reduction inorganic compounds withmolecular …jb.asm.org/content/84/4/647.full.pdf · reduction of...
TRANSCRIPT

REDUCTION OF INORGANIC COMPOUNDS WITH MOLECULARHYDROGEN BY MICROCOCCUS LACTILYTICUS
I. STOICHIOMETRY WITH COMPOUNDS OF ARSENIC, SELENIUM, TELLURIUM,TRANSITION AND OTHER ELEMENTS
C. A. WOOLFOLK AND H. R. WHITELEYDepartnment of Microbiology, School of Medicine, University of Washington, Seattle, Washington
Received for publication April 30, 1962
ABSTRACT
WOOLFOLK, C. A. (University of Washington,Seattle) AND H. R. WHITELEY. Reduction ofinorganic compounds with molecular hydrogenby Micrococcus lactilyticus. I. Stoichiometry withcompounds of arsenic, selenium, tellurium,transition and other elements. J. Bacteriol.84:647-658. 1962.-Extracts of Micrococcuslactilyticus (Veillonella alcalescens) oxidize molec-ular hydrogen at the expense of certain com-pounds of arsenic, bismuth, selenium, tellurium,lead, thallium, vanadium, manganese, iron,copper, molybdenum, tungsten, osmium, ruthe-nium, gold, silver, and uranium, as well asmolecular oxygen. Chemical and manometricdata indicate that the following reductions areessentially quantitative: arsenate to arsenite,pentavalent and trivalent bismuth to the freeelement, selenite via elemental selenium toselenide, tellurate and tellurite to tellurium, leaddioxide and manganese dioxide to the divalentstate, ferric to ferrous iron, osmium tetroxide toosmate ion, osmium dioxide and trivalent osmiumto the metal, uranyl uranium to the tetravalentstate, vanadate to the level of vanadyl, and poly-molybdate ions to molybdenum blues with anaverage valence for molybdenum of +5. Theresults of a study of certain other hydrogenase-containing bacteria with respect to their abilityto carry out some of the same reactions are alsopresented.
The reduction of inorganic compounds in anatmosphere of hydrogen by cell suspensions andby extracts containing hydrogenase has beenreported for many bacteria. Thus, the hydro-genase-coupled reductions of inorganic sulfurcompounds by Desulfovibrio (Postgate, 1959;Peck, 1959) and of inorganic nitrogen com-
pounds by a variety of bacteria (Nason andTakahashi, 1958; Gest, 1954) have beenthoroughly investigated. The reduction of oxygenat the expense of molecular hydrogen ("Knallgasreaction") as a source of energy for the auto-trophic growth of members of the genus Hydro-genomonas has been studied in detail (Packer,1958). The latter reaction is also known to occurin a variety of other bacteria (Gest, 1954),including the obligate anaerobe, Desulfovibrio(Postgate, 1956).Evidence has also been presented for the
biological reduction of a number of other in-organic compounds, although molecular hydrogenhas not been shown to serve as the electron donor.For example, the production of elementalselenium (Levine, 1925; Zalokar, 1953) andtellurium (Corper, 1915; Morton and Anderson,1941), of molybdenum blue (Jan, 1939; Marchaland Girard, 1948), of arsenite (Green, 1918), andof phosphite and hypophosphite (Barrenscheenand Beckh-Widmanstetter, 1923; Rudakov, 1927)by microorganisms growing in media containingselenate, tellurite, molybdate, arsenate, orphosphate, respectively, has been reported.Reduced methylated derivatives of arsenate,arsenite, tellurite, selenate, and selenite can beproduced by various molds during growth(Challenger, 1951). In addition, the reduction ofmanganese dioxide (Mann and Quastel, 1946)and ferric iron (Bromfield, 1954) by soil bacteriahas been studied.The reductions of tellurite (Terai, Kamahora,
and Yamamura, 1958) and selenite (Falcone andNickerson, 1960) have been observed in cell-freepreparations of Mycobacterium avium andSaccharomyces cerevisiae, respectively, and someof the properties of the enzymes mediating thesereactions have been investigated. It has also beenshown that vanadate, selenite, and phospho-
647
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

WOOLFOLK AND WHITELEY
molybdate can oxidize reduced cytochrome C3obtained from Desulfovibrio (Ishimoto et al.,1957).A systematic survey of inorganic compounds
has disclosed that cell-free extracts of Micro-coccus lactilyticus (Foubert and Douglas, 1948;classified as Veillonella alcalescens Pr6vot inBergey's Manual, 7th ed., 1957) are capable ofutilizing molecular hydrogen for the reductionof a wide variety of inorganic compounds(Woolfolk, Ordal, and Whiteley, 1959; Woolfolk,1960; Woolfolk and Whiteley, 1961). This paperpresents data on the stoichiometry of reductionof certain of these compounds, on the ionicspecies used as substrates, and on the rates ofthese reductions. Certain other bacteria con-taining hydrogenase are compared with respectto their ability to reduce some of these samecompounds.
MATERIALS AND METHODS
Cultivation of bacteria and preparation ofextracts. Bacteria were grown in the followingmedia: 11. lactilyticus, in a complex mediumcontaining lactate (Whiteley and Douglas, 1951);Mlicrococcus aerogenes (Peptococcus aerogenes),in a peptone-yeast extract medium with glu-tamate (Whiteley, 1957); Escherichia coli strain Bon the basal medium of McNall and Atkinson(1956) containing 5 X 10-s M (NH4)2SO4; Azoto-bacter vinelandii, in Burk's medium as used byNewton, Wilson, and Burris (1953); Desulfovibriodesulfuricans, in a medium similar to that ofPostgate (1951) to which 0.4%O sodium lactatewas added; Clostridium pasteurianum, in themedium of Carnahan and Castle (1958) usingNH4Cl as a nitrogen source; and Rhodomicrobiumvannielii, in the medium of Murray and Douglas(1950) under illumination.
Cells were harvested by centrifugation, washedonce or twice with distilled water, suspended inone to two volumes of water, and exposed tosonic oscillation for 20 to 45 min in a Raytheon10-kc sonic oscillator. The resulting preparationwas centrifuged in the cold at 25,000 X g for20 min and the supernatant fraction was used asa "crude extract." Protein was determined by themethod of Lowry (Layne, 1957).
Preparation of substrates. Sodium phosphitewas obtained by the hydrolysis of phosphorustrichloride in water followed by neutralization ofthe solution with sodium hydroxide. Sodium
hypophosphate (Na2H2P206) was prepared bythe method of Leininsen and Chulski (1953).Potassium tellurate (chromatographically free oftellurite) was prepared by treatment of potassiumtellurite with hydrogen peroxide. Ferric hydroxideand cupric hydroxide suspensions were obtainedby neutralization of the appropriate chlorides.Commercial manganese dioxide and lead dioxidewere reduced by extracts of M. lactilyticus underhydrogen, but higher specific activities could beobtained with suspensions containing highlydispersed particles of these substances. Man-ganese dioxide and lead dioxide suspensions wereobtained by treatment of potassium perman-ganate or lead acetate with sodium sulfite orsodium hypochlorite solutions, respectively. Thesuspensions were purified by repeated centrifuga-tion and resuspension. Colloidal elementalselenium was prepared by the action of hy-droxylamine hydrochloride on sodium selenitesolutions in the presence of 5% gum arabic, andwas purified by repeated centrifugation. Bismuthhydroxide [Bi(OH)3] and bismuthyl hydroxide[BiO(OH)] were obtained by addition of excesssodium hydroxide to bismuth subcarbonatesuspensions at room temperature and 100 C,respectively (Charlot, 1954). Other compoundstested as substrates were obtained from com-mercial sources.
Manometric procedures. The reduction ofinorganic compounds was followed by measuringthe uptake of hydrogen in a conventional War-burg apparatus at 30 C. The vessels were flushedwith hydrogen from which oxygen had beenremoved by passage through a hot copper column.Reaction mixtures having a total volume of2.2 ml routinely contained from 5 to 50 j,moles ofsubstrate in one side arm, and 150 ,umoles oftris(hydroxymethyl)aminomethane buffer (pH7.0) and 1 ml of extract containing approximately30 mg of protein in the main compartment. In theexperiments where stoichiometric relationshipswere determined, the substrate was incubatedwith the reaction mixture until the uptake ofhydrogen gas had ceased, and the vessel contentswere then analyzed for the end products.When molecular oxygen was used as the sub-
strate, one side arm gas vent was replaced with aserum stopper to permit the introduction ofoxygen into the vessel by means of a hypodermicsyringe. The stoichiometry of oxygen uptakeduring the utilization of both oxygen and hy-
648 J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 84, 1962 INORGANIC COMPOUND REDUCTION BY M. LACTILYTICUS
drogen was determined by the method of Schatzand Bovell (1952).
Analytical methods. For the determination ofarsenite, proteins were removed from the reactionmixture by precipitation with trichloroaceticacid, and the supernatant fraction was neutralizedwith sodium bicarbonate and titrated with iodinein excess bicarbonate. Total arsenate plus arsenitewas estimated colorimetrically by the molyb-denum blue method of Sandell (1944), withhydrazine as the reducing agent. Arsine wasdetermined by flushing a reaction vessel withhydrogen and trapping the flushed material in anabsorption vessel as described by Sandell (1944).The contents of the absorption vessel were thenanalyzed by the molybdenum blue method.
Solutions of selenite and tellurite and suspen-sions of lead dioxide were determined quantita-tively by titration with standard permanganate(Charlot and Bezier, 1957). Osmium trichloridesolutions were similarly estimated, on theassumption that the osmium was oxidized to thetetroxide.
Elemental selenium and tellurium were deter-mined colorimetrically after dissolution inconcentrated sulfuric acid (Wiberley et al., 1953),using selenite and tellurite solutions (determinedas above) as standards. Tellurate could also bedetermined by this method after reduction tothe element with acidified stannous chloride.Hydrogen selenide produced by the reduction ofselenite and selenium was absorbed by additionof cadmium chloride to the center well of theWarburg vessel as described by Postgate (1951)for hydrogen sulfide. After the enzymatic reactionwith selenium or selenite had gone to completion,0.5 ml of 5 N H2SO4 was added to the reactionmixture from one of the side arms to decomposemetal selenides that might have precipitated inthe reaction mixture, and the vessel was incu-bated for an additional 30 min. The contents of thecenter well were then oxidized with excesspermanganate in 6 N HCl. After 10 min, excessacidified stannous chloride was added, andelemental selenium, and hence the selenideformed during the reduction, was then determinedcolorimetrically as above.To measure vanadyl vanadium, excess iodine
and sodium bicarbonate were added to samples ofreaction mixtures, which were then assayed withstandard arsenite solution. Hydroxide producedduring the enzymatic reduction of molybdate and
phosphotungstate was measured by addingsufficient standard acid to achieve the original pHvalue of the reaction mixture. Molybdenumblues formed by the reduction of phosphomolyb-date complexes can be distinguished spectro-photometrically from those formed by reductionof molybdate per se. This method was used forthe characterization of the product formed by M.lactilyticus from molybdate.
Arsenate and arsenite were separated bychromatography of reaction mixtures on paper,using the solvent system of Yamaguchi (1953).Ammoniacal silver nitrate was used as a spray tolocate these compounds as well as phosphate.Phosphate, phosphite, and hypophosphite wereseparated and detected by paper chromatog-raphy, as described by Bonnin and Sue (1952).Vanadyl vanadium was identified as the productof vanadate reduction by the chromatographicmethod of Stevens (1956), using 8-hydroxy-quinoline in 20% citric acid to develop thechromatograms. Paper chromatography was alsoused to determine that tellurate preparationswere free of tellurite (Ghosh Mazumdar andLederer, 1957).
RESULTS
Reduction of compounds of the nitrogen family ofelements. Extracts of Ml. lactilyticus are capableof reducing nitrate, nitrite, hydroxylamine, andhydrazine to ammonia at the expense of molecu-lar hydrogen (Woolfolk et al., 1959). In view ofthese reactions, it was considered of interest totest the ability of this organism to reduce com-pounds of arsenic, phosphorus, antimony, andbismuth, all members of the nitrogen family ofelements: the results of these investigations arediscussed here. Information on the stoichiometryand pathway of reduction of the nitrogen com-pounds will be presented in a separate communi-cation.
Arsenate, but not p-arsanilic acid or arsenite,can be reduced in a hydrogen atmosphere byextracts of M. lactilyticus. One mole of arsenite(identified by paper chromatography) wasproduced per mole of hydrogen taken up (Table1); the total concentration of arsenate plusarsenite remained constant during the reduction,and arsine could not be detected as an endproduct.Sodium bismuthate was reduced with an
uptake of hydrogen (Table 1), and a dark brown
649
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
WOOLFOLK AND WHITELEY
TABLE 1. Reduction of compounds of arsenic andbismuth by Micrococcus lactilyticus
SubstrateAmount Hydrogen Arseniteadded uptake
pmoles pmoles pmoles
Arsenate ......... 10.0 10.2 9.62Bismuthate .... . 10.0 25.4Bismuthyl ........ 10.0 16.7
TABLE 2. Reduction of molecular oxygen andcompounds of selenium and tellurium by
Micrococcus lactilyticus
Substrate adAmount H2 Selen- Sele- Tellu-Substrate added uptake ium nide rium
;imoles jimoles umoles jumoles pumoles
Oxygen ........... 6.5 12.8Selenite..... 7.8 14.8 7.6 - -Selenite ....... 7.8 23.8 7.6Selenium..... 8.6 8.5 8.6Tellurate ..... 2.5 7.3 2.6Tellurite ......... 6.5 13.4 - 6.7
suspended material, probably elemental bismuth,accumulated. A similar reaction was observedwhen a suspension of bismuthyl chloride was
added as a substrate. Preparations of whitebismuth hydroxide [Bi(OH),] and yellow bis-muthyl hydroxide [BiO(OH)] were also reduced toelemental bismuth; however, they could not beused for quantitative determinations because ofthe variable composition of the bismuth hy-droxide preparations. Although the initial rate ofbismuthate reduction was in excess of thatobserved with bismuth hydroxide (Table 5), therate of the reduction of the latter compound was
consistent with its formation as an intermediatein the conversion of bismuthate to bismuth.Bismuthyl hydroxide, bismuthyl chloride, andbismuth subcarbonate (bismuthyl carbonate), on
the other hand, are reduced much more slowly,probably owing to the limited solubilities of thesecompounds or the state of their dispersion.Antimonate and antimonyl compounds could notserve as substrates for reduction by M. lactilyticus.
There have been several reports (Barrenscheenand Beckh-Widmanstetter, 1923; Rudakov, 1927)indicating the reduction of inorganic phosphoruscompounds by bacteria. Orthophosphate, pyro-
phosphate, tetrametaphosphate, insoluble meta-phosphate, phosphite, hypophosphite, and hypo-phosphate (P2068) were tested as substrates with
hydrogen and extracts of M. lactilyticus. Utiliza-tion of hydrogen was not observed with thesecompounds, and examination of reaction mixturesby paper chromatography did not give anyevidence of reduction. It should be noted that,except for hypophosphate, the reduction of thesephosphorus compounds would not be expected onthe basis of thermodynamic considerations.Reduction of compounds of the oxygen family of
elements. The introduction of oxygen intoreaction vessels containing hydrogen and M.lactilyticus extract resulted in a rapid uptake ofgas (Table 2). No oxygen uptake was observedwhen nitrogen was substituted for hydrogen.
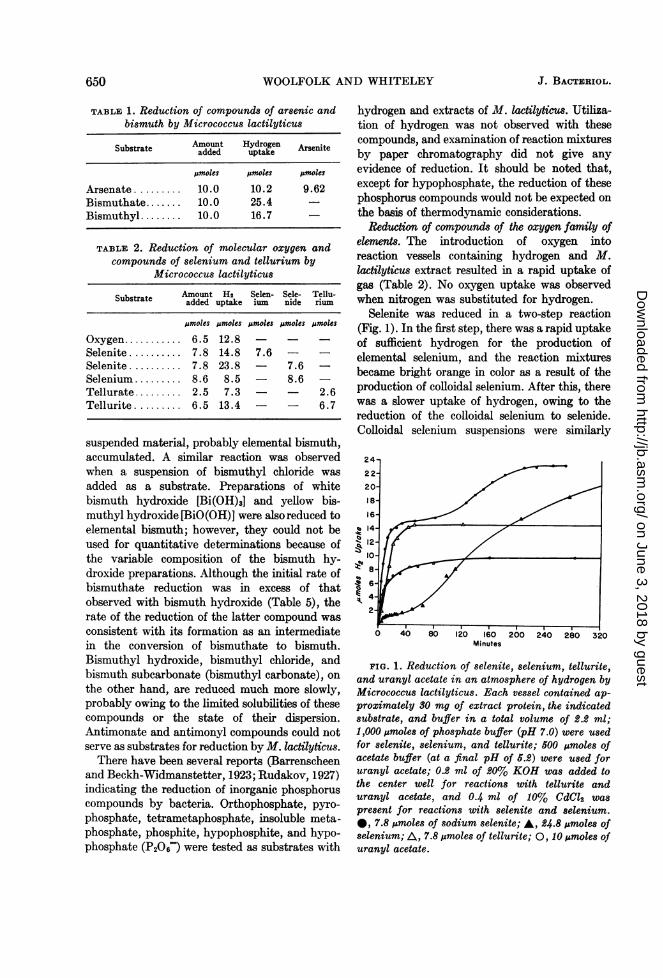
Selenite was reduced in a two-step reaction(Fig. 1). In the first step, there was a rapid uptakeof sufficient hydrogen for the production ofelemental selenium, and the reaction mixturesbecame bright orange in color as a result of theproduction of colloidal selenium. After this, therewas a slower uptake of hydrogen, owing to thereduction of the colloidal selenium to selenide.Colloidal selenium suspensions were similarly
24-
22-
20-
18-
16
q~14-12-
0 40 80 120 160 200 240 280 320Minutes
FIG. 1. Reduction of selenite, selenium, tellurite,and uranyl acetate in an atmosphere of hydrogen byMicrococcus lactilyticus. Each vessel contained ap-proximately 30 mg of extract protein, the indicatedsubstrate, and buffer in a total volume of 2.2 ml;1,000/,moles of phosphate buffer (pH 7.0) were usedfor selenite, selenium, and tellurite; 500 jumoles ofacetate buffer (at a final pH of 5.2) were used foruranyl acetate; 0.2 ml of 20%1 KOH was added tothe center well for reactions with tellurite anduranyl acetate, and 0.4 ml of 10% CdCl2 waspresent for reactions with selenite and selenium.*, 7.8 ;&moles of sodium selenite; A, 24.8 ,umoles ofselenium; A, 7.8,umoles of tellurite; 0, 10 ,umoles ofuranyl acetate.
650 J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 84, 1962 INORGANIC COMPOUND REDUCTION BY M. LACTILYTICUS
reduced to selenide (Fig. 1). The amountsof selenium formed during the first step of thisreaction, and the amounts of selenide ultimatelyproduced from selenite and colloidal selenium,are shown in Table 2. Selenate was not reduced.A number of sulfur compounds are reduced byextracts of M. lactilyticus; these data are pre-sented in the accompanying paper (Woolfolk,1962).
Tellurite (Fig. 1) and tellurate were convertedto dark brown colloidal tellurium, which was notreduced further. The quantities of hydrogenutilized and the amounts of tellurium formed withthese substrates are shown in Table 2. A com-parison of the specific activities with tellurateand tellurite (Table 5) suggests that tellurate isreduced to tellurium via the rate determiningformation of tellurite.
Reduction of compounds of the transitionelements. Vanadate may also serve as a substratefor reduction; the amount of hydrogen taken up(Table 3) was sufficient to account for theproduction of vanadyl compounds. A substancehaving the same mobility as vanadyl chloride wasdetected as a product of the reduction, andtitrimetric analysis indicated a quantitativereduction of vanadate to vanadyl (Table 3). Thereaction mixtures became light blue-green duringthe reduction, probably because of the formationof vanadyl hydroxide.When molybdate was added as a substrate in
reaction mixtures containing phosphate buffer(pH 5.5), there was a rapid uptake of sufficientmolecular hydrogen (Table 3) for reduction ofmolybdate to the pentavalent state. This re-action was accompanied by the formation of ablue color in the reaction mixtures, suggestingthat molybdate was reduced to a molybdenumblue compound. In general, the molybdenum in
TABLE 3. Reduction of vanadate, molybdate, andphosphotungstic acid by Micrococcus
lactilyticus
Substrate Amount H2 uptake Vanadyl Hydroxide
jmoles Mnmoles Amoles ;umoles
Vanadate.. 42.8 23.4 44.0Molybdate ...... 20 . Oa 10.6 - 19.8Phosphotungstic
acid.......... 10.0 2.6 2.9
aExpressed as MoO47.
TABLE 4. Reduction of compounds of osmium, lead,manganese, iron, and uranium by
Micrococcus lactilyticus
Substrate Amount H2 uptake
pmoles ,mol6s
Osmium tetroxide........... 15.6 12.0Osmium dioxide ............ 12.7 25.0Osmium (+3) chloride...... 7.3 10.7Lead dioxide ............... 6.25 6.15Manganese dioxide.......... 10.0 10.0Ferric hydroxide ............ 10.0 4.5Ferricyanide ................ 10.0 4.7Uranyl hydroxide........... 10.0 9.8
these uncharged complex oxides (molybdenumblues) has an average valence between +5 and+6, although several compounds of this typeare known with an average valence for molyb-denum of exactly +5 (Glemser and Lutz, 1947).If a less acid pH is used (i.e., 6.5), reactionmixtures turn from blue to red brown, probablybecause of the decomposition of molybdenumblue to molybdenyl hydroxide (Glemser andLutz, 1951).The formation of molybdenum blue was
probably due to the reduction of molybdate orisopolymolybdate ions, and not to the reductionof phosphomolybdate complexes. This con-clusion receives support from the fact thatphosphomolybdate complexes are stable only atmore acid pH values than those used in theenzymatic experiments. Furthermore, the spectraof molybdenum blues formed from molybdateproper and from phosphomolybdate are known todiffer (Charlot and Bezier, 1957), and thespectrum of the molybdenum blue formed by M.lactilyticus resembles the former.
Phosphotungstic acid was reduced by extractsof M. lactilyticus, although only a small amountof hydrogen was utilized (Table 3). This reduc-tion was also accompanied by the formation of ablue color (probably a tungsten blue) but thereaction was not investigated further.Osmium tetroxide could also serve as a sub-
strate for reduction (Table 4). The absence of theblack color characteristic of osmium dioxideindicates that this compound was not formed asthe end product, and the quantity of hydrogentaken up suggests that osmate ion was produced.Furthermore, osmium dioxide (added as a
651
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
WOOLFOLK AND WHITELEY
suspension) was reduced to elemental osmium, aswas trivalent osmium chloride.
Ferric hydroxide in suspension was convertedto ferrous hydroxide with an uptake of hydrogen,and ferricyanide was similarly reduced to ferro-cyanide (Table 4). Suspensions of manganesedioxide were reduced with the uptake of astoichiometric amount of hydrogen for theproduction of divalent manganese.Small uptakes of hydrogen were noted when
silver phosphate, ruthenium hydroxide (+3), orauric gold were the substrates; orange-coloredcolloidal silver, divalent ruthenium, and colloidalgold, respectively, were formed. Similarly, cuprichydroxide was reduced to cuprous hydroxide(hydrogen uptake only 70% of theoretical).
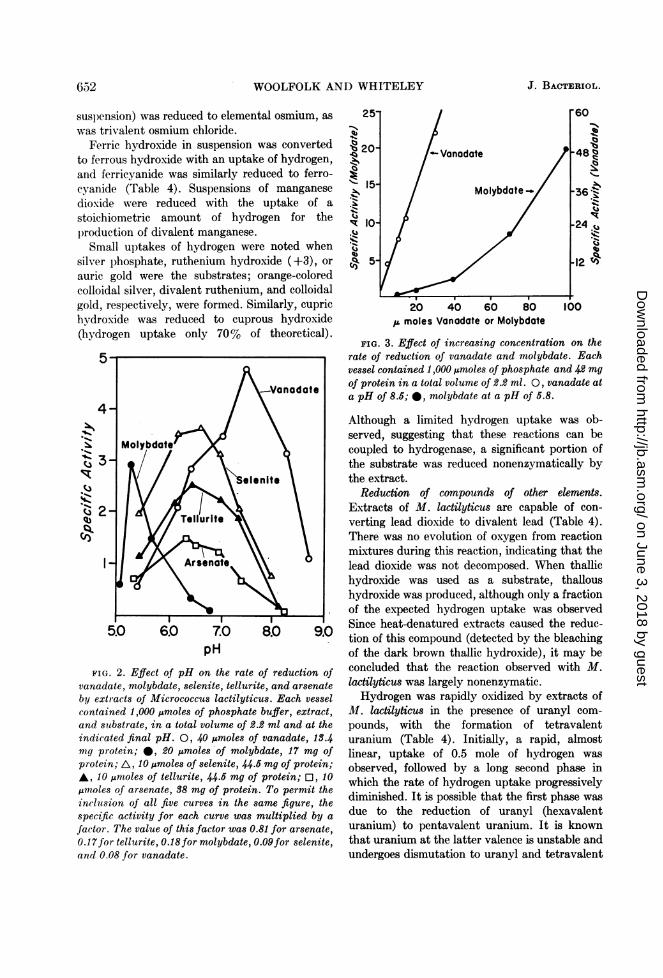
7.0
pH
FIG. 2. Effect of pH on the rate of reduction ofvanadate, molybdate, selenite, tellurite, and arsenateby extracts of Micrococcus lactilyticus. Each vesselcontained 1,000 ,umoles of phosphate buffer, extract,and sutbstrate, in a total volume of 2.2 ml and at theindicated final pH. 0, 40 ,umoles of vanadate, 13.4mg protein; 0, 20,umoles of molybdate, 17 mg ofprotein; A, 10 ,umoles of selenite, 44.5 mg of protein;A, 10 ,umoles of tellurite, 44.5 mg of protein; O, 10,umoles of arsenate, 38 mg of protein. To permit theinclusion of all five curves in the same figure, thespecific activity for each curve was multiplied by a
factor. The value of this factor was 0.81 for arsenate,0.17for tellurite, 0.18for molybdate, 0.09for selenite,and 0.08 for vanadate.
20 40 60 80fz moles Vanodate or Molybdote
S.
FIG. 3. Effect of increasing concentration on therate of reduction of vanadate and molybdate. Eachvessel contained 1,000 j,moles of phosphate and 42 mgof protein in a total volume of 2.2 ml. 0, vanadate ata pH of 8.5; 0, molybdate at a pH of 5.8.
Although a limited hydrogen uptake was ob-served, suggesting that these reactions can becoupled to hydrogenase, a significant portion ofthe substrate was reduced nonenzymatically bythe extract.
Reduction of compounds of other elements.Extracts of M. lactilyticus are capable of con-verting lead dioxide to divalent lead (Table 4).There was no evolution of oxygen from reactionmixtures during this reaction, indicating that thelead dioxide was not decomposed. When thallichydroxide was used as a substrate, thalloushydroxide was produced, although only a fractionof the expected hydrogen uptake was observedSince heat-denatured extracts caused the reduc-tion of this compound (detected by the bleachingof the dark brown thallic hydroxide), it may beconcluded that the reaction observed with M.lactilyticus was largely nonenzymatic.Hydrogen was rapidly oxidized by extracts of
AI. lactilyticus in the presence of uranyl com-pounds, with the formation of tetravalenturanium (Table 4). Initially, a rapid, almostlinear, uptake of 0.5 mole of hydrogen wasobserved, followed by a long second phase inwhich the rate of hydrogen uptake progressivelydiminished. It is possible that the first phase wasdue to the reduction of uranyl (hexavalenturanium) to pentavalent uranium. It is knownthat uranium at the latter valence is unstable andundergoes dismutation to uranyl and tetravalent
652 J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 84, 1962 INORGANIC COMPOUND REDUCTION BY M. LACTILYTICUS
uranium. The second step of hydrogen uptakemay be the result of the reduction of uranylproduced by this dismutation.
Kinetics of the reduction of a number of inorganicions. The effect of pH on the rate of reduction ofarsenate, selenite, tellurite, vanadate, andmolybdate is shown in Fig. 2. It is of interest thatthe reactions with arsenate, selenite, and telluritehad pH optima of approximately 6.5 to 7.0,and declined in rate to negligible values when thepH value approached 8.0. These ions have pKavalues of approximately 6.8 to 7.7, correspondingto the formation of the divalent ions from themonovalent ions. It is possible that the actualionic species reduced are the monovalent ions, andthat the activities decline with increasing pH asa result of the diminishing concentration of thisform of the substrate. On the other hand,vanadate, which does not have a pK in thisregion, was reduced readily at pH 8.0, indicatingthat the decline in activities with the other sub-strates was not due to inactivation of hydro-genase or possible intermediate electron carriers.The moderate rates observed for arsenate,selenite, and tellurite at pH 5.5, in contrast tothe negligible rate observed at pH 8.0, are inaccord with the hypothesis that the monovalentforms of these ions are reduced.
It is well known that the dissociation ofmolybdate and vanadate in solution is char-acterized by the formation of numerous poly-anions, particularly under acidic conditions. Theenzymatic reduction of molybdate was negligibleat pH 7.0, but reached an optimum at approxi-mately pH 5.5. The predominant ion in mo-lybdate solutions at the latter pH is Mo6021-6(Saxena and Saxena, 1961), and the increase inenzymatic activity noted at this pH may be dueto a requirement for this form of molybdate. Atthe more alkaline pH values, Mo6021-6 would bein equilibrium with less polymerized ions, forexample, Mo3O11-4. At a more alkaline pH,therefore, the concentration of Mo602j-6 wouldbe expected to increase exponentially withincreasing molybdate concentration. As shown inFig. 3, the rate of molybdate reduction increasedalmost exponentially with increasing totalmolybdate concentration, whereas vanadatereduction (discussed below) increased linearly.These results suggest that Mo602r76 is the actualionic species reduced in the reaction catalyzed by1M. lactilyticus extracts. As shown in Table 3,
titrimetric determinations indicate the produc-tion of 2 ,moles of base per Aumole of hydrogenuptake in molybdate reduction, as required bythe equation for Mo602 -6 reduction (shown inTable 5). In solutions having pH values between5 and 9, vanadate is predominantly in the ionicform, H2VO4-, which is in equilibrium withdivanadate and tetravanadate polyanions (Char-lot, 1954; Filipovic et al., 1954). The linearincrease in the rate of vanadate reduction by M.lactilyticus (Fig. 3) with increasing substrateconcentration suggests that the H2VO4- ion maybe the form of vanadate required in the reduction.When solutions of uranyl compounds are
buffered at neutrality, uranium is almost quanti-tatively precipitated in the form of uranylhydroxide [UO2(OH) 21 (Charlot, 1954). Therelatively high rate of reduction of uranyl byextracts of III. lactilyticus at pH 5.2 (Fig. 1)suggests the possibility that uranyl ion is theactual species reduced. In this connection, it isof interest that the reduction was even morerapid when citrate buffers were used. Citrate isknown to stabilize uranyl ion by the formationof a complex.
Rates of reduction. The specific activities ofmost of the reductions catalyzed by M. lactilyticusare shown in Table 5. These values were obtainedwith crude extracts without the addition ofelectron carriers, although compounds such asbenzyl viologen considerably enhanced the ratesof reduction of several of the compounds (Wool-folk et al., 1959). Cell suspensions were alsocapable of reducing most of the compounds listedin Table 5; however, the relative rates of thevarious reductions were not the same as withextracts (Woolfolk et al., 1959).
Inorganic reductions with other hydrogen donorsand other bacteria containing hydrogenase. Thereduction of inorganic compounds may be couplednot only to hydrogenase, but also to fermenta-tions which yield hydrogen. Thus, when pyruvate(Whiteley and Ordal, 1957) or hypoxanthine(Whiteley and Ordal, 1956) are fermented in thepresence of inorganic compounds, the productionof hydrogen is partially or completely inhibitedand the inorganic compound is reduced. Not allof the compounds listed in Table 5 have beentested, but many of the sulfur and nitrogencompounds, as well as arsenate, selenite, tellurite,and ferricyanide, are reduced as a result of thesefermentations. Di- and triphosphopyridine
653
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

WOOLFOLK AND WHITELEY
nucleotides could not serve as hydrogen donorsfor these reductions.
Certain other hydrogenase-containing bacteriawere examined with respect to their ability toreduce some of the above inorganic compounds.C. pasteurianum and D. desulfuricans reducedmany of the compounds rapidly (Table 6) but didnot reduce nitrate or arsenate. In addition to thecompounds listed in Table 6, Desulfovibrio was
also capable of reducing osmium tetroxide,bismuthate, ferricyanide, manganese dioxide,molybdate, and uranyl hydroxide. Extracts of E.coli catalyzed the reduction of hydroxylamine,
nitrate, and oxygen, as reported by McNall andAtkinson (1957) and Lascelles and Still (1946),but none of the other inorganic compoundstested. Extracts prepared from A. vinelandiireduced only oxygen (noted previously by Wilson,Lee, and Wilson, 1942), whereas extracts of R.vannielii, which have a hydrogenase capable ofreducing methylene blue, did not reduce any ofthe inorganic compounds tested. Suspensions ofyoung cells (approximately 10 hr of growth) ofM. aerogenes reduced most of the inorganic com-
pounds reduced by M. lactilyticus, but the specificactivities were low. These reactions were not
TABLE 5. Specific activities and Eo' values of the inorganic reductions catalyzed by extracts ofMicrococcus lactilyticus
Substrate Reaction catalyzed by extracts of M. lactilyticusa Specific activityb E,tc
Selenium Se + H2 -÷ HSe- + H+ 6.1 - .66Metabisulfite S205- + H2 -+ S204 + H20 83.0 -.48Benzyl viologen BV.. + 1½ H2 -- BVred 440 - .36dMolybdate Mo6021r6 + 3 H2 -* 6 M0O2.s + 6 OH- 58.0 -.32Phosphotungstate 23.0Uranyl hydroxide U02(OH)2 + H2 -k U(OH)4 18.5 -.26Sulfur S + H2 --1H2S 32.6 -.25Bismuth hydroxide Bi(OH)s + Y2 H2 Bi + 3 H20 3.4 -.15Ferric hydroxide Fe(OH)a + M H2 Fe(OH)2 + H20 24.1 -.12Tetrathionate S406- + H2 - 2 S203- + 2 H+ 1.4 -.08Tellurite HTeO32 + 2 H2 -- Te + 2 H20 + OH- 15.1 -.07Arsenate H2AsO4 + H2 -) HAsO2 + OH- + H20 2.7 0.00Vanadate H2VO4 + ½ H2 - VO(OH)2 + OH- 59.0 +.19Selenite HSeOs- + 2 H2 - Se + 2 H20 + OH- 39.4 +.23Osmium dioxide 0802 + 2 H2 0s + 2 H20 2.6 +.27Nitrite NO2- + 3 H2 NH4+ + 2 OH- 1.1 +.32Cupric hydroxide Cu(OH)2 + M1H2 -> Cu(OH) + H20 10.4 +.32Osmium tetroxide 0S04 + H2 -4 004-+ 2 H+ 21.0 +.36Manganese dioxide MnO2 + H2 - Mn+ + 2 OH- 10.4 +.40Nitrate NO2- + H2-) N027 + H20 1.6 +.41Ferricyanide Fe(CN)60 + M1H2 -- Fe(CN)o- + H+ 54.0 +.41Tellurate HTeO47 + 3 H2 - Te + 3 H20 + OH- 2.6 +.51Dithionite S2047 + H2 S203 + H20 7.7 +.52Lead dioxide PbO2 + H2 Pb(OH)2 0.7 +.63Hydrazine N2H5OH + H2 + H20 -- 2 NH4+ + 2 OH- 0.3 +.69Oxygen 02 + 2 H2 -- 2 H20 15.4 +.78Bismuthate BiO2- + %2 H2 Bi + 2 H20 + OH- 12.3 +.89Hydroxylamine NH30H+ + H2 NH4+ + H20 8.5 +.90a The equations show the various ions and compounds in the form that predominates at the pH where
reduction was observed (Charlot, 1954), except where data have been obtained suggesting that a minorcomponent was the actual species reduced (i.e., metabisulfite, Mo6021-6).
b Expressed as ,umoles of H2 X 108 per min per mg of protein.c The Eo' values for pH 7.0 given in this table are calculated from the normal oxidation potentials
given by Latimer (1952). The potential for the molybdate-molybdenum blue couple was approximatedwith the aid of information given by Saxena and Saxena (1961).
d Value of Michaelis and Hill (1933).
654 J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 84, 1962 INORGANIC COMPOUND REDUCTION BY M. LACTILYTICUS
TABLE 6. Reduction of inorganic compounds with molecular hydrogen by extracts of several hydrogenase-containing bacteriaa
Substrate M. lactilyticusb M. aerogenes' D. desulfuricans C. pasteurianum E. coli A. vinelandii
Benzyl viologen, methyl violo-gen, or methylene blued..... 440 238 1,006 1,170 20.8 132
Nitrate....................... 1.6 0.3 0.0 0.0 10.1 0.0Nitrite ....................... 1.1 0.0 43.0 85.4 0.0 0.0Hydroxylamine ............... 8.5 3.2 65.5 31.8 4.1 0.0Arsenate ..................... 2.7 0.7 0.0 0.0 0.0 0.0Oxygen....................... 15.4 0.6 17.4 38.0 47.0 52.6Metabisulfite ................. 83.0 0.3 37.0 230.0 0.0 0.0Tetrathionate ................. 1.4 1.1 31.2 9.9 0.0 0.0Selenite ...................... 39.4 0.7 24.8 141.0 0.0 0.0Tellurite ..................... 15.1 0.4 23.4 31.8 0.0 0.0Vanadate ..................... 59.0 57.5 171.0Sulfatee ...................... 0.0 1.0 - -
a Results are expressed as smoles of H2 X 103 per min per mg of protein.Data for M. lactilyticus taken from Table 5 and included for comparative purposes.Values for M. aerogenes are for cell suspensions. Specific activities were computed on a mg (dry wt)
basis.d Hydrogenase activity was measured as hydrogen evolution from reduced methyl viologen for M.
aerogenes, as hydrogen uptake with benzyl viologen for M. lactilyticus, D. desulfuricans, and C. pas-teurianum, and methylene blue for E. coli and A. vinelandii.
s Adenosine triphosphate (10,moles) and MgCl2 (10 ,umoles) were added.
observed with extracts of M. aerogenes or withsuspensions of older cells.
DISCUSSION
The inorganic reductions catalyzed by M.lactilyticus are summarized in Table 5. A numberof sulfur and nitrogen compounds can be reducedby other bacteria (Nason, 1962; Peck, 1962),both in assimilatory and dissimilatory reactions,and the mechanism of certain of these resemblesthe reductions mediated by M. lactilyticus. Theintermediates in the conversion of sulfur com-pounds by the latter organism are described inthe following paper (Woolfolk, 1962); thereduction of nitrogen compounds will be describedelsewhere. The reductions of selenite, tellurite,ferricyanide, ferric ion, manganese dioxide,oxygen, and benzyl viologen have also beenobserved for other bacteria. The hydrogenase-coupled reductions of selenite, selenium, mo-lybdate, tellurate, tellurite, phosphotungstate,arsenate, vanadate, osmium dioxide and te-troxide, bismuthate, hydroxides of uranyl, bis-muthyl, ferric, and cupric ions, and manganeseand lead dioxide have not been described pre-viously. In most of the reactions, a quantitativeone-electron or two-electron reduction was
observed. In some (bismuthate, bismuth hydrox-xide, tellurate, tellurite, selenite, and osmiumdioxide), the anion was reduced completely tothe free metal. The stoichiometry and rates ofreduction of bismuthate and tellurate are con-sistent with the formation of bismuth hydroxideand tellurite, respectively, as intermediates, andselenium was shown to be an intermediate in theconversion of selenite to selenide. Neither therates of the various reactions nor the extent ofreduction were related to the oxidation-reductionpotential.The present investigation has shown that
many of the above reactions can also be catalyzedby extracts of D. desulfuricans and C. pasteuri-anum. The reactions with Desulfovibrio mayresult from the combined action of cytochromeC3 and hydrogenase, as reported by Senez andPichinoty (1958a, b) for nitrite and hydroxyl-amine. lt is also possible that other naturallyoccurring carriers, such as the brown protein ofHori (1961), may participate in some of thereactions. Although M. lactilyticus does notpossess cytochromes, evidence has been presented(Whiteley and Ordal, 1957) that an iron-contain-ing intermediate electron carrier, probably aprotein, may function in another hydrogenase-
655
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

WOOLFOLK AND WHITELEY
coupled reduction, that of xanthine. This inter-mediate electron carrier has recently beenisolated and characterized (Mortenson, Valentine,and Carnahan, 1962; Valentine, Jackson, andWolfe, 1962). Possibly there are a number of"electron transport" proteins in this anaerobewhich may be conveniently investigated bymeans of an appropriate inorganic reduction. Therole of electron-transport proteins and themechanisms of some of the reductions catalyzedby .i. lactilyticus are currently under investiga-tion.The possible function of the reactions shown in
Table 5 in the metabolism of M. lactilyticus is ofinterest. Certain of the reactions (e.g., thereduction of sulfur and nitrogen compounds)may be of significance in providing reducedcompounds utilized for growth. The reactionsmight also be coupled to the oxidation of reducedcoenzymes such as flavin adenine dinucleotide,and thus influence over-all metabolism. It is alsopossible that certain of these reductions arecatalyzed nonspecifically by enzymes and co-enzymes which have another function in metab-olism. Thus, these enzymes or coenzymes couldbe isolated and studied by virtue of an inorganicreduction which is catalyzed only incidentally.Of interest in this connection is the observationthat the well-known diaphorase of animal tissueshas been identified as lipoyl dehydrogenase(Massey and Veeger, 1960).
ACKNOWLEDGMENTS
We wish to thank E. J. Ordal and H. C.Douglas, for helpful discussions in the course ofthis work, and Kayo Sunada, for assistance incertain of the experiments.
This investigated was carried out during thetenure of a Predoctoral Fellowship (awarded toC. A. W.) from the Division of General MedicalSciences, U.S. Public Health Service.
LITERATURE CITED
BARRENSCHEEN, H. K., AND H. A. BECKH-WID-MANSTETTER. 1923. Uber bakterielle Reduk-tion organisch gebundener Phosphorsaure.Biochem. Z. 140:279-83.
BONNIN, A., AND P. SUE. 1952. Separation parchromatographie sur papier des phosphates,phosphites, et hypophosphites. Compt. rend.234:960-961.
BROMFIELD, S. M. 1954. Reduction of ferric com-
pounds by soil bacteria. J. Gen. Microbiol.11:1-6.
CARNAHAN, J. E., AND J. E. CASTLE. 1958. Somerequirements of biological nitrogen fixation.J. Bacteriol. 75:121-124.
CHALLENGER, F. 1951. Biological methylation.Advances in Enzymol. 12:429-491.
CHARLOT, G. 1954. Qualitative inorganic analysis.John Wiley & Sons, Inc., New York.
CHARLOT, G., AND D. BEZIER. 1957. Quantitativeinorganic analysis. John Wiley & Sons, Inc.,New York.
CORPER, H. 1915. Sodium tellurite as a rapid testfor the viability of tubercle bacilli. J. In-fectious Diseases 16:47-53.
FALCONE, G., AND W. J. NICKERSON. 1960. Enzy-matic reduction of selenite. Bacteriol. Proc.,p. 152.
FILIPOVIC, I., Z. HAHL, Z. GASPARAC, AND V.KLEMENCIC. 1954. Polarographic character-istics of +5 vanadium in phosphate, borateand carbonate buffers. J. Am. Chem. Soc.76:2074-2080.
FOUBERT, E. L., AND H. C. DOUGLAS. 1948. Studieson the anaerobic micrococci. I. Taxonomicconsiderations. J. Bacteriol. 56:25-34.
GEST, H. 1954. Oxidation and evolution of molecu-lar hydrogen by microorganisms. Bacteriol.Rev. 18:43-73.
GHOSH MAZUMDAR, A. S., AND M. LEDERER. 1957.The separation of carrier-free tellurium tracesby paper chromatography. J. Inorg. & NuclearChem. 3:379-383.
GLEMSER, O., AND G. LUTZ. 1947. Zur Kenntnisder Molybdauoxydes und der Molybdaublau.Naturwissenschaften 34:215.
GLEMSER, O., AND G. LUTZ. 1951. Uber Molyb-daublau. Z. anorg. u. allgem. Chem. 264:17-33.
GREEN, H. S. 1918. Description of a bacterium iso-lated from a cattle-dipping tank, which re-duced arsenate to arsenite. Union S. Africa,Dept. Agr., Rept. Director Vet. Research5/6:613.
HORI, K. 1961. Electron transporting componentsparticipating in nitrate and respirations froma halo-tolerant micrococcus. II. Properties ofcytochrome C"1 and brown protein. J. Bio-chem. (Tokyo) 50:481-485.
ISHIMOTO, M., Y. KONDO, T. Kameyama, T. Yagi,AND M. SHIRAKI. 1957. The role of cytochromein enzyme system of sulfate-reducing bacteria.Intern. Symposium Enzyme Chem., Tokyo, p.212.
JAN, A. 1939. La reduction biologique du molyb-date d'ammonium par les bact6ries du genreSerratia. Bull. sci. pharmacol. 46:336-339.
LASCELES, J., AND J. L. STILL. 1946. Utilization
656 J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 84, 1962 INORGANIC COMPOUND REDUCTION BY M. LACTILYTICUS
of molecular hydrogen by bacteria. AustralianJ. Exptl. Biol. Med. Sci. 24:37.
LATIMER, W. M. 1952. The oxidation states of theelements and their potentials in aqueous solu-tions, 2nd ed. Prentice-Hall, Inc., New York.
LAYNE, E. 1957. Spectrometric and turbidometricmethods for measuring proteins, p. 447-454.In S. P. Colowick and N. 0. Kaplan [ed.],Methods in enzymology, vol. 3. AcademicPress, Inc., New York.
LEININSEN, E., AND T. CHULSKI. 1953. Disodiumdihydrogen hypophosphate. Inorg. Syntheses4:68-71.
LEVINE, V. E. 1925. The reducing properties ofmicroorganisms with special reference toselenium compounds. J. Bacteriol. 10:217-263.
MCNALL, E. G., AND D. E. ATKINSON. 1956. Ni-trate reduction. I. Growth of Escherichia coliwith nitrate as sole source of nitrogen. J.Bacteriol. 72:226-229.
McNALL, E. G., AND D. E. ATKINSON. 1957. Nitratereduction. II. Utilization of possible inter-mediates as nitrogen sources and as electronacceptors. J. Bacteriol. 74:60-66.
MANN, P. J. G., AND J. H. QUASTEL. 1946. Man-ganese metabolism in soils. Nature 158:154-156.
MARCHAL, J. G., AND T. H. GIRARD. 1948. Etudedu pouvoir reducteur de quelque souches decolibacille sur le molybdate d'ammoniaque.Trav. lab. microbiol. fac. pharm. Nancy 16:11-23.
MASSEY, V., AND C. VEEGER. 1960. On the reactionmechanism of lipoyl dehydrogenase. Biochim.et Biophys. Acta 40:185.
MICHAELIS, L., AND E. G. HILL. 1933. The viologenindicators. J. Gen. Physiol. 16:859-873.
MORTENSON, L. E., R. C. VALENTINE, AND J. E.CARNAHAN. 1962. An electron transportfactor from Clostridium pasteurianum. Bio-chem. Biophys. Research Communs. 7:448-452.
MORTON, H. E., AND T. F. ANDERSON. 1941. Elec-tron microscopic studies of biological reac-tions. I. Reaction of potassium tellurite byCorynebacterium diphtheriae. Proc. Soc. Exptl.Biol. Med. 46:272-276.
MURRAY, R. G. E., AND H. C. DOUGLAS. 1950.The reproductive mechanism of Rhodomicro-bium vannielii and the accompanying nuclearchanges. J. Bacteriol. 62:767-783.
NASON, A. 1962. Enzymatic pathways of nitrate,nitrite, and hydroxylamine metabolisms.Bacteriol. Rev. 26:16-41.
NASON, A., AND H. TAKAHASHI. 1958. Inorganicnitrogen metabolism. Ann. Rev. Microbiol.12:203-246.
NEWTON, J. W., P. W. WILSON, AND R. H. BURRIS.
1953. Direct demonstration of ammonia as anintermediate in nitrogen fixation by Azoto-bacter. J. Biol. Chem. 204:445-451.
PACKER, L. 1958. Respiratory carriers involved inthe oxidation of hydrogen and lactate in afacultative autotroph. Arch. Biochem. Bio-phys. 78:5465.
PECK, H. D., JR. 1959. The ATP-dependent reduc-tion of sulfate with hydrogen in extracts ofDesulfovibrio desulfuricans. Proc. Natl. Acad.Sci. U.S. 45:701-708.
PECK, H. D., JR. 1962. Comparative metabolismof inorganic sulfur compounds in microor-ganisms. Bacteriol. Rev. 26:67-94.
POSTGATE, J. R. 1951. The reduction of sulphurcompounds by Desulphovibrio desulphuricans.J. Gen. Microbiol. 5:725-738.
POSTGATE, J. R. 1956. Cytochrome c3 and desulpho-viridin; pigments of the anaerobe Desulpho-vibrio desulphuricans. J. Gen. Microbiol. 14:545-572.
POSTGATE, J. R. 1959. Sulphate reduction by bac-teria. Ann. Rev. Microbiol. 13:505-520.
RUDAKOV, K. I. 1927. Die Reduktion der minerali-schen Phosphate auf biologischem Wege. Ber.ges. Physiol. 41:507.
SANDELL, E. B. 1944. Colorimetric determinationof traces of metals. Interscience Pubs., Inc.,New York.
SAXENA, R. A., AND G. P. SAXENA. 1961. Electro-metric study on acid-molybdate system andthe formation of polyanions. Z. physik.Chem. (Frankfurt) 29:181-187.
SCHATZ, A., AND C. BOVELL. 1952. Growth andhydrogenase production of a new bacterium,Hydrogenomonasfacilis. J. Bacteriol. 63:87-98.
SENEZ, J. C., AND F. PICHINOTY. 1958a. Reductionde l'hydroxylamine liee 'a l'activite de l'hy-drogenase de Desulfovibrio desulfuricans. II.Nature du systeme enzymatique et du trans-porteur d'electrons interveant dans la reac-tion. Biochim. et Biophys. Acta 28:254.
SENEZ, J. C., AND F. PICHINOTY, 1958b. Sur lareduction du nitrite aux depens de l'hydrogenemoleculaire par Desulfovibrio desulfuricars etd'autres especes bacteriennes. Bull. soc. chim.biol. 40:2099-2177.
STEVENS, H. M. 1956. The preparation, detectionand separation of some valency states ofvanadium and cobalt by paper chromatog-raphy. Anal. Chim. Acta 15:51-75.
TERAI, T., T. KAMAHORA, AND Y. YAMAMURA.1958. Tellurite reductase from Mycobacteriumavium. J. Bacteriol. 75:535-539.
VALENTINE, R. C., R. L. JACKSON, AND R. S.WOLFE. 1962. Role of ferredoxin in hydrogenmetabolism of Micrococcus lactilyticus. Bio-
657
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

WOOLFOLK AND WHITELEY
chem. Biophys. Research Communs. 7:453-456.
WHITELEY, H. R. 1957. Fermentation of aminoacids by Micrococcus aerogenes. J. Bacteriol.74:324-330.
WHITELEY, H. R., AND H. C. DOUGLAS. 1951. Thefermentation of purines byMicrococcus lactilyt-icus. J. Bacteriol. 61:605-616.
WHITELEY, H. R., AND E. J. ORDAL. 1956. The pro-
duction of hydrogen by hypoxanthine oxida-tion, p. 521-538. In W. D. McElroy and B. Glass[ed.], Inorganic nitrogen metabolism. JohnsHopkins Press, Baltimore.
WHITELEY, H. R., AND E. J. ORDAL. 1957. Fer-mentation of alpha keto acids by Micrococcusaerogenes and Micrococcus lactilyticus. J.Bacteriol. 74:331-336.
WIBERLEY, S. E., L. G. BASSET, A. M. BURRILL,AND H. LYNG. 1953. Spectrophotometric de-termination of selenium and tellurium in con-
centrated sulfuric acid. Anal. Chem. 25:1586-1588.
WILSON, J. B., S. B. LEE, AND P. W. WILSON. 1942.Mechanism of biological nitrogen fixation.
IX. Properties of hydrogenase in azotobacter.J. Biol. Chem. 144:265-271.
WOOLFOLK, C. A. 1962. Reduction of inorganiccompounds with molecular hydrogen byMicrococcus lactilyticus. II. Stoichiometrywith inorganic sulfur compounds. J. Bacteriol.84:659-668.
WOOLFOLK, C. A., E. J. ORDAL, AND H. R. WHITE-LET. 1959. The reduction of arsenate and otherinorganic compounds by Micrococcus lactilyt-icus. Bacteriol. Proc., p. 116.
WOOLFOLK, C. A., E. J. ORDAL, AND H. R.WHITELEY. 1960. The reduction of inorganiccompounds by Micrococcus lactilyticus. Bac-teriol. Proc., p. 152.
WOOLFOLK, C. A., AND H. R. WHITELEY. 1961.Further studies on the reduction of inorganiccompounds by Micrococcus lactilyticus. Bac-teriol. Proc., p. 173.
YAMAGUCHI, K. 1953. Analysis of inorganic anionsby paper chromatography. J. Pharm. Soc.Japan 73:1258-1289.
ZALOKAR, M. 1953. Reduction of selenite byNeurospora. Arch. Biochem. Biophys. 44:330-337.
658 J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from