supplementary information supplementary … · supplementary information 2 | research supplementary...
TRANSCRIPT

W W W. N A T U R E . C O M / N A T U R E | 1
SUPPLEMENTARY INFORMATIONdoi:10.1038/nature12575
Supplementary Information
2
Fig. S1
Figure S1. Detailed Structure of Non-Oxidative Glycolysis (NOG). a) The phosphoketolase can either react directly with F6P or X5P. In the latter case, F6P input is first converted to X5P via transketolase. b) An example of carbon rearrangement networks involving SBP. The carbon color scheme illustrates carbon rearrangement. Hydroxyl groups are not shown. The red arrows indicate irreversible reactions. Metabolite abbreviations: D-sedoheptulose 7-phosphate (S7P), D-sedoheptulose 1,7-bisphosphate (SBP), D-fructose 6-phosphate (F6P), fructose 1,6-bisphosphate (FBP), D-xylulose 5-phosphate (X5P), D-ribose 5-phosphate (R5P), D-ribulose 5-phosphate (Ru5P), D-erythrose 4-phosphate (E4P), D-glyceraldehyde 3-phosphate (G3P), dihydroxyacetone phosphate (DHAP), acetyl-phosphate (AcP). Enzyme abbreviations are 1a:F6P phosphoketolase (Fpk), 1b: X5P phosphoketolase (Xpk), 2: transaldolase (Tal),3:Transketolase (Tkt),4:ribose-5-phosphate isomerase (Rpi),5:ribulose-5-phosphate epimerase (Rpe),6:Triose phosphate isomerase (Tpi),7:Fructose 1,6 bisphosphate aldolase (Fba),8: Fructose 1,6 bisphosphatase (Fbp), 9: Sedoheptulose-1,7-bisphosphate aldolase (Sba), 10:Sedoheptulose-bisphosphatase (Sbp).
AcP E4P
F6P
E4P X5P
F6P G3P
AcP G3P
Phosphoketolase system
1a1b
3
a.
or
DHAP
X5P E4P
F6P G3P
SBP
E4P X5P
G3P F6PS7P
R5P X5PRu5PX5P
3
3
3
10
9
5
E4P
4
6
b. Carbon rearrangement involving SBP
SUPPLEMENTARY INFORMATION
2 | W W W. N A T U R E . C O M / N A T U R E
RESEARCHSupplementary Information
3
Fig. S2
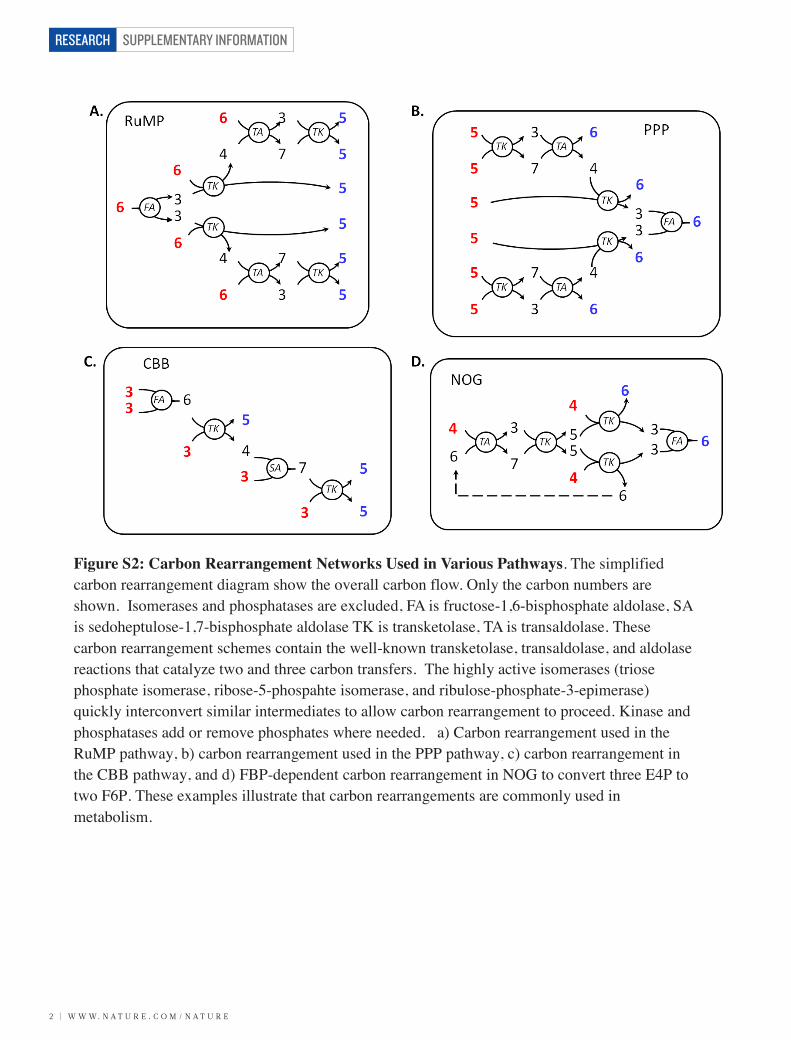
Figure S2: Carbon Rearrangement Networks Used in Various Pathways. The simplified carbon rearrangement diagram show the overall carbon flow. Only the carbon numbers are shown. Isomerases and phosphatases are excluded, FA is fructose-1,6-bisphosphate aldolase, SA is sedoheptulose-1,7-bisphosphate aldolase TK is transketolase, TA is transaldolase. These carbon rearrangement schemes contain the well-known transketolase, transaldolase, and aldolase reactions that catalyze two and three carbon transfers. The highly active isomerases (triose phosphate isomerase, ribose-5-phospahte isomerase, and ribulose-phosphate-3-epimerase) quickly interconvert similar intermediates to allow carbon rearrangement to proceed. Kinase and phosphatases add or remove phosphates where needed. a) Carbon rearrangement used in the RuMP pathway, b) carbon rearrangement used in the PPP pathway, c) carbon rearrangement in the CBB pathway, and d) FBP-dependent carbon rearrangement in NOG to convert three E4P to two F6P. These examples illustrate that carbon rearrangements are commonly used in metabolism.

W W W. N A T U R E . C O M / N A T U R E | 3
SUPPLEMENTARY INFORMATION RESEARCHSupplementary Information
4
Fig. S3
Figure S3. NOG with Pentose and Triose Sugar Phosphates. NOG can take any sugar as input molecules, as long as it can be converted to sugar phosphates that are present in the carbon rearrangement network. The pathways using R5P (A) and G3P (B) are shown. These pathways use F/Xpk dual activity, though similar pathways can be drawn using Fpk only or Xpk only. Carbon rearrangement can convert any sugar phosphate (triose to sedoheptulose) to stoichiometric amounts of F6P. Here the conversion of a ribose and triose are illustrated. Abbreviations and enzyme numbers are defined in Figure 1 legend.

SUPPLEMENTARY INFORMATION
4 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
Supplementary Information
5
Fig. S4
Figure S4: SDS-PAGE of HIS-tagged NOG proteins. SDS-PAGE denaturing gels of purified Rpe, Rpi, Tkt, Tal, F/Xpk, Ack, and Fbp. Sizes corresponded within theoretical values. These proteins were design with N-terminal polyhistidine tags and crude extracts were purified using a one-step affinity chromatography procedure.
Fig. S5
Figure S5: Expression of pIB4 in JCL118. Expression of F/Xpk and Fbp from JCL118 with pIB4. Lanes 1: Low Range Ladder, 2: Un-induced Crude Extract, 3: Induced Crude Extract, 4 and 5 are duplicates of 2 and 3. Lanes 6 through 9 are HIS-tag purified elutions of Lanes 2 through 5.

W W W. N A T U R E . C O M / N A T U R E | 5
SUPPLEMENTARY INFORMATION RESEARCH
Supplementary Information
6
Fig. S6

SUPPLEMENTARY INFORMATION
6 | W W W. N A T U R E . C O M / N A T U R E
RESEARCHSupplementary Information
7
Figure S6: In Vitro Enzyme Assay Schemes. A series of enzyme assays were performed to confirm the activity of purified NOG enzymes. For some enzymes (Ack, F/Xpk, and Fbp), both a colorimetric and a UV based assays were performed. Specific activities of all enzymes are listed in the Supplementary Table S1. In all enzyme-coupled assays, the key enzyme of interest was limiting by while the rest were added in excess. Glk (baker’s yeast), Zwf (baker’s yeast), Pfk (from Bacillus stearothermophilus), Tpi (rabbit muscle), Gpd (rabbit muscle), and Pgi (baker’s yeast) were purchased from Sigma-Alrich (St. Louis, MO).

W W W. N A T U R E . C O M / N A T U R E | 7
SUPPLEMENTARY INFORMATION RESEARCHSupplementary Information
8
Table S1: Calculation of Specific Activities of HIS-tag Purified Enzymes. One unit of enzyme activity (U) is defined as conversion of 1 micro mole of substrate per minute.
Assay Other Enzymes Added
Substrate Protein (mg) Units (µmole/min)
Specific Activity (µmole/min/mg protein)
Fbp – UV Pgi, Zwf 2 mM F6P 5.0*10-3 0.020 4.1 Xpk – UV Rpi, Rpe, Ack, Glk,
Zwf 10 mM R5P 2.5*10-3 0.00885 3.5
Xpk – Color Rpi, Rpe 10 mM R5P 2.85*10-2 0.1437 2.5 Fpk – UV Ack, Glk, Zwf 5 mM F6P 7.5*10-3 0.0053 0.71 Tkt – UV Rpe, Rpi, Tpi, Gpd 5 mM R5P 9.2*10-4 0.013 14.0 Tal – UV Rpe, Rpi, Tkt, Pgi,
Zwf 2 mM R5P 1.2*10-3 0.022 18
Ack – UV Glk, Zwf 5 mM AcP 8.5*10-5 0.065 764 Ack – Color Glk 5 mM AcP 1.3*10-4 0.27 1030

SUPPLEMENTARY INFORMATION
8 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
Supplementary Information
9
Figure S7

W W W. N A T U R E . C O M / N A T U R E | 9
SUPPLEMENTARY INFORMATION RESEARCHSupplementary Information
10
Figure S7 Various glucose-degrading (glycolytic) pathways. A) The Embden–Meyerhof–Parnas (EMP). In the most common glycolytic pathway, glucose is oxidized to two pyruvates which can then form two lactates. B) The Entner–Doudoroff (ED) pathway overlaps somewhat with the lower EMP pathway but differs in the initial utilization of glucose. C) The Phosphoketolase (PK) Pathway. The PK pathway produces a mixture of ethanol, lactate, and CO2. It utilizes a X5P-specific phosphoketolase unlike the Bifid Shunt and only produces 1 net ATP. D) The Bifid Shunt is specific to Bifidobacteria which contain a dual F6P/X5P phosphoketolase. This pathway produces a mixture of 1.5 acetate and lactate with a relatively high net of 2.5 ATP/glucose. Both the PK and Bifid Shunt are oxidative and non-cyclical. E) The Wood-Ljungdahl pathway (W-L) is used by some acetogens to produce a third AcCoA from wasted CO2 released by the oxidative decarboxylation of pyruvate. This complex pathway is highly-oxygen sensitive but can produce the highest net 4 ATP/glucose F) NOG contains well-established enzymes found in three distinct pathways: the pentose phosphate pathway (PPP), gluconeogenesis, and the phosphoketolase pathway1,2The NOG employs fewer enzymes which are oxygen-tolerant and can produce three acetates from glucose with a net of 2 ATP/glucose.

SUPPLEMENTARY INFORMATION
1 0 | W W W. N A T U R E . C O M / N A T U R E
RESEARCHSupplementary Information
11
Table S2. Overall reactions and Properties of Various Glycolytic Pathways
Name Products from Glucose Net ATP Net Redox O2 Sensitive? # Enzymes EMP 2 lactate 2 0 No 11
ED 2 lactate 1 0 No 11 PKP lactate + ethanol + CO2 1 0 No 15 B-S 1.5 acetate + lactate 2.5 0 No 14
EMP-WL 3 acetate 4 0 Yes >20
NOG* 3 acetate 2 0 No 11 *Theoretical conversion.

W W W. N A T U R E . C O M / N A T U R E | 1 1
SUPPLEMENTARY INFORMATION RESEARCHSupplementary Information
12
Table S3: Summary of Properties of NOG enzymes. One unit of enzyme activity (U) is defined as conversion of 1 micro mole of substrate per minute.
Name # Abbrev. EC# Source
Specific Activity (U/mg) Km Ref
F6P-Phosphoketolase 1a HIS-Fpk 4.1.2.22 B. adolescentis 0.5 N/A This work X5P-Phosphoketoalse 1b HIS-Xpk 4.1.2.9 B. adolescentis 3.5 N/A This work F6P-Phosphoketolase 1a Fpk 4.1.2.22 B. breve 14 9.6 3 X5P-Phosphoketoalse 1b Xpk 4.1.2.9 B. breve 29 N/A 3
Transaldolase B 2 Tal
2.2.1.2 E. coli 60 E4P-0.09 4 2.2.1.2 E. coli 60 F6P-1.2 4
HIS-Tal 2.2.1.2 E. coli 18 N/A This work
Transketolase A 3 Tkt
2.2.1.1 E. coli 50-110 G3P-2.1 5 2.2.1.1 E. coli
E4P-0.09 5
2.2.1.1 E. coli
R5P-1.4 5 2.2.1.1 E. coli
X5P-0.16 5
2.2.1.1 E. coli
F6P-1.1 5 2.2.1.1 E. coli
S7P-4.0 5
HIS-tkt 2.2.1.1 E. coli 14 N/A This work
Ribose-5-phosphate isomerase 4
Rpi 5.3.1.6 E. coli N/A R5P-3.1 6 5.3.1.6 E. coli 135 N/A 7 5.3.1.6 Yeast 2429 R5P-1.6 8
HIS-Rpi 5.3.1.6 E. coli >1000 N/A This work
Ribulose-3-phosphate epimerase 5
Rpe 5.1.3.1 E. coli N/A Ru5P-1.6 9 5.1.3.1 Rice 16000 N/A 10 5.1.3.1 Spinach 17000 Ru5P-0.22 11
HIS-Rpe 5.1.3.1 E. coli >1000 N/A This work
Triose Phosphate Isomerase 6 Tpi
5.3.1.1 O. cuniculus - liver 6400 G3P-0.42 12 5.3.1.1 O. cuniculus - liver
DHAP-0.75 12
5.3.1.1 O. cuniculus - muscle 7800 N/A 13
Fructose 1,6 bisphosphate Aldolase 7 Fba
4.1.2.13 O. cuniculus - muscle 14 N/A 14 4.1.2.13 O. cuniculus - muscle 11 N/A 15 4.1.2.13 O. cuniculus - liver 12 N/A 16 4.1.2.13 O. cuniculus - muscle 16 N/A 17 4.1.2.13 O. cuniculus - muscle N/A FBP-0.0013 18
Fructose 1,6 Bisphosphatase 8
Fbp 3.1.3.11 E. coli 24.2 FBP-0.015 19 3.1.3.11 E. coli 35 FBP-0.0017
HIS-Fbp 3.1.3.11 E. coli 4.1 N/A This work

SUPPLEMENTARY INFORMATION
1 2 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH Supplementary Information
13
Table S4. Properties of Wood-Ljungdahl and EMP Enzymes. One unit of enzyme activity (U) is defined as conversion of 1 micro mole of substrate per minute.
Name Abbrev. Specific Activity (U/mg)
O2 Sensitive Ref
Formate dehydrogenase (NADPH) Fdh 1050* Very 20 Formyl-THF-Synthetase Fts 780 N/A 21
230 N/A 22 Methenyl-THF Cyclohydrolase Mtc 310 N/A 23
Methylene-THF Dehydrogenase (NADPH) Mtd 200 N/A 23 Methylene-THF Reductase Mtr 139 Very 24
Methyltransferase Mtf 150 N/A 25 CO Dehydrogenase/Acetyl-CoA Synthase Complex Cod 0.75* Very 26
0.82* Very 27 Glyceraldehyde-3-Phosphate Dehydrogenase Gap 205 N/A 28
Phosphoglycerate kinase Pyk 800 N/A 28 Phosphoglycerate mutase Gpm 2000 N/A 28
Enolase Eno 160 N/A 28 Pyruvate Kinase Pyk 280 N/A 28
Pyruvate Dehydrogenase Complex Pdh 1791 N/A 29 Phosphotransacetylase Pta >1000 N/A
*Assayed in the reverse direction
Figure S8. Overlapping enzymes in the two aceotogenic pathways: NOG and EMP+ WL .

W W W. N A T U R E . C O M / N A T U R E | 1 3
SUPPLEMENTARY INFORMATION RESEARCHSupplementary Information
14
Table S5. Kinetic parameters of various F6P and X5P Phosphoketolases
Organism Km Specific Activity (U/mg) Ref
Bifidobacterium adolescentis N/A X5P-3.5 This work Bifidobacterium adolescentis N/A F6P-0.7 This work
Bifidobacterium longum BB 536 F6P-26 N/A 30 Bifidobacterium dentium F6P-23 N/A 30
Bifidobacterium animalis. lactis F6P-11.5 N/A 30 Bifidobacterium globosum F6P-12.5 N/A 30
Bifidobacterium breve st. 203 X5P-N/A X5P-29.0 3 F6P-9.7 F6P-14.0 3
Bifidobacterium globosum RU 230 F6P-1.4 F6P-24 31 Bifidobacterium dentium B 764 F6P-39 F6P-30 31
Lactobacillus paraplantarum F6P-5.1 F6P-147.3 32 Lactobacillus pentosus N/A X5P-4.5 33
Lactobacillus plantarum X5P-3.6 X5P-4.1 34 F6P-24 F6P-1.8 34
Bifidobacterium animalis. lactis X5P-45 X5P-27 35 F6P-10 F6P-5.2 35
Acetobacter xylinum F6P-2.5 F6P-0.267 36

SUPPLEMENTARY INFORMATION
1 4 | W W W. N A T U R E . C O M / N A T U R E
RESEARCHSupplementary Information
15
Table S6: Comparison of acetyl-CoA synthesis using CBB+NOG and CBB+EMP pathways. The CBB+NOG pathway requires 2 CO2 turnover by Rubisco and 6 ATP, while the CBB+EMP pathway requires 3 CO2 turnover and 7 ATP.

W W W. N A T U R E . C O M / N A T U R E | 1 5
SUPPLEMENTARY INFORMATION RESEARCH
Supplementary Information
16
Table S7. Primers used to Clone NOG enzymes and Create pIB4.
N-Terminal HIS-Tagged Template Primers
pQE9-HIS-Fxpk B. adolescentis
206
207
TTTTATTTGATGCCTCTAGATCACTCGTTATCGCCAGCGG
AAAACCTGTATTTTCAGGGAATGACGAGTCCTGTTATTGGCACC
pQE9-HIS-Tkt E. coli (JCL16)
235
236
AAAACCTGTATTTTCAGGGAATGTCCTCACGTAAAGAGCTTGC
TTTTATTTGATGCCTCTAGATTACAGCAGTTCTTTTGCTTTCGC
pQE9-HIS-Tal E. coli (JCL16)
233
234
AAAACCTGTATTTTCAGGGAATGACGGACAAATTGACCTCCCTT
TTTTATTTGATGCCTCTAGATTACAGCAGATCGCCGATCATTTTTTC
pQE9-HIS-Rpe E. coli (JCL16)
202
204
AAAACCTGTATTTTCAGGGAATGAAACAGTATTTGATTGCCCCCTC
TTTTATTTGATGCCTCTAGATTATTCATGACTTACCTTTGCCAGTTC
pQE9-HIS-Rpi E. coli (JCL16)
199
201
AAAACCTGTATTTTCAGGGAATGACGCAGGATGAATTGAAAAAAGC
TTTTATTTGATGCCTCTAGATCATTTCACAATGGTTTTGACACCG
pQE9-HIS-Ack E. coli (JCL16)
229
230
AAAACCTGTATTTTCAGGGAATGTCGAGTAAGTTAGTACTGGTTC
TTTTATTTGATGCCTCTAGATCAGGCAGTCAGGCGGC
pQE9-HIS-Fbp E. coli (JCL16)
211
213
AAAACCTGTATTTTCAGGGAATGAAAACGTTAGGTGAATTTATTGTCG
TTTTATTTGATGCCTCTAGATTACGCGTCCGGGAACTCAC
Vector Backbone pQE9 (Qiagen)
94
005F
TCTAGAGGCATCAAATAAAACGAAAGGC
TCCCTGAAAATACAGGTTTT
pIB4:
Vector Backbone pZE12-luc
316
005F
CTGGCGATAACGAGTGATCTAGAGGCATCAAATAAAACGAAAGGC
TCCCTGAAAATACAGGTTTT
Fbp pQE9-HIS-Fbp
211
314
AAAACCTGTATTTTCAGGGAATGAAAACGTTAGGTGAATTTATTGTCG
TCTCATAGTTAATTTCTCCTCTTTAATTTACGCGTCCGGGAACTCAC
Fpk pQE9-HIS-Fxpk
315
206
CGTAAATTAAAGAGGAGAAATTAACTATGAGAGGATCGCATCACCATCAC
TTTTATTTGATGCCTCTAGATCACTCGTTATCGCCAGCGG

SUPPLEMENTARY INFORMATION
1 6 | W W W. N A T U R E . C O M / N A T U R E
RESEARCH
Supplementary Information
17
Table S8. Strains and Plasmids Used
Strains
JCL16 BW25113/F’ [traD36, proAB+, lacIq, ZΔM15 (TetR)]
37
JCL166 JCL16 , but ΔadhE, ΔldhA, ΔfrdBC 37
JCL166/pIB4 JCL166 with pIB4 This work
JCL118 JCL16 , but ΔadhE, ΔldhA, ΔfrdBC, ΔpflB This work
JCL118/pIB4 JCL118 with pIB4 This work
Plasmids
pZE12-luc ColE1 ori; AmpR;PLlacO1::luc(PP) 38
pIB4 ColE1 ori; AmpR;PLlacO1::fbp(EC)-fxpk(BA) This work
pQE9 ColE1 ori; AmpR; PT5lacO:: Qiagen (Chatsworth, CA)
pQE9-HIS-F/Xpk From pQE9, PT5lacO::fxpk(BA) This work
pQE9-HIS-Ack From pQE9, PT5lacO::ackA(EC) This work
pQE9-HIS-Tkt From pQE9, PT5lacO::tktA(EC) This work
pQE9-HIS-Tal From pQE9, PT5lacO::talB(EC) This work
pQE9-HIS-Fbp From pQE9, PT5lacO::fbp(EC) This work
pQE9-HIS-Rpe From pQE9, PT5lacO::rpe(EC) This work
pQE9-HIS-Rpi From pQE9, PT5lacO::rpiA(EC) This work EC denotes Escherichia coli. PP denotes Photinus pyralis. BA denotes Bifidobacterium adolescentis.

W W W. N A T U R E . C O M / N A T U R E | 1 7
SUPPLEMENTARY INFORMATION RESEARCHSupplementary Information
18
References:
1. Heath, E., Hurwitz, J. & Ginsburg, A. Pentose fermentation by Lactobacillus plantarum I. The cleavage of xylulose 5-phosphate by phosphoketolase. Journal of Biological Chemistry (1958).
2. Suzuki, R. et al. Crystal structures of phosphoketolase: thiamine diphosphate-dependent dehydration mechanism. The Journal of biological chemistry 285, 34279–87 (2010).
3. Suzuki, R. et al. Overexpression, crystallization and preliminary X-ray analysis of xylulose-5-phosphate/fructose-6-phosphate phosphoketolase from Bifidobacterium breve. Acta crystallographica. Section F, Structural biology and crystallization communications 66, 941–3 (2010).
4. Sprenger, G. a, Schörken, U., Sprenger, G. & Sahm, H. Transaldolase B of Escherichia coli K-12: cloning of its gene, talB, and characterization of the enzyme from recombinant strains. Journal of bacteriology 177, 5930–6 (1995).
5. Sprenger, G. a, Schörken, U., Sprenger, G. & Sahm, H. Transketolase A of Escherichia coli K12. Purification and properties of the enzyme from recombinant strains. European journal of biochemistry / FEBS 230, 525–32 (1995).
6. Zhang, R. et al. Structure of Escherichia coli Ribose-5-Phosphate Isomerase:: A Ubiquitous Enzyme of the Pentose Phosphate Pathway and the Calvin Cycle. Structure 11, 31–42 (2003).
7. Hove-Jensen, B. & Maigaard, M. Escherichia coli rpiA gene encoding ribose phosphate isomerase A. Journal of bacteriology 175, 5628–35 (1993).
8. Reuter, R., Naumann, M., Bär, J., Miosga, T. & Kopperschläger, G. Ribose-5-phosphate isomerase from Saccharomyces cerevisiae: purification and molecular analysis of the enzyme. Bioseparation 7, 107–15 (1998).
9. Sobota, J. M. & Imlay, J. a. Iron enzyme ribulose-5-phosphate 3-epimerase in Escherichia coli is rapidly damaged by hydrogen peroxide but can be protected by manganese. Proceedings of the National Academy of Sciences of the United States of America 108, 5402–7 (2011).
10. Kopriva, S., Koprivova, a & Süss, K. H. Identification, cloning, and properties of cytosolic D-ribulose-5-phosphate 3-epimerase from higher plants. The Journal of biological chemistry 275, 1294–9 (2000).
11. Chen, Y., Hartman, F., Lu, T. & Larimer, F. D-Ribulose-5-phosphate 3-epimerase: cloning and heterologous expression of the spinach gene, and purification and characterization of the recombinant enzyme. Plant physiology 199–207 (1998).
12. Krietsch, W. Triosephosphate isomerase from rabbit liver. Methods in enzymology 438–442 (1975).

SUPPLEMENTARY INFORMATION
1 8 | W W W. N A T U R E . C O M / N A T U R E
RESEARCHSupplementary Information
19
13. Norton, I. L., Pfuderer, P., Stringer, C. D. & Hartman, F. C. Isolation and characterization of rabbit muscle triose phosphate isomerase. Biochemistry 9, 4952–8 (1970).
14. Ginsburg, a & Mehler, a H. Specific anion binding to fructose diphosphate aldolase from rabbit muscle. Biochemistry 5, 2623–34 (1966).
15. Shapiro, S., Enser, M., Pugh, E. & Horecker, B. L. The effect of pyridoxal phosphate on rabbit muscle aldolase. Archives of biochemistry and biophysics 128, 554–62 (1968).
16. Chappel, A., Hoogenraad, N. & Holmes, R. Purification and properties of the native form of rabbit liver aldolase. Evidence for proteolytic modification after tissue extraction. Biochemical Journal 175, 377–382 (1978).
17. Grazi, E. Fructose 1, 6-diphosphate aldolase from rabbit muscle. Effect of pH on the rate of formation and on the equilibrium concentration of the carbanion intermediate. Biochemical Journal 91, 167–172 (1975).
18. MacDonald, J. & Storey, K. Purification and characterization of fructose bisphosphate aldolase from the ground squirrel, Spermophilus lateralis: enzyme role in mammalian hibernation. Archives of biochemistry and biophysics 408, 279–285 (2002).
19. Kelley-loughnane, N. et al. Purification, kinetic studies, and homology model of Escherichia coli fructose-1,6-bisphosphatase. Biochimica et biophysica acta 1594, 6–16 (2002).
20. Yamamoto, I., Saiki, T., Liu, S. M. & Ljungdahl, L. G. Purification and properties of NADP-dependent formate dehydrogenase from Clostridium thermoaceticum, a tungsten-selenium-iron protein. The Journal of biological chemistry 258, 1826–32 (1983).
21. Radfar, R. et al. Cation binding and thermostability of FTHFS monovalent cation binding sites and thermostability of N10-formyltetrahydrofolate synthetase from Moorella thermoacetica. Biochemistry 39, 14481–6 (2000).
22. Ljungdahl, L., Brewer, J. M., Neece, S. H. & Fairwell, T. Purification, stability, and composition of formyltetrahydrofolate synthetase from Clostridium thermoaceticum. The Journal of biological chemistry 245, 4791–7 (1970).
23. Ljungdahl, L., O’Brien, W., Moore, M. & Liu, M. Methylenetetrahydrofolate dehydrogenase from Clostridium formicoaceticum and methylenetetrahydrofolate dehydrogenase, methenyltetrahydrofolate. Methods in enzymology 66, 599–609 (1980).
24. Clark, J. E. & Ljungdahl, L. G. Purification and properties of 5,10-methylenetetrahydrofolate reductase, an iron-sulfur flavoprotein from Clostridium formicoaceticum. The Journal of biological chemistry 259, 10845–9 (1984).
25. Roberts, D. L. E. E., Zhao, S., Doukov, T. & Ragsdale, S. W. The Reductive Acetyl Coenzyme A Pathway : Sequence and Heterologous Expression of Active Methyltetrahydrofolate : Corrinoid / Iron-Sulfur Protein Methyltransferase from Clostridium thermoaceticum. (1994).

W W W. N A T U R E . C O M / N A T U R E | 1 9
SUPPLEMENTARY INFORMATION RESEARCHSupplementary Information
20
26. Ragsdale, S. & Clark, J. Properties of purified carbon monoxide dehydrogenase from Clostridium thermoaceticum, a nickel, iron-sulfur protein. Journal of Biological Chemistry 258, 2364–2369 (1983).
27. Roberts, J., Lu, W. & Ragsdale, S. Acetyl-coenzyme A synthesis from methyltetrahydrofolate, CO, and coenzyme A by enzymes purified from Clostridium thermoaceticum: attainment of in vivo rates and identification of rate-limiting steps. Journal of bacteriology (1992).
28. Pawluk, A., Scopes, R. K. & Griffiths-smith, K. Isolation and properties of the glycolytic enzymes from Zymomonas mobilis. 281, 275–281 (1986).
29. Saumweber, H., Binder, R. & Bisswanger, H. Pyruvate dehydrogenase component of the pyruvate dehydrogenase complex from Escherichia coli K12. Purification and characterization. European journal of biochemistry / FEBS 114, 407–11 (1981).
30. Grill, J. P., Crociani, J. & Ballongue, J. Characterization of fructose 6 phosphate phosphoketolases purified from Bifidobacterium species. Current microbiology 31, 49–54 (1995).
31. Sgorbati, B., Lenaz, G. & Casalicchio, F. Purification and properties of two fructose-6-phosphate phosphoketolases in Bifidobacterium. Antonie Van Leeuwenhoek 42, 49–57 (1976).
32. Jeong, D. W., JUNG, M. I. N. L. E. E. & HYONG, J. O. O. L. E. E. Cloning and characterization of a gene encoding phosphoketolase in a Lactobacillus paraplantarum isolated from Kimchi. Journal of microbiology and biotechnology 17, 822–829 (2007).
33. Posthuma, C. C. et al. Expression of the xylulose 5-phosphate phosphoketolase gene, xpkA, from Lactobacillus pentosus MD363 is induced by sugars that are fermented via the phosphoketolase pathway and is repressed by glucose mediated by CcpA and the mannose phosphoenolpyruvate p. Applied and environmental microbiology 68, 831–837 (2002).
34. Yevenes, A. & Frey, P. a. Cloning, expression, purification, cofactor requirements, and steady state kinetics of phosphoketolase-2 from Lactobacillus plantarum. Bioorganic chemistry 36, 121–7 (2008).
35. Meile, L., Rohr, L. M., Geissmann, T. A., Herensperger, M. & Teuber, M. Characterization of the D-xylulose 5-phosphate/D-fructose 6-phosphate phosphoketolase gene (xfp) from Bifidobacterium lactis. Journal of Bacteriology 183, 2929–2936 (2001).
36. Racker, E. Fructose-6-phosphate phosphoketolase from Acetobacter xylinum. Methods in enzymology 1283, 276–280 (1962).
37. Atsumi, S. et al. Metabolic engineering of Escherichia coli for 1-butanol production. Metabolic engineering 10, 305–11 (2008).
38. Lutz, R. & Bujard, H. Independent and tight regulation of transcriptional units in Escherichia coli via the LacR/O, the TetR/O and AraC/I1-I2 regulatory elements. Nucleic acids research 25, 1203 (1997).