reassessing the biogenicity of earth s oldest trace search ... · reassessing the biogenicity of...
TRANSCRIPT

Reassessing the biogenicity of Earth’s oldest tracefossil with implications for biosignatures in thesearch for early lifeEugene G. Grosch1 and Nicola McLoughlin
Department of Earth Science and Centre for Geobiology, University of Bergen, N-5007 Bergen, Norway
Edited* by Norman H. Sleep, Stanford University, Stanford, CA, and approved April 28, 2014 (received for review February 10, 2014)
Microtextures in metavolcanic pillow lavas from the Barbertongreenstone belt of South Africa have been argued to representEarth’s oldest trace fossil, preserving evidence for microbial lifein the Paleoarchean subseafloor. In this study we present new insitu U–Pb age, metamorphic, and morphological data on thesetitanite microtextures from fresh drill cores intercepting the typelocality. A filamentous microtexture representing a candidate bio-signature yields a U–Pb titanite age of 2.819 ± 0.2 Ga. In the samedrill core hornfelsic-textured titanite discovered adjacent to a localmafic sill records an indistinguishable U–Pb age of 2.913 ± 0.31 Ga,overlapping with the estimated age of intrusion. Quantitative mi-croscale compositional mapping, combined with chlorite thermo-dynamic modeling, reveals that the titanite filaments are bestdeveloped in relatively low-temperature microdomains of thechlorite matrix. We find that the microtextures exhibit a morpho-logical continuum that bears no similarity to candidate biotexturesfound in the modern oceanic crust. These new findings indicatethat the titanite formed during late Archean ca. 2.9 Ga thermalcontact metamorphism and not in an early ca. 3.45 Ga subseafloorenvironment. We therefore question the syngenicity and biogenic-ity of these purported trace fossils. It is argued herein that thetitanite microtextures are more likely abiotic porphyroblasts ofthermal contact metamorphic origin that record late-stage retro-grade cooling in the pillow lava country rock. A full characteriza-tion of low-temperature metamorphic events and alternativebiosignatures in greenstone belt pillow lavas is thus requiredbefore candidate traces of life can be confirmed in Archeansubseafloor environments.
Archean habitats | astrobiology | Archean Earth | ichnofossil | bioalteration
F ilamentous titanite microtextures in ca. 3.472–3.432 Gametavolcanic pillow lavas of the Barberton greenstone belt
(BGB), South Africa, have been argued to represent Earth’soldest trace fossils (1–4). Subsequent work in other Archeanterrains, such as the Pilbara Craton of Western Australia and theAbitibi greenstone belt of Canada, have made similar claims fortrace fossils in early metabasaltic pillow lavas (5, 6). The currentparadigm for these titanite microtextures involves a complexbioalteration model in which endolithic microbes form hollowmicrotunnels by etching fresh volcanic glass in submarine envi-ronments on the early Earth (1–7). Subseafloor hydrothermalalteration is envisioned not only to provide a suitable environ-ment for microbial activity, but also to result in mineralization ofthe hollow tubes by titanite (CaTiSiO5) preserving the proposedtrace fossils (1–3). Thus, it has been suggested that volcanichabitats represent a previously unexplored geological setting inwhich life may have thrived and possibly originated on the earlyEarth (1–3). It has also been argued that similar microtextures inaltered extraterrestrial basalts may provide a useful biosignaturein the search for life on Mars and beyond (8).The principal argument for a biogenic origin of the purported
Archean trace fossils is based on their apparent similarity in size,shape, and distribution to partially mineralized microtubules of
argued biogenic origin from pillow lavas of the in situ oceaniccrust and ophiolites (1, 2, 9). In young, in situ pillow lavas,abundant microbial DNA and geochemical signatures supportthe presence of a deep subseafloor biosphere (10, 11). Giventherefore the major implications that the Archean titanitemicrotextures may hold for the earliest evidence of life on Earth,we have conducted syngenicity and biogenicity tests to evaluatea proposed subseafloor bioalteration model for their origin. Wereport in situ U–Pb dating of the titanite, quantitative microscalemapping of metamorphic conditions, and morphological evi-dence to evaluate the biogenicity of the titanite microtextures.
ResultsThe titanite microtextures were originally described from pillowlavas of the ca. 3.472-Ga Hooggenoeg Formation of the BGBthat stratigraphically overlies ca. 3.482 Ga ultramafic komatiitesof the Komati Formation (Fig. S1). The current study includessamples from drill core KD2b of the Barberton Scientific Dril-ling Project (BSDP) that intercepted the pillow lava type sectionfor the proposed biotextures (12, 13) (Fig. S1). Petrographicinvestigation of the drill core samples reveals that the originaltitanite bioalteration textures (1) form part of a morphologicalcontinuum that was previously only partially illustrated. The size,morphology, and distribution of these titanite microtextures areevaluated against biogenicity criteria proposed for endolithicmicroborings (14, 15). The simplest morphological endmemberis hornfelsic titanite spheres (Fig. 1 A–E) that may coalesce toform irregular bands. The spheres and bands develop filamentous
Significance
It has been argued that Archean subseafloor pillow lavasequences provide an environment in which to seek evidencefor the earliest traces of life. Candidate titanite biosignaturesof microbial activity have been reported in ∼3.45-Ga meta-volcanic glass from the Barberton greenstone belt of SouthAfrica. In this paper we present new in situ U–Pb age data,metamorphic constraints, and morphological observations onthese titanite microtextures. Our data challenges a biologicalorigin for these oldest purported trace fossils, with implicationsfor the ecological niches where life may have first emerged. Wetherefore suggest alternative biosignatures and approachesshould be considered in the search for subsurface life on earlyEarth and in extraterrestrial mafic–ultramafic rocks, for exam-ple, in martian basalts.
Author contributions: E.G.G. designed research; E.G.G. and N.M. performed research; E.G.G.and N.M. contributed new reagents/analytic tools; E.G.G. and N.M. analyzed data; andE.G.G. wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.
Freely available online through the PNAS open access option.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1402565111/-/DCSupplemental.
8380–8385 | PNAS | June 10, 2014 | vol. 111 | no. 23 www.pnas.org/cgi/doi/10.1073/pnas.1402565111
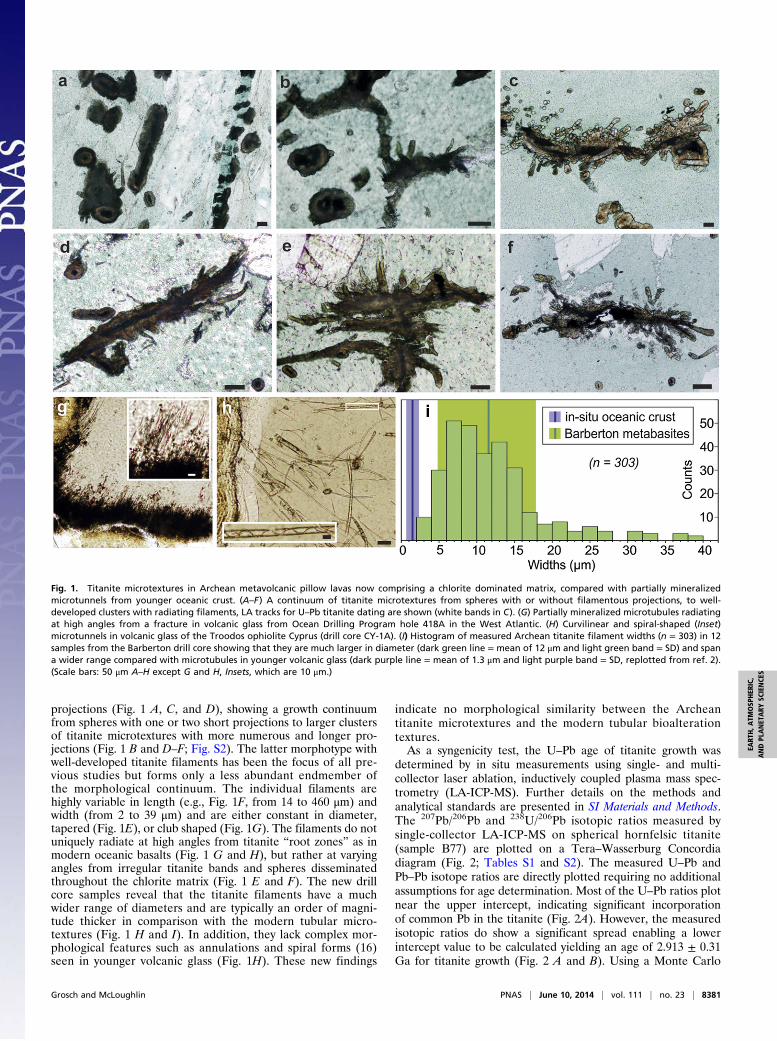
projections (Fig. 1 A, C, and D), showing a growth continuumfrom spheres with one or two short projections to larger clustersof titanite microtextures with more numerous and longer pro-jections (Fig. 1 B and D–F; Fig. S2). The latter morphotype withwell-developed titanite filaments has been the focus of all pre-vious studies but forms only a less abundant endmember ofthe morphological continuum. The individual filaments arehighly variable in length (e.g., Fig. 1F, from 14 to 460 μm) andwidth (from 2 to 39 μm) and are either constant in diameter,tapered (Fig. 1E), or club shaped (Fig. 1G). The filaments do notuniquely radiate at high angles from titanite “root zones” as inmodern oceanic basalts (Fig. 1 G and H), but rather at varyingangles from irregular titanite bands and spheres disseminatedthroughout the chlorite matrix (Fig. 1 E and F). The new drillcore samples reveal that the titanite filaments have a muchwider range of diameters and are typically an order of magni-tude thicker in comparison with the modern tubular micro-textures (Fig. 1 H and I). In addition, they lack complex mor-phological features such as annulations and spiral forms (16)seen in younger volcanic glass (Fig. 1H). These new findings
indicate no morphological similarity between the Archeantitanite microtextures and the modern tubular bioalterationtextures.As a syngenicity test, the U–Pb age of titanite growth was
determined by in situ measurements using single- and multi-collector laser ablation, inductively coupled plasma mass spec-trometry (LA-ICP-MS). Further details on the methods andanalytical standards are presented in SI Materials and Methods.The 207Pb/206Pb and 238U/206Pb isotopic ratios measured bysingle-collector LA-ICP-MS on spherical hornfelsic titanite(sample B77) are plotted on a Tera–Wasserburg Concordiadiagram (Fig. 2; Tables S1 and S2). The measured U–Pb andPb–Pb isotope ratios are directly plotted requiring no additionalassumptions for age determination. Most of the U–Pb ratios plotnear the upper intercept, indicating significant incorporationof common Pb in the titanite (Fig. 2A). However, the measuredisotopic ratios do show a significant spread enabling a lowerintercept value to be calculated yielding an age of 2.913 ± 0.31Ga for titanite growth (Fig. 2 A and B). Using a Monte Carlo
Fig. 1. Titanite microtextures in Archean metavolcanic pillow lavas now comprising a chlorite dominated matrix, compared with partially mineralizedmicrotunnels from younger oceanic crust. (A–F) A continuum of titanite microtextures from spheres with or without filamentous projections, to well-developed clusters with radiating filaments, LA tracks for U–Pb titanite dating are shown (white bands in C). (G) Partially mineralized microtubules radiatingat high angles from a fracture in volcanic glass from Ocean Drilling Program hole 418A in the West Atlantic. (H) Curvilinear and spiral-shaped (Inset)microtunnels in volcanic glass of the Troodos ophiolite Cyprus (drill core CY-1A). (I) Histogram of measured Archean titanite filament widths (n = 303) in 12samples from the Barberton drill core showing that they are much larger in diameter (dark green line = mean of 12 μm and light green band = SD) and spana wider range compared with microtubules in younger volcanic glass (dark purple line = mean of 1.3 μm and light purple band = SD, replotted from ref. 2).(Scale bars: 50 μm A–H except G and H, Insets, which are 10 μm.)
Grosch and McLoughlin PNAS | June 10, 2014 | vol. 111 | no. 23 | 8381
EART
H,A
TMOSP
HER
IC,
ANDPL
ANET
ARY
SCIENCE
S
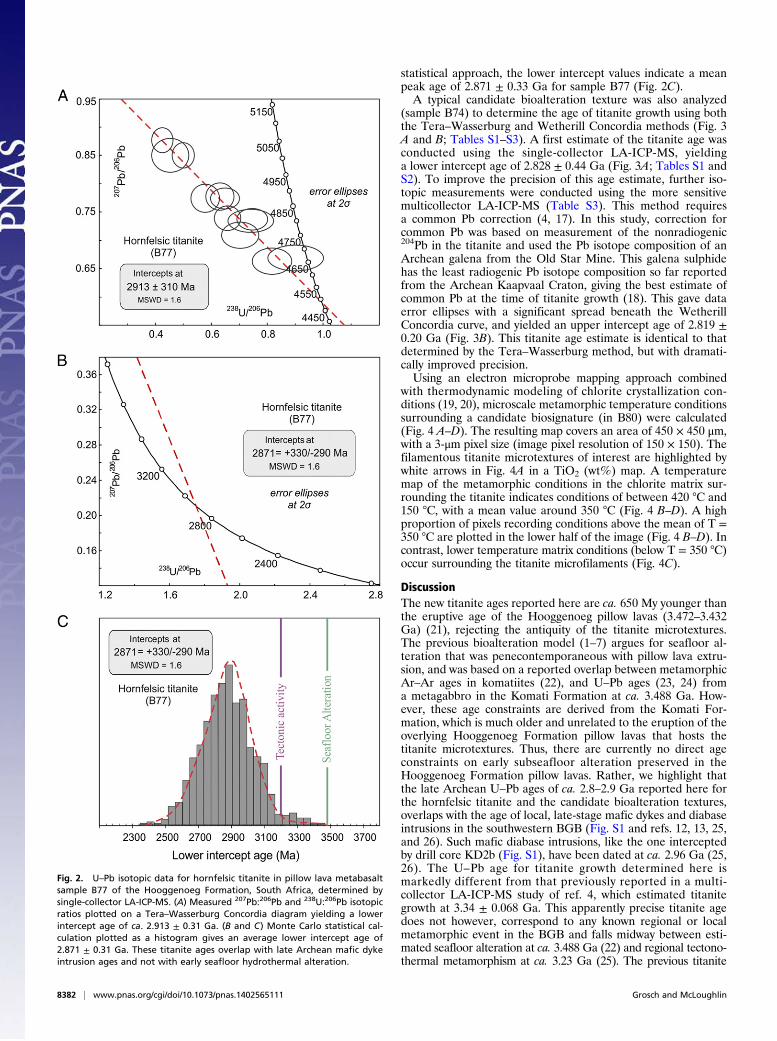
statistical approach, the lower intercept values indicate a meanpeak age of 2.871 ± 0.33 Ga for sample B77 (Fig. 2C).A typical candidate bioalteration texture was also analyzed
(sample B74) to determine the age of titanite growth using boththe Tera–Wasserburg and Wetherill Concordia methods (Fig. 3A and B; Tables S1–S3). A first estimate of the titanite age wasconducted using the single-collector LA-ICP-MS, yieldinga lower intercept age of 2.828 ± 0.44 Ga (Fig. 3A; Tables S1 andS2). To improve the precision of this age estimate, further iso-topic measurements were conducted using the more sensitivemulticollector LA-ICP-MS (Table S3). This method requiresa common Pb correction (4, 17). In this study, correction forcommon Pb was based on measurement of the nonradiogenic204Pb in the titanite and used the Pb isotope composition of anArchean galena from the Old Star Mine. This galena sulphidehas the least radiogenic Pb isotope composition so far reportedfrom the Archean Kaapvaal Craton, giving the best estimate ofcommon Pb at the time of titanite growth (18). This gave dataerror ellipses with a significant spread beneath the WetherillConcordia curve, and yielded an upper intercept age of 2.819 ±0.20 Ga (Fig. 3B). This titanite age estimate is identical to thatdetermined by the Tera–Wasserburg method, but with dramati-cally improved precision.Using an electron microprobe mapping approach combined
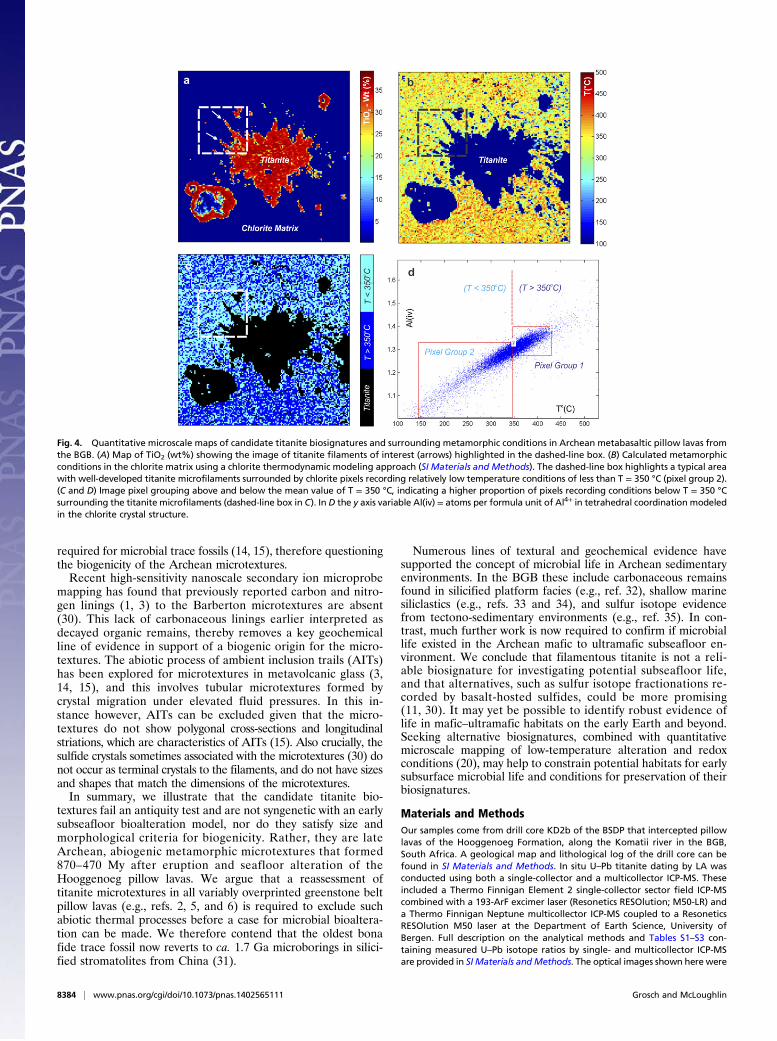
with thermodynamic modeling of chlorite crystallization con-ditions (19, 20), microscale metamorphic temperature conditionssurrounding a candidate biosignature (in B80) were calculated(Fig. 4 A–D). The resulting map covers an area of 450 × 450 μm,with a 3-μm pixel size (image pixel resolution of 150 × 150). Thefilamentous titanite microtextures of interest are highlighted bywhite arrows in Fig. 4A in a TiO2 (wt%) map. A temperaturemap of the metamorphic conditions in the chlorite matrix sur-rounding the titanite indicates conditions of between 420 °C and150 °C, with a mean value around 350 °C (Fig. 4 B–D). A highproportion of pixels recording conditions above the mean of T =350 °C are plotted in the lower half of the image (Fig. 4 B–D). Incontrast, lower temperature matrix conditions (below T = 350 °C)occur surrounding the titanite microfilaments (Fig. 4C).
DiscussionThe new titanite ages reported here are ca. 650 My younger thanthe eruptive age of the Hooggenoeg pillow lavas (3.472–3.432Ga) (21), rejecting the antiquity of the titanite microtextures.The previous bioalteration model (1–7) argues for seafloor al-teration that was penecontemporaneous with pillow lava extru-sion, and was based on a reported overlap between metamorphicAr–Ar ages in komatiites (22), and U–Pb ages (23, 24) froma metagabbro in the Komati Formation at ca. 3.488 Ga. How-ever, these age constraints are derived from the Komati For-mation, which is much older and unrelated to the eruption of theoverlying Hooggenoeg Formation pillow lavas that hosts thetitanite microtextures. Thus, there are currently no direct ageconstraints on early subseafloor alteration preserved in theHooggenoeg Formation pillow lavas. Rather, we highlight thatthe late Archean U–Pb ages of ca. 2.8–2.9 Ga reported here forthe hornfelsic titanite and the candidate bioalteration textures,overlaps with the age of local, late-stage mafic dykes and diabaseintrusions in the southwestern BGB (Fig. S1 and refs. 12, 13, 25,and 26). Such mafic diabase intrusions, like the one interceptedby drill core KD2b (Fig. S1), have been dated at ca. 2.96 Ga (25,26). The U–Pb age for titanite growth determined here ismarkedly different from that previously reported in a multi-collector LA-ICP-MS study of ref. 4, which estimated titanitegrowth at 3.34 ± 0.068 Ga. This apparently precise titanite agedoes not however, correspond to any known regional or localmetamorphic event in the BGB and falls midway between esti-mated seafloor alteration at ca. 3.488 Ga (22) and regional tectono-thermal metamorphism at ca. 3.23 Ga (25). The previous titanite
Fig. 2. U–Pb isotopic data for hornfelsic titanite in pillow lava metabasaltsample B77 of the Hooggenoeg Formation, South Africa, determined bysingle-collector LA-ICP-MS. (A) Measured 207Pb:206Pb and 238U:206Pb isotopicratios plotted on a Tera–Wasserburg Concordia diagram yielding a lowerintercept age of ca. 2.913 ± 0.31 Ga. (B and C) Monte Carlo statistical cal-culation plotted as a histogram gives an average lower intercept age of2.871 ± 0.31 Ga. These titanite ages overlap with late Archean mafic dykeintrusion ages and not with early seafloor hydrothermal alteration.
8382 | www.pnas.org/cgi/doi/10.1073/pnas.1402565111 Grosch and McLoughlin
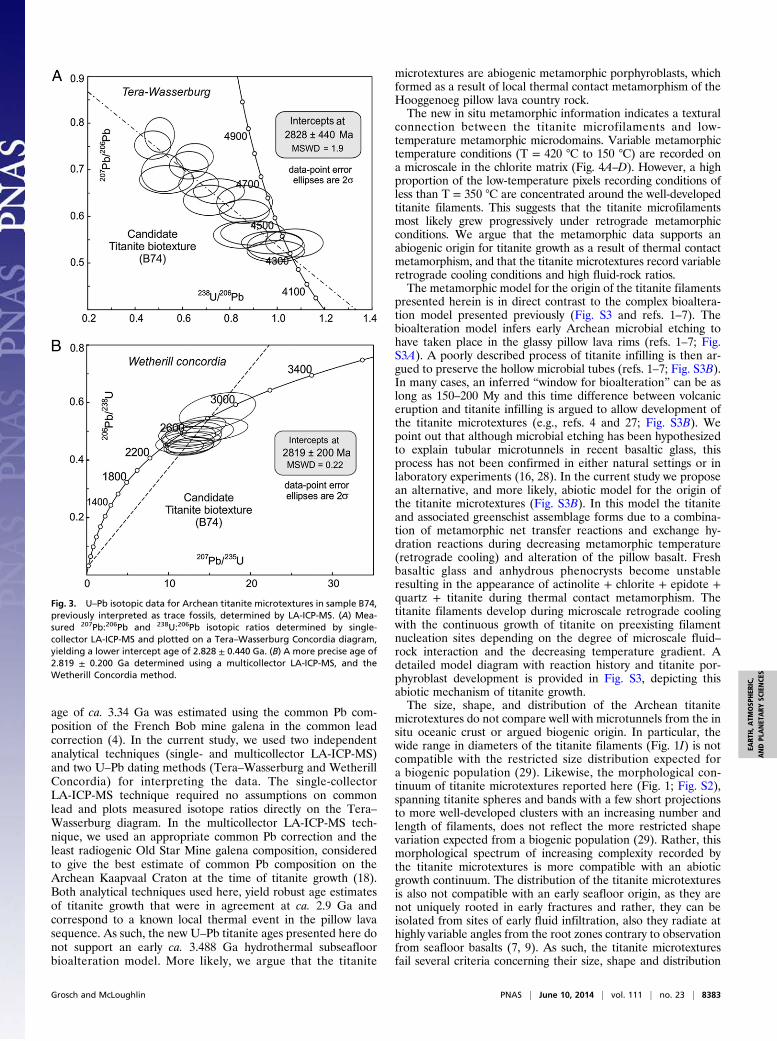
age of ca. 3.34 Ga was estimated using the common Pb com-position of the French Bob mine galena in the common leadcorrection (4). In the current study, we used two independentanalytical techniques (single- and multicollector LA-ICP-MS)and two U–Pb dating methods (Tera–Wasserburg and WetherillConcordia) for interpreting the data. The single-collectorLA-ICP-MS technique required no assumptions on commonlead and plots measured isotope ratios directly on the Tera–Wasserburg diagram. In the multicollector LA-ICP-MS tech-nique, we used an appropriate common Pb correction and theleast radiogenic Old Star Mine galena composition, consideredto give the best estimate of common Pb composition on theArchean Kaapvaal Craton at the time of titanite growth (18).Both analytical techniques used here, yield robust age estimatesof titanite growth that were in agreement at ca. 2.9 Ga andcorrespond to a known local thermal event in the pillow lavasequence. As such, the new U–Pb titanite ages presented here donot support an early ca. 3.488 Ga hydrothermal subseafloorbioalteration model. More likely, we argue that the titanite
microtextures are abiogenic metamorphic porphyroblasts, whichformed as a result of local thermal contact metamorphism of theHooggenoeg pillow lava country rock.The new in situ metamorphic information indicates a textural
connection between the titanite microfilaments and low-temperature metamorphic microdomains. Variable metamorphictemperature conditions (T = 420 °C to 150 °C) are recorded ona microscale in the chlorite matrix (Fig. 4A–D). However, a highproportion of the low-temperature pixels recording conditions ofless than T = 350 °C are concentrated around the well-developedtitanite filaments. This suggests that the titanite microfilamentsmost likely grew progressively under retrograde metamorphicconditions. We argue that the metamorphic data supports anabiogenic origin for titanite growth as a result of thermal contactmetamorphism, and that the titanite microtextures record variableretrograde cooling conditions and high fluid-rock ratios.The metamorphic model for the origin of the titanite filaments
presented herein is in direct contrast to the complex bioaltera-tion model presented previously (Fig. S3 and refs. 1–7). Thebioalteration model infers early Archean microbial etching tohave taken place in the glassy pillow lava rims (refs. 1–7; Fig.S3A). A poorly described process of titanite infilling is then ar-gued to preserve the hollow microbial tubes (refs. 1–7; Fig. S3B).In many cases, an inferred “window for bioalteration” can be aslong as 150–200 My and this time difference between volcaniceruption and titanite infilling is argued to allow development ofthe titanite microtextures (e.g., refs. 4 and 27; Fig. S3B). Wepoint out that although microbial etching has been hypothesizedto explain tubular microtunnels in recent basaltic glass, thisprocess has not been confirmed in either natural settings or inlaboratory experiments (16, 28). In the current study we proposean alternative, and more likely, abiotic model for the origin ofthe titanite microtextures (Fig. S3B). In this model the titaniteand associated greenschist assemblage forms due to a combina-tion of metamorphic net transfer reactions and exchange hy-dration reactions during decreasing metamorphic temperature(retrograde cooling) and alteration of the pillow basalt. Freshbasaltic glass and anhydrous phenocrysts become unstableresulting in the appearance of actinolite + chlorite + epidote +quartz + titanite during thermal contact metamorphism. Thetitanite filaments develop during microscale retrograde coolingwith the continuous growth of titanite on preexisting filamentnucleation sites depending on the degree of microscale fluid–rock interaction and the decreasing temperature gradient. Adetailed model diagram with reaction history and titanite por-phyroblast development is provided in Fig. S3, depicting thisabiotic mechanism of titanite growth.The size, shape, and distribution of the Archean titanite
microtextures do not compare well with microtunnels from the insitu oceanic crust or argued biogenic origin. In particular, thewide range in diameters of the titanite filaments (Fig. 1I) is notcompatible with the restricted size distribution expected fora biogenic population (29). Likewise, the morphological con-tinuum of titanite microtextures reported here (Fig. 1; Fig. S2),spanning titanite spheres and bands with a few short projectionsto more well-developed clusters with an increasing number andlength of filaments, does not reflect the more restricted shapevariation expected from a biogenic population (29). Rather, thismorphological spectrum of increasing complexity recorded bythe titanite microtextures is more compatible with an abioticgrowth continuum. The distribution of the titanite microtexturesis also not compatible with an early seafloor origin, as they arenot uniquely rooted in early fractures and rather, they can beisolated from sites of early fluid infiltration, also they radiate athighly variable angles from the root zones contrary to observationfrom seafloor basalts (7, 9). As such, the titanite microtexturesfail several criteria concerning their size, shape and distribution
Fig. 3. U–Pb isotopic data for Archean titanite microtextures in sample B74,previously interpreted as trace fossils, determined by LA-ICP-MS. (A) Mea-sured 207Pb:206Pb and 238U:206Pb isotopic ratios determined by single-collector LA-ICP-MS and plotted on a Tera–Wasserburg Concordia diagram,yielding a lower intercept age of 2.828 ± 0.440 Ga. (B) A more precise age of2.819 ± 0.200 Ga determined using a multicollector LA-ICP-MS, and theWetherill Concordia method.
Grosch and McLoughlin PNAS | June 10, 2014 | vol. 111 | no. 23 | 8383
EART
H,A
TMOSP
HER
IC,
ANDPL
ANET
ARY
SCIENCE
S
required for microbial trace fossils (14, 15), therefore questioningthe biogenicity of the Archean microtextures.Recent high-sensitivity nanoscale secondary ion microprobe
mapping has found that previously reported carbon and nitro-gen linings (1, 3) to the Barberton microtextures are absent(30). This lack of carbonaceous linings earlier interpreted asdecayed organic remains, thereby removes a key geochemicalline of evidence in support of a biogenic origin for the micro-textures. The abiotic process of ambient inclusion trails (AITs)has been explored for microtextures in metavolcanic glass (3,14, 15), and this involves tubular microtextures formed bycrystal migration under elevated fluid pressures. In this in-stance however, AITs can be excluded given that the micro-textures do not show polygonal cross-sections and longitudinalstriations, which are characteristics of AITs (15). Also crucially, thesulfide crystals sometimes associated with the microtextures (30) donot occur as terminal crystals to the filaments, and do not have sizesand shapes that match the dimensions of the microtextures.In summary, we illustrate that the candidate titanite bio-
textures fail an antiquity test and are not syngenetic with an earlysubseafloor bioalteration model, nor do they satisfy size andmorphological criteria for biogenicity. Rather, they are lateArchean, abiogenic metamorphic microtextures that formed870–470 My after eruption and seafloor alteration of theHooggenoeg pillow lavas. We argue that a reassessment oftitanite microtextures in all variably overprinted greenstone beltpillow lavas (e.g., refs. 2, 5, and 6) is required to exclude suchabiotic thermal processes before a case for microbial bioaltera-tion can be made. We therefore contend that the oldest bonafide trace fossil now reverts to ca. 1.7 Ga microborings in silici-fied stromatolites from China (31).
Numerous lines of textural and geochemical evidence havesupported the concept of microbial life in Archean sedimentaryenvironments. In the BGB these include carbonaceous remainsfound in silicified platform facies (e.g., ref. 32), shallow marinesiliclastics (e.g., refs. 33 and 34), and sulfur isotope evidencefrom tectono-sedimentary environments (e.g., ref. 35). In con-trast, much further work is now required to confirm if microbiallife existed in the Archean mafic to ultramafic subseafloor en-vironment. We conclude that filamentous titanite is not a reli-able biosignature for investigating potential subseafloor life,and that alternatives, such as sulfur isotope fractionations re-corded by basalt-hosted sulfides, could be more promising(11, 30). It may yet be possible to identify robust evidence oflife in mafic–ultramafic habitats on the early Earth and beyond.Seeking alternative biosignatures, combined with quantitativemicroscale mapping of low-temperature alteration and redoxconditions (20), may help to constrain potential habitats for earlysubsurface microbial life and conditions for preservation of theirbiosignatures.
Materials and MethodsOur samples come from drill core KD2b of the BSDP that intercepted pillowlavas of the Hooggenoeg Formation, along the Komatii river in the BGB,South Africa. A geological map and lithological log of the drill core can befound in SI Materials and Methods. In situ U–Pb titanite dating by LA wasconducted using both a single-collector and a multicollector ICP-MS. Theseincluded a Thermo Finnigan Element 2 single-collector sector field ICP-MScombined with a 193-ArF excimer laser (Resonetics RESOlution; M50-LR) anda Thermo Finnigan Neptune multicollector ICP-MS coupled to a ResoneticsRESOlution M50 laser at the Department of Earth Science, University ofBergen. Full description on the analytical methods and Tables S1–S3 con-taining measured U–Pb isotope ratios by single- and multicollector ICP-MSare provided in SI Materials andMethods. The optical images shown here were
Fig. 4. Quantitative microscale maps of candidate titanite biosignatures and surrounding metamorphic conditions in Archean metabasaltic pillow lavas fromthe BGB. (A) Map of TiO2 (wt%) showing the image of titanite filaments of interest (arrows) highlighted in the dashed-line box. (B) Calculated metamorphicconditions in the chlorite matrix using a chlorite thermodynamic modeling approach (SI Materials and Methods). The dashed-line box highlights a typical areawith well-developed titanite microfilaments surrounded by chlorite pixels recording relatively low temperature conditions of less than T = 350 °C (pixel group 2).(C and D) Image pixel grouping above and below the mean value of T = 350 °C, indicating a higher proportion of pixels recording conditions below T = 350 °Csurrounding the titanite microfilaments (dashed-line box in C). In D the y axis variable Al(iv) = atoms per formula unit of Al4+ in tetrahedral coordination modeledin the chlorite crystal structure.
8384 | www.pnas.org/cgi/doi/10.1073/pnas.1402565111 Grosch and McLoughlin

obtained using a Nikon LV100Pol polarizing microscope and photographedusing a DS-Fi1 color camera with 5.24-megapixel resolution combined withNIS-Elements BR 2.30 software. This software includes a microscalemeasuring tool that was used in the titanite morphological analysis. ACameca SX100 electron microprobe with five wavelength-dispersivespectrometers at the Department of Geosciences, University of Oslo (Oslo,Norway), was used to acquire point analyses and X-ray compositional maps.The X-ray maps were treated using the new software XMapTools to derive
quantitative compositional and metamorphic chlorite temperature maps (SIMaterials and Methods).
ACKNOWLEDGMENTS. We dedicate this paper to the memory of Prof. JanKošler, our friend and colleague. Prof. Košler provided technical assistance withthe U–Pb dating of titanite. This project was financially supported by the Nor-wegian Research Council through the Centre for Geobiology (E.G.G. and N.M.),and also by Bergens Forskningsstiftelse and the University of Bergen.
1. Furnes H, Banerjee NR, Muehlenbachs K, Staudigel H, de Wit MJ (2004) Early liferecorded in Archean pillow lavas. Science 304(5670):578–581.
2. Furnes H, et al. (2007) Comparing petrographic signatures of bioalteration in recentto Mesoarchean pillow lavas: Tracing subsurface life in oceanic igneous rocks. PrecamRes 158(3-4):156–176.
3. Banerjee NR, Furnes H, Muehlenbachs K, Staudigel H, de Wit MJ (2006) Preservationof ca. 3.4 - 3.5 Ga microbial biomarkers in pillow lavas and hyaloclastites from theBarberton Greenstone Belt, South Africa. Earth Planet Sci Lett 241(3-4):707–722.
4. Fliegel D, et al. (2010) In situ dating of earth’s oldest trace fossil at 3.34 Ga. EarthPlanet Sci Lett 299(3-4):290–298.
5. Banerjee NR, et al. (2007) Direct dating of Archean microbial ichnofossils. Geology35(6):487–490.
6. Bridge NJ, Banerjee NR, Mueller W, Muehlenbachs K, Chacko TA (2010) Volcanichabitat for early life preserved in the Abitibi greenstone belt, Canada. Precam Res179(1-4):88–98.
7. Staudigel H, et al. (2008) 3.5 billion years of glass bioalteration: Volcanic rocks asa basis for microbial life? Earth Sci Rev 89(3-4):156–176.
8. Izawa MRM, Banerjee NR, Flemming RL, Bridge NJ, Scultz C (2010b) Basaltic glass asa habitat for microbial life: Implications for astrobiology and planetary exploration.Planet Space Sci 58(4):583–591.
9. Furnes H, et al. (2008) Oceanic pillow lavas and hyaloclastites as habitats for microbiallife through time—a review. Links Between Geological Processes, Microbial Activitiesand Evolution of Life, eds Dilek Y, Furnes H, Muehlenbachs K (Springer, Heidelberg),pp 1–68.
10. Santelli CM, et al. (2008) Abundance and diversity of microbial life in ocean crust.Nature 453(7195):653–656.
11. Lever MA, et al. (2013) Evidence for microbial carbon and sulfur cycling in deeplyburied ridge flank basalt. Science 339(6125):1305–1308.
12. Grosch EG, McLoughlin N, de Wit MJ, Furnes H (2009a) Drilling for the archean rootsof life and tectonic earth in the Barberton Mountains. Sci Drill 8:24–28.
13. Grosch EG, McLoughlin N, de Wit MJ, Furnes H (2009b) Deciphering earth’s deephistory: Drilling in Africa’s oldest greenstone belt. Eos 90(40):350–351.
14. McLoughlin N, Brasier MD, Wacey D, Green OR, Perry RS (2007) On biogenicity criteriafor endolithic microborings on early Earth and beyond. Astrobiology 7(1):10–26.
15. McLoughlin N, Staudigel H, Furnes H, Eickmann B, Ivarsson M (2010) Mechanisms ofmicrotunneling in rock substrates: Distinguishing endolithic biosignatures from abi-otic microtunnels. Geobiology 8(4):245–255.
16. Fisk MR, McLoughlin N (2013) Atlas of alteration textures in volcanic glass from theocean basins. Geosphere 9(2):317–341.
17. Andersen T (2002) Correction of common lead in U-Pb analyses that do not report204Pb. Chem Geol 192(1-2):59–79.
18. Saager R, Köppel V (1976) Lead Isotopes and trace elements from sulphides ofArchean greenstone belts in South Africa—A contribution to the knowledge of theoldest known mineralizations. Econ Geol 71(1):44–57.
19. Lanari P, et al. (2014) XMapTools: A MATLAB©-based program for electron micro-probe X-ray image processing and geothermobarometry. Comput Geosci 62:227–240.
20. Grosch EG, McLoughlin N, Lanari P, Erambert M, Vidal O (2014) Microscale mappingof alteration conditions and potential biosignatures in basaltic-ultramafic rocks onearly Earth and beyond. Astrobiology 14(3):216–228.
21. Grosch EG, et al. (2011) Paleoarchean detrital zircon ages from the earliest tectonicbasin in the Barberton greenstone belt, Kaapvaal Craton, South Africa. Precam Res191(1-2):85–99.
22. López-Martínez M, York D, Hanes JA (1992) A 40Ar/39Ar geochronological study ofkomatiites and komatiitic basalts from the Lower Onverwacht Volcanics: BarbertonMountain Land, South Africa. Precambrian Res 57(1-2):91–119.
23. Armstrong RA, Compston W, de Wit MJ, Williams IS (1990) The stratigraphy of the3.5–3.2 Ga Barberton Greenstone Belt revisited: A single zircon ion microprobe study.Earth Planet Sci Lett 101(1):90–106.
24. Kamo S, Davis DW (1994) Reassessment of Archaean crustal development in theBarberton Mountain Land, South Africa, based on U-Pb dating. Tectonics 13(1):167–192.
25. de Wit MJ, Furnes H, Robins B (2011) Geology and tectonostratigraphy of theOnverwacht Suite, Barberton greenstone belt, South Africa. Precam Res 186(1-4):1–27.
26. Olsson JR, Söderlund U, Klausen MB, Ernst RE (2010) U-Pb baddeleyite ages linkingmajor Archaean dyke swarms to volcanic-rift forming events in the Kaapvaal Craton(South Africa), and a precise age for the Bushveld Complex. Precam Res 183(3):490–500.
27. Fliegel D, et al. (2011) Tubular textures in pillow lavas from a Caledonian westNorwegian ophiolite: A combined TEM, LA-ICP-MS, and STXM study. Geochem Geo-phys Geosyst 12:Q02010.
28. Fisk MR, Crovisier JL, Honnorez J (2013) Experimental abiotic alteration of igneousand manufactured glasses. C R Geosci 345(4):176–184.
29. Brasier MD, McLoughlin N, Green OR, Wacey D (2006) A fresh look at the fossil evi-dence for early Archaean cellular life: The taphonomic potential of sandstones. PhilTrans Roy Soc B 361(1470):887–902.
30. McLoughlin N, Grosch EG, Kilburn MR, Wacey D (2012) Sulfur isotope evidence for aPaleoarchean subseafloor biosphere, Barberton, South Africa. Geology 40(11):1031–1034.
31. Zhang Z, Goloubic S (1987) Endolithic microfossils (cyanophyta) from early ProterozoicStromatolites, Hebei China. Acta Micropaleontol Sin 4:1–12.
32. Tice MM, Lowe DR (2004) Photosynthetic microbial mats in the 3,416-Myr-old ocean.Nature 431(7008):549–552.
33. Javaux EJ, Marshall CP, Bekker A (2010) Organic-walled microfossils in 3.2-billion-year-old shallow-marine siliciclastic deposits. Nature 463(7283):934–938.
34. Noffke N, Eriksson KA, Hazen RM, Simpson EL (2006) A new window into EarlyArchean Life: Microbial mats in Earth’s oldest siliclastic tidal deposits (3.2 Ga MoodiesGroup, South Africa). Geology 34(4):253–256.
35. Grosch EG, McLoughlin N (2013) Paleoarchean sulfur cycle and biogeochemical sur-face conditions on the early Earth, Barberton, South Africa. Earth Planet Sci Lett377–378:142–154.
Grosch and McLoughlin PNAS | June 10, 2014 | vol. 111 | no. 23 | 8385
EART
H,A
TMOSP
HER
IC,
ANDPL
ANET
ARY
SCIENCE
S