rats in antiquity
DESCRIPTION
Unwelcome companions: ancient rats reviewedTRANSCRIPT

Unwelcome companions: ancient rats reviewed
PHILIP L. ARMITAGE”
The commensal rats - notably the black rat Rattus rattus and the brown R. norvegicus - are among mankind’s most destructive and dangerous enemies, and have spread relent-
lessly with humans across the globe. A decade after an important ratty meeting at the Natural History Museum, London, in 1981, this noxious rodent pest is again reviewed.
This paper is submitted in honour of Juliet Clutton-Brock (Natural History Museum, London).
Introduction In October, 1981, at the instigation of Dr David E. Davis of California, a small group of archaeozoologists and one historian met at the Natural History Museum, London, in order to review what was then known concerning the distribution of Rattus rattus in medieval Brit- ain. This event proved ‘particularly stimulating’ (Twigg 1984: preface), and rekindled interest in the subject, especially the question of the rat’s presence in Britain before the medieval period; a possibility raised by the discovery of their skel- etal remains in a Roman well in York (Rackham 1979). It was the conclusion of the group that the York rats were unlikely to be an isolated phe- nomenon, an observation demonstrated two years later (in 1983) when further Roman rat bones (FIGURE 1) were recovered by Museum of London archaeologists (directed by Fredericke Hammer and Ken Steedman) from a securely dated 3rd-century well fill at Fenchurch Street in the City of London.
Discovery of the Fenchurch Street black rat remains prompted Barbara West and myself (both then employed by the Museum of Lon- don) to collaborate in investigating their wider signifiance. West gathered together published and unpublished archaeological records of R. rattus in Britain, and drew up a provisional his- torical distribution map, while I collated archaeo- logical records from Europe and further afield to see how the black rat spread from its presumed homeland in southern Asia to northern Europe during the Roman era (Armitage et al. 1984).
Further reports of early European rats came from Roman sites in central Europe (Teichert 1985), from 2nd-century BC Menorca (Spain) (Reumer 1986), from Roman York (O’Connor 1988b) and from Picardie (northern France), where Vigne & Femolant (1991) found black rat as early as the 1st century AD.
As O’Connor (1991: 318) noted, in just over a decade, then, archaeozoologists in Britain and on the continent have largely rewritten the early history of R. rattus. But despite this ad- vance our knowledge is still incomplete; and this paper reviews chronology and pattern in the spread of R. rattus, identifying where in- formation is either patchy or lacking. Among the prioirity areas meriting further study are: 1 The pre-Roman spread of R. rattus from
southeast Asia; 2 The apparent extinction in Dark Age north-
ern Europe of R. rattus; 3 The reintroduction into northern Europe of
R. rclttus c. late 9th century/early loth century;
The apparent extraordinary large size of R. rattus in northern Europe during the later medieval period.
4
Evidence for the pre-Roman spread of Rattus rattus from southeast Asia Among responses to Armitage et al. (1984) was the observation that I had neglected evidence for black rats moving from southeast Asia much earlier than the 4th century BC. In my original (tentative) model I had suggested the develop-
* 1972 Roseate Lane, Sanibel Island FL 33957, USA.
ANTIQUITY 68 (1994): 231-40
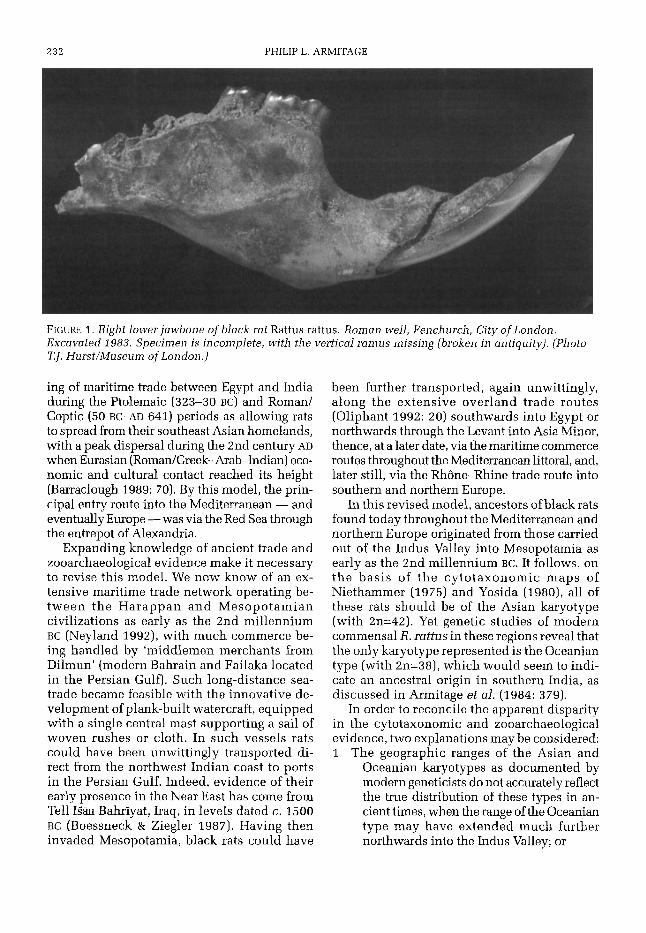
232 PHILIP L. ARMITAGE
FIGURE 1. Right lower jawbone of black rat Rattus rattus. Roman well, Fenchurch, City of London. Excavated 1983. Specimen is incomplete, with the vertical ramus missing (broken in antiquity). (Photo TJ. Hurst/Museum of London.)
ing of maritime trade between Egypt and India during the Ptolemaic (323-30 BC) and Roman/ Coptic (50 BC-AD 641) periods as allowing rats to spread from their southeast Asian homelands, with a peak dispersal during the 2nd century AD when Eurasian (Roman/Greek-ArabIndian) eco- nomic and cultural contact reached its height (Barraclough 1989: 70). By this model, the prin- cipal entry route into the Mediterranean - and eventually Europe -was via the Red Sea through the entrepot of Alexandria.
Expanding knowledge of ancient trade and zooarchaeological evidence make it necessary to revise this model. We now know of an ex- tensive maritime trade network operating be- tween the Harappan and Mesopotamian civilizations as early as the 2nd millennium BC (Neyland 1992), with much commerce be- ing handled by ‘middlemen merchants from Dilmun’ (modern Bahrain and Failaka located in the Persian Gulf). Such long-distance sea- trade became feasible with the innovative de- velopment of plank-built watercraft, equipped with a single central mast supporting a sail of woven rushes or cloth. In such vessels rats could have been unwittingly transported di- rect from the northwest Indian coast to ports in the Persian Gulf. Indeed, evidence of their early presence in the Near East has come from Tell ISa Bahriyat, Iraq, in levels dated c. 1500 BC (Boessneck & Ziegler 1987). Having then invaded Mesopotamia, black rats could have
been further transported, again unwittingly, along the extensive overland trade routes (Oliphant 1992: 20) southwards into Egypt or northwards through the Levant into Asia Minor, thence, at a later date, via the maritime commerce routes throughout the Mediterranean littoral, and, later still, via the RhBne-Rhine trade route into southern and northern Europe.
In this revised model, ancestors of black rats found today throughout the Mediterranean and northern Europe originated from those carried out of the Indus Valley into Mesopotamia as early as the 2nd millennium BC. It follows, on the basis of the cytotaxonomic maps of Niethammer (1975) and Yosida (1980), all of these rats should be of the Asian karyotype (with 2n=42). Yet genetic studies of modern commensal R. rattus in these regions reveal that the only karyotype represented is the Oceanian type (with 2n=38), which would seem to indi- cate an ancestral origin in southern India, as discussed in Armitage et al. (1984: 379).
In order to reconcile the apparent disparity in the cytotaxonomic and zooarchaeological evidence, two explanations may be considered: 1 The geographic ranges of the Asian and
Oceanian karyotypes as documented by modern geneticists do not accurately reflect the true distribution of these types in an- cient times, when the range of the Oceanian type may have extended much further northwards into the Indus Valley; or

UNWELCOME COMPANIONS: ANCIENT RATS REVIEWED 233
2 Asian type rats did spread from northwest India to southwest Asia and Asia Minor during the 2nd millennium BC, but failed to establish themselves permanently.
Some support for the second scenario comes from the Greek ‘Dark Age’ (Early Iron Age) site at Nichoria in southwest Greece, where R. rattus was noticeably absent from an otherwise very rich small mammal faunal assemblage that was excavated from apithos (storage jar) which had served as a pit-fall trap for unwary small creatures living on the acropolis c. 1050-975 BC (see Sloan & Duncan 1978: 75). Perhaps, then, there was no continuity in the dispersal of R. rattus from the 2nd millennium BC through to the Roman period, and it was a much later, second wave of rats from southern India that established a permanent rat popula- tion in Egypt/Mediterranean, which in the Roman period came to invade Europe.
More problematic is the claim by Tchernov (1984) that commensal R. rattus has occupied the coastal region of northern Israel since the Natufian ( c . 9500-7500 BC). Certainly the Natufian adoption of a sedentary way of life centred on proto-villages with storage pits for grain harvested from the wild, created condi- tions favouring infestation by rats. But where did they come from? If commensal R. rattus originated elsewhere (in sub-tropical southeast Asia), it is difficult to explain them in the Le- vant at so early a period. Tchernov overcomes this problem by suggesting that R. rattus is ‘na- tive’ to the southern Levant, where it has lived since the end of the Pleistocene. Before accept- ing this, however, we should re-examine the stratigraphic integrity of the ‘Natufian’ rodent bones, to be sure they are not contaminated from more recent levels.
Extinction in the Dark Ages While R. rattus is known to have been present in Roman Britain, its fine-scale distribution there is poorly understood. In Britain, and throughout northern Europe, was R. rattus mainly confined to the ports and larger urban centres, less common in, or even entirely ab- sent from, rural districts? Reumer (1986) thinks so, as rat was absent from a ‘large sample of small mammals’ he examined from the Roman levels in the town of Tiddington near Stratford, yet he acknowledges the inland spread of this species by the 5th century AD (there were rat
bones in the sub-Roman levels at Wroxeter: Armitage et al. 1984).
Our reconstruction of rat distribution in later Roman times will have to take account of the economic upheavals of that period. As early as the mid 2nd century AD, southern and cen- tral urban centres such as Londinium experi- enced decline, with greatly reduced densities of human occupation (Sheldon 1975; Merri- field 1983: 147; Marsden & West 1992), this was accompanied by a shift in socio-economic emphasis to the rural (villa) estates which func- tioned as self-reliant working farms and/or luxurious country houses (Hills 1986: 130), all of which probably affected rat populations of the urban centres. Although the rat bones in 4th-century deposits at Crosswall in the City of London (Armitage et al. 1984) indicates con- tinuity of rat occupancy, actual rat numbers in London and other port cities may nevertheless have been declining, while the construction of villas may have provided opportunities for rats to settle in rural districts, a process that could have been facilitated by a warmer climate (for the later Roman climate in Britain, see Addyman et al. 1976). Black rats, unwittingly transported by the Romans to their villas, could have moved into nearby native (British) settlement sites. The same diffusion may have taken place in Gaul where a similar shift of economic emphasis from urban (city) to rural (villa) sites occurred.
By the terminology and conceptual models of biological invasions of Hengeveld (1989: 18- 19), this would have been a ‘hierarchical dif- fusion’, featuring, first, long-distance jump dispersal from the primary foci (major urban centres/ports) to isolated, independently propagating bridgeheads (villas); then, a fur- ther spread into the surrounding countryside (native settlements) by ‘neighbourhood diffu- sion’, this shorter-distance secondary spread depending on local topography, settlement pattern and human density.
Only through the systematic collection and detailed interpretation of zooarchaeological evidence from rural Romano-British sites will this model be substantiated or refuted. Of spe- cial interest will be just how far west and north had the spread of R. rattus reached, and whether it survived in these regions into the sub-Roman (5th century AD) period, when the Roman way of life was replaced by an ‘impov- erished version of Iron Age society’ - when

234 PHILIP L. ARMITAGE
the human population retreated ‘from former Roman centres to the hillforts their ancestors had fortified centuries before’ (Hills 1986: 138). For the warmth-loving black rat these elevated hillfort locations were probably ill-suited as habitats - cool, wet and exposed.
By the ‘Dark Ages’ (6th-8th centuries AD) the rats had gone, as first revealed by West’s survey (in Armitage et al. 1984). The supposed 8th-century specimens from Coppergate, York, included by West were later confirmed as of the late 9th century, leaving a clear gap in Brit- ain between the 6th and 8th centuries in which no rats have been found. Some would argue that this absence of rats is more apparent than real, reflecting inadequate sampling strategies for small mammal remains. O‘Connor’s meticu- lous work at York, however, provides irrefuta- ble evidence for the absence of rat in that city during the Anglian period, a conclusion reached after extensive sieving and the recov- ery of the bones of many other small mammals (see O’Connor 1988a & 1991). Similar negative evidence from elsewhere in northern and cen- tral Europe reveals rat extinction to have been widespread throughout what had been the Western Roman Provinces. Only in the East- ern Roman Empire (later the Byzantine Empire) is there definite proof for a continuity of R. rattus from the 6th-9th centuries AD: their bones have been recorded from 6th-century Corsica (Vigne & Marinval-Vigne 1985), in lev- els dated 6th-7th century AD at Naples, Italy (A. King pers. comm. 1984), in Byzantine depos- its at Kalapodi, Boeotia, Greece (Stanzel 1991: 121) and at Apamea, northern Syria, in levels dated to 6th-7th century AD (Gautier 1984). In the Byzantine Empire, the Roman economy and lifestyle continued largely uninterrupted, while northern Europe suffered barbarian incursions, urban decay and - perhaps more importantly for rats - the virtual severance of commercial contact with the Mediterranean (Byzantine) world (see Lewis 1958: 38 & 93).
This enforced isolation of the old Roman At- lantic Provinces, including Britain and western Gaul, meant there would be no further replen- ishment of their rat populations, vulnerable from their being at the extreme climatic limits to the geographic range of this subtropical species. The continued survival of R. rattus in northern lati- tudes had depended on close association with humans, which the breakdown in heated life-
styles threatened. One contributory factor may have been the replacement of Roman stone buildings, many heated by hypocaust systems, with wooden huts that were ‘little better than temporary shelters’ (Myers, of Anglo-Saxon houses at Sutton Courtney, referenced by Loyn 1962: 42). Draughty and cold, these would have provided poor harbourage for the warmth- loving black rat. Nevertheless, the sunken houses in York, from the time of Viking settle- ment, hear certain similarities in mode and p lan of construct ion wi th Anglo-Saxon Grubenhauser (Hills 1986: 194) and these were infested with rats (see below). An additional factor must therefore have brought about the demise of the black rat in northern Europe sometime during the 6th century AD. It could, perhaps, have been a deterioration in climate: proxy records from five upland blanket mire sites in the British Isles show cooler tempera- tures and prolonged wetness c. 1400 BP (radio- carbon calibrated date range AD 415-790) according to Blackford & Chambers (1991), and climatic deterioration in the mid 6th century is also shown by Irish tree-ring studies (Baillie & Munro 1988).
Rats and the Viking trade revival Armitage et al. (1984) concluded that the re- appearance of R. rattus in Britain was coinci- dent wi th the revival and expansion of international trade in the Anglo-Scandinavian1 later Anglo-Saxon periods, when trading ports were established along the eastern and south- ern coasts and on navigable rivers (Reynolds 1977). O’Connor (1991: 319) further considered this aspect, believing the presence of rats in Vi- king York - as evidenced by their remains at Coppergate, in 9th century levels - had ‘resulted from occupation of the city by Scandinavian set- tlers . . . [when] the city was receiving a substan- tial input of people and cargoes, and the means of introduction was certainly available’.
If, as now seems proven, there was a new invasion of black rats into Britain during the late 9th century - rather than an expansion (recovery phase) of relict pockets (surviving isolated colonies) of the Roman rat population - where did these animals come from? If R. rattus became extinct throughout northern Europe in the Dark Ages, the source(s) must have been elsewhere, in the Byzantine Empire or perhaps even further eastwards, in the Mus-

UNWELCOME COMPANIONS: ANCIENT RATS REVIEWED 235
lim world - and Viking traders had by the 9th century established direct trade links to both these civilizations.
Viking voyages made in the 9th century along the western seaboard of Europe and into the Mediterranean appear to have been raid- ing/exploratory rather than trading ventures; they therefore probably provided little oppor- tunity for transport of rats from the Mediterra- nean. We should look instead to the principal commercial route to Byzantium, by way of Kievan Russia. As discussed by Martin (1987: 128) Swedish merchant mariners ‘penetrated by way of the Baltic and through the heart of Russia to Constantinople dragging their ships overland between the navigable waterways of- fered by the great river systems’. This route extended along the river systems of the Divana and Drueper, and thence to the Black Sea via Konugard (Kiev). From the Black Sea port of Berezany, merchants could sail direct to Mikligard (Constantinople), the administrative and commercial hub of the Byzantine Empire.
Another Viking trade route crossed the Bal- tic to connect with Holmgard (Novgorod) in Russia, then followed the Volga river system to the Caspian Sea, across which lay the port of Gurgan and the overland caravan route to Baghdad, then the largest city in the world and the heart of Muslim civilization. This trade link may hold great significance for the origins of the second population of black rats, as Mus- lim merchants had themselves established a vigorous commerce with the peoples of south- east Asia and the Far East (the presumed home- land of R. rattus!). Expansion of the Islamic faith after AD 632 had by the 9th century brought stability in central Asia and restored ‘contacts between the eastern and western halves of the Euroasian heartland’ previously disrupted by the barbarian incursions (Barra- clough 1989: 109). Trading voyages were un- dertaken by Arab dhows (sturdy lateen-rigged ocean-going ships) between Arabia, India and China, in an extensive maritime activity which would have given opportunities for further in- flux of R. rattus from southeast Asia into south- west Asia via the Persian Gulf. Having reached and infested Baghdad, these rats could have been transported through central Russia and into the Baltic region, eventually to spread along the maritime trade routes to the north- ern European countries including Britain. Arab
traders during this same period (8th-10th cen- tury) also probably were responsible for first introducing black rats into eastern and south- ern Africa, as evidenced by R. rattus bones found at Pont Drift, northern Transvaal, and Ndondondwane, Natal (Plug & Dippenaar 1979: 82; Voigt & von den Driesch 1984: 100).
There is zooarchaeological evidence for the passage of black rats along either or both of the major Viking trade routes during the 9th cen- tury. The seaport of Birka in Sweden, during this period, served as the key Baltic trading settlement, the main destination of goods brought to Scandinavia from Byzantium and the Islamic east (Graham-Campbell & Kidd 1980: 45). Excavations in Birka yielded skel- etal remains of R. rattus from levels dated 800- 900 AD (Lepiksaar referred to in de Bruyn 1981: 64). Once in Birka, rats had an opportunity of crossing the sea westwards to York, and other English east-coast ports. Given that these Vi- king trade routes also reached Ireland, there is an inexplicable lack of evidence for R. rattus there: no rat bones were found by McCormick in the large samples of faunal material from loth- to 11th-century levels in Viking Dublin, or from the 11th-century urban site at Water- ford (McCormick pers. comm. 1992). Only in the Early Christian level at Ruthmullan, a raised rath site on the coast in County Down (Lynn 1982: 154), does R. rattus appear in Ire- land prior to the Norman occupation of AD 1169 (McCormick 1991).
An equally inexplicable absence occurs at the important early medieval emporium of Dorestad (present day Duurstede in the Neth- erlands) and at the flourishing Anglo-Saxon trading settlement of Hamwic (early medieval Southampton) (Prummel 1983: 245-6). These settlements pre-date the Viking trade with By- zantium and the Islamic east, yet reappraisal of northern maritime history shows the possi- bility of an earlier reintroduction of this rodent species. Haywood (1991) sees that ‘the capa- bilities of the pre-Viking Germanic seafarers [have] been underestimated’. In fact, Frankish naval forces were employed widely from the late 7th-late 8th centuries: on inland water- ways in support of military operations against the Slavs, Saxons and Avars; on the open seas, in the western Mediterranean campaigning against Muslim pirates from Spain and Africa, and as far east as the Adriatic fighting against

236 PHILIP L. ARMITAGE
Byzantium for control of Venice. All this na- val activity in the Mediterranean must surely have provided some opportunity for rats to be transported into northern Europe in Frankish vessels returning home. Before its destruction by raiding Vikings in AD 863, Dorestad was the principal centre of activity of Friesian com- merce in western Europe; it continued as an important trading settlement under Frankish control throughout the late 8th/early 9th cen- tury (see Loyn 1962: 82-3). Had Frankish war- ships brought rats from the Mediterranean, they should have been present in Dorestad, from where they could have spread to York, Hamwic, London and other English trading settlements. Part of the answer may be found in the organi- zation of the Frankish fleets, if those operating in the Mediterranean sailed from ports along the southern coast of the Frankish Kingdom, isolated from those stationed in the North Sea. Frankish seafaring activity in the Mediterra- nean was essentially naval rather than com- mercial; as Lewis (1958: 243) observed, ‘the world of the Carolingians faced north rather than south. . . . Its economic centre also was moving to the northeast towards Germany’.
Regional disputes and fragmentation in Carolingian France had by AD 900 resulted in the ‘economy of Poitou, the Seine and the in- terior of Picardy [becoming] essentially local in nature , , . and almost no commerce existed between [these regions] and the Mediterranean’ (Lewis 1958: 294). The internal isolation of much of France probably explains the late re- introduction of R. rattus which seems to have been no earlier than the 11th century, some two centuries after its reappearance in Britain: in this connection, the 11th-century record of black rat from the monastic site at La CharitB- sur-Loire (the earliest for medieval France) is of significance (Audoin-Rouzeau 1986: 44).
In marked contrast, 9th-century England seems to have maintained contact with the Medi- terranean, and by the loth century there was considerable commerce between the two regions. Italian merchants traded in England, whose own merchants, in turn, visited Italy (see Lewis 1958: 299,301,305). Thismay explainthelate 10th-cen- tury record of R. mf fus in London, an immature tibia at St Magnus in the City (Annitage 1979).
Opportunities for rats to spread from the Mediterranean to northern Europe were greatly facilitated from the 11th century onward when
the Normans extended their sphere of influ- ence into the Mediterranean, opening again the old Roman commerce routes, including the important RhGne-Rhine route. Throughout western Europe, phenomenal human popula- tion growth in the High Medieval period (11th- 13th centuries) provided optimal conditions for proliferation of rats. Larger populations, rising rural prosperity and resurgent international trade precipitated a remarkable phase in urban expansion (Butler 1975; Platt 1976: 21).
All this would have favoured rat infestation, and the archaeological record is beginning to confirm that R. rattus did extend its geographic range throughout northern Europe. Rat remains at Abingdon, Ascot Doilly, Tetsworth, Exeter, Reigate, Middleton Stoney, Stamford, South Witham, Ludgershall, York, Kilton Castle and Lincoln (West in Armitage et al. 1984) indi- cate that in Britain R. rattus was well estab- lished by the High Medieval period. Giraldus Cambrensis’ reference in Itinerarium Kam- beriae of c. 1191 AD (Public Record Office 1868: 111) further shows that rats had spread as far westwards as Wales, thereby re-establishing their Roman distribution pattern.
Morphological changes and the ‘Little Ice Age’ Workers in the early 1980s noticed the extraor- dinarily large size apparent in later medieval/ early Tudor rat bones from British archaeologi- cal sites. It seemed that modern black rats were smaller. This size difference may have arisen out of the lack of interspecific competition: R. rattus entering northern Europe in medieval times found an unoccupied niche for, until the introduction of R. norvegicus sometime in the early 18th century, it enjoyed an exclusive op- portunity to exploit the food resources and harbourage of human settlements. Without ri- val rodent species or any real threat from hu- mans (whose eradication measures were largely ineffectual), R. rattus was, by this model, able to attain full optimal body size.
Another factor worthy of consideration is linked to Bergmann’s ecogeographical rule (Mayr 1963: 318-21). Was the spread of this sub-tropical rodent into cooler and wetter northern latitudes accompanied by adaptive physiological and morphological changes?
Coat colour adaptations Marked changes in the original wild-type

UNWELCOME COMPANIONS: ANCIENT RATS REVIEWED 237
FIGURE 2. Black rut as depicted b y Swiss naturalist-physician Gesner (1551).
pelage are evident in Swiss naturalist Konrad Gesner’s description of the local rats he saw, which were: ‘colore subniger, vel fuscus, qui ventrem versus dilutior est’ (Gesner 1551) (FIG- URE 2). This melanic phenotype then, as now, was apparently predominant throughout much of northernlwestern Europe; according to de 1’Isle (1865), it arose from two mutational trans- formations of the original wild-type coat col- our, partly as a result of climatic influences. The white belly of the wild type changed to grey when R. rattus entered Egypt and the Mediterranean region; the second change (in the colour of the back and flanks), from brown to black, followed when its range later extended even further, into central and northern Europe (Tomich & Komi 1966 is a useful synopsis of the genetic basis of these changes).
The coat colour mutations within the commensal form of R. rattus created much con- fusion among taxonomists, who in the late 19th/early 20th century accorded each of the three main pelage types full subspecific sta- tus, as follows: Rattusmttusmttus (Linnaeus 1758), the ‘true’black
rat (also known as house rat, wharf rat & ship rat). Black dorsum with slate-grey belly.
Rattus rattus alexandrinus (Geoffroy 1803). Agouti (brown) dorsum with grey belly.
Rattus rattus frugivorous (Rafinesque 1814), the fruit rat. Original ‘wild-type’ pelage: agouti dorsum with white belly.
Under this classification, each ‘subspecies’ was believed to exhibit distinctive distributional1 behavioural traits: R . r. rat tus and R . r. alexandrinus as strictly indoor rats (obligate commensals), while R. r. frugivorous was peridomestic, rarely found indoors, but the
most common form on ocean-going ships (Schwarz & Schwarz 1967).
Modern field observations question these distinctions, whether in Cardiff between the wars (Matheson 1931), or southwest Florida today (survey by the present author) and the three main colour forms are nowadays viewed as polymorphic variants of R. rattus rather than as true subspecies (Armitage 1993). What is important, however, is the prevalence of the black (melanic) form in northern Europe, with Twigg (1984: 86) commenting, ‘like all melanics . . . [this form] is hardier in a cold climate’. Another climatic influence may have been the high rainfall levels in northern Europe and the prevailing year-round high humiditylwetness - it is interesting to note that in Hawaii Tomich (1968) found a high prevalence of the melanic form in the higher and wetter parts of the island.
Size adaptations Besides determining the pelage colour, the wet temperate climate of northern Europe may have also resulted in other physiological adaptations in R. rattus. By Bergmann’s rule, European black rats would be more robust and larger in body size than the gracile, smaller animals of sub-tropical latitudes. Modern osteometric data does not support this notion however: in body size and cranial dimensions, R. rattus speci- mens collected in sub-tropical southwest Florida and examined by the author between 1988 and 1991 appear similar to their counter- parts in northern Europe. One explanation for this unexpected finding is that modern black rats in Europe may be relatively recent arriv- als, which do not therefore carry the traits char- acteristic of the medieval population. In Britain
238 PHILIP L. ARMITAGE
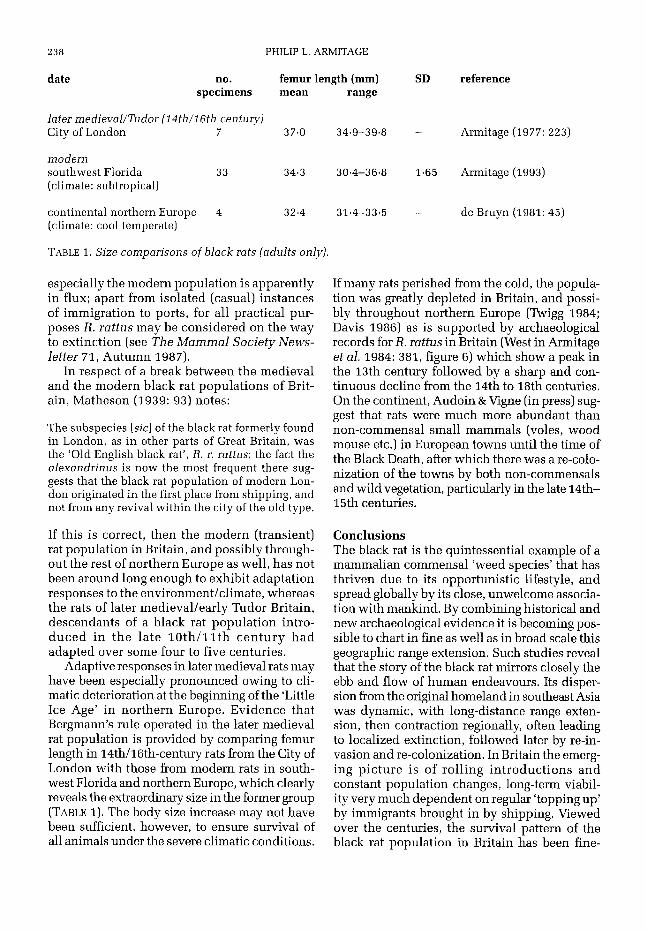
date no. femur length (mm) SD specimens mean range
later medievallTudor [ I 4th/l6th century) City of London 7 37.0 34.9-39.8 -
modern
(climate: subtropical) southwest Florida 33 34.3 30.4-36.8 1.65
continental northern Europe 4 32.4 31.4-33.5 - (climate: cool temperate)
TABLE 1. Size comparisons of black nits [adults only).
reference
Armitage (1977: 223)
Armitage (1993)
de Bruyn (1981: 45)
especially the modern population is apparently in flux; apart from isolated (casual) instances of immigration to ports, for all practical pur- poses R. rattus may be considered on the way to extinction (see The Mammal Society News- letter 71, Autumn 1987).
In respect of a break between the medieval and the modern black rat populations of Brit- ain, Matheson (1939: 93) notes:
The subspecies [sic] of the black rat formerly found in London, as in other parts of Great Britain, was the ‘Old English black rat’, R. r. rattus; the fact the alexandrinus is now the most frequent there sug- gests that the black rat population of modern Lon- don originated in the first place from shipping, and not from any revival within the city of the old type.
If this is correct, then the modern (transient) rat population in Britain, and possibly through- out the rest of northern Europe as well, has not been around long enough to exhibit adaptation responses to the environment/climate, whereas the rats of later medieval/early Tudor Britain, descendants of a black rat population intro- duced in the late 1 0 t h / l l t h century had adapted over some four to five centuries.
Adaptive responses in later medieval rats may have been especially pronounced owing to cli- matic deterioration at the beginning of the ‘Little Ice Age’ in northern Europe. Evidence that Bergmann’s rule operated in the later medieval rat population is provided by comparing femur length in 14th/l6th-century rats from the City of London with those from modern rats in south- west Florida and northern Europe, which clearly reveals the extraordinary size in the former group (TABLE 1). The body size increase may not have been sufficient, however, to ensure survival of all animals under the severe climatic conditions.
If many rats perished from the cold, the popula- tion was greatly depleted in Britain, and possi- bly throughout northern Europe (Twigg 1984; Davis 1986) as is supported by archaeological records for R. rattus in Britain (West in Armitage et al. 1984: 381, figure 6) which show a peak in the 13th century followed by a sharp and con- tinuous decline from the 14th to 18th centuries. On the continent, Audoin & Vigne (in press) sug- gest that rats were much more abundant than non-commensal small mammals (voles, wood mouse etc.) in European towns until the time of the Black Death, after which there was a re-colo- nization of the towns by both non-commensals and wild vegetation, particularly in the late 14th- 15th centuries.
Conclusions The black rat is the quintessential example of a mammalian commensal ‘weed species’ that has thriven due to its opportunistic lifestyle, and spread globally by its close, unwelcome associa- tion with mankind. By combining historical and new archaeological evidence it is becoming pos- sible to chart in fine as well as in broad scale this geographic range extension. Such studies reveal that the story of the black rat mirrors closely the ebb and flow of human endeavours. Its disper- sion from the original homeland in southeast Asia was dynamic, with long-distance range exten- sion, then contraction regionally, often leading to localized extinction, followed later by re-in- vasion and re-colonization. In Britain the emerg- ing picture is of rolling introductions and constant population changes, long-term viabil- ity very much dependent on regular ‘topping up’ by immigrants brought in by shipping. Viewed over the centuries, the survival pattern of the black rat population in Britain has been fine-
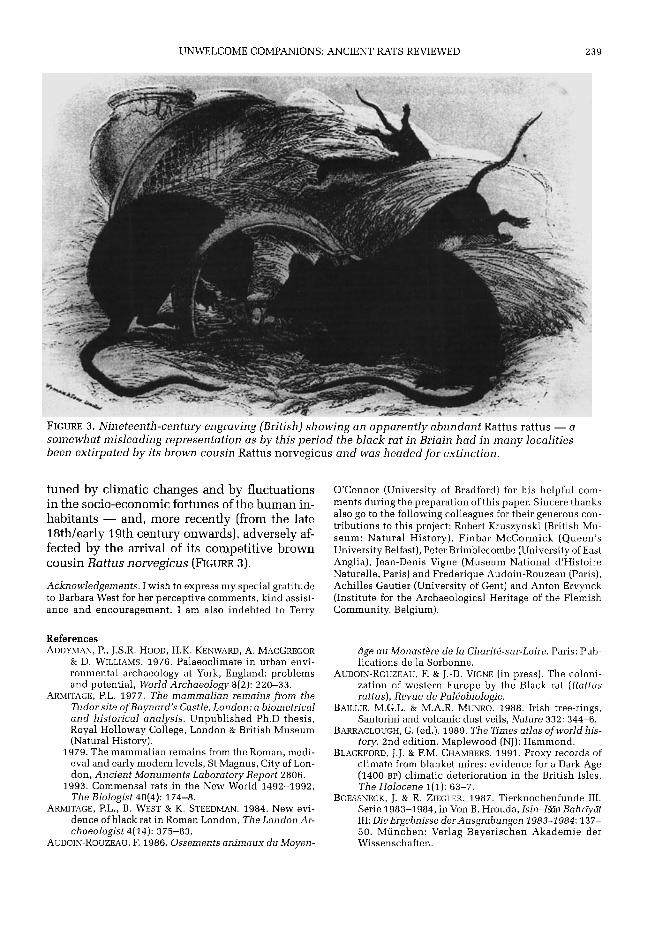
UNWELCOME COMPANIONS: ANCIENT RATS REVIEWED 239
FIGURE 3. Nineteenth-century engraving (British) showing an apparently abundant Rattus rattus - a somewhat misleading representation as b y this period the black rat in Briain had in many localities been extirpated b y its brown cousin Rattus norvegicus and was headed for extinction.
tuned by climatic changes and by fluctuations in the socio-economic fortunes of the human in- habitants - and, more recently (from the late 18th/early 19th century onwards), adversely af- fected by the arrival of its competitive brown cousin Rattus norvegicus (FIGURE 3).
Acknowledgements. I wish to express my special gratitude to Barbara West for her perceptive comments, kind assist- ance and encouragement. I a m also indebted to Terry
References ADDYMAN, P., J.S.R. HOOD, H.K. KENWARD, A. MACGREGOR
& D. WILLIAMS. 1976. Palaeoclimate in urban envi- ronmental archaeology at York, England: problems and potential, World Archaeology 8(2): 220-33.
ARMITAGE, P.L. 1977. The mammalian remains from the Tudor site of Baynard’s Castle, London: a biornetrical and historical analysis. Unpublished Ph.D thesis, Royal Holloway College, London & British Museum (Natural History).
1979. The mammalian remains from the Roman, medi- eval and early modern levels, St Magnus, City of Lon- don, Ancient Monuments Laboratory Report 2806.
1993. Commensal rats in the New World 1492-1992, The Biologist 40(4): 174-8.
ARMITAGE, P.L., B. WEST & K. STEEDMAN. 1984. New evi- dence of black rat in Roman London, The London Ar- chaeologist 4(14): 375-83.
AUDOIN-ROUZEAU, F. 1986. Ossements animaux du Moyen-
O’Connor (University of Bradford) for his helpful com- ments during the preparation of this paper. Sincere thanks also go to the following colleagues for their generous con- tributions to this project: Robert Kruszynski (British Mu- seum: Natural History), Finbar McCormick (Queen’s University Belfast), Peter Brimblecombe (University of East Anglia), Jean-Denis Vigne (Museum National d’Histoire Naturelle, Paris) and Frederique Audoin-Rouzeau (Paris), Achilles Gautier (University of Gent) and Anton Ervynck (Institute for the Archaeological Heritage of the Flemish Community, Belgium).
6ge au Monasthre de la CharitB-sur-Loire. Paris: Pub- lications de la Sorbonne.
AUDOIN-ROLIZEAU, F. & J.-0. VIGNE ( in press). The coloni- zation of western Europe by the Black rat (Rattus rattus), Revue de Palhbiologie.
BAILLIE, M.G.L. & M.A.R. MUNRO. 1988. Irish tree-rings, Santorini and volcanic dust veils, Nature 332: 344-6.
BARRACLOUGH, G. (ed.). 1989. The Times atlas ofworld his- tory. 2nd edition. Maplewood (NJ): Hammond.
BLACKFORD, 7.7. & F.M. CHAMBERS. 1991. Proxy records of climate from blanket mires: evidence for a Dark Age (1400 BP) climatic deterioration in the British Isles, The Holocene 1(1): 63-7.
BOESSNECK, J. & R. ZIEGLER. 1987. Tierknochenfunde 111. Serie 1983-1984, in Von B. Hrouda, Isin-IEn Bahriydt 111: Die Ergebnisse der Ausgrabungen 1983-1984: 137- 50. Miinchen: Verlag Bayerischen Akademie der Wissenschaften.

240 PHILIP L. ARMITAGE
BRUYN, T. DE. 1981. Huissrat (Rattus rottus) en Bruine rat (R. norvegicus) in archeozoologische context. Unpub- lished dissertation, Rijksuniversiteit Gent.
BUTLER, L. 1975. The evolution of towns: planted towns after 1066, in M.W. Barley (ed.), The plans and to- pography of medieval towns in England and Wales: 32-48. London: Council for British Archaeology. Re- search report 14.
DAVIS, D.E. 1986. The scarcity of rats and the Black Death: an ecological history, Journal of Interdisciplinary His- tory 16(3): 455-70.
GAUTIER, A. 1984. La faune de quelques maisons d’Apamee, i n J . Balty (ed . ) , Aparnge de Syrie. Bilan des recherches archgologiques 1973-1979: 305-37. Bruxelles: Centre Belge de Recherches Archbo- logiques & Apam6e de Syrie.
GESNER, K. 1551. Historia Animalium. Zurich. GRAHAM-CAMPBELL, J. & D. KIDD. 1980. The Vikings. Lon-
don: British Museum Publications. HAYWOOD, J. 1991. Dark Age naval power: 11 reassessment
of Frankish and Anglo-Saxon seafaring activity. Lon- don: Routledge.
HENGEVELD, R. 1989. Dynamics of biological invasions. New York (NY): Chapman & Hall.
HILLS, C. 1986. Blood of the British from Ice Age to Nor- man Conquest. London: Guild.
ISLE, A. DE L’. 1865. De l’existence d’une race negre chez le rat, ou de l’identite specifique d u Mils rattus et du Mus alexandrinus, Annales de Sciences Naturelles serie 5(4): 173-222.
LEWIS, A.R. 1958. The northern seas: shipping and com- merce in Northern Europe A D 300-1 100. Princeton (NJ): Princeton University Press.
LOYN, H.R. 1962. Anglo-Saxon England and the Norman Conquest. London: Longmans, Green.
LYNN, C.J. 1982. The excavation of Rathmullan, a raised rath and motte in County Down, Ulster Journal of Archaeology 44-5 (1981-82): 65-171.
MARSDEN, P. & B. WEST. 1992. Population change in Ro- man London, Britannia 23: 133-40.
MARTIN, C. 1987. The Viking world, i n P. Throckmorton (ed.), The sea remembers: shipwrecks and archae- ology: 128-33. New York (NY): Weidenfeld & Nicolson.
MATHESON, C. 1931. TheBrown and theBlackRatin Wales. Cardiff: National Museum of Wales.
1939. A survey of the status of Rattus rattus and its subspecies in the seaports of Great Britain and Ire- land, Journal of Animal Ecology 8: 76-93.
MAYR, E. 1963. Animal species and evolution. Cambridge (MA): Harvard University Press.
MCCORMICK, F. 1991. The effect of the Anglo-Norman set- tlement on Ireland’s wild and domesticated fauna, in P.J. Crabtree & K. Ryan (ed.), Animal use and culture change: 41-52. Philadelphia (PA): MASCA Research Papers in Science and Archaeology. Supplement to volume 8 (1991).
MERRIFIELD, R. 1983. London: city of the Romans. London: Ernest Benn.
NEYLAND, R.S. 1992. The seagoing vessels on Dilmun seals, in D.H. Keith & T.L. Carrel1 (ed.), Underwaterarchae- ologyproceedings of the Society for Historical Archae- ology Conference at Kingston, Jamaica 1992: 68-74. Tucson (AZ): Society for Historical Archaeology.
NIETHAMMER, J . VON 1975. Zur taxonomie u n d Aus- breitungsgeschichte der Hausratte (Rattus rattus), Zoologischer Anzeiger, Jena 194: 405-15.
O’CONNOR, T.P. 1988a. The case of the absent rat, Archae- ologyin York Interim 13(4): 39-41.
1988h. Bones from the General Accident site, Tanner Row, Archaeology of York 15(2): 61-136.
1991. On the lack of bones of the ship rat Rattus rattus from Dark Age York, Journal of Zoology, London 224:
OLIPHANT, M. 1992. The atlas of the ancient world. New York (NY): Simon & Schuster.
PLATT, C. 1976. TheEnglish medieval town. London: Secker & Warburg.
PLUG, I. & N.J. DIPPENAAR. 1979. Evidence of Rattus rattus (House rat) from Pont Drift, an Iron Age site in North- ern Transvaal, South Afiican Journal of Science 75: 82.
PRUMMEL, W. 1983. Excavations at Dorestad 2: Earlymedieval Darestad: an archaeozoological study. Amersfoort: ROB.
PUBLIC RECORD OFFICE. 1868. Rolls Series VI: Book 11. RACKHAM, J. 1979. Rattus rattus: the introduction of the
black rat into Britain, Antiquity 53: 112-20. REUMER, J.W.F. 1986. Note on the spread of the black rat,
Rattus rattus, Mammalia 50(1): 118-19. REYNOLDS, S. 1977. A n introduction to the history of Eng-
lish medieval towns. Oxford: Clarendon Press. SCHWARZ, E. & H.K. SCHWARZ. 1967. A monograph of the
Rattus rattus group, Anales de la Escuela Nacional de Ciencias Biologicas, Mexico 14: 79-178.
SHELDON, H. 1975. A decline in the London settlement AD
150-250? London Archaeologist 2(11): 278-84. SLOANE, R.E. & M.A. DUNCAN, 1978. Zooarchaeology of
Nichoria, in G.Rapp & S.E. Aschenbrenner (ed.), Ex- cavations at Nichoria in Southwest Greece Vol.1: Site, environs and techniques: 60-77. Minneapolis (MN): University of Minnesota Press.
STANZEL, M. 1991. Die Tierreste aus dem Artemis-/Apollon- Heiligtum bei Kalapodi in Bootien/Griechenland. Unpublished dissertation, Universitat Miinchen.
TEICHERT, M. VON 1985. Beitrag zur Faunengeschichte der Hausratte, Rattus rattus (L.), Zeitschrift fur Archao- logie 19: 263-9.
TCHERNOV, E. 1984. Commensal animals and human sedentism in the Middle East, in J. Clutton-Brock & C. Grigson (ed.), Animalsin archaeology3: Earlyherd- ers and theirflocks: 91-115. Oxford: British Archaeo- logical Reports. International series S202.
TOMICH, P.Q. 1968. Coat color in wild populations of the roof rat in Hawaii, Journal of Mammalogy49(1): 74-82.
TOMM, P.Q. & H.T. KAMI. 1966. Coat color inheritance of the roof rat in Hawaii, Journal of Mammalogy47(3): 423-31.
TWIGG, G. 1984. The Black Death: a biological reappraisal. London: Batsford.
VIGNE, J.-D. & J.-M. FEMOLANT. 1991. Premiere observation de la presence d u rat noir (Rattus rattus) en France septentrionale a la periode gallo-romaine (Picardie, Ier siecle apres J.-C.), Mammalia 55(2): 319-21.
VIGNE, J.-D. & M.-C. MARINVAL-VIGNE. 1985. Le rat en Corse au 6 siecle apres J.-C.? Mammalia 49(1): 138-9.
VOIGT, E.A. & A. VON DEN DRIESCH. 1984. Preliminary re- port on the faunal assemblage from Ndondondwane, Natal, Annals of the Natal Museum 26(1): 95-104.
YOSIDA, T.H. 1980. Cytogenetics of the Black Rat. Tokyo: University of Tokyo Press.
318-20.