B) (D) (F) and (J)) Scalebar 25 120583m Inset of (B) (G) (H) and (K) scale bar 10 120583m Images counterstaining with hematoxylin
6 International Journal of Endocrinology
Table 1 AR expression in pituitary PD of male viscachas
Distribution Immunostaining pattern Morphological characteristicsMP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cellsLP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cells
Immature Isolated or in small groups Homogeneous and heterogeneous cytoplasmsome intensely stained nuclei Cells oval in shape
Prepubertal Isolated or in groups Homogeneous cytoplasmintensely nuclear staining Irregular cells
Adult-Rep P Isolated or in small groups Nuclear immunoreactivityscarce immunostained cytoplasms Oval spherical or irregular cells
Adult-Reg P Isolated Nuclear immunoreactivityscarce immunostained cytoplasms Small and spherical cytoplasm
Adult-Rec P Isolated or in small groups Nuclear immunoreactivityintense cytoplasmic staining
Spherical oval or irregular cytoplasmsand nuclei
short cytoplasmic prolongation
variations the highest value was found in the medial regionin relation to the ventral region and caudal end (119875 lt001) whereas the lowest percentage was found at the caudalend (Figure 2(a)) The immunostaining of ARn-ir cells wasintense in the dorsal region and the highest percentage ofthese cells was found in the medial region in relation to thedorsal and ventral regions and rostral end (119875 lt 005 Figures2(b) and 3(I)ndash3(K))
In PD of adult animals the AR expression mainly hada nuclear localization In the reproductive period severalpositive nuclei were found at the caudal end and dorsalregion as well as on the dorsal and ventral edges of PDThe percentage of ARn-ir cells was significantly higher atthe caudal end in relation to the other PD zones (119875 lt005 Figure 2(b)) A small number of ARc-ir cells werefrequently located in the medial region The cytoplasmicimmunolabeling pattern was homogeneous and sometimesintense In addition some cells presented heterogeneouscytoplasmic staining and positivity on the nuclear periphery(Figures 4(A)ndash4(E)) No significant changes were observedin the percentage of ARc-ir cells between different PD zones(119875 gt 005 Figure 2(a))
In the gonadal regression period AR-ir cells were mainlyfound in the medial region of PD The immunostainedcytoplasms were small with medial or eccentric nucleusThere were no significant differences in ARc-ir cells betweendifferent PD zones (119875 gt 005 Figure 2(a)) whereas ARn-ircells were more numerous in the medial region in relation tothe ventral region and the rostral end (119875 lt 005 Figure 2(b))
In the gonadal recovery period an increase of AR-ir cellsthroughout PD parenchyma was observed In this periodthe cytoplasmic labeling was more intense and homogeneousin relation to the gonadal regression period (Figures 4(F)ndash4(I)) The percentage of ARc-ir cells was not significantlydifferent between the PD zones (119875 gt 005 Figure 2(a)) Thepercentage of ARn-ir cells was higher at the caudal end inrelation to the rostral end and the medial region (119875 lt 001Figure 2(b)) Table 1 shows a summary of themain cytologicalcharacteristics
The morphometric variations of the AR expression inrelation to development and growth in viscachas PD werestudied by comparing the data obtained from fetal pituitaryduring mid- and late pregnancy and from immature prepu-bertal and adult pituitary in the reproductive period Therewas an increase in ARn-ir cells percentage and a significantdecrease in the percentage of ARc-ir cells in immatureanimals as compared to fetus and in adult animals duringthe reproductive period as compared to prepubertal animalsThe percentage of ARc-ir cells increased significantly inprepubertal in relation to immature viscachas Statisticalanalysis is shown in Figure 5
In order to study the seasonal expression of AR the threeperiods of the reproductive cycle were analyzed The totalpercentages of ARn-ir and ARc-ir cells obtained from threeperiods of the reproductive cycle of adult male viscachaswere statistically analyzed (Figure 5)The percentage of ARn-ir cells showed a significant decrease during the gonadalregression period (119875 lt 0001) No variations were observedin the percentage of ARc-ir cells throughout the reproductivecycle (119875 gt 005)
The testosterone serum concentrations of immature ani-mals were below the detection limit (lt020 ngmL) and toolow to reliably reveal differences from the other groupsThe testosterone serum concentrations of prepubertal andadult viscachas in the gonadal regression period were lowerthan those in adult viscachas during their reproductive andgonadal recovery periods (119875 lt 0001 Figure 6)
4 Discussion
The analysis of the AR expression in the viscacha pituitarygland is essential for a better understanding of the regulatorysystems since androgens are involved in the control ofthe reproductive function This study represents the firstdescription of variations in the regionalization and in theimmunostaining pattern of pituitary AR in relation to thedevelopment growth and reproductive cycle of male vis-cachas
International Journal of Endocrinology 7
(A) (B)
(C) (D) (E)
(F) (G)
(H) (I)
Figure 4 Seasonal expression of AR in pituitary during the reproductive cycle of adult viscachas ((A)ndash(E)) Reproductive period ((A) (B))Images of caudal end the AR were expressed mainly in the nuclei of PD cells (arrows) ((C) (D) and (E)) AR expression at the rostral endventral region and dorsal region respectively The arrowhead indicated a cell with cytoplasmic labeling for AR ((F) (G)) A few AR-ir cellsat caudal end during the gonadal regression period ((H) (I)) AR-ir cells at caudal end during the gonadal recovery period Caudal endce blood vessels v follicular structures f ((A) (F) and (H)) Scale bar 100 120583m ((B) (C) (D) (E) (G) and (I)) Scale bar 25 120583m Imagescounterstaining with hematoxylin
Themain findings of this study can be summarized as fol-lows (1) the regionalization and immunostaining pattern ofpituitary AR vary in relation to growth (2) the AR expressionwas mainly cytoplasmic (ARc-ir) in fetuses (3) a significant
increase of ARc-ir cells was observed in prepubertal animals(4) the nuclear AR (ARn-ir) expression increased in relationto development reaching the highest values in adult animals(5) the ARn-ir cells varied seasonally with maximum values
8 International Journal of Endocrinology
0123456789
10
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
ARn-ir cells ()ARc-ir cells ()
Pitu
itary
PD
()
lowastlowastlowastlowastlowastlowast
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 5 Total percentage of ARn-ir cells and ARc-ir cells infetus (from mid- and late pregnancy) immature prepubertaland adult viscachas (during reproductive gonadal regression andgonadal recovery periods) The result revealed statistically signifi-cant changes lowastlowastlowast119875 lt 0001 immature versus LP-FETUS prepu-bertal versus immature Adult-Rep P versus prepubertal Adult-Reg P versus Adult-Rep P Adult-Rec P versus Adult-Reg P
0
1
2
3
4
5
6
7
Prepubertal Adult-Rep P Adult-Reg P Adult-Rec P
Seru
m te
stoste
rone
conc
entr
atio
n (n
gm
L)
lowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 6 Testosterone serum concentrations The values areexpressed as mean plusmn SEM (119899 = 4) lowastlowast119875 lt 001 Adult-Rep Pversus Adult-Reg P lowastlowastlowast119875 lt 0001 prepubertal versus Adult-RepP Adult-Reg P versus Adult-Rec P
during the reproductive period and minimum values duringthe gonadal regression period (6) the amount of ARn-ir cellswas directly related to testosterone serum concentration
There are still many questions about the relations ofAR expression with the pituitary development and hormonesecretion Wu et al [12] did not observe AR in the pituitarygland during late gestation and suggested that the lack of
feedback by way of AR at pituitary levels may have importantconsequences on the fetal pituitary function More recentlyYuan et al [18] have reported in sheep that AR emerges inthe anterior pituitary before day 60 of gestation and thepercentage of the total anterior pituitary cells expressing thereceptor did not change throughout gestation In additionother studies have concluded that the fetal brain and pitu-itary are target sites for circulating androgens or androgenprecursors in fetal plasma and the increase of androgensin hypothalamic action immediately prior to birth might becritical to the timing of parturition [13] Huang and Yuan[42] have reported that AR expression reflects the functionalstatus of androgen in the pituitary of male sheep lambTheseauthors suggest that during sexual maturity the feedback ofsex hormones on the pituitary is at low level and stimulatesgonadotropin secretion
Some reproductive characteristics of female viscachassuch as polyovulation implantation and offspring birth havebeen described in previous studies Gestation lasts approx-imately 154 days an unusually long period for a rodentand it finishes in spring when the survival probabilities areideal for the mother and offspring [38 43 44] The resultsobtained in this work demonstrate AR in fetal pituitary frommidpregnancy indicating that AR are present around days75ndash80 of gestation in the pituitary cells These receptors arepotentially in position to participate in the regulation of thefetal androgen function Furthermore the analysis of thedistribution morphometry and immunostaining pattern ofAR-ir cells in the parenchyma of fetal glands suggests thatthese cells contain variable amounts of AR and they aremorphologically related to blood supply and other endocrinecells of PD parenchyma In late pregnancy of viscacha thepercentages of ARc-ir and ARn-ir cells did not change withrespect to the values found during midpregnancy Theseresults suggested that fetal pituitary AR might have anyspecific function since midgestation in viscacha
Thehypothalamic-pituitary-gonadal axis is activated dur-ing puberty when hormonalmodulation of sexual behavior isorganized [45] During male development the masculiniza-tion is given by the hormones produced by the fetal testesprimarily testosterone [46 47] The gonadal steroids feedback to the central nervous system to exert a key role inthe central control of the reproductive axis by regulatingthe release of the gonadotropin-releasing hormone (GnRH)[48] However the neurons that synthesize GnRH did notexpress AR and there are sex-steroids sensitive neuronsupstream the GnRH cells where the feedback signals ofthe steroid might be processed to the GnRH cells [49] Theneurons of hypothalamic nuclei that expressed kisspeptin(K) and neurokinin B (NKB) may sense circulating levelsof testosterone through the expression of AR [49 50] Inrodents sheep and primates K and NKB have recently beenimplicated in the control of puberty and the maintenanceof the reproductive function in adulthood [51ndash53] In thepituitary the expression of AR has been mainly observedin gonadotrophs [18 54 55] and gonadal steroid hormonesaffect the gonadotroph population in order to regulate theiractivity
International Journal of Endocrinology 9
In immature male viscachas a significant increase ofnuclear AR and an important decrease of cytoplasmic ARwere observed In prepubertal viscachas more nuclei andcytoplasm appear and there is a clear increase of ARc-ir cellsmainly in the medial region suggesting that more cells beginto express AR in preparation for sexual maturation In adultthe immunostaining is mainly nuclear and regionalized atthe caudal end of PD suggesting that AR are active in thePD cells The present results demonstrate variations in theregionalization and immunostaining pattern of AR duringsexual maturity probably in relation to the contribution ofandrogens reaching the different pituitary zones throughblood vessels Thus the nuclear AR expression in pituitaryPD of viscacha varies according to gonadal developmentand testosterone serum concentration Our results are inagreement with those found by Wood and Newman [24]suggesting that a subset of unoccupied receptors is located inthe cell cytoplasm in the absence of ligand
Bibliographic reports referring to seasonal variations ofRA are scarce Lu et al [56] have recently demonstrated astronger expression ofAR in scented glands during the breed-ing season suggesting that androgens may directly influ-ence the glandular function of the muskrats and courtshipbehavior The viscacha is a seasonal breeding rodent andprevious studies have demonstrated that testosterone acts ongonadotrophs whether directly or indirectly regulating theiractivity in the pituitary gland [28] Castrated viscachamodelshave been used to examine the withdrawal effects of circulat-ing androgens on pituitary cells mainly on the gonadotrophsdue to their involvement in the reproductive processes Inaddition castration resulted in a significant decrease in thepercentage of cells expressing AR throughout the PD [37]The present results demonstrate changes in the amount ofpituitary AR-ir cells during the annual reproductive cycleof adult male viscacha which might be due to variations inthe supply of ligands across blood irrigation The maximumvalues of AR expression were found in the reproductiveperiod and the minimum values in the regression period inrelation to androgen concentrationsThus ARmight regulatethe activity of the pituitary cells involved in the reproductivecycle of adult male Lagostomus
In conclusion our results demonstrate thatAR expressionis related to the pituitary histophysiological changes duringthe growth and sexual maturity of male viscacha Moreoverthey are likely to participate in the seasonal reproductioncontrol in adulthoodHowever the precise actionmechanismof AR in the pituitary viscacha needs further study
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Mrs A Bernardi and Mr JArroyuelo for their technical participation This work wassupported by Project 2-2614 Secretaria de Ciencia y TecnicaUniversidad Nacional de San Luis
References
[1] A K Roy Y Lavrovsky C S Song et al ldquoRegulation ofandrogen actionrdquo Vitamins amp Hormones vol 55 pp 309ndash3521999
[2] S Denayer C Helsen L Thorrez A Haelens and F ClaessensldquoThe rules of DNA recognition by the androgen receptorrdquoMolecular Endocrinology vol 24 no 5 pp 898ndash913 2010
[3] E V Jensen G L Greene L E Closs E R DeSombre and MNadji ldquoReceptors reconsidered a 20-year perspectiverdquo RecentProgress in Hormone Research vol 38 pp 1ndash40 1982
[4] B D SchanbacherM P Johnson andD J Tindall ldquoAndrogenicregulation of luteinizing hormone secretion relationship toandrogen binding in sheep pituitaryrdquo Biology of Reproductionvol 36 no 2 pp 340ndash350 1987
[5] R J Handa D L Reid and J A Resko ldquoAndrogen receptorsin brain and pituitary of female rats cyclic changes andcomparisons with the malerdquo Biology of Reproduction vol 34no 2 pp 293ndash303 1986
[6] J V A Choate and J A Resko ldquoEffects of androgen on brain andpituitary androgen receptors and LH secretion of male guineapigsrdquoThe Journal of Steroid Biochemistry andMolecular Biologyvol 59 no 3-4 pp 315ndash322 1996
[7] T J Spady R Shayya V GThackray L Ehrensberger J S Bai-ley and P L Mellon ldquoAndrogen regulates follicle-stimulatinghormone 120573 gene expression in an activin-dependent manner inimmortalized gonadotropesrdquo Molecular Endocrinology vol 18no 4 pp 925ndash940 2004
[8] P B Connolly J V A Choate and J A Resko ldquoEffects ofexogenous androgen on brain androgen receptors of the fetalrhesus monkeyrdquoNeuroendocrinology vol 59 no 3 pp 271ndash2761994
[9] J A Resko A Perkins C E Roselli J A Fitzgerald J V AChoate and F Stormshak ldquoEndocrine correlates of partnerpreference behavior in ramsrdquo Biology of Reproduction vol 55no 1 pp 120ndash126 1996
[10] S E Abdelgadir C E Roselli J V A Choate and J A ReskoldquoAndrogen receptor messenger ribonucleic acid in brains andpituitaries of male Rhesus monkeys studies on distributionhormonal control and relationship to luteinizing hormonesecretionrdquo Biology of Reproduction vol 60 no 5 pp 1251ndash12561999
[11] R W Bonsall H D Rees and R P Michael ldquoThe distributionnuclear uptake and metabolism of [3H]dihydrotestosterone inthe brain pituitary gland and genital tract of the male rhesusmonkeyrdquo Journal of Steroid Biochemistry vol 23 no 4 pp 389ndash398 1985
[12] S S Wu P W Nathanielsz and T J McDonald ldquoImmunocy-tochemical distribution of androgen receptors in the hypotha-lamus and pituitary of the fetal baboon in late gestationrdquoDevelopmental Brain Research vol 84 no 2 pp 278ndash281 1995
[13] C E Wood and M Keller-Wood ldquoOntogeny of androgenreceptor expression in the ovine fetal central nervous systemand pituitaryrdquo Neuroscience Letters vol 439 no 2 pp 153ndash1562008
[14] J V A Choate O D Slayden and J A Resko ldquoImmuno-cytochemical localization of androgen receptors in brains ofdeveloping and adult male rhesus monkeysrdquo Endocrine vol 8no 1 pp 51ndash60 1998
10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
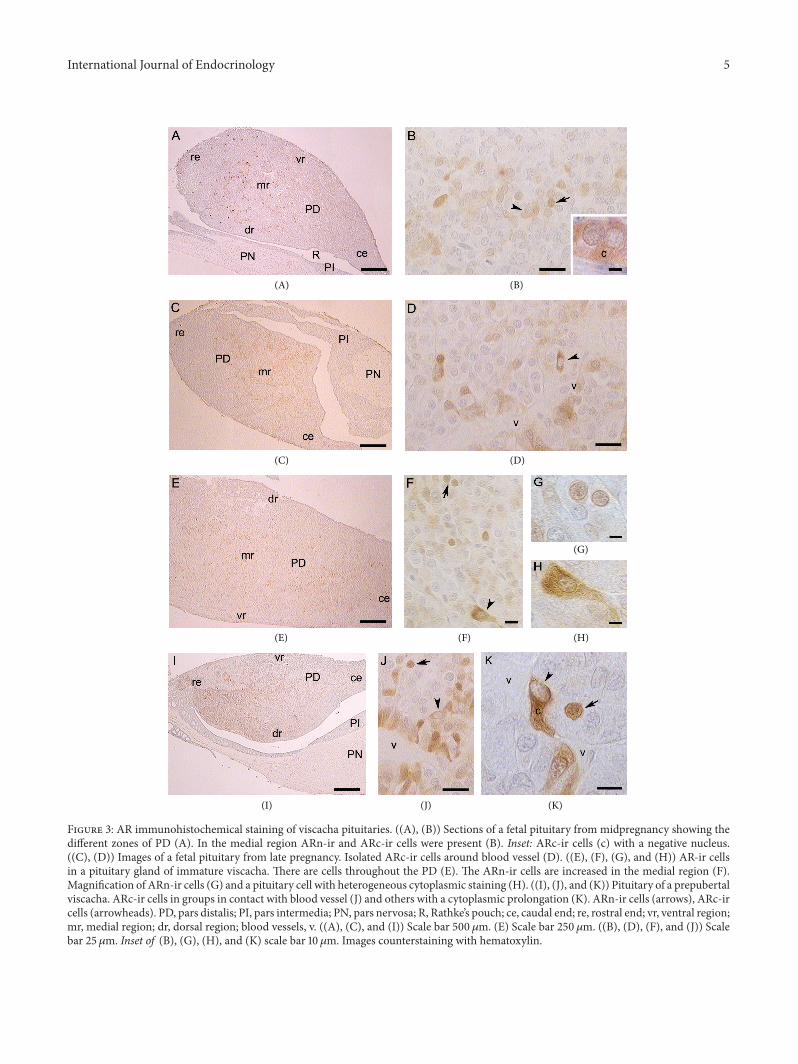
International Journal of Endocrinology 5
(A) (B)
(C)
(E) (F) (H)
(I) (J) (K)
(G)
(D)
Figure 3 AR immunohistochemical staining of viscacha pituitaries ((A) (B)) Sections of a fetal pituitary from midpregnancy showing thedifferent zones of PD (A) In the medial region ARn-ir and ARc-ir cells were present (B) Inset ARc-ir cells (c) with a negative nucleus((C) (D)) Images of a fetal pituitary from late pregnancy Isolated ARc-ir cells around blood vessel (D) ((E) (F) (G) and (H)) AR-ir cellsin a pituitary gland of immature viscacha There are cells throughout the PD (E) The ARn-ir cells are increased in the medial region (F)Magnification of ARn-ir cells (G) and a pituitary cell with heterogeneous cytoplasmic staining (H) ((I) (J) and (K)) Pituitary of a prepubertalviscacha ARc-ir cells in groups in contact with blood vessel (J) and others with a cytoplasmic prolongation (K) ARn-ir cells (arrows) ARc-ircells (arrowheads) PD pars distalis PI pars intermedia PN pars nervosa R Rathkersquos pouch ce caudal end re rostral end vr ventral regionmr medial region dr dorsal region blood vessels v ((A) (C) and (I)) Scale bar 500 120583m (E) Scale bar 250 120583m ((B) (D) (F) and (J)) Scalebar 25 120583m Inset of (B) (G) (H) and (K) scale bar 10 120583m Images counterstaining with hematoxylin
6 International Journal of Endocrinology
Table 1 AR expression in pituitary PD of male viscachas
Distribution Immunostaining pattern Morphological characteristicsMP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cellsLP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cells
Immature Isolated or in small groups Homogeneous and heterogeneous cytoplasmsome intensely stained nuclei Cells oval in shape
Prepubertal Isolated or in groups Homogeneous cytoplasmintensely nuclear staining Irregular cells
Adult-Rep P Isolated or in small groups Nuclear immunoreactivityscarce immunostained cytoplasms Oval spherical or irregular cells
Adult-Reg P Isolated Nuclear immunoreactivityscarce immunostained cytoplasms Small and spherical cytoplasm
Adult-Rec P Isolated or in small groups Nuclear immunoreactivityintense cytoplasmic staining
Spherical oval or irregular cytoplasmsand nuclei
short cytoplasmic prolongation
variations the highest value was found in the medial regionin relation to the ventral region and caudal end (119875 lt001) whereas the lowest percentage was found at the caudalend (Figure 2(a)) The immunostaining of ARn-ir cells wasintense in the dorsal region and the highest percentage ofthese cells was found in the medial region in relation to thedorsal and ventral regions and rostral end (119875 lt 005 Figures2(b) and 3(I)ndash3(K))
In PD of adult animals the AR expression mainly hada nuclear localization In the reproductive period severalpositive nuclei were found at the caudal end and dorsalregion as well as on the dorsal and ventral edges of PDThe percentage of ARn-ir cells was significantly higher atthe caudal end in relation to the other PD zones (119875 lt005 Figure 2(b)) A small number of ARc-ir cells werefrequently located in the medial region The cytoplasmicimmunolabeling pattern was homogeneous and sometimesintense In addition some cells presented heterogeneouscytoplasmic staining and positivity on the nuclear periphery(Figures 4(A)ndash4(E)) No significant changes were observedin the percentage of ARc-ir cells between different PD zones(119875 gt 005 Figure 2(a))
In the gonadal regression period AR-ir cells were mainlyfound in the medial region of PD The immunostainedcytoplasms were small with medial or eccentric nucleusThere were no significant differences in ARc-ir cells betweendifferent PD zones (119875 gt 005 Figure 2(a)) whereas ARn-ircells were more numerous in the medial region in relation tothe ventral region and the rostral end (119875 lt 005 Figure 2(b))
In the gonadal recovery period an increase of AR-ir cellsthroughout PD parenchyma was observed In this periodthe cytoplasmic labeling was more intense and homogeneousin relation to the gonadal regression period (Figures 4(F)ndash4(I)) The percentage of ARc-ir cells was not significantlydifferent between the PD zones (119875 gt 005 Figure 2(a)) Thepercentage of ARn-ir cells was higher at the caudal end inrelation to the rostral end and the medial region (119875 lt 001Figure 2(b)) Table 1 shows a summary of themain cytologicalcharacteristics
The morphometric variations of the AR expression inrelation to development and growth in viscachas PD werestudied by comparing the data obtained from fetal pituitaryduring mid- and late pregnancy and from immature prepu-bertal and adult pituitary in the reproductive period Therewas an increase in ARn-ir cells percentage and a significantdecrease in the percentage of ARc-ir cells in immatureanimals as compared to fetus and in adult animals duringthe reproductive period as compared to prepubertal animalsThe percentage of ARc-ir cells increased significantly inprepubertal in relation to immature viscachas Statisticalanalysis is shown in Figure 5
In order to study the seasonal expression of AR the threeperiods of the reproductive cycle were analyzed The totalpercentages of ARn-ir and ARc-ir cells obtained from threeperiods of the reproductive cycle of adult male viscachaswere statistically analyzed (Figure 5)The percentage of ARn-ir cells showed a significant decrease during the gonadalregression period (119875 lt 0001) No variations were observedin the percentage of ARc-ir cells throughout the reproductivecycle (119875 gt 005)
The testosterone serum concentrations of immature ani-mals were below the detection limit (lt020 ngmL) and toolow to reliably reveal differences from the other groupsThe testosterone serum concentrations of prepubertal andadult viscachas in the gonadal regression period were lowerthan those in adult viscachas during their reproductive andgonadal recovery periods (119875 lt 0001 Figure 6)
4 Discussion
The analysis of the AR expression in the viscacha pituitarygland is essential for a better understanding of the regulatorysystems since androgens are involved in the control ofthe reproductive function This study represents the firstdescription of variations in the regionalization and in theimmunostaining pattern of pituitary AR in relation to thedevelopment growth and reproductive cycle of male vis-cachas
International Journal of Endocrinology 7
(A) (B)
(C) (D) (E)
(F) (G)
(H) (I)
Figure 4 Seasonal expression of AR in pituitary during the reproductive cycle of adult viscachas ((A)ndash(E)) Reproductive period ((A) (B))Images of caudal end the AR were expressed mainly in the nuclei of PD cells (arrows) ((C) (D) and (E)) AR expression at the rostral endventral region and dorsal region respectively The arrowhead indicated a cell with cytoplasmic labeling for AR ((F) (G)) A few AR-ir cellsat caudal end during the gonadal regression period ((H) (I)) AR-ir cells at caudal end during the gonadal recovery period Caudal endce blood vessels v follicular structures f ((A) (F) and (H)) Scale bar 100 120583m ((B) (C) (D) (E) (G) and (I)) Scale bar 25 120583m Imagescounterstaining with hematoxylin
Themain findings of this study can be summarized as fol-lows (1) the regionalization and immunostaining pattern ofpituitary AR vary in relation to growth (2) the AR expressionwas mainly cytoplasmic (ARc-ir) in fetuses (3) a significant
increase of ARc-ir cells was observed in prepubertal animals(4) the nuclear AR (ARn-ir) expression increased in relationto development reaching the highest values in adult animals(5) the ARn-ir cells varied seasonally with maximum values
8 International Journal of Endocrinology
0123456789
10
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
ARn-ir cells ()ARc-ir cells ()
Pitu
itary
PD
()
lowastlowastlowastlowastlowastlowast
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 5 Total percentage of ARn-ir cells and ARc-ir cells infetus (from mid- and late pregnancy) immature prepubertaland adult viscachas (during reproductive gonadal regression andgonadal recovery periods) The result revealed statistically signifi-cant changes lowastlowastlowast119875 lt 0001 immature versus LP-FETUS prepu-bertal versus immature Adult-Rep P versus prepubertal Adult-Reg P versus Adult-Rep P Adult-Rec P versus Adult-Reg P
0
1
2
3
4
5
6
7
Prepubertal Adult-Rep P Adult-Reg P Adult-Rec P
Seru
m te
stoste
rone
conc
entr
atio
n (n
gm
L)
lowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 6 Testosterone serum concentrations The values areexpressed as mean plusmn SEM (119899 = 4) lowastlowast119875 lt 001 Adult-Rep Pversus Adult-Reg P lowastlowastlowast119875 lt 0001 prepubertal versus Adult-RepP Adult-Reg P versus Adult-Rec P
during the reproductive period and minimum values duringthe gonadal regression period (6) the amount of ARn-ir cellswas directly related to testosterone serum concentration
There are still many questions about the relations ofAR expression with the pituitary development and hormonesecretion Wu et al [12] did not observe AR in the pituitarygland during late gestation and suggested that the lack of
feedback by way of AR at pituitary levels may have importantconsequences on the fetal pituitary function More recentlyYuan et al [18] have reported in sheep that AR emerges inthe anterior pituitary before day 60 of gestation and thepercentage of the total anterior pituitary cells expressing thereceptor did not change throughout gestation In additionother studies have concluded that the fetal brain and pitu-itary are target sites for circulating androgens or androgenprecursors in fetal plasma and the increase of androgensin hypothalamic action immediately prior to birth might becritical to the timing of parturition [13] Huang and Yuan[42] have reported that AR expression reflects the functionalstatus of androgen in the pituitary of male sheep lambTheseauthors suggest that during sexual maturity the feedback ofsex hormones on the pituitary is at low level and stimulatesgonadotropin secretion
Some reproductive characteristics of female viscachassuch as polyovulation implantation and offspring birth havebeen described in previous studies Gestation lasts approx-imately 154 days an unusually long period for a rodentand it finishes in spring when the survival probabilities areideal for the mother and offspring [38 43 44] The resultsobtained in this work demonstrate AR in fetal pituitary frommidpregnancy indicating that AR are present around days75ndash80 of gestation in the pituitary cells These receptors arepotentially in position to participate in the regulation of thefetal androgen function Furthermore the analysis of thedistribution morphometry and immunostaining pattern ofAR-ir cells in the parenchyma of fetal glands suggests thatthese cells contain variable amounts of AR and they aremorphologically related to blood supply and other endocrinecells of PD parenchyma In late pregnancy of viscacha thepercentages of ARc-ir and ARn-ir cells did not change withrespect to the values found during midpregnancy Theseresults suggested that fetal pituitary AR might have anyspecific function since midgestation in viscacha
Thehypothalamic-pituitary-gonadal axis is activated dur-ing puberty when hormonalmodulation of sexual behavior isorganized [45] During male development the masculiniza-tion is given by the hormones produced by the fetal testesprimarily testosterone [46 47] The gonadal steroids feedback to the central nervous system to exert a key role inthe central control of the reproductive axis by regulatingthe release of the gonadotropin-releasing hormone (GnRH)[48] However the neurons that synthesize GnRH did notexpress AR and there are sex-steroids sensitive neuronsupstream the GnRH cells where the feedback signals ofthe steroid might be processed to the GnRH cells [49] Theneurons of hypothalamic nuclei that expressed kisspeptin(K) and neurokinin B (NKB) may sense circulating levelsof testosterone through the expression of AR [49 50] Inrodents sheep and primates K and NKB have recently beenimplicated in the control of puberty and the maintenanceof the reproductive function in adulthood [51ndash53] In thepituitary the expression of AR has been mainly observedin gonadotrophs [18 54 55] and gonadal steroid hormonesaffect the gonadotroph population in order to regulate theiractivity
International Journal of Endocrinology 9
In immature male viscachas a significant increase ofnuclear AR and an important decrease of cytoplasmic ARwere observed In prepubertal viscachas more nuclei andcytoplasm appear and there is a clear increase of ARc-ir cellsmainly in the medial region suggesting that more cells beginto express AR in preparation for sexual maturation In adultthe immunostaining is mainly nuclear and regionalized atthe caudal end of PD suggesting that AR are active in thePD cells The present results demonstrate variations in theregionalization and immunostaining pattern of AR duringsexual maturity probably in relation to the contribution ofandrogens reaching the different pituitary zones throughblood vessels Thus the nuclear AR expression in pituitaryPD of viscacha varies according to gonadal developmentand testosterone serum concentration Our results are inagreement with those found by Wood and Newman [24]suggesting that a subset of unoccupied receptors is located inthe cell cytoplasm in the absence of ligand
Bibliographic reports referring to seasonal variations ofRA are scarce Lu et al [56] have recently demonstrated astronger expression ofAR in scented glands during the breed-ing season suggesting that androgens may directly influ-ence the glandular function of the muskrats and courtshipbehavior The viscacha is a seasonal breeding rodent andprevious studies have demonstrated that testosterone acts ongonadotrophs whether directly or indirectly regulating theiractivity in the pituitary gland [28] Castrated viscachamodelshave been used to examine the withdrawal effects of circulat-ing androgens on pituitary cells mainly on the gonadotrophsdue to their involvement in the reproductive processes Inaddition castration resulted in a significant decrease in thepercentage of cells expressing AR throughout the PD [37]The present results demonstrate changes in the amount ofpituitary AR-ir cells during the annual reproductive cycleof adult male viscacha which might be due to variations inthe supply of ligands across blood irrigation The maximumvalues of AR expression were found in the reproductiveperiod and the minimum values in the regression period inrelation to androgen concentrationsThus ARmight regulatethe activity of the pituitary cells involved in the reproductivecycle of adult male Lagostomus
In conclusion our results demonstrate thatAR expressionis related to the pituitary histophysiological changes duringthe growth and sexual maturity of male viscacha Moreoverthey are likely to participate in the seasonal reproductioncontrol in adulthoodHowever the precise actionmechanismof AR in the pituitary viscacha needs further study
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Mrs A Bernardi and Mr JArroyuelo for their technical participation This work wassupported by Project 2-2614 Secretaria de Ciencia y TecnicaUniversidad Nacional de San Luis
References
[1] A K Roy Y Lavrovsky C S Song et al ldquoRegulation ofandrogen actionrdquo Vitamins amp Hormones vol 55 pp 309ndash3521999
[2] S Denayer C Helsen L Thorrez A Haelens and F ClaessensldquoThe rules of DNA recognition by the androgen receptorrdquoMolecular Endocrinology vol 24 no 5 pp 898ndash913 2010
[3] E V Jensen G L Greene L E Closs E R DeSombre and MNadji ldquoReceptors reconsidered a 20-year perspectiverdquo RecentProgress in Hormone Research vol 38 pp 1ndash40 1982
[4] B D SchanbacherM P Johnson andD J Tindall ldquoAndrogenicregulation of luteinizing hormone secretion relationship toandrogen binding in sheep pituitaryrdquo Biology of Reproductionvol 36 no 2 pp 340ndash350 1987
[5] R J Handa D L Reid and J A Resko ldquoAndrogen receptorsin brain and pituitary of female rats cyclic changes andcomparisons with the malerdquo Biology of Reproduction vol 34no 2 pp 293ndash303 1986
[6] J V A Choate and J A Resko ldquoEffects of androgen on brain andpituitary androgen receptors and LH secretion of male guineapigsrdquoThe Journal of Steroid Biochemistry andMolecular Biologyvol 59 no 3-4 pp 315ndash322 1996
[7] T J Spady R Shayya V GThackray L Ehrensberger J S Bai-ley and P L Mellon ldquoAndrogen regulates follicle-stimulatinghormone 120573 gene expression in an activin-dependent manner inimmortalized gonadotropesrdquo Molecular Endocrinology vol 18no 4 pp 925ndash940 2004
[8] P B Connolly J V A Choate and J A Resko ldquoEffects ofexogenous androgen on brain androgen receptors of the fetalrhesus monkeyrdquoNeuroendocrinology vol 59 no 3 pp 271ndash2761994
[9] J A Resko A Perkins C E Roselli J A Fitzgerald J V AChoate and F Stormshak ldquoEndocrine correlates of partnerpreference behavior in ramsrdquo Biology of Reproduction vol 55no 1 pp 120ndash126 1996
[10] S E Abdelgadir C E Roselli J V A Choate and J A ReskoldquoAndrogen receptor messenger ribonucleic acid in brains andpituitaries of male Rhesus monkeys studies on distributionhormonal control and relationship to luteinizing hormonesecretionrdquo Biology of Reproduction vol 60 no 5 pp 1251ndash12561999
[11] R W Bonsall H D Rees and R P Michael ldquoThe distributionnuclear uptake and metabolism of [3H]dihydrotestosterone inthe brain pituitary gland and genital tract of the male rhesusmonkeyrdquo Journal of Steroid Biochemistry vol 23 no 4 pp 389ndash398 1985
[12] S S Wu P W Nathanielsz and T J McDonald ldquoImmunocy-tochemical distribution of androgen receptors in the hypotha-lamus and pituitary of the fetal baboon in late gestationrdquoDevelopmental Brain Research vol 84 no 2 pp 278ndash281 1995
[13] C E Wood and M Keller-Wood ldquoOntogeny of androgenreceptor expression in the ovine fetal central nervous systemand pituitaryrdquo Neuroscience Letters vol 439 no 2 pp 153ndash1562008
[14] J V A Choate O D Slayden and J A Resko ldquoImmuno-cytochemical localization of androgen receptors in brains ofdeveloping and adult male rhesus monkeysrdquo Endocrine vol 8no 1 pp 51ndash60 1998
10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 International Journal of Endocrinology
Table 1 AR expression in pituitary PD of male viscachas
Distribution Immunostaining pattern Morphological characteristicsMP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cellsLP-FETUS Isolated or in small groups Cytoplasmic and homogeneous Oval or spherical cells
Immature Isolated or in small groups Homogeneous and heterogeneous cytoplasmsome intensely stained nuclei Cells oval in shape
Prepubertal Isolated or in groups Homogeneous cytoplasmintensely nuclear staining Irregular cells
Adult-Rep P Isolated or in small groups Nuclear immunoreactivityscarce immunostained cytoplasms Oval spherical or irregular cells
Adult-Reg P Isolated Nuclear immunoreactivityscarce immunostained cytoplasms Small and spherical cytoplasm
Adult-Rec P Isolated or in small groups Nuclear immunoreactivityintense cytoplasmic staining
Spherical oval or irregular cytoplasmsand nuclei
short cytoplasmic prolongation
variations the highest value was found in the medial regionin relation to the ventral region and caudal end (119875 lt001) whereas the lowest percentage was found at the caudalend (Figure 2(a)) The immunostaining of ARn-ir cells wasintense in the dorsal region and the highest percentage ofthese cells was found in the medial region in relation to thedorsal and ventral regions and rostral end (119875 lt 005 Figures2(b) and 3(I)ndash3(K))
In PD of adult animals the AR expression mainly hada nuclear localization In the reproductive period severalpositive nuclei were found at the caudal end and dorsalregion as well as on the dorsal and ventral edges of PDThe percentage of ARn-ir cells was significantly higher atthe caudal end in relation to the other PD zones (119875 lt005 Figure 2(b)) A small number of ARc-ir cells werefrequently located in the medial region The cytoplasmicimmunolabeling pattern was homogeneous and sometimesintense In addition some cells presented heterogeneouscytoplasmic staining and positivity on the nuclear periphery(Figures 4(A)ndash4(E)) No significant changes were observedin the percentage of ARc-ir cells between different PD zones(119875 gt 005 Figure 2(a))
In the gonadal regression period AR-ir cells were mainlyfound in the medial region of PD The immunostainedcytoplasms were small with medial or eccentric nucleusThere were no significant differences in ARc-ir cells betweendifferent PD zones (119875 gt 005 Figure 2(a)) whereas ARn-ircells were more numerous in the medial region in relation tothe ventral region and the rostral end (119875 lt 005 Figure 2(b))
In the gonadal recovery period an increase of AR-ir cellsthroughout PD parenchyma was observed In this periodthe cytoplasmic labeling was more intense and homogeneousin relation to the gonadal regression period (Figures 4(F)ndash4(I)) The percentage of ARc-ir cells was not significantlydifferent between the PD zones (119875 gt 005 Figure 2(a)) Thepercentage of ARn-ir cells was higher at the caudal end inrelation to the rostral end and the medial region (119875 lt 001Figure 2(b)) Table 1 shows a summary of themain cytologicalcharacteristics
The morphometric variations of the AR expression inrelation to development and growth in viscachas PD werestudied by comparing the data obtained from fetal pituitaryduring mid- and late pregnancy and from immature prepu-bertal and adult pituitary in the reproductive period Therewas an increase in ARn-ir cells percentage and a significantdecrease in the percentage of ARc-ir cells in immatureanimals as compared to fetus and in adult animals duringthe reproductive period as compared to prepubertal animalsThe percentage of ARc-ir cells increased significantly inprepubertal in relation to immature viscachas Statisticalanalysis is shown in Figure 5
In order to study the seasonal expression of AR the threeperiods of the reproductive cycle were analyzed The totalpercentages of ARn-ir and ARc-ir cells obtained from threeperiods of the reproductive cycle of adult male viscachaswere statistically analyzed (Figure 5)The percentage of ARn-ir cells showed a significant decrease during the gonadalregression period (119875 lt 0001) No variations were observedin the percentage of ARc-ir cells throughout the reproductivecycle (119875 gt 005)
The testosterone serum concentrations of immature ani-mals were below the detection limit (lt020 ngmL) and toolow to reliably reveal differences from the other groupsThe testosterone serum concentrations of prepubertal andadult viscachas in the gonadal regression period were lowerthan those in adult viscachas during their reproductive andgonadal recovery periods (119875 lt 0001 Figure 6)
4 Discussion
The analysis of the AR expression in the viscacha pituitarygland is essential for a better understanding of the regulatorysystems since androgens are involved in the control ofthe reproductive function This study represents the firstdescription of variations in the regionalization and in theimmunostaining pattern of pituitary AR in relation to thedevelopment growth and reproductive cycle of male vis-cachas
International Journal of Endocrinology 7
(A) (B)
(C) (D) (E)
(F) (G)
(H) (I)
Figure 4 Seasonal expression of AR in pituitary during the reproductive cycle of adult viscachas ((A)ndash(E)) Reproductive period ((A) (B))Images of caudal end the AR were expressed mainly in the nuclei of PD cells (arrows) ((C) (D) and (E)) AR expression at the rostral endventral region and dorsal region respectively The arrowhead indicated a cell with cytoplasmic labeling for AR ((F) (G)) A few AR-ir cellsat caudal end during the gonadal regression period ((H) (I)) AR-ir cells at caudal end during the gonadal recovery period Caudal endce blood vessels v follicular structures f ((A) (F) and (H)) Scale bar 100 120583m ((B) (C) (D) (E) (G) and (I)) Scale bar 25 120583m Imagescounterstaining with hematoxylin
Themain findings of this study can be summarized as fol-lows (1) the regionalization and immunostaining pattern ofpituitary AR vary in relation to growth (2) the AR expressionwas mainly cytoplasmic (ARc-ir) in fetuses (3) a significant
increase of ARc-ir cells was observed in prepubertal animals(4) the nuclear AR (ARn-ir) expression increased in relationto development reaching the highest values in adult animals(5) the ARn-ir cells varied seasonally with maximum values
8 International Journal of Endocrinology
0123456789
10
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
ARn-ir cells ()ARc-ir cells ()
Pitu
itary
PD
()
lowastlowastlowastlowastlowastlowast
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 5 Total percentage of ARn-ir cells and ARc-ir cells infetus (from mid- and late pregnancy) immature prepubertaland adult viscachas (during reproductive gonadal regression andgonadal recovery periods) The result revealed statistically signifi-cant changes lowastlowastlowast119875 lt 0001 immature versus LP-FETUS prepu-bertal versus immature Adult-Rep P versus prepubertal Adult-Reg P versus Adult-Rep P Adult-Rec P versus Adult-Reg P
0
1
2
3
4
5
6
7
Prepubertal Adult-Rep P Adult-Reg P Adult-Rec P
Seru
m te
stoste
rone
conc
entr
atio
n (n
gm
L)
lowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 6 Testosterone serum concentrations The values areexpressed as mean plusmn SEM (119899 = 4) lowastlowast119875 lt 001 Adult-Rep Pversus Adult-Reg P lowastlowastlowast119875 lt 0001 prepubertal versus Adult-RepP Adult-Reg P versus Adult-Rec P
during the reproductive period and minimum values duringthe gonadal regression period (6) the amount of ARn-ir cellswas directly related to testosterone serum concentration
There are still many questions about the relations ofAR expression with the pituitary development and hormonesecretion Wu et al [12] did not observe AR in the pituitarygland during late gestation and suggested that the lack of
feedback by way of AR at pituitary levels may have importantconsequences on the fetal pituitary function More recentlyYuan et al [18] have reported in sheep that AR emerges inthe anterior pituitary before day 60 of gestation and thepercentage of the total anterior pituitary cells expressing thereceptor did not change throughout gestation In additionother studies have concluded that the fetal brain and pitu-itary are target sites for circulating androgens or androgenprecursors in fetal plasma and the increase of androgensin hypothalamic action immediately prior to birth might becritical to the timing of parturition [13] Huang and Yuan[42] have reported that AR expression reflects the functionalstatus of androgen in the pituitary of male sheep lambTheseauthors suggest that during sexual maturity the feedback ofsex hormones on the pituitary is at low level and stimulatesgonadotropin secretion
Some reproductive characteristics of female viscachassuch as polyovulation implantation and offspring birth havebeen described in previous studies Gestation lasts approx-imately 154 days an unusually long period for a rodentand it finishes in spring when the survival probabilities areideal for the mother and offspring [38 43 44] The resultsobtained in this work demonstrate AR in fetal pituitary frommidpregnancy indicating that AR are present around days75ndash80 of gestation in the pituitary cells These receptors arepotentially in position to participate in the regulation of thefetal androgen function Furthermore the analysis of thedistribution morphometry and immunostaining pattern ofAR-ir cells in the parenchyma of fetal glands suggests thatthese cells contain variable amounts of AR and they aremorphologically related to blood supply and other endocrinecells of PD parenchyma In late pregnancy of viscacha thepercentages of ARc-ir and ARn-ir cells did not change withrespect to the values found during midpregnancy Theseresults suggested that fetal pituitary AR might have anyspecific function since midgestation in viscacha
Thehypothalamic-pituitary-gonadal axis is activated dur-ing puberty when hormonalmodulation of sexual behavior isorganized [45] During male development the masculiniza-tion is given by the hormones produced by the fetal testesprimarily testosterone [46 47] The gonadal steroids feedback to the central nervous system to exert a key role inthe central control of the reproductive axis by regulatingthe release of the gonadotropin-releasing hormone (GnRH)[48] However the neurons that synthesize GnRH did notexpress AR and there are sex-steroids sensitive neuronsupstream the GnRH cells where the feedback signals ofthe steroid might be processed to the GnRH cells [49] Theneurons of hypothalamic nuclei that expressed kisspeptin(K) and neurokinin B (NKB) may sense circulating levelsof testosterone through the expression of AR [49 50] Inrodents sheep and primates K and NKB have recently beenimplicated in the control of puberty and the maintenanceof the reproductive function in adulthood [51ndash53] In thepituitary the expression of AR has been mainly observedin gonadotrophs [18 54 55] and gonadal steroid hormonesaffect the gonadotroph population in order to regulate theiractivity
International Journal of Endocrinology 9
In immature male viscachas a significant increase ofnuclear AR and an important decrease of cytoplasmic ARwere observed In prepubertal viscachas more nuclei andcytoplasm appear and there is a clear increase of ARc-ir cellsmainly in the medial region suggesting that more cells beginto express AR in preparation for sexual maturation In adultthe immunostaining is mainly nuclear and regionalized atthe caudal end of PD suggesting that AR are active in thePD cells The present results demonstrate variations in theregionalization and immunostaining pattern of AR duringsexual maturity probably in relation to the contribution ofandrogens reaching the different pituitary zones throughblood vessels Thus the nuclear AR expression in pituitaryPD of viscacha varies according to gonadal developmentand testosterone serum concentration Our results are inagreement with those found by Wood and Newman [24]suggesting that a subset of unoccupied receptors is located inthe cell cytoplasm in the absence of ligand
Bibliographic reports referring to seasonal variations ofRA are scarce Lu et al [56] have recently demonstrated astronger expression ofAR in scented glands during the breed-ing season suggesting that androgens may directly influ-ence the glandular function of the muskrats and courtshipbehavior The viscacha is a seasonal breeding rodent andprevious studies have demonstrated that testosterone acts ongonadotrophs whether directly or indirectly regulating theiractivity in the pituitary gland [28] Castrated viscachamodelshave been used to examine the withdrawal effects of circulat-ing androgens on pituitary cells mainly on the gonadotrophsdue to their involvement in the reproductive processes Inaddition castration resulted in a significant decrease in thepercentage of cells expressing AR throughout the PD [37]The present results demonstrate changes in the amount ofpituitary AR-ir cells during the annual reproductive cycleof adult male viscacha which might be due to variations inthe supply of ligands across blood irrigation The maximumvalues of AR expression were found in the reproductiveperiod and the minimum values in the regression period inrelation to androgen concentrationsThus ARmight regulatethe activity of the pituitary cells involved in the reproductivecycle of adult male Lagostomus
In conclusion our results demonstrate thatAR expressionis related to the pituitary histophysiological changes duringthe growth and sexual maturity of male viscacha Moreoverthey are likely to participate in the seasonal reproductioncontrol in adulthoodHowever the precise actionmechanismof AR in the pituitary viscacha needs further study
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Mrs A Bernardi and Mr JArroyuelo for their technical participation This work wassupported by Project 2-2614 Secretaria de Ciencia y TecnicaUniversidad Nacional de San Luis
References
[1] A K Roy Y Lavrovsky C S Song et al ldquoRegulation ofandrogen actionrdquo Vitamins amp Hormones vol 55 pp 309ndash3521999
[2] S Denayer C Helsen L Thorrez A Haelens and F ClaessensldquoThe rules of DNA recognition by the androgen receptorrdquoMolecular Endocrinology vol 24 no 5 pp 898ndash913 2010
[3] E V Jensen G L Greene L E Closs E R DeSombre and MNadji ldquoReceptors reconsidered a 20-year perspectiverdquo RecentProgress in Hormone Research vol 38 pp 1ndash40 1982
[4] B D SchanbacherM P Johnson andD J Tindall ldquoAndrogenicregulation of luteinizing hormone secretion relationship toandrogen binding in sheep pituitaryrdquo Biology of Reproductionvol 36 no 2 pp 340ndash350 1987
[5] R J Handa D L Reid and J A Resko ldquoAndrogen receptorsin brain and pituitary of female rats cyclic changes andcomparisons with the malerdquo Biology of Reproduction vol 34no 2 pp 293ndash303 1986
[6] J V A Choate and J A Resko ldquoEffects of androgen on brain andpituitary androgen receptors and LH secretion of male guineapigsrdquoThe Journal of Steroid Biochemistry andMolecular Biologyvol 59 no 3-4 pp 315ndash322 1996
[7] T J Spady R Shayya V GThackray L Ehrensberger J S Bai-ley and P L Mellon ldquoAndrogen regulates follicle-stimulatinghormone 120573 gene expression in an activin-dependent manner inimmortalized gonadotropesrdquo Molecular Endocrinology vol 18no 4 pp 925ndash940 2004
[8] P B Connolly J V A Choate and J A Resko ldquoEffects ofexogenous androgen on brain androgen receptors of the fetalrhesus monkeyrdquoNeuroendocrinology vol 59 no 3 pp 271ndash2761994
[9] J A Resko A Perkins C E Roselli J A Fitzgerald J V AChoate and F Stormshak ldquoEndocrine correlates of partnerpreference behavior in ramsrdquo Biology of Reproduction vol 55no 1 pp 120ndash126 1996
[10] S E Abdelgadir C E Roselli J V A Choate and J A ReskoldquoAndrogen receptor messenger ribonucleic acid in brains andpituitaries of male Rhesus monkeys studies on distributionhormonal control and relationship to luteinizing hormonesecretionrdquo Biology of Reproduction vol 60 no 5 pp 1251ndash12561999
[11] R W Bonsall H D Rees and R P Michael ldquoThe distributionnuclear uptake and metabolism of [3H]dihydrotestosterone inthe brain pituitary gland and genital tract of the male rhesusmonkeyrdquo Journal of Steroid Biochemistry vol 23 no 4 pp 389ndash398 1985
[12] S S Wu P W Nathanielsz and T J McDonald ldquoImmunocy-tochemical distribution of androgen receptors in the hypotha-lamus and pituitary of the fetal baboon in late gestationrdquoDevelopmental Brain Research vol 84 no 2 pp 278ndash281 1995
[13] C E Wood and M Keller-Wood ldquoOntogeny of androgenreceptor expression in the ovine fetal central nervous systemand pituitaryrdquo Neuroscience Letters vol 439 no 2 pp 153ndash1562008
[14] J V A Choate O D Slayden and J A Resko ldquoImmuno-cytochemical localization of androgen receptors in brains ofdeveloping and adult male rhesus monkeysrdquo Endocrine vol 8no 1 pp 51ndash60 1998
10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

International Journal of Endocrinology 7
(A) (B)
(C) (D) (E)
(F) (G)
(H) (I)
Figure 4 Seasonal expression of AR in pituitary during the reproductive cycle of adult viscachas ((A)ndash(E)) Reproductive period ((A) (B))Images of caudal end the AR were expressed mainly in the nuclei of PD cells (arrows) ((C) (D) and (E)) AR expression at the rostral endventral region and dorsal region respectively The arrowhead indicated a cell with cytoplasmic labeling for AR ((F) (G)) A few AR-ir cellsat caudal end during the gonadal regression period ((H) (I)) AR-ir cells at caudal end during the gonadal recovery period Caudal endce blood vessels v follicular structures f ((A) (F) and (H)) Scale bar 100 120583m ((B) (C) (D) (E) (G) and (I)) Scale bar 25 120583m Imagescounterstaining with hematoxylin
Themain findings of this study can be summarized as fol-lows (1) the regionalization and immunostaining pattern ofpituitary AR vary in relation to growth (2) the AR expressionwas mainly cytoplasmic (ARc-ir) in fetuses (3) a significant
increase of ARc-ir cells was observed in prepubertal animals(4) the nuclear AR (ARn-ir) expression increased in relationto development reaching the highest values in adult animals(5) the ARn-ir cells varied seasonally with maximum values
8 International Journal of Endocrinology
0123456789
10
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
ARn-ir cells ()ARc-ir cells ()
Pitu
itary
PD
()
lowastlowastlowastlowastlowastlowast
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 5 Total percentage of ARn-ir cells and ARc-ir cells infetus (from mid- and late pregnancy) immature prepubertaland adult viscachas (during reproductive gonadal regression andgonadal recovery periods) The result revealed statistically signifi-cant changes lowastlowastlowast119875 lt 0001 immature versus LP-FETUS prepu-bertal versus immature Adult-Rep P versus prepubertal Adult-Reg P versus Adult-Rep P Adult-Rec P versus Adult-Reg P
0
1
2
3
4
5
6
7
Prepubertal Adult-Rep P Adult-Reg P Adult-Rec P
Seru
m te
stoste
rone
conc
entr
atio
n (n
gm
L)
lowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 6 Testosterone serum concentrations The values areexpressed as mean plusmn SEM (119899 = 4) lowastlowast119875 lt 001 Adult-Rep Pversus Adult-Reg P lowastlowastlowast119875 lt 0001 prepubertal versus Adult-RepP Adult-Reg P versus Adult-Rec P
during the reproductive period and minimum values duringthe gonadal regression period (6) the amount of ARn-ir cellswas directly related to testosterone serum concentration
There are still many questions about the relations ofAR expression with the pituitary development and hormonesecretion Wu et al [12] did not observe AR in the pituitarygland during late gestation and suggested that the lack of
feedback by way of AR at pituitary levels may have importantconsequences on the fetal pituitary function More recentlyYuan et al [18] have reported in sheep that AR emerges inthe anterior pituitary before day 60 of gestation and thepercentage of the total anterior pituitary cells expressing thereceptor did not change throughout gestation In additionother studies have concluded that the fetal brain and pitu-itary are target sites for circulating androgens or androgenprecursors in fetal plasma and the increase of androgensin hypothalamic action immediately prior to birth might becritical to the timing of parturition [13] Huang and Yuan[42] have reported that AR expression reflects the functionalstatus of androgen in the pituitary of male sheep lambTheseauthors suggest that during sexual maturity the feedback ofsex hormones on the pituitary is at low level and stimulatesgonadotropin secretion
Some reproductive characteristics of female viscachassuch as polyovulation implantation and offspring birth havebeen described in previous studies Gestation lasts approx-imately 154 days an unusually long period for a rodentand it finishes in spring when the survival probabilities areideal for the mother and offspring [38 43 44] The resultsobtained in this work demonstrate AR in fetal pituitary frommidpregnancy indicating that AR are present around days75ndash80 of gestation in the pituitary cells These receptors arepotentially in position to participate in the regulation of thefetal androgen function Furthermore the analysis of thedistribution morphometry and immunostaining pattern ofAR-ir cells in the parenchyma of fetal glands suggests thatthese cells contain variable amounts of AR and they aremorphologically related to blood supply and other endocrinecells of PD parenchyma In late pregnancy of viscacha thepercentages of ARc-ir and ARn-ir cells did not change withrespect to the values found during midpregnancy Theseresults suggested that fetal pituitary AR might have anyspecific function since midgestation in viscacha
Thehypothalamic-pituitary-gonadal axis is activated dur-ing puberty when hormonalmodulation of sexual behavior isorganized [45] During male development the masculiniza-tion is given by the hormones produced by the fetal testesprimarily testosterone [46 47] The gonadal steroids feedback to the central nervous system to exert a key role inthe central control of the reproductive axis by regulatingthe release of the gonadotropin-releasing hormone (GnRH)[48] However the neurons that synthesize GnRH did notexpress AR and there are sex-steroids sensitive neuronsupstream the GnRH cells where the feedback signals ofthe steroid might be processed to the GnRH cells [49] Theneurons of hypothalamic nuclei that expressed kisspeptin(K) and neurokinin B (NKB) may sense circulating levelsof testosterone through the expression of AR [49 50] Inrodents sheep and primates K and NKB have recently beenimplicated in the control of puberty and the maintenanceof the reproductive function in adulthood [51ndash53] In thepituitary the expression of AR has been mainly observedin gonadotrophs [18 54 55] and gonadal steroid hormonesaffect the gonadotroph population in order to regulate theiractivity
International Journal of Endocrinology 9
In immature male viscachas a significant increase ofnuclear AR and an important decrease of cytoplasmic ARwere observed In prepubertal viscachas more nuclei andcytoplasm appear and there is a clear increase of ARc-ir cellsmainly in the medial region suggesting that more cells beginto express AR in preparation for sexual maturation In adultthe immunostaining is mainly nuclear and regionalized atthe caudal end of PD suggesting that AR are active in thePD cells The present results demonstrate variations in theregionalization and immunostaining pattern of AR duringsexual maturity probably in relation to the contribution ofandrogens reaching the different pituitary zones throughblood vessels Thus the nuclear AR expression in pituitaryPD of viscacha varies according to gonadal developmentand testosterone serum concentration Our results are inagreement with those found by Wood and Newman [24]suggesting that a subset of unoccupied receptors is located inthe cell cytoplasm in the absence of ligand
Bibliographic reports referring to seasonal variations ofRA are scarce Lu et al [56] have recently demonstrated astronger expression ofAR in scented glands during the breed-ing season suggesting that androgens may directly influ-ence the glandular function of the muskrats and courtshipbehavior The viscacha is a seasonal breeding rodent andprevious studies have demonstrated that testosterone acts ongonadotrophs whether directly or indirectly regulating theiractivity in the pituitary gland [28] Castrated viscachamodelshave been used to examine the withdrawal effects of circulat-ing androgens on pituitary cells mainly on the gonadotrophsdue to their involvement in the reproductive processes Inaddition castration resulted in a significant decrease in thepercentage of cells expressing AR throughout the PD [37]The present results demonstrate changes in the amount ofpituitary AR-ir cells during the annual reproductive cycleof adult male viscacha which might be due to variations inthe supply of ligands across blood irrigation The maximumvalues of AR expression were found in the reproductiveperiod and the minimum values in the regression period inrelation to androgen concentrationsThus ARmight regulatethe activity of the pituitary cells involved in the reproductivecycle of adult male Lagostomus
In conclusion our results demonstrate thatAR expressionis related to the pituitary histophysiological changes duringthe growth and sexual maturity of male viscacha Moreoverthey are likely to participate in the seasonal reproductioncontrol in adulthoodHowever the precise actionmechanismof AR in the pituitary viscacha needs further study
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Mrs A Bernardi and Mr JArroyuelo for their technical participation This work wassupported by Project 2-2614 Secretaria de Ciencia y TecnicaUniversidad Nacional de San Luis
References
[1] A K Roy Y Lavrovsky C S Song et al ldquoRegulation ofandrogen actionrdquo Vitamins amp Hormones vol 55 pp 309ndash3521999
[2] S Denayer C Helsen L Thorrez A Haelens and F ClaessensldquoThe rules of DNA recognition by the androgen receptorrdquoMolecular Endocrinology vol 24 no 5 pp 898ndash913 2010
[3] E V Jensen G L Greene L E Closs E R DeSombre and MNadji ldquoReceptors reconsidered a 20-year perspectiverdquo RecentProgress in Hormone Research vol 38 pp 1ndash40 1982
[4] B D SchanbacherM P Johnson andD J Tindall ldquoAndrogenicregulation of luteinizing hormone secretion relationship toandrogen binding in sheep pituitaryrdquo Biology of Reproductionvol 36 no 2 pp 340ndash350 1987
[5] R J Handa D L Reid and J A Resko ldquoAndrogen receptorsin brain and pituitary of female rats cyclic changes andcomparisons with the malerdquo Biology of Reproduction vol 34no 2 pp 293ndash303 1986
[6] J V A Choate and J A Resko ldquoEffects of androgen on brain andpituitary androgen receptors and LH secretion of male guineapigsrdquoThe Journal of Steroid Biochemistry andMolecular Biologyvol 59 no 3-4 pp 315ndash322 1996
[7] T J Spady R Shayya V GThackray L Ehrensberger J S Bai-ley and P L Mellon ldquoAndrogen regulates follicle-stimulatinghormone 120573 gene expression in an activin-dependent manner inimmortalized gonadotropesrdquo Molecular Endocrinology vol 18no 4 pp 925ndash940 2004
[8] P B Connolly J V A Choate and J A Resko ldquoEffects ofexogenous androgen on brain androgen receptors of the fetalrhesus monkeyrdquoNeuroendocrinology vol 59 no 3 pp 271ndash2761994
[9] J A Resko A Perkins C E Roselli J A Fitzgerald J V AChoate and F Stormshak ldquoEndocrine correlates of partnerpreference behavior in ramsrdquo Biology of Reproduction vol 55no 1 pp 120ndash126 1996
[10] S E Abdelgadir C E Roselli J V A Choate and J A ReskoldquoAndrogen receptor messenger ribonucleic acid in brains andpituitaries of male Rhesus monkeys studies on distributionhormonal control and relationship to luteinizing hormonesecretionrdquo Biology of Reproduction vol 60 no 5 pp 1251ndash12561999
[11] R W Bonsall H D Rees and R P Michael ldquoThe distributionnuclear uptake and metabolism of [3H]dihydrotestosterone inthe brain pituitary gland and genital tract of the male rhesusmonkeyrdquo Journal of Steroid Biochemistry vol 23 no 4 pp 389ndash398 1985
[12] S S Wu P W Nathanielsz and T J McDonald ldquoImmunocy-tochemical distribution of androgen receptors in the hypotha-lamus and pituitary of the fetal baboon in late gestationrdquoDevelopmental Brain Research vol 84 no 2 pp 278ndash281 1995
[13] C E Wood and M Keller-Wood ldquoOntogeny of androgenreceptor expression in the ovine fetal central nervous systemand pituitaryrdquo Neuroscience Letters vol 439 no 2 pp 153ndash1562008
[14] J V A Choate O D Slayden and J A Resko ldquoImmuno-cytochemical localization of androgen receptors in brains ofdeveloping and adult male rhesus monkeysrdquo Endocrine vol 8no 1 pp 51ndash60 1998
10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
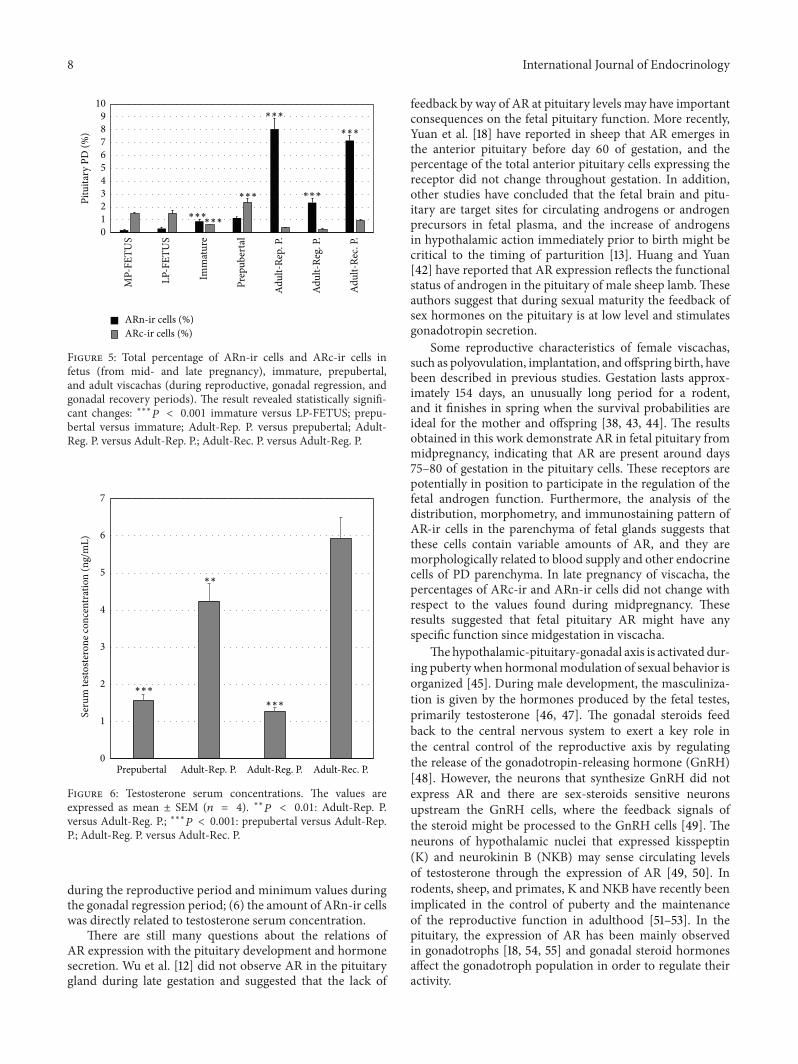
8 International Journal of Endocrinology
0123456789
10
MP-
FETU
S
LP-F
ETU
S
Imm
atur
e
Prep
uber
tal
Adul
t-Rep
P
Adul
t-Reg
P
Adul
t-Rec
P
ARn-ir cells ()ARc-ir cells ()
Pitu
itary
PD
()
lowastlowastlowastlowastlowastlowast
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 5 Total percentage of ARn-ir cells and ARc-ir cells infetus (from mid- and late pregnancy) immature prepubertaland adult viscachas (during reproductive gonadal regression andgonadal recovery periods) The result revealed statistically signifi-cant changes lowastlowastlowast119875 lt 0001 immature versus LP-FETUS prepu-bertal versus immature Adult-Rep P versus prepubertal Adult-Reg P versus Adult-Rep P Adult-Rec P versus Adult-Reg P
0
1
2
3
4
5
6
7
Prepubertal Adult-Rep P Adult-Reg P Adult-Rec P
Seru
m te
stoste
rone
conc
entr
atio
n (n
gm
L)
lowastlowast
lowastlowastlowast
lowastlowastlowast
Figure 6 Testosterone serum concentrations The values areexpressed as mean plusmn SEM (119899 = 4) lowastlowast119875 lt 001 Adult-Rep Pversus Adult-Reg P lowastlowastlowast119875 lt 0001 prepubertal versus Adult-RepP Adult-Reg P versus Adult-Rec P
during the reproductive period and minimum values duringthe gonadal regression period (6) the amount of ARn-ir cellswas directly related to testosterone serum concentration
There are still many questions about the relations ofAR expression with the pituitary development and hormonesecretion Wu et al [12] did not observe AR in the pituitarygland during late gestation and suggested that the lack of
feedback by way of AR at pituitary levels may have importantconsequences on the fetal pituitary function More recentlyYuan et al [18] have reported in sheep that AR emerges inthe anterior pituitary before day 60 of gestation and thepercentage of the total anterior pituitary cells expressing thereceptor did not change throughout gestation In additionother studies have concluded that the fetal brain and pitu-itary are target sites for circulating androgens or androgenprecursors in fetal plasma and the increase of androgensin hypothalamic action immediately prior to birth might becritical to the timing of parturition [13] Huang and Yuan[42] have reported that AR expression reflects the functionalstatus of androgen in the pituitary of male sheep lambTheseauthors suggest that during sexual maturity the feedback ofsex hormones on the pituitary is at low level and stimulatesgonadotropin secretion
Some reproductive characteristics of female viscachassuch as polyovulation implantation and offspring birth havebeen described in previous studies Gestation lasts approx-imately 154 days an unusually long period for a rodentand it finishes in spring when the survival probabilities areideal for the mother and offspring [38 43 44] The resultsobtained in this work demonstrate AR in fetal pituitary frommidpregnancy indicating that AR are present around days75ndash80 of gestation in the pituitary cells These receptors arepotentially in position to participate in the regulation of thefetal androgen function Furthermore the analysis of thedistribution morphometry and immunostaining pattern ofAR-ir cells in the parenchyma of fetal glands suggests thatthese cells contain variable amounts of AR and they aremorphologically related to blood supply and other endocrinecells of PD parenchyma In late pregnancy of viscacha thepercentages of ARc-ir and ARn-ir cells did not change withrespect to the values found during midpregnancy Theseresults suggested that fetal pituitary AR might have anyspecific function since midgestation in viscacha
Thehypothalamic-pituitary-gonadal axis is activated dur-ing puberty when hormonalmodulation of sexual behavior isorganized [45] During male development the masculiniza-tion is given by the hormones produced by the fetal testesprimarily testosterone [46 47] The gonadal steroids feedback to the central nervous system to exert a key role inthe central control of the reproductive axis by regulatingthe release of the gonadotropin-releasing hormone (GnRH)[48] However the neurons that synthesize GnRH did notexpress AR and there are sex-steroids sensitive neuronsupstream the GnRH cells where the feedback signals ofthe steroid might be processed to the GnRH cells [49] Theneurons of hypothalamic nuclei that expressed kisspeptin(K) and neurokinin B (NKB) may sense circulating levelsof testosterone through the expression of AR [49 50] Inrodents sheep and primates K and NKB have recently beenimplicated in the control of puberty and the maintenanceof the reproductive function in adulthood [51ndash53] In thepituitary the expression of AR has been mainly observedin gonadotrophs [18 54 55] and gonadal steroid hormonesaffect the gonadotroph population in order to regulate theiractivity
International Journal of Endocrinology 9
In immature male viscachas a significant increase ofnuclear AR and an important decrease of cytoplasmic ARwere observed In prepubertal viscachas more nuclei andcytoplasm appear and there is a clear increase of ARc-ir cellsmainly in the medial region suggesting that more cells beginto express AR in preparation for sexual maturation In adultthe immunostaining is mainly nuclear and regionalized atthe caudal end of PD suggesting that AR are active in thePD cells The present results demonstrate variations in theregionalization and immunostaining pattern of AR duringsexual maturity probably in relation to the contribution ofandrogens reaching the different pituitary zones throughblood vessels Thus the nuclear AR expression in pituitaryPD of viscacha varies according to gonadal developmentand testosterone serum concentration Our results are inagreement with those found by Wood and Newman [24]suggesting that a subset of unoccupied receptors is located inthe cell cytoplasm in the absence of ligand
Bibliographic reports referring to seasonal variations ofRA are scarce Lu et al [56] have recently demonstrated astronger expression ofAR in scented glands during the breed-ing season suggesting that androgens may directly influ-ence the glandular function of the muskrats and courtshipbehavior The viscacha is a seasonal breeding rodent andprevious studies have demonstrated that testosterone acts ongonadotrophs whether directly or indirectly regulating theiractivity in the pituitary gland [28] Castrated viscachamodelshave been used to examine the withdrawal effects of circulat-ing androgens on pituitary cells mainly on the gonadotrophsdue to their involvement in the reproductive processes Inaddition castration resulted in a significant decrease in thepercentage of cells expressing AR throughout the PD [37]The present results demonstrate changes in the amount ofpituitary AR-ir cells during the annual reproductive cycleof adult male viscacha which might be due to variations inthe supply of ligands across blood irrigation The maximumvalues of AR expression were found in the reproductiveperiod and the minimum values in the regression period inrelation to androgen concentrationsThus ARmight regulatethe activity of the pituitary cells involved in the reproductivecycle of adult male Lagostomus
In conclusion our results demonstrate thatAR expressionis related to the pituitary histophysiological changes duringthe growth and sexual maturity of male viscacha Moreoverthey are likely to participate in the seasonal reproductioncontrol in adulthoodHowever the precise actionmechanismof AR in the pituitary viscacha needs further study
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Mrs A Bernardi and Mr JArroyuelo for their technical participation This work wassupported by Project 2-2614 Secretaria de Ciencia y TecnicaUniversidad Nacional de San Luis
References
[1] A K Roy Y Lavrovsky C S Song et al ldquoRegulation ofandrogen actionrdquo Vitamins amp Hormones vol 55 pp 309ndash3521999
[2] S Denayer C Helsen L Thorrez A Haelens and F ClaessensldquoThe rules of DNA recognition by the androgen receptorrdquoMolecular Endocrinology vol 24 no 5 pp 898ndash913 2010
[3] E V Jensen G L Greene L E Closs E R DeSombre and MNadji ldquoReceptors reconsidered a 20-year perspectiverdquo RecentProgress in Hormone Research vol 38 pp 1ndash40 1982
[4] B D SchanbacherM P Johnson andD J Tindall ldquoAndrogenicregulation of luteinizing hormone secretion relationship toandrogen binding in sheep pituitaryrdquo Biology of Reproductionvol 36 no 2 pp 340ndash350 1987
[5] R J Handa D L Reid and J A Resko ldquoAndrogen receptorsin brain and pituitary of female rats cyclic changes andcomparisons with the malerdquo Biology of Reproduction vol 34no 2 pp 293ndash303 1986
[6] J V A Choate and J A Resko ldquoEffects of androgen on brain andpituitary androgen receptors and LH secretion of male guineapigsrdquoThe Journal of Steroid Biochemistry andMolecular Biologyvol 59 no 3-4 pp 315ndash322 1996
[7] T J Spady R Shayya V GThackray L Ehrensberger J S Bai-ley and P L Mellon ldquoAndrogen regulates follicle-stimulatinghormone 120573 gene expression in an activin-dependent manner inimmortalized gonadotropesrdquo Molecular Endocrinology vol 18no 4 pp 925ndash940 2004
[8] P B Connolly J V A Choate and J A Resko ldquoEffects ofexogenous androgen on brain androgen receptors of the fetalrhesus monkeyrdquoNeuroendocrinology vol 59 no 3 pp 271ndash2761994
[9] J A Resko A Perkins C E Roselli J A Fitzgerald J V AChoate and F Stormshak ldquoEndocrine correlates of partnerpreference behavior in ramsrdquo Biology of Reproduction vol 55no 1 pp 120ndash126 1996
[10] S E Abdelgadir C E Roselli J V A Choate and J A ReskoldquoAndrogen receptor messenger ribonucleic acid in brains andpituitaries of male Rhesus monkeys studies on distributionhormonal control and relationship to luteinizing hormonesecretionrdquo Biology of Reproduction vol 60 no 5 pp 1251ndash12561999
[11] R W Bonsall H D Rees and R P Michael ldquoThe distributionnuclear uptake and metabolism of [3H]dihydrotestosterone inthe brain pituitary gland and genital tract of the male rhesusmonkeyrdquo Journal of Steroid Biochemistry vol 23 no 4 pp 389ndash398 1985
[12] S S Wu P W Nathanielsz and T J McDonald ldquoImmunocy-tochemical distribution of androgen receptors in the hypotha-lamus and pituitary of the fetal baboon in late gestationrdquoDevelopmental Brain Research vol 84 no 2 pp 278ndash281 1995
[13] C E Wood and M Keller-Wood ldquoOntogeny of androgenreceptor expression in the ovine fetal central nervous systemand pituitaryrdquo Neuroscience Letters vol 439 no 2 pp 153ndash1562008
[14] J V A Choate O D Slayden and J A Resko ldquoImmuno-cytochemical localization of androgen receptors in brains ofdeveloping and adult male rhesus monkeysrdquo Endocrine vol 8no 1 pp 51ndash60 1998
10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

International Journal of Endocrinology 9
In immature male viscachas a significant increase ofnuclear AR and an important decrease of cytoplasmic ARwere observed In prepubertal viscachas more nuclei andcytoplasm appear and there is a clear increase of ARc-ir cellsmainly in the medial region suggesting that more cells beginto express AR in preparation for sexual maturation In adultthe immunostaining is mainly nuclear and regionalized atthe caudal end of PD suggesting that AR are active in thePD cells The present results demonstrate variations in theregionalization and immunostaining pattern of AR duringsexual maturity probably in relation to the contribution ofandrogens reaching the different pituitary zones throughblood vessels Thus the nuclear AR expression in pituitaryPD of viscacha varies according to gonadal developmentand testosterone serum concentration Our results are inagreement with those found by Wood and Newman [24]suggesting that a subset of unoccupied receptors is located inthe cell cytoplasm in the absence of ligand
Bibliographic reports referring to seasonal variations ofRA are scarce Lu et al [56] have recently demonstrated astronger expression ofAR in scented glands during the breed-ing season suggesting that androgens may directly influ-ence the glandular function of the muskrats and courtshipbehavior The viscacha is a seasonal breeding rodent andprevious studies have demonstrated that testosterone acts ongonadotrophs whether directly or indirectly regulating theiractivity in the pituitary gland [28] Castrated viscachamodelshave been used to examine the withdrawal effects of circulat-ing androgens on pituitary cells mainly on the gonadotrophsdue to their involvement in the reproductive processes Inaddition castration resulted in a significant decrease in thepercentage of cells expressing AR throughout the PD [37]The present results demonstrate changes in the amount ofpituitary AR-ir cells during the annual reproductive cycleof adult male viscacha which might be due to variations inthe supply of ligands across blood irrigation The maximumvalues of AR expression were found in the reproductiveperiod and the minimum values in the regression period inrelation to androgen concentrationsThus ARmight regulatethe activity of the pituitary cells involved in the reproductivecycle of adult male Lagostomus
In conclusion our results demonstrate thatAR expressionis related to the pituitary histophysiological changes duringthe growth and sexual maturity of male viscacha Moreoverthey are likely to participate in the seasonal reproductioncontrol in adulthoodHowever the precise actionmechanismof AR in the pituitary viscacha needs further study
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
The authors wish to thank Mrs A Bernardi and Mr JArroyuelo for their technical participation This work wassupported by Project 2-2614 Secretaria de Ciencia y TecnicaUniversidad Nacional de San Luis
References
[1] A K Roy Y Lavrovsky C S Song et al ldquoRegulation ofandrogen actionrdquo Vitamins amp Hormones vol 55 pp 309ndash3521999
[2] S Denayer C Helsen L Thorrez A Haelens and F ClaessensldquoThe rules of DNA recognition by the androgen receptorrdquoMolecular Endocrinology vol 24 no 5 pp 898ndash913 2010
[3] E V Jensen G L Greene L E Closs E R DeSombre and MNadji ldquoReceptors reconsidered a 20-year perspectiverdquo RecentProgress in Hormone Research vol 38 pp 1ndash40 1982
[4] B D SchanbacherM P Johnson andD J Tindall ldquoAndrogenicregulation of luteinizing hormone secretion relationship toandrogen binding in sheep pituitaryrdquo Biology of Reproductionvol 36 no 2 pp 340ndash350 1987
[5] R J Handa D L Reid and J A Resko ldquoAndrogen receptorsin brain and pituitary of female rats cyclic changes andcomparisons with the malerdquo Biology of Reproduction vol 34no 2 pp 293ndash303 1986
[6] J V A Choate and J A Resko ldquoEffects of androgen on brain andpituitary androgen receptors and LH secretion of male guineapigsrdquoThe Journal of Steroid Biochemistry andMolecular Biologyvol 59 no 3-4 pp 315ndash322 1996
[7] T J Spady R Shayya V GThackray L Ehrensberger J S Bai-ley and P L Mellon ldquoAndrogen regulates follicle-stimulatinghormone 120573 gene expression in an activin-dependent manner inimmortalized gonadotropesrdquo Molecular Endocrinology vol 18no 4 pp 925ndash940 2004
[8] P B Connolly J V A Choate and J A Resko ldquoEffects ofexogenous androgen on brain androgen receptors of the fetalrhesus monkeyrdquoNeuroendocrinology vol 59 no 3 pp 271ndash2761994
[9] J A Resko A Perkins C E Roselli J A Fitzgerald J V AChoate and F Stormshak ldquoEndocrine correlates of partnerpreference behavior in ramsrdquo Biology of Reproduction vol 55no 1 pp 120ndash126 1996
[10] S E Abdelgadir C E Roselli J V A Choate and J A ReskoldquoAndrogen receptor messenger ribonucleic acid in brains andpituitaries of male Rhesus monkeys studies on distributionhormonal control and relationship to luteinizing hormonesecretionrdquo Biology of Reproduction vol 60 no 5 pp 1251ndash12561999
[11] R W Bonsall H D Rees and R P Michael ldquoThe distributionnuclear uptake and metabolism of [3H]dihydrotestosterone inthe brain pituitary gland and genital tract of the male rhesusmonkeyrdquo Journal of Steroid Biochemistry vol 23 no 4 pp 389ndash398 1985
[12] S S Wu P W Nathanielsz and T J McDonald ldquoImmunocy-tochemical distribution of androgen receptors in the hypotha-lamus and pituitary of the fetal baboon in late gestationrdquoDevelopmental Brain Research vol 84 no 2 pp 278ndash281 1995
[13] C E Wood and M Keller-Wood ldquoOntogeny of androgenreceptor expression in the ovine fetal central nervous systemand pituitaryrdquo Neuroscience Letters vol 439 no 2 pp 153ndash1562008
[14] J V A Choate O D Slayden and J A Resko ldquoImmuno-cytochemical localization of androgen receptors in brains ofdeveloping and adult male rhesus monkeysrdquo Endocrine vol 8no 1 pp 51ndash60 1998
10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 International Journal of Endocrinology
[15] J E Woods C M Blandin and R C Thommes ldquoOncogenyof immunocytochemically demonstrable androgen- and andro-gen receptor-containing cells in the hypothalamus and ade-nohypophysis of the chick embryordquo Growth Development andAging vol 58 no 1 pp 21ndash31 1994
[16] A Crocoll C C Zhu A C B Cato and M Blum ldquoExpressionof androgen receptor mRNA during mouse embryogenesisrdquoMechanisms of Development vol 72 no 1-2 pp 175ndash178 1998
[17] O Naess E Haug A Attramadal A Aakvaag V Hansson andF French ldquoAndrogen receptors in the anterior pituitary andcentral nervous system of the androgen lsquoinsensitiversquo (Tfm) ratcorrelation between receptor binding effects of androgens ongonadotropin secretionrdquo Endocrinology vol 99 no 5 pp 1295ndash1303 1976
[18] X J Yuan Y Q He J L Liu H S Luo J H Zhang and SCui ldquoExpression of androgen receptor and its co-localizationwith estrogen receptor-alpha in the developing pituitary glandof sheep fetusrdquo Histochemistry and Cell Biology vol 127 no 4pp 423ndash432 2007
[19] R J Nelson D L Denlinger and D E Somers PhotoperiodismTheBiological Calender OxfordUniversity PressNewYorkNYUSA 2010
[20] Y T ShaoMArvidsson S Trombley RW SchulzM Schmitzand B Borg ldquoAndrogen feedback effects on LH and FSH andphotoperiodic control of reproduction in male three-spinedsticklebacks Gasterosteus aculeatusrdquo General and ComparativeEndocrinology vol 182 pp 16ndash23 2013
[21] L Z Garamszegi M Eens S Hurtrez-Bousses and A PMoslashller ldquoTestosterone testes size and mating success in birdsa comparative studyrdquo Hormones amp Behavior vol 47 no 4 pp389ndash409 2005
[22] A Leska J Kiezun B Kaminska and L Dusza ldquoSeasonalchanges in the expression of the androgen receptor in the testesof the domestic goose (Anser anser f domestica)rdquo General andComparative Endocrinology vol 179 no 1 pp 63ndash70 2012
[23] E L Bittman D A Ehrlich J L Ogdahl and A E Jet-ton ldquoPhotoperiod and testosterone regulate androgen receptorimmunostaining in the Siberian hamster brainrdquo Biology ofReproduction vol 69 no 3 pp 876ndash884 2003
[24] R I Wood and S W Newman ldquoIntracellular partitioning ofandrogen receptor immunoreactivity in the brain of the maleSyrian hamster effects of castration and steroid replacementrdquoJournal of Neurobiology vol 24 no 7 pp 925ndash938 1993
[25] M J Tetel T C Ungar B Hassan and E L BittmanldquoPhotoperiodic regulation of androgen receptor and steroidreceptor coactivator-1 in Siberian hamster brainrdquo MolecularBrain Research vol 131 no 1-2 pp 79ndash87 2004
[26] S Domınguez R S Piezzi L Scardapane and J A Guzman ldquoAlight and electron microscopic study of the pineal gland of theviscacha (Lagostomus maximum maximus)rdquo Journal of PinealResearch vol 4 no 2 pp 211ndash219 1987
[27] L Fuentes M Moslashller E Munoz C Calderon and L PelzerldquoSeasonal variations in the expression of the mRNA encoding1205731-adrenoceptor and AA-NAT enzyme and in the AA-NATactivity in the pineal gland of vizcacha (Lagostomus max-imus maximus)mdashcorrelation with serum melatoninrdquo BiologicalRhythm Research vol 34 no 2 pp 193ndash206 2003
[28] V Filippa A Penissi and F Mohamed ldquoSeasonal variationsof gonadotropins in the pars distalis male viscacha pituitaryEffect of chronic melatonin treatmentrdquo European Journal ofHistochemistry vol 49 no 3 pp 291ndash300 2005
[29] V Filippa andFMohamed ldquoACTHcells of pituitary pars distalisof viscacha (Lagostomus maximus maximus) immunohisto-chemical study in relation to season sex and growthrdquo Generaland Comparative Endocrinology vol 146 no 3 pp 217ndash2252006
[30] V Filippa and F Mohamed ldquoImmunohistochemical studyof somatotrophs in pituitary pars distalis of male viscacha(Lagostomus maximus maximus) in relation to the gonadalactivityrdquoCells Tissues Organs vol 184 no 3-4 pp 188ndash197 2007
[31] V Filippa and F Mohamed ldquoImmunohistochemical and mor-phometric study of pituitary pars distalis thyrotrophs of maleviscacha (Lagostomus maximus maximus) seasonal variationsand effect of melatonin and castrationrdquoThe Anatomical Recordvol 291 no 4 pp 400ndash409 2008
[32] V Filippa and FMohamed ldquoMorphological andmorphometricchanges of pituitary lactotrophs of viscacha (Lagostomus max-imus maximus) in relation to reproductive cycle age and sexrdquoThe Anatomical Record vol 293 no 1 pp 150ndash161 2010
[33] E Munoz T Fogal S Dominguez L Scardapane J Guzmanand R S Piezzi ldquoSeasonal changes of the Leydig cells ofviscacha (Lagostomus maximus maximus) A light and electronmicroscopy studyrdquo Tissue and Cell vol 29 no 1 pp 119ndash1281997
[34] E M Munoz T Fogal S Dominguez L Scardapane and R SPiezzi ldquoUltrastructural and morphometric study of the sertolicell of the viscacha (Lagostomus maximus maximus) during theannual reproductive cyclerdquoTheAnatomical Record vol 262 no2 pp 176ndash185 2001
[35] E M Chaves C Aguilera-Merlo A Cruceno et al ldquoSeasonalmorphological variations and age-related changes of the sem-inal vesicle of viscacha (Lagostomus maximus maximus) anultrastructural and immunohistochemical studyrdquoThe Anatom-ical Record vol 295 no 5 pp 886ndash895 2012
[36] A M Cruceno J C de Rosas M Foscolo et al ldquoEffects ofphotoperiod one epididymal and sperm morphology in a wildrodent the viscacha (Lagostomus maximus maximus)rdquo ISRNAnatomy vol 2013 Article ID 128921 10 pages 2013
[37] V Filippa D Godoy E Perez and F Mohamed ldquoEffects ofcastration on androgen receptors and gonadotropins in thepituitary of adult male viscachasrdquo Reproduction Fertility andDevelopment vol 26 no 7 pp 991ndash1000 2014
[38] V Filippa and F Mohamed ldquoThe pituitary of non-pregnant andpregnant viscachas (Lagostomus maximus maximus) a com-parative study by immunohistochemistry and morphometricanalysisrdquo Zoology vol 113 no 6 pp 361ndash372 2010
[39] N P Leopardo F Jensen M A Willis M B Espinosa andA D Vitullo ldquoThe developing ovary of the South Americanplains vizcacha Lagostomus maximus (Mammalia Rodentia)massive proliferation with no sign of apoptosis-mediated germcell attritionrdquo Reproduction vol 141 no 5 pp 633ndash641 2011
[40] A C Llanos and J A Crespo ldquoEcologıa de la vizcacha(Lagostomus maximus maximus) en el nordeste de la provinciade Entre Rıosrdquo Revista de Investigaciones Agrıcolas Extra NuevaSerie no 10 pp 5ndash95 1954
[41] V Filippa M Acosta and F Mohamed ldquoCellular associationsof pituitary gonadotrophs in a rodent (Lagostomus maximusmaximus) with photoperiod-dependent reproductionrdquo Tissueand Cell vol 44 no 6 pp 351ndash357 2012
[42] L B Huang and X J Yuan ldquoExpression of androgen receptorand estrogen receptor-alpha in the developing pituitary glandof male sheep lambrdquo Animal Reproduction Science vol 127 no3-4 pp 164ndash168 2011
International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

International Journal of Endocrinology 11
[43] B J Weir ldquoThe reproductive organs of the female plainsviscacha Lagostomus maximusrdquo Journal of Reproduction andFertility vol 25 no 3 pp 365ndash373 1971
[44] D L Keefe and F W Turek ldquoCircadian time keeping processesin mammalian reproductionrdquo Oxford Reviews of ReproductiveBiology vol 7 pp 346ndash400 1985
[45] B Shannon Z Stephens and K Wallen ldquoEnvironmental andsocial influences on neuroendocrine puberty and behaviorin macaques and other nonhuman primatesrdquo Hormones andBehavior vol 64 no 2 pp 226ndash239 2013
[46] D J MacLeod R M Sharpe M Welsh et al ldquoAndrogen actionin themasculinization programming window and developmentof male reproductive organsrdquo International Journal of Androl-ogy vol 33 no 2 pp 279ndash287 2010
[47] T NrsquoTumba-Byn D Moison M Lacroix et al ldquoDifferentialeffects of bisphenol A and diethylstilbestrol on human rat andmouse fetal Leydig cell functionrdquo PLoS ONE vol 7 no 12Article ID e51579 2012
[48] G Fink ldquoNeuroendocrine regulation of pituitary functiongeneral principlesrdquo in Neuroendocrinology in Physiology andMedicine P M Conn and M E Freeman Eds pp 107ndash134Humana Press Totowa NJ USA 2000
[49] A S Kauffman ldquoGonadal and nongonadal regulation of sexdifferences in hypothalamic kiss1 neuronesrdquo Journal ofNeuroen-docrinology vol 22 no 7 pp 682ndash691 2010
[50] P Ciofi O C Lapirot and G Tramu ldquoAn androgen-dependentsexual dimorphism visible at puberty in the rat hypothalamusrdquoNeuroscience vol 146 no 2 pp 630ndash642 2007
[51] N E Rance and T R Bruce ldquoNeurokinin B gene expressionis increased in the arcuate nucleus of ovariectomized ratsrdquoNeuroendocrinology vol 60 no 4 pp 337ndash345 1994
[52] R L Goodman M N Lehman J T Smith et al ldquoKisspeptinneurons in the arcuate nucleus of the ewe express both dynor-phin A and neurokinin Brdquo Endocrinology vol 148 no 12 pp5752ndash5760 2007
[53] V M Navarro ldquoInteractions between kisspeptins and neu-rokinin BrdquoAdvances in Experimental Medicine and Biology vol784 pp 325ndash347 2013
[54] N Kimura A Mizokami T Oonuma H Sasano and HNagura ldquoImmunocytochemical localization of androgen recep-tor with polyclonal antibody in paraffin-embedded humantissuesrdquoThe Journal of Histochemistry amp Cytochemistry vol 41no 5 pp 671ndash678 1993
[55] B W Scheithauer K Kovacs S Zorludemir R V LloydS Erdogan and J Slezak ldquoImmunoexpression of androgenreceptor in the nontumorous pituitary and in adenomasrdquoEndocrine Pathology vol 19 no 1 pp 27ndash33 2008
[56] L Lu S Liu Q Li et al ldquoSeasonal expression of androgenreceptor in scented gland of muskrat (Ondatra zibethicus)rdquoGeneral and Comparative Endocrinology vol 204 pp 1ndash7 2014
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom