recombinant ns3 serine protease from dengue virus … · 1.3 status of dengue therapy ... figure...
TRANSCRIPT

RECOMBINANT NS3 SERINE PROTEASE FROM DENGUE VIRUS 2 AS A SCREEN FOR SMALL MOLECULES
by
NUROHAIDA BINTI AB AZIZ
Thesis submitted in fulfillment of the requirements for the degree of
Master of Science
August 2011

847785
'('0
~ I<A ~44-U4 ~ :q~-t 4
"10 \ \

ACKNOWLEDGMENTS
First and foremost I would like to express my deepest gratitude to my main
supervisor, Professor Maqsudul Alam, who had contributed fruitful ideas, comments,
continuous motivation and excellent technical assistance towards the completion of
this thesis. I would like to thank my co-supervisor, Dr. Jennifer Saito for her
assistance, guidance and supervision throughout this study. I would also like to thank
my collaborator, Dr. Irene Newhouse from University of Hawaii for the collaboration
work in in silica drug design. I would also like to thank Dr. Azat Mukhametov from
the Centre for Chemical Biology, Universiti Sains Malaysia for performing
additional structural analysis.
I am thankful to Professor Nazalan Najimudin and Dr. Rashidah for their
guidance and support. I would like to express my sincere gratitude to my lab
members especially Dr. Teh Aik Hong, Dr. Masaomi Kanbe, Suria, Su Yean,
Luqman, Beng Soon, Patrick, Lingsze, Sheri, Bee Feong and friends for the
motivation and help during my Master of science program. I would also like to thank
the Centre for Chemical Biology management team including Mr. Larry, Ms. Roslina,
Ms. Nithi and Ms. Komala for all of their help throughout the years.
Most of all, I want to thank my husband, Mohd Aatif for his love,
encouragement and support in completing this thesis. Finally, I am grateful to my
parents and family r:1embers for their constant encouragement and support during
this endeavor.
11

TABLE OF CONTENTS
ACKNOWLEDGEMENTS ......................................................................................... ii
TABLE OF CONTENTS ............................................................................................ iii
LIST OF TABLES .................................................................................................... viii
LIST OF FIGURES .................................................................................................... ix
LIST OF SYMBOLS AND ABBREVIA nONS ..................................................... xiii
ABSTRAK ................................................................................................................ xvi
ABSTRACT ............................................................................................................ xviii
CHAPTER 1 - Introduction
1.1 Dengue virus infection ........................................................................................... 1
1.2 Transmission, prevalence, and consequences ........................................................ 2
1.3 Status of dengue therapy ........................................................................................ 6
1.4 Molecular biology of dengue virus ........................................................................ 9
1.5 NS2B-NS3: The two-component protease of dengue virus ................................. 15
1.6 Structure cifDENV2 NS3 serine protease ............................................................ 20
1.7 Serine protease ..................................................................................................... 24
l.8 Strategies for dengue antiviral drug discovery .................................................... 27
1.9 Objectives ............................................................................................................. 30
CHAPTER 2 - Materials and methods
2.1 Over/iew of research methodology ..................................................................... 31
2.21n silica drug design ............................................................................................. 32
111

2.2.1 Building protein and ligand for computational docking experiments ....... 33 ,
2.2.2 Glide docking ............................................................................................. 33
2.2.3 Adsorption distribution metabolism and excretion (ADME) related
properties ............................................................................................................ 35
2.3 Recombinant DNA methods ................................................................................ 36
2.3.1 DNA Materials and vectors ....................................................................... 37
2.3.2 Primer design ............................................................................................. 38
2.3.3 Synthetic gene construction ...................................................................... 39
2.3.4 Construction of recombinant DNA ............................................................ 39
2.3.5 Purification of PCR product ...................................................................... 40
2.3.6 Preparative enzyme digestion .................................................................... 41
2.3.7 Extraction of DNA from agarose gels ....................................................... 42
2.3.8 Ligation of DNA ........................................................................................ 44
2.3.9 Isolation of plasmid DNA .......................................................................... 44
2.3.10 Restriction enzyme digestion (mini digestion) ........................................ 46
2.3.11 Site-directed mutagenesis ........................................................................ 46
2.4 Microbiological methods ..................................................................................... 47
2.4.1 Media and solutions ................................................................................... 47
2.4.2 Bacterial strains and growth conditions ..................................................... 47
2.4.3 Freezing and storage of E. coli cells .......................................................... 48
2.4.4 Transformation of E. coli competent cells ................................................. 48
2.5 Recombinant protein expression, purification, and analysis ................................ 49
IV

3.2.1 PCR amplification of gene fragments encoding for NS2B hydrophilic, NS3
protease domain, and chimeric NS2B-NS3pro ................................................... 75
3.2.2 Cloning ofNS2B-NS3pro chimeric gene into pET-14b vector (Novagen)
and sequence verification ................................................................................... 77
3.2.3 Expression of recombinant NS2B-NS3pro and recombinant NS2B-
NS3H51A inE. coli ................................................... ......................................... 80
3.2A Purification ofrecomhinant NS2B-NS3pro and recombinant NS2B-
NS3H5lA proteins by IMAC and gel filtration chromatography ...................... 81
3.3 Establishing the ill vitro functional assay ............................................................. 88
3.3.1 Autoprocessing activity of recombinant NS2B-NS3pro and recombinant
NS2B-NS3H51A at NS2BINS3 junction ........................................................... 88
3.3.2 Activity of recombinant NS2B-NS3pro toward GRR-AMC. .................... 89
3.4 Detem1ination of the in vitro activity of 4 high-scoring water soluble,
commercially available compounds ........................................................................... 91
3 A.l Solubility of compounds 1, 2, 3 and 4 ....................................................... 91
3 A.2 Protease inhibition activity of compounds 1, 2, 3 and 4 ............................ 93
3A.3 Inhibition mechanism of compound 4 against recombinant NS2B-NS3pro
............................................................................................................................ 96
CHAPTER 4 - Discussion ......................................................................................... 98
CHAPTER 5 - Conclusion
5.1 Final remarks ...................................................................................................... 106
5.2 Future research direction .................................................................................... 106
VI

REFERENCES ......................................................................................................... 108
APPENDIX A - Media and solutions
APPENDIX B - Potential anti-protease compounds selected by in silica drug design
APPENDIX C - Calibration curve of7-amino-4-methyl coumarin (AMC)
Vll

LIST OF TABLES
Table 1.1 Status of DENY vaccine developments ...................................................... 7
Table 1.2 Representative DENY NS2B-NS3 protease inhibitors ............................. 28
Table 2.1 DNA and vectors used in this study .......................................................... 37
Table 2.2 Primers used for construction of the recombinant protease ....................... 38
Table 2.3 peR reaction setup .................................................................................... 40
Table 2.4 Preparative digestion setup ....................................................................... 42
Table 2.5 Ligation reaction setup .............................................................................. 44
Table 2.6 Mini-digestion setup ................................................................................. 46
Table 2.7 Site-directed mutagenesis reaction setup .................................................. 47
Table 2.8 E. coli strains used in this study ................................................................ 48
Table 2.9 Recipe for polyacrylamide separating gel ............................. ··················· 55
Table 3.1 Structure of compounds selected for in vitro protease inhibition assay ... 72
Table 3.2 Predicted ADME-related properties of the 4 small molecule inhibitors .... 73
Table 3.3 Percentage of protease inhibition activity of 4 compounds ....................... 95
Vlll

LIST OF FIGURES
Figure 1.1 Transmission of DENY by Aedes aegypti and/or Aedes albopitus ............ 3
Figure 1.2 Approximate distributions of dengue cases in 2008 .................................. 4
Figure 1.3 Model for antibody-dependent enhancement of DENY replication .......... 5
Figure 1.4 Schematic representations of DENY genome organization and polyprotein
processing .................................................................................................................. 19
Figure 1.5 The DENY replication cycle ................................................................... 13
Figure 1.6 Structure and organization of the DENY NS2B-NS3 protease ............... 15
Figure 1.7 Sequences of DENY NS3 serine protease domain .................................. 16
Figure 1.8 Sequences of DENV NS2B ..................................................................... 17
Figure 1.9 Schematic representation of the structures NS2B-NS3 protease ............. 20
Figure 1.10 Comparison of the DENY NS3 protease (PDB identifier 1 BEF) and the
DENY NS2B-NS3 protease (PDB identifier 2FOM) ............................................... 23
Figure 1.11 Schechter and Berger System of Nomenclature .................................... 25
Figure 1.12 Catalytic mechanisms of DENY NS2B-NS3 serine protease ................ 26
Figure 2.1 Overview of research methodology .......................................................... 31
Figure 2.2 Schematic overview of the in silica-in vitro screening process ............... 32
Figure 2.3 Flowchart for processing, docking and scoring the ligand binding .......... 35
Figure 2.4 Flow chart for recombinant DNA methodology ....................................... 37
Figure 2.5 Flowchart for purification ofPCR product using QIAquick peR
Purification Kit protocol (Qiagen) ....................................................... .................... 41
ix

Figure 2.6 Flowchart for preparative enzyme digestion method (Saito, 2008) ......... 42
Figure 2.7 Flowchart for Extraction of DNA from agarose gels using Geneclean Spin
Kit protocol (Qbiogenc) ............................................................................................. 43
Figure 2.8 Flowchart for isolation of plasmid DNA using QIAprep Spin Miniprep Kit
protocol (Qiagen) ....................................................................................................... 45
Figure 2.9 Flowchart for transformation of E. coli competent cells (Saito, 2008) .... 49
Figure 2.10 Flowchart protein expression and purification methodology ................. 50
Figure 2.11 Flowchart expression of recombinant protein (D'arcy et al., 2006) ....... 52
Figure 2.12 Flowchart for protein purification by IMAC (D'arcy et al., 2006) ........ 53
Figure 2.13 Flowchart for protein purification by gel filtration chromatography
(D'arcy et al., 2006) ................................................................................................... 54
Figure 2.14 Flowchart for protein analysis by SDS-PAGE (Saito, 2008) ................. 57
Figure 2.15 Flowchart for protein analysis by immunoblot(Saito, 2008) .................. 59
Figure 2.16 Flowchart for in vitro functional assay methodology ............................. 60
Figure 2.17 Flowchart for determination of recombinant protease autocatalytic
activity (Niyomrattanakit et al., 2004) ....................................................................... 61
Figure 2.18 Flowchart to generate calibration curve of AMC (Chanprapaph, 2005) 62
Figure 2.19 Flowchart to determine the recombinant protease kinetic activity
(Chanprapaph et al., 2005) ......................................................................................... 63
Figure 2.20 Flowchart to determine the compound solubility (Tomlinson et al.,
2009a) ......................................................................................................................... 65
Figure 2.21 Flowchart for protease inhibitory assay (Tomlinson et al., 2009a) ........ 66
x

Figure 2.22 Flowchart for kinetic inhibitory assay (Chanprapaph et al., 2005) ........ 67
Figure 3.1 The active site of DENY protease ............................................................ 69
Figure 3.2 PCR amplification of gene fragments encoding for NS2B hydrophilic and
NS3 protease domains ................................................................................................ 76
Figure 3.3 PCR amplification of gene fragment encoding for chimeric NS2B-NS3pro
.................................................................................................................................... 76
Figure 3.4 Ncal and BamHI double digestion of recombinant pET-14b NS2B-
NS3pro ....................................................................................................................... 77
Figure 3.5 Multiple sequence alignment.. .................................................................. 78
Figure 3.6 Expression of recombinant protease ......................................................... 81
Figure 3.7 Purification of recombinant NS2B-NS3pro by Ni2+ affinity
chrolnatography ......................................................................................................... 82
Figure 3.8 Purification of recombinant NS2B-NS3H51A by Ni2+ affinity
chromatography ......................................................................................................... 84
Figure 3.9 Purification of recombinant NS2B-NS3pro by gel filtration
chromatography ......................................................................................................... 85
Figure 3.10 Purification of recombinant NS2B-NS3 H51 A by gel filtration
chrolnatography ................................................................................... , ..................... 87
Figure 3.11 SOS-PAGE showing absence of auto processing activity of recombinant
NS2B-NS3pro and recombinant NS2B-NS3H51A precursor at NS2BINS3 junction
.................................................................................................................................... 89
Figure 3.12 Recombinant NS2B-NS3pro enzymatic activity assays with GRR-AMC
fluorogenic peptide substrate ..................................................................................... 90
xi

Figure 3.13 Solubility of all 4 tested compounds in 50 mM Tris, pH 8.5 aqueous
cleavage buffer ........................................................................................................... 92
Figure 3.141n vitro recombinant DENV NS2B-NS3 protease inhibition assays ...... 94
Figure 3.15 The effect of compound 4 on the recombinant NS2B-NS3pro catalyzed
reaction ....................................................................................................................... 96
Figure 4.1 View of compound 4 at the allosteric binding site ofNS2B-NS3 protease
.................................................................................................................................. 103
xii

LIST OF SYMBOLS AND ABBREVIATIONS
A
AMC
bp
cDNA
cm
C-terminal
CY
DENY
DENY I, DENY2, DENV3, and
DENV4
OF
DHF
DNA
DSS
E. coli
ER
FcyR
g
GRR-AMC
HCY
HTS
IMAC
angstrom
7-amino-4-methyl coumarin
base pair (s)
complementary DNA
centimetre
carboxyl terminal
column volume
dengue virus
dengue virus serotype I, 2, 3, and 4
dengue fever
dengue hemorrhagic fever
deoxyribonucleic acid
dengue shock syndrome
Escherichia coli
endoplasmic reticulum
Fcy receptors
gram
Boc-G lye ine-Arginine-Arginine-4-
methy lcoumary 1-7 -amide
hepatitis C virus
high-throughput screening
Immobilized metal affinity chromatography
xiii

IPTG
kb
kDa
L
M
mg
mm
ml
mM
mm
ng
NGC
nm
nM
NS2B
NS3pro
N-terminal
°C
00
ORF
PCR
PDB
pmol
PVDF
RdRp
isopropyl-~-D-thiogalactopyranoside
kilobase pair (s)
kilodalton (s)
litre
molar
milligram
minute (s)
millilitre
millimolar
millimetre
nanogram
New Guinea C
nanometer (s)
nanomolar
non structural protein 2B
nonstructural protein 3 protease domain
amino terminal
degree Celsius
optical density
open reading frame
polymerase chain reaction
Protein Data Bank
picomolar
PolyVinyliDene Fluoride
RNA-dependent RNA polymerase
XIV

RNA
s
SAM
SAR
SOS-PAGE
TeA
TGN
TM
U
UTR
UV
V
v/v
VP
WNV
xg
~g
~I
~M
ribonucleic acid
second (s)
S-adenosy I methy Itransferase
structure-activity relationship
sodium dodecyl sulfate polyacrylamide gel
electrophoresis
Trichloroacetic Acid
trans-Golgi network
transmembrane regions
unit (s)
untranslated region
ultraviolet
voltage
volume/volume
vesicle packets
West Nile virus
g-force
microgram
microlitre
Micromolar
xv

REKOMBINAN PROTEASE SERINA NS3 DARIPADA VIRUS DENGGI 2
SEBAGAI PENYARING UNTUK MOLEKUL KECIL
ABSTRAK
Jangkitan denggi adalah muneul semula sebagai satu penyakit utama dunia
dan diklasifikasikan sebagai patogen utama kategori A. Setiap tahun, dianggarkan
50-100 juta manusia dijangkiti virus denggi dan dianggap sebagai penyebab kepada
salah satu penyakit virus baw(lan arthropoda paling penting dari segi kematian dan
kemorbidan manusia. Penjangkitan virus berlaku melalui gigitan nyamuk Aedes
aegypti dan separuh daripada populasi dunia berisiko kepada jangkitan. Sehingga
sekarang masih tiada lagi drug antivirus atau vaksin diluluskan yang berkesan untuk
menetang virus denggi. Fokus tesis ini adalah untuk menggabungkan diantara kuasa
pengkomputeran berprestasi tinggi dengan eksperimen makmal dimana rekombinan
protease serina NS3 daripada virus denggi 2 sebagai penyaring molekul keeil
antivirus yang boleh digunakan untuk menghalang atau merawat jangkitan virus
denggi.
Kerjasama dengan Dr. Irene Newhouse, Advance Studies· for Genomies,
Proteomies and Bioinformaties (ASGPB), University of Hawaii, model molekul dan
penyaringan secara in silica telah dijalankan keatas perpustakaan sebatian molekul
keeil daripada pangkalan data National Cancer Institute (NCI) dan ZINC untuk
sebatian keeil yang mengedok ke dalam tapak ikatan protease DENV2 NS2B-NS3.
Daripada himpunan calon-calon yang menunjukkan suaian terbaik (53 perencat
perencat molekul keeil yang berpotensi), 4 sebatian larut air yang menunjukkan skor
tertinggi, boleh didapati secara komersial telah dipilih untuk penilaian secara in vitro.
XVl

Gen protease serina NS2B-NS3 daripada virus denggi serotip 2 telah diklon
dan diekspresi dalam E. coli sebagai protein rekombinan berpenanda heksahistidina.
Protease NS2B-NS3 telah ditulen menggunakan kromatografi affinity dan penurasan
gel. Asai in vitro menunjukkan aktiviti protease terhadap substrat peptida fluorogenik
yang mengandungi dua residual berbes. Kesemua 4 sebatian larut alr yang
menunjukkan skor tertinggi diuji dan mempamerkan aktiviti perencatan secara in
vitro terhadap rekombinan protease serina NS2B-NS3. Sebatian 4 didapati memberi
kesan perencatan paling aktif dimana kadar perencatan sebanyak 64% pada
kepekatan 100 J.lM.
Sebagai kesimpulan, kajian tesis 1111 membuktikan bahawa rekombinan
protease senna NS3 daripada virus denggi 2 bolch digunakan sebagai penyanng
molekul kecil antivirus secara in silica dan in vitro. Sebatian 4 adalah penemuan
berharapan dan berpotensi untuk dibangunkan sebagai dnlg anti-denggi.
XVII

RECOMBINANT NS3 SERINE PROTEASE FROM DENGUE VIRUS 2 AS A
SCREEN FOR SMALL MOLECULES
ABSTRACT
Dengue infection is re-emerging as a major global disease and is classified as
a Category A priority pathogen. Dengue viruses are estimated to infect 50-100
million people annually and are considered to cause one of the most important
arthropod-borne viral diseases in terms of human morbidity and mortality. Virus
transmission occurs through the bite of the Aedes aebYJpti mosquito and half the
world's population is at risk for infection. There is presently no approved vaccine or
antiviral drug that is effective against dengue viruses. The focus of this thesis is to
combine the power of high perfonnance computing with wet lab experiments for the
recombinant NS3 serine protease from dengue virus type 2 as a screen for antiviral
small molecules that can be used either to prevent or treat dengue virus infections.
In collaboration with Dr. Irene Newhouse, Advance Studies for Genomics,
Proteomics and Bioinformatics (ASGPB), University of Hawaii, molecular
modelling and in silica screening of small molecule compound libraries from the
National Cancer Institute (NCI) and ZINC databases that dock into the DENV2
NS2B-NS3 protease binding site was carried out. From the pool of best-fit candidates
(53 potential small molecule inhibitors), the 4 high-scoring water-soluble,
commercially available compounds were selected for in vitro assessment.
The NS2B-NS3 serine protease gene from dengue virus serotype 2 was
cloned and expressed in E. coli as a recombinant hexahistidine tagged protein. The
NS2B-NS3 protease was purified using affinity and gel filtration chromatography. In
XVlll

vitro assay revealed protease activity toward a fluorogenic peptide substrate
containing two basic ammo acid residues. All 4 high-scoring water-soluble
compounds were tested and exhibited in vitro inhibition activity on the recombinant
NS2B-NS3 serine protease. Compound 4 was found to be most active inhibitor with
64% inhibition at 100 )lM concentration.
In summary, this thesis project has established that the purified recombinant
NS3 serine protease from dengue virus type 2 can be used to screen antiviral small
molecules in silico and in vitro. Compound 4 is a promising finding for further
development as an anti-dengue drug.
XIX

CHAPTER 1
Introduction
1.1 Dengue virus infection
Dengue virus (DENY) is the most important human viral disease transmitted
by an arthropod vector, with an estimated annual infection rate in excess of 50
million. The majority of infections are silent with no obvious ~linical symptoms.
Nevertheless, a significant minority of infected individuals develop a mild febrile
illness, dengue fever (DF), or even life-threatening dengue hemorrhagic fever
(DHF)/dengue shock syndrome (OSS) which has an increasing incidence in tropical
and sUbtropical countries.
The first reported epidemics of DF occurred In 1779-1780 in Asia, Africa,
and North America. During that time, OF was considered a benign, nonfatal disease
of visitors to the tropics. The disease was confined to relatively small geographic
regions and the four different serotypes of DENY remained isolated. On the contrary,
the global prevalence of DENY is now increasing dramatically and DENY epidemics
caused by multiple serotypes (hyperendemicity) are more frequent (Gubler, 1998;
Rigau-Perez et aI., 1998; Gubler, 2002).
The disease is caused by four antigenically related but distinct serotypes of
DENY: DENYl, DENY2, DENY3, and DENY4. Despite being an age-old disease,
there is no effective treatment for DENY infection. Researchers have endeavored to
develop a vaccine for many years with very little success. The reason is that an
effective vaccine would have to protect against all four serotypes of DENY.

Considerable efforts are now contributed to the development of antiviral compounds
(Sampath and Padmanabhan, 2009).
Efficient and accurate diagnosis of DENY is of primary importance for
clinical care. It includes epidemiological consideration (season of the year, travel
history), physical examination (high body temperature, blood pressure, evidence of
bleeding in the skin or other sites, hydration status, evidence of increased vascular
permeability, and tourniquet test), and clinical laboratory tests (virus isolation,
nucleic acid detection, detection of antigens, serological tests, and haematological
tests) (WHO, 2009).
1.2 Transmission, prevalence, and consequences
Mosquitoes, humans, and lower primates such as chimpanzees, gibbons, and
macaques are all considered to be the natural hosts for DENY infections. However,
humans are the main amplifying host of the virus (Henchal and Putnak, 1990).
DENY is transmitted to humans through the bite of infected female mosquitoes,
either Aedes aegypti or Aedes albopitus, which can usually be found near or in
human dwellings. The species is day-active, with most biting activity occurring in
the early morning or late afternoon.
The transmission cycle of DENY by the mosquito begins with a DENY
infected person (Gubler, 1998). The person will have virus circulating in the blood
for approximately 4 to 7 days. This beginning state is called viremia. During this
period, if other uninfected female mosquitoes bite the ill person, those mosquitoes
may become infected and becomes infective after an obligatory extrinsic incubation
period of 10 to 12 days. After the mosquito becomes infective, it may transmit
2
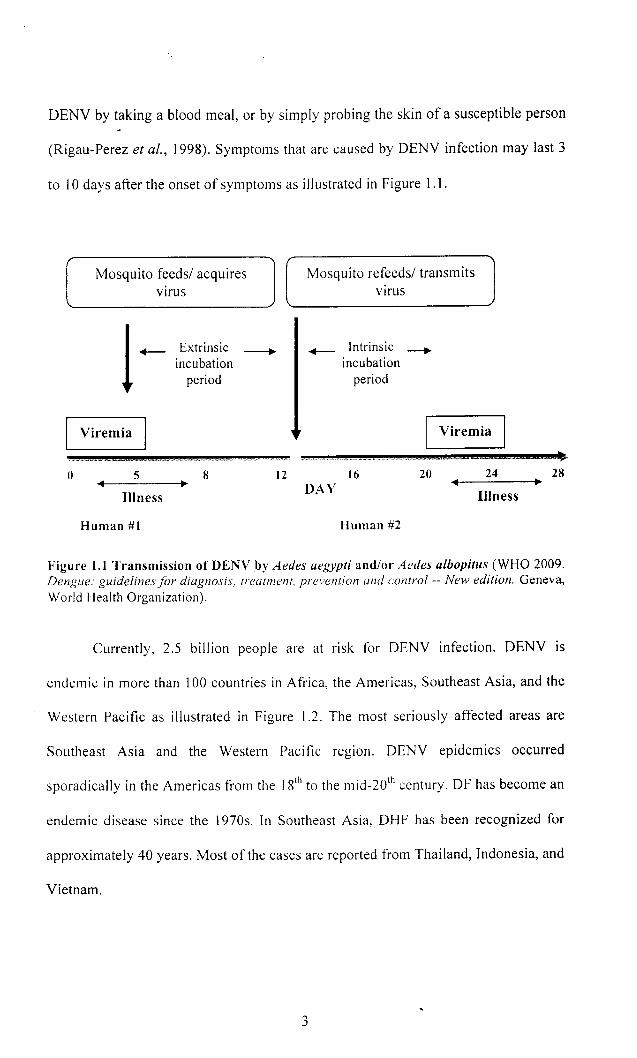
DENV by taking a blood meal, or by simply probing the skin of a susceptible person
(Rigau-Perez et al., 1998). Symptoms that are caused by DENV infection may last 3
to 10 days after the onset of symptoms as illustrated in Figure 1.1.
Mosquito feeds/ acquires virus
Viremia
Extrinsic incubation
period
o 5 8
Illness
Human #1
Mosquito refeeds/ transmits virus
~ Intrinsic ~ incubation
period
Viremia
12 16 20 24 DAY Illness
Human #2
--28
Figure 1.1 Transmission of DENY by Aedes aegypti and/or Aedes albopitus (WHO 2009. Dengue: guidelines for diagnosis, treatment, prevention and control -- New edition. Geneva, World Health Organization).
Currently, 2.5 billion people are at risk for DENV infection. DENV is
endemic in more than 100 countries in Africa, the Americas, Southeast Asia, and the
Western Pacific as illustrated in Figure 1.2. The most seriously affected areas are
Southeast Asia and the Western Pacific region. DENV epidemics occurred
sporadically in the Americas from the 18th to the mid-20th century. DF has become an
endemic disease since the 1970s. In Southeast Asia, DHF has been recognized for
approximately 40 years. Most of the cases are reported from Thailand, Indonesia, and
Vietnam.
3
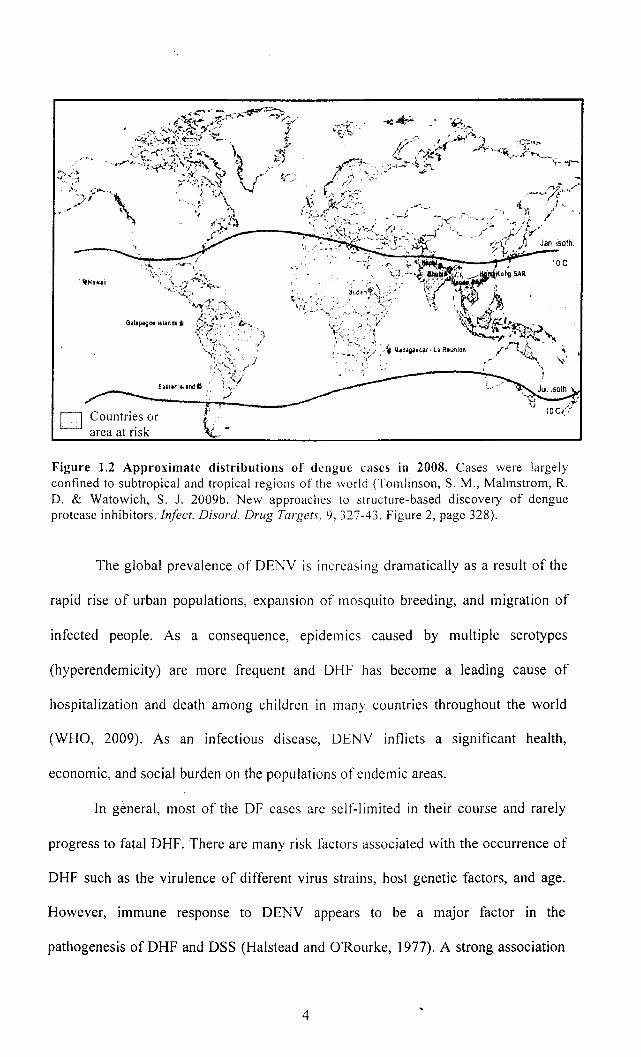
'tHawail
D Countries or area at risk
Figure 1.2 Approximate distributions of dengue cases in 2008. Cases were largely confined to subtropical and tropical regions of the world (Tomlinson, S. M., Malmstrom, R. D. & Watowich, S. J. 2009b. New approaches to structure-based discovery of dengue protease inhibitors. Infect. Disord. Drug Targets, 9,327-43. Figure 2, page 328).
The global prevalence of DENY is increasing dramatically as a result of the
rapid rise of urban populations, expansion of mosquito breeding, and migration of
infected people. As a consequence, epidemics caused by mUltiple serotypes
(hyperendemicity) are more frequent and DHF has become a leading cause of
hospitalization and death among children in many countries throughout the world
(WHO, 2009). As an infectious disease, DENY inflicts a significant health,
economic, and social burden on the populations of endemic areas.
In general, most of the OF cases are self-limited in their course and rarely
progress to fatal DHF. There are many risk factors associated with the occurrence of
DHF such as the virulence of different virus strains, host genetic factors, and age.
However, immune response to DENY appears to be a major factor in the
pathogenesis of DHF and DSS (Halstead and O'Rourke, 1977). A strong association
4
of severe disease in humans undergoing a heterotypic secondary infection has been
established4
(Halstead et al .. 1970; Yaughn et aI.. 1997).
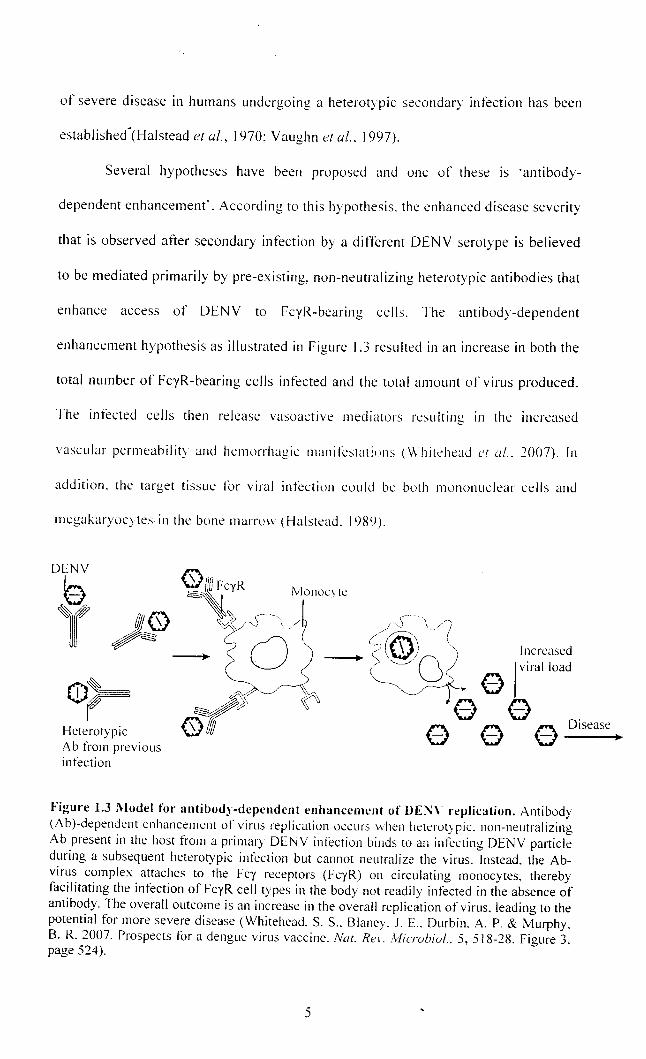
Several hypotheses have been proposed and one of these is 'antibody-
dependent enhancement'. According to this hypothesis. the enhanced disease severity
that is observed atter secondary infection by a different DENY serotype is believed
to be mediated primarily by pre-existing, non-neutralizing heterotypic antibodies that
enhance access of DENY to FcyR-bearing cells. The antibody-dependent
enhancement hypothesis as illustrated in Figure 1.3 resulted in an increase in both the
total number of FcyR-bearing cells infected and the total amount of virus produced.
The infected cells then release vasoactive mediators resulting in the increased
vascular permeability and hemorrhagic manitCstaliuns (Whitehead ('[ ul .. :2007). In
addition. the target tissue lor viral infection could be buth munonuclear cells and
megakaryocytes in the bone marnm (Halstead. 1989).
DENY e l(
Increased
Figure 1.3 Model for antibody-dependent enhancement of DE~\' replication. Antibody (A b)-dependent enhancement of virus replication occurs \\hen heterotypic. non-neutralizing Ab present in the host from a primary DENY infection binds to an infecting DENY panicle during a subsequent heterotypic infection but cannot neutralize the virus. Instead. the Abvirus complex attaches to the Fey receptors (FeyR) on circulating monocytes, thereby facilitating the infection of FeyR cell types in the body not readily infected in the absence of antibody. The overall outcome is an increase in the overall replication of virus. leading to the potential for more severe disease (Whitehead. S. S .. Blaney. J. E .. Durbin. A. P. & Murphy. B. R. 2007. Prospects for a dengue virus vaccine. Nal. Rei'. ill/ie-rubial .. 5, 518-28. Figure 3. page 524).
5

There are 3 criteria that must be fulfilled in order to meet the case definition
for severe dengue (WHO, 2009). The criteria are severe plasma leakage leading to
shock, accumulation of fluid and respiratory distress, severe hemorrhagic
manifestations, and severe organ impairment which mainly involve the liver, heart,
and central nervous system.
1.3 Status of dengue therapy
There is no specific treatment for DENV infection. The only treatment
available is symptomatic treatment vvith careful clinical management by experienced
physicians :md nurses. This can often save the lives ofDHF patients (WHO, 2009).
Despite considerable work over the years, a licensed vaccine against DENV
is still elusive and even today there are only candidate DENV vaccines. A successful
vaccine must be tetravalent, capable of simultaneously inducing a high level of long
lasting immunity to all four DENV serotypes (Ray and Shi, 2006). The immune
enhancement phenomenon underlying disease pathogenesis and the lack of suitable
animal models to evaluate candidate DENV vaccines are the major challenges to
vaccine development (Johnson and Roehrig, 1999; Lei et aI., 2001).
Various strategies have been used to develop DENV vaccines: live attenuated
viruses, chimeric live attenuated viruses, inactivated or sub-unit vaccines, and
nucleic acid-based vaccines (Halstead and Deen, 2002). However, efforts to develop
a DENV vaccine have focused mainly on live attenuated virus vaccines, inactivated
virus vaccine, and subunit virus vaccines (Table 1.1).
6

Table 1.1 Status of DENV vaccine developments
Vaccine Developer
Live attenuated tetravalent vaccine (LAya) WRAIR, GSK*
Intertypic chimeric vaccine (6.30a) NIAID, NIH*
Chimeric vaccine (ChimeriYaxa) Acambis, Sanofi-A ventis
Chimeric vaccine CDC*
Flavivirus-based recombinant DNA vaccine
Non-flavivirus based recombinant DNA vaccine (E (ecto) proteinb
)
Navy Medical Research Center
Hawaii Biotech
(Whitehead, S. S., Blaney, J. E., Durbin, A. P. & Murphy, B. R. 2007. Prospects for a dengue virus vaccine. Nat. Rev. Microbiol.. 5,518-7 8. Table I, page 525)
Phase
1 III
II
II
I
* Walter Reed Army Institute of Research (WRAIR); GlaxoSmithKline Biologicals (GSK); National Institute of Allergy and Infectious Diseases (NIAID); National Institutes of Health (NIH); Centers for Disease control and Prevention (CDC) a LA V, 6.30 and ChimeriVax are being tested as tetravalent vaccines (targeting all four DENY serotypes simultaneously) b Recombinant DENYI E protein ectodomain (N-terminal 80%) formulated in alum
In all these instances, the vaccine viruses are monovalent in that each one is
specific to one DENY serotyp~s. A tetravalent DENY vaccine is based on producing
vaccine formulations by mixing all four monovalent vaccine viruses. Studies show
that both the live attenuated and the ChimeriYax tetravalent DENY vaccine
formulations elicit unbalanced immune response due to viral interference
(Swaminathan and Khanna, 2010). The occurrence of this phenomenon, which tends
to skew the immune response predominantly towards one serotype, emphasizes the
limitations and the risks associated with mixing four monovalent vaccine viruses to
create a tetravalent vaccine. However, tetravalent formulations and immunization
7

schedules are being optimized, so as to confer similar levels of protection against all
four DENV serotypes (Ray and Shi, 2006).
In the absence of vaccines, drugs for specific therapy are needed, but no
antiviral medications are approved for use against DENV. The proteins required for
the fitness of the virus provide several potential targets against which to develop
antiviral drugs. Strategies for DENV antiviral drug discovery include structure-based
approaches, modulating the host immune response, high-throughput screening (HTS)
using virus replication cell-based assays, or HTS specifically targeting viral
morphogenesis, the 3' UTR, viral absorption, or assembly and maturation
(Tomlinson et al., 2009b).
A challenge for inhibitors discovered with virus replication cell-based HTS is
determining the mechanism of inhibition. Testing natural products identified several
lead compounds that inhibit DENV replication in cell-culture (Parida et al., 2002;
Kiat et al., 2006; Jain et al., 2008), with the components offingerroot (Boesenbergia
rotunda) reported to inhibit the DENV protease with a flM inhibition constant (Kiat
et al., 2006). A very general strategy utilizes compounds identified from other viral
studies and tests them for inhibitory activity against DENV replication. There have
been a few discoveries utilizing these various strategies, however, no leads have
progressed to clinical trials.
In recent years, health authorities have emphasized disease prevention and
mosquito control through community-based programs. Such programs are proper
solid waste disposal, improved water storage practices, covering containers to
prevent access by egg laying female mosquitoes, and the use of chemical and
biological insecticides (WHO, 2009). These programs are very demanding in terms
8

of time, expertise, and financial resources. Therefore, there are only of limited
usefulness for the control of DEN V diseases.
1.4 Molecular biology of dengue virus
DENV is a vector borne member of the genus Flavivirus and the family
Flaviviridae (Westaway et aI., 1985). The genus Flavivirus contains more than 70
members, including yellow fever virus (YFV), Japanese encephalitis virus (lEV),
tick-borne encephalitis virus (TBEV), and West Nile virus (WNV). The complete
nucleotide sequences of several tlaviviruses have been reported and sequence
comparison among the tlavivirus polyproteins suggested that despite divergences in
amino acid sequence, their hydrophobicity profiles are highly conserved, especially
within the NS I, NS3, and NS5 proteins (Chambers et aI., 1990a).
DENV displays four antigenically related but distinct serotypes: DENV 1,
DENV2, DENV3, and DENV4. The fOllr serotypes are almost indistinguishable in
terms of clinical and pathological symptoms they cause, but they can be identified by
neutralization tests, monoclonal antibodies, and polymerase chain reaction (peR)
(Morita et aI., 1991). These serotypes also vary in their degree of virulence and
infection with one DENV serotype provides lifelong immunity to that virus, but there
is no cross-protective immunity to the other serotypes. Each of the four serotypes can
cause severe and fatal disease (Rigau-Perez et aI., 1998).
DENV is a smooth and spherical enveloped virus with a diameter of 500 A.
The virus contains a single-stranded positive-sense RNA genome of 10,723
nucleotides. The genome is enclosed in the viral capsid which is surrounded by a
9
host-derived lipid bilayer envelope (Kuhn et al .. 2002). The RNA genome has a type
I cap at the S' end. but is lacking a poly(A) tract at the 3' end.
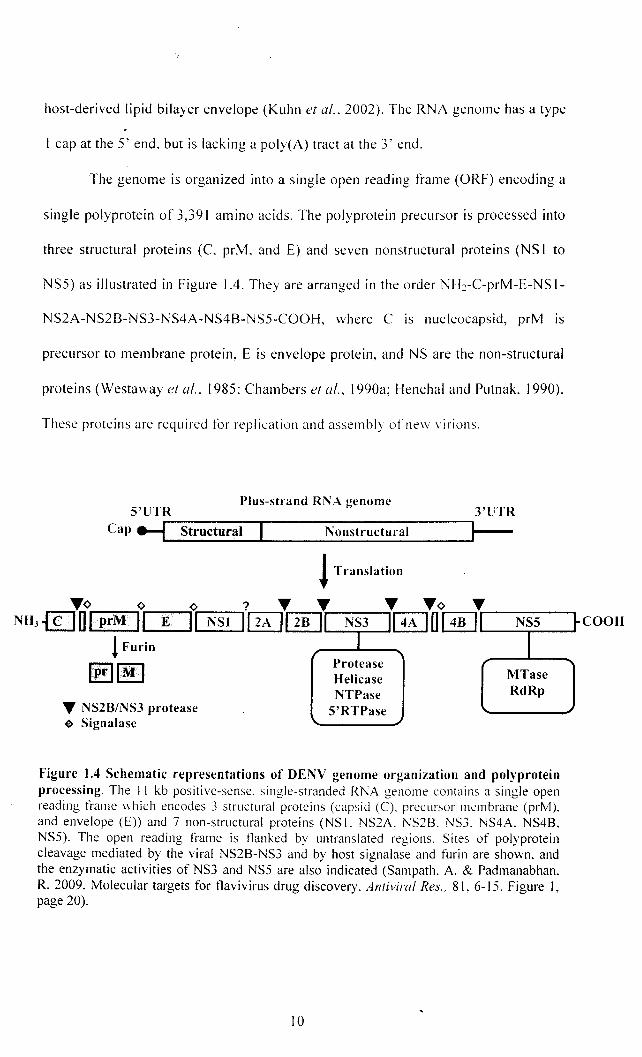
The genome is organized into a single open reading frame (ORF) encoding a
single polyprotein of 3,391 amino acids. The polyprotein precursor is processed into
three structural proteins (C. prM. and E) and seven nonstructural proteins (NS I to
NSS) as illustrated in Figure 104. They are arranged in the order NH~-C-prM-E-NS 1-
NS2A-NS2B-NS3-NS4A-NS4B-NSS-COOH, where C is nucleocapsid, prM is
precursor to membrane protein, E is envelope protein, and NS are the non-structural
proteins (Westaway et al.. 1985: Chambers el al.. I 990a; Henchal and Putnak, 1990).
These proteins are required for replication and assembly ofne\\ virions.
Plus-strand RNA genume S'UTR 3'UTR
C ~~------~--------------~~ ap ----.1.._S_tr_u_c_t_u_ra_l_.&-____ N_o_n_s_tt_·u_c_'t_ur_a_I ___ --Ir-
! Translatiun
'Y¢ ¢ ¢ ? 'Y 'Y NH3 {£] U IprM ,., ..... -E---'" NSI IIzA , ~
'Y 'Y¢ ... ~ U [ill'r----NS- S--, COOH II NS3 I 4
! Furin
~IM' 'Y NSZB/NS3 protease ~ Signalase
I Protease Helicase NTPase
S'RTPase
I MTase RdRp
Figure 1.4 Schematic representations of DENV genome organization and polyprotein processing. The 11 kb positive-sense. single-stranded RNA genome contains a single open reading frame which encodes 3 structural proteins (capsid (C). precursor membrane (prM). and envelope (E» and 7 non-structural proteins (NS 1. NS2A. NS2B. NS3. NS4A. NS4B. NS5). The open reading frame is tlanked by untranslated regions. Sites of polyprotein cleavage mediated by the viral NS2B-NS3 and by host signalase and furin are shO\vn. and the enzymatic activities of NS3 and NS5 are also indicated (Sampath. A. & Padmanabhan. R. 2009. Molecular targets for tlavivirus drug discovery. Antiviral Res .. 81. 6-\S. Figure I, page 20).
10

The C protein consists of ~ 120 amino acids and is involved with packaging of
the viral genome and forming the nucleocapsid (NC) core. This protein is the first
viral polyprotein synthesized during translation, has a molecular weight of about 13.5
kDa, and is rich in lysine and arginine residues. This highly basic character probably
enables it to interact with the virion RNA (Henchal and Putnak, 1990).
prM (~165 amino acids) and E (~495 amino acids) are glycoproteins, each of
which contains two transmembrane helices. Before it is cleaved during particle
maturation to yield the pr peptide and the M protein (~75 amino acids), the prM
protein might function as a chaperone for folding and assembly of the E protein.
The E protein contains a cellular receptor-binding site(s) and a fusion peptide
(Mukhopadhyay et al., 2005). E is associated \vith viral hemagglutination and
neutralization activity, and interacts with cellular receptors to mediate viral
attachment and entry (Crill and Roehrig, 2001).
NS I (46 kDa) is required for tlavivirus replication and is presumably
involved in negative-strand synthesis by an unknown mechanism. A large deletion in
YFY NS 1 abolished viral replication but can be complemented in trans by functional
expression from Sindbis virus vector (Lindenbach and Rice, 1997).
NS2A (22 kDa) is a small hydrophobic transmembrane protein that is
involved in production of virus particles and in generation of virus-induced
membranes during virus assembly (Leung et aI., 2008)
NS3 (70 kOa) and NS5 (104 kDa) are the best characterized non structural
proteins, with multiple enzyme activities that are required for viral replication. NS3
has three distinct activities: serine protease together with the cofactor NS2B, required
for polyprotein processing; helicaselNTPase activity, required for unwinding the
double-stranded replicative form of RNA; and RNA triphosphatase, required for
11

capping nascent viral RNA (Falgout et al., 1991; Zhang et al., 1992; Li et al., 1999).
Mutations that affect each activity impair viral replication (Matusan et al., 2001).
NS5 is the largest and most highly conserved flaviviral protein, with greater
than 75% sequence identity across all DENY serotypes. It contains two distinct
enzymatic activities, separated by an inter domain region: an S-adenosyl
methyl transferase (SAM) (Egloff et al., 2002) and an RNA-dependent RNA
polymerase (RdRp) (Guyatt et al., 2001). NS4A (16 kDa) is an integral membrane
nprotein which may induce membrane rearrangements to form the viral replication
complex. NS4B (27 kDa) inhibits the type I interferon response of host cells, and
may modulate viral replication via its interaction with NS3 (Sampath and
Padmanabhan, 2009).
DENY replicates In the cytoplasm of susceptible host cells, including
monocytes, macrophages, and dendritic cells. A specific receptor for internalization
of DENY into the host cell has not yet been identified. Several cellular molecules
capable of mediating virus attachment are known, but none have been conclusively
shown to function as virus receptors (Tassaneetrithep et al., 2003; Lozach et al.,
2005; Krishnan et aI., 2007; Miller et al., 2008).
As illustrated in Figure 1.5, during virus entry, E proteins forming the
glycoprotein shell bind to cell surface receptors that assist in the internalization of the
virus through clathrin-mediated endocytosis. Following internalization, the acidic
environment of the endosome triggers an irreversible trimerization of the E protein
that results in fusion of the viral and cell membrane. This leads to the release of the
viral RNA into the cytoplasm (Mukhopadhyay et aI., 2005; Krishnan et aI., 2007;
van der Schaar et al., 2007).
12
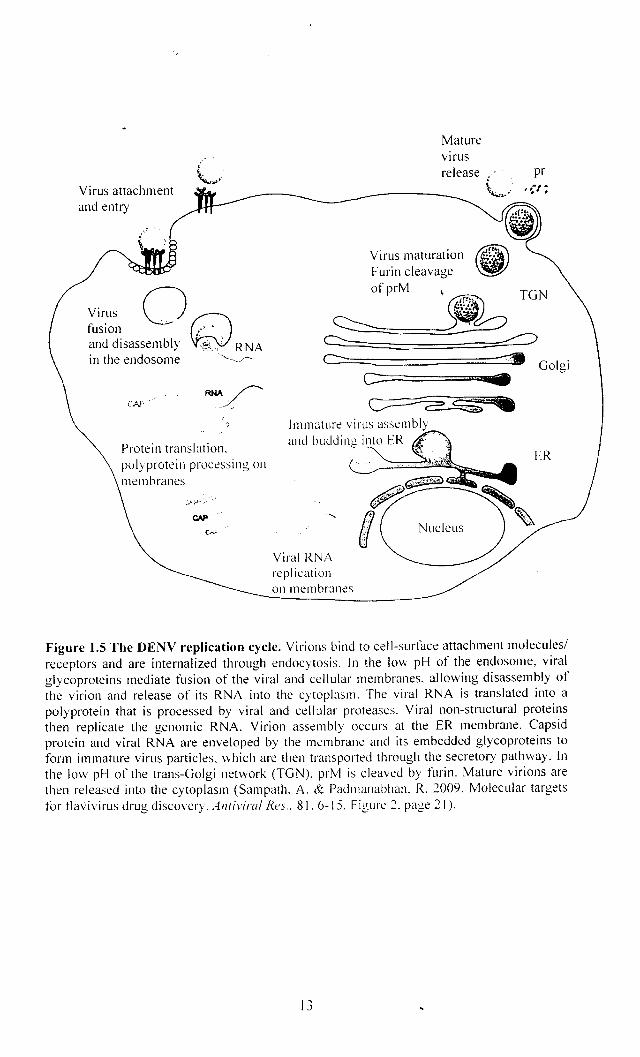
Virus attachment and entry
~ir.us o~ tuslon .. '" and disassembly ',~,,' RNA in the endosome '''--.. /''
Protein translation. polyprotein processing on membranes
Mature vIrus release .' pr
\-~~-~.' , ~-I ;
tI2 Virus maturation ~ Furin cleavage ~ ofprM ~ '~TGN ~~~====~ ~
c---c:= • ~.
Immature virus assembly
Viral RNA repl icatiol1 on membranes
Nucleus
Golgi
ER
Figure 1.5 The DENY replication cycle. Virions bind to cell-surface attachment molecules! receptors and are internalized through endocytosis. In the low pH of the endosome, viral glycoproteins mediate fusion of the viral and cellular membranes. allowing disassembly of the virion and release of its RNA into the cytoplasm. The viral RNA is translated into a polyprotein that is processed by viral and cellular proteases. Viral non-structural proteins then replicate the genomic RNA. Virion assembly occurs at the ER membrane. Capsid protein and viral RNA are enveloped by the membrane and its embedded glycoproteins to form immature virus pmticles. which are then transported through the secretory pathway. [n the low pH of the trans-Golgi network (TGN). prM is cleaved by furin. Mature virions are then released into the cytoplasm (Sampath. A. & Padmanabhan. R. 2009. Molecular targets for tlavivirus drug discovery. Antiviral Rt>s .. 81.6-15. Figure 2. page 21).
13

The viral RNA is directly translated into a single polyprotein by the host's
translational machinery. The processing of the polyprotein precursor occurs both
cotranslationally and post-translationally by host cell and virus-encoded proteases.
Host cell signalase located in the luminal side of the endoplasmic reticulum
(ER) is responsible for the cleavages at the C-prM, prM-E, E-NS1, and NS4A-NS4B
junctions (Chambers et at., 1990a; Henchal and Putnak, 1990). Previous work
suggest that NS I-NS2A cleavage occurs in the ER and NS2A is required to permit a
host ER-resident protease, possibly signalase to effect cleavage (Falgout and
Markoff, 1995).
The virus-encoded trypsin-like serine protease, a complex ofNS2B and NS3,
cleaves at a number of sites including the NS2A-NS2B, NS2B-NS3, NS3-NS4A, and
NA4B-NS5 junctions (Preugschat et af., 1990) (Figure 1.4). In addition, it is also
responsible for the cleavage within the viral protein C, NS4A, and within NS3 itself
(Teo and Wright, 1997). The viral RNA replication is catalyzed by a replication
complex which is composed of NS5, the RNA-dependent RNA polymerase, and
other viral and host factors in the rough ER and in Golgi-derived membranes called
vesicle packets (VP) (Mackenzie, 2005).
Newly synthesized RNA encapsulated by C protein is then enveloped by
glycoproteins prM and E to assemble immature virus particles that bud into the ER.
These immature particles are transported through the secretory pathway to the Golgi
apparatus. In the low pH environment of the trans-Golgi, furin-mediated cleavage of
prM to M drives maturation of the virus. prM processing destabilizes the prM-E
interaction and promotes the formation of E homodimers present in mature infectious
virions. Finally, progeny virus particles are released from the cell by exocytosis
(Henchal and Putnak, 1990; Perera et af., 2008; Sampath and Padmanabhan, 2009).
14

1.5 NS2B-NS3: The two-component protease of dengue virus
The NS2B-NS3 protease is a two-component protease. This heterodimeric
complex ofNS2B and NS3 is responsible for cleavage of the newly translated DENV
polyprotein at the NS2A-NS2B, NS2B-NS3, NS3-NS4A, and NS4B-NS5 sites as
well as internal sites within the viral protein C, NS2A, NS3, and NS4A (Chambers et
al., 1990a; Chambers et al., 1990b; Preugschat et al., 1990; Falgout et al., 1991;
Zhang et al., 1992; Clum et al., 1997; Yusof et al., 2000; Bera et al., 2007).
NS3 is a multifunctional protein as illustrated in Figure 1.6. The virus
encoded protease lies within the N-terminal 180 ammo acid residues of the 618
residue protein. The C-terminal region comprises the RNA-stimulated nucleoside
triphosphatase (NTPase) and RNA helicase activities. The functional domains of the
protease and NTPase overlap within a region of 20 amino acid residues (residues 160
to 180) (Li et al., 1999).
NS2814kDa NS3 69 kDa
NS3pro
NS28 NS3pro
* * * His51 Asp 75 Ser135 domain
! NS2B·NS3 cleavage ,ite
Figure 1.6 Structure and organization of the DENV NS2B-NS3 protease. (A) Scheme for viral enzymes, showing the cofactor domain of NS2B in black and the NS3pro in gray with the catalytic triad: His51, Asp75, andSerl35. (NaIL T. A .. Chappell, K. 1., Stoermer, M. J., Fang, N. X., Tyndall, 1. D., Young, P. R. & Fairlie, D. P. 2004. Enzymatic characterization and homology model of a catalytically active recombinant West Nile virus NS3 protease. J. Bioi. Chem., 279,48535-42. Figure 1, page 48536).
15

The serine protease domain of NS3 was identified based on sequence
homology to known senne proteases (Bazan and Fletterick, 1989). Four separate
regions of significant conservation were identified (boxes 1,2,3, and 4). Boxes 1,2,
and 3 encompass the catalytic triad (HisS I , Asp75, and Ser 135) and boxes 3 and 4
contain residues that are involved in substrate binding and recognition (Figure 1.7).
Subsequent biochemical studies confirmed the protease activity within the N-
terminal 180 amino acid residues of NS3 (Chambers et aI., 1990b; Preugschat et aI.,
1990). Site-directed mutagenesis experiments performed with YFV showed that
replacement of the putative catalytic triad residues abolished protease activity in
vitro. and when the changes were incurporated into the infectious full-length cDNA
clone. virus \\as not recovered (Chambers ('I (fl.. 1990b).
,-' "':"1) B c"] ,.:' #
DENV 1 ,'i~.,D,,:' E'IER :':~DD = ,R:!:, R __ R ,'/ ',' ',' . E,.'/,,~ l·;,ij'rR
DENV 2 '!~"D'i:"':';EKE~ED "RIK.R ~_ ',i " ':i:n·' .. H'rR DElN 3 : '/~"D'::'::E~,.K.E~EE '/R:K" = K ,'i ',' 'i. '/:,'U{"H'rR
\'J, '. KILE, ,,,
KRIEe),
rl tt
-. , "
"E E.RE ~',.:,.K
EE
EE EE
DENV 4 L,D'/~". ~,IL>E 'i.RIl-!.R __ re,'! - DK.,DKEED
D 150 1f,) 1'7,) lEi #
DENV 1 E~ lr.~IU!;:~~:',Ir.;E E'i ~ ,:1)'K:. j':!i.RE"V L: t: 'N ~~ C':V~.j.~,K-,o~,E , DENV 2 Ee K:~R,1!:~1C' I:rC,i~ ':1 r,'rcLD: c: - ~fIiDRK' IV LUW R3~ __ V~.-J-,. ~EK)IED' DEWl 3 EE lC'K:i::':'I·r;: 1: ,~<c~EII ,LD: f{;
DEWl 4 Ee K:; f\H,r :C'f{;~: ~ L ~ E:'.'-:- LD: Ire ~: I, iRE N L::: 'N K;,VC l :,;:~:,,1:rD E
,,= :,RK' \'I ~, :. V'i K, o.'.'or;.1:RI ErO
Figure 1.7 Sequences of DENV NSJ set-ine protease domain. Multiple sequence alignment of DENV protease domain from 4 serotypes. The catalytic triad residues: His51. Asp75. and Ser 135. are labeled \vith the symbol # are found in boxes labeled A. B. and C. The boxes labeled A. B. C. and D identifies regions of significant similarity surrounding the catalytic triad residues and residues that might form the substrate-binding pocket. (Aleshin. A. E .. Shiryaev, S. A .. Strongin. A, Y. & Liddington, R. C. 2007. Structural evidence for regulation and specificity of tlavivira! proteases and evolution of the Flaviviridae fold. Protein Sci" 16, 795-806. Figure I, page 796).
16
.11

NS2B is an ER-resident integral membrane protein. The protein contains 130
amino acid residues with the molecular mass of 14 kDa. The hydrophobicity plot of
NS2B shows that NS2B contains a central hydrophilic domain flanked by t\\'o
hydrophobic domains at the N-terminus (I and II) and C-terminus (III and IV)
(Figure 1.8) (Clum e{ al.. 1997). The hydrophobic sequences are essential for co-
translational insertion of the protease cofactor into ER membranes for etlicient
cleavage of the NS2B/NS3 junction (Clum et al .. 1997) but they are dispensable for
protease activity. The central hydrophilic region contains 40 amino acids which are
conserved among tlaviviruses. The individual NS3pro domain, lacking the NS2B
part. is catalytically inert (Murthy I!l al .. 1999: Murthy et ul .. 2(00).
DENV 1
DEN'l 2
DEN'! 3
DEN'! 4
DIl " =1= , X(RBHt netb¥ . "'1f1Pi"D-*~
_,E ULV IV,E,:":"':":"K D'!;:" ~:..: 1,::"L C 'J:' ~ E-.II!.'! },;'/':.::...- - ·:....~R_,D=.l!~ : -',- :..:.. ','C ','_
:".E 'n,;.v '- .. ,. _ _:"R D'," j,: _:.. = C " :
. :.. ,E 1M V :"V':":" :":"K DV: :..
R D 5:'
D 5:,
,-_______________ , nere ,mn DEN'l 1 _' LEK·c.EV:,;.EEEEH' 'h:,Ic.'iEV.DD "j,XIRDEE DENV 2 ' :"ER. DVR"ED;'EI' '::":LI'ED :1·>IKEEE . :""::":R ::':'
DEN'! 3 !·lURDDE'.
DB DH
DENV 1 ·.~:"LVS 'l.bL'oL-.':'UV.::, ; .. KKK:R :3J
DENV 2 L~VI.s V~ [\rSI~· I~·.·~··_ -.'" _ L,,'E'lKK~.R ::SCI
DElN 3 ,LLIVS It ~'. ;,r;,·,':'LL'!,.rl",.;,:.K: ':,:R 13'J DENV 4 _ .~I-:-·1/.s L ... [-L~_I-~r~'l·!'Y'L·;:~l,t:·:·.·VJC,_R :3·:!
Figure 1.8 Sequences of DENY NS2B. The box indicates the minimal cofactor segment required for activation of the NS3 protease ill vilro. TM 1-TM4 are predicted transmembrane regions (Aleshin. A. E.. Shiryaev. S. A .. Strongin. A. Y. & Liddington. R. C. 2007. Structural evidence for regulation and specificity of tlaviviral proteases and evolution of the Flaviviridae fold. Prulein Sci .. 16, 795-806. Figure I. page 796).
Results from co-expression studies showed that the proteolytic activity of the
NS3 protease was critically dependent upon the presence of its cotactor, NS2B
protein (Falgout et 01.. 1991). Truncation studies in DENV2 showed that the central
40 amino acid hydrophilic domain is sufficient for protease activity (Chambers et at.,
17

1993; Falgout et al., 1993; Clum et al., 1997; Niyomrattanakit et al., 2004). The
presence of NS28 resulted in a several thousand-fold activation of the NS3 protease
towards dibasic peptide substrates (Yusof et al., 2000). The flanking hydrophobic
domains within NS28 are likely to function in promoting membrane association of
NS28-NS3 (Clum et al., 1997).
The kinetic parameters and substrate specificity of DENY protease were
reported (Yusof et al., 2000; Leung et al., 2001; Khumthong et al., 2002;
Chanprapaph et al., 2005; Shiryaev et al., 2007a; Iempridee et aI., 2008). The
precursor devoid of the hydrophobic regions but containing the conserved NS28
hydrophilic domain linked to the NS3 protease domain through a carboxy terminal
region of NS28 containing the NS28-NS3 cleavage site was expressed in E.coli
(Yusof et al., 2000). The precursor, expressed as insoluble inclusion bodies, was
purified by denaturation and refolding.
The expression of soluble and active protease was achieved when the
hydrophilic portion of the NS28 viral cofactor spanning residues 49-95 (hereafter
named CF40) of either WNY or DENY2 was fused to residues 1-169 of the NS3
protein via a flexible (GlY4-Ser-GlY4) linker, thus obviating the denaturation and
refolding steps in the purification of the protease (Leung et al., 2001).
A number of in vitro assays for the viral proteases have been described in
several studies (Clum et al., 1997; Yusof et al., 2000; Leung et aI., 2001; Walker and
Lynas, 2001; Khumthong et al., 2002; Tong, 2002). Either virus-encoded polyprotp.in
or synthetic peptides have been utilized as the substrates. Important information on
the regulation and requirements for the viral polyprotein processing were obtained
from the assay with virus-encoded polyprotein. The assay with the synthetic peptides
18

would provide the information on the substrate specificity of the enzymes and were
used in the inhibitor screening.
The viral protease has a preference for two basic amino acid residues (Arg
Arg, Arg-Lys, Lys-Arg, or occasionally Gln-Arg) at the P2 and PI positions
preceding the cleavage sites, followed by Gly, Ala, or Ser at the PI' position. The
earliest report for the DENY protease in vitro assay had used commercially available
fluorogenic peptides as the substrates.
All of these peptides contain two basic amino acid residues (Arg-Arg, Arg
Lys, Lys-Arg) at the PI and P2 positions preceding the cleavage site. None of the
peptides contain an amino acid residue at the PI' position, but rather the PI' residue
is replaced by a tluorogenic moiety. Their result revealed that the substrate Gly-Arg
Arg-MCA, which contains a Gly residue at the P3 position, is the most active of the
four substrates tested (Yusof et al., 2000).
Li et al. (2005) cloned and expressed the protease from all four DENY
serotypes (DENYI-4 CF40-GlY4-Ser-GlY4-NS3pro) and adapted the in vitro assay
described by Yusof et al. (2000) to screen tetrapeptide and octapeptide libraries
comprising ~ 13,000 substrates.
The tetrapeptide benzoyl-norleucine (P4 )-Iysine (P3)-arginine (P2)-arginine
(P1)-ACMC (Bz-Nle-Lys-Arg-Arg-ACMC) was identified as the optimal substrate
with the steady state kinetics parameter kca/Km of 51,800 M'ls'l which is >150-fold
more sensitive than other published peptides. The sensitivity enabled miniaturization
of the assay for high-throughput screening (Keller et al., 2006). Moreover, this ideal
tetrapeptide sequence formed the basis for the peptidomimetic approach for finding
potent substrate-based inhibitors (Yin et al., 2006a; Yin et al., 2006b).
19

1.6 Structure of DEN V NS3 serine protease
The high-quality crystal structure of active DENY NS2B-NS3 protease (1.5
A; PDB identifier 2FOM) and WNY NS2B-NS3 protease in the complex with the
substrate-based inhibitor Bz-Nle-Lys-Arg-Arg-H (1.68 A; PDB identifier 2FP7) were
resolved (D'Arcy et ([/.. 2006; Erbel et a/.. 2006). The NS3 protease domains in both
structures adopt chymotrypsin-like serine protease folds with two ~-barrels, each
formed by six ~-strands. and the catalytic triad (HisS 1 , Asp75. and Ser 135) located at
the cleft between the two ~-barrels (Figure 1.9). Sharing 50% sequence identity. the
two NS2B-NS3 protease structures have close structural similarity.
[ill
F2·' B2tJ C2
B2a
E1b
DENV2 NS2B-NS3pro
88
76 85
E1b
I~ ,,' F 1 .19 E i Cl
• • " r11./ 01 f\!'f-' ~!. C 1
E2"t.,'., ' In tt--, ,I» ~1t
W')I' ,,1 t ~11 1'J B2t> r·;d. E2d A1 0.2' D2 J'
WNV NS2B-NS3pro
Figure 1.9 Schematic representation of the structures NS2B-NS3 protease, (Left) The apo-enzyme from DENV2 with the NS28 cofactor in yellow and the catalytic triad represented as sticks. (Right) Complex of WNV NS28-NS3 protease with the KKRR tetrapeptide. The N-terminal part of the NS28 cofactor is sufficient to stabilize the enzyme (Erbel. P .. Schiering, N., D'arcy. A .. Renatus. M .. Kroemer, M .. Lim. S. P., Yin. Z., Keller. T. H .. Vasudevan. S. G. & Hommel. U. 2006. Structural basis for the activation of tlaviviral NS3 proteases from dengue and West Nile virus. Nat, StruC{, Mol. Bio/., 13,372-3, Figure I. page 372).
20

The structures of WNY and DENY NS2B-NS3 proteases reveal residues of
NS2B that are important for the stabilization of the NS3 protease fold. Similarly to
the HCY NS4A-NS3 protease, the N-terminal part of the cofactor contributes one p
strand (p-strand 1, NS2B residues 51-57 in DENY) to the N-terminal p-barrel of the
protease, which conceals hydrophobic residues from the solvent and provides
stabilization to this domain.
This explains the observed strong tendency for NS3 protease and full-length
NS3 to aggregate when the strand contributed by NS2B is absent in synthetic
constructs. In this respect the N-terminal of NS2B has a chaperone-like role in
stabilizing NS3.
On the other hand. the fold adopted by the C-terminal part of the NS2B
cofactor shows marked differences between the unliganded DENY NS2B-NS3pro
and inhibitor-bound WNY NS2B-NS3pro complexes. In the inhibitor-bound protease
complex, a large rearrangement brings residues 67-88 ofNS2B in close proximity to
the substrate-like inhibitor, forming a belt around the NS3 protease. Residues Arg78-
Leu87 of the NS2B cofactor forms a P-Ioop which interacts with the N-terminal
barrel of the NS3 protease, affecting the formation of the active site and substrate
recognition.
The contribution of the NS2B cofactor to stabilize both the N- and C-terminal
barrels and complete the substrate-binding site is indeed unique to flaviviruses. It
differs substantially from those observed with other cofactor-activated viral proteases
such as HCY NS4A-NS3pro which requires a short fragment of NS4A to form the
active enzyme (Erbel et al., 2006; Lescar et al., 2008).
The unprecedented way in which the NS2B cofactor region forms a belt
around the protease domain was confirmed in a second structure that was reported
21

for the WNY enzyme as a complex with the aprotinin/BPTI inhibitor (Aleshin et al.,
2007). The aprotinin occupied all the specificity pockets of the protease and induced
a fully formed oxyanion hole, which allowed Aleshin et al. (2007) to provide a
complete view of the enzyme substrate Michaelis complex for a flavivirus protease.
These structures open up new opportunities for discovering flavivirus-specific drugs
that could function by interfering with protein-protein interactions that are needed for
the activation of the protease in addition to active site directed competitive inhibitors.
The DENY NS3 protease structure in the absence of the NS2B cofactor
deviates substantially from DENY NS2B-NS3 protease structures. These differences
are observed throughout the entire enzyme and affect the length and location of
secondary structure elements (Erbel et at., 2006). Although the protease catalytic site
residues (His51, Asp75, and Serl35) were arranged similarly in the NS3 and NS2B
NS3 protease crystal structures, numerous large conformational differences were
evident.
For instance, overlaying the catalytic regions of the two structures resulted in
position differences of 14A and 35 A for Leu31 and Asn1I9, respectively (Figure
1.10). Of relevance to DENY protease inhibitor design are the large differences in
the substrate-binding region between the two structures. The S 1 site within the NS3
substrate-binding region formed a deep pocket that could accommodate long
positively charged PI side chains of the substrate. However, in the NS2B-NS3
protease structure, the S 1 site forms only a shallow depression. Structure-based drug
discovery approaches must consider the differences between the NS3 and NS2B-NS3
structures since small molecules may interact differently with the active sites ofNS3
and NS2B-NS3 (Tomlinson et al., 2009b).
22
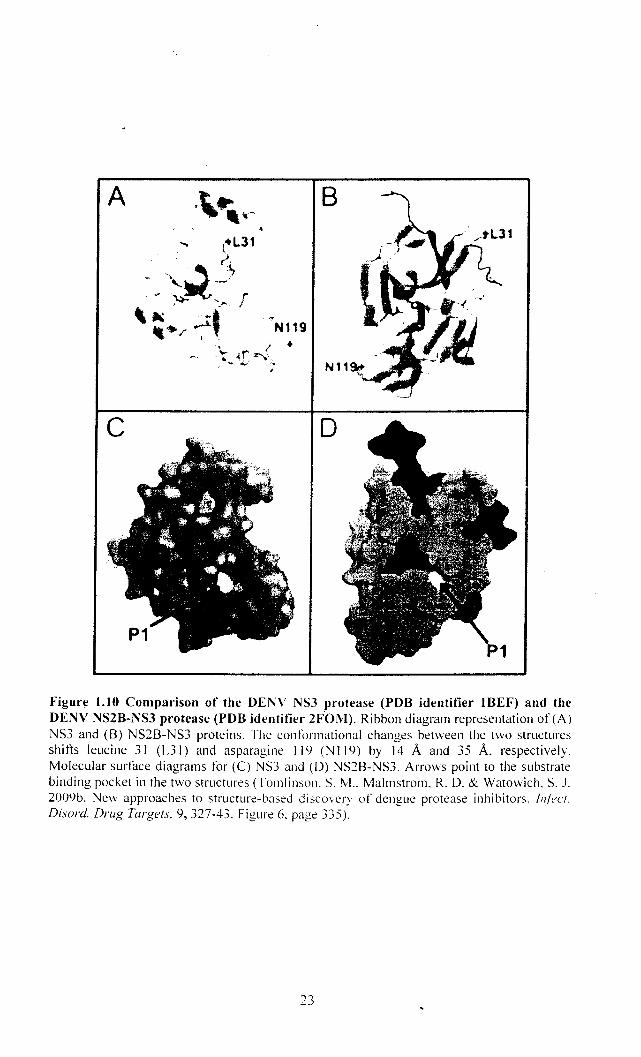
A
c
~N119
\.. I .. ~r ~.' -..;../ .... , •
o
Figure l.1O Comparison of the DENV NS3 protease (PDB identifier IBEF) and the DENV NS2B-NS3 protease (PDB identifier 2FOM). Ribbon diagram representation of (A) NS3 and (B) NS2B-NS3 proteins. The conformational changes between the two structures shifts leucine 31 (L3I) and asparagine 119 (NI19) by 14 A and 35 A. respectively. Molecular surface diagrams for (e) NS3 and (D) NS2B-NS3. Arrows point to the substrate binding pocket in the two structures (Tomlinson. S. M .. Malmstrom. R. O. & Watowich. S. J. 2009b. Nevv approaches to structure-based discovery of dengue protease inhibitors. IIl(ecT.
Disord. Drug Targets. 9,327-43. Figure 6. page 335).

1. 7 Serine protease
Proteases have long been recognized as attractive targets in the drug
discovery processes. Serine proteases are the most widely studied group of proteins
in biology (Walker and Lynas, 200 I). The important role of serine proteases has been
elucidated in the pathology of viral infections (Tong, 2002). Many crystal structures
and their complexes with either substrates or inhibitors have been resolved (Kim et
aI., 1996; Erbel et aI., 2006; Aleshin et al., 2007).
In humans, serine proteases are involved in many important physiological
processes such as inflammation, fibrinolysis, immune response, digestion, blood
coagulation, and fertilization. Hence, it is critical that whenever an inhibitor of viral
serine proteases is utilized for the treatment of disease in humans, such inhibitor
must have a high selectively for the viral protease to minimize the risk of any adverse
effects.
Proteases are enzymes that selectively catalyze the hydrolysis of peptide
bonds. Proteases are classified into five major classes based on their mechanism of
action. These classes are serine proteases, cysteine proteases, aspartic proteases,
threonine proteases, and metallo proteases. The classification is made owing to the
critical residues used in catalysis.
Serine proteases are characterized chiefly by the presence of an active site
serine (Ser) residue, the y hydroxyl group of which acts as a nucleophile during the
hydrolytic process. Two other amino acid residues that are directly involved in the
catalytic mechanism are histidine (His) and aspartate (Asp) that together form the
catalytic triad (Walker and Lynas, 2001; Hedstrom, 2002). In addition, the enzyme
possesses an oxyanion binding site that is made from the backbone amide NH groups
24